Mar Biol (2008) 153:953–963 DOI 10.1007/s00227-007-0867-x 123 RESEARCH ARTICLE Circadian shelter occupancy patterns and predator–prey interactions of juvenile Caribbean spiny lobsters in a reef lagoon Howard M. Weiss · Enrique Lozano-Álvarez · Patricia Briones-Fourzán Received: 8 August 2007 / Accepted: 8 November 2007 / Published online: 27 November 2007 © Springer-Verlag 2007 Abstract The spiny lobster, Panulirus argus, is predomi- nantly nocturnal, remaining inside shelters during the day and foraging outside at night, presumably to minimize pre- dation risk. Predation risk generally decreases with increas- ing lobster size. Therefore, this study examined the hypothesis that size would inXuence this basic circadian pattern. Video cameras continuously recorded the shelter occupancy of juvenile lobsters (n = 72) having a carapace length (CL) of 30–62 mm that were tethered to shelters in a shallow reef lagoon. The lobsters’ shelter occupancy was 100% during the day, but declined linearly from shortly before sunset to a minimum of 50% shortly after midnight and then increased linearly, reaching 100% by 1 h after sunrise. The percent time the lobsters spent in the shelters followed a similar trend, but there was wide variability at night (0–100%) for individual lobsters. Lobsters left their shelters 2–30 times night ¡1 , with a majority of excursions lasting <10 min. These results suggest that juvenile P. argus minimize predation risk by remaining in their shel- ters as long as possible but oVset the energetic cost of this behavior by foraging close to their shelters for several short periods at night. This emergence pattern contrasts with those of early benthic phase lobsters (<15 mm CL), which seldom leave their shelters, and adults (>80 mm CL), which have a dusk/early evening peak in activity and leave the shelter for extended periods of time during the night. Fur- thermore, a minimum shelter occupancy in the middle of the night appears especially well adapted to avoid expo- sure to daytime predators. Videotaped observations also included interactions between lobsters and two dominant lobster predators, the triggerWsh, Balistes capriscus, and the octopus Octopus cf. vulgaris. Lobsters responded diVer- ently to these predators: remaining in the shelter when attacked by a triggerWsh and Xeeing the shelter when attacked by an octopus. TriggerWsh were nearly twice as likely to attack a lobster that was outside of the shelter than inside. Once under attack, however, a lobster had nearly the same chance of surviving if it was inside or outside. Results suggest that the patterns of shelter use and emergence change as lobsters grow, probably reXecting the interplay between perception of predation risk and the need to forage. Introduction Many marine animals dwell in shelters during some portion of the day or night. Shelters provide protection from preda- tors as well as a micro-habitat where harsh external envi- ronmental conditions such as intense sunlight, strong waves or currents, extreme temperatures, and toxic chemicals can be avoided or modulated (Sih 1980). However, most animals must spend some time outside of their shelters to forage for food, locate a mate, migrate, or carry out other essential biological functions. Thus, the advantages of being in a shelter must be balanced with the necessities of Communicated by J.P. Grassle. Electronic supplementary material The online version of this article (doi:10.1007/s00227-007-0867-x) contains supplementary material, which is available to authorized users. H. M. Weiss (&) Project Oceanology, Avery Point, Groton, CT 06355, USA e-mail: [email protected] E. Lozano-Álvarez · P. Briones-Fourzán Unidad Académica Puerto Morelos, Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, PO Box 1152, Cancún, Quintana Roo 77500, Mexico

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Mar Biol (2008) 153:953–963
DOI 10.1007/s00227-007-0867-xRESEARCH ARTICLE
Circadian shelter occupancy patterns and predator–prey interactions of juvenile Caribbean spiny lobsters in a reef lagoon
Howard M. Weiss · Enrique Lozano-Álvarez · Patricia Briones-Fourzán
Received: 8 August 2007 / Accepted: 8 November 2007 / Published online: 27 November 2007© Springer-Verlag 2007
Abstract The spiny lobster, Panulirus argus, is predomi-nantly nocturnal, remaining inside shelters during the dayand foraging outside at night, presumably to minimize pre-dation risk. Predation risk generally decreases with increas-ing lobster size. Therefore, this study examined thehypothesis that size would inXuence this basic circadianpattern. Video cameras continuously recorded the shelteroccupancy of juvenile lobsters (n = 72) having a carapacelength (CL) of 30–62 mm that were tethered to shelters in ashallow reef lagoon. The lobsters’ shelter occupancy was100% during the day, but declined linearly from shortlybefore sunset to a minimum of 50% shortly after midnightand then increased linearly, reaching 100% by 1 h aftersunrise. The percent time the lobsters spent in the sheltersfollowed a similar trend, but there was wide variability atnight (0–100%) for individual lobsters. Lobsters left theirshelters 2–30 times night¡1, with a majority of excursionslasting <10 min. These results suggest that juvenile P.argus minimize predation risk by remaining in their shel-ters as long as possible but oVset the energetic cost of this
behavior by foraging close to their shelters for several shortperiods at night. This emergence pattern contrasts withthose of early benthic phase lobsters (<15 mm CL), whichseldom leave their shelters, and adults (>80 mm CL), whichhave a dusk/early evening peak in activity and leave theshelter for extended periods of time during the night. Fur-thermore, a minimum shelter occupancy in the middleof the night appears especially well adapted to avoid expo-sure to daytime predators. Videotaped observations alsoincluded interactions between lobsters and two dominantlobster predators, the triggerWsh, Balistes capriscus, and theoctopus Octopus cf. vulgaris. Lobsters responded diVer-ently to these predators: remaining in the shelter whenattacked by a triggerWsh and Xeeing the shelter whenattacked by an octopus. TriggerWsh were nearly twice aslikely to attack a lobster that was outside of the shelter thaninside. Once under attack, however, a lobster had nearly thesame chance of surviving if it was inside or outside. Resultssuggest that the patterns of shelter use and emergencechange as lobsters grow, probably reXecting the interplaybetween perception of predation risk and the need to forage.
Introduction
Many marine animals dwell in shelters during some portionof the day or night. Shelters provide protection from preda-tors as well as a micro-habitat where harsh external envi-ronmental conditions such as intense sunlight, strong wavesor currents, extreme temperatures, and toxic chemicals canbe avoided or modulated (Sih 1980). However, mostanimals must spend some time outside of their shelters toforage for food, locate a mate, migrate, or carry out otheressential biological functions. Thus, the advantages ofbeing in a shelter must be balanced with the necessities of
Communicated by J.P. Grassle.
Electronic supplementary material The online version of this article (doi:10.1007/s00227-007-0867-x) contains supplementary material, which is available to authorized users.
H. M. Weiss (&)Project Oceanology, Avery Point, Groton, CT 06355, USAe-mail: [email protected]
E. Lozano-Álvarez · P. Briones-FourzánUnidad Académica Puerto Morelos, Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, PO Box 1152, Cancún, Quintana Roo 77500, Mexico
123

954 Mar Biol (2008) 153:953–963
being outside. The amount of time and the hours of the dayin which species are inside versus outside their sheltershave presumably evolved to optimize their survival (Sih1980, 1987; Vannini and Cannicci 1995).
Spiny lobsters (Palinuridae) are nocturnal, tending toremain inside shelters during the day and foraging outsidethe shelters at night (see Phillips et al. 1980). Numerous Weldand laboratory studies have reported this basic circadianpattern for the Caribbean spiny lobster, Panulirus argus(Latreille, 1804) (e.g., SutcliVe Jr 1956; Herrnkind 1980;Childress and Herrnkind 1994). In general, individuals tendto emerge from shelters shortly after dusk and return to shel-ter prior to dawn. This basic circadian pattern, however, maybe modulated by many factors, such as the time of sunriseand sunset (Kanciruk and Herrnkind 1973; Childress andHerrnkind 1994), the season (Kanciruk and Herrnkind1973), the phase of the moon (SutcliVe Jr 1956), social inter-actions (Kanciruk and Herrnkind 1973), migrations (Herrn-kind 1980), mating behavior (Lipcius et al. 1983), themolting cycle (Lipcius and Herrnkind 1982), the life-cyclestage/size of the lobster (Childress and Herrnkind 1994), andpredation risk (Lozano-Álvarez 1996; Lozano-Álvarez andSpanier 1997; Lozano-Álvarez and Briones-Fourzán 2001).
Amongst all these factors, predation risk has possibly thegreatest inXuence on the shelter occupancy patterns of lob-sters. For example, spiny lobsters kept in sea enclosureswithout predators show an increase in daytime activity(Glaholt 1990; Lozano-Álvarez 1996). Moreover, becausepredation risk usually declines with increasing size of prey(Werner and Gilliam 1984; Wahle 1992), early benthicstages of lobsters are usually more cryptic and have smallerforaging ranges than later stages. Newly settled clawed lob-sters (Homarus americanus) remain in their burrows dayand night, feeding on suspended organic particles (Lavalliand Barshaw 1989). As they grow, they switch to feedingon benthic prey and increase their foraging range (Wahle1992). Newly settled P. argus (6–15 mm carapace length,CL) take shelter among the marine vegetation of shallowbay and lagoon habitats, feeding on available small prey(Childress and Herrnkind 1994). After a few months, juve-nile P. argus switch to crevice dwelling across these shallowhabitats. Because these juveniles have not yet reached a sizerefuge against predators, they sustain higher levels of preda-tion (Smith and Herrnkind 1992; Acosta and Butler 1997;Briones-Fourzán et al. 2007) and have smaller foragingranges than larger conspeciWcs (Herrnkind 1980; Briones-Fourzán et al. 2007). By contrast, adult, non-migratory indi-viduals of P. argus (80 to >200 mm CL) that dwell in coralreef habitats forage at night hundreds of meters away fromshelter (Herrnkind et al. 1975) and may also leave the shelterduring the day to search for mates (Lipcius et al. 1983).
Predators of juvenile P. argus include numerous speciesof Wshes and octopuses (Weiss et al. 2006). Some of these
species also have distinctive day-night activity patterns(Helfman 1986). In Caribbean habitats, potential lobsterpredators such as toadWshes (Batrachoididae), nurse sharks(Ginglymostomidae), grunts (Haemulidae), and adult snap-pers (Lutjanidae) are active during the night and reducetheir activity during the day (Starck II and Davis 1966;Ogden and Ziemann 1977; Robblee and Ziemann 1984).Others, such as triggerWshes (Balistidae), juvenile snappers,and some groupers (Serranidae) are active only duringthe daytime (Hobson 1965; Lozano-Álvarez and Spanier1997). Still others, such as moray eels (Muraenidae), somegroupers, and octopuses (Forsythe and Hanlon 1997;Meisel et al. 2003; Brock et al. 2006; Weiss et al. 2006) areactive during both phases of the light–dark cycle. In addi-tion, some of these species are searching predators (e.g.,triggerWshes, moray eels, octopuses) whereas others are sit-and-wait predators (e.g., toadWshes, groupers). The preda-tion pressure exerted by such a wide variety of organismswith diVerent activity schedules and hunting tactics, inaddition to the relatively scarce structured refuges in theshallow habitats where juvenile P. argus dwell (Acosta andButler 1997; Briones-Fourzán and Lozano-Álvarez 2001),likely increase the perception of danger by these juveniles.Lobsters perceive the presence of predators through tactile,visual, chemical, and other sensory cues (Barshaw et al.2003; Berger and Butler 2001; Horner et al. 2006).
The present study explored the day–night shelter occu-pancy patterns of juvenile P. argus in a natural habitat wherethese lobsters were subjected to resident predators. Unlikeprevious studies that have examined these patterns in lobsterscontained in mesocosms or through short periods of observa-tions in the Weld, we used continuous, 24 h in-situ video mon-itoring of lobster shelters, using red lights at night. Weiss et al(2006) found that this technique does not signiWcantly aVectthe activity patterns of juvenile lobsters and associated spe-cies. We measured the time and duration of shelter occupancyby juveniles tethered individually to shelters. We hypothe-sized that juveniles would remain in their shelters throughoutthe daytime and would be active outside their shelters onlyduring a short period at night, more likely shortly after dusk.This hypothesis was based on activity patterns reported inlarger P. argus, which invariably show a sunset/early eveningpeak (Kanciruk and Herrnkind 1973; Herrnkind et al. 1975;Lipcius and Herrnkind 1982). Video monitoring also allowedus to identify the prevalence of day–night predators and pred-ator attacks, and to describe the behavioral interactions ofjuvenile lobsters and their predators.
Materials and methods
Our study site was located in Puerto Morelos, Mexico(20°51�N, 86°55�W), in the reef lagoon between the shore
123

Mar Biol (2008) 153:953–963 955
and the Mesoamerican Reef, which extends along theCaribbean coast of the Yucatan Peninsula. Our observa-tions were conducted in a seagrass habitat about 150 mfrom shore and 2.5 m deep. This habitat has been describedin detail by Briones-Fourzán and Lozano-Álvarez (2001).In particular, natural crevice shelters are extremely scarceacross the Puerto Morelos reef lagoon, causing a populationbottleneck due to high mortality rates of juvenile P. argus.
Four artiWcial lobster shelters were placed on the bottom7 m apart from each other. Each shelter consisted of acement building block 40 cm wide £ 16 cm high £ 20 cmdeep with three openings 8.5 cm wide £ 8.5 cm high £20 cm deep. A solid cement block the same size was placedbehind each building block to close oV the back end of theopenings and to anchor a tether support frame. A videocamera and red light were placed 1.5 m in front of eachshelter (Fig. 1). The cameras were connected to a multi-plexor and time-lapse video recorder located in a weather-proof shed at the end of a dock. Weiss et al. (2006)provided detailed speciWcations for these electronic compo-nents and lights.
Experimental lobsters were collected from a habitat sim-ilar to but distant from our Weld study site. All lobsters(n = 72) were juveniles, with a mean (§SD) CL of44.5 § 6.5 mm and had hard exoskeletons indicative of anintermolt condition. They were placed in a large holdingtank and fed ad libitum for a few days until their use in theexperiment. One lobster was tethered to each shelter bylocking a plastic cable tie around its cephalothorax,between the second and third walking legs, and attaching a0.55 mm diameter monoWlament line to the cable tie usinga swivel clip. The monoWlament line was tied to a swivelclipped to a cable tie at the end of a 2.0 cm outside diameterPVC pipe that was suspended over the shelter and sup-ported by a PVC pipe frame anchored to the rear cementblock. The entire tether, including line, cable tie and swiv-els was 62 cm long. The end of the pipe where the tether
line was attached was 55 cm above the seaXoor and 30 cmin front of the shelter. The tether allowed the lobsters toenter and go to the back end of the refuge openings as wellas to walk around on top of the shelter and on the seaXoorwithin a circular area having a radius of »40 cm (Fig. 1).Tethered lobsters were replaced with new lobsters each daybetween 15:00 and 16:00 h. This tethering technique is sim-ilar to that used in other lobster studies in which speciWedbehavioral responses of tethered lobsters in the Weld wereshown to be similar to those of untethered lobsters observedin the Weld (e.g., Barshaw and Able 1990; Eggleston et al.1992; Smith and Herrnkind 1992; Childress and Herrnkind2001; Oliver et al. 2005). Because most of the natural shel-ters in our study area harbor one single juvenile (Briones-Fourzán et al. 2007), our investigation was conducted usinga single lobster tethered at each shelter. Juvenile P. argusmay forage several meters away from their shelter(Herrnkind 1980), but the artiWcial shelters were placed in ahabitat rich in potential food for juvenile P. argus (Briones-Fourzán et al. 2003) and tethering allowed for continuousobservations on the sheltering behavior of individual juve-niles.
Activity at the shelters was recorded continuously, 24 hper day, for 18 days (12–22 January and 3–11 March,2003). Ambient seawater temperature during the studyperiod ranged from 24.5 to 26.0°C. Twenty videotapes,each containing time-lapse recordings of up to four sheltersfor periods of 24–36 h, were viewed at 10£ speed overtheir entire length. The time and duration of each lobstershelter occupancy and visit to the shelters by a predator orother organism were logged onto computer spreadsheetWles. Activities of particular interest, such as predatorattacks and lobster defense behaviors, were viewed andanalyzed at actual speed or in slow motion and descriptionsof these activities were entered into the video logs.
Shelter occupancy (i.e., the proportion of lobsters in theshelters) was determined by noting the location of each lob-ster at instantaneous sample times chosen using randomnumbers to the nearest second within each half-hour inter-val before and after the time of sunrise and sunset. We stan-dardized observation times as hours relative to sunrise andsunset because, over the study period, sunrise occurredbetween 06:01 and 06:28 h, and sunset between 17:21 and17:55 h. We considered a lobster to be “in” the shelter ifany part of its body was within the shelter. Proportions ofshelter occupancy were arc-sine square-root transformed tomeet assumptions of normality and homogeneity of vari-ance (Zar 1999). Regression analyses were thus carried outon the transformed data as a function of time in hoursbefore (¡) and after (+) sunrise (Hsr), and before (¡) andafter (+) sunset (Hss).
The total amount of time each tethered lobster was in theshelter was also measured within half-hour intervals over a
Fig. 1 Image of shelter, camera, and red light lamp used in study. Thefour black lines represent the approximate range of movement of atethered lobster
123
956 Mar Biol (2008) 153:953–963
24 h period. These times, in minutes, were divided by 30 tocalculate the proportion of time each lobster occupied ashelter. These proportions were arc-sine square-root trans-formed. Because several lobsters were observed duringeach time interval, the transformed data were subjected toan analysis of regression with replication (Zar 1999) as afunction of time in hours before (¡) and after (+) sunrise(Hsr), and before (¡) and after (+) sunset (Hss). We alsocounted the number of excursions from the shelter per-formed by the 20 lobsters that remained in view throughoutan entire night and measured the total duration of eachexcursion.
Free-ranging lobsters, which were part of the existing nat-urally occurring population, also occasionally occupied theshelters. The shelter occupancy patterns of these untetheredlobsters were not included in our analysis. Separate observa-tions of the same predator at the same shelter occurringwithin 5 min of each other were counted as a single visit.
We also conducted an experiment to compare the behav-ior and survival of tethered lobsters that were able or notable to enter our shelters. Over a 7 day period (March 4–11,2003) we inserted sponge plugs into the openings of two ofthe cement building blocks and left the entrances open inthe other two blocks. The shelters that were open or closedwere selected daily using random numbers and the entranceplugs were changed each night accordingly. We hypothe-sized that the lobsters unable to enter the shelters wouldbe attacked and killed by predators more frequently thanlobsters able to enter the shelters. Because most attacks onlobsters during this period were conducted by triggerWsh
(see Sect. ”Results”), comparisons between lobsters insideand outside the shelters included the proportion of observa-tions in which lobsters were attacked by triggerWsh and theproportion of lobsters that survived a triggerWsh attack.These data were compared with two-sided Fisher exacttests (FET).
Results
Shelter occupancy patterns
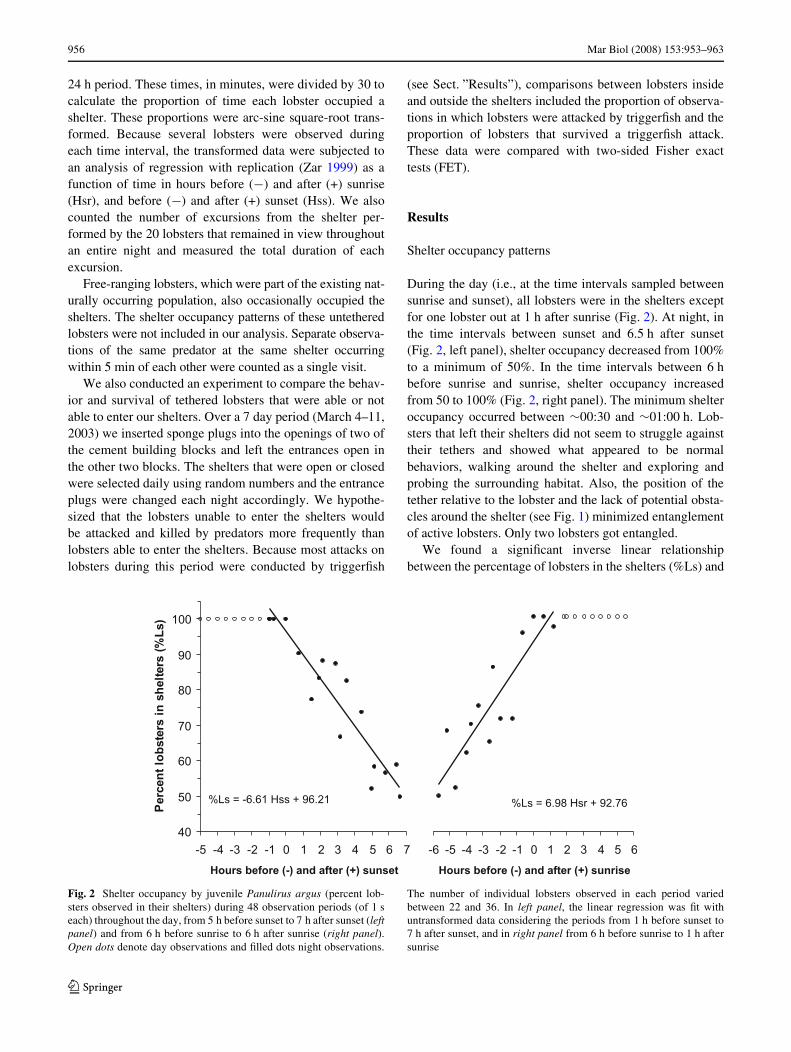
During the day (i.e., at the time intervals sampled betweensunrise and sunset), all lobsters were in the shelters exceptfor one lobster out at 1 h after sunrise (Fig. 2). At night, inthe time intervals between sunset and 6.5 h after sunset(Fig. 2, left panel), shelter occupancy decreased from 100%to a minimum of 50%. In the time intervals between 6 hbefore sunrise and sunrise, shelter occupancy increasedfrom 50 to 100% (Fig. 2, right panel). The minimum shelteroccupancy occurred between »00:30 and »01:00 h. Lob-sters that left their shelters did not seem to struggle againsttheir tethers and showed what appeared to be normalbehaviors, walking around the shelter and exploring andprobing the surrounding habitat. Also, the position of thetether relative to the lobster and the lack of potential obsta-cles around the shelter (see Fig. 1) minimized entanglementof active lobsters. Only two lobsters got entangled.
We found a signiWcant inverse linear relationshipbetween the percentage of lobsters in the shelters (%Ls) and
Fig. 2 Shelter occupancy by juvenile Panulirus argus (percent lob-sters observed in their shelters) during 48 observation periods (of 1 seach) throughout the day, from 5 h before sunset to 7 h after sunset (leftpanel) and from 6 h before sunrise to 6 h after sunrise (right panel).Open dots denote day observations and Wlled dots night observations.
The number of individual lobsters observed in each period variedbetween 22 and 36. In left panel, the linear regression was Wt withuntransformed data considering the periods from 1 h before sunset to7 h after sunset, and in right panel from 6 h before sunrise to 1 h aftersunrise
123
Mar Biol (2008) 153:953–963 957
the number of hours elapsed from 1 h before sunset to 6.5 hafter sunset (r2 = 0.788; df = 1, 13, P < 0.001). The resultantequation was: %Ls (in degrees) = 79.150 ¡ 5.297 £ h relativeto sunset. In addition, we found a signiWcant positive linearrelationship from 6 h before sunrise to 1 h after sunrise(r2 = 0.761; df = 1, 12; P < 0.001) and the respectiveequation was %Ls (in degrees) = 79.911 + 6.551 £ h relativeto sunrise. In Fig. 2, the least-squares best Wt lines for thepercentages (untransformed) of lobsters in the shelters arepresented as a function of hours relative to sunset (leftpanel), and as a function of hours relative to sunrise (rightpanel).
The absolute values of the slopes for the regressionlines of these relationships indicate that the rate at whichthe lobsters left the shelters during the Wrst half of thenight (6.6% per hour) was similar to the rate at which theyreturned to the shelters during the second half of the night(7.0% per hour). Calculations using the regression equa-tions suggest that those lobsters that left their sheltersstarted doing so at »35 min before sunset (Hss = ¡0.57)and Wnished returning to the shelters at »60 min after sun-rise (Hsr = 1.04).
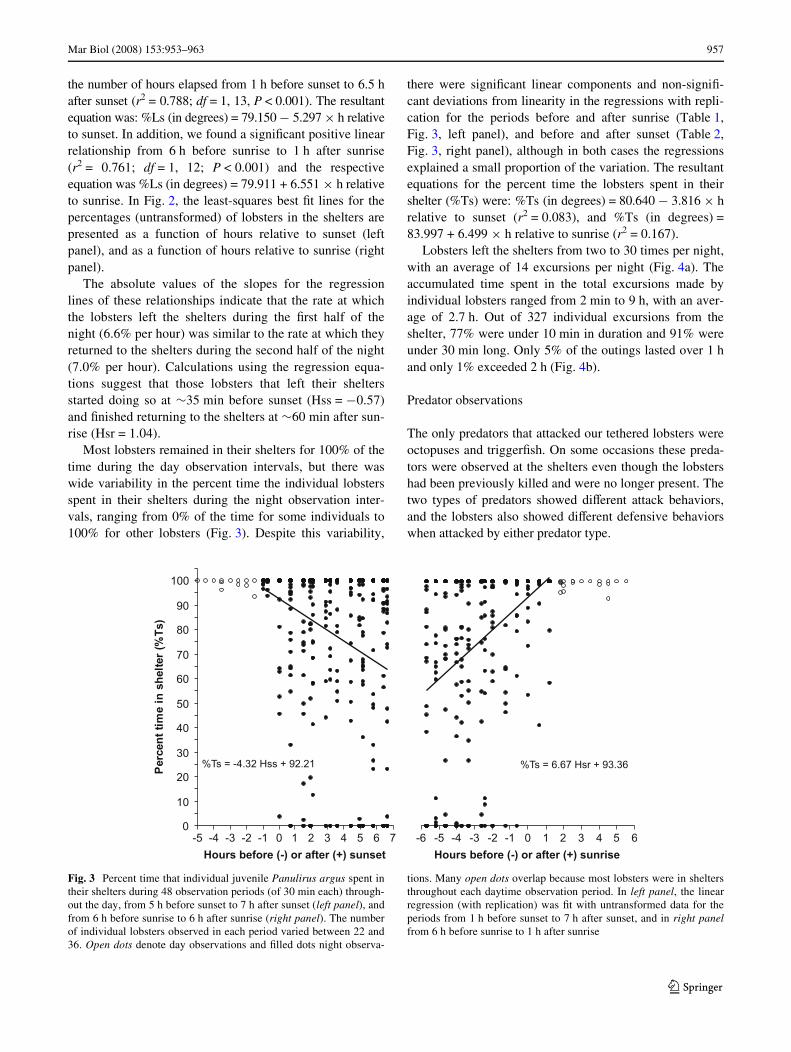
Most lobsters remained in their shelters for 100% of thetime during the day observation intervals, but there waswide variability in the percent time the individual lobstersspent in their shelters during the night observation inter-vals, ranging from 0% of the time for some individuals to100% for other lobsters (Fig. 3). Despite this variability,
there were signiWcant linear components and non-signiW-cant deviations from linearity in the regressions with repli-cation for the periods before and after sunrise (Table 1,Fig. 3, left panel), and before and after sunset (Table 2,Fig. 3, right panel), although in both cases the regressionsexplained a small proportion of the variation. The resultantequations for the percent time the lobsters spent in theirshelter (%Ts) were: %Ts (in degrees) = 80.640 ¡ 3.816 £ hrelative to sunset (r2 = 0.083), and %Ts (in degrees) =83.997 + 6.499 £ h relative to sunrise (r2 = 0.167).
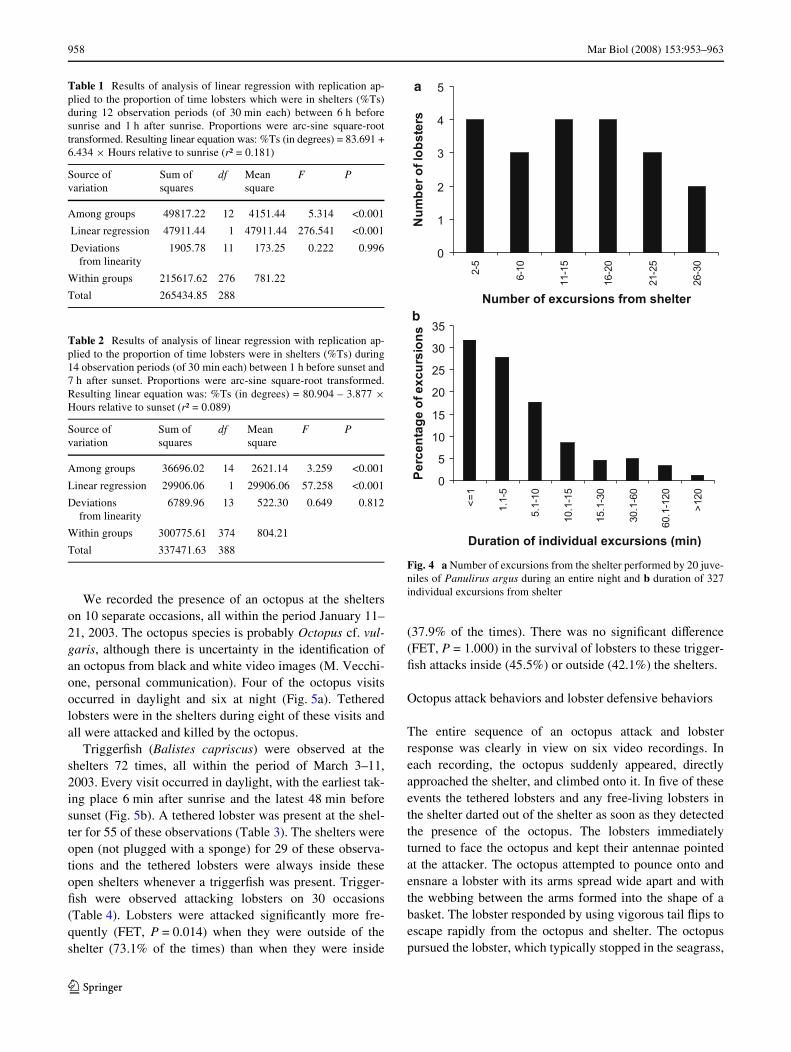
Lobsters left the shelters from two to 30 times per night,with an average of 14 excursions per night (Fig. 4a). Theaccumulated time spent in the total excursions made byindividual lobsters ranged from 2 min to 9 h, with an aver-age of 2.7 h. Out of 327 individual excursions from theshelter, 77% were under 10 min in duration and 91% wereunder 30 min long. Only 5% of the outings lasted over 1 hand only 1% exceeded 2 h (Fig. 4b).
Predator observations
The only predators that attacked our tethered lobsters wereoctopuses and triggerWsh. On some occasions these preda-tors were observed at the shelters even though the lobstershad been previously killed and were no longer present. Thetwo types of predators showed diVerent attack behaviors,and the lobsters also showed diVerent defensive behaviorswhen attacked by either predator type.
Fig. 3 Percent time that individual juvenile Panulirus argus spent intheir shelters during 48 observation periods (of 30 min each) through-out the day, from 5 h before sunset to 7 h after sunset (left panel), andfrom 6 h before sunrise to 6 h after sunrise (right panel). The numberof individual lobsters observed in each period varied between 22 and36. Open dots denote day observations and Wlled dots night observa-
tions. Many open dots overlap because most lobsters were in sheltersthroughout each daytime observation period. In left panel, the linearregression (with replication) was Wt with untransformed data for theperiods from 1 h before sunset to 7 h after sunset, and in right panelfrom 6 h before sunrise to 1 h after sunrise
123
958 Mar Biol (2008) 153:953–963
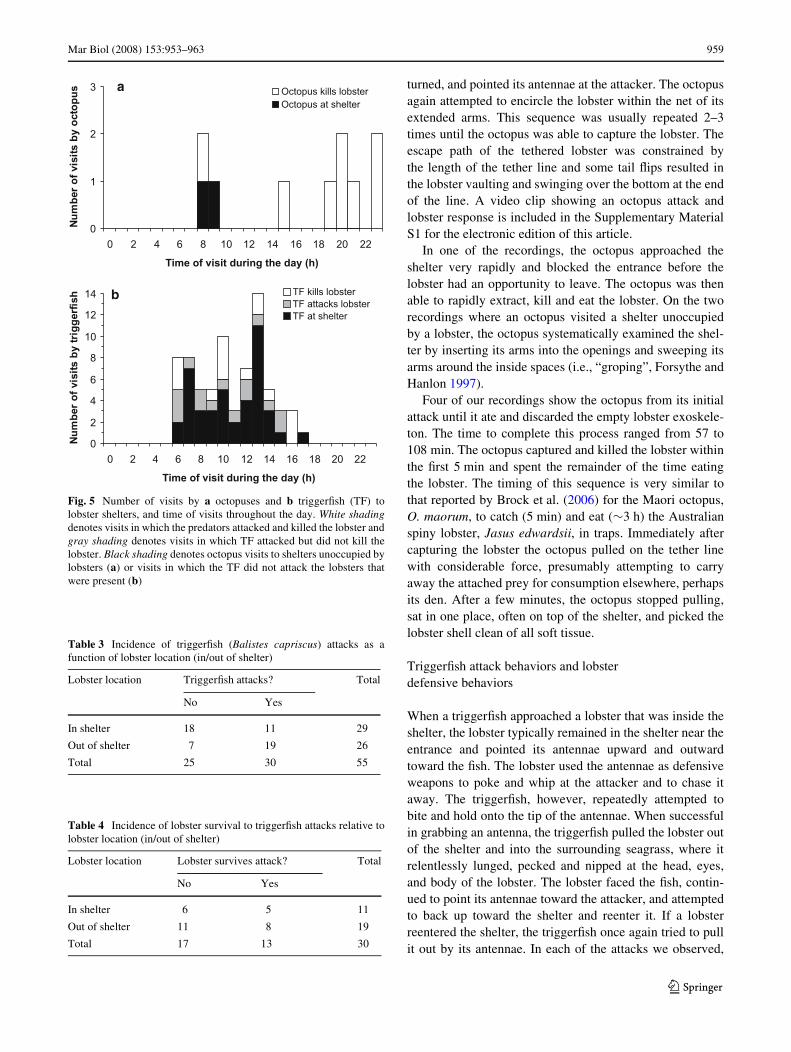
We recorded the presence of an octopus at the shelterson 10 separate occasions, all within the period January 11–21, 2003. The octopus species is probably Octopus cf. vul-garis, although there is uncertainty in the identiWcation ofan octopus from black and white video images (M. Vecchi-one, personal communication). Four of the octopus visitsoccurred in daylight and six at night (Fig. 5a). Tetheredlobsters were in the shelters during eight of these visits andall were attacked and killed by the octopus.
TriggerWsh (Balistes capriscus) were observed at theshelters 72 times, all within the period of March 3–11,2003. Every visit occurred in daylight, with the earliest tak-ing place 6 min after sunrise and the latest 48 min beforesunset (Fig. 5b). A tethered lobster was present at the shel-ter for 55 of these observations (Table 3). The shelters wereopen (not plugged with a sponge) for 29 of these observa-tions and the tethered lobsters were always inside theseopen shelters whenever a triggerWsh was present. Trigger-Wsh were observed attacking lobsters on 30 occasions(Table 4). Lobsters were attacked signiWcantly more fre-quently (FET, P = 0.014) when they were outside of theshelter (73.1% of the times) than when they were inside
(37.9% of the times). There was no signiWcant diVerence(FET, P = 1.000) in the survival of lobsters to these trigger-Wsh attacks inside (45.5%) or outside (42.1%) the shelters.
Octopus attack behaviors and lobster defensive behaviors
The entire sequence of an octopus attack and lobsterresponse was clearly in view on six video recordings. Ineach recording, the octopus suddenly appeared, directlyapproached the shelter, and climbed onto it. In Wve of theseevents the tethered lobsters and any free-living lobsters inthe shelter darted out of the shelter as soon as they detectedthe presence of the octopus. The lobsters immediatelyturned to face the octopus and kept their antennae pointedat the attacker. The octopus attempted to pounce onto andensnare a lobster with its arms spread wide apart and withthe webbing between the arms formed into the shape of abasket. The lobster responded by using vigorous tail Xips toescape rapidly from the octopus and shelter. The octopuspursued the lobster, which typically stopped in the seagrass,
Table 1 Results of analysis of linear regression with replication ap-plied to the proportion of time lobsters which were in shelters (%Ts)during 12 observation periods (of 30 min each) between 6 h beforesunrise and 1 h after sunrise. Proportions were arc-sine square-roottransformed. Resulting linear equation was: %Ts (in degrees) = 83.691 +6.434 £ Hours relative to sunrise (r² = 0.181)
Source of variation
Sum of squares
df Mean square
F P
Among groups 49817.22 12 4151.44 5.314 <0.001
Linear regression 47911.44 1 47911.44 276.541 <0.001
Deviations from linearity
1905.78 11 173.25 0.222 0.996
Within groups 215617.62 276 781.22
Total 265434.85 288
Table 2 Results of analysis of linear regression with replication ap-plied to the proportion of time lobsters were in shelters (%Ts) during14 observation periods (of 30 min each) between 1 h before sunset and7 h after sunset. Proportions were arc-sine square-root transformed.Resulting linear equation was: %Ts (in degrees) = 80.904 – 3.877 £Hours relative to sunset (r² = 0.089)
Source of variation
Sum of squares
df Mean square
F P
Among groups 36696.02 14 2621.14 3.259 <0.001
Linear regression 29906.06 1 29906.06 57.258 <0.001
Deviations from linearity
6789.96 13 522.30 0.649 0.812
Within groups 300775.61 374 804.21
Total 337471.63 388Fig. 4 a Number of excursions from the shelter performed by 20 juve-niles of Panulirus argus during an entire night and b duration of 327individual excursions from shelter
a
b
123
Mar Biol (2008) 153:953–963 959
turned, and pointed its antennae at the attacker. The octopusagain attempted to encircle the lobster within the net of itsextended arms. This sequence was usually repeated 2–3times until the octopus was able to capture the lobster. Theescape path of the tethered lobster was constrained bythe length of the tether line and some tail Xips resulted inthe lobster vaulting and swinging over the bottom at the endof the line. A video clip showing an octopus attack andlobster response is included in the Supplementary MaterialS1 for the electronic edition of this article.
In one of the recordings, the octopus approached theshelter very rapidly and blocked the entrance before thelobster had an opportunity to leave. The octopus was thenable to rapidly extract, kill and eat the lobster. On the tworecordings where an octopus visited a shelter unoccupiedby a lobster, the octopus systematically examined the shel-ter by inserting its arms into the openings and sweeping itsarms around the inside spaces (i.e., “groping”, Forsythe andHanlon 1997).
Four of our recordings show the octopus from its initialattack until it ate and discarded the empty lobster exoskele-ton. The time to complete this process ranged from 57 to108 min. The octopus captured and killed the lobster withinthe Wrst 5 min and spent the remainder of the time eatingthe lobster. The timing of this sequence is very similar tothat reported by Brock et al. (2006) for the Maori octopus,O. maorum, to catch (5 min) and eat (»3 h) the Australianspiny lobster, Jasus edwardsii, in traps. Immediately aftercapturing the lobster the octopus pulled on the tether linewith considerable force, presumably attempting to carryaway the attached prey for consumption elsewhere, perhapsits den. After a few minutes, the octopus stopped pulling,sat in one place, often on top of the shelter, and picked thelobster shell clean of all soft tissue.
TriggerWsh attack behaviors and lobster defensive behaviors
When a triggerWsh approached a lobster that was inside theshelter, the lobster typically remained in the shelter near theentrance and pointed its antennae upward and outwardtoward the Wsh. The lobster used the antennae as defensiveweapons to poke and whip at the attacker and to chase itaway. The triggerWsh, however, repeatedly attempted tobite and hold onto the tip of the antennae. When successfulin grabbing an antenna, the triggerWsh pulled the lobster outof the shelter and into the surrounding seagrass, where itrelentlessly lunged, pecked and nipped at the head, eyes,and body of the lobster. The lobster faced the Wsh, contin-ued to point its antennae toward the attacker, and attemptedto back up toward the shelter and reenter it. If a lobsterreentered the shelter, the triggerWsh once again tried to pullit out by its antennae. In each of the attacks we observed,
Fig. 5 Number of visits by a octopuses and b triggerWsh (TF) tolobster shelters, and time of visits throughout the day. White shadingdenotes visits in which the predators attacked and killed the lobster andgray shading denotes visits in which TF attacked but did not kill thelobster. Black shading denotes octopus visits to shelters unoccupied bylobsters (a) or visits in which the TF did not attack the lobsters thatwere present (b)
a
b
Table 3 Incidence of triggerWsh (Balistes capriscus) attacks as afunction of lobster location (in/out of shelter)
Lobster location TriggerWsh attacks? Total
No Yes
In shelter 18 11 29
Out of shelter 7 19 26
Total 25 30 55
Table 4 Incidence of lobster survival to triggerWsh attacks relative tolobster location (in/out of shelter)
Lobster location Lobster survives attack? Total
No Yes
In shelter 6 5 11
Out of shelter 11 8 19
Total 17 13 30
123

960 Mar Biol (2008) 153:953–963
the lobsters were able to reenter the shelter at least threetimes and sometimes up to 10 times. The lobster eventuallyappeared to become tired and perhaps weakened by the lossof blood from any wounds inXicted by the triggerWsh bites.Usually within 2–4 min after the initial attack, the lobsterbecame passive, no longer able to defend itself or to reenterthe shelter. The triggerWsh often continued to peck at theinactive lobster for 5–10 min after the lobster stoppedstruggling. Whenever a lobster was killed by a triggerWsh alarge aggregation of Wshes composed of a variety of specieswas attracted to the site and participated in a feedingfrenzy. A video clip showing a triggerWsh attack and lobsterresponse is included in the Supplementary Material S2 forthe electronic edition of this article.
Lobsters that were outside the closed shelters (withplugged openings) typically stayed next to the sides of thecement blocks during the daylight hours. When a triggerWshWrst appeared in view, the lobster usually remained inac-tive, perhaps depending on being inconspicuous and unno-ticed by the triggerWsh. However, if the triggerWsh attackedthe lobster, it actively defended itself in the same manner asdescribed above for the lobsters that were pulled out of theunplugged shelters. A similar attack sequence of triggerWshon lobsters without access to shelter was described by Bar-shaw et al. (2003) and Briones-Fourzán et al. (2006). Thelobster often attempted to return to be next to the cementblock if the triggerWsh pulled it away.
All of the lobsters under attack by triggerWsh seemed toprefer to return to the shelters rather than to move into thecover provided by the surrounding seagrass beds. They alsopredominantly used their antennae to defend against thetriggerWsh attacks and rarely attempted to escape from theWsh by tailXipping.
Discussion and conclusions
As expected, juvenile P. argus remained in their sheltersthroughout the daytime, but surprisingly, shelter occupancydecreased linearly from 100% shortly before sunset to 50%shortly after midnight and then increased linearly untilreaching 100% shortly after sunrise. The percent time thelobsters spent in the shelters during the night followed asimilar trend, but varied widely for diVerent individuals:from 0–100%. Furthermore, they left and returned to theirshelter 2–30 times night¡1 with a majority of excursionslasting <10 min. These results suggest that juvenile P.argus remain in their shelters as much as possible —even inthe middle of the night— to minimize exposure to preda-tors, but that they oVset the energetic cost of remaining intheir shelters by foraging close to their shelters for severalshort periods during the night rather than for one longerperiod immediately after dusk. Some individuals may
accept greater amounts of risk than others (Lima and Dill1990; Wahle 1992), reXecting individual diVerences in stateof hunger, size, reaction to handling, stage in molt cycle,and perception of predation risk, as well as diVerences inthe near-shelter food supply. This pattern was also observedby Segura-García et al. (2004) among individuals ofP. guttatus, a reef-obligate species that is more vulnerableto predation than coexisting reef-dwelling individuals ofP. argus (Lozano-Álvarez et al. 2007).
Out-of-shelter activity of juvenile lobsters did notabruptly increase at dusk, remain constantly high during thenight, and decrease abruptly at dawn. Thus, shelter occu-pancy was not a simple response to a threshold light inten-sity. Instead, the minimum shelter occupancy in the middleof the night suggests that the selective pressure of diurnaland crepuscular (active at dawn and dusk) predators makesa strong contribution to the shelter occupancy pattern ofjuvenile P. argus. Weiss et al. (2006) found that these pre-dators were responsible for over 70% of the mortality ofjuvenile lobsters tethered outside of shelters. Other factors,such as the periodicity of lobster prey availability and noc-turnal predator activity, may also contribute to this pattern.We found, for example, that lobsters actually prefer tovacate their shelters in the presence of octopuses. Thus,being outside of the shelter at night may confer the dualadvantage of allowing the lobster to forage and to minimizeoctopus predation.
The shelter occupancy pattern shown by our juvenilelobsters may reXect their level of perceived predation risk.Díaz et al. (2005) found that predation on juvenile spinylobsters was more intense during the hours of daylight thanat night and Oliver et al. (2005) found that it was moreintense during crepuscular periods, times of transitionbetween diurnal and nocturnal Wsh predators. The shelteroccupancy pattern of our juvenile P. argus suggests thatthey perceived a lower predation risk at night than duringcrepuscular or diurnal periods. However, activity patterns ofjuvenile P. argus changed from exclusively nocturnal tonocturnal/diurnal after about 2 weeks of captivity in enclo-sures without predators (Lozano-Álvarez 1996), suggestinga decline in perception of risk by those lobsters. We hypoth-esize that these declines in perceived risk may be due, inpart, to the immediate cessation of direct visual and tactilealarm signals as well as a gradual reduction in the chemicalcues remaining after predators are no longer nearby.
Spiny lobsters undergo an ontogenetic shift in their habi-tat selection, social interactions, and activity patterns asthey make the transition from the early benthic phase(EBP), living in the vegetation of shallow lagoons, to cre-vice-sheltering juveniles, and Wnally to adults living incoral reefs (Herrnkind et al. 1975; Childress and Herrnkind,1994). The shelter occupancy pattern we found for juvenileP. argus was intermediate to that of EBP and mature
123

Mar Biol (2008) 153:953–963 961
lobsters. Whereas recently settled P. argus rarely leave theirshelters, dwelling and feeding inside the interstices of mac-roalgae (Smith and Herrnkind, 1992), most of the juvenilelobsters in our study left the shelters for at least some timeduring the night. Moreover, unlike larger P. argus, whichmay leave their shelter during daylight hours (Lipcius et al.1983; Lozano-Álvarez and Spanier 1997; Lozano-Álvarezand Briones-Fourzán 2001), our experimental juvenilesoccupied their shelters throughout the daytime and the min-imum shelter occupancy of juveniles at night was 50%,contrasting with minimum shelter occupancies of 0–30%recorded for larger subadult and adult P. argus (Lozano-Álvarez and Spanier 1997; Lozano-Álvarez and Briones-Fourzán 2001). Adult lobsters invariably show a sunset/early evening peak in activity (Kanciruk and Herrnkind1973; Herrnkind et al. 1975; Lipcius and Herrnkind 1982;Lozano-Álvarez and Briones-Fourzán 2001). Instead, wefound that the out-of-shelter activity of juveniles peakedshortly after midnight. These ontogenetic changes in theshelter occupancy pattern may reXect a decrease in per-ceived predation risk as the lobsters increase in size.
Kanciruk and Herrnkind (1973) suggested that the earlyevening activity peak they observed was associated withincreased social interactions among lobsters. Our investiga-tion was conducted using a single lobster tethered at eachshelter. Although free-living lobsters also occasionallyoccupied the shelters, social interactions were infrequentand this may be an additional factor to account for theabsence of an evening activity peak.
In multipredator environments, variations in how preda-tors forage may aVect antipredator decision-making by prey(Sih et al. 1998; Lima 2002). Also, predator behavior, inconjunction with other factors such as the scaling betweenshelter size and lobster size (Eggleston and Lipcius 1992;Mintz et al. 1994) and the chemical environment, maydetermine the refuge value of a particular shelter. Forexample, free-ranging individuals of P. argus tend to avoidshelters with chemical scents of octopuses (Berger andButler 2001; Horner et al. 2006) and, in our study, tetheredlobsters usually vacated the shelters upon the approach of anoctopus. In South Africa, Berry (1971) observed an identi-cal response in sheltered P. homarus when approached byan octopus (O. granulosus), with the lobsters ultimatelyescaping by tailXipping if the octopus advanced. Thisescape response was constrained in our lobsters by thelength of the tether, but Lozano-Álvarez et al. (unpub-lished) observed that octopuses were usually unable tocatch free-ranging lobsters in large experimental tanks andwe observed that an octopus was adept at using its arms toextract a lobster from a shelter. By contrast, octopusesquickly subdued and killed lobsters constrained in traps(Brock et al. 2006). Thus, the chances for lobsters to sur-vive an octopus attack may be better outside than inside a
shelter and their behavioral responses to the octopus mayhave evolved accordingly, avoiding shelters with octopusscents and vacating shelters when an octopus approaches.
In contrast, lobsters remained in the shelters when a trig-gerWsh was present and repeatedly attempted to return tothe shelter if pulled out by the Wsh. A triggerWsh was nearlytwice as likely to attack a lobster that was outside of theshelter than a lobster inside; thus, lobsters in the open maybe more visible to a triggerWsh than lobsters inside a refuge.Surprisingly, once under attack by a triggerWsh, a lobsterhad a nearly equal chance of surviving the attack if it wasinside or outside the shelter. The triggerWsh were veryskilled at removing lobsters from the shelters by pulling ontheir antennae. The shelter openings were large enough sothat sometimes a triggerWsh would poke its head partiallyinto the den to grab the lobster’s antennae, but this was usu-ally unnecessary. Also, although the shelters were deepenough for a lobster to fully retract inside, the lobsters typi-cally remained near the shelter entrance where they weremore able to use their antennae as defensive weapons, butthis also made them more readily accessible to the Wsh.Thus, the failure of the shelters to enable the lobsters tofully avoid triggerWsh and octopus predation was probablydue to the shelter dimensions, the behavior of both the lob-sters and the predators, and the tethering technique. In par-ticular, although the tether did not prevent our lobsters fromentering the shelters, it did constrain the distance to whichthe lobsters could escape. Consequently, tethering had amuch greater impact on the ability of P. argus to avoid pre-dation by the octopus than the triggerWsh.
All the triggerWsh visits to our shelters occurred duringthe day, whereas six of the octopus visits to our sheltersoccurred during the night and four during the day. Theseobservations, although few, are consistent with other recentstudies, which suggest that octopuses are active in the dayand night (Meisel et al. 2003; Brock et al. 2006). Becauseoctopuses use chemotactile rather than visual search meth-ods to Wnd prey (Forsythe and Hanlon 1997; Brock et al.2006), there may not be a strong selection for either diurnalor nocturnal activity. In addition, octopuses learn the loca-tion of prey (Forsythe and Hanlon 1997) and may use refer-ence memory for places where food might be found(Hochner et al. 2006). Reef Wshes also appear to use cogni-tive maps to get access to their prey and triggerWshes inparticular can remember speciWc food items (Bshary et al.2002). The octopus predation we observed was concen-trated in about a 2-week time period. The triggerWsh preda-tion events also occurred within a few days of each other.Based on size and general appearance, we suspect that eachof these predator species may have been represented by asingle individual that returned on consecutive days to ourshelters. If so, this would suggest that these predatorsquickly learned where to go to Wnd a lobster to prey upon.
123

962 Mar Biol (2008) 153:953–963
We observed that crevice-dwelling juvenile P. argusexhibited a circadian shelter occupancy pattern diVerentfrom and intermediate to that reported for early benthicjuveniles and adults. This pattern may be modulated by theinterplay between the energetic demands of these juvenilesand their level of perception of predation risk. Thus, juve-nile P. argus tend to remain in their shelter for as muchtime as possible and to oVset the energetic cost of avoidingpredation by foraging close to their shelters for severalshort periods at night, with some individuals acceptingmore risk than others. Our observations also show how thedefensive behavior of juvenile P. argus varies in reaction todiVerent predator species.
Acknowledgments This work was funded in part by a J. W. Ful-bright Senior Scholar Grant and by Universidad Nacional Autónomade México (UNAM). Fernando Negrete-Soto and Cecilia Barradas-Ortiz provided invaluable technical assistance throughout this study.Michael Vecchione of the National Museum of Natural History identi-Wed the species of octopus in the videos. The experiments comply withthe current laws of Mexico. Annual permits to collect and use spinylobsters were issued by Comisión Nacional de Acuacultura y Pesca,México.
References
Acosta CA, Butler MJ IV (1997) Role of mangrove habitat as a nurseryfor juvenile spiny lobster, Panulirus argus, in Belize. Mar FreshwRes 48:721–727
Barshaw DE, Able KW (1990) Tethering as a technique for assessingpredation rates in diVerent habitats: an evaluation using juvenilelobsters Homarus americanus. Fish Bull 88:415–417
Barshaw DE, Lavalli KL, Spanier E (2003) OVense versus defense:responses of three morphological types of lobsters to predation.Mar Ecol Prog Ser 256:171–182
Berger DK, Butler MJ IV (2001) Octopuses inXuence den selection byjuvenile Caribbean spiny lobster. Mar Freshw Res 52:1049–1053
Berry PF (1971) The spiny lobsters (Palinuridae) of the east coast ofSouthern Africa: distribution and ecological notes. Invest RepOceanogr Res Inst Durban 27:1–23
Briones-Fourzán P, Lozano-Álvarez E (2001) EVects of artiWcial shel-ters (Casitas) on the abundance and biomass of juvenile spiny lob-sters Panulirus argus in a habitat-limited tropical reef lagoon.Mar Ecol Prog Ser 221:221–232
Briones-Fourzán P, Castañeda-Fernández de Lara V, Lozano-ÁlvarezE, Estrada-Olivo J (2003) Feeding ecology of the three juvenilephases of the spiny lobster Panulirus argus in a tropical reeflagoon. Mar Biol 142:855–865
Briones-Fourzán P, Pérez-Ortiz M, Lozano-Álvarez E (2006) Defensemechanisms and antipredator behavior in two sympatric speciesof spiny lobster, Panulirus argus and P. guttatus. Mar Biol149:227–239
Briones-Fourzán P, Lozano-Álvarez E, Negrete-Soto F, Barradas-Ortiz C (2007) Enhancement of juvenile Caribbean spiny lobsters:an evaluation of changes in multiple response variables with theaddition of large artiWcial shelters. Oecologia 151:401–416
Brock DJ, Saunders TM, Ward TM, Linnane AJ (2006) A two-cham-bered trap reduces within-trap predation by octopus on rock lob-sters in aquarium trials. Fish Res 80:129–135
Bshary R, Wickler W, Fricke H (2002) Fish cognition: a primate’s eyeview. Anim Cogn 5:1–13
Childress MJ, Herrnkind WF (1994) The behavior of juvenile Carib-bean spiny lobster in Florida Bay: seasonality, ontogeny and soci-ality. Bull Mar Sci 54:819–827
Childress MJ, Herrnkind WF (2001) The guide eVect inXuence on thegregariousness of juvenile Caribbean spiny lobsters. Anim Behav62:465–472
Díaz D, Zabala M, Linares C, Hereu B, Abelló P (2005) Increased pre-dation of juvenile European spiny lobster (Palinurus elephas) ina marine protected area. N Z J Mar Freshw Res 39:447–453
Eggleston DB, Lipcius RN (1992) Shelter selection by spiny lobstersunder variable predation risk, social conditions, and shelter size.Ecology 73:992–1011
Eggleston DB, Lipcius RN, Miller DL (1992) ArtiWcial shelters and thesurvival of juvenile Caribbean spiny lobster Panulirus argus:Spatial, habitat, and lobster size eVects. Fish Bull 90:691–702
Forsythe JW, Hanlon RT (1997) Foraging and associated behavior byOctopus cyanea Gray, 1849 on a coral atoll, French Polynesia.J Exp Mar Biol Ecol 209:15–31
Glaholt RD (1990) Social behaviour and habitat use of captive juvenilespiny lobster, Panulirus argus (Latreille, 1804) (Decapoda, Pali-nuridae). Crustaceana 58:200–206
Helfman GS (1986) Fish behavior by day, night, and twilight. In:Pitcher TJ (ed) The behavior of teleost Wshes. Croom-Helm, Lon-don, pp 366–387
Herrnkind WF (1980) Movement patterns in Palinurid lobsters. In:Cobb JS, Phillips BF (eds) The biology and management of lob-sters, vol 1. Academic, New York, pp 349–407
Herrnkind WF, Vanderwalker JA, Barr L (1975) Population dynamics,ecology and behavior of spiny lobsters Panulirus argus from StJohn, U.S.V.I. IV: habitation, patterns of movement, and generalbehavior. Nat Hist Mus Los Ang Cty Sci Bull 20:31–45
Hobson ES (1965) Diurnal-nocturnal activity of some inshore Wshes inthe Gulf of California. Copeia 3:291–302
Hochner B, Shomrat T, Fiorito G (2006) The octopus: a model forcomparative analysis of the evolution of learning and memorymechanisms. Biol Bull 210:308–317
Horner AJ, Nickles SP, Weissburg MJ, Derby CD (2006) Source andspeciWcity of chemical cues mediating shelter preference ofCaribbean spiny lobsters (Panulirus argus). Biol Bull 211:128–139
Kanciruk P, Herrnkind WF (1973) Preliminary investigations of thedaily and seasonal locomotor activity rhythms of the spiny lob-ster, Panulirus argus. Mar Behav Physiol 1:351–359
Lavalli KL, Barshaw DE (1989) Post-larval American lobsters (Hom-arus americanus) living in burrows may be suspension feeding.Mar Behav Physiol 15:255–264
Lima SL (2002) Putting predators back into behavioral predator–preyinteractions. Trends Ecol Evol 17:70–75
Lima SL, Dill LM (1990) Behavioral decisions made under the risk ofpredation: a review and prospectus. Can J Zool 68:619–640
Lipcius RN, Herrnkind WF (1982) Molt cycle alterations in behavior,feeding and diel rhythms of a decapod crustacean, the spiny lob-ster Panulirus argus. Mar Biol 68:241–252
Lipcius RN, Edwards ML, Herrnkind WF, Waterman SA (1983) In situmating behavior of the spiny lobster Panulirus argus. J Crust Biol3:217–222
Lozano-Álvarez E (1996) Ongrowing of juvenile spiny lobsters, Pan-ulirus argus (Latreille, 1804) (Decapoda, Palinuridae), in portablesea enclosures. Crustaceana 69:958–973
Lozano-Álvarez E, Briones-Fourzán P (2001) Den choice and occupa-tion patterns of shelters by two sympatric lobster species, Panuli-rus argus and Panulirus guttatus, under experimental conditions.Mar Freshwater Res 52:1145–1155
Lozano-Álvarez E, Spanier E (1997) Behaviour and growth of captivespiny lobsters (Panulirus argus) under the risk of predation. MarFreshwater Res 48:707–713
123

Mar Biol (2008) 153:953–963 963
Lozano-Álvarez E, Briones-Fourzán P, Osorio-Arciniegas A, Negrete-Soto F, Barradas-ortiz C (2007) Coexistence of congeneric spinylobsters on coral reefs: diVerential use of shelter resources andvulnerability to predators. Coral Reefs 26:361–373
Meisel DV, Byrne RA, Kuba M, Griebel U, Mather JA (2003) Circa-dian rhythms in Octopus vulgaris. In: Warnke K, Keupp H, Bole-tzky SV (eds) Coleoid cephalopods through time, vol 3. BerlinerPaläobiol Abh, Berlin, pp 171–177
Mintz JD, Lipcius RN, Eggleston DB, Seebo MS (1994) Survival ofjuvenile Caribbean spiny lobster: eVects of shelter size, geo-graphic location and conspeciWc abundance. Mar Ecol Prog Ser112:255–266
Ogden JC, Ziemann JC (1977) Ecological aspects of coral reef-seagrass bed contacts in the Caribbean. Proceedings of the 3rdinternational coral reef symposium, Miami, pp 377–382
Oliver MD, Stewart R, Mills D, MacDiarmid AB, Gardner C (2005)Stock enhancement of rock lobsters (Jasus edwardsii): timing ofpredation on naïve juvenile lobsters immediately after release.N Z J Mar Freshw Res 39:391–397
Phillips BF, Cobb JS, George RW (1980) General biology. In: CobbJS, Phillips BF (eds) The biology and management of lobsters, vol1. Academic, New York, pp 1–72
Robblee MB, Ziemann JC (1984) Diel variation in the Wsh fauna of atropical seagrass feeding ground. Bull Mar Sci 34:335–345
Segura-García I, Lozano-Álvarez E, Briones-Fourzán P (2004) With-in-shelter behavior of the spotted spiny lobster Panulirus guttatus(Latreille), in simulated communal dens: an exploratory study.Mar Freshw Behav Physiol 37:17–30
Sih A (1980) Optimal foraging: can foragers balance two conXictingdemands? Science 210:1041–1043
Sih A (1987) Predators and prey lifestyles: an evolutionary and ecolog-ical overview. In: Kerfoot WC, Sih A (eds) Predation: direct andindirect impacts on aquatic communities. University Press ofNew England, Hanover, pp 203–224
Sih A, Englund G, Wooster D (1998) Emergent impacts of multiplepredators on prey. Trends Ecol Evol 13:350–355
Smith KN, Herrnkind WF (1992) Predation on early juvenile spinylobsters Panulirus argus (Latreille): inXuence of size and shelter.J Exp Mar Biol Ecol 157:3–18
Starck WA II, Davis WP (1966) Night habits of Wshes on AlligatorReef, Florida. Ichthyologica 38:313–355
SutcliVe WH Jr (1956) EVect of light intensity on the activity of theBermuda spiny lobster, Panulirus argus. Ecology 37:200–201
Vannini M, Cannicci S (1995) Homing behaviour and possible cogni-tive maps in crustacean decapods. J Exp Mar Biol Ecol 193:67–91
Wahle RA (1992) Body-size dependent anti-predator mechanisms ofthe American lobster. Oikos 65:52–60
Weiss HM, Lozano-Álvarez E, Briones-Fourzán P, Negrete-Soto F(2006) Using red light with Wxed-site video cameras to study thebehavior of the spiny lobster, Panulirus argus, and associated ani-mals at night and inside their shelters. Mar Tech Soc J 40(3):86–95
Werner EE, Gilliam JF (1984) The ontogenetic niche and species inter-actions in size-structured populations. Annu Rev Ecol Syst15:393–425
Zar JH (1999) Biostatistical analysis, 4th edn. Prentice-Hall, UpperSaddle River
123
Related Documents











