Mol Gen Genet (1995) 246:590-599 © Springer-Verlag 1995 Felix L6pez de Felipe • Christian Magni Diego de Mendoza • Paloma L6pez Citrate utilization gene cluster of the Lactococcus lactis biovar diacetylactis: organization and regulation of expression Received: 1 July 1994 / Accepted: 22 September 1994 Abstract The transport of citrate in Lactococcus lactis biovar diacetylactis is mediated by the citrate permease P. This polypeptide is encoded by the citP gene carried by plasmid pCIT264. In this report, we characterize the citP transcript, identify a cluster of two genes cotran- scribed with citP and describe their post-transcriptional regulation. The transcriptional promoter is located 1500 nucleotides upstream of the citP gene and the tran- scriptional terminator is positioned next to the Y-end of this gene. The DNA sequence was determined of the region upstream of the citP gene, including the pro- moter. Two partially overlapping open reading frames, citQ and citR were identified, which could encode polypeptides of 3.9 and 13 kDa respectively. These two genes, together with citP, constitute the cit cluster. Moreover, an IS-like element located between the cit promoter and the citQ open reading frame was identi- fied. This element includes an open reading frame ORF1, which could encode a 33 kDa polypeptide. A translational fusion between the citP and a cat reporter gene showed that translation ofcitR and citP is coupled, and regulated by CitR. The cit mRNA was subjected to specific cleavage after addition of rifampicin to the bac- terial cultures. We propose that expression of the cit cluster is controlled at the post-transcriptional level by mRNA processing at a putative complex secondary structure and by translational repression mediated by CitR. Key words citP gene - Citrate permease Lactococcus lactis. Gene expression. Transcription Communicated by B. J. Kilbey F.L. de Felipe • P. Ldpez ([~) Centro de Investigaciones Biol6gicas, C.S.I.C., Velfizquez, 144, 28006 Madrid, Spain C. Magni ' D. de Mendoza Departamento de Microbiologla, Facultad de Ciencias Bioquimicas y Farmacefiticas, Universidad Nacional de Rosario, Suipacha 531, 2000 Rosario, Argentina Introduction Citrate is abundant in the natural world being a con- stituent of all living cells. Most species of Enterobacteri- aceae are able to utilize citrate as a sole carbon source, as they possess a citrate transport system which medi- ates the uptake of this compound into the cells (Ishiguro and Sato 1985; Ishiguro et al. 1992). By contrast, the citrate-fermenting lactic acid bacterium designated Lac- tococcus lactis subsp, lactis biovar diacetylactis, requires in addition to citrate another source of metabolic ener- gy for the transport of citrate (reviewed by Hugenholtz 1993). The ability of L. lactis biovar diacetylactis to transport citrate is linked to an 8.0 kb plasmid, which appears to be present in all citrate-fermenting L. lactis strains analysed so far (Kempler and McKay 1981). The citrate permease P (CitP), encoded by the citP gene and carried on this plasmid, is the only essential requirement for the transport of citrate in this bacterium (Magni et al. 1994) and in the heterologous host Escherichia coli (David et al. 1990). The citP gene product was identified independently in two laboratories (Sesma et al. 1990; David et al. 1990). The amino acid sequence of CitP (David et al. 1990) is homologous to that of the sodium-dependent citrate permease S (CitS) of Klebsiella pneumoniae, which is in- duced by citrate (van der Rest et al. 1992). This homolo- gy would suggest that the CitP and CitS transport sys- tems might have the same ion requirements for citrate translocation, and might be regulated by the same effec- tor. However, citrate uptake appears to be sodium-inde- pendent in L. lactis biovar diacetylactis (van der Rest et al. 1992), and expression of the citP gene is not induced by citrate (Magni et al. 1994). The mechanisms which regulate the expression of CitP are currently unknown, although the existence of two citP mRNA species has been reported (Magni et. al. 1994). In this study, we have analysed the expression of the citP gene in L. lactis at the transcriptional and post-transcriptional level. We propose that expression is controlled by the CitR regu- latory polypeptide and by an mRNA processing mecha- nism.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Mol Gen Genet (1995) 246:590-599 © Springer-Verlag 1995
Felix L6pez de Felipe • Christian Magni Diego de Mendoza • Paloma L6pez
Citrate utilization gene cluster of the Lactococcus lactis biovar diacetylactis: organization and regulation of expression
Received: 1 July 1994 / Accepted: 22 September 1994
Abstract The transport of citrate in Lactococcus lactis biovar diacetylactis is mediated by the citrate permease P. This polypeptide is encoded by the citP gene carried by plasmid pCIT264. In this report, we characterize the citP transcript, identify a cluster of two genes cotran- scribed with citP and describe their post-transcriptional regulation. The transcriptional promoter is located 1500 nucleotides upstream of the citP gene and the tran- scriptional terminator is positioned next to the Y-end of this gene. The DNA sequence was determined of the region upstream of the citP gene, including the pro- moter. Two partially overlapping open reading frames, citQ and citR were identified, which could encode polypeptides of 3.9 and 13 kDa respectively. These two genes, together with citP, constitute the cit cluster. Moreover, an IS-like element located between the cit promoter and the citQ open reading frame was identi- fied. This element includes an open reading frame ORF1, which could encode a 33 kDa polypeptide. A translational fusion between the citP and a cat reporter gene showed that translation ofcitR and citP is coupled, and regulated by CitR. The cit mRNA was subjected to specific cleavage after addition of rifampicin to the bac- terial cultures. We propose that expression of the cit cluster is controlled at the post-transcriptional level by mRNA processing at a putative complex secondary structure and by translational repression mediated by CitR.
Key words citP gene - Citrate permease Lactococcus lactis. Gene expression. Transcription
Communicated by B. J. Kilbey
F.L. de Felipe • P. Ldpez ([~) Centro de Investigaciones Biol6gicas, C.S.I.C., Velfizquez, 144, 28006 Madrid, Spain
C. Magni ' D. de Mendoza Departamento de Microbiologla, Facultad de Ciencias Bioquimicas y Farmacefiticas, Universidad Nacional de Rosario, Suipacha 531, 2000 Rosario, Argentina
Introduction
Citrate is abundant in the natural world being a con- stituent of all living cells. Most species of Enterobacteri- aceae are able to utilize citrate as a sole carbon source, as they possess a citrate transport system which medi- ates the uptake of this compound into the cells (Ishiguro and Sato 1985; Ishiguro et al. 1992). By contrast, the citrate-fermenting lactic acid bacterium designated Lac- tococcus lactis subsp, lactis biovar diacetylactis, requires in addition to citrate another source of metabolic ener- gy for the transport of citrate (reviewed by Hugenholtz 1993). The ability of L. lactis biovar diacetylactis to transport citrate is linked to an 8.0 kb plasmid, which appears to be present in all citrate-fermenting L. lactis strains analysed so far (Kempler and McKay 1981). The citrate permease P (CitP), encoded by the citP gene and carried on this plasmid, is the only essential requirement for the transport of citrate in this bacterium (Magni et al. 1994) and in the heterologous host Escherichia coli (David et al. 1990). The citP gene product was identified independently in two laboratories (Sesma et al. 1990; David et al. 1990).
The amino acid sequence of CitP (David et al. 1990) is homologous to that of the sodium-dependent citrate permease S (CitS) of Klebsiella pneumoniae, which is in- duced by citrate (van der Rest et al. 1992). This homolo- gy would suggest that the CitP and CitS transport sys- tems might have the same ion requirements for citrate translocation, and might be regulated by the same effec- tor. However, citrate uptake appears to be sodium-inde- pendent in L. lactis biovar diacetylactis (van der Rest et al. 1992), and expression of the citP gene is not induced by citrate (Magni et al. 1994). The mechanisms which regulate the expression of CitP are currently unknown, although the existence of two citP mRNA species has been reported (Magni et. al. 1994). In this study, we have analysed the expression of the citP gene in L. lactis at the transcriptional and post-transcriptional level. We propose that expression is controlled by the CitR regu- latory polypeptide and by an mRNA processing mecha- nism.

Table 1 Bacterial strains and plasmids used in this work
Designation Characteristics/genotype Reference or source
591
Bacterial strains
L. lactis subsp, lactis MG1363 L. lactis biovar diacetylactis CRL264 Streptococcus pneumoniae 708 Escherichia coli MC1061
Bacillus subtilis MBI1
Plasmids pCIT264
pFS21
pFU5
pFU8
pC194 pLS1 pMS41
pFL10 pFL12
pFL13 pFL14
pFL15 pAlter-1 pET1 ld pCM6 pCM7
pIL253 pFI20
Lac, Cit, plasmid-free derivative of 712 Lac +, Pro +, Cit +, harboring pCIT264 end-l, exo-2, trt-1, hex-4, realM594 hsdR, mcrB, lacX74, galU, galK, rpsL, thi, araD A(araABC-leu) Iys-3 metB10 hisH2
Cit + plasmid from L. Iactis biovar diacetylactis Hybrid plasmid composed of pUC18 and pCIT264 pUC18 derivative containing the 3.5 kb EcoRI-BglII DNA fragment from pCIT264 pUC18 derivative containing the 2.0 kb EcoRV-EcoRV DNA fragment from pCIT264 Plasmid containing the cat gene Broad-host range multicopy plasmid Tc R pLS1 derivative containing the pneumococcal polA gene pC194 cat reporter vector based on pLS1 pLS1 derivative containing citQ, citR and citP-cat translational fusion all under the control of cit promoter pFL12 containing frameshift mutation in citR pLS1 derivative containing citR, and citP-cat translational fusion both under the control of polA promoter pFL14 containing frameshift mutation in citR E. coli phagemid cloning vector E. coil expression vector citR cloned into p E T l l d pCM6 containing polA promoter upstream of citR Gram-positive plasmid vector pIL253 derivative containing citR under the control of polA promoter
Gasson (1983) Sesma et al. (1990), Lacks (1970) M. Casadaban
Lacks et al. (1986)
Magni et al. (1994)
Sesma et al. (1990)
This work
Magni et al. (1994)
Horinouchi and Weisblum (1982) Lacks et al. (1986) Diaz et al. (1992)
Ldpez de Felipe et al. (1991) This work
This work This work
This work Promega Studier et al. (1989) This work This work
Simon and Chopin (1988) This work
Materials and methods Bacterial strains, growth media and plasmids
The bacterial strains used in this work are listed in Table 1. L. lactis strains were grown at 30 ° C without shaking, in M17 medi- um (Gasson 1983) supplemented with 0.5% glucose. The transfer of plasmids to L. lactis was performed by electroporation follow- ing the procedure of Dornan and Collins (1989). Streptococcus pneumoniae and Bacillus subtilis strains were grown and trans- formed as previously described (Martinez et al. 1987). The plas- mids used in this work are also listed in Table 1.
Construction of derivatives of plasmid pCIT264
The 3.5 kb EcoRI-BglII DNA fragment from pCIT264 was cloned into the EcoRI-BamHI site of pUC18 to construct pFU5.
An in-frame fusion of the citP gene and the pC194 cat gene was constructed as follows. An EcoRI site was introduced at the translational start codon of the citP gene by subcloning the filled- in 6.5 kb EcoRI-CtaI fragment from pCIT264 into the Smal site of the pAlter-1 vector. Site-directed mutagenesis was performed by using a 19mer oligonucleotide (5 ' -ATGCGGGTGAATTCAT- CAT-3') and a Promega kit. The region located upstream of the citP gene, including its transcriptional promoter, was released from the recombinant plasmid by digestion with EcoRI, and sub-
cloned into the unique EcoRI site (located at the translational start codon of the cat gene) of the reporter vector pFL10. The resulting plasmid, designated pFL12 (Fig. 1), was established in S. pneumoniae. This plasmid contains a mutated cat gene under the control of the transcriptional and translational start signals of citP, and encodes an altered chloramphenicol acetyltransferase with an extra amino acid, Ser3.
Plasmid pFI20 (Fig. 1) was constructed as follows. Firstly, the oligomers (5 ' -TTTAAACCATGGCGACAACAAAAA-Y) and (5 '-AAAGGATCCTGTCTTATATTCCTAG-3') were synthe- sized so that polymerase chain reaction (PCR) technology could be used to amplify a segment of DNA including the citR gene (coordinates 1220 to 1563 in Fig. 2). Oligomers were synthesized so that the 360 bp product contained a NcoI site overlapping the citR translational start codon and a BamHI site downstream of the citR gene. The PCR product was cloned into the NcoI-BamHI sites of pETl ld , giving rise to the pCM6 plasmid. This plasmid contains the citR gene under the control of the transcriptional and translational signals of the T7 gene 10. Synthesis of the CitR polypeptide encoded by the plasmid was verified with the E. coli BL21(DE3) host-vector expression system as previously described (Ports et al. 1991; results not shown). Secondly, the citR gene was placed under the control of the polA promoter as follows. The pCM6 plasmid was linearized by digestion with XbaI, and after filling-in the recessed ends, was blunt-end ligated to the filled-in 1.1 kb BglII-NheI fragment of pSM41, including the polA pro- moter. A plasmid called pCM7, carrying the promoter in the right

592
BgLll ATGc;tR SOcitP
EcoRI PLSDeitR X ~ /,/ATGcitpr.TcltP EcoRI pCIT26/~ -]
6RF P Tcctt Hind I11 EcoRI
pFLI2 ~ ''I "IF', ~,,~, .................. , ....... ,,,,,,,I
GATC cat' tetL '
PpotA SD~10 I I I , d II pFL1/-, ,\ \\\\\\\\,v~ .........................................
GATC I TI~7 II II pFL15 k\\~\\\\\\,~ illlll[ll H[IIIIIIIIIIi illll[lilllliH II
pF120 ...... J\\,JJ ~ , MLS"
Fig. 1 Physical map of pCIT264 and its derivatives. Genes are transcribed in the direction indicated by the arrows. The frameshift mutation present in plasmids pFL13 and pFL15 is indi- cated. The positions of the Shine-Dalgarno sequence (SD) and the translational start codons (ATG) of citR and citP are indicated. P1, cit promoter; Ppo~A, polA promoter; Tcitp, cit transcriptional terminator; Tea,, cat transcriptional terminator. Open boxes repre- sent DNA segments from pCIT264 lactococcal plasmid, closed boxes, pC194 cat gene, vertically shaded boxes, plasmid pLS1; left diagonal boxes, pneumococcal insert; right diagonal boxes, DNA segment from p E T l l d including Shine-Dalgarno sequence (SDqb~o) of T7 gene 10; single line, plasmid pIL253. Only relevant restriction sites are depicted
orientation, was established in E. coli. Third, the region contain- ing the polA promoter and the citR gene (flanked by the ribosomal binding site and the transcriptional terminator from the T7 gene 10) was released from pCM7 by digestion with EcoRV and cloned into the BamHI site of the gram-positive vector pIL253 by blunt- end ligation. The resulting plasmid pFI20 (Fig. 1) was cloned in B. subtilis.
Plasmid pFL14 (Fig. 1) was constructed as follows. Plasmid pFI20 was digested with BglII and the 1341 bp fragment, contain- ing the poIA promoter and the 5'-one-third of the citR gene, was purified. This fragment was ligated to the 972 bp BglII-HindIII fragment from pFL12, including the 3'-two-thirds of citR and the citP-cat fusion gene. The ligation mixture was digested with HindIII and after treatment with Klenow enzyme, the resulting DNA molecules were blunt-end ligated to pLS1 linearized with EcoRI and treated with Klenow. The resulting plasmid pFL14 was established in S. pneumoniae. This plasmid contains the citR and citP-cat genes under the control of the polA promoter and lacks the cit promoter and the putative mRNA processing site adjacent to the citR gene. To disrupt the citR gene, the plasmids pFL12 and pFL14 were linearized with BglII, and after filling-in the cohesive ends were recircularized by ligation. The resulting plasmids called pFL13 and pFL15, respectively (Fig. 1), were es- tablished in S. pneumoniae.
DNA sequencing and sequence analysis
The DNA nucleotide sequence of the region located upstream of the citP gene was determined by the dideoxy chain termination method of Sanger et al. (1977) with the T7 polymerase sequencing kit (Pharmacia) and [35S]dATP. Double stranded plasmids pFS21, pFU5 and pFU8 for templates were prepared as described by Chen and Seeburg (1985). The sequence of both strands of 1583 bp located upstream of citP gene (Fig. 2) was determined with 17mer oligonucleotide primers available commercially or synthesized in
a Gene Assembler (Pharmacia) at the Centro de Investigaciones Biol6gicas, Madrid, Spain. The search for open reading frames (ORFs) and for homologies of CitQ, CitR and ORF1 (see the Results) with the NBRF databank resources was performed using DNASTAR computer programs. Predictions of secondary struc- tures of mRNA were performed with the aid of the FOLD pro- gram (Zuker and Stiegler 1981) of the University of Wisconsin GCG package (Devereux et al. 1984).
RNA isolation and Northern hybridization
Total RNA was isolated from L. lactis strains grown in M17 medium as previously described (Magni et al. 1994). For the mRNA decay experiment, a culture grown to an absorbance value at 660 nm of 0.4 was treated with rifampicin at 200 gg/ml. Por- tions were then collected at the indicated times and the RNA was extracted as above. For Northern blotting, samples containing 7.5 gg of total RNA were prepared, electrophoresed on 1% agarose gel and blotted onto nitrocellulose filters following the procedure of Hirschorn et al. (1984). To visualize the specific mRNAs, the membranes were prehybridized for 5 h at 42 ° C and then incubated for 18 h with heat-denatured DNA fragments, la- belled in the noncoding strand with 3ap by primer extension. The probes used were: (1) for mRNA decay experiments, the Bg/II- BamHI fragment of pCIT264 encompassing the 5'-end of citP (coordinates - 129 to 621 of the gene according to David et al. 1990), or the 2.0 kb EcoRV-EcoRV fragment of pCIT264 encom- passing the distally located region of the P1 promoter of citP mRNA1 (located 1153 nucleotides upstream of the gene), and (2) for detection of the cit-cat mRNA, the BglI-Sau3A fragment from pC194 (coordinates 970 to 2001 according to Horinouchi and Weisblum 1982) containing the cat gene. For autoradiography, blotted filters were exposed to Kodak X-Omat S films. Quantita- tion of the nitrocellulose bands was performed with the Phosphor- Imager system (Molecular Dynamics).
Mapping of citP transcripts
Primer extension analysis was performed according to van Rooi- jen and de Vos (1990). One picomole of the 17mer oligonucleotide (5 ' -GAAATTAGAGATGATAC-3') complementary to mRNAI from nucleotides 234 to 218 (coordinates in Fig. 2) was annealed to 15 gg of RNA obtained from the L. lactis biovar diacetylactis CRL264 strain grown to an absorbance of 0.4 at 660 nm. Primer extension reactions were performed by incubation of the anneal- ing mixture with 20 U of M-MLV reverse transcriptase at 37 ° C for 30 min. Size determination of the reaction products was car- ried out in 8% polyacrylamide gel containing 7 M urea.
Nuclease SI mapping was performed essentially as previously described (Ldpez et al. 1989). DNA probes used for detection of the start site of transcription of mRNA1 were: (a) the 3.5 kb EcoRI-BglII DNA fragment from pCIT264, 32P-labelled at the 5' end generated by Bg/II, and (b) a 234 bp DNA fragment from pCIT264 amplified by PCR using the oligomers 5'-CCGCTTA- GATTTACTTG-3' (coordinates 1-17 in Fig. 2) and 5'-GAAA- TTAGAGATGATAC-3 ' 32P-labelled at the 5' end (complemen- tary to nucleotides 234 to 218, coordinates in Fig. 2). For detection of the termination site of transcription, the probe used was a 3181 bp BanI-EcoRI DNA fragment from pCIT264 32p-labelled by filling-in the T-recessed-end generated by BanI (coordinate 1279 of citP gene, according to David et al. 1990). Size detemina- tion of the Sl-protected fragments was carried out on 8% poly- acrylamide containing 7 M urea.
Determination of chloramphenicol acetyltransferase (CAT) activity
The extracts were prepared from cultures of L. lactis subsp, lactis MG1363 harbouring various plasmids, grown to an absorbance

of 0.4 at 660 nm. Cultures were sedimented by centrifugation, washed by suspension in buffer A (50 mM T~Is-HC1, 1 mM ED- TA, 0.1 mM phenylmethylsulphonyl fluoride, 1 gM dithiothreitol) centrifuged again, and concentrated 20 fold by suspension in buffer A. The L. lactis cells were disrupted by sonication. CAT activity was determined as described by Shaw (1975). One unit of enzymatic activity is defined as the amount of enzyme catalysing the acetylation of 1 nmol of chloramphenicol/min at 37 ° C.
Results
Nucleotide sequence of the region located upstream of citP
Transcription of the region upstream of citP generates two m R N A species (mRNA1 and mRNA2) beginning approximately 1500 and 500 nucleotides upstream from citP (Magni et al. 1994). In a search for transcriptional initiation sites, the sequence of 1583 nucleotides located upstream of the citP gene was determined. One putative promoter site, which could drive the synthesis of mRNA1 was observed and designated P1 (Fig. 2). In addition, two partially overlapping ORFs were ob- served, and designated citQ and ci~R. These two puta- tive genes could encode two polypeptides of 3876 and
Fig. 2 Nucleotide and deduced amino acid sequences of the 1583 nucleotide DNA region located upstream of the lactococcal citP gene. The asterisks at nucleotides 104 and 106 indicate the main transcriptional start sites for cit mRNA. The putative - 1 0 and - 3 5 hexamers in the cit promoter (P1) are overlined. The putative Shine-Dalgarno sequences (SD) for the open reading frames are underlined. Arrows indicate inverted repeats. The two 17mer shad- ed boxes indicate the inverted repeat flanking the 1 kb IS-like element
593
13 087 Da, respectively. The citR initiation codon (posi- tion 1220 in Fig. 2) is included in a potential secondary structure with a AG of -8 .7 kcal/mol (Tinoco et al. 1973). The short length of the citQ ORF indicates that this is not likely to be a protein coding sequence. How- ever, citQ is preceded by a putative ribosomal binding site (5 ' -UGAAACGAGGT-3 ~) complementary to the 3'-end of the 16S rRNA from L. lactis (3 ' -UCUUUC- CUCCA-5') (Ludwig et al. 1985). This ribosomal bind- ing site has a predicted free energy of binding (AG) to the 3'-end of the lactococcal 16S rRNA of -9 .7 kcal/ mol, according to the rules of Schurr et al. (1993). On this basis, it is expected that the lactococcal ribosomes should be able to translate citQ. When the predicted amino acid sequences of citQ and citR were used to search the NBRF database, no significant homology was detected with any of the proteins contained in the databank.
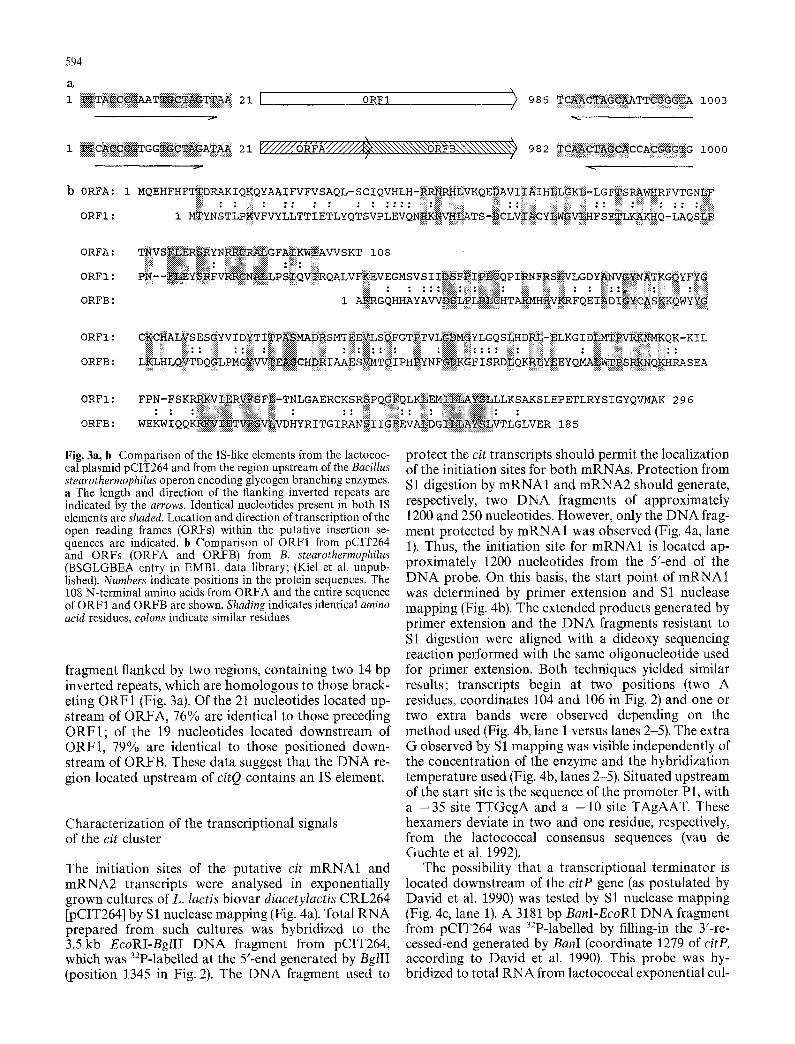
The analysis of the DNA sequence located upstream of citQ revealed the presence of two identical inverted repeats (coordinates 107-123 and 1091-1107 in Fig. 2). Furthermore, these sequences bracket an open reading frame of 296 codons (designated ORF1) located in the opposite orientation to citP. A search of the NBRF databank revealed homology of ORF1 to two overlap- ping ORFs (ORFA and ORFB) of a putative IS element located upstream of the g/gC gene from Bacillus stearothermophilus (Fig. 3b). Of the 108 amino acids in the amino-terminal segment of ORFA, 27% are identi- cal and another 24% are similar to the first 107 amino acids of the ORF1 product. The carboxy-terminal re- gion of ORFI shows 35% identity and 16% similarity to the 185 amino acids of ORFB. Moreover, the B. stearothermophilus ORFs are located in a 960 bp DNA
-35 P1 -10 . . ~CGCTTAGATTTACTTGCT~TCAGTATGT~TTTAAAATA~CACAATGTT~AAAATACGT~AGcTTATTATTTGCGAAAC.AAAACAGAAA~GAATAGAAT~AAAAT~~GATTATTTAGCCATGACTTGATACcCG 150
EndLysAlaMetVBtGtnTyrGlyl
ATAGAATATCTTAAAGTCTCTGGTTCCAGTGATTTAGCTGATTTTAACAGTAAAGAATACGCTAAAAGTATCAT~TCTAATTTCAATTGAAAACCTTGAGGCGAACGA~TTTTACAACGcTCAGCTCCTAGATTTGTCAAAAAAGAGAAA 300 TATCTTATAGAATTTCAGAGAcCAAGGT~ACTAAATCGACTAAAATTGTCATTTCTTATGCGATTTTCATAGTAGAGATTAAAGTTAA~TTTTGGAACTcCGCTTGCTGAAAATGTTGcGAGT~GAGGATCTAAACAGTTTTTTCTCTTT [e~erTyrArgLeuThrG~u~r~Glu~eu~erLysALa$erL~sLeuLeuLeu~erT~rA~aLeu~euI~eMetGLuLeuL~sLeuGLnPheGLyGlnPr~erArg~erLysC~sArgG~uAIaGL~LeuAsnThrLeuPheSerPhev
A~TCG`CTCAATCACTTTTCTACGTTTTGAAAAATTAGGGAAAAGGATTTTC~TTT~TTCATGTTCTTCCTGACAGGTGTCATTAGAT~AATTcCTTTTAATTcCAGC~TATCATGCAGTGACTGA~CTAGATATCCCATATCTCCAAGG 450 TGAGCGAGTTAGTGAAAAGATGCAAAACTTTTTAATCCCTTTTCCTA/U~AG/UU~CGAAGTACAAGAAGGACTGTCCACAGTAATCTAGTTAAGGAAAATTAAGGTCGGATAGTA~GTCACTGACTGGATCTATACCGTATAGAGGTTCC a~ArgG~uI~eVa~LysArgArgLys~erPheAsnPr~PheLeuI~eLysLysG~nLysMetAsn~ysArgValPr~ThrMetLeuAspILeGLyLysLeuGLu~euArgAspH~sLeu~erG~nGL~LeuTyrGlYMetAspG~y~euV
ACTGTTGGTGTcCCAAATTGACTCAACACTT0CTCGGTcATTGAACGATCAGCCATTGAAGCAGGAGTAATTGTGTAGTCTATGACATAGCCTGATTCACTGACTAAAGCATGAcATTTACAGCcATAGAAGTACTGTCCCTTTGTAGCA 600 TGACAACCACAGGGTTTAACTGAGTTGTGAAGGAGCcAGTAACTTGcTAGTCGGTAACTT•GTCCTCATTAACACATCAGATACTGTATCGGACTAAGTGACTGATTTCGTACTGTAAATGTCGGTATCTTCATGAcAGGGAAACATCGT a~ThrPr~ThrG~yPheG~nSerLeuVa~GLuG~uThrMet~erArgAspA~aMet$erALaPr~Thr~[eThrTyrAspI~eVa~TyrG~ySerGtu$er~eLLeuAlaH~sCysLysCysG~yTyrPheTyrG~nG~yLysThrA(aA
TTGTAGcCAA~ATTTGCATAATCT~CAAGAA~TTTGCTTCTGAAATTACGAATAGGCTGACACAAA~GAATGGGGAAGCTGTCAATAATGGATACACTCATTCCTTCAACCTCTTTAAAGA~GAGTGCTTGGCGAATGACTTGGATACTC 750 AA~ATCG~TTGTAAACGTATTAGAGGTTCTTGAAACGAAGACTTTAATGCTTATCCGACTGTGTTTCCTTACCCCTTCGACAGTTATTACCTATGTGAGTAAGGAAGTTGGAGAAATTTCTGCTCACGAA~CGCTTAC1GAA~CTAT~AG snTyrGLyV~AsnA~TyrAsp~tyLeuVa~s~erArgPhe^snArgI~ePr~G~nCysLeuPr~]LePr~Phe~erAspI~e~[e~erVaLSerMetG~yG~uVaLG~uLysPheV~LeuA~eG~nArgI~eV~LG(nI~eSerP
GGTAAGAGGGCATTACAA~GG~GGACAAAGCGAGAATATTcTAGGAAATTAGGAAATAAACTTTGAGC~AATTGGTGCTTAGCTTTAAGCGTTTCACTAAAATGCAGTACGCcCCATAGGTAACAAGCGATAACTAAACAGT~AGATGTT 900 CCATTC~CCCGTAATGTTGCCGCCTGTTTCGCTCTTATAAGATCCTTTAATCCTTTATTTGAAACTCGGTTAACCA~GAATCGAAATTCGCAAAGTGATTTTACGTCATGCGGGGTATCcATTGTTCGCTATTGATTTGTCAGTCTACAA r~LeuLeuA~aAsnC~sArgArgVaL~heArg~erT~rGLu~euPheAsnPr~PheLeu~erG~nALaLeuGLnH~sLYsA~aLysLeuThrG~u~erPheHisLeuVaLG~TrpLeuTyrCysA~aI[eVaLLeuCysAspSerThrA
GCAAGATGGACGTT~TTTcGGTTTTGAA~CTCAAGGGG~A~ACTCGTTTG~TM~AGCGTCT~ATGGTTGTCAGTAAAT~AA~AAAAA~TTTTGGAAGTGTGCTATTATAAGTCATATAAGTCGTGAGCTTTCTAATGCTTAGTG~TTTA 1050 ~GTTcTAC~TG~AAGAAAGC~CTTGGAGTTCCccTTGTGAG~AAACTATTTCGCAGAGTTACCAACAGTCATTTATTTGTTTTTGAAAACCTTCACACGATAATATT~AGTATATTCAGCA~TCGAAAGATTACGAATCACGAAAT ~a~euHisVaIAsnLysArgAsnGtn~atG~uLeuPr~Va(SerThrGlnTyrLe~ThrGLuIIeThrThrLeuLeuT~rVaLPheVaLLysPr~euThr~erAsnTyrThrMet <<ORF1 SD
SO AGATTAGGATAGcACGACTTATTTATTTTCcAATGAAATT~i~AAATATCAAGTAcAGAAGTCAGCGAcCGAGTGAAACGAGGTCAATGcGCACTTAcACCCCA~cAAATAATT~TGGGGTAAATTGTGCTAAAA 1200 TCTAATCCTATCGTGCTGAATAAATAAAAGGTTACTTTA~~ ~ > MetArgThrTyrThrProProAsnAsnCysG[yValAsnCysAlakysT
SD cCTGTATCAGAAGTcGC~TAT~CGAcAACAAAAAcCAAAAAcAAAACCCTAAAAGAAAGGAGACAACAACATGAAAGTCATTT~TTTGATGCAACATTAAACGCGCCTTTGTTGGCGAATCATGCGCTGCCAGATTGGAGACAAACGCT 1350 hrCysIleArgSerArgLeuTrpArgGlnGlnLysProLysThrLysProEnd
CitR>> MetA~aThrThr~ysThr~ysAsnL~sThrkeu~sGLuAr~ArgGlnGLnHisG~u~erH~sPhePheAspA~aThrLeuAsnA[@r~LeuLeuALaA~nHisA~aLeuPr~AspTr~ArgGLnThrLe CGCAGATCTTTCTATCCCACAATTAATAGTGACTGGAGAACGGTCTCCAATCTGGAAGCATGATTTTTGTCGCTATTGTGAAAAAGTTATCAATGAAAAATCTCAGGTCGTCACCATTAATCAGGTTGGTCACATGCCACAGATAGAAGT 1500 uAtaAspLeu~er~LePr~GLnLeuILeVaLThrGLYGLuAr~SerPr~ILeTrpL~sHisAspPheCysAr~TYrCYsGLuLYsVaL]LeAsnGLuLysSerGLnVaLVaLThr~LeAsnGLnVaLGLyH~MetPr~G~n~LeG~u~a
SD ACCTGAAAAATTTAATGAAA•AACATGGAATTTTATTCAACACAAACTAGGAATATAAGACAGATATAAATGGAGATAGAATTATG 1586 [ProGtuLysPheAsnGLulteThrTrpAsnPheILeGLnHisLysLeuGlyIleEnd -- -- CitP>> Met
594
8.
i ~ T ~ g ~ T ~ T ~ 211 0RFI > 985 ~ ~ T T ~ A 1003
:.x.x.x.: :::::::::::::::::::::::::::::::::::::: :~.x.x.:~::.x.:.:+:.:+:,x.x ::::::::::::::::::::::: / / ooo
b 0RFA: 1 MQEHFHFT~DRAKIQ~QYAAIFVFVSAQL-SCIQVHLH-~VKQED)WI~I~L~K~-LGF~IS~WHRFVTGN~ i!i!i::i ~ • • ~: ' : ~ ~ - - - ~ ~ ' : ~ i i i ~ i . . . . . . . . . . . i!i~::~.:~::::iil iii::ii!::{::iii::ii :i'!:,ii . . ::!iiii::~!ill ~i?::~ ::iiiii! ::!ii::i:~i . . i::!ii .::::i::;:~i ~i~i::i:: . . . . ::i::iil;~i~i
ORFA:
ORFI:
ORFB:
~ s ~ ~ ~ ! a v v s ~ ~.o i~iii! $iiiiiNiiiii ~.iii!!!i!: !~liIIIiiiiiiiii ii!iiii!!!I! :i!i!~!!: iiiii?.
ORFI:
ORFB:
ORFI:
ORFB:
FPN-FSK~i~F~-TNT,GA~RCKS~PQ~QL~.M~N~T,U<Sa~S~EPETLaYS I G Y Q ~ 296
Fig. 3a, b Comparison of the IS-like elements from the lactococ- cal plasmid pCIT264 and from the region upstream of the Bacillus stearothermophilus operon encoding glycogen branching enzymes. a The length and direction of the flanking inverted repeats are indicated by the arrows. Identical nucleotides present in both IS elements are shaded. Location and direction of transcription of the open reading frames (ORFs) within the putative insertion se- quences are indicated, b Comparison of ORF1 from pCIT264 and ORFs (ORFA and ORFB) from B. stearothermophilus (BSGLGBEA entry in EMBL data library; (Kiel et al. unpub- lished). Numbers indicate positions in the protein sequences. The 108 N-terminal amino acids from ORFA and the entire sequence of ORF1 and ORFB are shown. Shading indicates identical amino acid residues, colons indicate similar residues
fragment flanked by two regions, containing two 14 bp inverted repeats, which are homologous to those brack- eting ORF1 (Fig. 3a). Of the 21 nucleotides located up- stream of ORFA, 76% are identical to those preceding ORF1; of the 19 nucleotides located downstream of ORF1, 79% are identical to those positioned down- stream of ORFB. These data suggest that the D NA re- gion located upstream of citQ contains an IS element.
Characterization of the transcriptional signals of the cit cluster
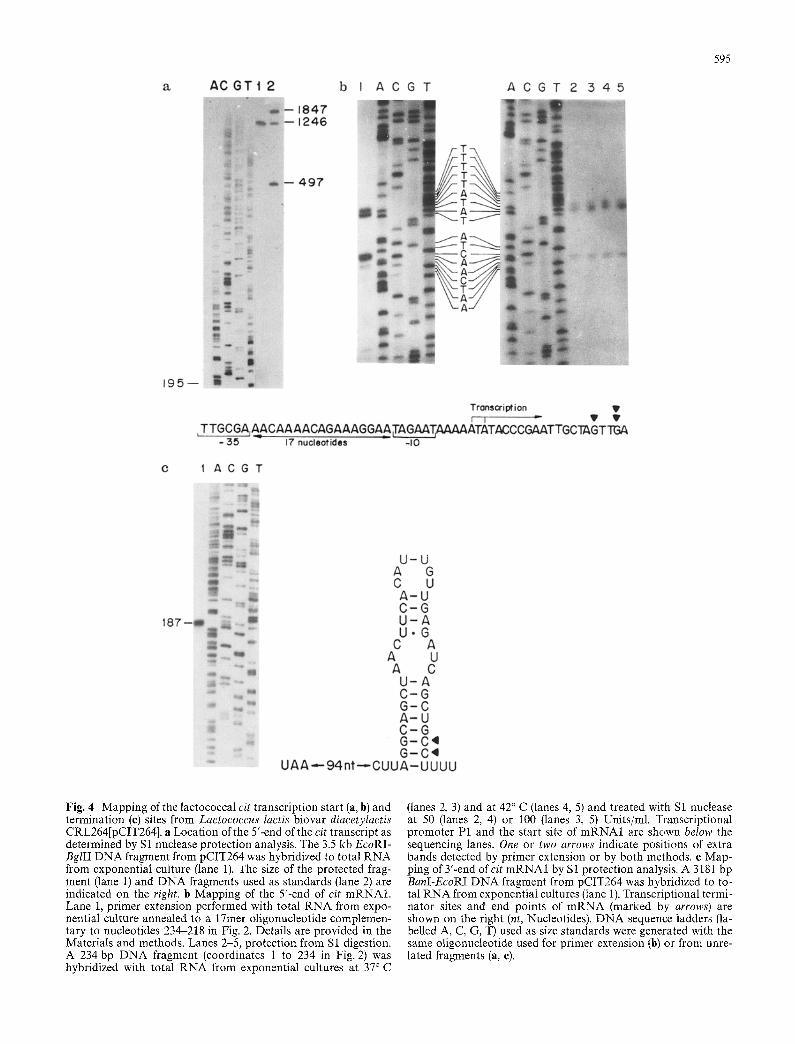
The initiation sites of the putative cit mRNA1 and mRNA2 transcripts were analysed in exponentially grown cultures of L. lactis biovar diacetylactis CRL264 [pCIT264] by S 1 nuclease mapping (Fig. 4a). Total R NA prepared from such cultures was hybridized to the 3.5 kb EcoRI-BgIII DNA fragment from pCIT264, which was 32p-labelled at the 5'-end generated by Bg/II (position 1345 in Fig. 2). The DNA fragment used to
protect the cit transcripts should permit the localization of the initiation sites for both mRNAs. Protection from S1 digestion by mRNA1 and mRNA2 should generate, respectively, two DNA fragments of approximately 1200 and 250 nucleotides. However, only the DN A frag- ment protected by mRNA1 was observed (Fig. 4a, lane 1). Thus, the initiation site for mRNA1 is located ap- proximately 1200 nucleotides from the 5'-end of the DNA probe. On this basis, the start point of mRNA1 was determined by primer extension and S1 nuclease mapping (Fig. 4b). The extended products generated by primer extension and the DNA fragments resistant to S1 digestion were aligned with a dideoxy sequencing reaction performed with the same oligonucleotide used for primer extension. Both techniques yielded similar results; transcripts begin at two positions (two A residues, coordinates 104 and 106 in Fig. 2) and one or two extra bands were observed depending on the method used (Fig. 4b, lane 1 versus lanes 2-5). The extra G observed by S 1 mapping was visible independently of the concentration of the enzyme and the hybridization temperature used (Fig. 4b, lanes 2-5). Situated upstream of the start site is the sequence of the promoter P1, with a - 3 5 site TTGcgA and a - 1 0 site TAgAAT. These hexamers deviate in two and one residue, respectively, from the lactococcal consensus sequences (van de Guchte et al. 1992).
The possibility that a transcriptional terminator is located downstream of the citP gene (as postulated by David et al. 1990) was tested by S1 nuclease mapping (Fig. 4c, lane 1). A 3181 bp BanI-EcoRI DNA fragment from pCIT264 was riP-labelled by filling-in the 3'-re- cessed-end generated by BanI (coordinate 1279 of citP, according to David et al. 1990). This probe was hy- bridized to total R NA from lactococcal exponential cul-
El,
195 - -
AC G T I b A C G T A C G T 2 5 4 5
1 8 4 7 1246
4 9 7
13
Tronscriplion • l ] • V
, T TGC GAI~AC AA AACAGAAAGGAA ~AGAATI/k&&AATATACCCGAATTGCTAGT T ~ - 35 17 nucleotide$ - I0
I A C G T
595
187
U - U A G ¢ U
A - U C - G U - A U . G
~ C A • o ~ : A U
~ ~ C - G . . . . . G- c
A - U ;i~ C - G
,,,~ G- C 4
.... U A A " - 94nt - - - -CUUA- U UUU
Fig. 4 Mapping of the lactococcal cit transcription start (a, b) and termination (e) sites from Lactococcus Iactis biovar diacetylactis CRL264[pCIT264]. a Location of the 5'-end of the cit transcript as determined by Sl nuclease protection analysis. The 3.5 kb EcoRI- BglII DNA fragment from pCIT264 was hybridized to total RNA from exponential culture (lane 1). The size of the protected frag- ment (lane 1) and DNA fragments used as standards (lane 2) are indicated on the right, b Mapping of the 5'-end of cit mRNA1. Lane 1, primer extension performed with total RNA from expo- nential culture annealed to a 17met oligonucleotide complemen- tary to nucleotides 234-218 in Fig. 2. Details are provided in the Materials and methods. Lanes 2-5, protection from S1 digestion. A 234 bp DNA fragment (coordinates 1 to 234 in Fig. 2) was hybridized with total RNA from exponential cultures at 37°C
(lanes 2, 3) and at 42 ° C (lanes 4, 5) and treated with S1 nuclease at 50 (lanes 2, 4) or 100 (lanes 3, 5) Units/ml. Transcriptional promoter P1 and the start site of mRNA1 are shown below the sequencing lanes. One or two arrows indicate positions of extra bands detected by primer extension or by both methods, e Map- ping of 3'-end of cit mRNA1 by S1 protection analysis. A 3181 bp BanI-EcoRI DNA fragment from pCIT264 was hybridized to to- tal RNA from exponential cultures (lane 1). Transcriptional termi- nator sites and end points of mRNA (marked by arrows) are shown on the right (nt, Nucteotides). DNA sequence ladders (la- belled A, C, G, T) used as size standards were generated with the same oligonucleotide used for primer extension (b) or from unre- lated fragments (a, e).
596
ture and hybrids were treated with nuclease $1. Two protected fragments of 186 and 187 nucleotides were detected (Fig. 4c, lane 1). Thus, the termination site of the transcript was placed near the 3'-end of a potential hairpin in the m R N A located 98 nucleotides of the UAA translation termination codon of the eitP gene.
Processing of cit mRNA1
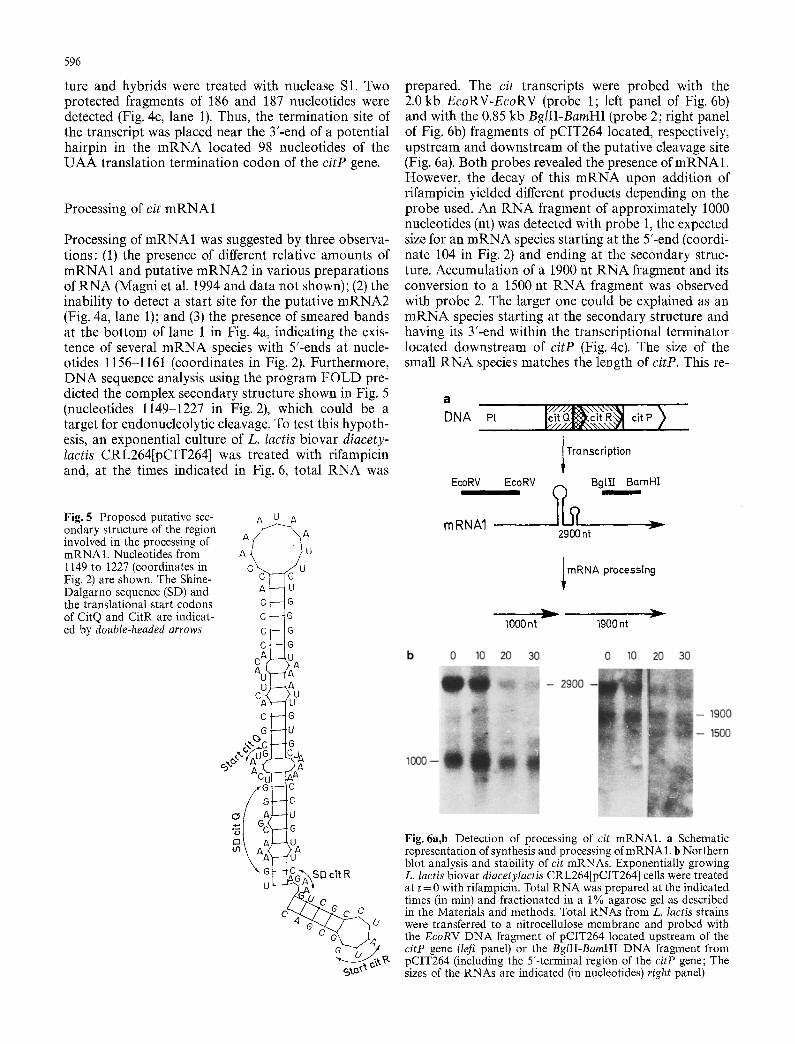
Processing of mRNA1 was suggested by three observa- tions: (1) the presence of different relative amounts of mRNA1 and putative mRNA2 in various preparations of RNA (Magni et al. 1994 and data not shown); (2) the inability to detect a start site for the putative mRNA2 (Fig. 4a, lane 1); and (3) the presence of smeared bands at the bot tom of lane 1 in Fig. 4a, indicating the exis- tence of several m R N A species with 5'-ends at nucle- otides 1156-1161 (coordinates in Fig. 2). Furthermore, DNA sequence analysis using the program FOLD pre- dicted the complex secondary structure shown in Fig. 5 (nucleotides 1149-1227 in Fig. 2), which could be a target for endonucleolytic cleavage. To test this hypoth- esis, an exponential culture of L. lactis biovar diacety- lactis CRL264[pCIT264] was treated with rifampicin and, at the times indicated in Fig. 6, total RNA was
Fig. 5 Proposed putative sec- A ondary structure of the region j r involved in the processing of A mRNA1. Nucleotides from A 1149 to 1227 (coordinates in ( Fig. 2) are shown. The Shine- c Dalgarno sequence (SD) and the translational start codons of CitQ and CitR are indicat- ed by double-headed arrows
" G A U C G C G C G C G A U
u ,u A
C" G G' U
~e, .G
A lu
O 4 - '
SD cit R
c *'-.o
5&a~ t
prepared. The cit transcripts were probed with the 2.0 kb EcoRV-EcoRV (probe 1 ; left panel of Fig. 6b) and with the 0.85 kb BgIII-BamHI (probe 2; right panel of Fig. 6b) fragments of pCIT264 located, respectively, upstream and downstream of the putative cleavage site (Fig. 6a). Both probes revealed the presence of mRNA1. However, the decay of this m R N A upon addition of rifampicin yielded different products depending on the probe used. An RNA fragment of approximately 1000 nucleotides (nt) was detected with probe 1, the expected size for an m R N A species starting at the 5'-end (coordi- nate 104 in Fig. 2) and ending at the secondary struc- ture. Accumulation of a 1900 nt RNA fragment and its conversion to a 1500 nt RNA fragment was observed with probe 2. The larger one could be explained as an m R N A species starting at the secondary structure and having its 3'-end within the transcriptional terminator located downstream of citP (Fig. 4c). The size of the small RNA species matches the length of citP. This re-
a
DNA Pt
EcoRV EcoRV
m RNA1
V////4 N ~\" -," ,,\\ ,x ,~ ] \ Nit O]~t, cit R ~ t i t P ) [ / ( ' / / { \ \ \ \ ' ~ ¢ I /
I Tronseription
BgtTf BarnHI
2900 nt
I mRNA processing
y F
1000 nt 1900 nt
b 0 10 20 30 0 10 20 30
1000 -
- 2900
1900
1500
Fig. 6a,b Detection of processing of cit mRNA1, a Schematic representation of synthesis and processing of m R N A 1. b Northern blot analysis and stability of cit mRNAs. Exponentially growing L. lactis biovar diacetylactis CRL264[pCIT264] cells were treated at t = 0 with rifampicin. Total RNA was prepared at the indicated times (in min) and fractionated in a 1% agarose gel as described in the Materials and methods, Total RNAs from L. lactis strains were transferred to a nitrocellulose membrane and probed with the EcoRV DNA fragment of pCIT264 located upstream of the citP gene (left panel) or the BglII-BamHI DNA fragment from pCIT264 (including the 5'-terminal region of the citP gene; The sizes of the RNAs are indicated (in nucleotides) right panel)

sult could indicate that after the initial processing of the mRNA1 into two pieces the Y-end fragment is subject- ed to further nucleolytic cleavage.
Role of CitR on the expression of the cit gene cluster
We have previously shown by deletion analysis that CitP is the only plasmid-encoded protein required for the transport of citrate in L. lactis (Magni et al. 1994). This result excluded an involvement of CitR in the up- take of citrate, and suggested to us the possibility of a regulatory function for CitR in citP expression. To ex- plore this possibility, at the transcriptional and post- transcriptional levels, plasmids pFL12 and pFI20 were constructed and transferred to the L. lactis MG1363 strain, pFL12 contains the citP gene fused to the pC194 cat reporter gene under the control of promoter P1 (Fig. 1) and plasmid pFI20 encodes CitR transcribed from the polA promoter (Fig. 1). L. lactis MG1363 har- bouring pFL12 synthesizes the expected cit-cat m R N A 2200 nt in length, as revealed by Northern hybridization (Fig. 7). The presence of plasmid pFI20 did not affect either the copy number ofpFL12 (eight copies/cell, data not shown) nor the synthesis of the cit-cat mRNA en- coded by pFLI2 (Fig. 7). However, overexpression of CitR produces a fourfold decrease in CAT activity en- coded by pFL12, indicating an inhibitory trans effect, at the post-transcriptional level, of CitR on CitP synthesis (Fig. 7). To detect possible cis effects of citR on citP ex- pression, plasmid pFL13 (Fig. 1) was generated from pFL12, by introduction of a frameshift mutation in citR.
O
e ~
v..q w=4
Plasmids CAT activity (Ulmg protein)
pFL12 1408 + 175
pFL12, pFI20 356 +49
Fig. 7 Influence of CitR on expression of the citP-cat fusion. L. lactis MG1363 harbouring the plasmids indicated was grown to an absorbance of 0.4 at 660 nm. Total RNA from these cultures was prepared and analysed as described in Fig. 6. The citP-cat mRNA was revealed by hybridization with a cat probe as de- scribed in the Materials and methods. Crude extracts were pre- pared from the aforementioned cultures and the chloramphenicol acetyltransferase (CAT) activities determined; each value is the average of at least three independent experiments
597
Table 2 Chloramphenicol acetyltransferase (CAT) activity direct- ed by citP-cat translational fusions
Plasmids CAT activity a (U/rag protein)
pFL10 < 1 pFL13 665_+ 37 pFL13, pFI20 661 _+ 55 pFL14 759 + 50 pFL14, pFI20 716_+43 pFL15 383_+ 12
L. lactis MG1363 harbouring the plasmids indicated was grown to an absorbance of 0.4 at 660 nm, and the CAT activities were determined as described in the Materials and methods. Each value is the average of at least three independent determinations
Interruption of citR in pFL13 resulted in a twofold de- crease in CAT activity compared to the level produced by the parental plasmid pFL12 (Fig. 7; TaNe 2), and d id not affect the synthesis of cit-cat m R N A (data not shown). This result indicates a coupling of the transla- tion of citR and citP, which enhances the synthesis of CitP. Moreover, the presence of pFI20 did not signifi- cantly affect the CAT activity encoded by pFL13 (Table 2), suggesting that the repression exerted by CitR sup- plied in trans requires coupled translation of citR and citP.
To test this hypothesis further, plasmids pFL14 and pFL15 were constructed. Both plasmids contain the citP-cat fusion under the control of the potA promoter (Fig. 1), but pFL15 also contains a frameshift mutation in citR. Comparison of the CAT activity encoded by cells bearing pFL14 or pFL15 (Table 2) further con- firms, a cis effect of citR translation on CitP synthesis, which is independent of the transcriptional promoter used. Furthermore, as the trans effect of CitR was not observed to act on the CAT activity encoded by pFL14 (Table 2), it seems likely that the target of CitR is no longer present in this plasmid.
Discussion
The results presented in this paper provide evidence that the lactococcal plasmid pCIT264 contains a cluster of genes involved in the transport of citrate in L. lactis. Transcription of the cit region from the lactococcal plas- mid produces the mRNA1 transcript of 21900 nt. The 5'- and 3'-ends of this m R N A were mapped precisely, and found to include the citR and citP genes and the putative citQ open reading frame. Transcription begins at promoter P1 located 1 kb upstream of the citQ gene and terminates at a hairpin structure typical of prokary- otic terminators (Rosenberg and Court 1979). The long leader sequence present in mRNA1 presumably origi- nated from a transposition event, resulting in the intro- duction of an IS-like sequence between the P1 promoter and the beginning of the citQ gene.

598
The transcription of the cit gene cluster is constitu- tive and is not induced by citrate (Magni et al. 1994); however, production of CitR and CitP seems to be con- trolled at the post-transcriptional level. Our results show that translation of the citP gene is enhanced by translation of the upstream citR gene. One may assume that ribosomes, after finishing translation of the citR gene, can bind to the Shine-Dalgarno sequence of citP, resulting in a higher efficiency of its utilization and, as a consequence, synthesis of higher levels of CitP.
The question therefore arises of how the levels of CitR are controlled in the cells. Our results indicate that specific endonucleolytic cleavage of mRNA1 generates a 1100 nt 5'-RNA untranslated region, and a 1800 nt 3'-RNA, which could encode CitR and CitP. The region surrounding the processing site is included in a complex secondary structure with a predicted free energy of -31.2 kcal/mol (Fig. 5), containing a large loop of 9 nucleotides (coordinates 1149 to 1227 in Fig. 2). This kind of structure has been characterized as a target for endonucleolytic cleavage in other bacterial systems (Gamper et al. 1992; Lesage et al. 1992; Liu et al., 1992). It is tempting to assume that the mRNA secondary structure and processing of the cit mRNA are involved in the post-transcriptional control of citR expression. The AUG start codon of CitR is positioned in the loop of a putative hairpin structure located at the 3'-end of the secondary structure (coordinates 1213 to 1227 in Fig. 2); also the putative ribosomal binding site of CitR (AGAAG), complementary to the 3'-end of L. lactis 16SrRNA (Ludwig et al. 1985), is partially blocked in the two stems of the structure. Formation of the sec- ondary structure on intact mRNA could interfere with the translation of citR, since these kinds of structures appear to influence the efficiency of translation initia- tion in L. lactis (reviewed by van de Guchte et al. 1992). In addition, we can envisage competition between the ribosomes translating the putative citQ open reading frame and the ribonucleases processing the mRNA1, as proposed for the lac operon of E. coli (Yarchuk et al. 1992). Translation of citQ from an intact mRNA1 could inhibit the formation of the secondary structure and subsequent processing of this mRNA, and thus could favour the expression of citR by coupled translation or by ribosomal frameshifting. This type of translational regulatory mechanism is functional in other bacterial systems, for example, the Staphylococcus aureus ermC gene (Mayford and Weisblum 1989) and the E. coli oper- on encoding translational initiation factor IF3 and the ribosomal proteins L35 and L20 (Lesage et al. 1992).
What is the role of CitR? This polypeptide seems negatively to regulate expression of citP. Our results suggest that the repression is exerted at the post-tran- scriptional level and that the target of this repressor is not included within the coding sequence of citR and citP. The operator sequence for CitR could be the puta- tive secondary structure that includes the translational start codon of citR. The binding of the repressor to its operator should stabilize the secondary structure and
will block initiation of translation of citR (a common mechanism for translational repressors; see review by McCarthy and Gualerzi 1990). Therefore, the coupled translation of citR and citP will not take place, and thus, synthesis of the CitP polypeptide will decrease. In sum- mary, we report a complex post-transcriptional regula- tion of expression of a citrate permease gene mediated by a regulatory protein in bacteria. Since citrate is not an inducer of the expression of the cit operon (Magni et al. 1994), the environmental conditions responsible for the control of CitP levels in the cell remain obscure at present.
Acknowledgements We thank M. Espinosa for helpful discussions and critical reading of the manuscript and J. Sarraseca and M.A. Corrales for technical assistance. Research at the Centro de Inves- tigaciones Bioldgicas was under the auspices of the Consejo Supe- rior de Investigaciones Cientificas (CSIC), Spain and was support- ed by the Comisi6n Interministerial de Ciencia y Tecnologia grants BIO091-0691 and BIO094-1029. This work and F.L.F. were supported by the Commision of the European Communities grant CI1".0772. C.M. is a fellow from the Consejo Nacional de Investigaciones Cientificas y T6cnicas de la Republica Argentina (CONICET).
References
Chen EJ, Seeburg PH (1985) Supercoil sequencing a fast method for sequencing DNA plasmid. DNA 4:165-170
David S, van der Rest ME, Driessen AJM, Simmons G, de Vos WM (1990) Nucleotide sequence and expression in Escherichia coli of the Lactococcus lactis citrate permease gene. J Bacteriol 172:5789-5794
Devereux J, Haeverli P, Smithies O (1984) A comprehensive set of programs for the Vax. Nucleic Acids Res 12:387-395
Diaz A, Pons ME, Lacks SA, L6pez P (1992) Streptococcus pneu- moniae DNA polymerase I lacks 3'-to-5' exonuclease activity: localization of the 5'-to-3' exonucleolytic domain. J Bacteriol 174:2014-2024
Dornan S, Collins MA (1990) High efficiency electroporation of L. lactis susp. lactis LM0230. (Letters) Appl Microbiol 11:62-64
Gamper M, Ganter B, Polito MA, Haas D (1992) RNA processing modulates the expression of the arcDABC operon in Pseu- domonas aeruginosa. J Mol Biol 226:943-957
Gasson MJ (1983) Plasmid complements o f Streptococcus lactis NCDO 712 and other streptococci after protoplast-induced curing. J Bacteriol 154:1-9
Hirschorn RR, Aller P, Yuan ZA, Gibson CW, Basenga R (1984) Cell-cycle specific cDNAs from mammalian cells temperature sensitive for growth. Proc Natl Acad Sci USA 81:6004-6008
Horinouchi S, Weisblum B (1982) Nucleotide sequence and func- tional map of pC194, a plasmid that specifies inducible chlo- ramphenicol resistance. J Bacteriol 150:815-825
Hugenholtz J (1993) Citrate metabolism in lactic acid bacteria. FEMS Microbiol Rev 12:165-178
Ishiguro N, Sato G (1985) Nucleotide sequence of the gene deter- mining plasmid-mediated citrate utilization. J Bacteriol 164:977-982
Ishiguro N, Izawa H, Shinagawa M, Shimamoto T, Tsuchiya T (1992) Cloning and nucleotide sequence of the gene (citC) en- coding a citrate carrier from several Salmonella serovars. J Biol Chem 267:9559-9564
Kempler GM, McKay LL (1981) Biochemistry and genetics of citrate utilization in Streptococcus lactis subsp, diacetylactis. J Dairy Sei 64:1527-1539
Lacks SA (1970) Mutants of Diplococcus pneumoniae that lack deoxyribonucleases and other activities possibly pertinent to genetic transformation. J Bacteriol 101:373-383

599
Lacks SA, L6pez P, Greenberg B, Espinosa M (1986) Identifica- tion and analysis of genes for tetracycline resistance and repli- cation functions in the broad-host-range plasmid pLS1. J Mol Biol 192:753-765
Lesage P, Chiaruttini C, Graffe M, Dondon J, Miler M, Springer M (1992) Messenger RNA secondary structure and trans- lational coupling in Escherichia coIi operon encoding trans- lation initiation factor IF3 and the ribosomal proteins, L35 and L20. J Mol Biol 228:366-385
Liu J, Baruell WO, Conway T (1992) The polycistronic mRNA of the Zygomonas mobilis glf-zwf-edd-glk operon is subjected to complex transcript processing. J Bacteriol 174:2824-2833
L6pez P, Diaz A, Espinosa M, Lacks SA (1989) Characterization of the poIA gene of Streptococcus pneumoniae and comparison of the DNA polymerase I it encodes to homologous enzymes from E. coli and phage T7. J Biol Chem 264:4255-4263
L6pez de Felipe F, Corrates MA, Ldpez P (1994) Comparative analysis of gene expression in Streptococcus lactis and Lacto- coccus lactis. FEMS Microbiol Lett 122:289-296
Ludwig W, Seewaldt E, Klipper-Balz R, Schleifer KH, Magrum L, Woese CR, Fox, GE, Stackebrandt E (1985) The phytogenetic position of Streptococcus and Enterococcus J Gen Microbiol 131:543-551
Magni C, L6pez de Felipe F, Sesma F, L6pez P, de Mendoza D (1994) Citrate transport in Lactococcus lactis subsp, lactis bio- vat diacetylactis. Expression of the citrate permease P. FEMS Microbiol Lett 118:75-82
Martinez S, L6pe'z P, Espinosa M, Lacks SA (1987) Complemen- tation of B. subtilis poIA mutants by DNA polymerase I from Streptococcus pneumoniae. Mol Gen Genet 210:203-210
Mayford M, Weisblum B (1989) Conformational alterations in the ermC transcript in vivo during induction. EMBO J 8:4307- 4314
McCarthy JEG, Gualerzi C (1990) Translational control of prokaryotic gene expression. Trends Genet 6:78-85
Pons ME, Diaz A, Lacks, SA, L6pez P (1991) The polymerase domain of Streptococcus pneumoniae DNA polymerase I. High expression, purification and characterization. Eur J Biochem 201:147 155
Rosenberg M, Court D (1979) Regulatory sequences involved in the promotion and termination of RNA transcription. Annu Rev Genet 13:319-353
Sanger F, Nicklen S, Coulson AR (1977) DNA sequencing with chain-terminating inhibitors. Proc Nat1 Acad Sci USA 74: 5463-5467
Schurr T, Nadir E. Margalit H (1993) Identification and character- ization of the Escherichia coli ribosomal binding sites by free energy computation. Nucleic Acids Res 21:4019-4023
Sesma F, Gardiol D, de Ruiz Holgado AP, de Mendoza D (1990) Cloning and expression of the citrate permease gene of Lacto- coccus lactis subsp, lactis biovar diacetylactis in Escherichia coli. Appl Environ Microbiol 56:2099-2103
Shaw WV (1975) Chloramphenicol acetyltransferase from chlo- ramphenicol-resistance bacteria. Methods Enzymol 43:737- 755
Simon D, Chopin A (1988) Construction of a vector plasmid fam- ily and its use for molecular cloning in Streptococcus Iactis. Biochimie 70:559-566
Studier FW, Rosenberg AH, Dunn JJ (1989) Use of the T7 RNA polymerase to direct expression of cloned genes~ Methods En- zymol 185: 60-89
Tinoco I, Borer PN, Dengler B, Levine MD (1973) Improved estimation of secondary structure in ribonucleic acids. Nature 246: 40~1
Van de Guchte M, Kok J, Venema G (1992) Gene expression in Lactococcus lactis. FEMS Microbiol Rev 88:73-92
Van der Rest ME, Siewe RM, Abee T, Schwarz E, Oesterhelt D, Konings WN (1992) Nucleotide sequence and functional prop- erties of a sodium-dependent citrate transport system from KlebsielIa pneumoniae. J Biol Chem 267:8971-8976
Van Rooijen RJ, de Vos WM (1990) Molecular cloning, transcrip- tional analysis and nucleotide sequence of lacR, a gene encod- ing the repressor of the lactose phosphotransferase system of Lactococcus lactis. J Biol Chem 265:18499-18503
Yarchuk O, Jacques N, Guillerez J, Dreyfus M (1992) Interdepen- dence of translation, transcription and mRNA degradation in the lacZ gene. J Mol Biol 226:581-596
Zuker M, Stiegler P (1981) Optimal computer folding of large RNA sequences using thermodynamics and auxiliary informa- tion. Nucleic Acids Res 9:133-148
Related Documents











