This article was downloaded by: [Academia Sinica - Taiwan] On: 19 December 2014, At: 01:13 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Click for updates Cell Cycle Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/kccy20 Regulation and roles of Cdc7 kinase under replication stress Masayuki Yamada a , Hisao Masai b & Jiri Bartek ac a Institute of Molecular and Translational Medicine; Faculty of Medicine and Dentistry; Palacky University; Olomouc, Czech Republic b Genome Dynamics Project; Department of Genome Medicine; Tokyo Metropolitan Institute of Medical Science; Tokyo, Japan c Danish Cancer Society Research Center; Copenhagen, Denmark Published online: 19 May 2014. To cite this article: Masayuki Yamada, Hisao Masai & Jiri Bartek (2014) Regulation and roles of Cdc7 kinase under replication stress, Cell Cycle, 13:12, 1859-1866, DOI: 10.4161/cc.29251 To link to this article: http://dx.doi.org/10.4161/cc.29251 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http:// www.tandfonline.com/page/terms-and-conditions

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article was downloaded by: [Academia Sinica - Taiwan]On: 19 December 2014, At: 01:13Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Click for updates
Cell CyclePublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/kccy20
Regulation and roles of Cdc7 kinase under replicationstressMasayuki Yamadaa, Hisao Masaib & Jiri Bartekac
a Institute of Molecular and Translational Medicine; Faculty of Medicine and Dentistry;Palacky University; Olomouc, Czech Republicb Genome Dynamics Project; Department of Genome Medicine; Tokyo Metropolitan Instituteof Medical Science; Tokyo, Japanc Danish Cancer Society Research Center; Copenhagen, DenmarkPublished online: 19 May 2014.
To cite this article: Masayuki Yamada, Hisao Masai & Jiri Bartek (2014) Regulation and roles of Cdc7 kinase under replicationstress, Cell Cycle, 13:12, 1859-1866, DOI: 10.4161/cc.29251
To link to this article: http://dx.doi.org/10.4161/cc.29251
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
PersPective
www.landesbioscience.com cell cycle 1859
cell cycle 13:12, 1859–1866; June 15, 2014; © 2014 Landes Bioscience
PersPective PersPective PersPective
Cdc7 (cell division cycle 7) kinase together with its activation subunit
ASK (also known as Dbf4) play pivotal roles in DNA replication and contribute also to other aspects of DNA metabo-lism such as DNA repair and recombina-tion. While the biological significance of Cdc7 is widely appreciated, the molecu-lar mechanisms through which Cdc7 kinase regulates these various DNA transactions remain largely obscure, including the role of Cdc7-ASK/Dbf4 under replication stress, a condition associated with diverse (patho)physi-ological scenarios. In this review, we first highlight the recent findings on a novel pathway that regulates the sta-bility of the human Cdc7-ASK/Dbf4 complex under replication stress, its interplay with ATR-Chk1 signaling, and significance in the RAD18-depen-dent DNA damage bypass pathway. We also consider Cdc7 function in a broader context, considering both physiological conditions and pathologies associated with enhanced replication stress, par-ticularly oncogenic transformation and tumorigenesis. Furthermore, we inte-grate the emerging evidence and propose a concept of Cdc7-ASK/Dbf4 contribut-ing to genome integrity maintenance, through interplay with RAD18 that can serve as a molecular switch to dictate DNA repair pathway choice. Finally, we discuss the possibility of targeting Cdc7, particularly in the context of the Cdc7/RAD18-dependent translesion synthe-sis, as a potential innovative strategy for treatment of cancer.
Background
The serine–threonine kinase Cdc7 is essential for chromosomal DNA replica-tion in a wide spectrum of species, being conserved from yeast to human.1-4 The activity of the catalytic subunit Cdc7 is positively regulated by a complex forma-tion with its activation subunit, the Dbf4 protein, also known as ASK (activator of S-phase kinase in human).5-10 The Cdc7–ASK/Dbf4 complex is therefore also commonly referred to as the Dbf4-dependent kinase, or DDK in short. DDK regulates the timing of DNA rep-lication origin firing throughout S phase mainly by phosphorylation of MCM proteins, the major components of repli-cative helicase.11-13
When the replication fork stalls, due either to endogenous obstacles or upon exposure of a cell to various genotoxic insults, the DNA replication checkpoint pathway becomes activated14 and sup-presses further origin firing or slows down the fork progression. In budding yeast Saccharomyces cerevisiae, Rad53 checkpoint kinase (an ortholog of human Chk2) directly phosphorylates Dbf4 upon fork stalling to block late origin firing, most likely through downregula-tion of the DDK activity,15-18 although this is not proven yet. Similarly, an in vitro study using Xenopus egg extracts revealed that DDK activity is inhibited upon DNA damage caused by etoposide treatment in an ATR kinase-dependent manner.19 However, later work using a similar experimental system presented a
Regulation and roles of Cdc7 kinase under replication stress
Masayuki Yamada1,*, Hisao Masai2, and Jiri Bartek1,3,*1Institute of Molecular and Translational Medicine; Faculty of Medicine and Dentistry; Palacky University; Olomouc, Czech Republic; 2Genome Dynamics
Project; Department of Genome Medicine; Tokyo Metropolitan Institute of Medical Science; Tokyo, Japan; 3Danish Cancer Society Research Center;
Copenhagen, Denmark
Keywords: Cdc7 kinase, DDK, RAD18, TLS, DNA damage bypass, replication checkpoint, DNA repair pathway choice
Abbreviations: APC/C, anaphase promoting complex/cyclosome; ATR, ataxia telangiectasia and Rad3-related protein; CDK, cyclin-dependent kinase; DDK, Dbf4-dependent kinase; DDR, DNA damage response; FA, Fanconi anemia; HR, homologous recombination; PIP box, PCNA-interacting protein box; RING domain, really interesting new gene domain; TLS, translesion synthesis
Submitted: 05/05/2014
Accepted: 05/15/2014
Published Online: 05/19/2014
http://dx.doi.org/10.4161/cc.29251
*Correspondence to: Jiri Bartek; Email: [email protected]; Masayuki Yamada; Email: [email protected]
Dow
nloa
ded
by [
Aca
dem
ia S
inic
a -
Tai
wan
] at
01:
13 1
9 D
ecem
ber
2014

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
1860 cell cycle volume 13 issue 12
contradictory result, namely preserved DDK activity upon DNA damage.20 It is also controversial whether the human DDK is inactivated under genotoxic stress conditions. Previously, it was reported that Cdc7 kinase activity in budding yeast is downregulated in response to hydroxy-urea treatment that arrests cells in early S phase through inhibition of ribonucleo-tide reductase.18 Earlier work with human cells also concluded that the DDK activ-ity is inhibited upon etoposide treatment through the ATR-dependent inhibition of formation of the human DDK complex.21 However, more recent reports proposed that the human DDK is not inactivated under various genotoxic treatments.20,22 Recent reports showed that the human ASK/Dbf4 is a direct substrate of the upstream DNA damage response kinases ATM and/or ATR, and that such phos-phorylation of ASK/Dbf4 is required for activation of the so-called intra-S phase checkpoint that suppresses DNA repli-cation upon exposure of S-phase cells to ionizing radiation.23 This is consistent with the possibility that the human DDK function is downregulated upon genotoxic stress, which would lead to inhibition of the late origin firing. Interestingly, how-ever, the authors found that the phos-phorylation of ASK/Dbf4 did not affect the kinase activity of Cdc7, indicating that this regulatory phosphorylation may impact a currently unknown mechanism, such as regulation of some protein–pro-tein interactions that may facilitate sup-pression of late origin firing in human cells.23 It should be also noted that the human DDK activity is required for Chk1 activation, most probably through phos-phorylating Claspin, an adaptor protein essential for ATR-dependent phosphoryla-tion of Chk1,24,25 consistent with the idea that the DDK is active under replication stress.
Taken together, such divergent and partly contradictory reports highlight the important questions to be addressed regarding the human DDK: (1) Is the human DDK activity preserved under genotoxic/replication stress or not? (2) If it does remain active, is it only a certain subset of Cdc7 that is active, and what is the role of the DDK activity under such conditions?
In addition, although cell cycle-dependent regulation of transcription was reported for the components of the mam-malian DDK,4,26,27 it has been unclear whether and how the protein abundance of the DDK subunits is regulated upon replication block, and whether there are any differences in such regulatory mecha-nisms between unperturbed conditions vs. replication stress. Physiological and path-ological roles of the DDK activity under genotoxic stress should also be addressed, considering that DDK has been pro-posed as a promising target of anticancer treatment.28-35
Stabilization of DDK on Chromatin in Response
to Replication Stress
Recently, fresh insights have been pro-vided into these important issues. New results showed that under replication stress conditions, human DDK accumu-lates as an active complex on chromatin, and that such stabilization of the DDK protein components is mediated by the ATR–Chk1 kinase pathway-dependent inactivation of the anaphase-promoting complex/cyclosome-Cdh1(APC/CCdh1).36 The APC/CCdh1 ubiquitin ligase is one of the major cell cycle-regulatory machines, which becomes activated in later mitosis, remains active throughout the following G
1 phase, and then is switched off by the
enhanced CDK kinase activity (CDK1 and CDK2 in human cells) from the G
1/S
boundary until early M phase.37-40 Since the replication stress-responsive ATR–Chk1 signaling inhibits CDK activity (including CDK2 in S phase) via check-point pathways, such silencing of CDK could, in principle, lead to re-activation of the APC/CCdh1 at a “wrong” cell cycle phase, for example in S phase, thereby grossly deregulating the DNA replication factories and other S-phase factors. As the recent work clarified,36 this conundrum is solved through concomitant Chk1-medietad inactivation of the APC/CCdh1 complex, through promoting autodegra-dation of Cdh1 under replication stress conditions. Thus, the ATR–Chk1 axis silences the CDKs via checkpoints to delay replication, and at the same time ensures
that the APC/CCdh1- mediated proteolysis is not inappropriately activated in S phase.
Interestingly, and in contrast to replica-tion stress response in S phase, the APC/CCdh1 pathway becomes active in response to DNA double-strand breaks induced by X-ray irradiation or doxorubicin treat-ment, and this plays an important role in G
2 checkpoint activation through degra-
dation of PLK1, an essential kinase that regulates mitotic entry.41,42
It should be noted that DNA repair factors such as 53BP1 and Rap80 are among candidate substrates targeted for degradation by the APC/CCdh1 ubiquitin ligase,43,44 suggesting that the APC/CCdh1 pathway may be also involved in regula-tion of homologous recombination (HR), given a role of these proteins in restrict-ing DNA end resection, an essential step in HR.45,46
Furthermore, the DDK activity itself is required for the stabilization of the human Dbf4/ASK protein, suggesting that one of the important roles of the DDK activity under replication stress is to stabilize ASK/Dbf4, and thereby the DDK complex.36
Central Role of DDK in TLS-Mediated DNA Damage Repair:
Concept of the DDK-RAD18 Interaction as a Molecular Switch
in DNA Repair Pathway Choice
So, what could be the biological advan-tage for the active DDK to be stabilized on chromatin under replication stress? We believe, based on our own data36 and work done mainly on yeast models (see below), that the answer is in the choice of DNA repair pathway, in order to preserve genomic integrity while preventing collapse of the assembled replication fork machin-eries under conditions of replication stress. Previous studies in yeast strongly sug-gested that the DDK plays a crucial role in translesion synthesis (TLS),47,48 a mecha-nism that enables continued DNA replica-tion across the encountered DNA lesion. More recently, human DDK was shown to phosphorylate a cluster of serine residues on RAD18, and this positively regulates interaction between RAD18 and DNA polymerase η (Pol η).49,50 Furthermore, motif-C, a conserved C2H2-type zinc
Dow
nloa
ded
by [
Aca
dem
ia S
inic
a -
Tai
wan
] at
01:
13 1
9 D
ecem
ber
2014
©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
www.landesbioscience.com cell cycle 1861
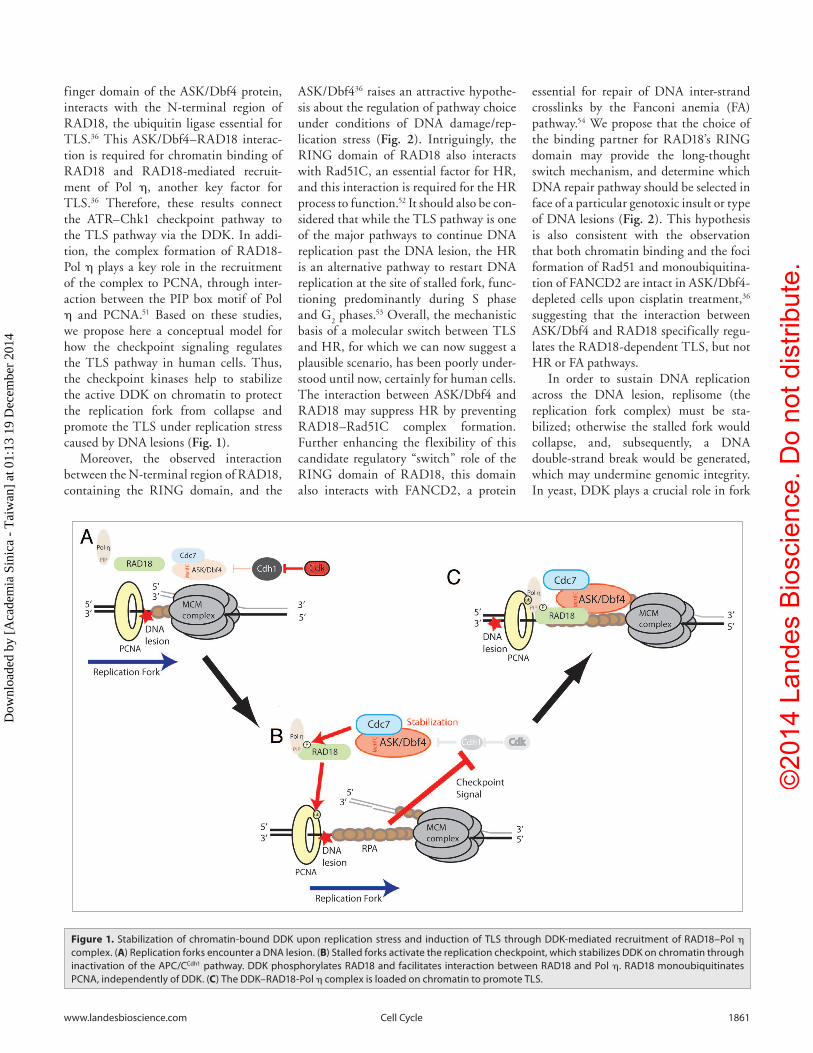
finger domain of the ASK/Dbf4 protein, interacts with the N-terminal region of RAD18, the ubiquitin ligase essential for TLS.36 This ASK/Dbf4–RAD18 interac-tion is required for chromatin binding of RAD18 and RAD18-mediated recruit-ment of Pol η, another key factor for TLS.36 Therefore, these results connect the ATR–Chk1 checkpoint pathway to the TLS pathway via the DDK. In addi-tion, the complex formation of RAD18-Pol η plays a key role in the recruitment of the complex to PCNA, through inter-action between the PIP box motif of Pol η and PCNA.51 Based on these studies, we propose here a conceptual model for how the checkpoint signaling regulates the TLS pathway in human cells. Thus, the checkpoint kinases help to stabilize the active DDK on chromatin to protect the replication fork from collapse and promote the TLS under replication stress caused by DNA lesions (Fig. 1).
Moreover, the observed interaction between the N-terminal region of RAD18, containing the RING domain, and the
ASK/Dbf436 raises an attractive hypothe-sis about the regulation of pathway choice under conditions of DNA damage/rep-lication stress (Fig. 2). Intriguingly, the RING domain of RAD18 also interacts with Rad51C, an essential factor for HR, and this interaction is required for the HR process to function.52 It should also be con-sidered that while the TLS pathway is one of the major pathways to continue DNA replication past the DNA lesion, the HR is an alternative pathway to restart DNA replication at the site of stalled fork, func-tioning predominantly during S phase and G
2 phases.53 Overall, the mechanistic
basis of a molecular switch between TLS and HR, for which we can now suggest a plausible scenario, has been poorly under-stood until now, certainly for human cells. The interaction between ASK/Dbf4 and RAD18 may suppress HR by preventing RAD18–Rad51C complex formation. Further enhancing the flexibility of this candidate regulatory “switch” role of the RING domain of RAD18, this domain also interacts with FANCD2, a protein
essential for repair of DNA inter-strand crosslinks by the Fanconi anemia (FA) pathway.54 We propose that the choice of the binding partner for RAD18’s RING domain may provide the long-thought switch mechanism, and determine which DNA repair pathway should be selected in face of a particular genotoxic insult or type of DNA lesions (Fig. 2). This hypothesis is also consistent with the observation that both chromatin binding and the foci formation of Rad51 and monoubiquitina-tion of FANCD2 are intact in ASK/Dbf4-depleted cells upon cisplatin treatment,36 suggesting that the interaction between ASK/Dbf4 and RAD18 specifically regu-lates the RAD18-dependent TLS, but not HR or FA pathways.
In order to sustain DNA replication across the DNA lesion, replisome (the replication fork complex) must be sta-bilized; otherwise the stalled fork would collapse, and, subsequently, a DNA double-strand break would be generated, which may undermine genomic integrity. In yeast, DDK plays a crucial role in fork
Figure 1. stabilization of chromatin-bound DDK upon replication stress and induction of tLs through DDK-mediated recruitment of rAD18–Pol η complex. (A) replication forks encounter a DNA lesion. (B) stalled forks activate the replication checkpoint, which stabilizes DDK on chromatin through inactivation of the APc/ccdh1 pathway. DDK phosphorylates rAD18 and facilitates interaction between rAD18 and Pol η. rAD18 monoubiquitinates PcNA, independently of DDK. (C) the DDK–rAD18-Pol η complex is loaded on chromatin to promote tLs.
Dow
nloa
ded
by [
Aca
dem
ia S
inic
a -
Tai
wan
] at
01:
13 1
9 D
ecem
ber
2014

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
1862 cell cycle volume 13 issue 12
stabilization upon genotoxic insults.55,56 DDK-mutant yeast cells feature sponta-neous Rad52 foci, suggesting the ongo-ing rescue of collapsed forks by HR.56 An analogous role was also proposed for mammalian DDK,23,57 hence this func-tion of DDK in replisome stabilization to continue DNA replication across lesions appears to be conserved in all eukaryotes. Given that excessive HR can also compro-mise genome integrity, we speculate that by regulating pathway choice, DDK might also help to actively prevent untimely HR, a process that could be accomplished by modulating helicase activities of PARI, FBH1, or RTEL1, for example.58-60
Suppression of Late Origin Firing in Response to Replication Stress
At this moment, it is still unclear whether the entire pool of DDK stays active after exposure to replication stress. Although the bulk activity of DDK may be unaltered after the replication stress, it is possible that checkpoint kinase-mediated
phosphorylation may inactivate or pro-mote dissociation of at least a certain subset of Cdc7–ASK/Dbf4 complexes or those associated with the early-firing pre-RC, and thus render it unavailable for activation of late dormant origins.
If the human DDK remains active under replication stress conditions, how can the firing of late replication origins be suppressed without inhibiting the DDK function? Recently, a potential relevant role of a phosphatase was reported in Xenopus. In the Xenopus cell-free system, protein phosphatase 1 (PP1)-mediated dephosphorylation of MCM proteins, which is triggered by the checkpoint kinases, counteracts the DDK-dependent origin firing without modulating the DDK kinase activity.61 Thus, it might be an intriguing possibility that the check-point kinases activate PP1, which then counteracts the DDK-dependent phos-phorylation of the MCM complex to pre-vent late origin firing. However, there is currently no evidence either in Xenopus egg extracts or in any other systems that depletion or inactivation of phosphatases
override the checkpoint-mediated inhibi-tion of late origin firing. Nor is there any evidence that PP1 is indeed recruited to late origins upon replication stress. These issues need to be examined in the future to test this hypothesis.
Recently, Rif1, a telomere-binding fac-tor in yeasts, was found to be a major reg-ulator of replication timing in both yeasts and mammals.62-64 In the absence of Rif1, replication timing changes dramatically both in yeast and mammalian cells dur-ing normal cell cycle progression. In both species, timing and efficiency of pre-RC assembly is unaffected, but that of Cdc45 changes in the absence of Rif1.62,63 In fis-sion yeast, Rif1 binds close to the late/dor-mant origins and suppresses the firing of nearby origins62 (Kanoh et al. unpublished data). It is unknown whether mammalian Rif1 shows a similar binding preference. Nevertheless, the similarity is striking, in that the absence of Rif1 causes general loss of replication timing regulation through-out the genome.65 It is unlikely that Rif1 is involved in cellular responses to replica-tion stress, since rif1-null yeast cells and
Figure 2. A model for rAD18-dependent DNA repair pathway choice in response to genotoxic insults. rAD18 may function as a molecular switch for DDr pathway choice through alternating partner factors that bind to its riNG domain.
Dow
nloa
ded
by [
Aca
dem
ia S
inic
a -
Tai
wan
] at
01:
13 1
9 D
ecem
ber
2014

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
www.landesbioscience.com cell cycle 1863
Rif1-depleted human cells are resistant to HU.62,65 However, it is of interest that PP1 interacts with Rif1, and that this interac-tion may play a role in Rif1-mediated reg-ulation of replication timing in budding yeast.66
DDK and Cancer
Last but not least, we wish to discuss DDK in the context of tumorigenesis, and also as a candidate target for innovative cancer therapy. The major reason why the DDK-mediated response to replication stress is highly relevant for cancer is that activation of diverse oncogenes and loss of some tumor suppressors evoke replica-tion stress and consequent DNA damage that triggers the checkpoint responses of the ATR–Chk1 and ATM–Chk2 signal-ing cascades, as exemplified in cell culture experiments and documented by analy-ses of clinical specimens from a range of human malignancies.67-75 This anti-cancer barrier provided by the DNA damage checkpoints precedes the activation of the ARF tumor suppressor, the second anti-cancer barrier,76-80 and helps delay or prevent tumor progression by trigger-ing senescence or cell death among the nascent cancer cells.67,68,70,71 However, in those tumors that eventually do progress, the ATM-Chk2-p53 and other check-points are commonly bypassed or inac-tivated,67,72,73 while the ATR–Chk1 axis, which generally helps cells to cope with replication stress (as discussed above), remains operational, as cancer cells need to deal with the omnipresent challenge of enhanced replication stress.81 In such scenario, and given the above discussed roles of DDK under replication stress, the DDK activity (including the ASK/Dbf4-RAD18 role in TLS) may provide one of the stress-support functions on which can-cer cells depend more than their normal counterpart cells.
In addition, we suggest that the occur-rence of much greater endogenous replica-tion stress in cancer cells would provide opportunities for therapeutic intervention by DDK inhibitors. Indeed, several such scenarios could be proposed, as would be rationalized on the basis of the synthetic lethality principle, best documented by
the selective sensitivity of HR- (e.g., BRCA1/2-) defective tumors to PARP inhibitors.82-85 Since the HR pathway is impaired in BRCA-deficient tumors, small-molecule inhibitors of Cdc7 kinase activity (and/or inhibitors of RAD18 ubiquitin ligase, yet to be developed) could be useful as an alternative of PARP inhibi-tors. Indeed, it has been shown that the HR pathway is required for the viability of rad18Δ yeast cells upon chronic low-dose UV light exposure.86,87 It was also reported that double mutants for RAD18 and RAD54, a gene required for HR, exhibit synthetic lethality in chicken DT40 cells, in contrast to only modest impact of either mutation alone.88 Moreover, human cells deficient in the TLS pathway, including RAD18 deficiency, are hypersensitive to cisplatin treatment,89 whose tolerance is also dependent on the HR pathway. Since the FA pathway is proposed to be involved in HR,90-95 the Cdc7 kinase inhibitors could be also effective for tumors in which the FA pathway is impaired.
In conclusion, targeting the DDK activity under conditions of enhanced replication stress and the nodal point of RAD18 interactions in DNA repair pathway choice may inspire novel treat-ment strategies in oncology. Such DDK/RAD18-antagonizing drugs could be used as single treatment in subsets of patients selected based on suitable biomarker infor-mation on the DNA repair pathway status and extent of replication stress.67,70,81,96,97 Alternatively, they could be combined with standard-of-care genotoxic che-motherapeutics or ionizing radiation to obtain higher sensitization of cancer cells.
Despite the undisputable progress in recent years, a great deal remains to be done to mechanistically understand the cellular responses to replication stress in general, and the role(s) of the DDK in checkpoint responses and DNA repair in particular. Given the numerous emerg-ing fertile avenues of this research, it is clear that this fascinating field will move even more rapidly, and into the forefront of basic and likely also translational bio-medical research.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
The work in the authors’ labora-tories is supported by grants from the Danish Cancer Society, European Commission (projects DDResponse and Biomedreg: CZ.1.05/2.1.00/01.0030), the Grant Agency of the Czech Republic (GA13-17555S), the Internal Grant Agency of the Czech Ministry of Health (grant NT11065-5), the Danish National Research Foundation, the Lundbeck Foundation, the Novo Nordisk Foundation, the Kellner Family Foundation, and the Danish Research Council.
References1. Hartwell LH. Genetic control of the cell divi-
sion cycle in yeast. II. Genes controlling DNA replication and its initiation. J Mol Biol 1971; 59:183-94; PMID:5283752; http://dx.doi.org/10.1016/0022-2836(71)90420-7
2. Masai H, Miyake T, Arai K. hsk1+, a Schizosaccharomyces pombe gene related to Saccharomyces cerevisiae CDC7, is required for chro-mosomal replication. EMBO J 1995; 14:3094-104; PMID:7621824
3. Sato N, Arai K, Masai H. Human and Xenopus cDNAs encoding budding yeast Cdc7-related kinases: in vitro phosphorylation of MCM subunits by a putative human homologue of Cdc7. EMBO J 1997; 16:4340-51; PMID:9250678; http://dx.doi.org/10.1093/emboj/16.14.4340
4. Kim JM, Sato N, Yamada M, Arai K, Masai H. Growth regulation of the expression of mouse cDNA and gene encoding a serine/threonine kinase related to Saccharomyces cerevisiae CDC7 essential for G1/S transition. Structure, chromosomal localiza-tion, and expression of mouse gene for s. cerevisiae Cdc7-related kinase. J Biol Chem 1998; 273:23248-57; PMID:9722556; http://dx.doi.org/10.1074/jbc.273.36.23248
5. Kumagai H, Sato N, Yamada M, Mahony D, Seghezzi W, Lees E, Arai K, Masai H. A novel growth- and cell cycle-regulated protein, ASK, activates human Cdc7-related kinase and is essential for G1/S transition in mammalian cells. Mol Cell Biol 1999; 19:5083-95; PMID:10373557
6. Jiang W, McDonald D, Hope TJ, Hunter T. Mammalian Cdc7-Dbf4 protein kinase complex is essential for initiation of DNA replication. EMBO J 1999; 18:5703-13; PMID:10523313; http://dx.doi.org/10.1093/emboj/18.20.5703
7. Takeda T, Ogino K, Matsui E, Cho MK, Kumagai H, Miyake T, Arai K, Masai H. A fission yeast gene, him1(+)/dfp1(+), encoding a regulatory subunit for Hsk1 kinase, plays essential roles in S-phase initiation as well as in S-phase checkpoint control and recovery from DNA damage. Mol Cell Biol 1999; 19:5535-47; PMID:10409743
8. Brown GW, Kelly TJ. Cell cycle regulation of Dfp1, an activator of the Hsk1 protein kinase. Proc Natl Acad Sci U S A 1999; 96:8443-8; PMID:10411894; http://dx.doi.org/10.1073/pnas.96.15.8443
9. Jackson AL, Pahl PM, Harrison K, Rosamond J, Sclafani RA. Cell cycle regulation of the yeast Cdc7 protein kinase by association with the Dbf4 protein. Mol Cell Biol 1993; 13:2899-908; PMID:8474449
10. Matthews LA, Guarné A. Dbf4: the whole is greater than the sum of its parts. Cell Cycle 2013; 12:1180-8; PMID:23549174; http://dx.doi.org/10.4161/cc.24416
Dow
nloa
ded
by [
Aca
dem
ia S
inic
a -
Tai
wan
] at
01:
13 1
9 D
ecem
ber
2014

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
1864 cell cycle volume 13 issue 12
11. Matsumoto S, Masai H. Regulation of chromosome dynamics by Hsk1/Cdc7 kinase. Biochem Soc Trans 2013; 41:1712-9; PMID:24256280
12. Labib K. How do Cdc7 and cyclin-dependent kinases trigger the initiation of chromosome replica-tion in eukaryotic cells? Genes Dev 2010; 24:1208-19; PMID:20551170; http://dx.doi.org/10.1101/gad.1933010
13. Kim JM, Yamada M, Masai H. Functions of mam-malian Cdc7 kinase in initiation/monitoring of DNA replication and development. Mutat Res 2003; 532:29-40; PMID:14643427; http://dx.doi.org/10.1016/j.mrfmmm.2003.08.008
14. Bartek J, Lukas C, Lukas J. Checking on DNA dam-age in S phase. Nat Rev Mol Cell Biol 2004; 5:792-804; PMID:15459660; http://dx.doi.org/10.1038/nrm1493
15. Zegerman P, Diffley JF. Checkpoint-dependent inhibition of DNA replication initiation by Sld3 and Dbf4 phosphorylation. Nature 2010; 467:474-8; PMID:20835227; http://dx.doi.org/10.1038/nature09373
16. Lopez-Mosqueda J, Maas NL, Jonsson ZO, Defazio-Eli LG, Wohlschlegel J, Toczyski DP. Damage-induced phosphorylation of Sld3 is important to block late origin firing. Nature 2010; 467:479-83; PMID:20865002; http://dx.doi.org/10.1038/nature09377
17. Kihara M, Nakai W, Asano S, Suzuki A, Kitada K, Kawasaki Y, Johnston LH, Sugino A. Characterization of the yeast Cdc7p/Dbf4p complex purified from insect cells. Its protein kinase activity is regulated by Rad53p. J Biol Chem 2000; 275:35051-62; PMID:10964916; http://dx.doi.org/10.1074/jbc.M003491200
18. Weinreich M, Stillman B. Cdc7p-Dbf4p kinase binds to chromatin during S phase and is regulated by both the APC and the RAD53 checkpoint path-way. EMBO J 1999; 18:5334-46; PMID:10508166; http://dx.doi.org/10.1093/emboj/18.19.5334
19. Costanzo V, Shechter D, Lupardus PJ, Cimprich KA, Gottesman M, Gautier J. An ATR- and Cdc7-dependent DNA damage checkpoint that inhibits initiation of DNA replication. Mol Cell 2003; 11:203-13; PMID:12535533; http://dx.doi.org/10.1016/S1097-2765(02)00799-2
20. Tsuji T, Lau E, Chiang GG, Jiang W. The role of Dbf4/Drf1-dependent kinase Cdc7 in DNA-damage checkpoint control. Mol Cell 2008; 32:862-9; PMID:19111665; http://dx.doi.org/10.1016/j.molcel.2008.12.005
21. Dierov J, Dierova R, Carroll M. BCR/ABL translo-cates to the nucleus and disrupts an ATR-dependent intra-S phase checkpoint. Cancer Cell 2004; 5:275-85; PMID:15050919; http://dx.doi.org/10.1016/S1535-6108(04)00056-X
22. Tenca P, Brotherton D, Montagnoli A, Rainoldi S, Albanese C, Santocanale C. Cdc7 is an active kinase in human cancer cells undergoing replication stress. J Biol Chem 2007; 282:208-15; PMID:17062569; http://dx.doi.org/10.1074/jbc.M604457200
23. Lee AY, Chiba T, Truong LN, Cheng AN, Do J, Cho MJ, Chen L, Wu X. Dbf4 is direct downstream target of ataxia telangiectasia mutated (ATM) and ataxia telangiectasia and Rad3-related (ATR) protein to regulate intra-S-phase checkpoint. J Biol Chem 2012; 287:2531-43; PMID:22123827; http://dx.doi.org/10.1074/jbc.M111.291104
24. Kim JM, Kakusho N, Yamada M, Kanoh Y, Takemoto N, Masai H. Cdc7 kinase mediates Claspin phosphorylation in DNA replication checkpoint. Oncogene 2008; 27:3475-82; PMID:18084324; http://dx.doi.org/10.1038/sj.onc.1210994
25. Rainey MD, Harhen B, Wang GN, Murphy PV, Santocanale C. Cdc7-dependent and -indepen-dent phosphorylation of Claspin in the induction of the DNA replication checkpoint. Cell Cycle 2013; 12:1560-8; PMID:23598722; http://dx.doi.org/10.4161/cc.24675
26. Yamada M, Sato N, Taniyama C, Ohtani K, Arai K, Masai H. A 63-base pair DNA segment containing an Sp1 site but not a canonical E2F site can confer growth-dependent and E2F-mediated transcriptional stimulation of the human ASK gene encoding the regulatory subunit for human Cdc7-related kinase. J Biol Chem 2002; 277:27668-81; PMID:12015319; http://dx.doi.org/10.1074/jbc.M202884200
27. Wu X, Lee H. Human Dbf4/ASK promoter is acti-vated through the Sp1 and MluI cell-cycle box (MCB) transcription elements. Oncogene 2002; 21:7786-96; PMID:12420215; http://dx.doi.org/10.1038/sj.onc.1205914
28. Natoni A, Coyne MR, Jacobsen A, Rainey MD, O’Brien G, Healy S, Montagnoli A, Moll J, O’Dwyer M, Santocanale C. Characterization of a Dual CDC7/CDK9 Inhibitor in Multiple Myeloma Cellular Models. Cancers (Basel) 2013; 5:901-18; PMID:24202326; http://dx.doi.org/10.3390/cancers5030901
29. Woods KW, Lai C, Miyashiro JM, Tong Y, Florjancic AS, Han EK, Soni N, Shi Y, Lasko L, Leverson JD, et al. Aminopyrimidinone cdc7 kinase inhibi-tors. Bioorg Med Chem Lett 2012; 22:1940-3; PMID:22326396; http://dx.doi.org/10.1016/j.bmcl.2012.01.041
30. Koltun ES, Tsuhako AL, Brown DS, Aay N, Arcalas A, Chan V, Du H, Engst S, Ferguson K, Franzini M, et al. Discovery of XL413, a potent and selec-tive CDC7 inhibitor. Bioorg Med Chem Lett 2012; 22:3727-31; PMID:22560567; http://dx.doi.org/10.1016/j.bmcl.2012.04.024
31. Natoni A, Murillo LS, Kliszczak AE, Catherwood MA, Montagnoli A, Samali A, O’Dwyer M, Santocanale C. Mechanisms of action of a dual Cdc7/Cdk9 kinase inhibitor against quiescent and prolifer-ating CLL cells. Mol Cancer Ther 2011; 10:1624-34; PMID:21768328; http://dx.doi.org/10.1158/1535-7163.MCT-10-1119
32. Swords R, Mahalingam D, O’Dwyer M, Santocanale C, Kelly K, Carew J, Giles F. Cdc7 kinase - a new tar-get for drug development. Eur J Cancer 2010; 46:33-40; PMID:19815406; http://dx.doi.org/10.1016/j.ejca.2009.09.020
33. Sawa M, Masai H. Drug design with Cdc7 kinase: a potential novel cancer therapy target. Drug Des Devel Ther 2009; 2:255-64; PMID:19920912
34. Vanotti E, Amici R, Bargiotti A, Berthelsen J, Bosotti R, Ciavolella A, Cirla A, Cristiani C, D’Alessio R, Forte B, et al. Cdc7 kinase inhibitors: pyrrolopyr-idinones as potential antitumor agents. 1. Synthesis and structure-activity relationships. J Med Chem 2008; 51:487-501; PMID:18201066; http://dx.doi.org/10.1021/jm700956r
35. Montagnoli A, Valsasina B, Croci V, Menichincheri M, Rainoldi S, Marchesi V, Tibolla M, Tenca P, Brotherton D, Albanese C, et al. A Cdc7 kinase inhibitor restricts initiation of DNA replication and has antitumor activity. Nat Chem Biol 2008; 4:357-65; PMID:18469809; http://dx.doi.org/10.1038/nchembio.90
36. Yamada M, Watanabe K, Mistrik M, Vesela E, Protivankova I, Mailand N, Lee M, Masai H, Lukas J, Bartek J. ATR-Chk1-APC/CCdh1-dependent sta-bilization of Cdc7-ASK (Dbf4) kinase is required for DNA lesion bypass under replication stress. Genes Dev 2013; 27:2459-72; PMID:24240236; http://dx.doi.org/10.1101/gad.224568.113
37. Zachariae W, Schwab M, Nasmyth K, Seufert W. Control of cyclin ubiquitination by CDK-regulated binding of Hct1 to the anaphase promoting complex. Science 1998; 282:1721-4; PMID:9831566; http://dx.doi.org/10.1126/science.282.5394.1721
38. Lukas C, Sørensen CS, Kramer E, Santoni-Rugiu E, Lindeneg C, Peters JM, Bartek J, Lukas J. Accumulation of cyclin B1 requires E2F and cyclin-A-dependent rearrangement of the anaphase-promoting complex. Nature 1999; 401:815-8; PMID:10548110; http://dx.doi.org/10.1038/44611
39. Listovsky T, Zor A, Laronne A, Brandeis M. Cdk1 is essential for mammalian cyclosome/APC regulation. Exp Cell Res 2000; 255:184-91; PMID:10694434; http://dx.doi.org/10.1006/excr.1999.4788
40. Sørensen CS, Lukas C, Kramer ER, Peters JM, Bartek J, Lukas J. A conserved cyclin-binding domain determines functional interplay between anaphase-promoting complex-Cdh1 and cyclin A-Cdk2 during cell cycle progression. Mol Cell Biol 2001; 21:3692-703; PMID:11340163; http://dx.doi.org/10.1128/MCB.21.11.3692-3703.2001
41. Sudo T, Ota Y, Kotani S, Nakao M, Takami Y, Takeda S, Saya H. Activation of Cdh1-dependent APC is required for G1 cell cycle arrest and DNA damage-induced G2 checkpoint in vertebrate cells. EMBO J 2001; 20:6499-508; PMID:11707420; http://dx.doi.org/10.1093/emboj/20.22.6499
42. Bassermann F, Frescas D, Guardavaccaro D, Busino L, Peschiaroli A, Pagano M. The Cdc14B-Cdh1-Plk1 axis controls the G2 DNA-damage-response check-point. Cell 2008; 134:256-67; PMID:18662541; http://dx.doi.org/10.1016/j.cell.2008.05.043
43. Adams MM, Carpenter PB. Tying the loose ends together in DNA double strand break repair with 53BP1. Cell Div 2006; 1:19; PMID:16945145; http://dx.doi.org/10.1186/1747-1028-1-19
44. Cho HJ, Lee EH, Han SH, Chung HJ, Jeong JH, Kwon J, Kim H. Degradation of human RAP80 is cell cycle regulated by Cdc20 and Cdh1 ubiquitin ligases. Mol Cancer Res 2012; 10:615-25; PMID:22426463; http://dx.doi.org/10.1158/1541-7786.MCR-11-0481
45. Hu Y, Scully R, Sobhian B, Xie A, Shestakova E, Livingston DM. RAP80-directed tuning of BRCA1 homologous recombination function at ionizing radi-ation-induced nuclear foci. Genes Dev 2011; 25:685-700; PMID:21406551; http://dx.doi.org/10.1101/gad.2011011
46. Coleman KA, Greenberg RA. The BRCA1-RAP80 complex regulates DNA repair mechanism utiliza-tion by restricting end resection. J Biol Chem 2011; 286:13669-80; PMID:21335604; http://dx.doi.org/10.1074/jbc.M110.213728
47. Njagi GD, Kilbey BJ. cdc7-1 a temperature sensi-tive cell-cycle mutant which interferes with induced mutagenesis in Saccharomyces cerevisiae. Mol Gen Genet 1982; 186:478-81; PMID:6752657; http://dx.doi.org/10.1007/BF00337951
48. Pessoa-Brandão L, Sclafani RA. CDC7/DBF4 func-tions in the translesion synthesis branch of the RAD6 epistasis group in Saccharomyces cerevisiae. Genetics 2004; 167:1597-610; PMID:15342501; http://dx.doi.org/10.1534/genetics.103.021675
49. Day TA, Palle K, Barkley LR, Kakusho N, Zou Y, Tateishi S, Verreault A, Masai H, Vaziri C. Phosphorylated Rad18 directs DNA polymerase η to sites of stalled replication. J Cell Biol 2010; 191:953-66; PMID:21098111; http://dx.doi.org/10.1083/jcb.201006043
50. Vaziri C, Masai H. Integrating DNA replication with trans-lesion synthesis via Cdc7. Cell Cycle 2010; 9:4818-23; PMID:21150323; http://dx.doi.org/10.4161/cc.9.24.14241
51. Durando M, Tateishi S, Vaziri C. A non-catalytic role of DNA polymerase η in recruiting Rad18 and promoting PCNA monoubiquitination at stalled rep-lication forks. Nucleic Acids Res 2013; 41:3079-93; PMID:23345618; http://dx.doi.org/10.1093/nar/gkt016
52. Huang J, Huen MS, Kim H, Leung CC, Glover JN, Yu X, Chen J. RAD18 transmits DNA damage sig-nalling to elicit homologous recombination repair. Nat Cell Biol 2009; 11:592-603; PMID:19396164; http://dx.doi.org/10.1038/ncb1865
53. Hochegger H, Sonoda E, Takeda S. Post-replication repair in DT40 cells: translesion polymerases versus recombinases. Bioessays 2004; 26:151-8; PMID:14745833; http://dx.doi.org/10.1002/bies.10403
Dow
nloa
ded
by [
Aca
dem
ia S
inic
a -
Tai
wan
] at
01:
13 1
9 D
ecem
ber
2014

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
www.landesbioscience.com cell cycle 1865
54. Williams SA, Longerich S, Sung P, Vaziri C, Kupfer GM. The E3 ubiquitin ligase RAD18 regulates ubiq-uitylation and chromatin loading of FANCD2 and FANCI. Blood 2011; 117:5078-87; PMID:21355096; http://dx.doi.org/10.1182/blood-2010-10-311761
55. Sommariva E, Pellny TK, Karahan N, Kumar S, Huberman JA, Dalgaard JZ. Schizosaccharomyces pombe Swi1, Swi3, and Hsk1 are components of a novel S-phase response pathway to alkyla-tion damage. Mol Cell Biol 2005; 25:2770-84; PMID:15767681; http://dx.doi.org/10.1128/MCB.25.7.2770-2784.2005
56. Matsumoto S, Ogino K, Noguchi E, Russell P, Masai H. Hsk1-Dfp1/Him1, the Cdc7-Dbf4 kinase in Schizosaccharomyces pombe, associates with Swi1, a component of the replication fork protection complex. J Biol Chem 2005; 280:42536-42; PMID:16263721; http://dx.doi.org/10.1074/jbc.M510575200
57. Kim JM, Nakao K, Nakamura K, Saito I, Katsuki M, Arai K, Masai H. Inactivation of Cdc7 kinase in mouse ES cells results in S-phase arrest and p53-dependent cell death. EMBO J 2002; 21:2168-79; PMID:11980714; http://dx.doi.org/10.1093/emboj/21.9.2168
58. Moldovan GL, Dejsuphong D, Petalcorin MI, Hofmann K, Takeda S, Boulton SJ, D’Andrea AD. Inhibition of homologous recombination by the PCNA-interacting protein PARI. Mol Cell 2012; 45:75-86; PMID:22153967; http://dx.doi.org/10.1016/j.molcel.2011.11.010
59. Fugger K, Mistrik M, Danielsen JR, Dinant C, Falck J, Bartek J, Lukas J, Mailand N. Human Fbh1 helicase contributes to genome maintenance via pro- and anti-recombinase activities. J Cell Biol 2009; 186:655-63; PMID:19736316; http://dx.doi.org/10.1083/jcb.200812138
60. Barber LJ, Youds JL, Ward JD, McIlwraith MJ, O’Neil NJ, Petalcorin MI, Martin JS, Collis SJ, Cantor SB, Auclair M, et al. RTEL1 maintains genomic stability by suppressing homologous recom-bination. Cell 2008; 135:261-71; PMID:18957201; http://dx.doi.org/10.1016/j.cell.2008.08.016
61. Poh WT, Chadha GS, Gillespie PJ, Kaldis P, Blow JJ. Xenopus Cdc7 executes its essential function early in S phase and is counteracted by checkpoint-regulated protein phosphatase 1. Open Biol 2014; 4:130138; PMID:24403013; http://dx.doi.org/10.1098/rsob.130138
62. Hayano M, Kanoh Y, Matsumoto S, Renard-Guillet C, Shirahige K, Masai H. Rif1 is a global regulator of timing of replication origin firing in fission yeast. Genes Dev 2012; 26:137-50; PMID:22279046; http://dx.doi.org/10.1101/gad.178491.111
63. Yamazaki S, Ishii A, Kanoh Y, Oda M, Nishito Y, Masai H. Rif1 regulates the replication timing domains on the human genome. EMBO J 2012; 31:3667-77; PMID:22850674; http://dx.doi.org/10.1038/emboj.2012.180
64. Cornacchia D, Dileep V, Quivy JP, Foti R, Tili F, Santarella-Mellwig R, Antony C, Almouzni G, Gilbert DM, Buonomo SB. Mouse Rif1 is a key regulator of the replication-timing programme in mammalian cells. EMBO J 2012; 31:3678-90; PMID:22850673; http://dx.doi.org/10.1038/emboj.2012.214
65. Yamazaki S, Hayano M, Masai H. Replication tim-ing regulation of eukaryotic replicons: Rif1 as a global regulator of replication timing. Trends Genet 2013; 29:449-60; PMID:23809990; http://dx.doi.org/10.1016/j.tig.2013.05.001
66. Hiraga S, Alvino GM, Chang F, Lian HY, Sridhar A, Kubota T, Brewer BJ, Weinreich M, Raghuraman MK, Donaldson AD. Rif1 controls DNA replication by directing Protein Phosphatase 1 to reverse Cdc7-mediated phosphorylation of the MCM complex. Genes Dev 2014; 28:372-83; PMID:24532715; http://dx.doi.org/10.1101/gad.231258.113
67. Bartkova J, Horejsí Z, Koed K, Krämer A, Tort F, Zieger K, Guldberg P, Sehested M, Nesland JM, Lukas C, et al. DNA damage response as a candidate anti-cancer barrier in early human tumorigenesis. Nature 2005; 434:864-70; PMID:15829956; http://dx.doi.org/10.1038/nature03482
68. Gorgoulis VG, Vassiliou LV, Karakaidos P, Zacharatos P, Kotsinas A, Liloglou T, Venere M, Ditullio RA Jr., Kastrinakis NG, Levy B, et al. Activation of the DNA damage checkpoint and genomic instability in human precancerous lesions. Nature 2005; 434:907-13; PMID:15829965; http://dx.doi.org/10.1038/nature03485
69. Bartkova J, Bakkenist CJ, Rajpert-De Meyts E, Skakkebaek NE, Sehested M, Lukas J, Kastan MB, Bartek J. ATM activation in normal human tis-sues and testicular cancer. Cell Cycle 2005; 4:838-45; PMID:15846060; http://dx.doi.org/10.4161/cc.4.6.1742
70. Bartkova J, Rezaei N, Liontos M, Karakaidos P, Kletsas D, Issaeva N, Vassiliou LV, Kolettas E, Niforou K, Zoumpourlis VC, et al. Oncogene-induced senescence is part of the tumorigenesis bar-rier imposed by DNA damage checkpoints. Nature 2006; 444:633-7; PMID:17136093; http://dx.doi.org/10.1038/nature05268
71. Di Micco R, Fumagalli M, Cicalese A, Piccinin S, Gasparini P, Luise C, Schurra C, Garre’ M, Nuciforo PG, Bensimon A, et al. Oncogene-induced senescence is a DNA damage response triggered by DNA hyper-replication. Nature 2006; 444:638-42; PMID:17136094; http://dx.doi.org/10.1038/nature05327
72. Bartek J, Lukas J, Bartkova J. DNA damage response as an anti-cancer barrier: damage threshold and the concept of ‘conditional haploinsufficiency’. Cell Cycle 2007; 6:2344-7; PMID:17700066; http://dx.doi.org/10.4161/cc.6.19.4754
73. Halazonetis TD, Gorgoulis VG, Bartek J. An onco-gene-induced DNA damage model for cancer develop-ment. Science 2008; 319:1352-5; PMID:18323444; http://dx.doi.org/10.1126/science.1140735
74. Takacova S, Slany R, Bartkova J, Stranecky V, Dolezel P, Luzna P, Bartek J, Divoky V. DNA damage response and inflammatory signaling limit the MLL-ENL-induced leukemogenesis in vivo. Cancer Cell 2012; 21:517-31; PMID:22516260; http://dx.doi.org/10.1016/j.ccr.2012.01.021
75. Burrell RA, McClelland SE, Endesfelder D, Groth P, Weller MC, Shaikh N, Domingo E, Kanu N, Dewhurst SM, Gronroos E, et al. Replication stress links structural and numerical cancer chro-mosomal instability. Nature 2013; 494:492-6; PMID:23446422; http://dx.doi.org/10.1038/nature11935
76. Evangelou K, Bartkova J, Kotsinas A, Pateras IS, Liontos M, Velimezi G, Kosar M, Liloglou T, Trougakos IP, Dyrskjot L, et al. The DNA damage checkpoint precedes activation of ARF in response to escalating oncogenic stress during tumorigenesis. Cell Death Differ 2013; 20:1485-97; PMID:23852374; http://dx.doi.org/10.1038/cdd.2013.76
77. Velimezi G, Liontos M, Vougas K, Roumeliotis T, Bartkova J, Sideridou M, Dereli-Oz A, Kocylowski M, Pateras IS, Evangelou K, et al. Functional interplay between the DNA-damage-response kinase ATM and ARF tumour suppressor protein in human cancer. Nat Cell Biol 2013; 15:967-77; PMID:23851489; http://dx.doi.org/10.1038/ncb2795
78. Tavana O, Chen D, Gu W. Controlling ARF sta-bility: new players added to the team. Cell Cycle 2014; 13:497-8; PMID:24419106; http://dx.doi.org/10.4161/cc.27786
79. Kotsinas A, Papanagnou P, Galanos P, Schramek D, Townsend P, Penninger JM, Bartek J, Gorgoulis VG. MKK7 and ARF: New players in the DNA damage response scenery. Cell Cycle 2014; 13:1227-36; PMID:24675893; http://dx.doi.org/10.4161/cc.28654
80. Monasor A, Murga M, Lopez-Contreras AJ, Navas C, Gomez G, Pisano DG, Fernandez-Capetillo O. INK4a/ARF limits the expansion of cells suffering from replication stress. Cell Cycle 2013; 12:1948-54; PMID:23676215; http://dx.doi.org/10.4161/cc.25017
81. Bartek J, Mistrik M, Bartkova J. Thresholds of replication stress signaling in cancer development and treatment. Nat Struct Mol Biol 2012; 19:5-7; PMID:22218289; http://dx.doi.org/10.1038/nsmb.2220
82. Farmer H, McCabe N, Lord CJ, Tutt AN, Johnson DA, Richardson TB, Santarosa M, Dillon KJ, Hickson I, Knights C, et al. Targeting the DNA repair defect in BRCA mutant cells as a therapeutic strategy. Nature 2005; 434:917-21; PMID:15829967; http://dx.doi.org/10.1038/nature03445
83. Bryant HE, Schultz N, Thomas HD, Parker KM, Flower D, Lopez E, Kyle S, Meuth M, Curtin NJ, Helleday T. Specific killing of BRCA2-deficient tumours with inhibitors of poly(ADP-ribose) poly-merase. Nature 2005; 434:913-7; PMID:15829966; http://dx.doi.org/10.1038/nature03443
84. Jackson SP, Bartek J. The DNA-damage response in human biology and disease. Nature 2009; 461:1071-8; PMID:19847258; http://dx.doi.org/10.1038/nature08467
85. Dedes KJ, Wilkerson PM, Wetterskog D, Weigelt B, Ashworth A, Reis-Filho JS. Synthetic lethal-ity of PARP inhibition in cancers lacking BRCA1 and BRCA2 mutations. Cell Cycle 2011; 10:1192-9; PMID:21487248; http://dx.doi.org/10.4161/cc.10.8.15273
86. Hishida T, Kubota Y, Carr AM, Iwasaki H. RAD6-RAD18-RAD5-pathway-dependent toler-ance to chronic low-dose ultraviolet light. Nature 2009; 457:612-5; PMID:19079240; http://dx.doi.org/10.1038/nature07580
87. Hishida T, Hirade Y, Haruta N, Kubota Y, Iwasaki H. Srs2 plays a critical role in reversible G2 arrest upon chronic and low doses of UV irradiation via two distinct homologous recombination-dependent mechanisms in postreplication repair-deficient cells. Mol Cell Biol 2010; 30:4840-50; PMID:20713444; http://dx.doi.org/10.1128/MCB.00453-10
88. Yamashita YM, Okada T, Matsusaka T, Sonoda E, Zhao GY, Araki K, Tateishi S, Yamaizumi M, Takeda S. RAD18 and RAD54 cooperatively contribute to maintenance of genomic stability in vertebrate cells. EMBO J 2002; 21:5558-66; PMID:12374756; http://dx.doi.org/10.1093/emboj/cdf534
89. Hicks JK, Chute CL, Paulsen MT, Ragland RL, Howlett NG, Guéranger Q, Glover TW, Canman CE. Differential roles for DNA polymerases eta, zeta, and REV1 in lesion bypass of intrastrand ver-sus interstrand DNA cross-links. Mol Cell Biol 2010; 30:1217-30; PMID:20028736; http://dx.doi.org/10.1128/MCB.00993-09
90. Yamamoto K, Ishiai M, Matsushita N, Arakawa H, Lamerdin JE, Buerstedde JM, Tanimoto M, Harada M, Thompson LH, Takata M. Fanconi anemia FANCG protein in mitigating radiation- and enzyme-induced DNA double-strand breaks by homologous recombination in vertebrate cells. Mol Cell Biol 2003; 23:5421-30; PMID:12861027; http://dx.doi.org/10.1128/MCB.23.15.5421-5430.2003
Dow
nloa
ded
by [
Aca
dem
ia S
inic
a -
Tai
wan
] at
01:
13 1
9 D
ecem
ber
2014

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
1866 cell cycle volume 13 issue 12
91. Yamamoto K, Hirano S, Ishiai M, Morishima K, Kitao H, Namikoshi K, Kimura M, Matsushita N, Arakawa H, Buerstedde JM, et al. Fanconi anemia protein FANCD2 promotes immunoglobulin gene conversion and DNA repair through a mechanism related to homologous recombination. Mol Cell Biol 2005; 25:34-43; PMID:15601828; http://dx.doi.org/10.1128/MCB.25.1.34-43.2005
92. Nakanishi K, Yang YG, Pierce AJ, Taniguchi T, Digweed M, D’Andrea AD, Wang ZQ, Jasin M. Human Fanconi anemia monoubiquitination path-way promotes homologous DNA repair. Proc Natl Acad Sci U S A 2005; 102:1110-5; PMID:15650050; http://dx.doi.org/10.1073/pnas.0407796102
93. Niedzwiedz W, Mosedale G, Johnson M, Ong CY, Pace P, Patel KJ. The Fanconi anaemia gene FANCC promotes homologous recombination and error-prone DNA repair. Mol Cell 2004; 15:607-20; PMID:15327776; http://dx.doi.org/10.1016/j.molcel.2004.08.009
94. Hirano S, Yamamoto K, Ishiai M, Yamazoe M, Seki M, Matsushita N, Ohzeki M, Yamashita YM, Arakawa H, Buerstedde JM, et al. Functional rela-tionships of FANCC to homologous recombina-tion, translesion synthesis, and BLM. EMBO J 2005; 24:418-27; PMID:15616572; http://dx.doi.org/10.1038/sj.emboj.7600534
95. Nakanishi K, Cavallo F, Perrouault L, Giovannangeli C, Moynahan ME, Barchi M, Brunet E, Jasin M. Homology-directed Fanconi anemia pathway cross-link repair is dependent on DNA replication. Nat Struct Mol Biol 2011; 18:500-3; PMID:21423196; http://dx.doi.org/10.1038/nsmb.2029
96. Oplustilova L, Wolanin K, Mistrik M, Korinkova G, Simkova D, Bouchal J, Lenobel R, Bartkova J, Lau A, O’Connor MJ, et al. Evaluation of candi-date biomarkers to predict cancer cell sensitivity or resistance to PARP-1 inhibitor treatment. Cell Cycle 2012; 11:3837-50; PMID:22983061; http://dx.doi.org/10.4161/cc.22026
97. Fagerholm R, Sprott K, Heikkinen T, Bartkova J, Heikkilä P, Aittomäki K, Bartek J, Weaver D, Blomqvist C, Nevanlinna H. Overabundant FANCD2, alone and combined with NQO1, is a sensitive marker of adverse prognosis in breast can-cer. Ann Oncol 2013; 24:2780-5; PMID:23897704; http://dx.doi.org/10.1093/annonc/mdt290
Dow
nloa
ded
by [
Aca
dem
ia S
inic
a -
Tai
wan
] at
01:
13 1
9 D
ecem
ber
2014
Related Documents



![Human Mitogen-activated Protein Kinase Kinase 4 as a ......(CANCERRESEARCH57. 4177—4182,October 1, 1997] Advances in Brief Human Mitogen-activated Protein Kinase Kinase 4 as](https://static.cupdf.com/doc/110x72/6082557b7810d746a5071f39/human-mitogen-activated-protein-kinase-kinase-4-as-a-cancerresearch57.jpg)







