Circadian Clocks During Embryonic and Fetal Development Maria Seron-Ferre,* Gullermo J. Valenzuela, and Claudia Torres-Farfan Circadian rhythmicity is a fundamental characteristic of organisms, which helps ensure that vital functions occur in an appropriate and pre- cise temporal sequence and in accordance with cyclic environmental changes. Living beings are endowed with a system of biological clocks that measure time on a 24-hr basis, termed the circadian timing sys- tem. In mammals, the system is organized as a master clock located in the suprachiasmatic nucleus (SCN) of the hypothalamus, commanding peripheral clocks located in almost every tissue of the body. At the cell level, interlocking transcription/translation feedback loops of the genes Bmal-1, Clock, Per1–2, and Cry1–2, named clock genes, and their pro- tein products results in circadian oscillation of clock genes and of genes involved in almost every cellular function. During gestation, the concep- tus follows a complex and dynamic program by which it is simultane- ously fit to develop and live in a circadian environment provided by its mother and to prepare for the very different environment that it will ex- perience after birth. It has been known for a number of years that the mother tells the fetus the time of day and season of the year, and that the fetus uses this information to set the phase of fetal and neonatal cir- cadian rhythms. There is evidence that the maternal rhythm of melato- nin is one of the time signals to the fetus. In the last few years, the study of the development of the circadian system has turned to the investigation of the oscillatory expression of clock genes and their possi- ble role in development, and to answering questions on the organization of the fetal circadian system. Emerging evidence shows that clock genes are expressed in the oocyte and during early and late development in embryo/fetal organs in the rat and in a fetal primate. The data available raise the intriguing possibility that the fetal SCN and fetal tissues may be peripheral clocks commanded by separate maternal signals. The rapid methodological and conceptual advances on chronobiology may help to unravel how the developing embryo and fetus faces time in this plastic period of life. Birth Defects Research (Part C) 81:204–214, 2007. V C 2007 Wiley-Liss, Inc. INTRODUCTION The reliable (and therefore pre- dictable) environmental alterna- tions of day and night and the seasons originated from the fact that our planet turns about itself in 24 hr and moves around the sun in a year, led to evolution of biological clocks that measure time on about 24-hr basis (circa- dian clocks) in practically all living beings. These clocks also time reproduction in seasonal animals, ensuring the best chances for sur- vival to the offspring (Moore-Ede et al., 1982; Edery, 2000; Lincoln et al., 2003). Circadian clocks pro- vide a 24-hr framework for inter- nal temporal organization, sepa- rating incompatible physiological functions both at the cellular and also at the organism level. In mammals, the circadian timing system is organized as a hierarchy of oscillators located in most tis- sues of the body (peripheral oscil- lators), synchronized by a central rhythm generator located in the suprachiasmatic nucleus (SCN) of the hypothalamus (Fig. 1). The overt circadian rhythms of tem- perature, cortisol, activity, sleep, and heart rate to which we are familiarized can be viewed as the hands of this clock system. The SCN and peripheral clocks are run by a common transcriptional cir- cuitry of genes (clock genes) that generates rhythmic patterns of gene expression in themselves and on other genes (clock-con- trolled genes) that are involved in protein expression, metabolism, and/or function depending on the REVIEW Birth Defects Research (Part C) 81:204–214 (2007) M. Sero ´n-Ferre ´ is from the Programa de Fisiopatologı ´a, Instituto de Ciencias Biome ´dicas (ICBM) Facultad de Medicina, Universidad de Chile, Santiago, Chile, and from the Universidad de Tarapaca and Centro de Investigaciones del Hombre en el Desierto (CIHDE), Arica, Chile. G.J. Valenzuela is from the Department of Women’s Health, Arrowhead Regional Medical Center, Colton, CA. C. Torres-Farfan is from the Programa de Fisiopatologı ´a, Instituto de Ciencias Biome ´dicas (ICBM) Facultad de Medicina, Universidad de Chile, Santiago, Chile. Grant sponsor: Fondo Nacional de Desarrollo Cientı ´fico y Tecnolo ´gico, Fondecyt 106766, Santiago, Chile; Grant sponsor: San Bernar- dino Medical Foundation, Department of Women’s Health, Arrowhead Regional Medical Center, CA. *Correspondence to: Marı ´a Sero ´n-Ferre ´, Ph.D., Salvador 486, Providencia, Santiago, Chile, Programa de Fisiopatologı ´a, ICBM, Facul- tad de Medicina, Universidad de Chile, Casilla 16038, Santiago 9, Santiago, Chile. E-mail: [email protected] Published online in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/bdrc.20101 V C 2007 Wiley-Liss, Inc.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Circadian Clocks During Embryonic andFetal Development
Maria Seron-Ferre,* Gullermo J. Valenzuela, and Claudia Torres-Farfan
Circadian rhythmicity is a fundamental characteristic of organisms,which helps ensure that vital functions occur in an appropriate and pre-cise temporal sequence and in accordance with cyclic environmentalchanges. Living beings are endowed with a system of biological clocksthat measure time on a 24-hr basis, termed the circadian timing sys-tem. In mammals, the system is organized as a master clock located inthe suprachiasmatic nucleus (SCN) of the hypothalamus, commandingperipheral clocks located in almost every tissue of the body. At the celllevel, interlocking transcription/translation feedback loops of the genesBmal-1, Clock, Per1–2, and Cry1–2, named clock genes, and their pro-tein products results in circadian oscillation of clock genes and of genesinvolved in almost every cellular function. During gestation, the concep-tus follows a complex and dynamic program by which it is simultane-ously fit to develop and live in a circadian environment provided by itsmother and to prepare for the very different environment that it will ex-perience after birth. It has been known for a number of years that themother tells the fetus the time of day and season of the year, and thatthe fetus uses this information to set the phase of fetal and neonatal cir-cadian rhythms. There is evidence that the maternal rhythm of melato-nin is one of the time signals to the fetus. In the last few years, thestudy of the development of the circadian system has turned to theinvestigation of the oscillatory expression of clock genes and their possi-ble role in development, and to answering questions on the organizationof the fetal circadian system. Emerging evidence shows that clock genesare expressed in the oocyte and during early and late development inembryo/fetal organs in the rat and in a fetal primate. The data availableraise the intriguing possibility that the fetal SCN and fetal tissues maybe peripheral clocks commanded by separate maternal signals. Therapid methodological and conceptual advances on chronobiology mayhelp to unravel how the developing embryo and fetus faces time in thisplastic period of life. Birth Defects Research (Part C) 81:204–214,2007. VC 2007 Wiley-Liss, Inc.
INTRODUCTIONThe reliable (and therefore pre-dictable) environmental alterna-tions of day and night and the
seasons originated from the factthat our planet turns about itselfin 24 hr and moves around thesun in a year, led to evolution of
biological clocks that measuretime on about 24-hr basis (circa-dian clocks) in practically all livingbeings. These clocks also timereproduction in seasonal animals,ensuring the best chances for sur-vival to the offspring (Moore-Edeet al., 1982; Edery, 2000; Lincolnet al., 2003). Circadian clocks pro-vide a 24-hr framework for inter-nal temporal organization, sepa-rating incompatible physiologicalfunctions both at the cellular andalso at the organism level. Inmammals, the circadian timingsystem is organized as a hierarchyof oscillators located in most tis-sues of the body (peripheral oscil-lators), synchronized by a centralrhythm generator located in thesuprachiasmatic nucleus (SCN) ofthe hypothalamus (Fig. 1). Theovert circadian rhythms of tem-perature, cortisol, activity, sleep,and heart rate to which we arefamiliarized can be viewed as thehands of this clock system. TheSCN and peripheral clocks are runby a common transcriptional cir-cuitry of genes (clock genes) thatgenerates rhythmic patterns ofgene expression in themselvesand on other genes (clock-con-trolled genes) that are involved inprotein expression, metabolism,and/or function depending on the
REVIE
WBirth Defects Research (Part C) 81:204–214 (2007)
M. Seron-Ferre is from the Programa de Fisiopatologıa, Instituto de Ciencias Biomedicas (ICBM) Facultad de Medicina, Universidadde Chile, Santiago, Chile, and from the Universidad de Tarapaca and Centro de Investigaciones del Hombre en el Desierto (CIHDE),Arica, Chile.G.J. Valenzuela is from the Department of Women’s Health, Arrowhead Regional Medical Center, Colton, CA.C. Torres-Farfan is from the Programa de Fisiopatologıa, Instituto de Ciencias Biomedicas (ICBM) Facultad de Medicina, Universidadde Chile, Santiago, Chile.
Grant sponsor: Fondo Nacional de Desarrollo Cientıfico y Tecnologico, Fondecyt 106766, Santiago, Chile; Grant sponsor: San Bernar-dino Medical Foundation, Department of Women’s Health, Arrowhead Regional Medical Center, CA.
*Correspondence to: Marıa Seron-Ferre, Ph.D., Salvador 486, Providencia, Santiago, Chile, Programa de Fisiopatologıa, ICBM, Facul-tad de Medicina, Universidad de Chile, Casilla 16038, Santiago 9, Santiago, Chile. E-mail: [email protected]
Published online in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/bdrc.20101
VC 2007 Wiley-Liss, Inc.
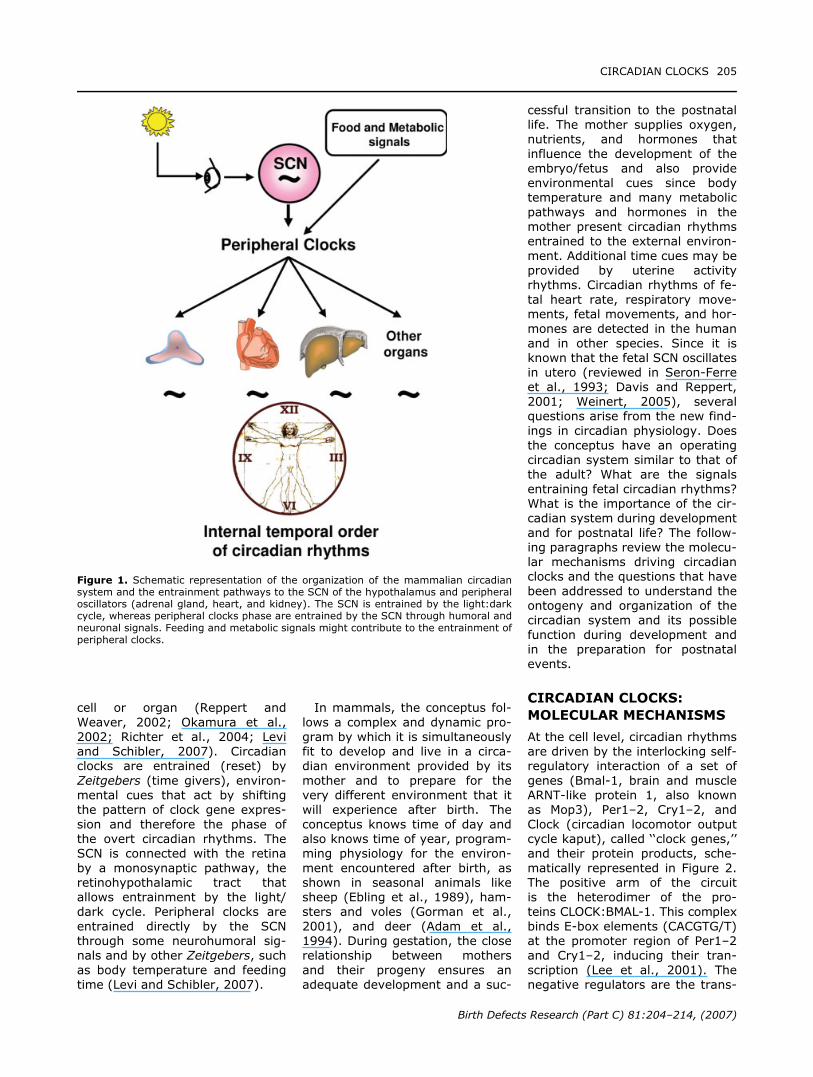
cell or organ (Reppert andWeaver, 2002; Okamura et al.,2002; Richter et al., 2004; Leviand Schibler, 2007). Circadianclocks are entrained (reset) byZeitgebers (time givers), environ-mental cues that act by shiftingthe pattern of clock gene expres-sion and therefore the phase ofthe overt circadian rhythms. TheSCN is connected with the retinaby a monosynaptic pathway, theretinohypothalamic tract thatallows entrainment by the light/dark cycle. Peripheral clocks areentrained directly by the SCNthrough some neurohumoral sig-nals and by other Zeitgebers, suchas body temperature and feedingtime (Levi and Schibler, 2007).
In mammals, the conceptus fol-lows a complex and dynamic pro-gram by which it is simultaneouslyfit to develop and live in a circa-dian environment provided by itsmother and to prepare for thevery different environment that itwill experience after birth. Theconceptus knows time of day andalso knows time of year, program-ming physiology for the environ-ment encountered after birth, asshown in seasonal animals likesheep (Ebling et al., 1989), ham-sters and voles (Gorman et al.,2001), and deer (Adam et al.,1994). During gestation, the closerelationship between mothersand their progeny ensures anadequate development and a suc-
cessful transition to the postnatallife. The mother supplies oxygen,nutrients, and hormones thatinfluence the development of theembryo/fetus and also provideenvironmental cues since bodytemperature and many metabolicpathways and hormones in themother present circadian rhythmsentrained to the external environ-ment. Additional time cues may beprovided by uterine activityrhythms. Circadian rhythms of fe-tal heart rate, respiratory move-ments, fetal movements, and hor-mones are detected in the humanand in other species. Since it isknown that the fetal SCN oscillatesin utero (reviewed in Seron-Ferreet al., 1993; Davis and Reppert,2001; Weinert, 2005), severalquestions arise from the new find-ings in circadian physiology. Doesthe conceptus have an operatingcircadian system similar to that ofthe adult? What are the signalsentraining fetal circadian rhythms?What is the importance of the cir-cadian system during developmentand for postnatal life? The follow-ing paragraphs review the molecu-lar mechanisms driving circadianclocks and the questions that havebeen addressed to understand theontogeny and organization of thecircadian system and its possiblefunction during development andin the preparation for postnatalevents.
CIRCADIAN CLOCKS:
MOLECULAR MECHANISMS
At the cell level, circadian rhythmsare driven by the interlocking self-regulatory interaction of a set ofgenes (Bmal-1, brain and muscleARNT-like protein 1, also knownas Mop3), Per1–2, Cry1–2, andClock (circadian locomotor outputcycle kaput), called ‘‘clock genes,’’and their protein products, sche-matically represented in Figure 2.The positive arm of the circuitis the heterodimer of the pro-teins CLOCK:BMAL-1. This complexbinds E-box elements (CACGTG/T)at the promoter region of Per1–2and Cry1–2, inducing their tran-scription (Lee et al., 2001). Thenegative regulators are the trans-
Figure 1. Schematic representation of the organization of the mammalian circadiansystem and the entrainment pathways to the SCN of the hypothalamus and peripheraloscillators (adrenal gland, heart, and kidney). The SCN is entrained by the light:darkcycle, whereas peripheral clocks phase are entrained by the SCN through humoral andneuronal signals. Feeding and metabolic signals might contribute to the entrainment ofperipheral clocks.
CIRCADIAN CLOCKS 205
Birth Defects Research (Part C) 81:204–214, (2007)

lated CRY and PER proteins, thatcomplex with casein kinase e andtranslocate into the nucleus, inter-acting with the CLOCK:BMAL1complex and thus blocking theirown transcription. Phosphorylationof the PER and CRY proteins bycasein kinase e controls their pro-teasomal degradation, delayingthe formation of CRY:PER com-plexes and determining the lengthof the cycle (Lowrey et al., 2000;Vanselow et al., 2006). TheCLOCK:BMAL1 heterodimer alsoinduces the transcription of Rev-erba and Rora genes, which inter-act with Rev-erb/Ror elements(RREs) in the promoter of Bmal1,repressing and driving its tran-scription, respectively (Preitneret al., 2002). Additional regulatorsare a further negative loop gener-ated by the transcription of thedec1 (sharp2/stra13) and dec2(sharp1) genes, which are alsodriven by CLOCK:BMAL1 via E-boxes in their promoters (Gre-chez-Cassiau et al., 2004). DEC1and DEC2 proteins may block cir-cadian gene expression, in part bythe formation of a nonfunctionalheterodimer with BMAL1 (andhence inhibiting the expression ofall genes dependent on an E-box),as well as play a role in lightinduction of genes in the SCN(Honma et al., 2002). The circa-dian oscillation of clock geneexpression controls the expressionof genes involved in multiple cellu-lar functions in the 24-hr period(clock-controlled genes, CCGs) byat least two mechanisms, directinteraction with E-boxes in thepromoters of these genes, andthrough the regulation of otherCCGs that are in turn transcriptionfactors, like DBP (reviewed inReppert and Weaver, 2002; Oka-mura et al., 2002; Richter et al.,2004; Levi and Schibler, 2007).
An important breakthrough wasthe finding of circadian expressionof clock genes in practically everytissue studied (liver, pancreas,kidney, and heart) and also in celllines (Oishi et al., 1998a, 1998b).These tissues are referred to asperipheral oscillators or peripheralclocks. Importantly, recent experi-ments show that under culture
conditions, the clock gene oscilla-tory process lasts for weeks inthese tissues, underlying its en-dogenous nature (Yoo et al.,2004). Microarray analysis hasshown circadian oscillation in animportant part of the transcrip-tome involving almost every cellu-lar biochemical pathway (Pandaet al., 2002). The link betweenbiochemical processes and clockgenes is not completely under-stood; however, transcriptionalcoactivators like PGC-1a havebeen proposed to be involved (Liuet al., 2007). A consistent obser-vation is that in vivo, clock genesin the peripheral oscillators have aphase delay of 4- to 8-hr relativeto their oscillation in the SCN (Bal-salobre et al., 2000; Damiolaet al., 2000; Stokkan et al.,2001). In rats and mice, destruc-tion of the SCN eliminates circa-dian rhythms of clock geneexpression in peripheral organs(Sakamoto et al., 1998). However,the experiments cannot distin-guish whether SCN lesions abolishcircadian rhythms within an indi-vidual or results in asynchronousrhythms between individuals. SCNtransplant can restore the rhythmof locomotor activity but not endo-crine rhythms in hamster (Meyer-Bernstein et al., 1999), suggestingthat blood-borne and/or othernonneural (e.g., behavioral) rhy-
thms may be responsible for thecommunication of the SCN withsome but not all peripheral oscilla-tors. This is further supported inparabiosis experiments linkingSCN-lesioned mice to intact mice(Guo et al., 2005). In theseexperiments, parabiosis inducedrecovery of rhythms of mPer1,mPer2, and mBmal1 gene expres-sion in liver and kidney, but not inspleen, heart, or skeletal muscle,of the SCN-lesioned mice. Feedingmay provide a time cue for someperipheral clocks, as the phasedisplayed by clock genes in theliver, pancreas, kidney, and heartis modified in rodents subjected tofeeding restricted to a few hoursevery day (Balsalobre et al., 2000;Stokkan et al., 2001; Le Minhet al., 2001; Balsalobre, 2002).These animals increase both loco-motor activity and core body tem-perature in anticipation of thetimed daily meal. Therefore, inaddition to endocrine and meta-bolic signals, biophysical signalslike locomotor activity and bodytemperature may provide Zeitgeb-ers for peripheral clocks (reviewedby Stratmann and Schibler, 2006).In summary, as depicted in Figure1, the circadian timing system inmammals is organized as a hierar-chy of peripheral oscillators syn-chronized by the SCN of the hypo-thalamus (Guo et al., 2006). Theend result is the separation of vitalfunctions in an appropriate andprecise temporal sequence withinthe 24-hr period, hence ensuringpredictive adaptation to cyclicenvironmental changes (Pitten-drigh, 1981; Moore-Ede et al.,1982).
THE CIRCADIAN SYSTEM
AND CIRCADIAN CLOCKS
THROUGHOUT
DEVELOPMENT
Earlier studies on development ofthe circadian system were cen-tered on development of the mas-ter clock (SCN) and on investigat-ing the hands of the clock; i.e.,the presence of overt circadianrhythms either in the late gesta-tion fetus or in the newborns. Cir-
Figure 2. Schematic representation ofthe core components of a circadian clock.The positive arm of the clock, the hetero-dimer CLOCK:BMAL1, initiates transcrip-tion of the genes Per 1–3 and Cry 1–2.Upon translation, the complex of PER:CRY is phosphorylated by casein kinase e(CK e) and transported to the nucleus in-hibiting transcription. A second loop isformed by the transcription factorsREVERBa and ROR. [Redrawn from Mer-row et al. (2006).]
206 SERON-FERRE ET AL.
Birth Defects Research (Part C) 81:204–214, (2007)

cadian rhythms of several phy-siological functions are presentin fetuses of precocial specieslike human, rhesus monkey, andsheep: 1) in the human, fetalheart rate, fetal breathing, fetalmovements, and plasma cortisol;2) in rhesus fetuses circulatingplasma dehydroepiandrosteronesulfate (DHAS); and 3) in fetalsheep fetal breathing, plasma pro-lactin. and arginine vasopressin(AVP) in cerebrospinal fluid(reviewed in Seron-Ferre et al.,1993, 2001). In these species, fe-tal SCN neurogenesis and SCNinnervation by the retinohypotha-lamic tract (RHT) is completed bymidgestation (reviewed in Seron-Ferre et al., 2001) and the fetalSCN shows circadian rhythms ofmetabolic activity (using the14C-labelled 2-deoxy-D-glucoseuptake) (Reppert and Schwartz,1984) and of c-FOS (Constandilet al., 1995; Breen et al., 1996).In contrast, in altricial species(pups are very immature at birth),like rats and hamsters, overt cir-cadian rhythms of temperature,behavioral rhythms like locomotoractivity, drinking, and the plasmacorticosterone rhythm developpostnatally. In these species, SCNneurogenesis is completed close tobirth and innervation by RTH iscompleted postnatally (reviewedby Davis and Reppert, 2001);notwithstanding, the fetal SCNpresents day/night rhythms inmetabolic activity (Reppert andSchwartz, 1983; Davis and Gorski,1985), an abundance of messen-ger RNA (mRNA) for vasopressin(Reppert and Uhl, 1987), andof spontaneous neural activity(recorded in slice preparations;Shibata and Moore, 1987). Alto-gether, these evidences demon-strate oscillatory function of thefetal SCN in altricial and precocialspecies and the presence of overtcircadian rhythms during fetal lifein the latter. A characteristic com-mon to precocial and altricial spe-cies is that fetal SCN rhythmsappear synchronized to clock timein individuals from the same litterand between individuals fromdifferent mothers. Rhythms aresynchronized between twins in
human and sheep pregnancy (Par-raguez et al., 1996; Maeda et al.,2006). In addition, postnatalrhythms are synchronized be-tween newborns from the samelitter (mice, rats, and hamsters). Aseries of experiments in rats andhamsters demonstrate that fetalSCN rhythms persist but within-lit-ter synchrony is lost after mater-nal SCN ablation, indicating thatthe SCN rhythm is generatedendogenously in the fetus but isentrained by circadian signalscontrolled by the maternal SCN.Similarly, lesions of the maternalSCN result in desynchronizationof postnatal behavioral rhythmswithin a litter. To summarize,these series of experiments dem-onstrate that the fetal SCN hasendogenous oscillatory properties,and is entrained by a signal con-trolled by the maternal SCN. Asecond finding is that the phase ofpostnatal behavioral rhythms inrats and hamsters is set by signalsexperienced during gestation(reviewed by Davis and Reppert,2001). The underlying conceptwas of a fetal circadian systemorganized with the fetal SCN asmaster clock entrained by a non-photic signal derived from themother. The new tools provided bythe understanding of the molecu-lar events underlying circadianrhythms and the recent awarenessthat circadian clocks are present inmost peripheral tissues may chal-lenge this view.
Oscillation of circadian clocksaccompanies the growing oocyte,and thereafter. The clock genes,Per-2 and Bmal-1, and their pro-teins oscillate in follicles and cor-pus luteum in the rat ovary andmay play a role in steroidogenesis(Karman and Tischkau, 2006;Fahrenkrug et al., 2006). A fullcomplement of clock genes oscil-late in the rat oviduct (Johnsonet al., 2002; Kennaway et al.,2003), which carries the ovulatedoocyte through fertilization andearly embryonic development, andin the uterus, where it will implant(Johnson et al., 2002; Horardet al., 2004). The functional signifi-cance of these genes in early devel-opment and uterine preparation for
implantation is unknown (reviewedby Boden and Kennaway, 2006).
On the other hand, the unfertil-ized mouse oocyte, as shown inFigure 3, expresses the mRNAs ofthe six canonical clock genes (Per1 and 2, Cry 1 and 2, Clock, andBmal1). After fertilization, expres-sion of these mRNAs decreasesbetween two-cell and 16-cellstages to be reinitiated at theblastocyst stage (Ko et al., 2000;Johnson et al., 2002). What fol-lows after implantation is just be-ginning to be explored. Usingimaging techniques, Saxena et al.(2007) studied the expression ofthe clock gene Per-1 throughoutgestation in the uterus of pregnantrats (Fig. 3). Wild-type female ratswere mated with a heterozygoustransgenic male carrying the Per-1promoter coupled to firefly lucifer-ase; therefore, some of thefetuses expressed the Per-1::lucconstruct. Although the study wasaimed at establishing the feasibil-ity of the technique, some amaz-ing observations were produced.Using 120-sec exposure, Per-1::luc expression was detectedat dusk at 10 days of gestationand a dawn/dusk difference wasobserved at 12 days. A sharp risein fluorescence was detected atthis age, obligating reduced expo-sure time to 1 sec for mea-surements at subsequent ages.Overall, the authors found anexponential increase in Per-1::lucexpression from day 10 to day 21,the end of gestation, and in post-natal day 1. It is not known whichintrauterine tissues (placenta,embryo, or fetal organs) contrib-uted to the intrauterine fluores-cence observed. However, afterbirth, there was strong internalorgan fluorescence attributedmainly to the liver, a very largeorgan in the newborn (and also inthe fetus), besides weak skin fluo-rescence (Saxena et al., 2007). Itis of interest to note that fluores-cence was higher at dusk than atdawn at late gestation, whereasthe pattern is reversed in the new-born. This promising technique inaddition to more conventionaltechniques offers the possibility toidentify which fetal organ(s) is the
CIRCADIAN CLOCKS 207
Birth Defects Research (Part C) 81:204–214, (2007)

source of fluorescence at the dif-ferent gestational ages andwhether this fluorescence is ac-companied by oscillatory expres-sion of other clock genes.
The ontogeny of oscillatorymRNA expression of clock genes inthe fetal SCN and in a few fetalperipheral tissues has beenrecently studied in rat, mice, ham-ster, rabbit, and the capuchinmonkey. Studies in the rat andhamster demonstrate that thecore elements of the circadian sys-tem, the clock genes Per1, Per2,Cry1, Bmal1, and Clock, areexpressed in the rat and hamsterfetal SCN (Sladek et al., 2004; Liand Davis, 2005). However, there
was no clear circadian rhythm ofany of the clock gene mRNAs orPER1, PER2, and CRY1 proteins inthe fetal SCN during fetal life,whereas a clear oscillation ofmRNA and protein was detected atthree to 10 days of postnatal agein the rat and of Bmal1, Cry1, andPer1 also in the hamster SCN. Atthis postnatal age, the pattern ofexpression of clock genes was akinto that in the adult SCN. Similarobservations were obtained forPer1 and Per 2 in the rat and mice(Shimomura et al., 2001; Ohtaet al., 2002).The lack of oscillatoryexpression in the fetal rodent SCNis surprising, given the presenceof cyclic metabolic activity already
discussed. The source of this dis-crepancy remains to be estab-lished. The SCN of another altricialspecies, the rabbit, show oscilla-tory expression of clock genes atday 7 postnatal, the only timestudied (Caldelas et al., 2007).This is of interest since rabbit pupsshow a rhythm of body tempera-ture by day 2 of postnatal age,and show anticipatory activity tothe daily mother visit for feedingsoon after birth (reviewed in Gon-zalez-Mariscal, 2007). In contrastto the lack of oscillation of clockgenes in rats and hamster fetalSCN, as shown in Figure 4A, Bmal-1 and Per-2 expression in the cap-uchin fetal SCN is oscillatory at
Figure 3. A: Expression of clock genes at different embryonic stages. [Data from Johnson et al. (2002) and Ko et al. (2000)].B: Bioluminescence from heterozygous Per1::luc embryos imaged in utero throughout gestation and from two heterozygous new-borns and two wild-type newborns. [Modified from Saxena et al. (2007).]
208 SERON-FERRE ET AL.
Birth Defects Research (Part C) 81:204–214, (2007)

90% gestation (Torres-Farfanet al., 2006a). The peaks of Bmal-1 and Per-2 expression were inantiphase, Bmal-1 peaking at thebeginning of the night and Per-2peaking about 10 hr later, at theend of the night. This 10-hr inter-val between the increases ofBmal-1 and Per-2 in the fetal cap-uchin SCN is longer than thatreported in the SCN of adult ani-mals (Oishi et al., 1998a; Zylkaet al., 1998; Lincoln et al., 2002;
Johnston et al., 2005; Watanabeet al., 2006), and it is reminiscentof the antiphase pattern describedin most peripheral clocks men-tioned in the preceding section. Asecond fetal tissue studied wasthe fetal adrenal. As in the SCN,Bmal-1 and Per-2 oscillation wasdetected. A surprising findingwas that the phase of oscillationwas similar to that detected in thefetal SCN, in contrast with thephase delay between SCN and pe-
ripheral clocks in the adult (Oishiet al., 1998a; Zylka et al., 1998;Lincoln et al., 2002; Johnstonet al., 2005; Watanabe et al.,2006). In both fetal SCN and adre-nal glands, the clock gene oscilla-tion was accompanied by a circa-dian output. We found a rhythm ofDHAS, a steroid secreted by thefetal adrenal and rhythms ofexpression of the MT1 melatoninreceptor in the fetal SCN and ad-renal gland. In addition, another
Figure 4. A: Oscillatory expression of Bmal-1 and Per-2 measured by RT-PCR in the fetal capuchin monkey SCN and adrenal glandat the indicated hours. Fetal adrenal gland (right upper panels): mean 6 SE of Bmal-1 and Per-2 expression measured by RT-PCRat 0800 hr (n 5 5), 1400 hr (n 5 5), and 2000 hr (n 5 3). *Different from 0800 hr and 1400 hr, p \ 0.05, ANOVA and Tukey’stest; yDifferent from 1400 hr and 2000 hr, p \ 0.001, ANOVA, and Tukey’s test. In each graph the value at 0800 hr is repeated inthe next 24 hr. The dark bars indicate lights-off hours. B: Clock gene expression (Bmal-1, Per-2, Cry-2, and Clock) in capuchinmonkey pituitary, thyroid, brown fat, and pineal. Fetuses were euthanized at 0800, 1400, and 2000 hr. [Redrawn from Torres-Farfan et al. (2006a).]
CIRCADIAN CLOCKS 209
Birth Defects Research (Part C) 81:204–214, (2007)

marker of fetal adrenal function,expression of the steroidogenicenzyme 3b hydroxysteroid dehy-drogenase showed clock time–related changes (Torres-Farfanet al., 2006a). Of note, the pro-moter of the MT1 receptor has E-boxes and retinoid orphan recep-tor elements (ROREs), suggestingthat it could be a clock-controlledgene (Ueda et al., 2005; Johnstonet al., 2007). Thus, in the capu-chin, the fetal SCN and the fetaladrenal show circadian oscillationof two clock genes and evidence ofoscillatory function downstream ofthese genes. Recently, severalauthors reported the oscillatoryexpression of clock genes in theadrenal gland of adult rodents(Ishida et al., 2005; Watanabeet al., 2006; Torres-Farfan et al.,2006b) and primate (Lemos et al.,2006), providing strong evidencefor the presence of a peripheralclock in this tissue. This possibilityis highly relevant, in light of evi-dence supporting a role for plasmaglucocorticoids in the entrainmentof other oscillators (Balsalobreet al., 2000), as well as in controlof metabolic rhythms in otherorgans such as liver (Oishi et al.,2005) and adipocytes (Alonso-Vale et al., 2005). Expression ofclock genes in the adrenal glandmay participate in the generationof the corticosterone rhythm, asmutant mice defective in adrenal
clock genes (double Per2/Cry1knockout), show loss of hypothal-amus-pituitary-adrenal axis rhyth-micity. In these mice, transplantof adrenals from wild-type to mu-tant mice rescues the corticoster-one rhythm in light:dark condi-tions (Oster et al., 2006). Whetherthe fetal adrenal gland is a power-ful circadian clock remains to beinvestigated.
Clock genes were also expressedin the other capuchin fetal tissuesinvestigated (pituitary, thyroid,and brown adipose tissue) asshown in Figure 4B, although os-cillatory expression was not meas-ured. These are tissues that exertseveral functions in fetal life, ei-ther producing hormones (pitui-tary, thyroid) (Grumbach andKaplan, 1998; Polk and Fisher,1998) or accruing to serve asthermogenic substrate in the new-born (adipose tissue) (Cannon andNedergaard, 2004), whereas thefetal pineal starts secreting mela-tonin after birth (Kennaway et al.,1992). The fetal pineal did notexpress Per-2 but expressedBmal-1 and Clock and had a sig-nificantly higher expression ofCry-2 than the other tissuestested. Whether the absence ofPer-2 expression coupled to highCry-2 expression relates to thelack of pineal melatonin synthesisduring fetal life needs to beexplored. The only other fetal tis-
sue studied to date is the rat fetalliver. Ontogeny of oscillation ofthe clock genes, Per1, Per2, Rev-Erba, Cry1, Bmal1, and Clock, inthe rat fetal liver, was studied bySladek et al. (2007) finding prena-tal (20 days of gestational age)oscillation of Rev-Erba but not ofthe other genes. A more adult pat-tern of oscillation was attainedbetween 10 to 20 days of age. Ofnote, the authors show oscillationof Per-1 in the liver at postnatalday 2, coinciding with the findingsin neonatal rats of Figure 3 (fromSaxena et al., 2007). The emerg-ing picture from the data accruedin the last few years is that clockgenes are expressed very early indevelopment and most likely inmany embryonic/fetal tissues.Whether they represent functionalcircadian clocks in the sense thatwe understand in the adult or serveother functions is presentlyunknown. Parsimony is used widelyin biology, and it is conceivable thatduring development some of theclock genes may serve functionsnot related to circadian timekeeping(Miller et al., 2007).
MATERNAL SIGNALS
ENTRAINING FETAL AND
NEWBORN CIRCADIAN
RHYTHMS
As described before, the fetalrhythms in precocial species orneonatal rhythms in altricial spe-cies and the rhythms of expressionBmal-1 and Per-2 in the fetal cap-uchin SCN and fetal adrenal glandwere entrained to clock time. Arecent experiment, using clockgene mutant mice, showed loss ofsynchrony in the postnatal rhythmof activity in heterozygous pupsof arrhythmic double knock-out mPer1Brdn/Per2Brdnmice andmPer2Brdn/mCry1 mice (Jud andAlbrecht, 2006), confirming againthat the rhythm of activity is notinduced by the mother but itrequires maternal signals for syn-chronization. As mentioned before,fetuses are subject to a variety ofnonphotic stimuli originating fromthe mother, including circadian
Figure 5. Distribution of the mean hour of drinking in individual pups between threeand six weeks of age. Pups were born to dams sham operated (control), pinealectom-ized during pregnancy, and pinealectomized dams receiving daily melatonin replace-ment. Each symbol represents a pup. The length of the arrow indicates the degree ofsynchrony of each group of pups. An absent arrow indicates no synchrony. [Redrawnfrom Bellavia et al. (2006).]
210 SERON-FERRE ET AL.
Birth Defects Research (Part C) 81:204–214, (2007)

fluctuations in hormones, nu-trients, and uterine motility.Maternal signals are most likelyredundant, and may have differenttargets in the embryo/fetus duringdevelopment. There is evidencefor two maternal signals actingduring gestation on entrainment ofnewborn rhythms: maternal feed-ing time and maternal melatonin.In SCN-lesioned pregnant rats,restricting feeding to a 4-hr periodrestores synchronization of thepups’ drinking rhythms (Weaverand Reppert, 1989). As alreadydiscussed, the time of food avail-ability changes the phase of therhythms of corticosterone andlocomotor activity (Krieger et al.,1977), induces an anticipatoryincrease in body temperature(Recabarren et al., 2005), and inaddition changes the phase ofclock gene expression in liverwithout changing the phase in theSCN (Balsalobre, 2002). Thematernal metabolic signal has notbeen identified. However, themore recent findings that bodytemperature and corticosteroneshift the phase of clock genes infibroblasts (Balsalobre et al.,2000; Le Minh et al., 2001; Balsa-lobre, 2002) and in the liver (Bal-salobre et al., 2000) open inter-esting possibilities to identify theZeitgeber mediating the effects ofmaternal restricted feeding on thenewborns. The second maternalsignal studied, melatonin, is one ofthe few maternal hormones cross-ing the placenta without beingaltered. Melatonin is produced bythe mother in a circadian fashion.It has been named the hormone ofthe night, as it is secreted withalmost a square wave pattern,concentration very low at daytime,increasing sharply at the begin-ning of the night (when lights areoff), and decreasing abruptly atdawn (when lights are on). Theduration of the night increase inmelatonin, signals the length ofthe photoperiod and thus seasonof the year. Pineal melatonin syn-thesis starts postnatally in the rat(Deguchi, 1975), sheep (Nowaket al., 1990), and human (Kenn-away et al., 1992); however,maternal melatonin generates a
rhythm of melatonin in the fetalcirculation (Yellon and Longo,1988). Melatonin receptors arepresent in the human, capuchinmonkey, and rodent fetal SCN inaddition to other sites in thehuman fetal brain (Weaver andReppert, 1996; Thomas et al.,2002), fetal kidney (Drew et al.,1998), and as mentioned, in thecapuchin fetal adrenal gland andfetal SCN (Torres-Farfan et al.,2004, 2006a). Experimental evi-dence supports maternal melato-nin being an entraining signal forpostnatal behavioral rhythms inhamsters and rats and for therhythm of clock gene expression inthe capuchin SCN. Timed injection
of melatonin into SCN-lesionedmaternal hamsters restores syn-chrony in the rhythm of activity ofthe newborns (Davis and Mannion,1988). Similarly, maternal melato-nin replacement during late gesta-tion restores synchronization ofthe rhythm of drinking water in25-day-old pups of pinealectom-ized mothers (Bellavia et al.,2006) (Fig. 4). The site of actionof melatonin is presumed to bethe developing fetal SCN. Directevidence for maternal melatoninas a strong Zeitgeber for the fetalcapuchin SCN was obtained by usin the capuchin monkey (Torres-Farfan et al., 2006a). Suppressionof maternal melatonin secretion byexposing pregnant females to con-stant light during the last third ofgestation, shifted Bmal-1 and MT1melatonin receptor expression inthe fetal capuchin monkey SCNand this shift was reverted bydaily melatonin replacement. Alto-gether, these data support a rolefor maternal melatonin as a Zeit-geber for the fetal primate SCN,and rat and hamster SCN. How-ever, as usually happens inresearch, experiments answer onequestion and raise a few more. Inthe experiments in the capuchinjust described, neither adrenalclock gene expression nor markersof circadian fetal adrenal functionwere modified by suppressingmaternal melatonin. This observa-tion is in conflict with a vision ofthe fetal SCN being a master clockentrained by the mother that com-mands a putative fetal adrenalclock and probably other fetal pe-ripheral clocks not yet identified.It rather suggests that, at least inthe capuchin monkey, the fetalSCN is not a master clock for thefetal adrenal. More research isneeded to identify whether circa-dian clocks are present in other fe-tal organs and whether they arecommanded by the fetal SCN. Anintriguing possibility, that requiresfurther investigation, is that dur-ing fetal life the fetal SCN and dis-tinct fetal organs may be periph-eral clocks to the mother, andentrained by diverse maternal sig-nals, one of them being melatoninfor the fetal SCN (Fig. 6).
Figure 6. Schematic representation ofthe proposed entrainment pathway ofcapuchin monkey fetal SCN and fetal pe-ripheral oscillators. The fetal SCN isentrained by the rhythm of maternal mel-atonin, whereas the fetal adrenal andpossibly other fetal peripheral clocks arephase-entrained by the maternal SCNthrough humoral or metabolic signalsthat cross the placenta. [Based on dataof Torres-Farfan et al. (2006a).]
CIRCADIAN CLOCKS 211
Birth Defects Research (Part C) 81:204–214, (2007)

IMPORTANCE OF THE
CIRCADIAN SYSTEM
DURING DEVELOPMENT
AND FOR PREPARATION
FOR POSTNATAL LIFE
An important fact is that 20% ofthe working population is sub-jected to a shift-work schedule,with no exceptions being made forworking pregnant women. Therelationship between exposure tochanged photoperiod and preg-nancy and fetal outcome is notclear. However, an increased riskof preterm delivery and low weightat birth in women under systemshift-work schedule has beendemonstrated (Zhu et al., 2004;Croteau et al., 2006). Theincreased incidence of metabolicsyndrome in shift workers(Karlsson et al., 2001) may alterthe maternal environment. Addi-tionally, the growing evidence ofthe long-term impact of pregnancyenvironment programming sus-ceptibility to diseases that appearin adult life, such as diabetes,hypertension, and metabolic syn-drome (Barker et al., 2006; Ham-mer and Stewart, 2006; Fowdenet al., 2006; Nathanielsz, 2006),further stimulates the need tounderstand the development ofthe circadian system. Although thevery fact that a number of clockgene knockout animals are born,progress to adulthood, and repro-duce suggests that developmentwas not interrupted, the study ofthese clock gene mutant modelsindicates that the integrity of thecircadian system is paramount fornormal metabolic function(s) dur-ing adult life. For instance, lack ofClock expression is transduced toadipocyte hypertrophy and lipidengorgement of hepatocytes, withprominent glycogen accumulation,while plasma exhibits hypercholes-terolemia, hypertriglyceridemia,hyperglycemia, hypoinsulinemia,and lower corticosterone concen-tration than wild-type mice (Tureket al., 2005; Shimba et al., 2005).It is interesting to point out recentfindings in the human of associa-tion of clock gene defects toadvanced sleep syndrome (Jones
et al., 1999; Toh et al., 2001; Xuet al., 2007) and breast cancer(Chen et al., 2005). Altogether,the evidence available supportsthat the normal circadian systemis involved in several physiologicalprocesses and also in metabolicbalance; whether these mecha-nisms are operating in fetal liferemains to be investigated. More-over, understanding the circadianfactors helping life in utero haspractical importance for neonatalcare of preterm infants abruptlydeprived of circadian signals. Inthis regard, the positive impact ofexposing the preterm newborn toa light:dark cycle is being recog-nized, while the possible therapeu-tic use in preterm infants of mela-tonin is being proposed (Rivkees,2003; Jan et al., 2007). The circa-dian timing system is emerging asan important factor during devel-opment. Rapid methodologicaladvances combining in vivo and invitro experiments, together withthe incorporation of new findingson the mechanisms regulating cir-cadian clocks in the adult, mayhelp to unravel how the develop-ing embryo/fetus faces time in thisplastic period of life.
REFERENCES
Adam CL, Kyle CE, Young P. 1994.Influence of prenatal photoperiod onpostnatal reproductive developmentin male red deer (Cervus elaphus).J Reprod Fertil 100:607–611.
Alonso-Vale MI, Andreotti S, Peres SB,et al. 2005. Melatonin enhances lep-tin expression by rat adipocytes inthe presence of insulin. Am J PhysiolEndocrinol Metab 288:E805–E812.
Balsalobre A, Brown SA, Marcacci L,et al. 2000. Resetting of circadian timein peripheral tissues by glucocorticoidsignaling. Science 289:2344–2347.
Balsalobre A. 2002. Clock genes inmammalian peripheral tissues. CellTissue Res 309:193–199.
Barker DJ, Bagby SP, Hanson MA.2006. Mechanisms of disease: inutero programming in the pathoge-nesis of hypertension. Nat Clin PractNephrol 2:700–707.
Bellavia SL, Carpentieri AR, Vaque AM,et al. 2006. Pup circadian rhythmentrainment—effect of maternalganglionectomy or pinealectomy.Physiol Behav 89:342–349.
Boden MJ, Kennaway DJ. 2006. Circa-dian rhythms and reproduction.Reproduction 132:379–392.
Breen S, Rees S, Walker D. 1996. Thedevelopment of diurnal rhythmicityin fetal suprachiasmatic neurons asdemonstrated by fos immunohisto-chemistry.Neuroscience 74:917–926.
Cannon B, Nedergaard J. 2004. Brownadipose tissue: function and physio-logical significance. Physiol Rev 84:277–359.
Caldelas I, Tejadilla D, Gonzalez B,et al. 2007. Diurnal pattern of clockgene expression in the hypothalamusof the newborn rabbit. Neuroscience144:395–401.
Chen ST, Choo KB, Hou MF, et al. 2005.Deregulated expression of the PER1,PER2 and PER3 genes in breast can-cers. Carcinogenesis 26:1241–1246.
Constandil L, Parraguez VH, TorrealbaF, et al. 1995. Day-night changes inc-fos expression in the fetal sheepsuprachiasmatic nucleus at late ges-tation. Reprod Fertil Dev 7:411–413.
Croteau A, Marcoux S, Brisson C.2006. Work activity in pregnancy,preventive measures, and the risk ofdelivering a small-for-gestational-ageinfant. Am J Public Health96:846–855.
Davis FC, Gorski RA. 1985. Develop-ment of hamster circadian rhythms.I. Within-litter synchrony of motherand pups activity rhythms at wean-ing. Biol Reprod 33:353–362.
Davis FC, Mannion J. 1988. Entrain-ment of hamster pup circadianrhythms by prenatal melatonin injec-tions to the mother. Am J Physiol255:439–448.
Davis FC, Reppert SM. 2001. Deve-lopment of mammalian circadianrhythms. In: Takahashi JS, TurekFW, Moore RY, editors. Handbookof Behavioral Neurobiology. Vol 12.Circadian Clocks. New York: KluwerAcademic/Plenum Publishers. pp. 247–290.
Damiola F, Le Minh N, Preitner N, et al.2000. Restricted feeding uncouplescircadian oscillators in peripheral tis-sues from the central pacemaker inthe suprachiasmatic nucleus. GenesDev 14:2950–2961.
Deguchi T. 1975. Ontogenesis of a bio-logical clock for serotonin: acetylcoenzyme A N-acetyltransferase inpineal gland of rat. Proc Natl AcadSci USA 72:2814–2818.
Drew JE, Williams LM, Hannah LT,et al. 1998. Melatonin receptors inthe human fetal kidney: 2-[125I]iodomelatonin binding sitescorrelated with expression of Mel1aand Mel1b receptor genes. J Endocri-nol 156:261–267.
Ebling FJ, Wood RI, Suttie JM, et al.1989. Prenatal photoperiod influencesneonatal prolactin secretion in thesheep. Endocrinology 125:384–391.
Edery I. 2000. Circadian rhythms in anutshell. Physiol Genomics 3:59–74.
Fahrenkrug J, Georg B, Hannibal J, et al.2006. Diurnal rhythmicity of the clockgenes Per1 and Per2 in the rat ovary.Endocrinology 147:3769–3776.
212 SERON-FERRE ET AL.
Birth Defects Research (Part C) 81:204–214, (2007)

Fowden AL, Giussani DA, Forhead AJ.2006. Intrauterine programming ofphysiological systems: causes andconsequences. Physiology 21:29–37.
Gonzalez-Mariscal G. 2007. Mother rab-bits and their offspring: timing is every-thing. DevPsychobiol 49:71–76.
Gorman MR, Goldman BD, Zucker I.2001. Mammalian photoperiodism.In: Takahashi JS, Turek FW, MooreRY, editors.Handbook of BehavioralNeurobiology. Vol 12. Circadian Clocks.New York: Kluwer Academic/PlenumPublishers. pp. 481–528.
Grechez-Cassiau A, Panda S, LacocheS, et al. 2004. The transcriptionalrepressor STRA13 regulates a subsetof peripheral circadian outputs. J BiolChem 279:1141–1150.
Grumbach MM, Kaplan SL. 1998. Thepituitary. In: Polin RA, Fox WW, edi-tors. Fetal and Neonatal Physiology.2nd ed. Philadelphia: WB Saunders.pp. 2395–2442.
Guo H, Brewer JM, Champhekar A,et al. 2005. Differential control of pe-ripheral circadian rhythms by supra-chiasmatic-dependent neural signals.Proc Natl Acad Sci USA 102:3111–3116.
Guo H, Brewer JM, Lehman MN, BittmanEL. 2006. Suprachiasmatic regulationof circadian rhythms of gene expres-sion in hamster peripheral organs:effects of transplanting the pacemaker.J Neurosci 26:6406–6412.
Hammer F, Stewart PM. 2006. Cortisolmetabolism in hypertension. BestPract Res Clin Endocrinol Metab 20:337–353.
Honma S, Kawamoto T, Takagi Y, et al.2002. Dec1 and Dec2 are regulatorsof the mammalian molecular clock.Nature 419:841–844.
Horard B, Rayet B, Triqueneaux G,et al. 2004. Expression of the orphannuclear receptor ERRalpha is undercircadian regulation in estrogen-responsive tissues. J Mol Endocrinol33:87–97.
Ishida A, Mutoh T, Ueyama T, et al.2005. Light activates the adrenalgland: timing of gene expression andglucocorticoids release. Cell Metab2:297–307.
Jan JE, Wasdell MB, Freeman RD, BaxM. 2007. Evidence supporting theuse of melatonin in short gestationinfants. J Pineal Res 42:22–27.
Johnson MH, Lim A, Fernando D, DayML. 2002. Circadian clockwork genesare expressed in the reproductivetract and conceptus of the earlypregnant mouse. Reprod BiomedOnline 4:140–145.
Johnston JD, Ebling FJ, Hazlerigg DG.2005. Photoperiod regulates multiplegene expression in the suprachias-matic nuclei and pars tuberalis of theSiberian hamster (Phodopus sun-gorus). Eur J Neurosci 21:2967–2974.
Johnston JD, Schuster C, Barrett P,Hazlerigg DG. 2007. Regulation ofthe ovine MT1 melatonin receptor
promoter: interaction between multi-ple pituitary transcription factors atdifferent phases of development. MolCell Endocrinol 268:59–66.
Jones CR, Campbell SS, Zone SE, et al.1999. Familial advanced sleep-phasesyndrome: a short-period circadianrhythm variant in humans. Nat Med5:1062–1065.
Jud C, Albrecht U. 2006. Circadianrhythms in murine pups develop inabsence of a functional maternal cir-cadian clock. J Biol Rhythms 21:149–154.
Karlsson B, Knutsson A, Lindahl B.2001. Is there an associationbetween shift work and having ametabolic syndrome? Results from apopulation based study of 27,485 peo-ple. OccupEnvironMed 58:747–752.
Karman BN, Tischkau SA. 2006. Circa-dian clock gene expression in theovary: effects of luteinizing hormone.Biol Reprod 75:624–632.
Kennaway DJ, Stamp GE, Goble FC.1992. Development of melatoninproduction in infants and the impactof prematurity. J Clin EndocrinolMetab 75:367–379.
Kennaway DJ, Varcoe TJ, Mau VJ.2003. Rhythmic expression of clockand clock-controlled genes in the ratoviduct. Mol Hum Reprod 9:503–507.
Ko MS, Kitchen JR, Wang X, et al.2000. Large-scale cDNA analysisreveals phased gene expression pat-terns during preimplantation mousedevelopment. Development 127:1737–1749.
Krieger DT, Hauser H, Krey LC. 1977.Suprachiasmatic nuclear lesions donot abolish food-shifted circadian ad-renal and temperature rhythmicity.Science 197:398–399.
Le Minh N, Damiola F, Tronche F, et al.2001 Glucocorticoid hormones inhibitfood-induced phase-shifting of pe-ripheral circadian oscillators. EMBO J17:7128–136.
Lee C, Etchegaray JP, Cagampang FR,et al. 2001. Posttranslational mecha-nisms regulate the mammalian circa-dian clock. Cell 107:855–867.
Lemos DR, Downs JL, Urbanski HF.2006. Twenty-four hour rhythmicgene expression in the rhesus maca-que adrenal gland. Mol Endocrinol20:1164–1176.
Levi F, Schibler U. 2007. Circadianrhythms: mechanisms and therapeu-tic implications. Annu Rev PharmacolToxicol 47:593–628.
Li X, Davis FC. 2005. Developmentalexpression of clock genes in the Syr-ian hamster. Brain Res Dev Brain Res158:31–40.
Liu C, Li S, Liu T, et al. 2007. Tran-scriptional coactivator PGC-1alphaintegrates the mammalian clock andenergy metabolism. Nature 447:477–481.
Lincoln G, Messager S, Andersson H,Hazlerigg D. 2002. Temporal expres-
sion of seven clock genes in thesuprachiasmatic nucleus and thepars tuberalis of the sheep: evidencefor an internal coincidence timer.Proc Natl Acad Sci USA 99:13890–13895.
Lincoln GA, Andersson H, Hazlerigg D.2003. Clock genes and the long-termregulation of prolactin secretion: evi-dence for a photoperiod/circannualtimer in the pars tuberalis. J Neuro-endocrinol 15:390–397.
Lowrey PL, Shimomura K, Antoch MP,et al. 2000. Positional syntenic clon-ing and functional characterization ofthe mammalian circadian mutationtau. Science 288:483–492.
Maeda Y, Muro M, Shono M, et al.2006. Diurnal rhythms in fetal heartrate baseline and sustained fetaltachycardia in twin pregnancy. EarlyHum Dev 82:637–644.
Merrow M, Mazzotta G, Chen Z, Roen-neberg T. 2006. The right place atthe right time: regulation of dailytiming by phosphorylation. GenesDev 20:2629–2633.
Meyer-Bernstein EL, Jetton AE, Matsu-moto SI, et al. 1999. Effects ofsuprachiasmatic transplants on circa-dian rhythms of neuroendocrinefunction in golden hamsters. Endocri-nology 140:207–218.
Miller BH, McDearmon EL, Panda S, et al.2007. Circadian and CLOCK-controlledregulation of the mouse transcriptomeand cell proliferation. Proc Natl Acad SciUSA 104:3342–3347.
Moore-Ede MC, Sulzman FM, Fuller CA.1982. Characteristics of a circadiana-lock. In: The clocks that time us.Cambridge, MA: Harvard UniversityPress. pp. 30–133.
Nathanielsz PW. 2006. Animal modelsthat elucidate basic principles of thedevelopmental origins of adult dis-eases. ILAR J 47:73–82.
Nowak R, Young IR, McMillen IC. 1990.Emergence of the diurnal rhythm inplasma melatonin concentrations innewborn lambs delivered to intact orpinealectomized ewes. J Endocrinol125:97–102.
Ohta H, Honma S, Abe H, Honma K.2002. Effects of nursing mothers onrPer1 and rPer2 circadian expres-sions in the neonatal rat suprachias-matic nuclei vary with developmentalstage. Eur J Neurosci 15:1953–1960.
Oishi K, Sakamoto K, Okada T, et al.1998a. Antiphase circadian expres-sion between BMAL1 and periodhomologue mRNA in the suprachias-matic nucleus and peripheral tissuesof rats. Biochem Biophys Res Com-mun 253:199–203.
Oishi K, Sakamoto K, Okada T, et al.1998b Humoral signals mediate thecircadian expression of rat periodhomologue (rPer2) mRNA in periph-eral tissues. Neurosci Lett 256:117–119.
Oishi K, Amagai N, Shirai H, et al.2005. Genome-wide expression anal-
CIRCADIAN CLOCKS 213
Birth Defects Research (Part C) 81:204–214, (2007)

ysis reveals 100 adrenal gland-dependent circadian genes in themouse liver. DNA Res 12:191–202.
Okamura H, Yamaguchi S, Yagita K.2002. Molecular machinery of the cir-cadian clock in mammals. Cell TissueRes 309:47–56.
Oster H, Damerow S, Kiessling S, et al.2006. The circadian rhythm of gluco-corticoids is regulated by a gatingmechanism residing in the adrenalcortical clock. Cell Metab 4:163–173.
Panda S, Antoch MP, Miller BH, et al.2002. Coordinated transcription ofkey pathways in the mouse by thecircadian clock. Cell 109:307–320.
Parraguez VH, Valenzuela GJ, VergaraM, et al. 1996. Effect of constantlight on fetal and maternal prolactinrhythms in sheep. Endocrinology137:2355–2361.
Pittendrigh C. 1981. Circadian sys-tems: general perspective. In: Asch-off J, editor.Handbook of behavioralneurobiology. Vol. 4: Biological rhy-thms. New York: Plenum Press.pp. 57–80.
Polk DH, Fisher DA. 1998. The fetal ad-renal and fetal thyroid systems In:Polin RA, Fox WW, editors. Fetal andneonatal physiology. 2nd ed. Philadel-phia: WB Saunders. pp. 2460–2467.
Preitner N, Damiola F, Lopez-Molina L,et al. 2002. The orphan nuclear re-ceptor REV-ERBalpha controls circa-dian transcription within the positivelimb of the mammalian circadian os-cillator. Cell 110:251–260.
Recabarren MP, Valdes JL, Farias P,et al. 2005. Differential effects ofinfralimbic cortical lesions on tem-perature and locomotor activity re-sponses to feeding in rats. Neuro-science 134:1413–1422.
Reppert SM, Schwartz WJ. 1983.Maternal coordination of the fetalbiological clock in utero. Science220:969–971.
Reppert SM, Schwartz WJ. 1984. Func-tional activity of the suprachiasmaticnuclei in the fetal primate. NeurosciLett 46:145–149.
Reppert SM, Uhl GR. 1987 Vasopressinmessenger ribonucleic acid in supra-optic and suprachiasmatic nuclei:appearance and circadian regulationduring development. Endocrinology120:2483–2487.
Reppert SM, Weaver DR. 2002. Coordi-nation of circadian timing in mam-mals. Nature 418:935–941.
Richter HG, Torres-Farfan C, Rojas-Gar-cıa P, et al. 2004. The circadian timingsystem: making sense of day/nightgene expression. Biol Res 37:11–28.
Rivkees SA. 2003. Developing circa-dian rhythmicity in infants. PediatrEndocrinol Rev 1:38–45.
Sakamoto K, Nagase T, Fukui H, et al.1998. Multitissue circadian expres-sion of rat period homolog (rPer2)
mRNA is governed by the mamma-lian circadian clock, the suprachias-matic nucleus in the brain. J BiolChem 273:27039–27042.
Saxena MT, Aton SJ, Hildebolt C, et al.2007. Bioluminescence imaging ofperiod1 gene expression in utero.Mol Imaging 6:68–72.
Seron-Ferre M, Ducsay CA, ValenzuelaGJ. 1993. Circadian rhythms duringpregnancy. Endocr Rev 14:594–609.
Seron-Ferre M, Torres-Farfan C, Force-lledo ML, Valenzuela GJ. 2001. Thedevelopment of circadian rhythms inthe fetus and neonate. Semin Perina-tol 25:363–370.
Shibata S, Moore RY. 1987. Develop-ment of neuronal activity in the ratsuprachiasmatic nucleus. Brain Res431:311–315.
Shimba S, Ishii N, Ohta Y, et al. 2005.Brain and muscle Arnt-like protein-1(BMAL1), a component of the molecu-lar clock, regulates adipogenesis. ProcNatl Acad Sci USA 102:12071–12076.
Shimomura H, Moriya T, Sudo M, et al.2001. Differential daily expression ofPer1 and Per2 mRNA in the supra-chiasmatic nucleus of fetal and earlypostnatal mice. Eur J Neurosci13:687–693.
Sladek M, Sumova A, Kovacikova Z,et al. 2004. Insight into molecularcore clock mechanism of embryonicand early postnatal rat suprachias-matic nucleus. Proc Natl Acad SciUSA 101:6231–6236.
Sladek M, Jindrakova Z, Bendova Z,Sumova A. 2007. Postnatal ontogen-esis of the circadian clock within therat liver. Am J Physiol Regul IntegrComp Physiol 292:R1224–R1229.
Stokkan KA, Yamazaki S, Tei H, et al.2001. Entrainment of the circadianclock in the liver by feeding. Science291:490–493.
Stratmann M, Schibler U. 2006 Proper-ties, entrainment, and physiologicalfunctions of mammalian peripheraloscillators. J Biol Rhythms 21:494–506.
Thomas L, Purvis CC, Drew JE, et al.2002. Melatonin receptors in humanfetal brain: 2-[(125)I]iodomelatoninbinding and MT1 gene expression.J Pineal Res 33:218–224.
Toh KL, Jones CR, He Y, et al. 2001.An hPer2 phosphorylation site muta-tion in familial advanced sleepphase syndrome. Science 291:1040–1043.
Torres-Farfan C, Richter HG, GermainAM, et al. 2004. Maternal melatoninselectively inhibits cortisol productionin the primate fetal adrenal gland. JPhysiol 554:841–856.
Torres-Farfan C, Rocco V, Monso C,et al. 2006a. Maternal melatonineffects on clock gene expression in anonhuman primate fetus. Endocrinol-ogy 147:4618–4626.
Torres-Farfan C, Seron-Ferre M, DinetV, Korf HW. 2006b. Immunocyto-chemical demonstration of day/nightchanges of clock gene protein levelsin the murine adrenal gland: dif-ferences between melatonin-profi-cient (C3H) and melatonin-deficient(C57BL) mice. J Pineal Res 40:64–70.
Turek FW, Joshu C, Kohsaka A, et al.2005. Obesity and metabolic syn-drome in circadian Clock mutantmice. Science 308:1043–1045.
Ueda HR, Hayashi S, Chen W, et al.2005. System-level identification oftranscriptional circuits underlyingmammalian circadian clocks. NatGenet 37:187–192.
Vanselow K, Vanselow JT, WestermarkPO, et al. 2006. Differential effects ofPER2 phosphorylation: molecular ba-sis for the human familial advancedsleep phase syndrome (FASPS).Genes Dev 20:2660–2672.
Watanabe T, Kojima M, Tomida S,et al. 2006. Peripheral clock geneexpression in CS mice with bimodallocomotor rhythms. Neurosci Res54:295–301.
Weaver DR, Reppert SM. 1989. Peri-odic feeding of SCN-lesioned preg-nant rats entrains the fetal biologicalclock. Brain Res Dev Brain Res46:291–296.
Weaver DR, Reppert SM. 1996. TheMel1a melatonin receptor gene isexpressed in human suprachiasmaticnuclei. Neuroreport 8:109–112.
Weinert D. 2005. Ontogenetic de-velopment of the mammalian circadiansystem. Chronobiol Int 22:179–205.
Xu Y, Toh KL, Jones CR, et al. 2007.Modeling of a human circadianmutation yields insights into clockregulation by PER2. Cell. 128:59–70.
Yellon SM, Longo LD. 1988. Effect ofmaternal pinealectomy and reversephotoperiod on the circadian melato-nin rhythm in the sheep, fetus duringthe last trimester of pregnancy. BiolReprod 39:1093–1099.
Yoo SH, Yamazaki S, Lowrey PL, et al.2004. PERIOD2::LUCIFERASE real-time reporting of circadian dynamicsreveals persistent circadian oscilla-tions in mouse peripheral tissues.Proc Natl Acad Sci USA 101:5339–5346.
Zhu JL, Hjollund NH, Olsen J. 2004.Shift work, duration of pregnancy,and birth weight: the National BirthCohort in Denmark. Am J ObstetGynecol 191:285–291.
Zylka MJ, Shearman LP, Weaver DR,Reppert SM. 1998. Three periodhomologs in mammals: differentiallight responses in the suprachias-matic circadian clock and oscillatingtranscripts outside of brain. Neuron20:1103–1110.
214 SERON-FERRE ET AL.
Birth Defects Research (Part C) 81:204–214, (2007)
Related Documents











