52 Dengue Bulletin – Vol 27, 2003 C i rcul ati on of D engue Vi ruses i n North-W estern Peru, 2000-2001 € by Ysabel Montoya*, Susan Hol echek*, Omar Caceres*, Ana Pal acios**, James Burans***, Caroli na Guevara***, Fernando Qui ntana**, Victor Herrera Ö , Edwar Pozo ψ , Eli z abeth Anaya*, Enri que Mamani*, Victori a Guti erre z*, Adr i ana Ladron de Guevara ÖÖ , Eduardo Fernande z ÖÖÖ , Percy Asmat ÖÖÖ , Victor Alva- D aval os ¶ , Car l os Hogui n ¶ , V Al berto Laguna**, Ana M Moral es**, Percy Minaya** and Tadeusz Kochel*** # *N at i onal Inst i t ut e of H eal t h, Peru **G eneral O ff i ce of Epi demi ol ogy, Peru ***U .S. N aval Medi cal Research Cent er D etachment , Li ma , Peru Ö Mi nistry of H eal t h Regi onal Ref erenc e Laborat ory, Tumbes, Peru ÖÖ Mi nistry of H eal t h Regi onal Ref erenc e Laborat ory, Lambayeque, Peru ÖÖÖ Mi nistry of H eal t h Regi onal Ref erenc e Laborat ory, La Li berta d, Peru ¶ Mi nistry of H eal t h Regi onal Ref erenc e Laborat ory, Pi ura , Peru ψ Mi nistry of H eal t h CI CE-Luci ano Cast ill o Col onna Sub-Regi on, Querecot ill o, Peru Abstract Pri or t o 2000 , onl y DEN-1 a nd t he Ameri c a n genot ype DEN-2 vi ruses wer e i sol a t ed i n Peru . Duri ng t he dengue out bre ak from May 2000 t o May 2001 i n t he Nort h-west dep art ments of Peru , cl i ni c al d a t a a nd bl ood sa mpl es were obt a i ned from 1 , 652 p a t i ents who present ed cl i ni c al ma ni f est a t i ons c omp ara bl e wi t h t hose of dengue . Vi rus genome i sol at i ons were ma de from 278 (16 . 8%) of t he sa mpl es. Serot ype-spe ci f i c monocl on al a nt i bod i es a nd pol ymeras e c ha i n r e a ct i on (PCR) wer e us ed t o t ype isol at i ons. The genot ype of t he maj ori t y of t he DEN-2 isol at i ons was de t ermi ned by t he restri ct i on frag ment l engt h pol ymorphism (RFLP) a nal ysis. Members of e a ch serot ype a nd t wo genot ypes of DEN-2 vi rus were i sol at ed: DEN-1 (n=93) ; Asi a n genot ype DEN-2 (n=104) ; Ameri c a n genot ype DEN-2 (n=1) ; DEN-2 genot ype not de t ermi ned (n=53) ; DEN-3 (n=25) ; a nd DEN-4 (n=2). Represent at i ves of e a ch serot ype , a nd genot ype f or DEN-2 , wer e c onf i rmed by p art i al sequenc e a nal ysi s. Dengue a nt i bod i es as de t ermi ned by Pl a que Reduct i on Neutral i z at i on Test (PNT) were present i n t he acut e serum from t he si ngl e DHF Gra de-II pat i ent , i n 19 of 31 (61%) a c ut e sera from p a t i ents wi t h DHF Gra de-I a nd i n 3 of 45 (7%) a c ut e sera of DF p a t i ents, suggest i ng a n associ at i on be t ween pre-exi st i ng dengue a nt i body a nd d i s e as e s everi t y. Keywords: Dengue viruses, dengue transmissi on, dengue out break, Peru. € This rese arch was support ed by t he US N aval Medi cal Rese arch and Devel opment Command NMMC , Be t hesda , MD , Work Uni t No 62787A8701447. The opi ni ons and assert i ons cont ai ned here i n are t he pri vat e ones of t he aut hors and are not t o be construed as off i ci al or refl e ct i ng t he vi ews of t he N avy Depart ment of t he N avy Servi c e at l arge . # For correspondence : N aval Medi cal Rese arch Cent er, Si l ver Spri ng, Maryl and 20910-7500, USA; E-mai l : kochel t @nmrc . navy. mil

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

52 Dengue Bulletin – Vol 27, 2003
Circulation of Dengue Viruses in North-Western Peru, 2000-2001€
by Ysabel Montoya*, Susan Holechek*, Omar Caceres*, Ana Palacios**,
James Burans***, Carolina Guevara***, Fernando Quintana**, Victor HerreraÖ, Edwar Pozoψ, Elizabeth Anaya*, Enrique Mamani*, Victoria Gutierrez*, Adriana Ladron de GuevaraÖÖ, Eduardo FernandezÖÖÖ, Percy AsmatÖÖÖ,
Victor Alva-Davalos¶, Carlos Hoguin¶, V Alberto Laguna**, Ana M Morales**, Percy Minaya** and Tadeusz Kochel***#
*National Institute of Health, Peru **General O ffice of Epidemiology, Peru
***U .S. Naval Medical Research Center Detachment, Lima, Peru ÖMinistry of Health Regional Reference Laboratory, Tumbes, Peru
ÖÖMinistry of Health Regional Reference Laboratory, Lambayeque, Peru ÖÖÖMinistry of Health Regional Reference Laboratory, La Libertad, Peru
¶Ministry of Health Regional Reference Laboratory, Piura, Peru ψMinistry of Health CICE-Luciano Castillo Colonna Sub-Region, Querecotillo, Peru
Abstract Prior to 2000, only DEN-1 and the American genotype DEN-2 viruses were isolated in Peru. During the dengue outbreak from May 2000 to May 2001 in the North-west departments of Peru, clinical data and blood samples were obtained from 1,652 patients who presented clinical manifestations comparable w ith those of dengue. Virus genome isolations were made from 278 (16.8%) of the samples. Serotype-specific monoclonal antibodies and polymerase chain reaction (PCR) were used to type isolations. The genotype of the majority of the DEN-2 isolations was determined by the restriction fragment length polymorphism (RFLP) analysis. Members of each serotype and two genotypes of DEN-2 virus were isolated: DEN-1 (n = 93); Asian genotype DEN-2 (n = 104); American genotype DEN-2 (n = 1); DEN-2 genotype not determined (n = 53); DEN-3 (n = 25); and DEN-4 (n = 2). Representatives of each serotype, and genotype for DEN-2, were confirmed by partial sequence analysis. Dengue antibodies as determined by Plaque Reduction Neutralization Test (PNT) were present in the acute serum from the single D HF Grade-II patient, in 19 of 31 (61%) acute sera from patients w ith D HF Grade-I and in 3 of 45 (7%) acute sera of DF patients, suggesting an association between pre-existing dengue antibody and disease severity.
Keywords: Dengue viruses, dengue transmission, dengue outbreak, Peru.
€ This research was supported by the US Naval Medical Research and Development Command NMMC , Bethesda, M D , Work Unit No 62787A8701447. The opinions and assertions contained herein are the private ones of the authors and are not to be construed as official or reflecting the views of the Navy Department of the Navy Service at large. # For correspondence : Naval Medical Research Center, Silver Spring, Maryland 20910-7500, USA; E-mail: kochelt @nmrc.navy.mil

Dengue Viruses of Peru, 2000-2001
Dengue Bulletin – Vol 27, 2003 53
Introduction Dengue viruses are single-stranded, positive-sense RNA viruses that belong to the genus Flavivirus, family Flaviviridae and they are transmitted primarily by the Aedes aegypti mosquito(1). There are four serotypes of dengue viruses (DEN 1-4), each of which can cause disease(2). Over the past 40 years, the number of DF cases has increased at least twentyfold and the number of D HF cases has similarly increased over the same time-period(3).
W ith the re-establishment of the Aedes aegypti vector in central and south America, the region observed an increased dengue virus activity and all four serotypes of dengue viruses are now widely distributed(4,5).
Epidemics of DF fever caused by DEN-2 and DEN-3 were reported from the Caribbean and northern South America in the 1960s and 1970s, respectively, and with the introduction of DEN-1 into the region in 1977, widespread epidemics of DF occurred in many different countries throughout Latin America(4,5,6). Although multiple serotypes of dengue virus were circulating in the Americas during the 1970s, no outbreak of D HF occurred until 1981 when the Asian genotype DEN-2 virus was introduced into Cuba(5,6). Since then, this virus and D HF have spread across the Americas.
Epidemiological studies have shown that the severity of the disease can be dependent on the infecting virus and on the individual’s previous history of infection by dengue viruses(7). Evidence of both mechanisms of dengue pathogenesis can be seen in the Americas. A primary dengue infection will usually be either sub-clinical or cause DF and the individual will develop
immunity to the infecting serotype of virus. Heterotypic immunity is short-lived and sequential heterotypic infections are hypothesized to lead to an increased disease severity by an antibody-dependent enhancement mechanism (ADE)(8,9). Increased disease severity has been associated with all combinations of sequential infections, but a greater percentage of the cases with the sequential infection combination of DEN-1, followed by DEN-2, have led to increased disease severity(10,11). In Peru, DEN-1 and the American genotype, DEN-2 virus, have been circulating since the 1990s without D HF(12,13). The lack of D HF may be attributed to a heterologous neutralization phenomenon. Sera from DEN-1-immune individuals cross-neutralize American, but not Asian, genotype DEN-2 virus in vitro(14). This heterologous neutralization is unidirectional (DEN-1 to American genotype DEN-2 only), and it was expected that when the other serotypes (and genotypes for DEN-2) of dengue virus were introduced into Peru, no heterologous protection (other than against the American genotype DEN-2 virus) would be available to individuals and D HF would be observed. By 2000, all four serotypes of dengue virus, with the Asian genotype representing most isolates of DEN-2, were present in countries of the region, including Peru(15).
From May 2000 to May 2001, 28,815 cases of DF and 251 cases of D HF were reported by the Peruvian M inistry of Health in the Northwest Departments of Peru(16,17). Blood samples were collected from 1,652 clinically-presenting dengue patients for the purpose of identifying the etiological agent(s) of the outbreak and to further our understanding of the pathogenesis of dengue viruses.

Dengue Viruses of Peru, 2000-2001
54 Dengue Bulletin – Vol 27, 2003
Materials and methods
Sample collection
During the outbreak, the regional clinics and hospitals in north-western Peru collected blood samples from patients with symptoms consistent with dengue disease (n = 1,652), either dengue fever or a grade of dengue haemorrhagic fever as defined by the World Health O rganization(18). Sera were prepared and stored at –700C until they were delivered on dry ice to laboratories in Lima, Peru, for virus isolation and antibody testing. C linical and demographic data were collected from all sample-donating patients.
Virus isolation and identification
Sera were tested for virus in C6/36 cells propagated in T-25 cm2 flasks and viral isolates were identified by indirect fluorescent antibody testing using anti-dengue polyclonal and dengue serotype 1-4-specific monoclonal antibodies(19,20). The serotypes of virus isolates were confirmed by serotype-specific PCR(21). The genotypes of the DEN-2 virus isolates were determined by Restriction fragment length polymorphism (RFLP) analysis(22). The capsid, premembrane and envelope genes of representative DEN-1, DEN-3, DEN-4 and DEN-2 American and Asian genotype virus isolates were sequenced using the following protocol. PCR amplified envelope fragments were purified and ligated into the pGem-T vector (Promega). Recombinant plasmids were transformed in E. coli strain D H5α and purified by alkaline lysis. The nucleotide sequences of inserts were determined by primer extension dideoxy chain termination
method using AutoCycle Sequencing Kits and Taq Polymerase (Amersham Pharmacia Biotech) and an ALF Express Automatic D NA Sequencer.
Serology
Plaque Reduction Neutralization Test (PRNT)
A modified PRNT protocol of Morens et al was followed(23). Test sera were diluted twofold in media (E-MEM + Pen./Strep.) from 1:5 to 1:5,120. 200 ul media containing 40 to 80 PFU of assay virus was mixed with 200 ul diluted test serum and then incubated at 40C for 15 hours. In triplicate, 100 ul virus-serum mixture was added to 0.5 ml media containing 1.5 X 105 BHK21 cells and then added to a well of a 24-well tissue culture plate and incubated at 370C with 5% C O2 for three hours. The cells were then overlaid with 0.5 ml of overlay media (0.6% carboxymethyl cellulose, MEM w/o Phenol Red, 10% FBS, 0.075% NaH C O3 and Pen./Strep.) and incubated at 370C with 5% C O2 for five days. The media was then removed, the cells were rinsed with H2 O and stained with 0.5 ml/well stain solution (0.1% (w/v) Naphthol Blue Black, 1.36% (w/v) Sodium Acetate, and 6% (v/v) G lacial Acetic Acid) for 30 min. The stain was removed and the plaques were counted. The results were expressed as the serum dilution, determined by probit analysis that reduced the number of plaques by 50% compared to that of normal human serum at the same dilution. A PRNT50 titer < 1:20 was considered negative; a PRNT50 titer > 1:20 was considered positive.

Dengue Viruses of Peru, 2000-2001
Dengue Bulletin – Vol 27, 2003 55
PRNT and RFLP viruses
DEN-1 virus 16007, DEN-3 virus 16562 and Asian genotype DEN-2 virus 16681 were isolated in Thailand in 1964 from D HF cases(2). DEN–4 virus 1036 was obtained from ATCC . The American genotype DEN-2 virus IQT 2913 was isolated in Iquitos, Peru, in 1996, by the Naval Medical Research Center Detachment, Lima, Peru.
IgM ELISA
Dengue IgM antibody capture ELISA were performed following the protocol of Wu, et al(24), with the substitution of DEN-1 virus suckling mouse brain preparation for cell culture supernatants DEN-1 virus preparation as antigen(24). Acute specimens with optical density readings two standard deviations higher in dengue antigen wells than negative antigen wells were positive. For paired sera, the criterion for seroconversion was a fourfold increase in antibody titer in the convalescent specimen as compared to that of the acute specimen.
Results Sera samples were collected from 1,652 patients presenting symptoms consistent with dengue. 171 of the patients also provided convalescent sera samples. Based on signs and symptoms the specimens were divided into three categories, those that were from DF (1,481), from D HF Grade-I (170) and from D HF Grade-II (1) patients. The most frequently observed symptoms were fever, chills, eye pain and body pain (Table 1). Anti-dengue IgM antibodies were present in 18.7% of the acute sera and 52.1% of the paired sera exhibited dengue IgM seroconversion. Dengue viruses were isolated from 278 (16.8%) of the total sera samples.
Table 1. Clinical symptoms and presentation
Symptoms %
Fever 100 Headache 98 Chills 82 Eye pain 82 Myalgia 81 Joint pain 45 Nausea, vomiting 47 Sore throat 40 Cough 36 Nasal congestion 28 Rash 28 D iarrhea 22 Malaise 14 Muscle pain 14 Bloody nose 6 Bloody gums 4 Jaundice 3 Bloody feces 3 Lack of appetite 3 Bone pain 4 Bloody vomit 2 Vaginal hemorrhage 2 Bloody urine 1 Abdominal pain 2 Photophobia 0.6 Itching 0.5 Enlarged lymph nodes 0.3 Cold skin 0.3 Pale 0.1 Unconsciousness 0.07 Rapid pulse 0.07 Presentation Number %
DF 1,481 89.7 D HF Grade-I 170 10.3 D HF Grade-II 1 0.06
Dengue Viruses of Peru, 2000-2001
56 Dengue Bulletin – Vol 27, 2003
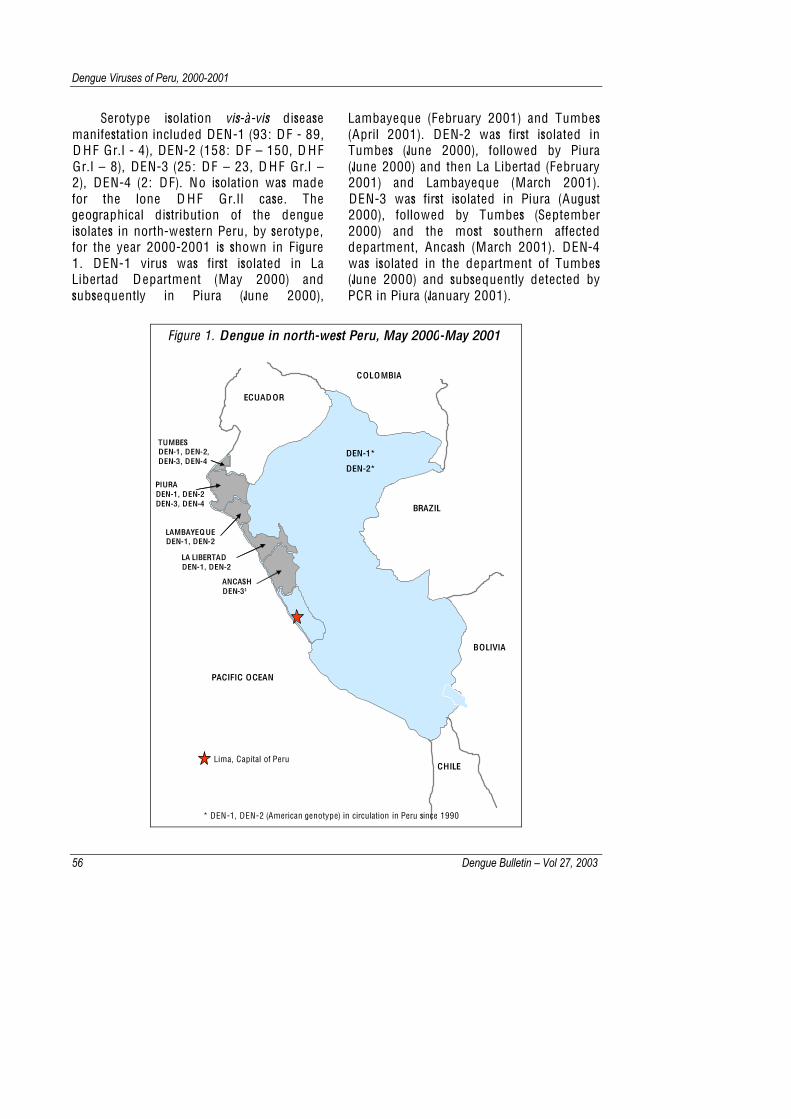
Serotype isolation vis-à-vis disease manifestation included DEN-1 (93: DF - 89, D HF Gr.I - 4), DEN-2 (158: DF – 150, D HF Gr.I – 8), DEN-3 (25: DF – 23, D HF Gr.I – 2), DEN-4 (2: DF). No isolation was made for the lone D HF Gr.II case. The geographical distribution of the dengue isolates in north-western Peru, by serotype, for the year 2000-2001 is shown in Figure 1. DEN-1 virus was first isolated in La Libertad Department (May 2000) and subsequently in Piura (June 2000),
Lambayeque (February 2001) and Tumbes (April 2001). DEN-2 was first isolated in Tumbes (June 2000), followed by Piura (June 2000) and then La Libertad (February 2001) and Lambayeque (March 2001). DEN-3 was first isolated in Piura (August 2000), followed by Tumbes (September 2000) and the most southern affected department, Ancash (March 2001). DEN-4 was isolated in the department of Tumbes (June 2000) and subsequently detected by PCR in Piura (January 2001).
BOLIVIA
TUMBESDEN-1, DEN-2,DEN-3, DEN-4
ECUADOR
COLOMBIA
BRAZIL
CHILE
PACIFIC OCEAN
PIURADEN-1, DEN-2DEN-3, DEN-4
LA LIBERTADDEN-1, DEN-2
Lima, Capital of Peru
LAMBAYEQUEDEN-1, DEN-2
ANCASHDEN-3/
DEN-1*
DEN-2*
* DEN-1, DEN-2 (American genotype) in circulation in Peru since 1990
Figure 1. Dengue in north-west Peru, May 2000-May 2001

Dengue Viruses of Peru, 2000-2001
Dengue Bulletin – Vol 27, 2003 57
Since American and Asian genotypes of DEN-2 virus have been in circulation in the southern hemisphere, RFLP analysis was performed on the majority of the DEN-2 isolates to determine their genotype. Representative restriction digests of Asian and an American genotype DEN-2 virus isolates are shown in Figure 2. The DEN-2 isolates of lanes 5-7 yield restriction patterns
identical to the marker Asian genotype virus, lane 3. The DEN-2 isolate of lane 8 has an identical restriction pattern to the American genotype marker virus of lane 4. Greater than 99% (104/105) of the samples tested were of the Asian genotype. O nly one isolate, from Piura, was of the American genotype.
Figure 2. RFLP Analysis of DEN-2 Isolates‡
1600 bp --
2400 bp --
100 bp --
200 bp --
400 bp --
800 bp --
1 2 3 4 5 6 7 8
1600 bp --
2400 bp --
100 bp --
200 bp --
400 bp --
800 bp --
1 2 3 4 5 6 7 8
[Lane 1: D NA molecular weight ladder, marker sizes are listed in the photo margin; lane 2: no template RT/PCR control reaction; lane 3: 295216 Asian DEN-2 marker virus; lane 4: IQ T2124 American genotype DEN-2 marker
virus; lane 5-8: O BT 396, O BT 1242, O BT 1243 and O BS 9638 viruses, respectively.]
‡ RT/PCR products of DEN-2 virus isolates were digested with Alu I, electrophoresed through a 2.0% agarose gel, stained with ethidium bromide and photographed.
To further identify the viruses of the outbreak, portions of the capsid, premembrane and envelope genes of representative isolates of each serotype, and genotype for DEN-2, were sequenced. The
sequence of the DEN-1 isolates was ninety-eight per cent identical to DEN-1 viruses that were isolated in Peru in 1997. Similarly, the sequence of the American genotype DEN-2 isolates was 98% identical to an American
Dengue Viruses of Peru, 2000-2001
58 Dengue Bulletin – Vol 27, 2003
genotype isolate from Iquitos, Peru, during 1996. The Asian DEN-2 isolates were 98% identical to the Asian DEN-2 isolates of Maracay, Venezuela, in 1998. The DEN-3 isolates were 98% identical to the DEN-3
isolates of Guatemala (1998) and DEN-4 isolates were 98% identical to the DEN-4 isolates of the Dominican Republic (1981) (Table 2).
Table 2. Identity of representative dengue virus isolates
Serotype Gene Accession
number and strain
Place and year of
isolation
Accession number and
strain
Country and year
Identity (%)
DEN-1 E AF378171 Peru 9808-00
Piura, 2000 AF378172 Peru 5189-97
Peru 1997
98.03
DEN-2 Asian Genotype
E AY079424 Sullana 6663-01
Piura, 2001 AF163096 PTC OL 96
L10041 C lone 2A
AY044442 Mara 3
Colombia 1996
Brazil 1990
Venezuela 1990
98.529
98.162
97.61
DEN-2 American Genotype
E AF378163 Peru 9638-00
Piura, 2000 AF100465 Ven 2 AF264054 PR 152 AF100468 IQT 2913
Venezuela 1987 Puerto Rico 1969 Peru 1996
98.32
98.13
98.13
DEN-3 C/PrM AY079175 Peru 6682-01
Piura, 2001 AB038475 Guate 98-2
Guatemala 1998
98
DEN-4 C/PrM AY079176 Peru 6682-01
Piura, 2001 AF326573 Dominica 81
Domincan Republic 1981
98
Since DEN-1 and the American
genotype DEN-2 viruses have been in circulation in Peru during the last decade, it was expected that some persons infected with dengue viruses during this outbreak were experiencing secondary infections. In order to determine if pre-existing dengue antibodies influenced the disease presentation, the acute sera dengue antibody status of individuals with DF (n = 45) and the more severe illnesses of D HF Grade-I (n = 31) or D HF Grade-II
(n = 1) were determined by PRNT. Primary dengue infections were identified by the absence of any dengue virus neutralizing antibody in the acute sera, while acute sera from secondarily infected individuals contained dengue neutralizing antibody against at least one serotype dengue virus. The acute sera that were selected for this analysis were dengue IgM antibody negative and were possible sera from which dengue virus had been isolated. Dengue viruses were
Dengue Viruses of Peru, 2000-2001
Dengue Bulletin – Vol 27, 2003 59
isolated from all 45 sera of DF-presenting patients, while dengue viruses were isolated from the sera of only four of the 31 patients that had D HF Grade-I, and no virus was isolated from the lone D HF Grade-II specimen that was available for analysis. By
PRNT, 7% (3 of 45) of the DF cases showed evidence of secondary heterologous serotype infections, while 61% (19 of 31) of the D HF Grade-I cases showed evidence of secondary heterologous serotype infections as did the D HF Grade-II case (Table 3).
Table 3. Dengue serostatus vs. disease severity
Disease 1o infection
number Isolated virus 2o infection
number Pre-existing
antibody Isolated
virus
DF 42 DEN-1 (n = 6) DEN-2 (n = 31) DEN-3 (n = 5)
3 DEN-1 (n = 3) DEN-2 (n = 2) DEN-3 (n = 1)
D HF Grade I
12 DEN-2 (n = 1) DEN-3 (n = 1)
19 DEN-1 (n = 5) DEN-2 (n = 14)
0 DEN-1 (n = 1) DEN-3 (n = 1)
D HF Grade II
0 0 1 DEN-2 (n = 1) 0
D iscussion Here we report the isolation and characterization of DEN-1, DEN-2 (Asian genotype), DEN-2 (American genotype), DEN-3 and DEN-4 viruses as the etiological agents of the May 2000–May 2001 dengue outbreak in north-western Peru. Along with the introduction of the DEN-2 Asian genotype, DEN-3 and DEN-4, Peru has seen her first D HF Grade-II cases.
The viruses that were isolated during this outbreak were not new to South America. DEN-1 and the American genotype DEN-2 viruses have been in circulation in Peru prior to this outbreak and the Asian genotype DEN-2, DEN-3 and DEN-4 viruses are genetically identical to those that were isolated in Ecuador during a dengue outbreak six months prior to the Peruvian
outbreak(25). We speculate that the ease of trans-border travel between Ecuador and Peru facilitated the transmission of these dengue viruses into north-west Peru. The geographical distribution of virus isolates demonstrated the co-circulation of multiple serotypes, and genotypes of DEN-2, of dengue virus during this outbreak. The greatest number of virus isolations was made in Piura where DEN-1, DEN-2 (American and Asian genotypes), DEN-3 and DEN-4 viruses were isolated. Piura was also the department with the greatest number of severe cases. DEN-4 was the least isolated virus of this outbreak with only two isolations in the Tumbes department and one identification by PCR in the Piura department. DEN-4 was not isolated from the southern departments, but we cannot rule out its presence there, since acute sera from only a fraction of the clinically-

Dengue Viruses of Peru, 2000-2001
60 Dengue Bulletin – Vol 27, 2003
presenting patients were available for virus testing. The same possibility exists for the other departments where a serotype of dengue was not isolated. It must be noted that DEN-1 cases were being seen in the most southern city of Casma in the southern department of Ancash during the preparation of this manuscript.
Since we have isolated the American DEN-2 virus from this outbreak and from the outbreak in Ecuador the year before, it raises other possibilities to the hypothesis that the Asian DEN-2 virus has displaced the American DEN-2 virus in South America(7). Possibly, the American DEN-2 virus is less virulent, so infected people do not seek medical attention, and hence it is isolated less often. Support for this hypothesis comes from this report where 73% (14/19) secondary infected individuals with D HF Grade-I and the lone D HF Grade-II individual had DEN-2 antibodies in their acute sera. These antibodies were presumably from American genotype DEN-2 infections, since the Asian genotype DEN-2 virus had not been previously isolated in Peru. To avoid misinterpreting PRNT results for the determination of pre-existing dengue antibody, only IgM negative specimens were considered for this portion of the study.
Explanations of the pathogenesis of dengue have focused on two non-exclusive hypotheses, viral determinants and ADE(26,27,28). Evidences for both the theories exist in the Americas. The American genotype DEN-2 virus is relatively avirulent, while the Asian genotype DEN-2 virus is associated with severe disease(7,13). DEN-1 antibody positive individuals secondarily infected with the American genotype DEN-2 virus did not develop D HF Grade-II in Iquitos, Peru, while in other dengue
outbreaks in Central and South America, DEN-1 antibody-positive individuals secondarily infected with the Asian genotype of DEN-2 did(4,29,30). Here we report that the D HF Grade-II case and 61% of the D HF Grade-I cases were the products of secondary infections, while only 7% of DF patients had pre-existing dengue antibodies. This finding is supportive of the hypothesis that secondary heterologous infections result in a higher frequency of severe dengue disease as compared to primary dengue infections, but it must be noted that we are reporting on patients with D HF Grade-I and only one D HF Grade-II case. However, the trend of disease severity with pre-existing antibodies is evident. Few isolations were made from specimens from D HF Grade-I patients (4/31) and no virus was isolated from the D HF Grade-II patient’s sera. Possibly those samples were collected in the post-viremic stages of the illness or the virus isolation process was complicated by pre-existing antibody. Unfortunately, for the majority of the D HF Grade-I specimens and that of the D HF Grade-II patient, only clinical presentations were available for assignment of dengue disease.
Future studies are aimed at furthering our understanding of the transmission and pathogenesis of dengue viruses in South America.
Acknowledgements We thank Zonia Rios, Vidal Felices and Angelica Espinoza for technical assistance with virus isolation, PCR and RFLP, and PRNT assays, respectively. We also thank Juan Perez for data management assistance and Curtis Hayes and Douglas Watts for a critical review of this manuscript.

Dengue Viruses of Peru, 2000-2001
Dengue Bulletin – Vol 27, 2003 61
References1. Chambers TJ, H ahn CS, Galler R and Rice CM .
Flavivirus genome organization, expression and replication. A. Rev. M icrobiol, 1990, 44: 649-688.
2. Leitmeyer KC , Vaughn D W , Watts DM , Salas R, Villalobos I de, Chacon Ramos C and Rico-Hesse R. Dengue virus structural differences that correlate with pathogenesis. J Virol, 1999, 73(6): 4738-4747.
3. World Health O rganization, Removing obstacles to health. World Health O rganization, 1999.
4. Pinheiro FP and Corber SJ. G lobal situation of dengue and dengue haemorrhagic fever and its emergence in the Americas. World Health Stat Q , 1997, 50(3-4): 161-169.
5. Gubler DJ. The global pandemic of dengue/dengue haemorrhagic fever: Current status and prospects for the future. Ann Acad Med Singapore, 1998, 27(2): 227-234.
6. Isturiz RE, Gubler DJ and Brea del Castillo J. Dengue and dengue haemorrhagic fever in Latin America and the Caribbean. Infect D is C lin North Am, 2000, 14(1): 121-140, ix.
7. Rico-Hesse R, H arrison L M , Salas RA, Tovar D , N isalak A, Ramos C , Boshell J, de Mesa MT, Nogueira RM and da Rosa AT. O rigins of dengue type 2 viruses associated with increased pathogenicity in the Americas. Virology, 1997, 230(2): 244-251.
8. H alstead SB and Simasthien P. O bservations related to the pathogenesis of dengue haemorrhagic fever II. Antigenic and biological properties of dengue viruses and their association with disease response in the host. Yale J Biol Med, 1970, 42: 276-292.
9. Kliks SC , N isalak A, Brandt WE, Wahl L and Burke DS. Antibody-dependent enhancement of dengue virus growth in human monocytes as a risk factor for dengue haemorrhagic fever. Am J Trop Med Hyg, 1989, 40(4): 444-451.
10. Sangkawibha N , Rojanasuphot S, Ahandrik S, Viriyapongse S, Jatanasen S, Salitul V, Phanthumachinda B and Halstead SB. Risk factors in dengue shock syndrome: A prospective epidemiologic study in Rayong, Thailand. The 1980 outbreak. Am J Epidemiol, 1984, 120(5): 653-669.
11. Guzman MG , Kouri G , Bravo J, Soler M and Martinez E. Sequential infection as risk factor for dengue haemorrhagic fever/dengue shock syndrome (D HF/DSS) during the 1981 dengue haemorrhagic Cuban epidemic. Mem Inst O swaldo Cruz, 1991, 86(3): 367.
12. Phillips I, Need J, Escamilla J, Colan E, Sanchez S, Rodriguez M , Vasquez L, Seminario J, Betz T and da Rosa AT. First documented outbreak of dengue in the Peruvian Amazon region. Bull Pan Am Health O rgan, 1992, 26(3): 201-207.
13. W atts D M , Porter KR, Putvatana P, Vasquez B, Calampa C , H ayes CG , H alstead SB. Failure of secondary infection with American genotype dengue 2 to cause dengue haemorrhagic fever. Lancet, 1999, 354(9188): 1431-1434.
14. Kochel TJ, Watts DM , H alstead SB, H ayes CG , Espinosa A, Felices V, Caceda R, Buatista CT, Montoya Y, Douglas S and Russell KL. Effect of dengue-1 antibodies on American dengue-2 viral infection and dengue haemorrhagic fever. Lancet, 2002, 360(27 Jul): 310-312.
15. PAH O , Dengue in the Americas 2000 and Dengue in the Americas 2001, in Pan American Health O rganization Travel Medicine Programme. 2002.
16. PAH O , Dengue Cases 2000.
17. PAH O , Dengue Cases 2001.
18. World Health O rganization, Technical guidelines for diagnosis, treatment and control of dengue haemorrhagic fever. 2nd ed, W H O , 1997, Geneva.
19. Tesh RB. A method for the isolation and identification of dengue viruses, using mosquito cell cultures. Am J Trop Med Hyg, 1979, 28(6): 1053-1059.
20. Igarashi A. Isolation of a Singh's Aedes albopictus cell clone sensitive to dengue and chikunggunya viruses. J Gen Virol, 1978, 40: 531-544.
21. Lanciotti RS, Calisher C H , Gubler DJ, Chang G and Vordam V. Rapid detection and typing of dengue viruses from clinical samples by using reverse transcriptase-polymerase chain reaction. J C lin M icrobiol, 1992, 30(3): 545-551.

Dengue Viruses of Peru, 2000-2001
62 Dengue Bulletin – Vol 27, 2003
22. Vorndam V, Kuno G and Rosado N . A PCR-restriction enzyme technique for determining dengue virus subgroups within serotypes. J Virol Meth, 1994, 48(2-3): 237-244.
23. Morens D M , H alstead SB, Repik PM , Putvatana R and Raybourne N . Simplified plaque reduction neutralization assay for dengue viruses by semimicro methods in BHK-21 cells: comparison of the BHK suspension test w ith standard plaque reduction neutralization. J C lin M icrobiol, 1985, 22(2): 250-254.
24. Wu SJ, H anson B, Paxton H , N isalak A, Vaughn D W , Rossi C , Henchal EA, Porter KR, W atts D M and Hayes CG . Evaluation of a dipstick enzyme-linked immunosorbent assay for detection of antibodies to dengue virus. C lin D iag Lab Immunol, 1997, 4(4): 452-457.
25. Kochel TJ. Unpublished Result. 2001.
26. H alstead SB. Pathogenesis of dengue: Challenges to molecular biology. Science, 1988, 239(4839): 476-481.
27. Rosen L. "The emperor's new clothes" revisited, or reflections on the pathogenesis of dengue haemorrhagic fever. J Trop Med Hyg, 1977, 26: 337-343.
28. Gubler DJ and Kuno G . Dengue and dengue haemorrhagic fever. 1997: CAB International.
29. Guzman MG , Kouri G , Valdes L, Bravo J, Vazquez S and H alstead SB. Enhanced severity of secondary dengue-2 infections: Death rates in 1981 and 1997 Cuban outbreaks. Rev Panam Salud Publica, 2002, 4(Apr 11): 223-227.
30. Guzman MG , Alvarez M , Rodriguez R, Rosario D , Vazquez S, Valdes L, Cabrera MV and Kouri G . Fatal dengue haemorrhagic fever in Cuba, 1997. Int J Infect D is, 1999, 3(3): 130-135.
Related Documents











