Arch Pharm Res Vol 35, No 5, 887-895, 2012 DOI 10.1007/s12272-012-0514-z 887 Chrysophanol-induced Necrotic-like Cell Death through an Impaired Mitochondrial ATP Synthesis in Hep3B Human Liver Cancer Cells Chien-Hang Ni 1,* , Po-Yuan Chen 2,* , Hsu-Feng Lu 3 , Jai-Sing Yang 4 , Hui-Ying Huang 5 , Shin-Hwar Wu 6,7 , Siu-Wan Ip 5 , Chin-Tung Wu 6 , Su-Yin Chiang 8 , Jaung-Geng Lin 9 , W. Gibson Wood 10 , and Jing-Gung Chung 2,11 1 Department of Chinese Medicine, E-DA Hospital/I-Shou University, Kaohsiung 824, Taiwan, 2 Department of Biological Science and Technology, China Medical University, Taichung 404, Taiwan, 3 Department of Clinical Pathology, Cheng Hsin General Hospital, Taipei 112, Taiwan, 4 Department of Pharmacology, China Medical University, Taichung 404, Taiwan, 5 Department of Nutrition, China Medical University, Taichung 404, Taiwan, 6 Department of Agronomy, National Chung Hsing University, Taichung 402, Taiwan, 7 Graduate Institute of Clinical Medical Science, China Medical University, Tai- chung 404, Taiwan, 8 Division of Critical Care Medicine, Department of Internal Medicine, Changhua Christian Hospital, Changhua 500, Taiwan, 9 School of Chinese Medicine, China Medical University, Taichung 404, Taiwan, 10 Department of Pharmacology, University of Minnesota, School of Medicine, Geriatric Research, Education and Clinical Center, VA Medi- cal Center, Minneapolis, MN 55455, USA, and 11 Department of Biotechnology, Asia University, Taichung 413, Taiwan (Received July 19, 2011/Revised October 7, 2011/Accepted November 7, 2011) Liver cancer is the most common form of cancer in Taiwan and it usually responds to chemo- therapy. However, patients often have side effects to the chemotherapeutic drugs. Thus new agents are urgently required to treat liver cancer. Chrysophanol, one of the anthraquinone derivatives, was reported to inhibit some human cancer cell growth which may be due to the induction of apoptosis similar to other anthraquinone derivatives though such actions have not been reported. In the present study, we reported that chrysophanol inhibits cell growth in Hep3B liver cancer cells based on the following observations: 1) induc cell morphological changes; 2) decreased percentage of viable cells; 3) induced S phase arrest of cell cycle progres- sion; 4) induced DNA damage as measured by comet assay and DAPI staining. Chrysophanol- induced cell death however, seems to be related to necrotic processes rather than typical apo- ptosis. Chrysophanol induced reactive oxygen species and Ca 2+ production and decreased mitochondrial membrane potential (∆Ψm) and ATP levels in Hep3B cells. No effects were observed on known protein regulators of apoptosis such as Bax and Bcl-2. Chrysophanol- induced cell death took place independently of caspase-8 and -9. Based on our findings, we propose that chrysophanol reduces cellular ATP levels causing a drop in energy resulting in necrotic-like cell death. Key words: Chrysophanol, Necrosis, Human liver cancer cells (Hep3B), Reactive oxygen spe- cies, Mitochondrial membrane potential INTRODUCTION Cancer is a major cause of death in populations worldwide. In Taiwan, liver cancer is the second leading cause of cancer death with 33.8 individuals per 100,000 dying of liver cancer in 2009 (Department of Health, Taiwan). Treatment options for liver cancer are not satisfactory and novel therapies are needed to treat this devastating disease. A type of novel therapies with traditional herbs or natural products has been used for the treatment of cancer patients (Aravindaram and Yang, 2010). There has been considerable interest in anthraquin- one derivatives including emodin, aloe-emodin, phy- scion, chrysophanol, rhein, danthron, emodin glycoside and chrysophanol glycoside (Verma et al., 2005; Huang et al., 2007). Emodin, aloe-emodin and rhein inhibited cell growth through cell cycle arrest and induction of *These authors contributed equally to this work. Correspondence to: Jing-Gung Chung, Department of Biological Science and Technology, China Medical University, Taichung 40402, Taiwan Tel: 886-4-22053366 ext 2161, Fax: 886-4-22053764 E-mail: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Arch Pharm Res Vol 35, No 5, 887-895, 2012
DOI 10.1007/s12272-012-0514-z
887
Chrysophanol-induced Necrotic-like Cell Death through an Impaired Mitochondrial ATP Synthesis in Hep3B Human Liver Cancer Cells
Chien-Hang Ni1,*, Po-Yuan Chen2,*, Hsu-Feng Lu3, Jai-Sing Yang4, Hui-Ying Huang5, Shin-Hwar Wu6,7,Siu-Wan Ip5, Chin-Tung Wu6, Su-Yin Chiang8, Jaung-Geng Lin9, W. Gibson Wood10, and Jing-Gung Chung2,11
1Department of Chinese Medicine, E-DA Hospital/I-Shou University, Kaohsiung 824, Taiwan, 2Department of BiologicalScience and Technology, China Medical University, Taichung 404, Taiwan, 3Department of Clinical Pathology, Cheng HsinGeneral Hospital, Taipei 112, Taiwan, 4Department of Pharmacology, China Medical University, Taichung 404, Taiwan,5Department of Nutrition, China Medical University, Taichung 404, Taiwan, 6Department of Agronomy, National ChungHsing University, Taichung 402, Taiwan, 7Graduate Institute of Clinical Medical Science, China Medical University, Tai-chung 404, Taiwan, 8Division of Critical Care Medicine, Department of Internal Medicine, Changhua Christian Hospital,Changhua 500, Taiwan, 9School of Chinese Medicine, China Medical University, Taichung 404, Taiwan, 10Department ofPharmacology, University of Minnesota, School of Medicine, Geriatric Research, Education and Clinical Center, VA Medi-cal Center, Minneapolis, MN 55455, USA, and 11Department of Biotechnology, Asia University, Taichung 413, Taiwan
(Received July 19, 2011/Revised October 7, 2011/Accepted November 7, 2011)
Liver cancer is the most common form of cancer in Taiwan and it usually responds to chemo-therapy. However, patients often have side effects to the chemotherapeutic drugs. Thus newagents are urgently required to treat liver cancer. Chrysophanol, one of the anthraquinonederivatives, was reported to inhibit some human cancer cell growth which may be due to theinduction of apoptosis similar to other anthraquinone derivatives though such actions havenot been reported. In the present study, we reported that chrysophanol inhibits cell growth inHep3B liver cancer cells based on the following observations: 1) induc cell morphologicalchanges; 2) decreased percentage of viable cells; 3) induced S phase arrest of cell cycle progres-sion; 4) induced DNA damage as measured by comet assay and DAPI staining. Chrysophanol-induced cell death however, seems to be related to necrotic processes rather than typical apo-ptosis. Chrysophanol induced reactive oxygen species and Ca2+ production and decreasedmitochondrial membrane potential (∆Ψm) and ATP levels in Hep3B cells. No effects wereobserved on known protein regulators of apoptosis such as Bax and Bcl-2. Chrysophanol-induced cell death took place independently of caspase-8 and -9. Based on our findings, wepropose that chrysophanol reduces cellular ATP levels causing a drop in energy resulting innecrotic-like cell death.
Key words: Chrysophanol, Necrosis, Human liver cancer cells (Hep3B), Reactive oxygen spe-cies, Mitochondrial membrane potential
INTRODUCTION
Cancer is a major cause of death in populations
worldwide. In Taiwan, liver cancer is the second
leading cause of cancer death with 33.8 individuals
per 100,000 dying of liver cancer in 2009 (Department
of Health, Taiwan). Treatment options for liver cancer
are not satisfactory and novel therapies are needed to
treat this devastating disease. A type of novel therapies
with traditional herbs or natural products has been
used for the treatment of cancer patients (Aravindaram
and Yang, 2010).
There has been considerable interest in anthraquin-
one derivatives including emodin, aloe-emodin, phy-
scion, chrysophanol, rhein, danthron, emodin glycoside
and chrysophanol glycoside (Verma et al., 2005; Huang
et al., 2007). Emodin, aloe-emodin and rhein inhibited
cell growth through cell cycle arrest and induction of
*These authors contributed equally to this work.
Correspondence to: Jing-Gung Chung, Department of Biological
Science and Technology, China Medical University, Taichung
40402, Taiwan
Tel: 886-4-22053366 ext 2161, Fax: 886-4-22053764
E-mail: [email protected]

888 C.-H. Ni et al.
apoptosis in many cancer cell lines (Su et al., 2005;
Lin SY et al., 2009; Aviello et al., 2010; Lin ML et al.,
2010). There are no reports showing that chrysopha-
nol induced apoptosis in human cancer cells. However,
chrysophanol was found to inhibit proliferation in leu-
kemia cells (Ueno et al., 1995), MDA-MB-231 human
breast cancer cells (Kang et al., 2008) and liver cancer
cells (Lu et al., 2010a). The purpose of the present
study was to determine if effects of chrysophanol on
human liver cells was associated with apoptosis or
necrosis. Chrysophanol induced the inhibition of cell
growth of Hep3B human liver cancer cells in vitro and
these effects were associated with chrysophanol-induced
necrotic-like cell death and but not apoptosis.
MATERIALS AND METHODS
Chemicals and reagentsChrysophanol, dimethyl sulfoxide (DMSO) and Tris-
HCl were purchased from Sigma Chemical Co. Chry-
sophanol was dissolved in DMSO and stored at −20oC.
Control cultures received the carrier solvent (0.5%
DMSO).
Cell cultureHuman hematoma cell line (Hep3B) were purchased
from the Food Industry Research and Development
Institute and were cultured in Dulbecco's modied
Eagle's medium (DMEM) supplemented with 10% fetal
bovine serum (FBS), 0.1 mg/mL streptomycin, and 100
units/mL penicillin at 37oC in an atmosphere of 5%
CO2. DMEM, fetal bovine serum, L-glutamine, penicillin-
streptomycin and trypsin-EDTA were obtained from
Invitrogen.
Morphology and cell viability assayHep3B cells (2×105 cells/well) were maintained in
12-well plates with DMEM for 24 h and then were
treated with chrysophanol at final concentrations of 0,
5, 10, 25, 50, 75, 100 and 120 µM at 37oC, 5% CO2 and
95% air for 24, 48 and 72 h. Cells were then examined
and photographed under a contrast-phase microscope
at 200× and cells from each groups were individually
harvested from each well by centrifugation. All isolated
cells from each treatment were stained with PI (5 µg/
mL) and analyzed by flow cytometry (Becton Dickinson
FACSCalibur) for determining the percentage of viable
cells as previously described (Lu et al., 2010a; Chiang
et al., 2011).
Cell cycle distribution assayHep3B cells (2×105 cells/well) in 12-well plates with
DMEM were treated with chrysophanol at 0, 1, 5, 10,
25, 50, 75 and 100 µM for 24 h before all samples in
each well were harvested. Treated and controlled cells
from each well were harvested, washed in cold phos-
phate-buffered saline (PBS), fixed in 70% ethanol, and
stored at 4oC. Cells in each tube were washed with
PBS and DNA was treated with RNase A (200 µg/mL)
at 37oC for 15 min and stained by PI (20 µg/mL) and
0.1% Triton X-100 in PBS in a dark room for 30 min
at 37oC. DNA content in each groups were assayed by
flow cytometer and cell fractions in G0/G1, S, and G2/
M phase were analyzed using flow cytometer equipped
with an argon ion laser at 488 nm wavelength (Lu CC
et al., 2010a; Lu HF et al., 2010b). The results were
analyzed by Mod Fit LT 3.0 software.
Comet assay and DAPI stainingIn order to examine whether or not chrysophanol
can induce DNA damage and cell death, Hep3B cells
at a density of 2 × 105 cells/well on 12-well plates were
treated with 0, 10, 25, 50, 75, 100 or 120 µM chryso-
phanol for 48 h or 4 µM hydrogen peroxide (H2O2) (a
positive control group). Cells from each treatment
were harvested and divided into two parts for comet
staining by PI and 4’-6-diamidino-2-phenylindole
(DAPI) staining and then were examined and photo-
graphed by using fluorescence microscopy as describ-
ed elsewhere (Chiang et al., 2006; Chen et al., 2011).
Data was carried out from three independent experi-
ments and have similar results. CometScore software
(Tritek Corp) was used to quantify the DNA damage
and PI-stained DNA tails and the results from comet
tail length were expressed (fold of control) in mean ±
S.D. and at least three independent samples as describ-
ed elsewhere (Chiang et al., 2011; Yu et al., 2011).
Flow cytometric analysis of reactive oxygen
species (ROS) production, mitochondrial mem-
brane potential (∆Ψm), and cytosolic Ca2+ releaseHep3B cells at a density of 2×105 cells/well on 12-
well plates were incubated with 50 µM chrysophanol
for indicated intervals of time. Production of ROS,
∆Ψm, and cytosolic Ca2+ levels were determined. At the
end of the incubation, cells from well were harvested,
washed twice by PBS, and then were re-suspended in
500 µL of DCFH-DA (10 µM) for ROS, in 500 µL of
DiOC6 (1 µmol/L) for the level of ∆Ψm, and in Indo 1/
AM (3 µg/mL) for cytosolic Ca2+ production in a dark
room for 30 min at 37oC. Cells in each treatment were
analyzed immediately by flow cytometry as previously
described (Lin et al., 2007; Lu et al., 2008).
Determination of ATP levelHep3B cells at a density of 1 × 104 cells/well were

Chrysophanol-triggered Necrosis of Hep3B Cells 889
maintained in 100 µL phenol red-free medium with
chrysophanol at different concentrations (0, 25, 50, 75,
100 and 120 µM) for 6 h in 96-well white microplates.
The intracellular ATP content level in each well was
measured using Luminescence ATP Detection Assay
by an ATPliteTM kit (PerkinElmer) as previously de-
scribed (Huan et al., 2006; Lu et al., 2010a) and resulting
luminescence was monitored by SynergyTM HT Multi-
Mode Microplate Reader (BioTek).
Determination of levels of proteins associated
with cell deathHep3B cells at a density of 1 × 106 cells/well on 6-
well plates were incubated with 50 µM chrysophanol
for 0, 6, 12, 24, and 48 h. Cells at the end of each
treatment period were harvested and extracted with
the PRO-PREPTM protein extraction solution (iNtRON
Biotechnology) (Lu et al., 2010a; Liu et al., 2011). The
lysates from each sample were centrifuged at 14000×g
for 30 min and the protein concentration in the super-
natant was determined with a BCA protein assay kit
as previously described (Ji et al., 2009; Lu et al.,
2010a). Equal amounts of protein (40 µg) from each
sample were separated by SDS-polyacrylamide gel
electrophoresis and then were electrotransferred to a
PVDF membrane. The membrane was blocked with a
solution containing 5% non-fat dry milk TBST buffer
(20 mM Tris-HCl, pH 7.4, 150 mM NaCl and 0.1%
Tween 20) for 1 h and washed with TBST buffer. The
primary antibodies [anti-cyclin D, -cyclin-dependent
kinase 2 (CDK2), thymidylate synthase, glutathione-
S-transferase (GST), catalase, superoxide dismutase
(SOD) (Mn), SOD (Cu), cytochrome c, poly(ADP-ribose)
polymerase (PARP), caspase-9, caspase-8, apoptotic
protease activating factor 1 (Apaf-1) and apoptosis-
inducing factor (AIF) (Santa Cruz Biotechnology Inc.)]
were applied and then washed twice followed by staining
with secondary antibody and detected by an ECL kit
(Millipore) and autoradiography using X-ray film.
Statistical analysisData are presented as mean ± S.D. Results are repre-
sentative of three independent experiments. Student’s
t-test for paired comparison was used for statistical
comparisons between the chrysophanol-treated and
control samples. A p value less than 0.05 was used to
denote statistical significance.
RESULTS
Chrysophanol-induced morphological changes
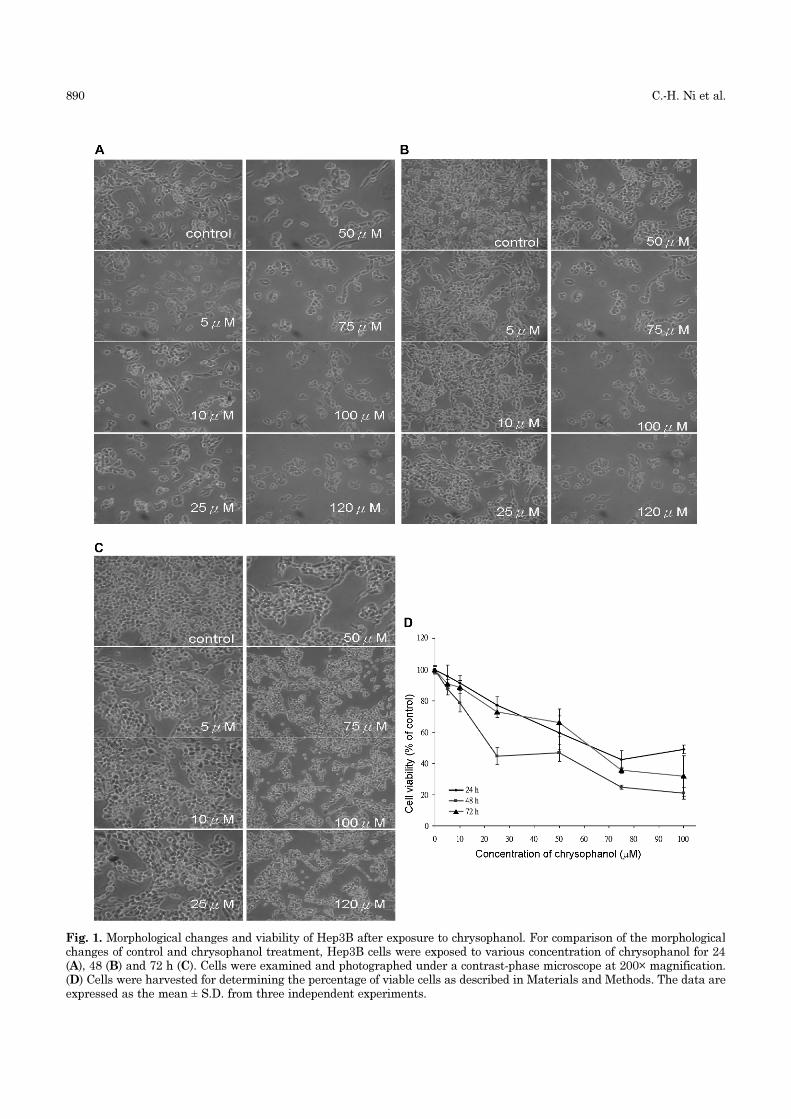
and decreased viability of Hep3BResults shown in Fig. 1A, B, C and D indicated that
chrysophanol induced cell morphological changes (in-
cluding a decrease in cell density, cell rounding, and
floating cells) and cell death. Chrysophanol treatment
caused a dose- and time-dependent decrease in the
viability of Hep3B cells. The concentration of chryso-
phanol required to cause a 50% inhibition of cell via-
bility (IC50) for Hep3B cells was 72 µM at 24 h, 50 µM
at 48 h and 68 µM at 72 h.
Chrysophanol-induced S phase arrest and
altered associated protein levelsIn order to determine whether chrysophanol could
block the cell cycle progression of Hep3B, the cell cycle
distribution of chrysophanol-treated and untreated
cells was evaluated by flow cytometric analysis and
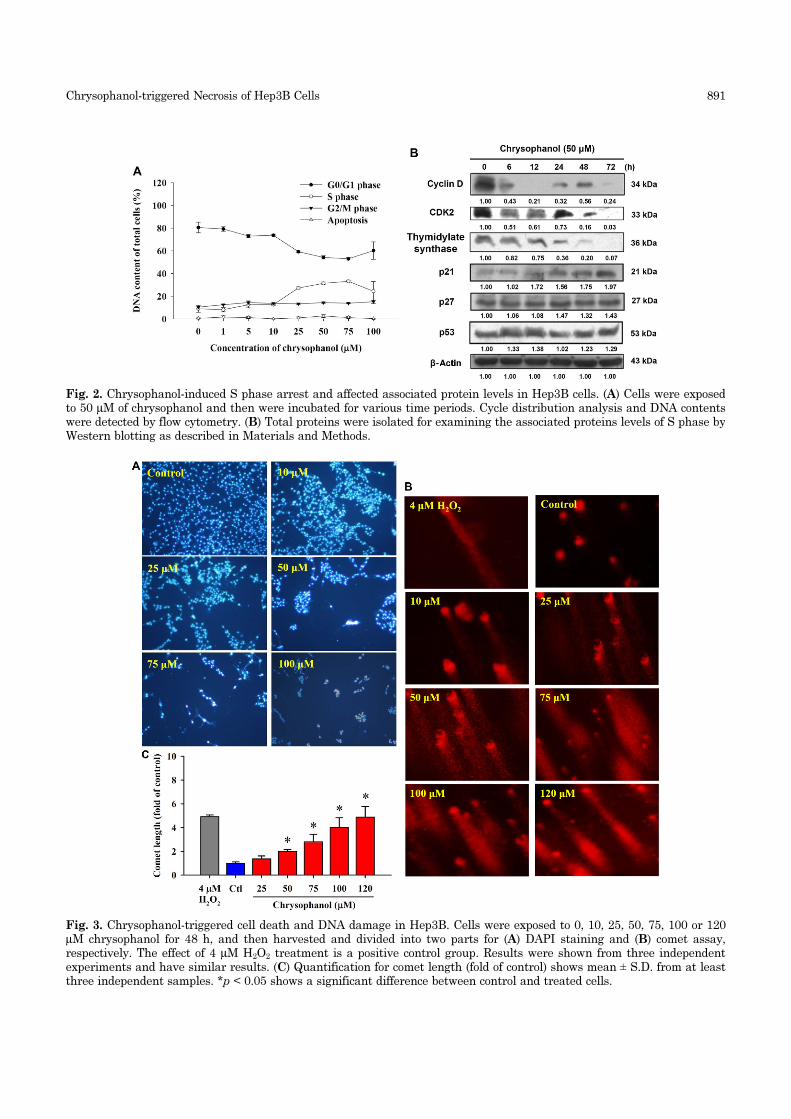
the results are shown in Fig. 2A. Chrysophanol treat-
ment induced cell cycle arrest at S phase in Hep3B
cells, and the percentage of Hep3B cells in the S phase
increased from 8% in the controls to 31% at 25 µM
and 37% at 50 µM, with a concurrent decline in the
G1 phase, in a doserelated fashion (Fig. 2A). To explore
the mechanisms underlying chrysophanol-induced S
phase arrest, the levels of S phase regulatory proteins
in Hep3B cells after 50 µM of chrysophanol treatment
for 0, 6, 12, 24 and 48 h were evaluated by Western
blotting. The results showed that chrysophanol treat-
ment resulted in substantial reductions in the levels
of cyclin D, CDK2 and thymidylate synthase proteins,
but increased the levels of p21 and p53 in Hep3B cells
(Fig. 2B).
Chrysophanol-induced DNA damageCells were isolated after chrysophanol treatment to
determine the DNA damage by DAPI staining and
comet assay, respectively. Fig. 3A indicates that chry-
sophanol treatment increases cell numbers with DNA
strand breaks (apoptotic morphological changes) in a
dose-related manner (Fig. 3A). Comet assay also
showed that chrysophanol induced DNA damage in
Hep3B cells in a dose-dependent manner (Fig. 3B).
Chrysophanol increases ROS level, Ca2+ pro-
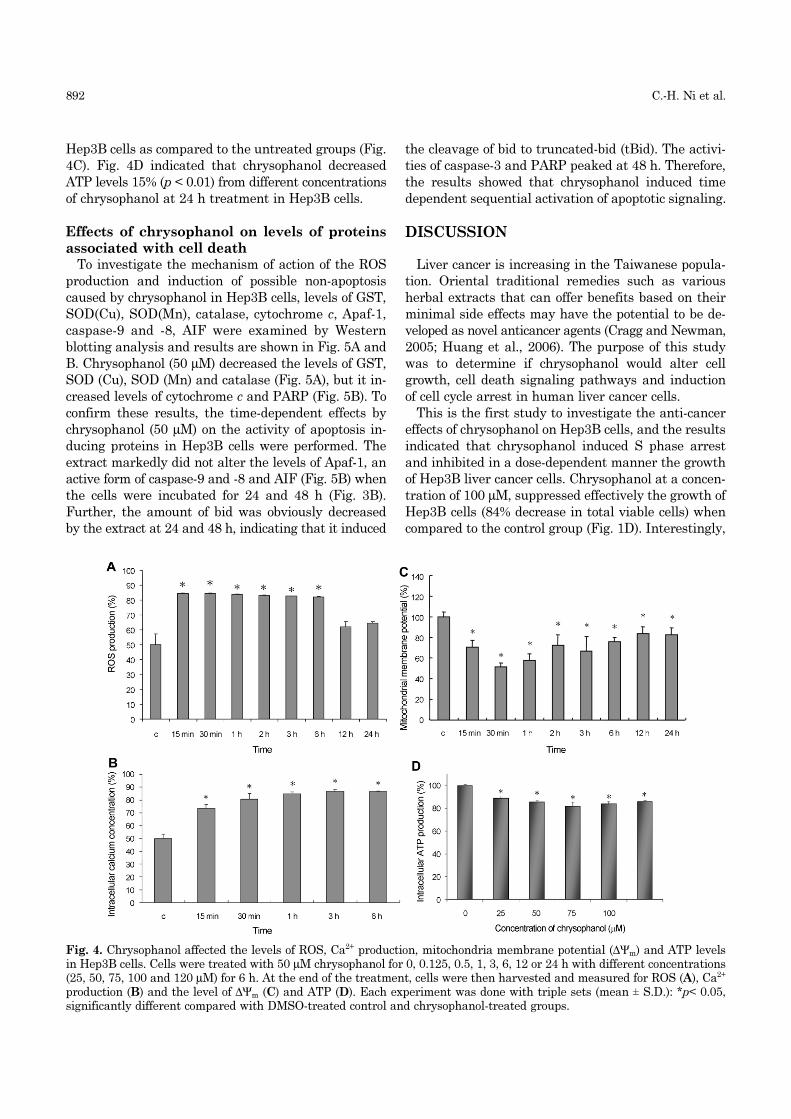
duction, and reduces ∆Ψm and ATP levelsResults indicated that a significant increase in
intracellular ROS (Fig. 4A) and cytosolic Ca2+ levels
(Fig. 4B), and a significant decrease in ∆Ψm level (Fig.
4C) and intracellular ATP levels were observed in
chrysophanol-treated cells when compared to the con-
trol cells. At the earlier treatment of chrysophanol
(0.125-24 h treatment), Hep3B cells had an initially
significantly increased ROS (Fig. 4A) and cytosolic
Ca2+ level (Fig. 4B) and significantly decreased the
levels of ∆Ψm (Fig. 4C) in 0.125-12 h treatment in
890 C.-H. Ni et al.
Fig. 1. Morphological changes and viability of Hep3B after exposure to chrysophanol. For comparison of the morphologicalchanges of control and chrysophanol treatment, Hep3B cells were exposed to various concentration of chrysophanol for 24(A), 48 (B) and 72 h (C). Cells were examined and photographed under a contrast-phase microscope at 200× magnification.(D) Cells were harvested for determining the percentage of viable cells as described in Materials and Methods. The data areexpressed as the mean ± S.D. from three independent experiments.
Chrysophanol-triggered Necrosis of Hep3B Cells 891
Fig. 2. Chrysophanol-induced S phase arrest and affected associated protein levels in Hep3B cells. (A) Cells were exposedto 50 µM of chrysophanol and then were incubated for various time periods. Cycle distribution analysis and DNA contentswere detected by flow cytometry. (B) Total proteins were isolated for examining the associated proteins levels of S phase byWestern blotting as described in Materials and Methods.
Fig. 3. Chrysophanol-triggered cell death and DNA damage in Hep3B. Cells were exposed to 0, 10, 25, 50, 75, 100 or 120µM chrysophanol for 48 h, and then harvested and divided into two parts for (A) DAPI staining and (B) comet assay,respectively. The effect of 4 µM H2O2 treatment is a positive control group. Results were shown from three independentexperiments and have similar results. (C) Quantification for comet length (fold of control) shows mean ± S.D. from at leastthree independent samples. *p < 0.05 shows a significant difference between control and treated cells.
892 C.-H. Ni et al.
Hep3B cells as compared to the untreated groups (Fig.
4C). Fig. 4D indicated that chrysophanol decreased
ATP levels 15% (p < 0.01) from different concentrations
of chrysophanol at 24 h treatment in Hep3B cells.
Effects of chrysophanol on levels of proteins
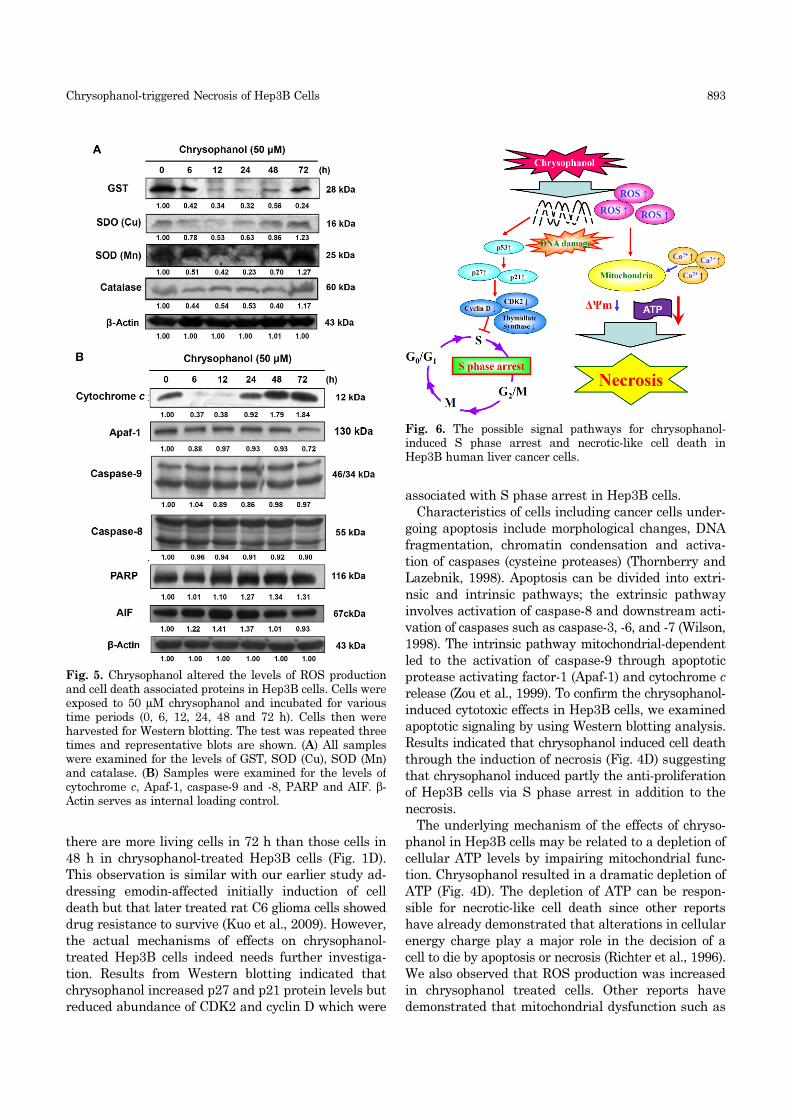
associated with cell deathTo investigate the mechanism of action of the ROS
production and induction of possible non-apoptosis
caused by chrysophanol in Hep3B cells, levels of GST,
SOD(Cu), SOD(Mn), catalase, cytochrome c, Apaf-1,
caspase-9 and -8, AIF were examined by Western
blotting analysis and results are shown in Fig. 5A and
B. Chrysophanol (50 µM) decreased the levels of GST,
SOD (Cu), SOD (Mn) and catalase (Fig. 5A), but it in-
creased levels of cytochrome c and PARP (Fig. 5B). To
confirm these results, the time-dependent effects by
chrysophanol (50 µM) on the activity of apoptosis in-
ducing proteins in Hep3B cells were performed. The
extract markedly did not alter the levels of Apaf-1, an
active form of caspase-9 and -8 and AIF (Fig. 5B) when
the cells were incubated for 24 and 48 h (Fig. 3B).
Further, the amount of bid was obviously decreased
by the extract at 24 and 48 h, indicating that it induced
the cleavage of bid to truncated-bid (tBid). The activi-
ties of caspase-3 and PARP peaked at 48 h. Therefore,
the results showed that chrysophanol induced time
dependent sequential activation of apoptotic signaling.
DISCUSSION
Liver cancer is increasing in the Taiwanese popula-
tion. Oriental traditional remedies such as various
herbal extracts that can offer benefits based on their
minimal side effects may have the potential to be de-
veloped as novel anticancer agents (Cragg and Newman,
2005; Huang et al., 2006). The purpose of this study
was to determine if chrysophanol would alter cell
growth, cell death signaling pathways and induction
of cell cycle arrest in human liver cancer cells.
This is the first study to investigate the anti-cancer
effects of chrysophanol on Hep3B cells, and the results
indicated that chrysophanol induced S phase arrest
and inhibited in a dose-dependent manner the growth
of Hep3B liver cancer cells. Chrysophanol at a concen-
tration of 100 µM, suppressed effectively the growth of
Hep3B cells (84% decrease in total viable cells) when
compared to the control group (Fig. 1D). Interestingly,
Fig. 4. Chrysophanol affected the levels of ROS, Ca2+ production, mitochondria membrane potential (∆Ψm) and ATP levelsin Hep3B cells. Cells were treated with 50 µM chrysophanol for 0, 0.125, 0.5, 1, 3, 6, 12 or 24 h with different concentrations(25, 50, 75, 100 and 120 µM) for 6 h. At the end of the treatment, cells were then harvested and measured for ROS (A), Ca2+
production (B) and the level of ∆Ψm (C) and ATP (D). Each experiment was done with triple sets (mean ± S.D.): *p< 0.05,significantly different compared with DMSO-treated control and chrysophanol-treated groups.
Chrysophanol-triggered Necrosis of Hep3B Cells 893
there are more living cells in 72 h than those cells in
48 h in chrysophanol-treated Hep3B cells (Fig. 1D).
This observation is similar with our earlier study ad-
dressing emodin-affected initially induction of cell
death but that later treated rat C6 glioma cells showed
drug resistance to survive (Kuo et al., 2009). However,
the actual mechanisms of effects on chrysophanol-
treated Hep3B cells indeed needs further investiga-
tion. Results from Western blotting indicated that
chrysophanol increased p27 and p21 protein levels but
reduced abundance of CDK2 and cyclin D which were
associated with S phase arrest in Hep3B cells.
Characteristics of cells including cancer cells under-
going apoptosis include morphological changes, DNA
fragmentation, chromatin condensation and activa-
tion of caspases (cysteine proteases) (Thornberry and
Lazebnik, 1998). Apoptosis can be divided into extri-
nsic and intrinsic pathways; the extrinsic pathway
involves activation of caspase-8 and downstream acti-
vation of caspases such as caspase-3, -6, and -7 (Wilson,
1998). The intrinsic pathway mitochondrial-dependent
led to the activation of caspase-9 through apoptotic
protease activating factor-1 (Apaf-1) and cytochrome c
release (Zou et al., 1999). To confirm the chrysophanol-
induced cytotoxic effects in Hep3B cells, we examined
apoptotic signaling by using Western blotting analysis.
Results indicated that chrysophanol induced cell death
through the induction of necrosis (Fig. 4D) suggesting
that chrysophanol induced partly the anti-proliferation
of Hep3B cells via S phase arrest in addition to the
necrosis.
The underlying mechanism of the effects of chryso-
phanol in Hep3B cells may be related to a depletion of
cellular ATP levels by impairing mitochondrial func-
tion. Chrysophanol resulted in a dramatic depletion of
ATP (Fig. 4D). The depletion of ATP can be respon-
sible for necrotic-like cell death since other reports
have already demonstrated that alterations in cellular
energy charge play a major role in the decision of a
cell to die by apoptosis or necrosis (Richter et al., 1996).
We also observed that ROS production was increased
in chrysophanol treated cells. Other reports have
demonstrated that mitochondrial dysfunction such as
Fig. 5. Chrysophanol altered the levels of ROS productionand cell death associated proteins in Hep3B cells. Cells wereexposed to 50 µM chrysophanol and incubated for varioustime periods (0, 6, 12, 24, 48 and 72 h). Cells then wereharvested for Western blotting. The test was repeated threetimes and representative blots are shown. (A) All sampleswere examined for the levels of GST, SOD (Cu), SOD (Mn)and catalase. (B) Samples were examined for the levels ofcytochrome c, Apaf-1, caspase-9 and -8, PARP and AIF. β-Actin serves as internal loading control.
Fig. 6. The possible signal pathways for chrysophanol-induced S phase arrest and necrotic-like cell death inHep3B human liver cancer cells.

894 C.-H. Ni et al.
ROS overproduction, intracellular [Ca2+] elevation,
and ATP reduction can damage mitochondrial dynamics
(Irrcher et al., 2003; Pletjushkina et al., 2006; Szabadkai
et al., 2006).
In conclusion, chrysophanol induced indirect changes
in the mitochondrial outer membrane permeability
then caused the release of cytochrome c and AIF from
mitochondria (Fig. 5B) and apart from uncoupling
which may add to an impairment of ATP synthesis for
leading to the decrease of ATP levels (Fig. 4D) and
finally led to necrosis in Hep3B cells.
ACKNOWLEDGEMENTS
This work was supported by a research grant
CMU100-NSC-01 from China Medical University,
Taichung, Taiwan.
REFERENCES
Aravindaram, K. and Yang, N. S., Anti-inflammatory plant
natural products for cancer therapy. Planta Med., 76,
1103-1117 (2010).
Aviello, G., Rowland, I., Gill, C. I., Acquaviva, A. M., Capasso,
F., McCann, M., Capasso, R., Izzo, A. A., and Borrelli, F.,
Anti-proliferative effect of rhein, an anthraquinone isolat-
ed from Cassia species, on Caco-2 human adenocarcinoma
cells. J. Cell. Mol. Med., 14, 2006-2014 (2010).
Chen, Y. L., Lu, H. F., Hung, F. M., Huang, A. C., Hsueh, S.
C., Liu, C. M., Yang, J. S., Yu, C. C., Chiang, J. H., Lu, C.
C., Chiu, T. H., and Chung, J. G., Danthron inhibits
murine WEHI-3 cells in vivo, and enhances macrophage
phagocytosis and natural killer cell cytotoxic activity in
leukemic mice. In Vivo, 25, 393-398 (2011).
Chiang, J. H., Yang, J. S., Ma, C. Y., Yang, M. D., Huang, H.
Y., Hsia, T. C., Kuo, H. M., Wu, P. P., Lee, T. H., and Chung,
J. G., Danthron, an anthraquinone derivative, induces
DNA damage and caspase cascades-mediated apoptosis in
SNU-1 human gastric cancer cells through mitochondrial
permeability transition pores and Bax-triggered path-
ways. Chem. Res. Toxicol., 24, 20-29 (2011).
Chiang, L. C., Ng, L. T., Lin, I. C., Kuo, P. L., and Lin, C. C.,
Anti-proliferative effect of apigenin and its apoptotic
induction in human Hep G2 cells. Cancer Lett., 237, 207-
214 (2006).
Cragg, G. M. and Newman, D. J., Plants as a source of anti-
cancer agents. J. Ethnopharmacol., 100, 72-79 (2005).
Huan, S. K., Lee, H. H., Liu, D. Z., Wu, C. C., and Wang, C.
C., Cantharidin-induced cytotoxicity and cyclooxygenase
2 expression in human bladder carcinoma cell line.
Toxicology, 223, 136-143 (2006).
Huang, Q., Lu, G., Shen, H. M., Chung, M. C., and Ong, C.
N., Anti-cancer properties of anthraquinones from
rhubarb. Med. Res. Rev., 27, 609-630 (2007).
Huang, Y. T., Huang, D. M., Chueh, S. C., Teng, C. M., and
Guh, J. H., Alisol B acetate, a triterpene from Alismatis
rhizoma, induces Bax nuclear translocation and apoptosis
in human hormone-resistant prostate cancer PC-3 cells.
Cancer Lett., 231, 270-278 (2006).
Irrcher, I., Adhihetty, P. J., Joseph, A. M., Ljubicic, V., and
Hood, D. A., Regulation of mitochondrial biogenesis in
muscle by endurance exercise. Sports Med., 33, 783-793
(2003).
Ji, B. C., Hsu, W. H., Yang, J. S., Hsia, T. C., Lu, C. C.,
Chiang, J. H., Yang, J. L., Lin, C. H., Lin, J. J., Suen, L. J.,
Gibson Wood, W., and Chung, J. G., Gallic acid induces
apoptosis via caspase-3 and mitochondrion-dependent
pathways in vitro and suppresses lung xenograft tumor
growth in vivo. J. Agric. Food Chem., 57, 7596-7604 (2009).
Kang, S. C., Lee, C. M., Choung, E. S., Bak, J. P., Bae, J. J.,
Yoo, H. S., Kwak, J. H., and Zee, O. P., Anti-proliferative
effects of estrogen receptor-modulating compounds isolated
from Rheum palmatum. Arch. Pharm. Res., 31, 722-726
(2008).
Kuo, T. C., Yang, J. S., Lin, M. W., Hsu, S. C., Lin, J. J., Lin,
H. J., Hsia, T. C., Liao, C. L., Yang, M. D., Fan, M. J., Wood,
W. G., and Chung, J. G., Emodin has cytotoxic and pro-
tective effects in rat C6 glioma cells: roles of Mdr1a and
nuclear factor kappaB in cell survival. J. Pharmacol. Exp.
Ther., 330, 736-744 (2009).
Lin, C. C., Yang, J. S., Chen, J. T., Fan, S., Yu, F. S., Yang,
J. L., Lu, C. C., Kao, M. C., Huang, A. C., Lu, H. F., and
Chung, J. G., Berberine induces apoptosis in human HSC-
3 oral cancer cells via simultaneous activation of the death
receptor-mediated and mitochondrial pathway. Anticancer
Res., 27, 3371-3378 (2007).
Lin, M. L., Lu, Y. C., Chung, J. G., Li, Y. C., Wang, S. G., NG,
S. H., Wu, C. Y., Su, H. L., and Chen, S. S., Aloe-emodin
induces apoptosis of human nasopharyngeal carcinoma
cells via caspase-8-mediated activation of the
mitochondrial death pathway. Cancer Lett., 291, 46-58
(2010).
Lin, S. Y., Lai, W. W., Ho, C. C., Yu, F. S., Chen, G. W., Yang,
J. S., Liu, K. C., Lin, M. L., Wu, P. P., Fan, M. J., and Chung,
J. G., Emodin induces apoptosis of human tongue squamous
cancer SCC-4 cells through reactive oxygen species and
mitochondria-dependent pathways. Anticancer Res., 29,
327-335 (2009).
Liu, K. C., Huang, A. C., Wu, P. P., Lin, H. Y., Chueh, F. S.,
Yang, J. S., Lu, C. C., Chiang, J. H., Meng, M., and Chung,
J. G., Gallic acid suppresses the migration and invasion of
PC-3 human prostate cancer cells via inhibition of matrix
metalloproteinase-2 and -9 signaling pathways. Oncol.
Rep., 26, 177-184 (2011).
Lu, C. C., Yang, J. S., Huang, A. C., Hsia, T. C., Chou, S. T.,
Kuo, C. L., Lu, H. F., Lee, T. H., Wood, W. G., and Chung,
J. G., Chrysophanol induces necrosis through the produc-
tion of ROS and alteration of ATP levels in J5 human liver
cancer cells. Mol. Nutr. Food Res., 54, 967-976 (2010a).
Lu, H. F., Chen, Y. S., Yang, J. S., Chen, J. C., Lu, K. W.,
Chiu, T. H., Liu, K. C., Yeh, C. C., Chen, G. W., Lin, H. J.,

Chrysophanol-triggered Necrosis of Hep3B Cells 895
and Chung, J. G., Gypenosides induced G0/G1 arrest via
inhibition of cyclin E and induction of apoptosis via
activation of caspases-3 and -9 in human lung cancer A-
549 cells. In Vivo, 22, 215-221 (2008).
Lu, H. F., Chen, Y. L., Yang, J. S., Yang, Y. Y., Liu, J. Y., Hsu,
S. C., Lai, K. C., and Chung, J. G., Antitumor activity of
capsaicin on human colon cancer cells in vitro and colo 205
tumor xenografts in vivo. J. Agric. Food Chem., 58, 12999-
13005 (2010b).
Pletjushkina, O. Y., Lyamzaev, K. G., Popova, E. N.,
Nepryakhina, O. K., Ivanova, O. Y., Domnina, L. V.,
Chernyak, B. V., and Skulachev, V. P., Effect of oxidative
stress on dynamics of mitochondrial reticulum. Biochim.
Biophys. Acta, 1757, 518-524 (2006).
Richter, C., Schweizer, M., Cossarizza, A., and Franceschi,
C., Control of apoptosis by the cellular ATP level. FEBS
Lett., 378, 107-110 (1996).
Su, Y. T., Chang, H. L., Shyue, S. K., and Hsu, S. L., Emodin
induces apoptosis in human lung adenocarcinoma cells
through a reactive oxygen species-dependent mitochon-
drial signaling pathway. Biochem. Pharmacol., 70, 229-
241 (2005).
Szabadkai, G., Simoni, A. M., Bianchi, K., De Stefani, D.,
Leo, S., Wieckowski, M. R., and Rizzuto, R., Mitochondrial
dynamics and Ca2+ signaling. Biochim. Biophys. Acta,
1763, 442-449 (2006).
Thornberry, N. A. and Lazebnik, Y., Caspases: enemies
within. Science, 281, 1312-1316 (1998).
Ueno, Y., Umemori, K., Niimi, E., Tanuma, S., Nagata, S.,
Sugamata, M., Ihara, T., Sekijima, M., Kawai, K., Ueno,
I., and Tashiro, F., Induction of apoptosis by T-2 toxin and
other natural toxins in HL-60 human promyelotic leukemia
cells. Nat. Toxins, 3, 129-137 (1995).
Verma, S. C., Singh, N. P., and Sinha, A. K., Determination
and locational variations in the quantity of hydroxyan-
thraquinones and their glycosides in rhizomes of Rheum
emodi using high-performance liquid chromatography. J.
Chromatogr. A, 1097, 59-65 (2005).
Wilson, M. R., Apoptotic signal transduction: emerging path-
ways. Biochem. Cell Biol., 76, 573-582 (1998).
Yu, F. S., Yang, J. S., Yu, C. S., Lu, C. C., Chiang, J. H., Lin, C.
W., and Chung, J. G., Safrole induces apoptosis in human
oral cancer HSC-3 cells. J. Dent. Res., 90, 168-174 (2011).
Zou, H., Li, Y., Liu, X., and Wang, X., An APAF-1.cytochrome
c multimeric complex is a functional apoptosome that
activates procaspase-9. J. Biol. Chem., 274, 11549-11556
(1999).
Related Documents











