Clathrin, AP-2, and the NPXY-binding subset of alternate endocytic adaptors facilitate FimH-mediated bacterial invasion of host cells Danelle S. Eto, Hannah B. Gordon, Bijaya K. Dhakal, Tiffani A. Jones and Matthew A. Mulvey* Division of Cell Biology and Immunology, Pathology Department, University of Utah, Salt Lake City, UT 84112-0565, USA. Summary The FimH adhesin, localized at the distal tips of type 1 pili, binds mannose-containing glycoprotein recep- tors like a3b1 integrins and stimulates bacterial entry into target host cells. Strains of uropathogenic Escherichia coli (UPEC), the major cause of urinary tract infections, utilize FimH to invade bladder epithe- lial cells. Here we set out to define the mechanism by which UPEC enters host cells by investigating four of the major entry routes known to be exploited by inva- sive pathogens: caveolae, clathrin, macropinocytosis and secretory lysosomes. Using pharmacological inhibitors in combination with RNA interference against specific endocytic pathway components, mutant host cell lines and a mouse infection model system, we found that type 1 pili-dependent bacterial invasion of host cells occurs via a cholesterol- and dynamin-dependent phagocytosis-like mechanism. This process did not require caveolae or secretory lysosomes, but was modulated by calcium levels, clathrin, and cooperative input from the primary clath- rin adaptor AP-2 and a subset of alternate adaptors comprised of Numb, ARH and Dab2. These alternate clathrin adaptors recognize NPXY motifs, as found within the cytosolic tail of b1 integrin, suggesting a functional link between the engagement of integrin receptors by FimH and the clathrin-dependent uptake of type 1-piliated bacteria. Introduction Strains of uropathogenic Escherichia coli (UPEC), the primary causative agent of urinary tract infections (UTIs), typically express filamentous surface adhesive organelles known as type 1 pili. At the distal tip of each type 1 pilus is an adhesin protein known as FimH that mediates attachment to and invasion of host bladder epithelial cells (Martinez et al., 2000). The FimH adhesin contains a carbohydrate-binding pocket that recognizes mannose and mannose-containing glycoproteins (Hung et al., 2002; Mulvey, 2002). Mouse UTI and cell culture model systems suggest that FimH-mediated bacterial invasion of host epithelial cells lining the urinary tract significantly contributes to the establishment, persistence and possibly the recurrence of UTIs (reviewed in Dhakal et al., 2008). Contact between type 1 pili and host cell receptors, such as a3b1 integrin, triggers localized actin rearrange- ments leading to the envelopment and internalization of bound bacteria (Mulvey et al., 1998; Martinez et al., 2000; Eto et al., 2007). This invasion process, in which the host plasma membrane zippers around adherent bacteria via contacts with the FimH-containing tips of type 1 pili, involves activation of a number of host-signalling mole- cules, including focal adhesion kinase, Src kinase, phos- phatidylinositide 3-kinase and Rho family GTPases like Rac-1 (Mulvey et al., 1998; Martinez et al., 2000; Martinez and Hultgren, 2002; Eto et al., 2007; Song et al., 2007; Wang et al., 2008). In addition, FimH interactions with host cells induce transient complex formation between a-actinin and vinculin, two proteins involved in the stabi- lization of actin filaments within focal adhesions (Martinez et al., 2000). These various host factors likely act in concert to stimulate and modulate actin and membrane dynamics at sites of bacterial internalization. Despite recent progress in identifying host factors associated with FimH-mediated UPEC invasion of host bladder epithelial cells, the specific entry pathway(s) used by type 1-piliated UPEC remains ambiguous. A number of intracellular microbial pathogens are known to usurp host endocytic pathways, facilitating their entry into non-professional phagocytes via caveolae- and clathrin-mediated endocytosis, macropinocytosis or the subversion of the secretory lysosome pathway (Gruen- berg and van der Goot, 2006). A major route of endocy- tosis in most eukaryotic cells is mediated by clathrin. Specific cargos are recognized and concentrated in clathrin-coated pits by different combinations of clathrin adaptor and accessory proteins. The canonical AP-2 Received 14 March, 2008; revised 14 August, 2008; accepted 16 August, 2008. *For correspondence. E-mail [email protected]; Tel. (+1) 801 581 5967; Fax (+1) 801 581 4517. Cellular Microbiology (2008) 10(12), 2553–2567 doi:10.1111/j.1462-5822.2008.01229.x First published online 10 September 2008 © 2008 The Authors Journal compilation © 2008 Blackwell Publishing Ltd

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Clathrin, AP-2, and the NPXY-binding subset ofalternate endocytic adaptors facilitate FimH-mediatedbacterial invasion of host cells
Danelle S. Eto, Hannah B. Gordon, Bijaya K. Dhakal,Tiffani A. Jones and Matthew A. Mulvey*Division of Cell Biology and Immunology, PathologyDepartment, University of Utah, Salt Lake City,UT 84112-0565, USA.
Summary
The FimH adhesin, localized at the distal tips of type 1pili, binds mannose-containing glycoprotein recep-tors like a3b1 integrins and stimulates bacterial entryinto target host cells. Strains of uropathogenicEscherichia coli (UPEC), the major cause of urinarytract infections, utilize FimH to invade bladder epithe-lial cells. Here we set out to define the mechanism bywhich UPEC enters host cells by investigating four ofthe major entry routes known to be exploited by inva-sive pathogens: caveolae, clathrin, macropinocytosisand secretory lysosomes. Using pharmacologicalinhibitors in combination with RNA interferenceagainst specific endocytic pathway components,mutant host cell lines and a mouse infection modelsystem, we found that type 1 pili-dependent bacterialinvasion of host cells occurs via a cholesterol- anddynamin-dependent phagocytosis-like mechanism.This process did not require caveolae or secretorylysosomes, but was modulated by calcium levels,clathrin, and cooperative input from the primary clath-rin adaptor AP-2 and a subset of alternate adaptorscomprised of Numb, ARH and Dab2. These alternateclathrin adaptors recognize NPXY motifs, as foundwithin the cytosolic tail of b1 integrin, suggesting afunctional link between the engagement of integrinreceptors by FimH and the clathrin-dependent uptakeof type 1-piliated bacteria.
Introduction
Strains of uropathogenic Escherichia coli (UPEC), theprimary causative agent of urinary tract infections (UTIs),typically express filamentous surface adhesive organelles
known as type 1 pili. At the distal tip of each type 1 pilusis an adhesin protein known as FimH that mediatesattachment to and invasion of host bladder epithelial cells(Martinez et al., 2000). The FimH adhesin contains acarbohydrate-binding pocket that recognizes mannoseand mannose-containing glycoproteins (Hung et al.,2002; Mulvey, 2002). Mouse UTI and cell culture modelsystems suggest that FimH-mediated bacterial invasion ofhost epithelial cells lining the urinary tract significantlycontributes to the establishment, persistence and possiblythe recurrence of UTIs (reviewed in Dhakal et al., 2008).
Contact between type 1 pili and host cell receptors,such as a3b1 integrin, triggers localized actin rearrange-ments leading to the envelopment and internalization ofbound bacteria (Mulvey et al., 1998; Martinez et al., 2000;Eto et al., 2007). This invasion process, in which the hostplasma membrane zippers around adherent bacteria viacontacts with the FimH-containing tips of type 1 pili,involves activation of a number of host-signalling mole-cules, including focal adhesion kinase, Src kinase, phos-phatidylinositide 3-kinase and Rho family GTPases likeRac-1 (Mulvey et al., 1998; Martinez et al., 2000; Martinezand Hultgren, 2002; Eto et al., 2007; Song et al., 2007;Wang et al., 2008). In addition, FimH interactions withhost cells induce transient complex formation betweena-actinin and vinculin, two proteins involved in the stabi-lization of actin filaments within focal adhesions (Martinezet al., 2000). These various host factors likely act inconcert to stimulate and modulate actin and membranedynamics at sites of bacterial internalization. Despiterecent progress in identifying host factors associated withFimH-mediated UPEC invasion of host bladder epithelialcells, the specific entry pathway(s) used by type 1-piliatedUPEC remains ambiguous.
A number of intracellular microbial pathogens areknown to usurp host endocytic pathways, facilitating theirentry into non-professional phagocytes via caveolae- andclathrin-mediated endocytosis, macropinocytosis or thesubversion of the secretory lysosome pathway (Gruen-berg and van der Goot, 2006). A major route of endocy-tosis in most eukaryotic cells is mediated by clathrin.Specific cargos are recognized and concentrated inclathrin-coated pits by different combinations of clathrinadaptor and accessory proteins. The canonical AP-2
Received 14 March, 2008; revised 14 August, 2008; accepted 16August, 2008. *For correspondence. E-mail [email protected];Tel. (+1) 801 581 5967; Fax (+1) 801 581 4517.
Cellular Microbiology (2008) 10(12), 2553–2567 doi:10.1111/j.1462-5822.2008.01229.xFirst published online 10 September 2008
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd

adaptor protein complex is a major regulator of clathrin-mediated internalization at the plasma membrane (Traub,2003). However, several other alternate adaptors havebeen identified that perform similar cargo sorting andclathrin assembly functions at the cell surface, actingeither independently or in association with AP-2 (Motleyet al., 2003; Traub, 2003). To date, host cell invasion byonly a few bacterial pathogens is known to require clathrin(Veiga et al., 2007). In contrast, caveolae are reported tofacilitate the uptake of several pathogens and bacterialtoxins (Duncan et al., 2002). Caveolae denote a subsetof plasma membrane microdomains known as lipid raftsthat are enriched in cholesterol and glycosphingolipids(Anderson, 1998). The presence of caveolin-1 (cav1) isthe primary distinguishing feature of caveolae, whichoften form flask-shaped invaginations at the cell surface.Previous studies have shown that cav1 is necessary forcaveolae formation and function (Drab et al., 2001;Razani et al., 2001).
Many endocytic pathways in mammalian cells, includ-ing those involving caveolae, clathrin-coated pits andphagocytosis, require dynamin (Conner and Schmid,2003). This large GTPase pinches and severs nascentendocytic vesicles from the plasma membrane. Dynaminalso interacts with a number of actin-binding proteins,possibly functioning in a regulatory role linking actindynamics with receptor-mediated endocytic pathways.One major endocytic pathway that does not depend ondynamin is macropinocytosis (Conner and Schmid, 2003;Miaczynska and Stenmark, 2008). Membrane rufflingcoincident with sizeable actin cytoskeletal changes ischaracteristic of macropinocytosis and results in the indis-criminateuptakeofmoleculesandsolutes into largehetero-geneous vesicles (Swanson, 2008). Several invasivebacterial pathogens are known to gain entry into host cellsby injecting effector molecules that trigger bacterialuptake by a macropinocytosis-like mechanism (Cossartand Sansonetti, 2004; Swanson, 2008).
Currently, the parasite Trypanosoma cruzi is the onlypathogen definitively shown to utilize secretory lysosomesto invade host cells. Secretory lysosomes, representing asubset of the classic lysosomal organelles that function inthe degradation of macromolecules, undergo calcium-regulated exocytosis and participate in the repair ofplasma membrane damage (Reddy et al., 2001; Blott andGriffiths, 2002). T. cruzi can hijack this system by inducingintracellular calcium fluxes that signal secretory lyso-somes to fuse with the plasma membrane at sites ofparasite attachment (Tardieux et al., 1992; Gruenbergand van der Goot, 2006). Lysosome exocytosis and fusionare thought to provide membrane for the generation of theso-called parasitophorous vacuole, which forms aroundthe invading parasites, likely promoting subsequent intra-cellular replication of T. cruzi (Andrade et al., 2004).
Recently, it was proposed that UPEC entry into bladderepithelial cells occurs via secretory lysosomes (Bishopet al., 2007), while other earlier work suggested thatUPEC entry is lipid raft-dependent and mediated throughcaveolae (Duncan et al., 2004). These reports raised thepossibility that type 1-piliated UPEC may invade hostbladder cells via multiple pathways. To address this issue,we investigated four major entry routes that are exploitedby invasive pathogens, considering the roles of bothcaveolae and secretory lysosomes in the invasionprocess along with macropinocytosis and clathrin. Ourresults indicate that FimH-mediated bacterial invasion ofhost cells can proceed independent of both caveolae andsecretory lysosomes via a phagocytosis-like mechanismthat is modulated by cholesterol, calcium, the hostGTPase dynamin, clathrin, AP-2, and at least three alter-nate clathrin adaptor proteins.
Results
Multiple endocytic pathway inhibitors decrease type 1pili-mediated bacterial invasion
To broadly assess the invasion mechanism used by type1-piliated bacteria, we employed bacterial cell associationand modified gentamicin protection-based invasionassays with the human bladder epithelial cell line desig-nated 5637 (Fig. 1). The bladder cells were pre-treated for30 min with the indicated endocytic pathway inhibitorsprior to infection with AAEC185/pSH2, a recombinantlaboratory K-12 E. coli strain that expresses type 1 piliencoded by the fim gene cluster within the pSH2 plasmid(Orndorff and Falkow, 1984; Blomfield et al., 1991;Mulvey et al., 1998). Without expression of functional type1 pili, AAEC185 does not invade host cells (Martinezet al., 2000). The inhibitors used were chosen based ontheir reported specificity for disrupting one or more of themajor endocytic pathways and for their innocuous effectson bacterial growth and viability. Methyl-beta-cyclodextrin(MbCD, 10 mM), a widely utilized lipid raft disruptor,depletes membrane cholesterol and consequently affectsseveral endocytic pathways including caveolae- andclathrin-mediated endocytosis and macropinocytosis(Subtil et al., 1999; Simons and Toomre, 2000; Grimmeret al., 2002). Monodansylcadaverine (MDC, 300 mM), atransglutaminidase inhibitor, interferes with clathrin-mediated endocytosis (Schlegel et al., 1982). Ethylisopro-pyl amiloride (EIPA, 100 mM), a specific Na+/H+ exchange(NHE) inhibitor, prevents macropinocytosis (West et al.,1989; Meier et al., 2002). Finally, the small moleculeinhibitor dynasore (80 mM) specifically inhibits theGTPase activity of dynamin and can thereby interfere withmultiple endocytic pathways, including caveolae- andclathrin-mediated endocytosis as well as phagocytosis
2554 D. S. Eto et al.
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Cellular Microbiology, 10, 2553–2567
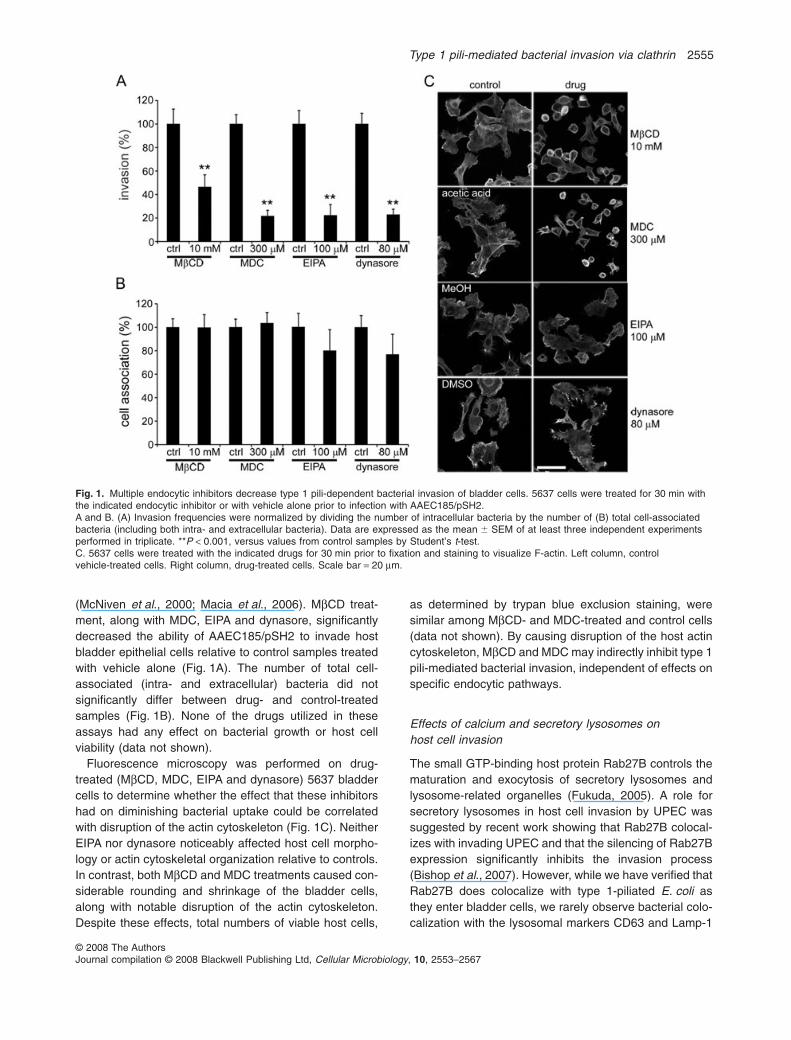
(McNiven et al., 2000; Macia et al., 2006). MbCD treat-ment, along with MDC, EIPA and dynasore, significantlydecreased the ability of AAEC185/pSH2 to invade hostbladder epithelial cells relative to control samples treatedwith vehicle alone (Fig. 1A). The number of total cell-associated (intra- and extracellular) bacteria did notsignificantly differ between drug- and control-treatedsamples (Fig. 1B). None of the drugs utilized in theseassays had any effect on bacterial growth or host cellviability (data not shown).
Fluorescence microscopy was performed on drug-treated (MbCD, MDC, EIPA and dynasore) 5637 bladdercells to determine whether the effect that these inhibitorshad on diminishing bacterial uptake could be correlatedwith disruption of the actin cytoskeleton (Fig. 1C). NeitherEIPA nor dynasore noticeably affected host cell morpho-logy or actin cytoskeletal organization relative to controls.In contrast, both MbCD and MDC treatments caused con-siderable rounding and shrinkage of the bladder cells,along with notable disruption of the actin cytoskeleton.Despite these effects, total numbers of viable host cells,
as determined by trypan blue exclusion staining, weresimilar among MbCD- and MDC-treated and control cells(data not shown). By causing disruption of the host actincytoskeleton, MbCD and MDC may indirectly inhibit type 1pili-mediated bacterial invasion, independent of effects onspecific endocytic pathways.
Effects of calcium and secretory lysosomes onhost cell invasion
The small GTP-binding host protein Rab27B controls thematuration and exocytosis of secretory lysosomes andlysosome-related organelles (Fukuda, 2005). A role forsecretory lysosomes in host cell invasion by UPEC wassuggested by recent work showing that Rab27B colocal-izes with invading UPEC and that the silencing of Rab27Bexpression significantly inhibits the invasion process(Bishop et al., 2007). However, while we have verified thatRab27B does colocalize with type 1-piliated E. coli asthey enter bladder cells, we rarely observe bacterial colo-calization with the lysosomal markers CD63 and Lamp-1
Fig. 1. Multiple endocytic inhibitors decrease type 1 pili-dependent bacterial invasion of bladder cells. 5637 cells were treated for 30 min withthe indicated endocytic inhibitor or with vehicle alone prior to infection with AAEC185/pSH2.A and B. (A) Invasion frequencies were normalized by dividing the number of intracellular bacteria by the number of (B) total cell-associatedbacteria (including both intra- and extracellular bacteria). Data are expressed as the mean � SEM of at least three independent experimentsperformed in triplicate. **P < 0.001, versus values from control samples by Student’s t-test.C. 5637 cells were treated with the indicated drugs for 30 min prior to fixation and staining to visualize F-actin. Left column, controlvehicle-treated cells. Right column, drug-treated cells. Scale bar = 20 mm.
Type 1 pili-mediated bacterial invasion via clathrin 2555
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Cellular Microbiology, 10, 2553–2567

at sites of bacterial entry (data not shown and Eto et al.,2006). Notably, Rab27B, and its isoform Rab27A, are alsopresent on non-lysosome-related organelles and likelymodulate a wide array of membrane trafficking events inaddition to lysosome exocytosis (Fukuda, 2005). Theseobservations suggested that secretory lysosomes mightnot be strictly required for UPEC entry. Furthermore,microbial invasion of non-professional phagocytic cells,such as epithelial cells and fibroblasts, via secretory lyso-somes requires intracellular calcium fluxes and a func-tional host microtubule network, but not actin (Tardieuxet al., 1992; Rodriguez et al., 1995; 1996; 1997). In con-trast, FimH-mediated UPEC entry into bladder cellsrequires both actin (Martinez et al., 2000) and microtu-bules (B.K. Dhakal and M.A. Mulvey, unpubl. obs.). Thecontribution of calcium to the FimH-mediated invasionprocess has not been described, leaving open the possi-bility that UPEC entry into host cells is at least in partmediated by the calcium-dependent trafficking of secre-tory lysosomes.
To test a role for calcium signalling, we utilized amembrane-permeable calcium chelator, BAPTA-AM, aswell as an extracellular calcium chelator, EGTA. By dis-rupting intracellular calcium fluxes, BAPTA-AM, but notnecessarily EGTA, is expected to interfere with bacterialuptake via secretory lysosomes (Andrews, 2002). Treat-ment of 5637 cells with BAPTA-AM instead resulted in asignificant, concentration-dependent rise in bacterialinvasion frequencies compared with control samples(Fig. 2A). The highest EGTA concentration used also ledto greatly enhanced bacterial invasion frequencies com-pared with controls (Fig. 2A). Neither BAPTA-AM norEGTA adversely affected bacterial adherence to the hostcells (Fig. 2B), and with the two lowest concentrations ofEGTA tested there was a slight, but significant, increase inthe numbers of cell-associated bacteria. We also testedthe effects of NiCl2, an inhibitor of transient T-type voltage-dependent calcium channels that has been shown toinhibit the efflux of UPEC from bladder cells, presumablyby interfering with the release of UPEC-containing secre-tory vesicles (Bishop et al., 2007). This compound, whichlikely does not perturb calcium levels to the same degreeas EGTA and BAPTA/AM, had no effect on either bacterialinvasion or cell association frequencies (Fig. 2A and B).
In contrast to results with the calcium chelators, artifi-cially increasing intracellular calcium levels using thecalcium ionophore ionomycin inhibited bacterial invasionfrequencies in a concentration-dependent fashion(Fig. 2C). The use of thapsigargin, a sarco/endoplasmicreticulum Ca2+ ATPase (SERCA) inhibitor that elevatescytosolic calcium levels, caused a similar decrease inintracellular bacterial titres at all drug concentrationsused. Like EGTA, thapsigargin and, to a lesser extent,ionomycin (at the highest concentration used) also
caused significant increases in bacterial adherence,perhaps by altering the availability or conformation ofFimH receptors. In total, these findings, and in particularthe results from BAPTA-AM-treated cells, are in contrastto those expected if type 1-piliated E. coli entered hostcells via a calcium-dependent secretory lysosome-mediated pathway.
To more directly address the role of secretory lyso-somes in type 1 pili-dependent bacterial entry, cell asso-ciation and invasion assays were performed with a mouseembryonic fibroblast cell line (MCHS cells derived frombeige-J mice), in which lysosome maturation and secre-tion are defective due to a mutation in the beige gene(Huynh et al., 2004). Comparisons were made withcontrol fibroblast cell lines from C57BL/6 (WT) or MCHScells complemented with the beige gene on a yeast arti-ficial chromosome (Big24) (Huynh et al., 2004). Similarnumbers of intracellular bacteria were observed within WTand Big24 cells, while substantially more bacteria weredetected within the lysosomal mutant MCHS cells(Fig. 2E). The total numbers of cell-associated bacteriawere similar for all three host cell lines (Fig. 2F). Theenhanced numbers of intracellular bacteria recoveredfrom the MCHS cells may reflect decreased levels ofbacterial trafficking back out of the host cells, increasedviability of internalized bacteria, and/or increased avail-ability of receptors that can mediate invasion, althoughsurface expression of the b1 integrin receptor was similaramong the MCHS and control cell lines (data not shown).
To assess the role of secretory lysosomes in mediatingbacterial invasion into mouse bladder epithelial cells invivo, adult female C57BL/6 (WT) or beige-J mice wereinoculated via transurethral catheterization with a wild-type UPEC cystitis isolate, UTI89. These bacteria invadebladder cells via a FimH-dependent mechanism likeAAEC185/pSH2 (Eto et al., 2007). After a 2 h infectionperiod, it was found that the UPEC isolate colonized andinvaded WT and beige-J mouse bladders similarly(Fig. 2G and H). These data, coupled with our cell culture-based observations, indicate that secretory lysosomesare not necessarily required for type 1 pili-dependent bac-terial invasion of host cells.
Type 1 pili-mediated bacterial invasion of host cellsdoes not require caveolae or flotillin-1
The inhibitory effect of dynasore on invasion (Fig. 1A)indicated that AAEC185/pSH2 enters host bladder cellsvia one of the dynamin-dependent pathways, whichinclude both caveolae- and clathrin-mediated endocyto-sis. Caveolae, in addition to their role in endocytosis,are important scaffolding domains that coordinatemany signalling events at membrane surfaces. Recentobservations that link cav1 phosphorylation with b1
2556 D. S. Eto et al.
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Cellular Microbiology, 10, 2553–2567
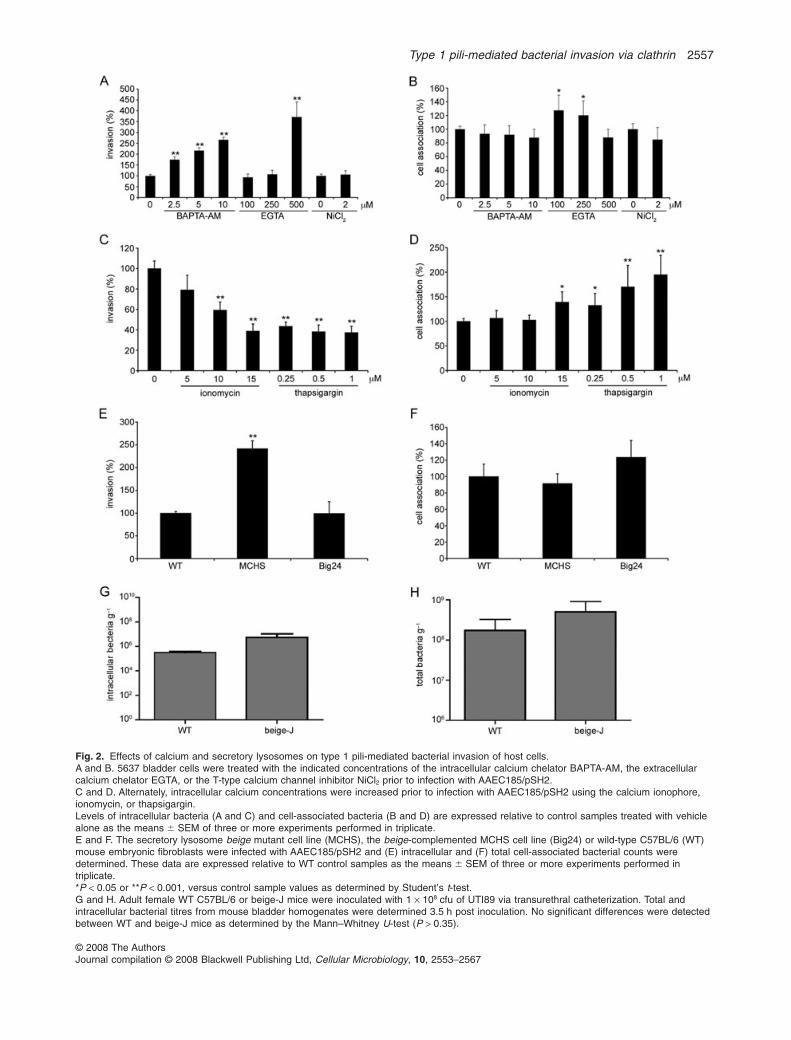
Fig. 2. Effects of calcium and secretory lysosomes on type 1 pili-mediated bacterial invasion of host cells.A and B. 5637 bladder cells were treated with the indicated concentrations of the intracellular calcium chelator BAPTA-AM, the extracellularcalcium chelator EGTA, or the T-type calcium channel inhibitor NiCl2 prior to infection with AAEC185/pSH2.C and D. Alternately, intracellular calcium concentrations were increased prior to infection with AAEC185/pSH2 using the calcium ionophore,ionomycin, or thapsigargin.Levels of intracellular bacteria (A and C) and cell-associated bacteria (B and D) are expressed relative to control samples treated with vehiclealone as the means � SEM of three or more experiments performed in triplicate.E and F. The secretory lysosome beige mutant cell line (MCHS), the beige-complemented MCHS cell line (Big24) or wild-type C57BL/6 (WT)mouse embryonic fibroblasts were infected with AAEC185/pSH2 and (E) intracellular and (F) total cell-associated bacterial counts weredetermined. These data are expressed relative to WT control samples as the means � SEM of three or more experiments performed intriplicate.*P < 0.05 or **P < 0.001, versus control sample values as determined by Student’s t-test.G and H. Adult female WT C57BL/6 or beige-J mice were inoculated with 1 ¥ 108 cfu of UTI89 via transurethral catheterization. Total andintracellular bacterial titres from mouse bladder homogenates were determined 3.5 h post inoculation. No significant differences were detectedbetween WT and beige-J mice as determined by the Mann–Whitney U-test (P > 0.35).
Type 1 pili-mediated bacterial invasion via clathrin 2557
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Cellular Microbiology, 10, 2553–2567
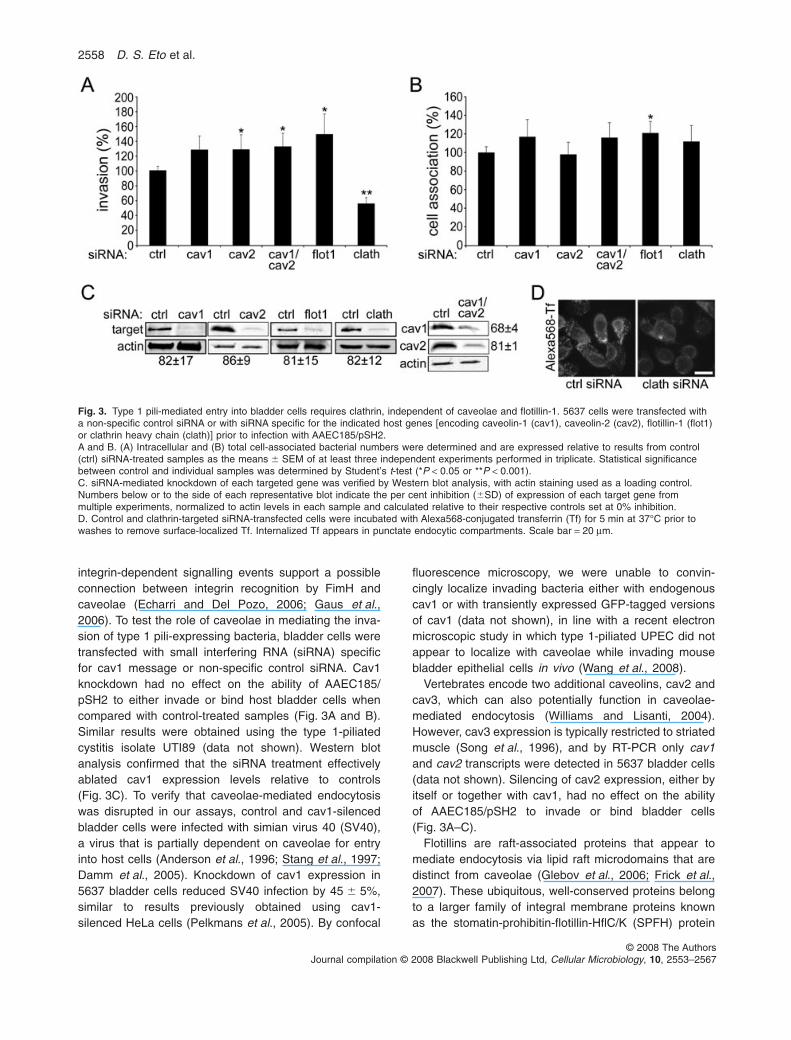
integrin-dependent signalling events support a possibleconnection between integrin recognition by FimH andcaveolae (Echarri and Del Pozo, 2006; Gaus et al.,2006). To test the role of caveolae in mediating the inva-sion of type 1 pili-expressing bacteria, bladder cells weretransfected with small interfering RNA (siRNA) specificfor cav1 message or non-specific control siRNA. Cav1knockdown had no effect on the ability of AAEC185/pSH2 to either invade or bind host bladder cells whencompared with control-treated samples (Fig. 3A and B).Similar results were obtained using the type 1-piliatedcystitis isolate UTI89 (data not shown). Western blotanalysis confirmed that the siRNA treatment effectivelyablated cav1 expression levels relative to controls(Fig. 3C). To verify that caveolae-mediated endocytosiswas disrupted in our assays, control and cav1-silencedbladder cells were infected with simian virus 40 (SV40),a virus that is partially dependent on caveolae for entryinto host cells (Anderson et al., 1996; Stang et al., 1997;Damm et al., 2005). Knockdown of cav1 expression in5637 bladder cells reduced SV40 infection by 45 � 5%,similar to results previously obtained using cav1-silenced HeLa cells (Pelkmans et al., 2005). By confocal
fluorescence microscopy, we were unable to convin-cingly localize invading bacteria either with endogenouscav1 or with transiently expressed GFP-tagged versionsof cav1 (data not shown), in line with a recent electronmicroscopic study in which type 1-piliated UPEC did notappear to localize with caveolae while invading mousebladder epithelial cells in vivo (Wang et al., 2008).
Vertebrates encode two additional caveolins, cav2 andcav3, which can also potentially function in caveolae-mediated endocytosis (Williams and Lisanti, 2004).However, cav3 expression is typically restricted to striatedmuscle (Song et al., 1996), and by RT-PCR only cav1and cav2 transcripts were detected in 5637 bladder cells(data not shown). Silencing of cav2 expression, either byitself or together with cav1, had no effect on the abilityof AAEC185/pSH2 to invade or bind bladder cells(Fig. 3A–C).
Flotillins are raft-associated proteins that appear tomediate endocytosis via lipid raft microdomains that aredistinct from caveolae (Glebov et al., 2006; Frick et al.,2007). These ubiquitous, well-conserved proteins belongto a larger family of integral membrane proteins knownas the stomatin-prohibitin-flotillin-HflC/K (SPFH) protein
Fig. 3. Type 1 pili-mediated entry into bladder cells requires clathrin, independent of caveolae and flotillin-1. 5637 cells were transfected witha non-specific control siRNA or with siRNA specific for the indicated host genes [encoding caveolin-1 (cav1), caveolin-2 (cav2), flotillin-1 (flot1)or clathrin heavy chain (clath)] prior to infection with AAEC185/pSH2.A and B. (A) Intracellular and (B) total cell-associated bacterial numbers were determined and are expressed relative to results from control(ctrl) siRNA-treated samples as the means � SEM of at least three independent experiments performed in triplicate. Statistical significancebetween control and individual samples was determined by Student’s t-test (*P < 0.05 or **P < 0.001).C. siRNA-mediated knockdown of each targeted gene was verified by Western blot analysis, with actin staining used as a loading control.Numbers below or to the side of each representative blot indicate the per cent inhibition (�SD) of expression of each target gene frommultiple experiments, normalized to actin levels in each sample and calculated relative to their respective controls set at 0% inhibition.D. Control and clathrin-targeted siRNA-transfected cells were incubated with Alexa568-conjugated transferrin (Tf) for 5 min at 37°C prior towashes to remove surface-localized Tf. Internalized Tf appears in punctate endocytic compartments. Scale bar = 20 mm.
2558 D. S. Eto et al.
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Cellular Microbiology, 10, 2553–2567
superfamily. SPFH proteins contain an evolutionarily con-served domain that appears to have affinity for lipid rafts.To examine the function of flotillin in host cell invasion bytype 1 pili-expressing bacteria, flotillin-1 was silenced in5637 bladder epithelial cells and the ability of AAEC185/pSH2 to invade the host cells was determined. Flotillin-1knockdown resulted in a slight, but significant, increase inboth bacterial invasion and adherence compared withcontrol transfected host cells (Fig. 3A and B). Silencing offlotillin-1 expression was verified by Western blot analysis(Fig. 3C). Cumulatively, these data indicate that FimH-mediated bacterial invasion of bladder cells can occurindependent of both caveolae and flotillin-1-associatedlipid raft domains.
Clathrin-mediated invasion of host cells by type1-piliated E. coli
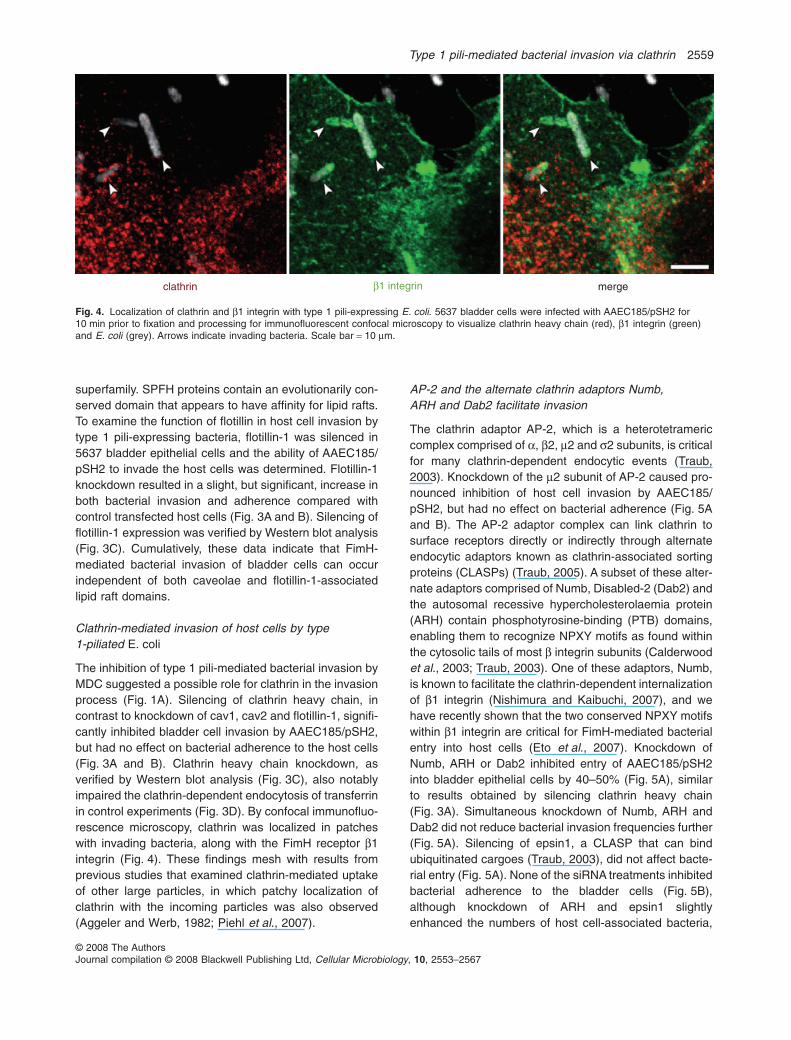
The inhibition of type 1 pili-mediated bacterial invasion byMDC suggested a possible role for clathrin in the invasionprocess (Fig. 1A). Silencing of clathrin heavy chain, incontrast to knockdown of cav1, cav2 and flotillin-1, signifi-cantly inhibited bladder cell invasion by AAEC185/pSH2,but had no effect on bacterial adherence to the host cells(Fig. 3A and B). Clathrin heavy chain knockdown, asverified by Western blot analysis (Fig. 3C), also notablyimpaired the clathrin-dependent endocytosis of transferrinin control experiments (Fig. 3D). By confocal immunofluo-rescence microscopy, clathrin was localized in patcheswith invading bacteria, along with the FimH receptor b1integrin (Fig. 4). These findings mesh with results fromprevious studies that examined clathrin-mediated uptakeof other large particles, in which patchy localization ofclathrin with the incoming particles was also observed(Aggeler and Werb, 1982; Piehl et al., 2007).
AP-2 and the alternate clathrin adaptors Numb,ARH and Dab2 facilitate invasion
The clathrin adaptor AP-2, which is a heterotetramericcomplex comprised of a, b2, m2 and s2 subunits, is criticalfor many clathrin-dependent endocytic events (Traub,2003). Knockdown of the m2 subunit of AP-2 caused pro-nounced inhibition of host cell invasion by AAEC185/pSH2, but had no effect on bacterial adherence (Fig. 5Aand B). The AP-2 adaptor complex can link clathrin tosurface receptors directly or indirectly through alternateendocytic adaptors known as clathrin-associated sortingproteins (CLASPs) (Traub, 2005). A subset of these alter-nate adaptors comprised of Numb, Disabled-2 (Dab2) andthe autosomal recessive hypercholesterolaemia protein(ARH) contain phosphotyrosine-binding (PTB) domains,enabling them to recognize NPXY motifs as found withinthe cytosolic tails of most b integrin subunits (Calderwoodet al., 2003; Traub, 2003). One of these adaptors, Numb,is known to facilitate the clathrin-dependent internalizationof b1 integrin (Nishimura and Kaibuchi, 2007), and wehave recently shown that the two conserved NPXY motifswithin b1 integrin are critical for FimH-mediated bacterialentry into host cells (Eto et al., 2007). Knockdown ofNumb, ARH or Dab2 inhibited entry of AAEC185/pSH2into bladder epithelial cells by 40–50% (Fig. 5A), similarto results obtained by silencing clathrin heavy chain(Fig. 3A). Simultaneous knockdown of Numb, ARH andDab2 did not reduce bacterial invasion frequencies further(Fig. 5A). Silencing of epsin1, a CLASP that can bindubiquitinated cargoes (Traub, 2003), did not affect bacte-rial entry (Fig. 5A). None of the siRNA treatments inhibitedbacterial adherence to the bladder cells (Fig. 5B),although knockdown of ARH and epsin1 slightlyenhanced the numbers of host cell-associated bacteria,
mergeβ1 integrinclathrin
Fig. 4. Localization of clathrin and b1 integrin with type 1 pili-expressing E. coli. 5637 bladder cells were infected with AAEC185/pSH2 for10 min prior to fixation and processing for immunofluorescent confocal microscopy to visualize clathrin heavy chain (red), b1 integrin (green)and E. coli (grey). Arrows indicate invading bacteria. Scale bar = 10 mm.
Type 1 pili-mediated bacterial invasion via clathrin 2559
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Cellular Microbiology, 10, 2553–2567
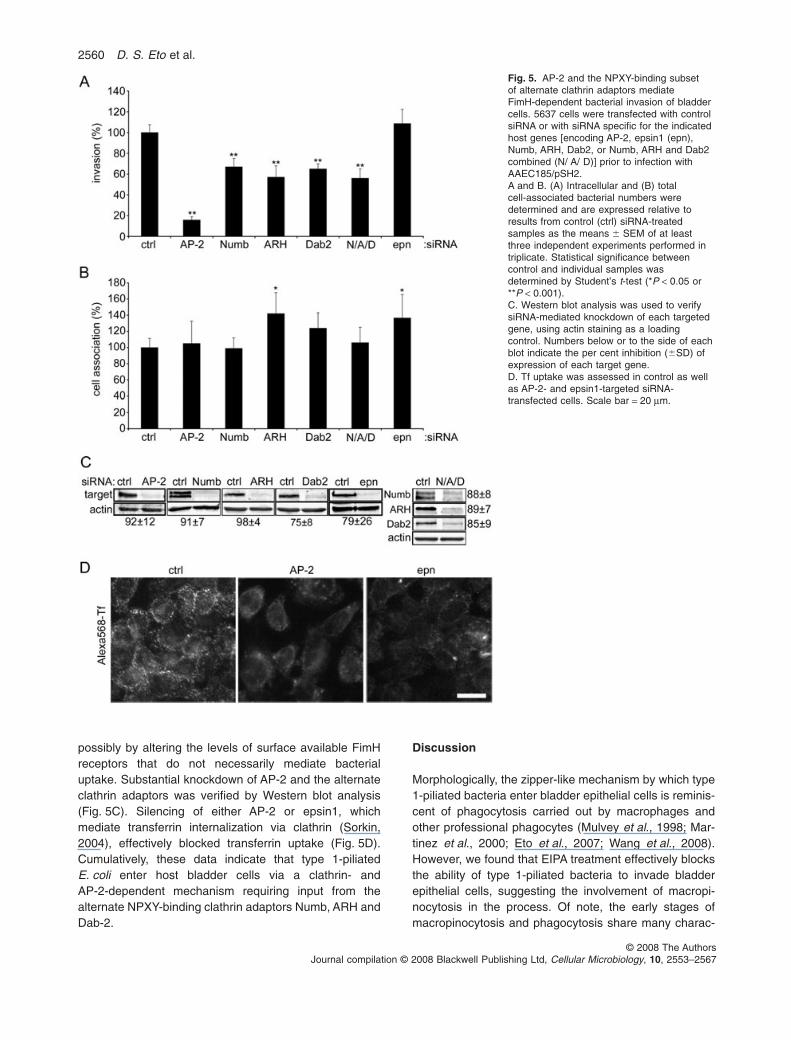
possibly by altering the levels of surface available FimHreceptors that do not necessarily mediate bacterialuptake. Substantial knockdown of AP-2 and the alternateclathrin adaptors was verified by Western blot analysis(Fig. 5C). Silencing of either AP-2 or epsin1, whichmediate transferrin internalization via clathrin (Sorkin,2004), effectively blocked transferrin uptake (Fig. 5D).Cumulatively, these data indicate that type 1-piliatedE. coli enter host bladder cells via a clathrin- andAP-2-dependent mechanism requiring input from thealternate NPXY-binding clathrin adaptors Numb, ARH andDab-2.
Discussion
Morphologically, the zipper-like mechanism by which type1-piliated bacteria enter bladder epithelial cells is reminis-cent of phagocytosis carried out by macrophages andother professional phagocytes (Mulvey et al., 1998; Mar-tinez et al., 2000; Eto et al., 2007; Wang et al., 2008).However, we found that EIPA treatment effectively blocksthe ability of type 1-piliated bacteria to invade bladderepithelial cells, suggesting the involvement of macropi-nocytosis in the process. Of note, the early stages ofmacropinocytosis and phagocytosis share many charac-
Fig. 5. AP-2 and the NPXY-binding subsetof alternate clathrin adaptors mediateFimH-dependent bacterial invasion of bladdercells. 5637 cells were transfected with controlsiRNA or with siRNA specific for the indicatedhost genes [encoding AP-2, epsin1 (epn),Numb, ARH, Dab2, or Numb, ARH and Dab2combined (N/ A/ D)] prior to infection withAAEC185/pSH2.A and B. (A) Intracellular and (B) totalcell-associated bacterial numbers weredetermined and are expressed relative toresults from control (ctrl) siRNA-treatedsamples as the means � SEM of at leastthree independent experiments performed intriplicate. Statistical significance betweencontrol and individual samples wasdetermined by Student’s t-test (*P < 0.05 or**P < 0.001).C. Western blot analysis was used to verifysiRNA-mediated knockdown of each targetedgene, using actin staining as a loadingcontrol. Numbers below or to the side of eachblot indicate the per cent inhibition (�SD) ofexpression of each target gene.D. Tf uptake was assessed in control as wellas AP-2- and epsin1-targeted siRNA-transfected cells. Scale bar = 20 mm.
2560 D. S. Eto et al.
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Cellular Microbiology, 10, 2553–2567

teristics, including their dependence on actin dynamicsand use of similar scaffolding and signalling moleculessuch as phosphatidylinositide 3-kinase (Cardelli, 2001;Vieira et al., 2002; Lefkir et al., 2004; Swanson, 2008).Consequently, use of EIPA, which likely interferes withmacropinocytosis by disrupting endosomal trafficking(Gekle et al., 2001; Meier et al., 2002), does not defini-tively implicate macropinocytosis in the invasion process.Furthermore, macropinocytosis does not necessarilyrequire the GTPase dynamin (Conner and Schmid, 2003),in contrast to the uptake of type 1-piliated E. coli, whichwe found to be significantly attenuated by the dynamininhibitor dynasore (see Fig. 1). This observation, coupledwith previous microscopic studies showing the plasmamembrane of bladder epithelial cells zippering aroundinvading UPEC (Mulvey et al., 1998; Martinez et al., 2000;Eto et al., 2007; Wang et al., 2008), argues against acentral role for macropinocytosis in type 1 pili-mediatedbacterial invasion of host cells.
The phagocytosis of large particles, like bacteria,requires the addition of membrane to the cell surface at thebase of the phagocytic cup (Braun and Niedergang, 2006;Swanson, 2008). In professional phagocytes, like macro-phages, this additional membrane is derived from theexocytic transport of endosomal compartments, lyso-somes, endoplasmic reticulum or other organelles. It isfeasible that the envelopment and internalization of type1-piliated E. coli by bladder epithelial cells also requires theaddition of membrane at sites of bacterial entry. The colo-calization of Rab27B with invading UPEC supports thispossibility (Bishop et al., 2007). This small GTP-bindingprotein, along with its closely related isoform Rab27A, canregulate multiple membrane trafficking events, includingthe calcium-induced exocytosis of secretory lysosomes(Fukuda, 2005). The colocalization of UPEC with Rab27B,coupled with observations that UPEC can trigger intracel-lular calcium fluxes and the release of lysosomal enzymes,suggested that UPEC might utilize secretory lysosomes togain entry into host cells, similar to T. cruzi (Bishop et al.,2007). However, while we do detect colocalization ofRab27B with invading E. coli, we could not validate a rolefor secretory lysosomes in the FimH-mediated invasionprocess. Specifically, the chelation of intracellular calciumby BAPTA-AM, which interferes with the exocytosis ofsecretory lysosomes (Rodriguez et al., 1997; Andrews,2000; Jaiswal et al., 2002), actually enhanced bacterialinvasion frequencies, while artificially elevating intracellu-lar calcium levels had the opposite effect. In addition, type1-piliated bacteria had no problem entering either fibro-blasts or bladder epithelial cells from beige mice, in whichthe exocytosis of secretory lysosomes is defective. Theseresults agree with our earlier observation that UPEC doesnot consistently localize with the lysosomal marker CD63until several hours after invading bladder epithelial cells
(Eto et al., 2006). Thus, while secretory lysosomes maymediate UPEC internalization under some conditions, ourdata indicate that they are not strictly required. Instead, wepropose that Rab27B facilitates the delivery of membranefrom other non-lysosomal compartments to sites of bacte-rial entry, aiding the envelopment and internalization of thebound microbes.
A key regulator of host cell invasion by many pathogensis cholesterol (Lafont and van der Goot, 2005). Here, wefound that sequestration of cholesterol using MbCD effec-tively inhibits bladder cell invasion by type 1-piliatedE. coli (see Fig. 1). Superficially, this observation sug-gests a role for cholesterol and lipid rafts in the invasionprocess, in agreement with previously published dataimplicating caveolae raft domains in the internalization ofUPEC (Duncan et al., 2004). However, in addition to itseffects on lipid rafts, cholesterol depletion by MbCD treat-ment can also disturb other host cell activities, includingactin dynamics and the localization of a key secondmessenger, phosphatidylinositol (4,5)-bisphosphate[PtdIns(4,5)P2] (Pike and Miller, 1998; Kwik et al., 2003).In our assays, we observed that MbCD treatment resultedin substantial alterations of the host actin cytoskeleton,suggesting that this drug may indirectly inhibit bacterialinvasion by disrupting the actin cytoskeleton rather thanlipid rafts per se. We also found that silencing of the raftdomain-associated proteins cav1, cav2 and flotillin-1 hadno inhibitory effect on type 1 pili-mediated bacterial inva-sion, despite the fact that the knockdown of cav1 effec-tively reduced caveolae-mediated internalization of SV40in our assays. Taken together, these results indicatethat neither caveolae nor flotillin-1-associated lipid raftdomains are absolutely essential for FimH-mediated bac-terial entry into bladder epithelial cells, although othercholesterol-rich raft domains may be required. Our find-ings do not exclude the possibility that FimH may be ableto direct bacterial invasion of host cells by more than onepathway, including the use of caveolae or secretory lyso-somes under some circumstances.
The observation that MDC, a known inhibitor of clathrin-dependent receptor-mediated endocytosis, substantiallydecreased bacterial invasion frequencies suggested afunctional link between clathrin and type 1 pili-dependentbacterial uptake by bladder cells. In addition to its inhibitoryeffects on receptor-mediated endocytosis via stabilizationof clathrin assemblies (Phonphok and Rosenthal, 1991),MDC also inactivates host transglutaminases, which arecalcium-dependent protein cross-linking enzymes thathave been implicated as modulators of receptor-mediatedinternalization (Davies and Murtaugh, 1984; Abe et al.,2000; Mehta et al., 2006). Because the enzymatic functionof these transglutaminases requires calcium, and consid-ering that the treatment of host bladder cells with calciumchelators does not negatively affect bacterial entry, it is
Type 1 pili-mediated bacterial invasion via clathrin 2561
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Cellular Microbiology, 10, 2553–2567

unlikely that type 1 pili-mediated bacterial uptake dependson host transglutaminase activity. Rather, MDC treatmentprobably inhibits type 1 pili-mediated bacterial invasion ofhost cells via alternate effects on clathrin or other hostfunctions. For example, MDC (similar to MbCD) causednotable disruption of the host actin cytoskeleton in ourassays, a phenomenon that likely contributes to the inhibi-tory effect of MDC on bacterial invasion.
Clathrin is typically associated with the endocytosis ofnutrients, receptors and other cell surface molecules(Ungewickell and Hinrichsen, 2007). However, a fewrecent studies have demonstrated that clathrin can alsomediate the internalization of much larger particles,including gap junction plaques and at least three invasivebacterial pathogens that enter host cells via zipper-likemechanisms (Veiga and Cossart, 2005; Piehl et al., 2007;Veiga et al., 2007). One of these pathogens, Listeriamonocytogenes, invades host cells by binding the recep-tors E-cadheren and Met (Gaillard et al., 1991; Lecuitet al., 1999; Shen et al., 2000), while the other two, Yers-inia pseudotuberculosis and Staphylococcus aureus,utilize b1 integrin receptors, similar to type 1-piliatedUPEC (Isberg and Leong, 1990; Grundmeier et al., 2004;Agerer et al., 2005; Eto et al., 2007). Extending our find-ings with MDC, we confirmed that type 1-piliated bacteriacould also hijack the clathrin-associated endocyticmachinery. By confocal microscopy, we observed patchesof clathrin localized with invading AAEC185/pSH2 alongwith b1 integrin (see Fig. 4). In addition, silencing expres-sion of either clathrin heavy chain or the major clathrinadaptor AP-2 significantly inhibited FimH-mediatedbacterial invasion (Figs 3 and 5). The more substantialinhibition of FimH-mediated bacterial invasion that weobserved with AP-2 versus clathrin knockdown may bedue to variations in how well these proteins were silencedas well as their initial relative abundance in the target hostcells. It is also possible that AP-2 effects during the inva-sion process extend beyond clathrin. In contrast to ourresults, the clathrin-dependent entry of L. monocytogenesinto host cells does not require AP-2 and instead relies onanother adaptor complex, AP-1 (Pizarro-Cerda et al.,2007; Veiga et al., 2007). Variances in the cellular machin-ery used during clathrin-mediated bacterial invasion ofhost cells may reflect differences in the host receptors thatare engaged to initiate the entry process.
In addition to AP-2, we found that FimH-mediated bac-terial invasion of bladder cells also requires a subset ofalternate clathrin adaptors comprised of Numb, ARH andDab2. Each of these adaptors has similar N-terminal PTBdomains that recognize both PtdIns(4,5)P2 and NPXYmotifs (Traub, 2003). The C-terminal domains of thesealternate adaptors bind AP-2 and, with the exception ofNumb, the clathrin heavy chain terminal domain. Dab2and ARH have been shown to mediate the internalization
of low-density lipoprotein receptor (LDLR) family mem-bers (Morris and Cooper, 2001; He et al., 2002; Mishraet al., 2002a,b; Nagai et al., 2003), while Numb is im-plicated in the endocytosis of b integrin subunits (Nishi-mura and Kaibuchi, 2007). In our assays, Numb, ARH andDab2 appear to act in a cooperative fashion to internalizetype 1-piliated E. coli. We speculate that each of theseadaptors works in conjunction with AP-2 to bind one ortwo NPXY motifs within the cytosolic tail of b1 integrinreceptors, stimulating the assembly of clathrin at sites ofbacterial entry. However, the mechanisms by which clath-rin and associated adaptor proteins drive bacterial entryinto host cells remain unclear. Of note, Dab2 has recentlybeen shown to mediate the clathrin-dependent internal-ization of gap junction plaques (Piehl et al., 2007), sug-gesting that members of the NPXY-binding subset ofalternate clathrin adaptors may have a general role in theclathrin-mediated uptake of large particles.
In conclusion, our data show that type 1-piliated bacte-ria like UPEC invade host cells via a phagocytosis-likeactin-dependent calcium-modulated zipper mechanismrequiring clathrin, dynamin, AP-2, and at least three alter-nate clathrin adaptors – Numb, ARH and Dab2. Genesilencing experiments suggest that these NPXY-bindingadaptors act cooperatively, and in a non-redundantfashion, during the invasion process. A more detailedanalysis of the temporal and spatial links among clathrin,adaptor proteins, cytoskeletal dynamics, and FimH-mediated recognition of integrins and other host receptorspromises to enlighten our understanding of UTIs as wellas the emerging role of clathrin in bacterial pathogenesis.
Experimental procedures
Cell lines and bacteria
The human bladder epithelial cell line 5637 (HTB-9; ATCC) wasmaintained in RPMI 1640 medium supplemented with 10%heat-inactivated fetal bovine serum (FBS) at 37°C (5% CO2).Fibroblasts from normal C57BL/6 mice (WT), beige-J micelacking a functional beige gene (MCHS), or beige-J MCHS cellscomplemented with a yeast artificial chromosome (YAC) carryingthe beige gene (Big24) (kindly provided by the J. Kaplan lab)were grown in DMEM plus 10% FBS. G418, at a concentration of1 mg ml-1, was used to maintain the stably transfected fibroblastcell line Big24. The recombinant type 1-piliated K-12 laboratoryE. coli strain AAEC185/pSH2 and the clinical cystitis isolateUTI89 have been described (Mulvey et al., 1998; Chen et al.,2006). Bacterial strains were grown statically for 48 h at 37°C in20 ml of Luria–Bertani (LB) broth to induce type 1 pili expression.
Gentamicin protection-based invasion assays,drugs and gene silencing
Modified gentamicin protection-based assays were performedessentially as described previously (Elsinghorst, 1994; Eto et al.,
2562 D. S. Eto et al.
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Cellular Microbiology, 10, 2553–2567

2007). Briefly, host cells were seeded into 24-well plates andgrown to confluency at 37°C (5% CO2). AAEC185/pSH2 wasused to infect the monolayers at a multiplicity of infection (moi)of 1–15 bacteria per host cell. In order to expedite and syn-chronize bacterial contact with the cell monolayers, plates werespun at 600 g for 5 min prior to a 2 h incubation at 37°C. Forcell association assays, host cell monolayers were washed fourtimes with PBS2+ (PBS plus Mg2+/Ca2+) before being lysed in1 ml of PBS2+ containing 0.4% Triton X-100. For invasionassays, extracellular bacteria were killed by addition of themembrane-impermeable antibiotic gentamicin (100 mg ml-1;Sigma). Two hours following addition of gentamicin, the cellmonolayers were washed four times with PBS2+ and lysed. Bac-terial titres from the lysates were determined by plating serialdilutions on LB agar plates. All results from the invasion assayswere normalized by dividing the number of intracellular bacteriaby the total number of cell-associated bacteria, accounting forany differences in host cell numbers. Levels of intracellular orcell-associated bacteria were expressed relative to controlsamples. All assays were repeated at least three times in trip-licate and significant differences between controls and experi-mental samples were determined by Student’s t-test (withP < 0.05 considered significant).
5637 bladder epithelial cells were pre-incubated with drugs orwith vehicle for 30 min prior to the addition of bacteria with theexception of thapsigargin, which was added to host cells 2 hprior to infection, and NiCl2, which was added 90 min beforeinfection. Drugs (or vehicle) were maintained during the entire2 h incubations post infection prior to washes or the addition ofgentamicin. MbCD, EIPA and EGTA were purchased fromSigma-Aldrich, BAPTA-AM was from Invitrogen, ionomycin andthapsigargin were obtained from Biomol, NiCl2 was fromScholAR Chemistry, and dynasore was kindly provided by T.Kirchhausen (Harvard). Dynasore, MbCD, BAPTA-AM, ionomy-cin and thapsigargin were solubilized in DMSO, while MDC wasdissolved in 1 N acetic acid, EIPA in MeOH, NiCl2 in ddH2O andEGTA in 1 N NaOH. All assays were performed using completemedium except for those employing MbCD and dynasore, forwhich serum-free RPMI was used. None of the drugs at thefinal concentrations used had any notable effects either on hostcell viability (as determined by trypan blue exclusion assays) oron bacterial growth and viability during the course of theassays.
For gene expression knockdown studies, 80% confluent 5637cells in T25 flasks were transfected with non-specific controlsiRNA or with gene-specific SmartPool siRNA (Dharmacon)directed against caveolin-1 (cav1), clathrin heavy chain (clath),epsin1 (epn), Dab2, ARH or flotillin-1 (flot1) using Dharmafect 1reagent (Dharmacon). This reagent allowed for transfection ofnearly all cells in each monolayer. The m2 subunit of AP-2 wasspecifically targeted with a custom oligo [AAGUGGAUGCCUUUCGGGUCA (Motley et al., 2003)], as was Numb [GGACCTCATAGTTGACCAG (Nishimura and Kaibuchi, 2007)]. Controlsamples for AP-2 and Numb silencing were transfectedwith scrambled siRNA oligos. The cell culture medium wasreplaced 10–24 h post transfection, and 24 h later the cellswere seeded into 24-well plates for use in invasion and cellassociation assays. These were performed 72 h post transfec-tion, except for AP-2- and clathrin-silenced cells, which wereused 96 h after the first of two serial transfections separated by48 h.
Mouse infections
Control C57BL/6 or beige-J mice (Jackson Laboratories) werebriefly sedated with isoflurane and transurethrally inoculated with1 ¥ 108 colony-forming units (cfu) of UTI89. Bladders were asep-tically removed 2 h post infection, splayed and incubated inRPMI � 100 mg ml-1 gentamicin. Ninety minutes later, bladderhalves were washed five times with PBS2+ and lysed in 0.025%Triton X-100/PBS using a tissue grinder. Total and gentamicin-protected (intracellular) bacterial titres were determined by serialdilution of lysates on LB agar plates. These experiments wereperformed twice with three or more mice in each group (n = 6 or7). All animal experiments were performed under accreditedconditions after approval by the local Animal Care and UseCommittee.
Western blot analysis
Gene expression knockdown by siRNA was verified as previouslydescribed (Eto et al., 2007). Briefly, 5637 cells were lysed inTNN buffer (50 mM Tris pH 7.4, 150 mM NaCl, 1% NP-40)supplemented with 1¥ complete protease inhibitors (Roche) and200 mM phenylmethanesulfonylfluoride (Sigma-Aldrich). Proteincontent within the post-nuclear supernatant was determinedusing the BCA reagent system (Pierce) and equivalent proteinamounts were resolved by SDS-PAGE, transferred to ImmobilonPVDF-FL membrane (Millipore), blocked, and incubated withrabbit anti-caveolin-1 (1:1000; Abcam), mouse anti-caveolin-2(1:250; BD Transduction Laboratories), rabbit anti-epsin (1:500;Santa Cruz), rabbit anti-Numb (1:1000; Cell Signaling), rabbitanti-Dab2 (1:250; Santa Cruz), mouse anti-ARH (1:500; Abcam),mouse anti-flotillin-1 (1:250; BD Transduction Laboratories),rabbit anti-clathrin heavy chain (1:500; Abcam), mouse anti-AP-2(1:250; BD Transduction Laboratories) or mouse anti-actin anti-bodies (1:400, Abcam). Blots were subsequently probed, visual-ized, and quantified using IRDye-labelled secondary antibodiesand a LI-COR Odyssey Infrared Imaging System (LI-COR Bio-sciences). Levels of each targeted protein were normalized toactin levels in each sample.
Fluorescence microscopy
To assess drug effects on host cell morphology and actin archi-tecture, 5637 bladder cells grown on 12 mm glass coverslipswere fixed in 3% paraformaldehyde/PBS for 20 min at roomtemperature following treatment with the indicated drugs orcarrier alone. Fixed cells were then washed with PBS and stainedfor 30 min using Alexa546-conjugated phalloidin (1:40; Invitro-gen). Alternately, 5637 cells were fixed after a 10 min infectionwith AAEC185/pSH2 (moi = 15), washed with PBS, incubated for20 min in blocking buffer (3% BSA/0.05% powdered milk/0.01%saponin/PBS), and stained using rat anti-b1 integrin (AIIB2,1:100; Developmental Studies Hybridoma Bank at the Universityof Iowa), rabbit anti-clathrin heavy chain (1:300; Cell Signaling)and goat anti-E. coli (1:500; BioDesign) primary antibodies alongwith appropriate Alexa fluor-conjugated secondary antibodies(1:500; Invitrogen). Following final washed in PBS, samples weremounted onto slides using FluorSave reagent (Calbiochem/EMDBiosciences). Samples were imaged using a 100¥ 1.4 OlympusPlanApo oil immersion objective on either an Olympus FV-300 ora FV-1000 confocal microscope utilizing Fluoview software.
Type 1 pili-mediated bacterial invasion via clathrin 2563
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Cellular Microbiology, 10, 2553–2567

Transferrin uptake by 5637 cells grown on 12 mm glasscoverslips was assessed 72–96 h post transfection with theindicated siRNA oligos. Transfected bladder cells were serumstarved overnight prior to a 15 min incubation on ice with 5 mg ml-1
Alexa568-conjugated transferrin (Tf, Invitrogen) in RPMI.Samples were then washed five times with cold RPMI to removeany unbound Tf prior to a 5 min incubation at 37°C to stimulate Tfuptake. Next, cells were washed twice in an acidic buffer [0.2 mMglycine (pH 2), 0.7% NaCl] to remove any remaining extracellularTf and subsequently fixed for 20 min in 2.5% paraformaldehyde/PBS at 37°C, washed and mounted. Samples were imaged usinga 40¥ UPlanF1 objective on an Olympus BX51 fluorescent micro-scope equipped with a Magnafire SP camera.
SV40 infection
Stocks of SV40 (VR-820, ATCC) were collected from infectedCV-1 African green monkey kidney cells and stored at -70°Cessentially as previously described (Levine and Teresky, 1970).5637 cells, treated with either cav1-specific or non-specific siRNAand grown in six-well plates, were infected with SV40 in 200 ml ofcomplete RPMI rocking on ice for 60 min. After three washes withcold medium to remove unbound virus, 1 ml of warm mediumwas added to each well and plates were incubated for 6 days at37°C prior to lysis and sonication in RIPA buffer [25 mM Tris-HCl(pH 7.4), 150 mM NaCl, 1% NP-40, 1% sodium deoxycholate,0.1% SDS] containing 1¥ complete protease inhibitors (Roche)and 200 mM phenylmethanesulfonylfluoride (Sigma-Aldrich).Protein content within the lysates was determined using the BCAreagent system (Pierce) and equivalent protein amounts wereresolved by SDS-PAGE, transferred to Immobilon PVDF-FLmembranes (Millipore), blocked, and probed using mouseanti-SV40 Large T antigen (1:200; Neomarkers), rabbit anti-caveolin-1 (1:1000; Abcam) and mouse anti-actin (1:400;Abcam). Blots from three independent experiments were visual-ized and quantified using a LI-COR Odyssey Infrared ImagingSystem (LI-COR Biosciences). Levels of large T antigen expres-sion, indicative of overall virus infection levels, were normalizedto actin levels in each sample.
Acknowledgements
We are grateful to J. Kaplan and D. Ward (University of Utah) forproviding the MEF cell lines. Dynasore, which was synthesized byH. Pelish, was kindly provided by the Kirchhausen lab (HarvardMedical School). We also thank C. Rodesch and the University ofUtah School of Medicine Cell Imaging Facility for help with themicroscopy. This work was supported by US National Institutesof Health (NIH) Microbial Pathogenesis Training Grant T32AI055434-01A1 (to D.S.E.) and by NIH Grant DK068585.
References
Abe, S., Yamashita, K., Kohno, H., and Ohkubo, Y. (2000)Involvement of transglutaminase in the receptor-mediatedendocytosis of mouse peritoneal macrophages. Biol PharmBull 23: 1511–1513.
Agerer, F., Lux, S., Michel, A., Rohde, M., Ohlsen, K., andHauck, C.R. (2005) Cellular invasion by Staphylococcus
aureus reveals a functional link between focal adhesionkinase and cortactin in integrin-mediated internalisation.J Cell Sci 118: 2189–2200.
Aggeler, J., and Werb, Z. (1982) Initial events during phago-cytosis by macrophages viewed from outside and insidethe cell: membrane–particle interactions and clathrin. J CellBiol 94: 613–623.
Anderson, R.G. (1998) The caveolae membrane system.Annu Rev Biochem 67: 199–225.
Anderson, H.A., Chen, Y., and Norkin, L.C. (1996) Boundsimian virus 40 translocates to caveolin-enriched mem-brane domains, and its entry is inhibited by drugs thatselectively disrupt caveolae. Mol Biol Cell 7: 1825–1834.
Andrade, L.O., and Andrews, N.W. (2004) Lysosomal fusionis essential for the retention of Trypanosoma cruzi insidehost cells. J Exp Med 200: 1135–1143.
Andrews, N.W. (2000) Regulated secretion of conventionallysosomes. Trends Cell Biol 10: 316–321.
Andrews, N.W. (2002) Lysosomes and the plasma mem-brane: trypanosomes reveal a secret relationship. J CellBiol 158: 389–394.
Bishop, B.L., Duncan, M.J., Song, J., Li, G., Zaas, D., andAbraham, S.N. (2007) Cyclic AMP-regulated exocytosis ofEscherichia coli from infected bladder epithelial cells. NatMed 13: 625–630.
Blomfield, I.C., McClain, M.S., and Eisenstein, B.I. (1991)Type 1 fimbriae mutants of Escherichia coli K12: charac-terization of recognized afimbriate strains and constructionof new fim deletion mutants. Mol Microbiol 5: 1439–1445.
Blott, E.J., and Griffiths, G.M. (2002) Secretory lysosomes.Nat Rev Mol Cell Biol 3: 122–131.
Braun, V., and Niedergang, F. (2006) Linking exocytosis andendocytosis during phagocytosis. Biol Cell 98: 195–201.
Calderwood, D.A., Fujioka, Y., de Pereda, J.M., Garcia-Alvarez, B., Nakamoto, T., Margolis, B., et al. (2003)Integrin beta cytoplasmic domain interactions withphosphotyrosine-binding domains: a structural prototypefor diversity in integrin signaling. Proc Natl Acad Sci USA100: 2272–2277.
Cardelli, J. (2001) Phagocytosis and macropinocytosis inDictyostelium: phosphoinositide-based processes, bio-chemically distinct. Traffic 2: 311–320.
Chen, S.L., Hung, C.S., Xu, J., Reigstad, C.S., Magrini, V.,Sabo, A., et al. (2006) Identification of genes subject topositive selection in uropathogenic strains of Escherichiacoli: a comparative genomics approach. Proc Natl Acad SciUSA 103: 5977–5982.
Conner, S.D., and Schmid, S.L. (2003) Regulated portals ofentry into the cell. Nature 422: 37–44.
Cossart, P., and Sansonetti, P.J. (2004) Bacterial invasion:the paradigms of enteroinvasive pathogens. Science 304:242–248.
Damm, E.M., Pelkmans, L., Kartenbeck, J., Mezzacasa, A.,Kurzchalia, T., and Helenius, A. (2005) Clathrin- andcaveolin-1-independent endocytosis: entry of simianvirus 40 into cells devoid of caveolae. J Cell Biol 168:477–488.
Davies, P.J., and Murtaugh, M.P. (1984) Transglutaminaseand receptor-mediated endocytosis in macrophages andcultured fibroblasts. Mol Cell Biochem 58: 69–77.
2564 D. S. Eto et al.
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Cellular Microbiology, 10, 2553–2567

Dhakal, B.K., Kulesus, R.R., and Mulvey, M.A. (2008)Mechanisms and consequences of bladder cell invasion byuropathogenic Escherichia coli. Eur J Clin Invest (in press)DOI: 10.1111/j.1365-2362.2008.01986.x
Drab, M., Verkade, P., Elger, M., Kasper, M., Lohn, M., Lau-terbach, B., et al. (2001) Loss of caveolae, vascular dys-function, and pulmonary defects in caveolin-1 gene-disrupted mice. Science 293: 2449–2452.
Duncan, M.J., Shin, J.S., and Abraham, S.N. (2002) Microbialentry through caveolae: variations on a theme. Cell Micro-biol 4: 783–791.
Duncan, M.J., Li, G., Shin, J.S., Carson, J.L., and Abraham,S.N. (2004) Bacterial penetration of bladder epitheliumthrough lipid rafts. J Biol Chem 279: 18944–18951.
Echarri, A., and Del Pozo, M.A. (2006) Caveolae internaliza-tion regulates integrin-dependent signaling pathways. CellCycle 5: 2179–2182.
Elsinghorst, E.A. (1994) Measurement of invasion by gen-tamicin resistance. Methods Enzymol 236: 405–420.
Eto, D.S., Sundsbak, J.L., and Mulvey, M.A. (2006) Actin-gated intracellular growth and resurgence of uropathogenicEscherichia coli. Cell Microbiol 8: 704–717.
Eto, D.S., Jones, T.A., Sundsbak, J.L., and Mulvey, M.A.(2007) Integrin-mediated host cell invasion by type1-piliated uropathogenic Escherichia coli. PLoS Pathog 3:e100.
Frick, M., Bright, N.A., Riento, K., Bray, A., Merrified, C., andNichols, B.J. (2007) Coassembly of flotillins induces forma-tion of membrane microdomains, membrane curvature,and vesicle budding. Curr Biol 17: 1151–1156.
Fukuda, M. (2005) Versatile role of Rab27 in membranetrafficking: focus on the Rab27 effector families. J Biochem137: 9–16.
Gaillard, J.L., Berche, P., Frehel, C., Gouin, E., and Cossart,P. (1991) Entry of L. monocytogenes into cells is mediatedby internalin, a repeat protein reminiscent of surfaceantigens from gram-positive cocci. Cell 65: 1127–1141.
Gaus, K., Le Lay, S., Balasubramanian, N., and Schwartz,M.A. (2006) Integrin-mediated adhesion regulates mem-brane order. J Cell Biol 174: 725–734.
Gekle, M., Freudinger, R., and Mildenberger, S. (2001)Inhibition of Na+–H+ exchanger-3 interferes with apicalreceptor-mediated endocytosis via vesicle fusion. J Physiol531: 619–629.
Glebov, O.O., Bright, N.A., and Nichols, B.J. (2006) Flotillin-1defines a clathrin-independent endocytic pathway in mam-malian cells. Nat Cell Biol 8: 46–54.
Grimmer, S., Van Deurs, B., and Sandvig, K. (2002) Mem-brane ruffling and macropinocytosis in A431 cells requirecholesterol. J Cell Sci 115: 2953–2962.
Gruenberg, J., and van der Goot, F.G. (2006) Mechanisms ofpathogen entry through the endosomal compartments. NatRev Mol Cell Biol 7: 495–504.
Grundmeier, M., Hussain, M., Becker, P., Heilmann, C.,Peters, G., and Sinha, B. (2004) Truncation of fibronectin-binding proteins in Staphylococcus aureus strain Newmanleads to deficient adherence and host cell invasion due toloss of the cell wall anchor function. Infect Immun 72:7155–7163.
He, G., Gupta, S., Yi, M., Michaely, P., Hobbs, H.H., andCohen, J.C. (2002) ARH is a modular adaptor protein that
interacts with the LDL receptor, clathrin, and AP-2. J BiolChem 277: 44044–44049.
Hung, C.S., Bouckaert, J., Hung, D., Pinkner, J., Widberg, C.,DeFusco, A., et al. (2002) Structural basis of tropism ofEscherichia coli to the bladder during urinary tract infection.Mol Microbiol 44: 903–915.
Huynh, C., Roth, D., Ward, D.M., Kaplan, J., and Andrews,N.W. (2004) Defective lysosomal exocytosis and plasmamembrane repair in Chediak-Higashi/beige cells. Proc NatlAcad Sci USA 101: 16795–16800.
Isberg, R.R., and Leong, J.M. (1990) Multiple beta 1 chainintegrins are receptors for invasin, a protein that promotesbacterial penetration into mammalian cells. Cell 60: 861–871.
Jaiswal, J.K., Andrews, N.W., and Simon, S.M. (2002) Mem-brane proximal lysosomes are the major vesicles respon-sible for calcium-dependent exocytosis in nonsecretorycells. J Cell Biol 159: 625–635.
Kwik, J., Boyle, S., Fooksman, D., Margolis, L., Sheetz, M.P.,and Edidin, M. (2003) Membrane cholesterol, lateralmobility, and the phosphatidylinositol 4,5-bisphosphate-dependent organization of cell actin. Proc Natl Acad SciUSA 100: 13964–13969.
Lafont, F., and van der Goot, F.G. (2005) Bacterial invasionvia lipid rafts. Cell Microbiol 7: 613–620.
Lecuit, M., Dramsi, S., Gottardi, C., Fedor-Chaiken, M., Gum-biner, B., and Cossart, P. (1999) A single amino acid inE-cadherin responsible for host specificity towards thehuman pathogen Listeria monocytogenes. EMBO J 18:3956–3963.
Lefkir, Y., Malbouyres, M., Gotthardt, D., Ozinsky, A., Cornil-lon, S., Bruckert, F., et al. (2004) Involvement of the AP-1adaptor complex in early steps of phagocytosis and mac-ropinocytosis. Mol Biol Cell 15: 861–869.
Levine, A.J., and Teresky, A.K. (1970) Deoxyribonucleic acidreplication in simian virus 40-infected cells. II. Detectionand characterization of simian virus 40 pseudovirions.J Virol 5: 451–457.
Macia, E., Ehrlich, M., Massol, R., Boucrot, E., Brunner, C.,and Kirchhausen, T. (2006) Dynasore, a cell-permeableinhibitor of dynamin. Dev Cell 10: 839–850.
McNiven, M.A., Cao, H., Pitts, K.R., and Yoon, Y. (2000) Thedynamin family of mechanoenzymes: pinching in newplaces. Trends Biochem Sci 25: 115–120.
Martinez, J.J., and Hultgren, S.J. (2002) Requirement of Rho-family GTPases in the invasion of Type 1-piliated uropatho-genic Escherichia coli. Cell Microbiol 4: 19–28.
Martinez, J.J., Mulvey, M.A., Schilling, J.D., Pinkner, J.S.,and Hultgren, S.J. (2000) Type 1 pilus-mediated bacterialinvasion of bladder epithelial cells. EMBO J 19: 2803–2812.
Mehta, K., Fok, J.Y., and Mangala, L.S. (2006) Tissue trans-glutaminase: from biological glue to cell survival cues.Front Biosci 11: 173–185.
Meier, O., Boucke, K., Hammer, S.V., Keller, S., Stidwill,R.P., Hemmi, S., and Greber, U.F. (2002) Adenovirus trig-gers macropinocytosis and endosomal leakage togetherwith its clathrin-mediated uptake. J Cell Biol 158: 1119–1131.
Miaczynska, M., and Stenmark, H. (2008) Mechanisms andfunctions of endocytosis. J Cell Biol 180: 7–11.
Type 1 pili-mediated bacterial invasion via clathrin 2565
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Cellular Microbiology, 10, 2553–2567

Mishra, S.K., Keyel, P.A., Hawryluk, M.J., Agostinelli, N.R.,Watkins, S.C., and Traub, L.M. (2002a) Disabled-2 exhibitsthe properties of a cargo-selective endocytic clathrinadaptor. EMBO J 21: 4915–4926.
Mishra, S.K., Watkins, S.C., and Traub, L.M. (2002b) Theautosomal recessive hypercholesterolemia (ARH) proteininterfaces directly with the clathrin-coat machinery. ProcNatl Acad Sci USA 99: 16099–16104.
Morris, S.M., and Cooper, J.A. (2001) Disabled-2 colocalizeswith the LDLR in clathrin-coated pits and interacts withAP-2. Traffic 2: 111–123.
Motley, A., Bright, N.A., Seaman, M.N., and Robinson, M.S.(2003) Clathrin-mediated endocytosis in AP-2-depletedcells. J Cell Biol 162: 909–918.
Mulvey, M.A. (2002) Adhesion and entry of uropathogenicEscherichia coli. Cell Microbiol 4: 257–271.
Mulvey, M.A., Lopez-Boado, Y.S., Wilson, C.L., Roth, R.,Parks, W.C., Heuser, J., and Hultgren, S.J. (1998) Induc-tion and evasion of host defenses by type 1-piliated uro-pathogenic Escherichia coli. Science 282: 1494–1497.
Nagai, M., Meerloo, T., Takeda, T., and Farquhar, M.G.(2003) The adaptor protein ARH escorts megalin to andthrough endosomes. Mol Biol Cell 14: 4984–4996.
Nishimura, T., and Kaibuchi, K. (2007) Numb controls integrinendocytosis for directional cell migration with aPKC andPAR-3. Dev Cell 13: 15–28.
Orndorff, P.E., and Falkow, S. (1984) Organization andexpression of genes responsible for type 1 piliation inEscherichia coli. J Bacteriol 159: 736–744.
Pelkmans, L., Fava, E., Grabner, H., Hannus, M., Haber-mann, B., Krausz, E., and Zerial, M. (2005) Genome-wideanalysis of human kinases in clathrin- and caveolae/raft-mediated endocytosis. Nature 436: 78–86.
Phonphok, Y., and Rosenthal, K.S. (1991) Stabilization ofclathrin coated vesicles by amantadine, tromantadine andother hydrophobic amines. FEBS Lett 281: 188–190.
Piehl, M., Lehmann, C., Gumpert, A., Denizot, J.P.,Segretain, D., and Falk, M.M. (2007) Internalizationof large double-membrane intercellular vesicles by aclathrin-dependent endocytic process. Mol Biol Cell 18:337–347.
Pike, L.J., and Miller, J.M. (1998) Cholesterol depletiondelocalizes phosphatidylinositol bisphosphate and inhibitshormone-stimulated phosphatidylinositol turnover. J BiolChem 273: 22298–22304.
Pizarro-Cerda, J., Payrastre, B., Wang, Y.J., Veiga, E., Yin,H.L., and Cossart, P. (2007) Type II phosphatidylinositol4-kinases promote Listeria monocytogenes entry intotarget cells. Cell Microbiol 9: 2381–2390.
Razani, B., Engelman, J.A., Wang, X.B., Schubert, W.,Zhang, X.L., Marks, C.B., et al. (2001) Caveolin-1 null miceare viable but show evidence of hyperproliferativeand vascular abnormalities. J Biol Chem 276: 38121–38138.
Reddy, A., Caler, E.V., and Andrews, N.W. (2001) Plasmamembrane repair is mediated by Ca(2+)-regulated exocy-tosis of lysosomes. Cell 106: 157–169.
Rodriguez, A., Rioult, M.G., Ora, A., and Andrews, N.W.(1995) A trypanosome-soluble factor induces IP3 forma-tion, intracellular Ca2+ mobilization and microfilament rear-rangement in host cells. J Cell Biol 129: 1263–1273.
Rodriguez, A., Samoff, E., Rioult, M.G., Chung, A., andAndrews, N.W. (1996) Host cell invasion by trypanosomesrequires lysosomes and microtubule/kinesin-mediatedtransport. J Cell Biol 134: 349–362.
Rodriguez, A., Webster, P., Ortego, J., and Andrews, N.W.(1997) Lysosomes behave as Ca2+-regulated exocyticvesicles in fibroblasts and epithelial cells. J Cell Biol 137:93–104.
Schlegel, R., Dickson, R.B., Willingham, M.C., and Pastan,I.H. (1982) Amantadine and dansylcadaverine inhibitvesicular stomatitis virus uptake and receptor-mediatedendocytosis of alpha 2-macroglobulin. Proc Natl Acad SciUSA 79: 2291–2295.
Shen, Y., Naujokas, M., Park, M., and Ireton, K. (2000) InIB-dependent internalization of Listeria is mediated by the Metreceptor tyrosine kinase. Cell 103: 501–510.
Simons, K., and Toomre, D. (2000) Lipid rafts and signaltransduction. Nat Rev Mol Cell Biol 1: 31–39.
Song, J., Bishop, B.L., Li, G., Duncan, M.J., and Abraham,S.N. (2007) TLR4-initiated and cAMP-mediated abrogationof bacterial invasion of the bladder. Cell Host Microbe 1:287–298.
Song, K.S., Scherer, P.E., Tang, Z., Okamoto, T., Li, S.,Chafel, M., et al. (1996) Expression of caveolin-3 in skel-etal, cardiac, and smooth muscle cells. Caveolin-3 is acomponent of the sarcolemma and co-fractionates withdystrophin and dystrophin-associated glycoproteins. J BiolChem 271: 15160–15165.
Sorkin, A. (2004) Cargo recognition during clathrin-mediatedendocytosis: a team effort. Curr Opin Cell Biol 16: 392–399.
Stang, E., Kartenbeck, J., and Parton, R.G. (1997) Majorhistocompatibility complex class I molecules mediateassociation of SV40 with caveolae. Mol Biol Cell 8: 47–57.
Subtil, A., Gaidarov, I., Kobylarz, K., Lampson, M.A., Keen,J.H., and McGraw, T.E. (1999) Acute cholesterol depletioninhibits clathrin-coated pit budding. Proc Natl Acad SciUSA 96: 6775–6780.
Swanson, J.A. (2008) Shaping cups into phagosomes andmacropinosomes. Nat Rev Mol Cell Biol 9: 639–649.
Tardieux, I., Webster, P., Ravesloot, J., Boron, W., Lunn, J.A.,Heuser, J.E., and Andrews, N.W. (1992) Lysosome recruit-ment and fusion are early events required for trypanosomeinvasion of mammalian cells. Cell 71: 1117–1130.
Traub, L.M. (2003) Sorting it out: AP-2 and alternate clathrinadaptors in endocytic cargo selection. J Cell Biol 163:203–208.
Traub, L.M. (2005) Common principles in clathrin-mediatedsorting at the Golgi and the plasma membrane. BiochimBiophys Acta 1744: 415–437.
Ungewickell, E.J., and Hinrichsen, L. (2007) Endocytosis:clathrin-mediated membrane budding. Curr Opin Cell Biol19: 417–425.
Veiga, E., and Cossart, P. (2005) Listeria hijacks the clathrin-dependent endocytic machinery to invade mammaliancells. Nat Cell Biol 7: 894–900.
Veiga, E., Guttman, J.A., Bonazzi, M., Boucrot, E., Toledo-Arana, A., Lin, A.E., et al. (2007) Invasive and adherentbacterial pathogens co-Opt host clathrin for infection. CellHost Microbe 2: 340–351.
2566 D. S. Eto et al.
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Cellular Microbiology, 10, 2553–2567

Vieira, O.V., Botelho, R.J., and Grinstein, S. (2002) Phago-some maturation: aging gracefully. Biochem J 366: 689–704.
Wang, H., Liang, F.X., and Kong, X.P. (2008) Characteristicsof the phagocytic cup induced by uropathogenic Escheri-chia coli. J Histochem Cytochem 56: 597–604.
West, M.A., Bretscher, M.S., and Watts, C. (1989) Distinctendocytotic pathways in epidermal growth factor-stimulated human carcinoma A431 cells. J Cell Biol 109:2731–2739.
Williams, T.M., and Lisanti, M.P. (2004) The caveolin pro-teins. Genome Biol 5: 214.
Type 1 pili-mediated bacterial invasion via clathrin 2567
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Cellular Microbiology, 10, 2553–2567
Related Documents











