Respiration Physiology 126 (2001) 233 – 243 Circadian patterns in men acclimatized to intermittent hypoxia Manuel Vargas a , Daniel Jime ´nez b , Fabiola Leo ´ n-Velarde c , Jorge Osorio b , Jacopo P. Mortola d, * a Centro de Inestigacio ´n en Medicina de Altura, Mutual de Seguridad C. Ch. C., Iquique, Chile b Compan ˜ia Minera Don ˜a Ine ´s de Collahuasi, Iquique, Chile c Uniersitad Cayetano Heredia, Departamento de Ciencias Fı ´siologicas, I. I. A., Lima, Peru d Department of Physiology, McGill Uniersity, 3655 Drummond Street, Montreal Quebec, Canada H3G 1Y6 Accepted 28 March 2001 Abstract Six men, normally working shifts of 7 days at high altitude (HA, 3800 m, 480 mm Hg barometric pressure) followed by 7 days of rest at sea level (SL), were studied during the last days of their HA and SL shifts with a 24-h constant routine protocol of sustained wakefulness and minimal activity. The amplitude of the circadian oscillations of oxygen consumption, breathing rate, thoracic skin temperature and diastolic pressure did not differ between HA and SL. At HA, the amplitude of the tympanic and calf temperature oscillations, were, respectively, lower and higher than at SL. End-tidal P CO 2 and systolic pressure had larger amplitude oscillations at HA than at SL. Hence, also in humans, as previously shown in animals, hypoxia can affect some circadian patterns, including those involved in thermoregulation. These effects of hypoxia could contribute to sleep disturbances at HA and in patients with cardiorespiratory diseases. © 2001 Elsevier Science B.V. All rights reserved. Keywords: Altitude, high, circadian oscillations; Hypoxia, chronic; Mammals, humans; Rhythm, circadian; Temperature, body, high altitude; Thermoregulation, high altitude www.elsevier.com/locate/resphysiol 1. Introduction The depressant effect of hypoxia on metabolic rate can occur within minutes, and its mechanisms and the implications on the control of breathing have attracted much attention over the last few years (Gautier, 1996). In young and adult mam- mals thermogenesis is blunted by hypoxia, with a lowering of the set point of thermoregulation (Mortola and Gautier, 1995). The metabolic re- sponse to cold is also depressed in adult high- landers (Blatteis and Lutherer, 1976) and infants born at high altitude (Frappell et al., 1998). Both oxygen consumption (V O 2 ) and body tem- perature (Tb) present daily oscillations. These cir- cadian patterns in the balance between heat * Corresponding author. Tel.: +1-514-3984335; fax +1- 514-3987452. E-mail address: [email protected] (J.P. Mortola). 0034-5687/01/$ - see front matter © 2001 Elsevier Science B.V. All rights reserved. PII:S0034-5687(01)00226-2

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Respiration Physiology 126 (2001) 233–243
Circadian patterns in men acclimatized to intermittenthypoxia
Manuel Vargas a, Daniel Jimenez b, Fabiola Leon-Velarde c, Jorge Osorio b,Jacopo P. Mortola d,*
a Centro de In�estigacion en Medicina de Altura, Mutual de Seguridad C. Ch. C., Iquique, Chileb Compania Minera Dona Ines de Collahuasi, Iquique, Chile
c Uni�ersitad Cayetano Heredia, Departamento de Ciencias Fısiologicas, I. I. A., Lima, Perud Department of Physiology, McGill Uni�ersity, 3655 Drummond Street, Montreal Quebec, Canada H3G 1Y6
Accepted 28 March 2001
Abstract
Six men, normally working shifts of 7 days at high altitude (HA, 3800 m, �480 mm Hg barometric pressure)followed by 7 days of rest at sea level (SL), were studied during the last days of their HA and SL shifts with a 24-hconstant routine protocol of sustained wakefulness and minimal activity. The amplitude of the circadian oscillationsof oxygen consumption, breathing rate, thoracic skin temperature and diastolic pressure did not differ between HAand SL. At HA, the amplitude of the tympanic and calf temperature oscillations, were, respectively, lower and higherthan at SL. End-tidal PCO2
and systolic pressure had larger amplitude oscillations at HA than at SL. Hence, also inhumans, as previously shown in animals, hypoxia can affect some circadian patterns, including those involved inthermoregulation. These effects of hypoxia could contribute to sleep disturbances at HA and in patients withcardiorespiratory diseases. © 2001 Elsevier Science B.V. All rights reserved.
Keywords: Altitude, high, circadian oscillations; Hypoxia, chronic; Mammals, humans; Rhythm, circadian; Temperature, body, highaltitude; Thermoregulation, high altitude
www.elsevier.com/locate/resphysiol
1. Introduction
The depressant effect of hypoxia on metabolicrate can occur within minutes, and its mechanismsand the implications on the control of breathinghave attracted much attention over the last few
years (Gautier, 1996). In young and adult mam-mals thermogenesis is blunted by hypoxia, with alowering of the set point of thermoregulation(Mortola and Gautier, 1995). The metabolic re-sponse to cold is also depressed in adult high-landers (Blatteis and Lutherer, 1976) and infantsborn at high altitude (Frappell et al., 1998).
Both oxygen consumption (V� O2) and body tem-
perature (Tb) present daily oscillations. These cir-cadian patterns in the balance between heat
* Corresponding author. Tel.: +1-514-3984335; fax +1-514-3987452.
E-mail address: [email protected] (J.P. Mortola).
0034-5687/01/$ - see front matter © 2001 Elsevier Science B.V. All rights reserved.
PII: S 0034 -5687 (01 )00226 -2

M. Vargas et al. / Respiration Physiology 126 (2001) 233–243234
production and heat loss occur even withoutchanges in activity levels (Refinetti and Menaker,1992; Decoursey et al., 1998), and could be at theorigin of numerous autonomic and behaviouralcircadian patterns (Refinetti and Menaker, 1992).Since hypoxia reduces thermogenesis, it wouldseem probable that also the circadian patterns ofTb and V� O2
could be depressed in hypoxic condi-tions. Indeed, the morning–night differences inTb and V� O2
were not observed in newborn rats inacute hypoxia (Saiki and Mortola, 1995). In adultrats, sustained hypoxia greatly reduced the ampli-tudes of the circadian oscillations of V� O2
and Tb(Mortola and Seifert, 2000). The same was ob-served in rats after denervation of the carotid andaortic chemoreceptors, suggesting that hypoxiamay act directly on the central nervous system,presumably at the level of the hypothalamic ther-moregulatory centers (Fenelon et al., 2000).
Data on the effect of hypoxia on circadianpatterns in humans are not available. It wouldseem important to extend to humans the studiespreviously performed on animals, because dailychanges in metabolism and Tb are believed to beassociated with changes in many body functions,including the duration and organization of sleep(Czeisler et al., 1980; Zulley et al., 1981). Hence,in this study we monitored tympanic and skintemperatures, in addition to some parameters ofcardiorespiratory functions, in six adult men athigh altitude (HA) and at sea level (SL), to testthe hypothesis of a difference between normoxicand hypoxic circadian patterns.
2. Methods
The experiments were conducted on six subjectsin the North of Chile. Tympanic and skin temper-atures (thoracic and calf), oxygen consumption,arterial oxygen saturation, blood pressure, andthe end-tidal partial pressure of CO2 were themain parameters measured either continuously orat fixed time-intervals for the 24 h of the ‘constantroutine’ protocol (see below). Measurements wereobtained at HA and SL. The HA location was aCollahuasi mining center, at �3800 m (baromet-ric pressure, Pb, �480 mm Hg, which corre-sponds to an inspired fractional concentration of�13% O2 in normobaric conditions), the SL loca-tion was Iquique (Pb�760 mm Hg).
2.1. Subjects and general protocol
The general characteristics of the six subjectsparticipating in the study are given in Table 1. Allof them had been exposed to chronic intermittenthypoxia by working at HA (between 3800 and4000 m) for periods of 1.5–4 years, with a 7 by 7pattern, i.e. 7 days of work at HA and 7 days ofrest at SL. Measurements were obtained duringthe last day of the high-altitude shift and duringthe last day of the week immediately following atSL. No attempt was made to control the level ofactivity during the days preceding the recordingsession.
The constant routine protocol (Mills et al.,1978) started at 20:00 h and terminated at 20:00 h
Table 1Characteristics of the six subjects
Age (years)Subject’s initials Weight (kg) Height (cm) Work at high altitude (years) Smoking habits
AT 44 68 167 3 10 per day34 70MA 160 4
CS �2 per day2169763832 63GP 164 1.5 �2 per day38 74HT 170 4 no smoking37 69 166 1.5 �2 per day
166 2.7Mean no smoking37 701SEM 0.52 2
M. Vargas et al. / Respiration Physiology 126 (2001) 233–243 235
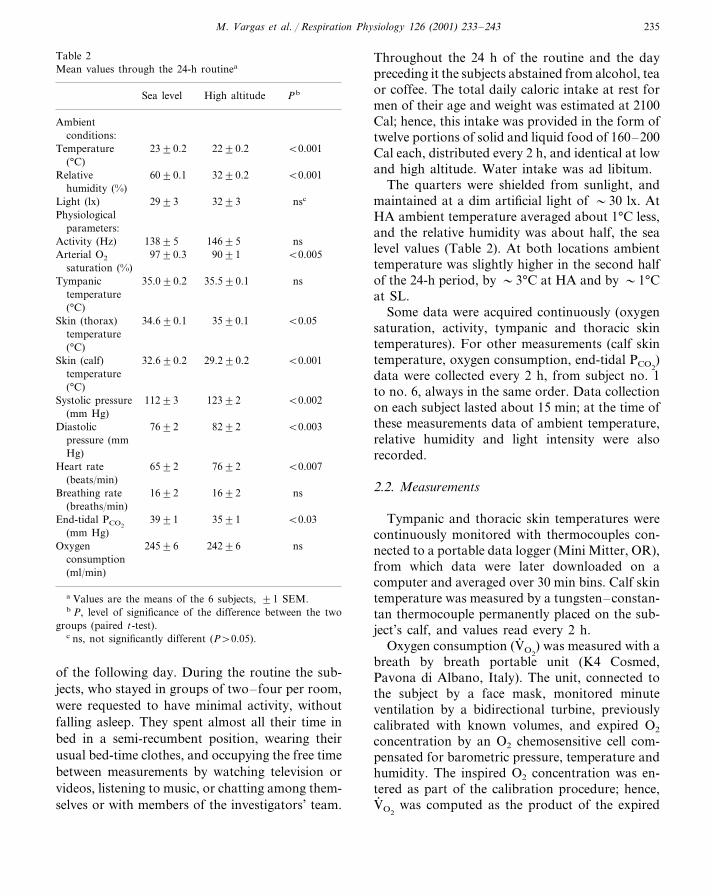
Table 2Mean values through the 24-h routinea
High altitude PbSea level
Ambientconditions:
23�0.2Temperature 22�0.2 �0.001(°C)
60�0.1Relative 32�0.2 �0.001humidity (%)
32�3Light (lx) nsc29�3Physiological
parameters:146�5Activity (Hz) ns138�5
97�0.3Arterial O2 90�1 �0.005saturation (%)
Tympanic 35.0�0.2 ns35.5�0.1temperature(°C)
Skin (thorax) 34.6�0.1 �0.0535�0.1temperature(°C)
�0.00132.6�0.2Skin (calf) 29.2�0.2temperature(°C)
Systolic pressure 112�3 123�2 �0.002(mm Hg)
76�2 82�2 �0.003Diastolicpressure (mmHg)
65�2Heart rate 76�2 �0.007(beats/min)
Breathing rate 16�2 16�2 ns(breaths/min)
39�1 35�1 �0.03End-tidal PCO2
(mm Hg)245�6 242�6Oxygen ns
consumption(ml/min)
a Values are the means of the 6 subjects, �1 SEM.b P, level of significance of the difference between the two
groups (paired t-test).c ns, not significantly different (P�0.05).
Throughout the 24 h of the routine and the daypreceding it the subjects abstained from alcohol, teaor coffee. The total daily caloric intake at rest formen of their age and weight was estimated at 2100Cal; hence, this intake was provided in the form oftwelve portions of solid and liquid food of 160–200Cal each, distributed every 2 h, and identical at lowand high altitude. Water intake was ad libitum.
The quarters were shielded from sunlight, andmaintained at a dim artificial light of �30 lx. AtHA ambient temperature averaged about 1°C less,and the relative humidity was about half, the sealevel values (Table 2). At both locations ambienttemperature was slightly higher in the second halfof the 24-h period, by �3°C at HA and by �1°Cat SL.
Some data were acquired continuously (oxygensaturation, activity, tympanic and thoracic skintemperatures). For other measurements (calf skintemperature, oxygen consumption, end-tidal PCO2
)data were collected every 2 h, from subject no. 1to no. 6, always in the same order. Data collectionon each subject lasted about 15 min; at the time ofthese measurements data of ambient temperature,relative humidity and light intensity were alsorecorded.
2.2. Measurements
Tympanic and thoracic skin temperatures werecontinuously monitored with thermocouples con-nected to a portable data logger (Mini Mitter, OR),from which data were later downloaded on acomputer and averaged over 30 min bins. Calf skintemperature was measured by a tungsten–constan-tan thermocouple permanently placed on the sub-ject’s calf, and values read every 2 h.
Oxygen consumption (V� O2) was measured with a
breath by breath portable unit (K4 Cosmed,Pavona di Albano, Italy). The unit, connected tothe subject by a face mask, monitored minuteventilation by a bidirectional turbine, previouslycalibrated with known volumes, and expired O2
concentration by an O2 chemosensitive cell com-pensated for barometric pressure, temperature andhumidity. The inspired O2 concentration was en-tered as part of the calibration procedure; hence,V� O2
was computed as the product of the expired
of the following day. During the routine the sub-jects, who stayed in groups of two– four per room,were requested to have minimal activity, withoutfalling asleep. They spent almost all their time inbed in a semi-recumbent position, wearing theirusual bed-time clothes, and occupying the free timebetween measurements by watching television orvideos, listening to music, or chatting among them-selves or with members of the investigators’ team.
M. Vargas et al. / Respiration Physiology 126 (2001) 233–243236
ventilation and the difference between inspiredand expired O2 concentration. The accuracy ofthe V� O2
measurement had been previouslychecked by delivering to the unit known flows ofgases of predefined inspired and expired composi-tions. In each subject V� O2
was measured for about4–5 min every 2 h, and the data stored for laterprocessing. Values are given at standard tempera-ture, pressure and dry (STPD) conditions (1 mlO2 STPD=0.0446 mmol O2).
The end-tidal partial pressure of CO2 was mea-sured with a calibrated infrared CO2 analyzer
(Criticare Systems Inc., Waukeska, WI) connectedto nasal lines, over a 4 min period every 2 h.Values are provided at body temperature, pres-sure, and water vapour saturation (BTPS) condi-tions. From the breath-by-breath CO2 signal,breathing rate was also measured.
Systolic and diastolic arterial pressures wereautomatically measured every 30 min by a pro-grammable unit controlling an inflatable brachialcuff, with data stored on a logger (AmbulatoryBlood Pressure recorder, model c RZ250,Rozinn Electronics Inc, Glendale, NY).
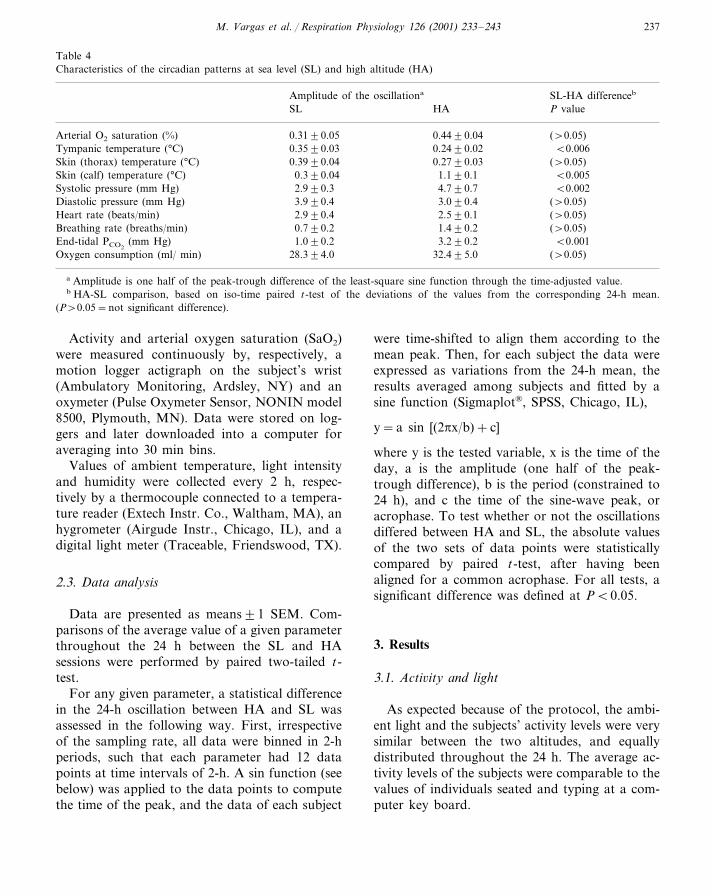
Table 3Hematocrit and hemoglobin in subjects with intermittent hypoxiaa
Sea level, before any After 8 month of intermittent hypoxia:hypoxia
At the end of the high-altitude weekAt the end of the low-altitude weekB CA
Number of 1436 19subjects
48�1b43�0.3 51�1b,cHematocrit, %Hemoglobin, g/dl 16.4�0.2b14.8�0.1 17.7�0.2b
a These data were obtained from a group of the subjects’ coworkers.b Significant difference from group Ac Significant difference from group B
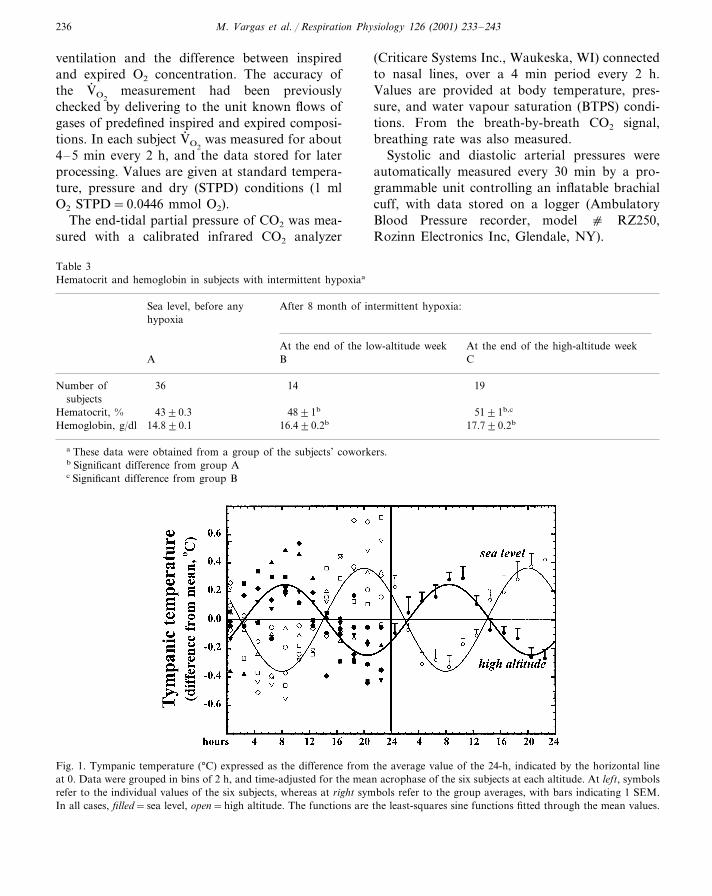
Fig. 1. Tympanic temperature (°C) expressed as the difference from the average value of the 24-h, indicated by the horizontal lineat 0. Data were grouped in bins of 2 h, and time-adjusted for the mean acrophase of the six subjects at each altitude. At left, symbolsrefer to the individual values of the six subjects, whereas at right symbols refer to the group averages, with bars indicating 1 SEM.In all cases, filled=sea level, open=high altitude. The functions are the least-squares sine functions fitted through the mean values.
M. Vargas et al. / Respiration Physiology 126 (2001) 233–243 237
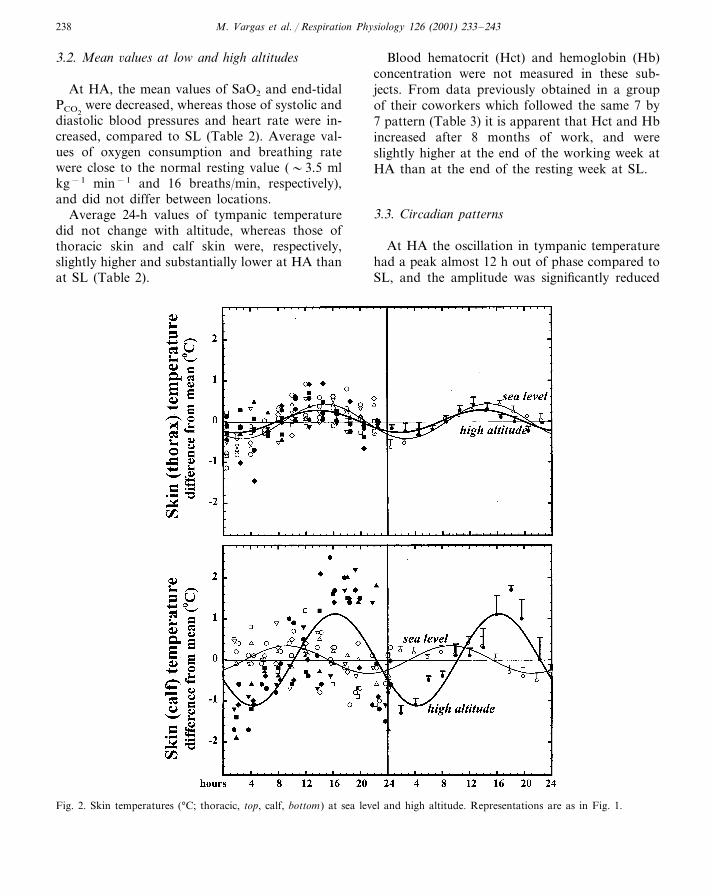
Table 4Characteristics of the circadian patterns at sea level (SL) and high altitude (HA)
Amplitude of the oscillationa SL-HA differenceb
SL HA P value
0.44�0.04Arterial O2 saturation (%) (�0.05)0.31�0.05Tympanic temperature (°C) 0.35�0.03 0.24�0.02 �0.006Skin (thorax) temperature (°C) 0.27�0.030.39�0.04 (�0.05)
1.1�0.10.3�0.04 �0.005Skin (calf) temperature (°C)4.7�0.7Systolic pressure (mm Hg) �0.0022.9�0.33.0�0.43.9�0.4 (�0.05)Diastolic pressure (mm Hg)
2.9�0.4Heart rate (beats/min) 2.5�0.1 (�0.05)1.4�0.20.7�0.2 (�0.05)Breathing rate (breaths/min)
1.0�0.2End-tidal PCO2(mm Hg) 3.2�0.2 �0.001
Oxygen consumption (ml/ min) 28.3�4.0 32.4�5.0 (�0.05)
a Amplitude is one half of the peak-trough difference of the least-square sine function through the time-adjusted value.b HA-SL comparison, based on iso-time paired t-test of the deviations of the values from the corresponding 24-h mean.
(P�0.05=not significant difference).
Activity and arterial oxygen saturation (SaO2)were measured continuously by, respectively, amotion logger actigraph on the subject’s wrist(Ambulatory Monitoring, Ardsley, NY) and anoxymeter (Pulse Oxymeter Sensor, NONIN model8500, Plymouth, MN). Data were stored on log-gers and later downloaded into a computer foraveraging into 30 min bins.
Values of ambient temperature, light intensityand humidity were collected every 2 h, respec-tively by a thermocouple connected to a tempera-ture reader (Extech Instr. Co., Waltham, MA), anhygrometer (Airgude Instr., Chicago, IL), and adigital light meter (Traceable, Friendswood, TX).
2.3. Data analysis
Data are presented as means�1 SEM. Com-parisons of the average value of a given parameterthroughout the 24 h between the SL and HAsessions were performed by paired two-tailed t-test.
For any given parameter, a statistical differencein the 24-h oscillation between HA and SL wasassessed in the following way. First, irrespectiveof the sampling rate, all data were binned in 2-hperiods, such that each parameter had 12 datapoints at time intervals of 2-h. A sin function (seebelow) was applied to the data points to computethe time of the peak, and the data of each subject
were time-shifted to align them according to themean peak. Then, for each subject the data wereexpressed as variations from the 24-h mean, theresults averaged among subjects and fitted by asine function (Sigmaplot®, SPSS, Chicago, IL),
y=a sin [(2�x/b)+c]
where y is the tested variable, x is the time of theday, a is the amplitude (one half of the peak-trough difference), b is the period (constrained to24 h), and c the time of the sine-wave peak, oracrophase. To test whether or not the oscillationsdiffered between HA and SL, the absolute valuesof the two sets of data points were statisticallycompared by paired t-test, after having beenaligned for a common acrophase. For all tests, asignificant difference was defined at P�0.05.
3. Results
3.1. Acti�ity and light
As expected because of the protocol, the ambi-ent light and the subjects’ activity levels were verysimilar between the two altitudes, and equallydistributed throughout the 24 h. The average ac-tivity levels of the subjects were comparable to thevalues of individuals seated and typing at a com-puter key board.
M. Vargas et al. / Respiration Physiology 126 (2001) 233–243238
3.2. Mean �alues at low and high altitudes
At HA, the mean values of SaO2 and end-tidalPCO2
were decreased, whereas those of systolic anddiastolic blood pressures and heart rate were in-creased, compared to SL (Table 2). Average val-ues of oxygen consumption and breathing ratewere close to the normal resting value (�3.5 mlkg−1 min−1 and 16 breaths/min, respectively),and did not differ between locations.
Average 24-h values of tympanic temperaturedid not change with altitude, whereas those ofthoracic skin and calf skin were, respectively,slightly higher and substantially lower at HA thanat SL (Table 2).
Blood hematocrit (Hct) and hemoglobin (Hb)concentration were not measured in these sub-jects. From data previously obtained in a groupof their coworkers which followed the same 7 by7 pattern (Table 3) it is apparent that Hct and Hbincreased after 8 months of work, and wereslightly higher at the end of the working week atHA than at the end of the resting week at SL.
3.3. Circadian patterns
At HA the oscillation in tympanic temperaturehad a peak almost 12 h out of phase compared toSL, and the amplitude was significantly reduced
Fig. 2. Skin temperatures (°C; thoracic, top, calf, bottom) at sea level and high altitude. Representations are as in Fig. 1.
M. Vargas et al. / Respiration Physiology 126 (2001) 233–243 239
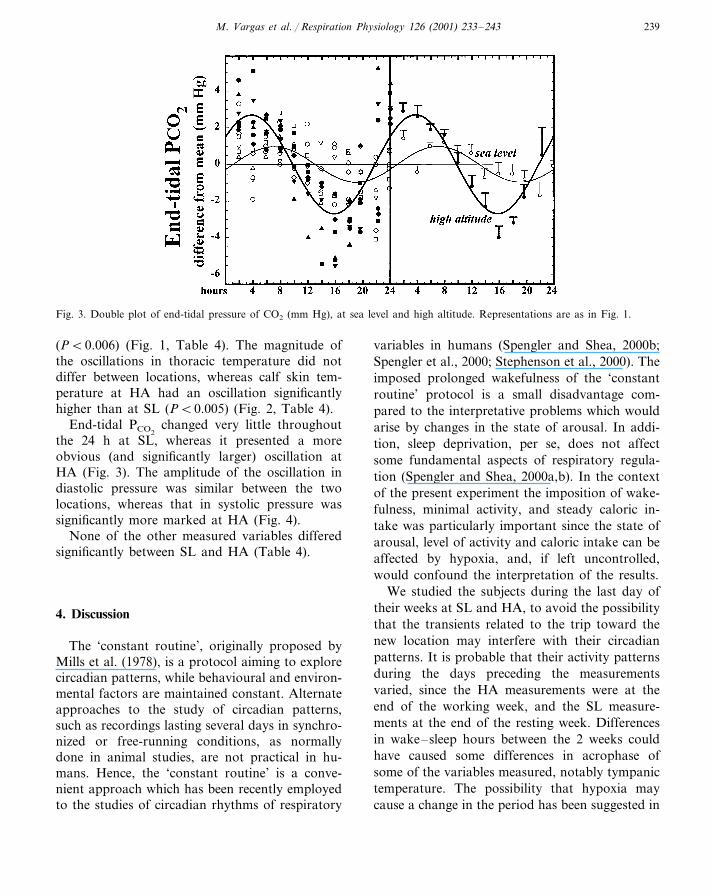
Fig. 3. Double plot of end-tidal pressure of CO2 (mm Hg), at sea level and high altitude. Representations are as in Fig. 1.
(P�0.006) (Fig. 1, Table 4). The magnitude ofthe oscillations in thoracic temperature did notdiffer between locations, whereas calf skin tem-perature at HA had an oscillation significantlyhigher than at SL (P�0.005) (Fig. 2, Table 4).
End-tidal PCO2changed very little throughout
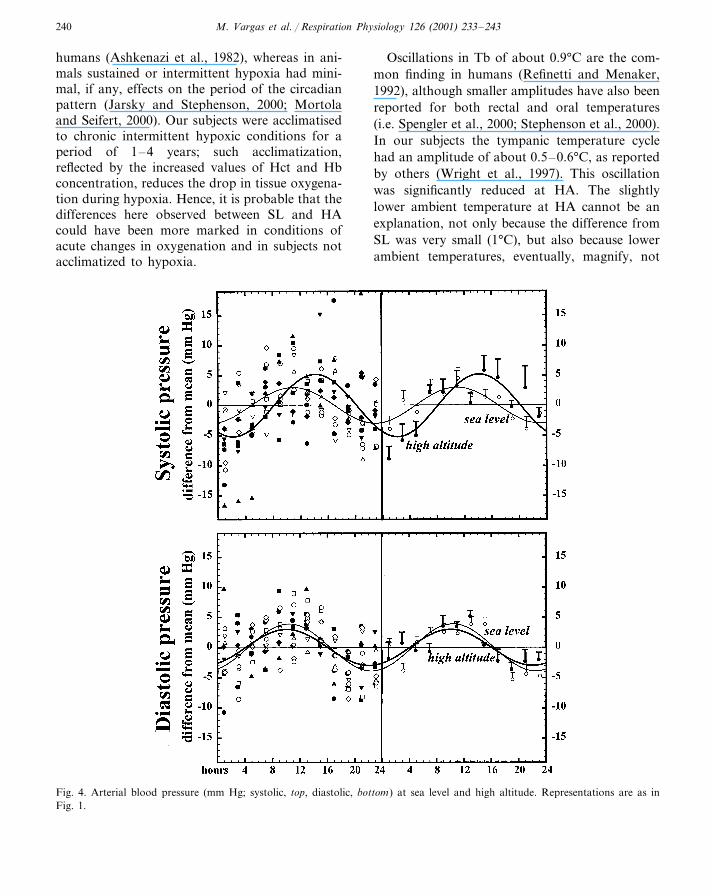
the 24 h at SL, whereas it presented a moreobvious (and significantly larger) oscillation atHA (Fig. 3). The amplitude of the oscillation indiastolic pressure was similar between the twolocations, whereas that in systolic pressure wassignificantly more marked at HA (Fig. 4).
None of the other measured variables differedsignificantly between SL and HA (Table 4).
4. Discussion
The ‘constant routine’, originally proposed byMills et al. (1978), is a protocol aiming to explorecircadian patterns, while behavioural and environ-mental factors are maintained constant. Alternateapproaches to the study of circadian patterns,such as recordings lasting several days in synchro-nized or free-running conditions, as normallydone in animal studies, are not practical in hu-mans. Hence, the ‘constant routine’ is a conve-nient approach which has been recently employedto the studies of circadian rhythms of respiratory
variables in humans (Spengler and Shea, 2000b;Spengler et al., 2000; Stephenson et al., 2000). Theimposed prolonged wakefulness of the ‘constantroutine’ protocol is a small disadvantage com-pared to the interpretative problems which wouldarise by changes in the state of arousal. In addi-tion, sleep deprivation, per se, does not affectsome fundamental aspects of respiratory regula-tion (Spengler and Shea, 2000a,b). In the contextof the present experiment the imposition of wake-fulness, minimal activity, and steady caloric in-take was particularly important since the state ofarousal, level of activity and caloric intake can beaffected by hypoxia, and, if left uncontrolled,would confound the interpretation of the results.
We studied the subjects during the last day oftheir weeks at SL and HA, to avoid the possibilitythat the transients related to the trip toward thenew location may interfere with their circadianpatterns. It is probable that their activity patternsduring the days preceding the measurementsvaried, since the HA measurements were at theend of the working week, and the SL measure-ments at the end of the resting week. Differencesin wake–sleep hours between the 2 weeks couldhave caused some differences in acrophase ofsome of the variables measured, notably tympanictemperature. The possibility that hypoxia maycause a change in the period has been suggested in
M. Vargas et al. / Respiration Physiology 126 (2001) 233–243240
humans (Ashkenazi et al., 1982), whereas in ani-mals sustained or intermittent hypoxia had mini-mal, if any, effects on the period of the circadianpattern (Jarsky and Stephenson, 2000; Mortolaand Seifert, 2000). Our subjects were acclimatisedto chronic intermittent hypoxic conditions for aperiod of 1–4 years; such acclimatization,reflected by the increased values of Hct and Hbconcentration, reduces the drop in tissue oxygena-tion during hypoxia. Hence, it is probable that thedifferences here observed between SL and HAcould have been more marked in conditions ofacute changes in oxygenation and in subjects notacclimatized to hypoxia.
Oscillations in Tb of about 0.9°C are the com-mon finding in humans (Refinetti and Menaker,1992), although smaller amplitudes have also beenreported for both rectal and oral temperatures(i.e. Spengler et al., 2000; Stephenson et al., 2000).In our subjects the tympanic temperature cyclehad an amplitude of about 0.5–0.6°C, as reportedby others (Wright et al., 1997). This oscillationwas significantly reduced at HA. The slightlylower ambient temperature at HA cannot be anexplanation, not only because the difference fromSL was very small (1°C), but also because lowerambient temperatures, eventually, magnify, not
Fig. 4. Arterial blood pressure (mm Hg; systolic, top, diastolic, bottom) at sea level and high altitude. Representations are as inFig. 1.

M. Vargas et al. / Respiration Physiology 126 (2001) 233–243 241
attenuate, the circadian oscillations of Tb(Refinetti and Menaker, 1992).
Hypoxia reduces thermogenesis, not only inanimals, but also in humans (Blatteis andLutherer, 1976; Frappell et al., 1998); sincechanges in thermoregulation are an importantmechanism for the circadian oscillation of Tb(Refinetti and Menaker, 1992), one may expectsome depressant effect of hypoxia on the circa-dian pattern of Tb. This indeed was apparentfrom data in young and adult rats (Fenelon et al.,2000; Mortola and Seifert, 2000; Saiki and Mor-tola, 1995). The present results are entirely consis-tent with these notions, and extend to humans theprevious results on animals. An alternate possibil-ity, namely that a reduction in the circadian pat-tern of Tb could be a non-specific response to thestress induced by the modifications in inspired gascomposition and the continuous hyperventilation,is improbable. In fact, in rats the circadian pat-terns were not affected by continuous exposure tohypercapnia (Mortola and Seifert, 2000), and thesame conclusion was reached from a study onhumans breathing 1.2% CO2 (Samel et al., 1998).
Skin temperatures in central locations, such asthe chest, tend to change in phase with coretemperature, whereas the peripheral skin tempera-tures such as those of hands and feet have analmost opposite pattern, because of the contribu-tion of the extremities in the control of Tb byvarying heat loss (Timbal et al., 1972). At highaltitude, the lower mean value of T skin(calf)likely reflected the peripheral vasoconstriction,which is part of the hypoxic redistribution ofblood toward the central organs (Kuwahira et al.,1993; Mortola et al., 1999). However, the exis-tence of a cycle would indicate that the peripheralvasoconstriction was still under the control of thenormal circadian pattern of heat loss. If thisinterpretation was correct, it would mean thathypoxia had a differential effect on the circadianpatterns of heat production and heat loss, a possi-bility not yet investigated in animal studies.
The mechanisms for the effects of hypoxia onthe circadian patterns of Tb are still unclear.Hypoxia can affect the daily patterns of neuro-transmitters of various brain regions including thesuprachiasmatic nuclei (Poncet et al., 1999), which
is the anatomical location of the biological clock(Van den Pol and Dudek, 1993). However, severalconsiderations suggest that this may not be themost probable site of action. The observation thatseveral circadian patterns are maintained whilethat of Tb is depressed is difficult to conciliatewith the idea of an hypoxic effect on the clockitself. In free-running conditions, i.e. with no dailysynchronization, the biological clock gets out ofphase with the chronological time by a predictableamount. The fact that the Tb cycle of rats infree-running conditions, upon return to normoxiaafter a few days in hypoxia, resumed at the ex-pected chronological time indicated that hypoxiadid not interfere with the clock itself (Fenelon etal., 2000; Mortola and Seifert, 2000). Finally, thenotions that the inhibitory effects of hypoxia aremanifest on all the various forms of heat control,shivering, non-shivering, and behavioural, thatthey are accompanied by a lowering of the ther-moregulatory set point, and that they do notrequire inputs from the peripheral chemorecep-tors, point toward the hypothalamic thermoregu-latory centers as a site of integration for theeffects of hypoxia (Mortola and Gautier, 1995).For all these reasons, the hypothalamic ther-moregulatory centers are also the best candidatefor the anatomical location responsible for theeffect of hypoxia on the circadian pattern of Tb.
In humans during ‘constant routine’ changes inresting metabolic rate and pulmonary ventilation(VE) throughout the 24 h are minimal and ap-proximately proportional to each other (Spengleret al., 2000; Stephenson et al., 2000); hence, thecircadian oscillations in end-tidal PCO2
are verysmall, within 1–2 mm Hg (Mills, 1953; Spengleret al., 2000). In behaving rats V� O2
had largeday-night differences, and so did VE, approxi-mately in proportion (Saiki and Mortola, 1995;Seifert et al., 2000, and unpublished measure-ments). We too found minimal changes in end-tidal PCO2
at SL; differently, at HA, end-tidal PCO2
showed a marked oscillation, with about 6 mmHg peak-trough difference. Also the oscillation insystolic pressure was significantly larger at HAthan at SL. Although not the primary interest ofthe study, these findings are intriguing and notreadily explainable.

M. Vargas et al. / Respiration Physiology 126 (2001) 233–243242
It could be more than a coincidence that thehours of hypoventilation (and of the decrease insystolic pressure) occurred in the first half of theroutine, i.e. almost in phase with the perceivedsleep hours, and the opposite during the perceivedday time. In other words, the patterns of end-tidalPCO2
and systolic pressure at HA, despite themaintained wakefulness, were reminiscent of whatis commonly observed at sea level during thenormal sleep-wakefulness cycle. The hypoventila-tion during sleep is attributed to the depression ofthe behavioural, in favour of the metabolic, orautonomic, factors stimulating V� E (Phillipson andBowes, 1986). Perhaps, the hypoventilation duringsleep hours may not be the effect of sleep per se,but of circadian changes in the magnitude ofmetabolic and behavioural stimuli on breathing,which would occur even during sustained wake-fulness (Mills, 1953). In such a case, hypoxia, bystimulating the autonomic responses, could fa-vour the manifestation of circadian oscillationsnot, or less, apparent during normoxia, such asthose of V� E and systolic pressure. In any case,irrespective of the explanation, the large oscilla-tion in end-tidal PCO2
must indicate that the nor-mal matching between ventilation and metabolicdemands at HA is not as effective as it is at SL.
In conclusion, the circadian patterns of tym-panic and skin temperatures differed between highand low altitudes. The results are qualitatively inagreement with what is observed in animal exper-iments during normobaric or hypobaric hypoxia,and could be interpreted as part of the generalphenomenon of the hypoxic depression of ther-mogenesis. The daily rhythm of Tb may be associ-ated with sleep duration and organization;specifically, at bedtime, the propensity for sleepand its characteristics correlated with the circa-dian phase of Tb, rather than the length of priorwakefulness (Czeisler et al., 1980; Zulley et al.,1981). Sleep is frequently altered in naive subjectsat HA (Weil, 1986; Anholm et al., 1992), and inpatients with cardiorespiratory diseases (Fleethamet al., 1982). Presumably, the modifications in thecircadian patterns of temperature regulation couldbe the mechanistic link between hypoxia and sleepdisturbances.
Acknowledgements
This study would have been impossible withoutthe collaboration of the personnel of the Compa-nia Minera Dona Ines de Collahuasi and of theCentro de Investigacion en Medicina de Altura,Mutual de Seguridad, Iquique, Chile. In particu-lar we own special thanks to G. Cortes, C. Hud-son, and A. Puig for their technical skills, and tothe six subjects for generously participating to thestudy.We are very grateful to the Mini MitterCompany (Sunriver, OR), Guy Jennings (Bio-Lynx Scientific Equipment, Montreal, PQ), SunnyDong (Sleep and Breathing Center, Vancouver,BC), and Stephen Cheung (School of Health andHuman Performance, Dalhousie University, Hali-fax, NS) for their kindness in lending the heartrate and temperature loggers.Financial contribu-tion was provided by funds from the CompaniaMinera Dona Ines de Collahuasi and the Centrode Investigacion en Medicina de Altura, Mutualde Seguridad C. Ch. C., Iquique, Chile, and theMedical Research Council, Canada.
References
Anholm, J.D., Powles, A.C.P., Downey, R., Houston, C.S.,Sutton, J.R., Bonnet, M.H., Cymerman, A., 1992. Opera-tion Everest II: arterial oxygen saturation and sleep atextreme simulated altitude. Am. Rev. Respir. Dis. 145,817–826.
Ashkenazi, I.E., Ribak, J., Avgar, D.M., Klepfish, A., 1982.Altitude and hypoxia as phase shift inducers. Av. Sp.Environ. Med. 53, 342–346.
Blatteis, C.M., Lutherer, L.O., 1976. Effect of altitude exposureon thermoregulatory response of man to cold. J. Appl.Physiol. 41, 848–858.
Czeisler, C.A., Weitzman, E.D., Moore-Ede, M.C., Zimmer-man, J.C., Knauer, R.S., 1980. Human sleep: its durationand organization depend on its circadian phase. Science 210,1264–1267.
Decoursey, P.J., Pius, S., Sandlin, C., Wethey, D., Schull, J.,1998. Relationship of circadian temperature and activityrhythms in two rodent species. Physiol. Behav. 65, 457–463.
Fenelon, K., Seifert, E.L., Mortola, J.P., 2000. Hypoxic depres-sion of circadian oscillations in sino-aortic denervated rats.Respir. Physiol. 122, 61–69.
Fleetham, J., West, P., Mezon, B., Conway, W., Roth, T.,Kryger, M., 1982. Sleep, arousal, and oxygen desaturationin chronic obstructive pulmonary disease. Am. Rev. Respir.Dis. 126, 429–433.

M. Vargas et al. / Respiration Physiology 126 (2001) 233–243 243
Frappell, P.B., Leon-Velarde, F., Aguero, L., Mortola, J.P.,1998. Response to cooling temperature in infants born atan altitude of 4,330 meters. Am. J. Respir. Crit. Care Med.158, 1751–1756.
Gautier, H., 1996. Interactions among metabolic rate, hy-poxia, and control of breathing. J. Appl. Physiol. 81,521–527.
Jarsky, T.M., Stephenson, R., 2000. Effects of hypoxia andhypercapnia on circadian rhythms in the golden hamster(Mesocricetus auratus). J. Appl. Physiol. 89, 2130–2138.
Kuwahira, I., Heisler, N., Piiper, J., Gonzalez, N.C., 1993.Effect of chronic hypoxia on hemodynamics, organ bloodflow and O2 supply in rats. Respir. Physiol. 92, 227–238.
Mills, J.N., 1953. Changes in alveolar carbon dioxide tensionby night and during sleep. J. Physiol, London 122, 66–80.
Mills, J.N., Minors, D.S., Waterhouse, J.M., 1978. Adaptationto abrupt time shifts on the oscillator(s) controlling humancircadian rhythms. J. Physiol. London 285, 455–470.
Mortola, J.P., Gautier, H., 1995. Interaction betweenmetabolism and ventilation: Effects of respiratory gasesand temperature. In: Dempsey, J.A., Pack, A.I. (Eds.),Regulation of Breathing, 2nd edn. Marcel Dekker, NY,pp. 1011–1064 (revised and expanded).
Mortola, J.P., Seifert, E.L., 2000. Hypoxic depression of circa-dian rhythms in adult rats. J. Appl. Physiol. 88, 365–368.
Mortola, J.P., Merazzi, D., Naso, L., 1999. Blood flow to thebrown adipose tissue of conscious young rabbits duringhypoxia in cold and warm conditions. Pflugers Arch. 437,255–260.
Phillipson, E.A., Bowes, G., 1986. Control of breathing duringsleep. In: Cherniack, N.S., Widdicombe, J.G. (Eds.),Handbook of Physiology, Section 3: The Respiratory Sys-tem. Control of Breathing, part 2, vol. II:. AmericanPhysiological Society, Bethesda, MD, pp. 649–689.
Poncet, L., Pequinot, J.-M., Cottet-Emard, J.-M., Dalmaz, Y.,Denoroy, L., 1999. Altered daily rhythms of brain andpituitary indolamines and neuropeptides in long-term hy-poxic rats. Am. J. Physiol. 277, R66–R75.
Refinetti, R., Menaker, M., 1992. The circadian rhythm ofbody temperature. Physiol. Behav. 51, 613–637.
Saiki, C., Mortola, J.P., 1995. Hypoxia abolishes the morn-ing–night differences of metabolism and ventilation in6-day-old rats. Can. J. Physiol. Pharmacol. 73, 159–164.
Samel, A., Vejvoda, M., Wittiber, K., Wenzel, J., 1998. Circa-dian rhythms and activity-rest cycle under different CO2
concentrations. Aviat. Space Environ. Med. 69, 501–505.Seifert, E.L., Knowles, J., Mortola, J.P., 2000. Continuous
circadian measurements of ventilation in behaving adultrats. Respir. Physiol. 120, 179–183.
Spengler, C.M., Shea, S.A., 2000a. Sleep deprivation per sedoes not decrease the hypercapnic ventilatory response inhumans. Am. J. Respir. Crit. Care Med. 161, 1124–1128.
Spengler, C.M., Shea, S.A., 2000b. Endogenous circadianrhythm of pulmonary function in healthy humans. Am. J.Respir. Crit. Care Med. 162, 1038–1046.
Spengler, C.M., Czeisler, C.A., Shea, S.A., 2000. An endoge-nous circadian rhythm of respiratory control in humans. J.Physiol. London 526, 683–694.
Stephenson, R., Mohan, R.M., Duffin, J., Jarsky, T.M., 2000.Circadian rhythms in the chemoreflex control of breathing.Am. J. Physiol. 278, R282–R286.
Timbal, J., Colin, J., Boutelier, C., Guieu, J.D., 1972. Bilanthermique de l’homme en ambiance controlee pendant 24houres. Pflugers Arch. 335, 97–108.
Van den Pol, A.N., Dudek, F.E., 1993. Cellular communica-tion in the circadian clock, the suprachiasmatic nucleus.Neuroscience 56, 793–811.
Weil, J.V., 1986. Ventilatory control at high altitude. In:Cherniack, N.S., Widdicombe, J.G. (Eds.), Handbook ofPhysiology, Section 3: The Respiratory System. Control ofBreathing, part 2, vol. II:. American Physiological Society,Bethesda, MD, pp. 703–727.
Wright, K.P., Badia, P., Myers, B.L., Plenzler, S.C., Hakel,M., 1997. Caffein and light effects on nighttime melatoninand temperature levels in sleep-deprived humans. BrainRes. 747, 78–84.
Zulley, J., Wever, R., Aschoff, J., 1981. The dependence ofonset and duration of sleep on the circadian rhythm ofrectal temperature. Pflugers Arch. 391, 314–318.
.
Related Documents











