Chronic Alcohol Consumption, Abstinence and Relapse: Brain Proton Magnetic Resonance Spectroscopy Studies in Animals and Humans Dieter J. Meyerhoff, Timothy C. Durazzo and Gabriele Ende Abstract This chapter summarizes the peer-reviewed literature of proton magnetic resonance spectroscopy ( 1 H MRS) studies on the effects of chronic and excessive alcohol consumption in both the animal and human brain. After a brief summary of the neuropathology of alcohol use disorders (AUD), we describe the primary brain metabolites measured by in vivo 1 H MRS. We then focus on published MRS studies of animal models of alcohol dependence and of treatment-seeking humans with AUD. We also summarize the scant MRS research on the much larger fraction of treatment-naïve individuals with AUD and the similarities and discrepancies relative to treatment-seekers. It is exceedingly apparent that premorbid and/or comorbid dis- orders/conditions, especially chronic smoking, among individuals with AUD con- tribute to the considerable variability in the pattern and magnitude of neurobio- logical and neurocognitive abnormalities in AUD. Therefore, we also review studies on the neurobiological consequences of the combined effects of chronic drinking and smoking in AUD. Finally, as AUD is characterized by a chronically relapsing/remitting course over lifetime and identification of those at greatest risk for relapse is important, we review 1 H MRS studies on brain spectroscopic mea- sures that contribute to the prediction of relapse in AUD. We conclude with an D. J. Meyerhoff (&) Á T. C. Durazzo Center for Imaging of Neurodegenerative Diseases, San Francisco Veterans Affairs Medical Center, San Francisco, CA, USA e-mail: [email protected] D. J. Meyerhoff Á T. C. Durazzo Department of Radiology and Biomedical Imaging, University of California San Francisco, San Francisco, CA, USA G. Ende Department of Neuroimaging, Central Institute of Mental Health, Mannheim, Germany Curr Topics Behav Neurosci DOI: 10.1007/7854_2011_131 Ó Springer-Verlag Berlin Heidelberg 2011

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Chronic Alcohol Consumption,Abstinence and Relapse: Brain ProtonMagnetic Resonance Spectroscopy Studiesin Animals and Humans
Dieter J. Meyerhoff, Timothy C. Durazzo and Gabriele Ende
Abstract This chapter summarizes the peer-reviewed literature of proton magneticresonance spectroscopy (1H MRS) studies on the effects of chronic and excessivealcohol consumption in both the animal and human brain. After a brief summary ofthe neuropathology of alcohol use disorders (AUD), we describe the primary brainmetabolites measured by in vivo 1H MRS. We then focus on published MRS studiesof animal models of alcohol dependence and of treatment-seeking humans withAUD. We also summarize the scant MRS research on the much larger fraction oftreatment-naïve individuals with AUD and the similarities and discrepancies relative totreatment-seekers. It is exceedingly apparent that premorbid and/or comorbid dis-orders/conditions, especially chronic smoking, among individuals with AUD con-tribute to the considerable variability in the pattern and magnitude of neurobio-logical and neurocognitive abnormalities in AUD. Therefore, we also reviewstudies on the neurobiological consequences of the combined effects of chronicdrinking and smoking in AUD. Finally, as AUD is characterized by a chronicallyrelapsing/remitting course over lifetime and identification of those at greatest riskfor relapse is important, we review 1H MRS studies on brain spectroscopic mea-sures that contribute to the prediction of relapse in AUD. We conclude with an
D. J. Meyerhoff (&) � T. C. DurazzoCenter for Imaging of Neurodegenerative Diseases,San Francisco Veterans Affairs Medical Center,San Francisco, CA, USAe-mail: [email protected]
D. J. Meyerhoff � T. C. DurazzoDepartment of Radiology and Biomedical Imaging,University of California San Francisco,San Francisco, CA, USA
G. EndeDepartment of Neuroimaging, Central Institute of Mental Health,Mannheim, Germany
Curr Topics Behav NeurosciDOI: 10.1007/7854_2011_131� Springer-Verlag Berlin Heidelberg 2011

overall assessment of the MRS research literature on brain alcohol effects, the roleof animal and human studies in understanding the disease, and discuss the need ofwidely integrative MRS studies of cohorts that include individuals with comorbi-dies that are reflective of the general population with AUD.
Keywords Alcoholism �Alcohol use disorders �Magnetic resonance spectroscopy �Neuroimaging � Brain metabolites � Recovery � Relapse � Nicotine � Smoking
AbbreviationsAUD Alcohol use disordersCNS Central nervous system.Cho Choline-containing compoundsCr Creatine and phosphocreatineCSF Cerebrospinal fluidGABA Gamma aminobutyric acid,Glu GlutamateGM Gray matterNAA N-acetylaspartateNAAG N-acetylaspartylglutamatemI MyoinositolMR Magnetic resonanceMRI Magnetic resonance imagingWM White matter1H MRS Proton magnetic resonance spectroscopy.1H MRSI Proton magnetic resonance spectroscopic imaging
Contents
1 Introduction..............................................................................................................................2 Neuropathology of AUD.........................................................................................................3 Proton MRS Methods and Commonly Measured Brain Metabolites....................................4 The Neurobiological Consequences of AUD and Changes During Abstinence from Alcohol5 Treatment-Seeking Versus Treatment Naïve Alcoholics and Common Comorbidities .......6 The Neurobiological Correlates of Relapse in Alcohol Use Disorders (AUD)....................7 Conclusions and Perspectives .................................................................................................References......................................................................................................................................
1 Introduction
This chapter summarizes the peer-reviewed literature on proton magnetic reso-nance spectroscopy (MRS) studies on the effects of chronic and excessivealcohol consumption in both the animal and human brain. After a brief summary
D. J. Meyerhoff et al.

of the neuropathology associated with alcohol use disorders (i.e., alcohol abuseand dependence), we describe the primary metabolites measured by in vivoproton MRS. We then focus on published MRS studies of animal models ofalcohol dependence and of humans with alcohol use disorders (AUD). Thehuman studies focus on individuals with uncomplicated AUD, i.e., without ahistory of Wernike-Korsakoff Syndrome or encephalopathy secondary to chronichepatic disease. The neuropathological, neuroimaging and neurocognitivecorollaries of Wernike-Korsakoff Syndrome and alcohol-induced hepaticencephalopathy have been reviewed elsewhere (Behar et al. 1999; Harper et al.2003; Hazell and Butterworth 1999; Oscar-Berman 2000; Sullivan 2000;Thomson and Marshall 2006).
It is estimated that only approximately 10% of individuals with AUD seektreatment at some point in their lives (Fein and Landman 2005; Moss et al.2007). However, most of what is known about the effects of AUD on the humanbrain is derived from studies with individuals recruited from inpatient and out-patient treatment programs (Fein et al. 2002), only a small fraction of whomparticipate in research studies. Given the possible bias from deriving potentiallyclinically useful data from a very small minority of affected individuals, we alsosummarize the scant MRS research on the much larger fraction of treatment-naïve individuals with AUD and the similarities and discrepancies relative totreatment-seekers.
In this context, it is widely recognized from large United States-basedepidemiological studies, such as the National Epidemiologic Survey on Alcoholand Related Conditions (NESARC), that a number of premorbid and/or comorbiddisorders/conditions are associated with AUD (Hasin et al. 2007; Mansell et al.2006; Mertens et al. 2003; Stinson et al. 2005). These together may promoteconsiderable variability in the pattern and magnitude of neurobiological andneurocognitive abnormalities demonstrated in AUD. Since chronic cigarettesmoking is the most common comorbidity (Daeppen et al. 2000; John et al. 2003;Room 2004), we also review studies on the neurobiological consequences of thecombined effects of chronic drinking and smoking.
Finally, AUD is characterized by a chronically relapsing/remitting course overlifetime (Dawson et al. 2007; Maisto and Connors 2006; Zywiak et al. 2006). Asubstantial amount of research has investigated the psychological, psychiatric,sociodemographic and behavioral correlates of relapse following treatment;however, the neurobiological factors contributing to relapse in humans have onlyrecently begun to be delineated, largely due to advances in in vivo neuroimagingtechniques (Durazzo et al. 2010b). We briefly summarize the applicability ofproton MRS to the prediction of relapse in AUD. We conclude with an overallassessment of the MRS research literature on brain alcohol effects, the role ofanimal and human studies in understanding the disease, and discuss the need ofMRS studies of cohorts that include individuals with comorbidities that arereflective of the general population of those with an AUD.
Chronic Alcohol Consumption, Abstinence and Relapse

2 Neuropathology of AUD
Postmortem examinations of individuals with uncomplicated AUD indicateneuronal loss primarily in the dorsolateral frontal cortex, hypothalamus and thecerebellum, with the hippocampi showing glial rather than neuronal loss (Hardinget al. 1997; Korbo 1999; Kril and Halliday 1999; Kril et al. 1996). Reduced glialcell density and size in the dorsolateral prefrontal cortex (Miguel-Hidalgo et al.2002) and lower neuronal and glial cell density in the orbitofrontal cortex havealso been reported (Miguel-Hidalgo et al. 2006). However, other investigatorsfound no abnormalities in neocortical neuronal cell volumes, neuronal and glialcell numbers or lobar and global neocortical surface area, thickness and volume inpostmortem studies of AUD (Fabricius et al. 2007; Jensen and Pakkenberg 1993).White matter (WM) loss in anterior brain regions has also been reported, whichmay involve disturbances in both myelin and axonal integrity (Harper 2009).According to a general model by Harper and Kril (1989), alcohol-related corticalbrain damage either constitutes loss of dendritic arbor and shrinkage of neuronalcell body volume or neuronal death and Wallerian degeneration of myelinatedaxons (e.g., Schwab and Bartholdi 1996) and occurs primarily in the frontal lobe,particularly in the superior frontal cortex (Harper 2009). With abstinence fromalcohol, dendritic arbor and neuronal cell body volume increases have beenreported, as have changes in myelin structure and increases in tissue density,particularly in the neocortical and subcortical gray matter (e.g., Dlugos andPentney 1997; Sullivan and Pfefferbaum 2005). This suggests that the brain doesrecover from alcohol-induced brain injury with extended sobriety.
The mechanisms of chronic alcohol-associated brain injury and neurocognitivedysfunction are hypothesized to involve glutamate and homocysteine-inducedexcitotoxicity, reduced levels of brain derived neurotrophic factors, increasedoxidative stress and free radical levels, thiamine and other nutritional deficiencies,increased acetaldehyde and aldehydes levels, hepatic dysfunction and geneticvulnerability (for review see Durazzo and Meyerhoff 2007). Excitotoxicity hasbeen suggested to be most prominent during withdrawal from alcohol (De Witte2004; Harris et al. 2003; Prendergast et al. 2000). In human AUD, reports ofassociations between level of alcohol consumption and structural, metabolic andfunctional brain injury are inconsistent (Durazzo and Meyerhoff 2007).
More recently, however, rodent studies have suggested that mechanisms ofalcohol-related brain injury involve oxidative stress secondary to proinflammatoryenzymes that are operative during intoxication rather than withdrawal (Crews andNixon 2009). In a rodent model, Crews et al. (2004) have shown that binge ethanoladministration induces brain damage demonstrated by agyrophilic silver staining.Brain damage increases progressively after 2 days beginning in the olfactory bulbfollowed by additional brain regions showing increasing damage with furtherbinge exposure. The rodent binge model shares both neurodegenerative and cog-nitive deficits found in the human alcoholic brain (Crews and Nixon 2009). Pro-longed alcohol dependence in rats has also been linked to long-term suppression of
D. J. Meyerhoff et al.

forebrain neurogenesis and loss of neuronal progenitor cells (Hansson et al. 2010).In studies of moderate alcohol consumption, alcohol-preferring rats were given thechoice of drinking 10% alcohol in water or pure water for 7 weeks (He et al.2009). During the subsequent abstinence period, hippocampal neurogenesisincreased as did differentiation of oligodendrocyte progenitors in the cingulate andproliferation of undifferentiated cells in the substantia nigra. Such cellularalterations associated with alcohol dependence and abstinence may contribute toalcohol-induced cortical dysfunction and neurocognitive deficits as well as to theirrecovery during prolonged sobriety.
All of the potential mechanisms mentioned above may work independently ortogether in AUD to alter cerebral cellular structures or organelles, membranephospholipids, myelin, DNA, gene expression, protein synthesis and cellularmetabolism.
3 Proton MRS Methods and Commonly Measured BrainMetabolites
Magnetic resonance spectroscopy (MRS) enables the non-invasive and concurrentquantitation of several metabolites from most brain regions. Proton MRS (1HMRS) enables the assessment of neurophysiological consequences of a disease/condition that may precede any associated gross morphological changes. Moststudies have been performed with single-volume approaches that acquire MRspectra from one or a few volumes of interest within the brain, with typicalvolumes between 4 and 16 cm3. The more technically and analytically demandingtwo- and three-dimensional MR spectroscopic imaging (MRSI, also calledchemical shift imaging or CSI) approaches use a combination of phase encodingand spectroscopy to acquire simultaneously MR spectra from up to hundreds ofvolumes from throughout most of the brain, with typical volumes of 0.8–2 cm3.The latter approach also allows reconstructing metabolite images, which displaythe distribution of metabolites throughout the imaged region. Most of the MRS-and MRSI-detectable brain proton metabolites addressed below are associatedwith neurocognition in normal aging, substance/alcohol use disorders, neurode-generative diseases, psychiatric conditions and traumatic brain injury (Babikianet al. 2006; Durazzo and Meyerhoff 2007; Martinez-Bisbal et al. 2006; Ohrmannet al. 2008; Ross and Sachdev 2004; Schuff et al. 2006; Yildiz-Yesiloglu andAnkerst 2006a, b; Zahr et al. 2008). These relationships demonstrate the functionalrelevance and clinical significance of MRS measurements (Steen et al. 2005).
At a magnetic field strength of 1.5 Tesla (T), the following brain metabolites aremost frequently measured: N-acetylaspartate (NAA), choline-containing com-pounds (Cho), creatine-containing compounds (Cr) and myo-inositol (mI; withshort echo time pulse sequences). At higher magnetic field strengths (e.g., [2T),the general metabolic pool of glutamine, glutamate (Glu) and gamma aminobu-tyric acid (GABA) can be measured. Figure 1 depicts an exemplary 1H MR
Chronic Alcohol Consumption, Abstinence and Relapse
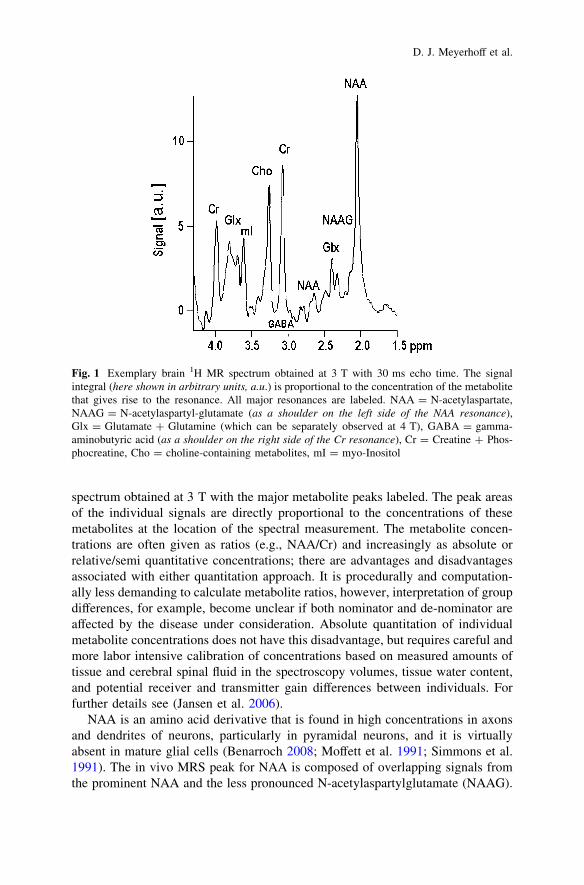
spectrum obtained at 3 T with the major metabolite peaks labeled. The peak areasof the individual signals are directly proportional to the concentrations of thesemetabolites at the location of the spectral measurement. The metabolite concen-trations are often given as ratios (e.g., NAA/Cr) and increasingly as absolute orrelative/semi quantitative concentrations; there are advantages and disadvantagesassociated with either quantitation approach. It is procedurally and computation-ally less demanding to calculate metabolite ratios, however, interpretation of groupdifferences, for example, become unclear if both nominator and de-nominator areaffected by the disease under consideration. Absolute quantitation of individualmetabolite concentrations does not have this disadvantage, but requires careful andmore labor intensive calibration of concentrations based on measured amounts oftissue and cerebral spinal fluid in the spectroscopy volumes, tissue water content,and potential receiver and transmitter gain differences between individuals. Forfurther details see (Jansen et al. 2006).
NAA is an amino acid derivative that is found in high concentrations in axonsand dendrites of neurons, particularly in pyramidal neurons, and it is virtuallyabsent in mature glial cells (Benarroch 2008; Moffett et al. 1991; Simmons et al.1991). The in vivo MRS peak for NAA is composed of overlapping signals fromthe prominent NAA and the less pronounced N-acetylaspartylglutamate (NAAG).
Fig. 1 Exemplary brain 1H MR spectrum obtained at 3 T with 30 ms echo time. The signalintegral (here shown in arbitrary units, a.u.) is proportional to the concentration of the metabolitethat gives rise to the resonance. All major resonances are labeled. NAA = N-acetylaspartate,NAAG = N-acetylaspartyl-glutamate (as a shoulder on the left side of the NAA resonance),Glx = Glutamate ? Glutamine (which can be separately observed at 4 T), GABA = gamma-aminobutyric acid (as a shoulder on the right side of the Cr resonance), Cr = Creatine ? Phos-phocreatine, Cho = choline-containing metabolites, mI = myo-Inositol
D. J. Meyerhoff et al.

NAA is synthesized in the neuronal mitochondria, from where it is exported to thecytosolic compartment. In the extracellular fluid it diffuses to oligodendrocytes,where it is rapidly hydrolyzed by amidohydrolase II (Baslow 2003). NAA acts asan organic osmolyte controlling cellular water distribution, which provides acritical source of acetate for myelin lipid synthesis in oligodendrocytes (myelin-ogenesis) and is involved in facilitating energy metabolism in neuronalmitochondria. NAA is also an immediate precursor for the enzyme-mediatedbiosynthesis of the neuronal dipeptide NAAG, which acts to regulate glutamateand dopamine release, most likely via activation of presynaptic mGluR2/3receptors. Abnormalities in NAA synthesis, transport and/or breakdown (i.e.,neuronal dysfunction) may contribute to an abnormal steady-state concentrationthat is measurable by 1H MRS/MRSI. The MRS signal from NAA is oftendescribed as a marker of neuronal viability or integrity (Baslow and Guilfoyle; DeStefano et al. 1995; Hugg et al. 1996; Schuff et al. 2001; Sullivan 2000; Vion-Duryet al. 1994). However, nominal NAA levels do not seem to be mandatory forneuronal viability or function because differentiated cultured neurons were stillviable in the absence of NAA and a progressive loss of NAA was detected inviable cultured organotypic brain slices (for review see Baslow 2003). Recentstudies in various pathologies have shown that NAA reduction can be at leastpartially reversible, which suggests that NAA does not necessarily reflect thedensity or concentration of neurons per se but is rather sensitive to the plasticity ofneuronal components and pathological processes affecting the metabolic func-tioning of neurons (Bertolino et al. 2003), particularly neuronal bioenergetics(Baslow and Guilfoyle 2007; Pan and Takahashi 2005). Additionally, postmortemstudies showed that NAA reduction is correlated with the overall volume ofneuronal soma size (Rajkowska et al. 1998; Selemon and Goldman-Rakic 1999).
NAA has been shown to be distributed homogeneously throughout the brain, at aconcentration of approximately 10 mmol/L, whereas NAAG increases rostral tocaudal (1.5–2.7 mmo/L) and exhibits higher concentrations in WM than in gray matter(GM) (Pouwels and Frahm 1998). Whether the NAA concentration is higher in GM orWM is a topic of continued debate and appears critically dependent on region andmethod. NAA tends to decrease globally throughout the brain with age (Maudsley et al.2009) and has been shown to decrease as a function of age in the temporal lobe(Riederer et al. 2007) and the medial prefrontal brain (Ende et al. 2000).
The MR detectable Cho represents the trimethyl ammonium resonance ofseveral choline-containing compounds. Most of the proton MRS resonance iscontributed from phosphocholine and glycerophosphocholine, with free cholinecontributing less than 5% and the neurotransmitter acetylcholine even less(Boulanger et al. 2000). The choline-containing compounds are intermediates inphospholipid (membrane) synthesis and breakdown, and it has been suggested thatdecreased phosphocholine and increased glycerophosphocholine levels (togetherwith other phosphoester alterations) reflect membrane breakdown (Pettegrew et al.1987, 1990). Phosphatidylcholine, the major choline-containing metabolite of thebrain and the main component of myelin, cell membranes, and other brain lipids isinvisible under normal MRS acquisition conditions, as it is restricted in its
Chronic Alcohol Consumption, Abstinence and Relapse

molecular mobility. However, in 1H MRS, (non membrane bound) choline-containing compounds cannot be distinguished—all are detected within one Chopeak. The compounds exhibit a marked regional variability with the highestconcentrations in the cerebellum and lowest levels and a strong rostral to caudaldecreasing gradient in GM (Pouwels and Frahm 1998). The Cho concentration isbetween 1.5 and 2.5 mmol/L and has been shown to increase with age (Maudsleyet al. 2009). A high Cho signal is thought to reflect increased cellular membraneturnover and density (Miller et al. 1996), myelin catabolism (Ross and Bluml2001) and/or inflammation (Brenner et al. 1993).
In its bioactive form, myo-inositol (mI) is a carbohydrate that structurallyresembles glucose. mI is a constituent of phosphatidylinositol, an importantcomponent of the phospholipid bilayer that constitutes all eukaryotic cell membranes.mI is synthesized predominantly by glia in the brain and is incapable of crossing theblood–brain barrier (Brand et al. 1993). The biological significance of mI has not yetbeen established with certainty. It has been suggested to be a glia-specific marker(Brand et al. 1993) and/or an osmolyte (Ross and Bluml 2001; Schweinsburg et al.2000) and to be involved in second messenger system functioning (Fisher et al.2002). mI elevations may reflect inflammation, astrocyte proliferation and/or anosmotic response to cell shrinkage (Rosen and Lenkinski 2007).
The MRS signal of Cr derives from creatine plus phosphocreatine. In normalbrain metabolism, phosphocreatine supplies a phosphate group to adenosinediphosphate (ADP), resulting in the production of adenosine triphosphate (ATP)and the release of creatine. Thus, total creatine should be a reliable marker of brainmetabolism, reflecting bioenergetics of neuronal and glial tissue (Ferguson et al.2002). Creatine, phosphocreatine, and their main precursor, guanidinoacetate, areprimarily synthesized in the liver and kidneys and then transported to the brain.The creatine concentration calculated from the Cr MRS signal is about 9 mmol/L.However, as Cr has an important buffer capacity in cellular energy metabolism, itsconcentration cannot be considered 100% stable; cerebral levels can vary acrossdifferent diseases or pathological states (Rosen and Lenkinski 2007; Ross andBluml 2001). Nevertheless, using the Cr signal from a spectrum as an ‘‘internalconcentration reference’’ when reporting metabolite levels from other resonancesof the same spectrum (e.g., NAA/Cr, Cho/Cr) is experimentally straightforward(and corrects for some experimental factors when comparing measures betweendifferent individuals); this simple approach has at least in early reports proven tobe of some diagnostic importance in many studies of various pathologies. Carefulstudies, however, have shown a brain activity-dependent change in Cr signalintensity (Ke et al. 2002), an age-dependent increase of Cr in WM (Maudsley et al.2009), and changes of absolute creatine concentrations in various pathologies, sothat Cr cannot be considered a useful concentration reference (Li et al. 2002; Rossand Michaelis 1994; Sartorius et al. 2008). The highest Cr levels are found incerebellum, parallel to the distribution of creatine kinase and energy-requiringprocesses in the brain (Pouwels and Frahm 1998).
Glutamate is the major excitatory neurotransmitter in the human brain,mediator of synaptic plasticity and is implicated in the initiation and maintenance
D. J. Meyerhoff et al.

of addictive disorders (Kalivas and O’Brien 2008; Spanagel 2009). It is linked tometabolism through a neurotransmitter cycle between neurons and astrocytes. Inthis cycle, neurotransmitter/neuromodulator molecules released by the neurons aretaken up by transporters in surrounding glial cells. In the glia, they are converted toglutamine which is released to the neuron, where it is used for the re-synthesis ofthe neurotransmitter. Glu is less concentrated in cortical WM than in GM (Pouwelsand Frahm 1998). Although Glu is present in the brain at even higher concen-tration than NAA (about 11 mmol/L in GM), the MR detection sensitivity is poordue to the Glu signal being spread over a large number of closely spaced multiplettresonances, due to cancellation of overlapping resonances secondary to phasedifferences at longer echo times often used for MRS, and due to spectral overlapwith equally complicated glutamine resonances. With the increasing availability of3 T and higher magnetic field strength instruments, acquisition schemes thatsimplify the Glu resonance signal, and spectral fitting routines that make use ofdetailed resonance information, the quantification of cerebral Glu has becomeincreasingly feasible.
Gamma-aminobutyric acid (GABA) is the major inhibitory neurotransmitter inthe mammalian brain with a concentration of approximately 1.2 mmol/L in normalhuman cortex for the entire metabolically active pool (Hetherington et al. 1998;Rothman et al. 1993a). Abnormalities of GABA ergic neurotransmission havebeen implicated in many disorders, including alcoholism and in neurologic andpsychological diseases such as epilepsy, Huntington’s disease, depression andschizophrenia. Due to its lower concentration, its complicated MR spectral pattern,and due to its co-resonance with much larger metabolite signals (especially Cr), invivo GABA quantitation is even more challenging than in vivo Glu quantitation.Special spectral editing methods are needed, the most frequently used of which isbased on J modulation inhibition (Rothman et al. 1993b). GABA also co-resonateswith unspecific macromolecules and it has been shown that 40–60% of the edited‘‘GABA resonance’’ may come from macromolecules (Choi et al. 2007). Recentpreliminary studies showed that the macromolecules are evenly distributedthroughout the brain, supporting the assumption that changes observed in theedited ‘‘GABA resonance’’ are largely attributable to GABA changes (Xin et al.2010). The potential, however, for inter-individual variability of the macro-molecule contribution, especially in AUDs that are known to be associatedwith membrane alterations, needs to be investigated.
4 The Neurobiological Consequences of AUD and ChangesDuring Abstinence from Alcohol
Since the early 1990s MRS in both humans and animals has been used to inves-tigate the consequences of chronic and excessive alcohol consumption on brainmetabolism. Most published studies examined (mostly alcohol-dependent) indi-viduals in various phases of AUD treatment, whereas only a few studies examined
Chronic Alcohol Consumption, Abstinence and Relapse
Tab
le1
1H
MR
Sst
udie
spe
rfor
med
inan
imal
mod
els
ofal
coho
lde
pend
ence
Ref
eren
ceA
nim
alm
odel
Exp
erim
enta
lde
sign
Mai
nfi
ndin
gs
Hir
akaw
aet
al.
1994
Rat
s,si
ngle
dose
ofet
hano
lF
ield
stre
ngth
and
met
hod
unkn
own
Low
NA
A4
haf
ter
etha
nol
expo
sure
Bra
unov
áet
al.
2000
Rat
s,20
%et
hano
lad
libi
dum
for
8an
d14
wee
ks(m
ean
3.2
g/kg
/day
)4.
7T
,16
ms
Ste
amL
owm
I/C
raf
ter
8w
eeks
ofet
hano
ldi
et
Lee
Het
al.
2003
bR
ats
20%
etha
nol
indr
inki
ngw
ater
for
16an
d60
wee
ks4.
8T
,S
team
68m
sH
igh
Cho
/NA
Aat
wee
k16
,lo
wat
wee
k60
Pfe
ffer
baum
etal
.20
07
Rat
s,al
coho
l-pr
efer
ring
(50t
hge
nera
tion
)an
dth
iam
ine
defi
cien
cyan
dpy
rith
iam
ine
adm
inis
trat
ion
3T
GE
who
lebo
dy,
J-re
solv
edM
RS
No
prim
ary
effe
ctor
inte
ract
ion
ofet
hano
l
Nic
hola
set
al.
2008
Rat
s,si
ngle
dose
ofet
hano
l(5
g/kg
)17
T,
exvi
vo(p
erch
lori
cac
idex
trac
ts)
No
chan
ges
inbr
ain
met
abol
ites
O’L
eary
-M
oore
etal
.20
08
Rat
s,bi
nge
like
-neo
nata
let
hano
lex
posu
re11
.7T
,hi
ghre
solu
tion
MR
Sex
vivo
Low
NA
Aan
dta
urin
ein
cere
bell
uman
dst
riat
um,
high
mI
ince
rebe
llum
;low
Glu
infe
mal
era
ts;l
owC
ho/C
rin
hipp
ocam
pus,
high
inst
riat
um.
Zah
ret
al.
2008
Rat
s,24
wee
ksof
vapo
rize
det
hano
l(1
4h
atni
ght)
3T
,ba
seli
ne,
at16
and
24w
eeks
Cho
incr
ease
saf
ter
16an
d24
wee
ks,
mI,
Glx
and
Glu
incr
ease
afte
r24
wee
ks;
tren
dsfo
rlo
wer
NA
Aan
dlo
wer
Cr
Zah
ret
al.
2010
Rat
s,bi
nge
mod
el:
5g/
kget
hano
lvi
aor
alga
vage
,th
ena
max
imum
of3
g/kg
ever
y8
hfo
r4
days
3T
,ba
seli
ne,
4da
ysbi
nge,
7da
ysre
cove
ry;
dors
alhi
ppoc
ampu
s
NA
Aan
dC
rde
crea
sean
dC
hoin
crea
ses
afte
r4
days
;no
rmal
izat
ion
afte
r7
days
ofab
stin
ence
Web
er-F
ahr
etal
.20
10
Rat
s,ch
roni
cin
term
itte
ntet
hano
lva
por
into
xica
tion
9.4
T,
mP
FC
and
hipp
ocam
pus
Low
mI
and
NA
A,h
igh
Cho
and
Glu
duri
ngin
toxi
cati
on;f
urth
erin
crea
sed
Glu
inea
rly
wit
hdra
wal
D. J. Meyerhoff et al.

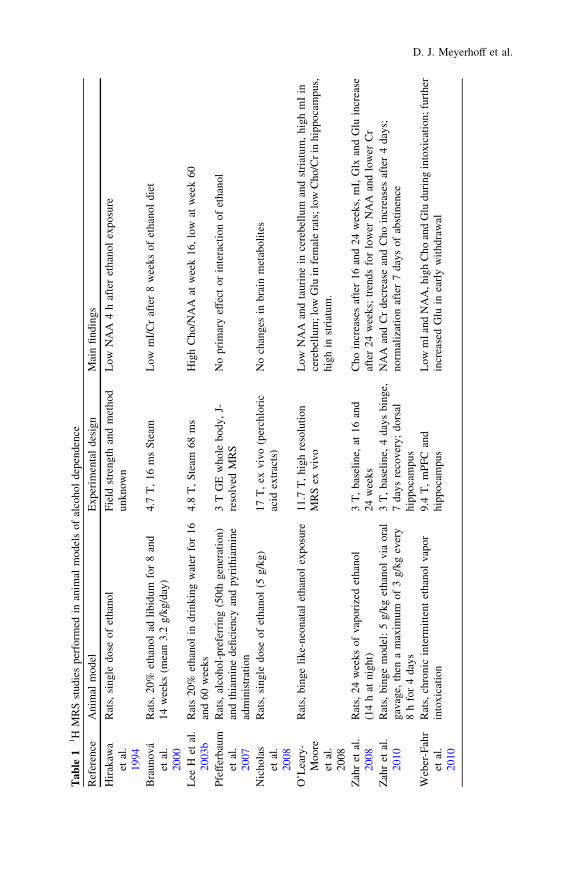
the larger pool of chronically drinking treatment-naïve individuals. Duration andlevel of alcohol consumption, age, nutritional status (including plasma thiaminelevels), family history of AUD and comorbid psychiatric and substance use dis-orders likely affect the magnitude and nature of brain metabolite abnormalitiesassociated with chronic alcohol consumption. Furthermore, duration of abstinencefrom alcohol critically affects metabolite levels. Table 1 presents all peer-reviewed1H MRS studies performed in animal models of alcohol dependence, whereasTable 2 lists peer-reviewed 1H MRS single-volume and 1H MRSI multi-volumestudies in humans.
The first MR study to suggest neuronal injury in the frontal cortex of abstinentalcoholics employed 1H MRSI (Fein et al. 1994). Subsequent research hasgenerally found reduced NAA levels in several brain regions of detoxified alcohol-dependent individuals, suggesting relatively widespread neuronal injury. Mostsingle-volume 1H MRS studies measured metabolites primarily in the frontal lobesand cerebellum of recovering alcoholics, brain regions most vulnerable to alcohol-induced tissue injury, and usually within 3–40 days of sobriety. They reported lowNAA in the frontal lobes (Bendszus et al. 2001; Jagannathan et al. 1996), thalamus(Jagannathan et al. 1996) and cerebellum (Parks et al. 2002; Seitz et al. 1999) ofindividuals with AUD suggesting neuronal injury, atrophied dendrites and/oraxons or derangement of metabolism. Other single-volume MRS studies reportedlower cerebellar Cho (Bendszus et al. 2001; Parks et al. 2002) and elevatedthalamic mI (Schweinsburg et al. 2000) relative to light-drinking controls,suggesting altered cell membrane metabolism and astrogliosis or osmotic changes.Lower concentrations of NAA in frontal WM and of NAA, Cho, and mI in thecerebellum correlated with lower neurocognitive and motor functioning (e.g.,Bendszus et al. 2001; Parks et al. 2002). Higher Cho/Cr was reported in theoccipital lobe of alcohol dependent patients (Modi et al. 2009), due to high Cho,low Cr concentrations or both. However, none of these human studies reported onsignificantly altered Cr concentrations in abstinent alcoholics.
Animal 1H MRS studies of the direct effects of alcohol on brain metabolism haveexclusively investigated rats. There are studies of the effects of single doses ofethanol (Hirakawa et al. 1994; Nicholas et al. 2008), rats bred for alcohol preference(Pfefferbaum et al. 2007), rats given 20% ethanol in water with ad libitum for atleast 8 weeks (Braunova et al. 2000; Lee et al. 2003a), binge drinking models byoral gavage (Zahr et al. 2010) and chronic ethanol exposure in vapor chambers(Weber-Fahr et al. 2010; Zahr et al. 2009). The first rodent MRS study by Hirakawa(1994) reported decreased NAA following a single dose of ethanol. This initialfinding was not replicated by other studies investigating single dose alcohol effects(Nicholas et al. 2008) or ad libitum ethanol consumption (Braunova et al. 2000) innon-selected rat populations or in genetically alcohol-preferring P rats (Pfefferbaumet al. 2007). Recent rodent MRS studies of 4 days of binge exposure (Zahr et al.2010) and chronic intermittent alcohol vapor exposure (Weber-Fahr et al. 2010)indicate that excessive alcohol intake is causally inked to decreased NAA. Bothstudies found decreased NAA in intoxicated animals as well as a rapid recovery ofNAA with discontinuation of alcohol exposure. Zahr et al. (2010) found normal
Chronic Alcohol Consumption, Abstinence and Relapse

Tab
le2
1H
MR
San
dM
RS
Ist
udie
spe
rfor
med
inhu
man
sw
ith
alco
hol
use
diso
rder
s(A
UD
)
Ref
eren
ceS
tudy
grou
psD
urat
ion
ofab
stin
ence
Maj
orfi
ndin
gs(a
llfi
ndin
gsst
atis
tica
lly
sign
ifica
ntfo
rpa
tien
tsco
mpa
red
toco
ntro
lsun
less
stat
edot
herw
ise)
Fei
net
al.
1994
11A
/9C
3–24
mon
ths
Gre
ater
fron
tal
than
pari
etal
cort
ical
NA
Alo
ssM
arti
net
al.
1995
10A
/9C
0–28
days
Incr
ease
ofC
ho/N
AA
ince
rebe
llar
verm
isw
ith
abst
inen
ce
Jaga
nnat
han
etal
.19
9610
A/
27C
1m
onth
Low
NA
A/C
ran
dN
AA
/Cho
ince
rebe
llum
,fr
onta
llo
bean
dth
alam
us
Sei
tzet
al.
1999
11A
/12
C4
days
Low
NA
A/C
rat
TE
=13
5m
san
dlo
wC
ho/C
rat
TE
=5
ms
Beh
aret
al.1
999
5A/1
0C5
wee
ksL
owG
AB
A?
hom
ocar
nosi
nein
occi
pita
llo
beS
chw
eins
burg
etal
.20
004A
/5C
5w
eeks
and
6ye
ars
Hig
hm
Iin
ante
rior
cing
ulat
eco
rtex
and
thal
amus
.N
AA
,C
hoan
dC
rno
tal
tere
d.m
Ino
rmal
at6
year
s.
Sch
wei
nsbu
rget
al.
2001
37A
/15
C1
mon
thL
owN
AA
infr
onta
lW
M,
high
mI
inW
M
Ben
dszu
set
al.
2001
17A
/12
C2
days
and
5w
eeks
Low
NA
A/C
rin
cere
bell
uman
dfr
onta
llob
e,lo
wC
ho/C
rin
cere
bell
um(d
ays
3–6)
and
norm
aliz
edva
lues
afte
r5
wee
ksP
arks
etal
.20
0231
A/
12C
4da
ysan
d3
mon
ths
Low
cere
bell
arN
AA
and
Cho
.N
AA
incr
ease
over
3m
onth
s(n
=11
).
Dur
azzo
etal
.20
0424
A/
26C
1w
eek
Low
NA
Aan
dC
hoin
fron
tall
obe,
low
Cho
inth
alam
usan
dpa
riet
allo
be.L
owN
AA
thro
ugho
utbr
ain
ofsm
okin
gvs
.no
n-sm
okin
gal
coho
lics
.C
ran
dm
Iun
chan
ged.
Mas
onet
al.
2006
12A
/8C
1w
eek
and
1m
onth
No
GA
BA
grou
pdi
ffer
ence
sin
occi
pita
lgr
aym
atte
r,bu
thi
gher
GA
BA
inno
n-sm
okin
gve
rsus
smok
ing
pati
ents
.H
igh
glut
amat
e?
glut
amin
ein
smok
ers.
GA
BA
decr
ease
inno
n-sm
okin
gal
coho
lics
over
1m
onth
.D
uraz
zoet
al.
2006
a25
A/
29C
1an
d4
wee
ksN
AA
and
Cho
incr
ease
infr
onta
lan
dpa
riet
allo
bes
over
4w
eeks
.m
Ian
dC
rin
fron
tal
whi
tem
atte
rin
crea
seov
er4
wee
ks.
Cha
nges
mor
epr
onou
nced
inno
n-sm
okin
gal
coho
lics
.E
nde
etal
.20
0533
A/
30C
3w
eeks
,3
and
6m
onth
sL
owC
hoin
cere
bell
uman
dfr
onta
llo
be,
low
NA
Ain
fron
tal
WM
;C
hoin
crea
ses
over
3m
onth
s(n
=14
);no
long
itud
inal
NA
Ach
ange
sov
er3
and
6m
onth
s(n
=11
)B
arts
chet
al.
2007
15A
/10
C5
days
and
6w
eeks
Low
fron
tom
esia
lN
AA
and
low
cere
bell
arC
ho.
Bot
hin
crea
seov
er6–
7w
eeks
(con
tinu
ed)
D. J. Meyerhoff et al.

Tab
le2
(con
tinu
ed)
Ref
eren
ceS
tudy
grou
psD
urat
ion
ofab
stin
ence
Maj
orfi
ndin
gs(a
llfi
ndin
gsst
atis
tica
lly
sign
ifica
ntfo
rpa
tien
tsco
mpa
red
toco
ntro
lsun
less
stat
edot
herw
ise)
Dur
azzo
etal
.20
0870
A1
mon
thL
owN
AA
inte
mpo
ral
gray
mat
ter
and
fron
tal
whi
tem
atte
ran
dlo
wC
hoin
fron
tal
gray
mat
ter
pred
ict
rela
pse
wit
hin
6–12
mon
ths
afte
rtr
eatm
ent
Gaz
dzin
ski
etal
.20
08a
35A
/32
HD
1w
eek
(alc
ohol
ics)
Low
NA
A,
Cho
,an
dm
Ith
roug
hout
the
brai
nof
trea
tmen
t-se
ekin
gal
coho
lics
vers
usno
n-tr
eate
dhe
avy
drin
kers
Gaz
dzin
ski
etal
.20
08b
24A
/14
C1
wee
kan
d4
wee
ksL
owN
AA
and
Cho
inm
edia
lte
mpo
ral
lobe
.N
AA
and
Cho
incr
ease
inno
n-sm
okin
gal
coho
lics
over
4w
eeks
.M
odi
etal
.20
099A
/13C
1w
eek
Hig
hC
ho/C
rin
occi
pita
llo
beD
uraz
zoet
al.
2010
b51
A/
26C
1w
eek
Low
NA
Aan
dC
rin
brai
nre
war
dsy
stem
ofth
ose
who
rela
pse
wit
hin
6–12
mon
ths
afte
rtr
eatm
entv
ersu
sab
stai
ners
and
cont
rols
Um
hau
etal
.20
1033
Ada
ys4
and
25of
med
icat
ion
Lon
gitu
dina
lde
crea
seof
Glu
/Cr
inan
teri
orci
ngul
ate
cort
exof
thos
etr
eate
dw
ith
acam
pros
ate
(N=
15)
vers
uspl
aceb
o(N
=18
)
A=
alco
hol
depe
nden
tpa
tien
ts,
C=
cont
rol
subj
ects
,H
=he
avy
drin
kers
Chronic Alcohol Consumption, Abstinence and Relapse

NAA levels after 7 days of recovery and Weber-Fahr et al. (2010) reported normallevels within 12 h of abstinence in the chronically exposed rats.
Abstinence from alcohol is associated with variable levels of recovery fromchronic alcohol-induced brain volume loss (atrophy) in humans (e.g., Pfefferbaumet al. 1995). Longitudinal 1H MRS studies during abstinence have the potential toilluminate some of the basic metabolic/cellular processes underlying such volumerecovery. Again, early studies focused primarily on the frontal lobes and cere-bellum. Martin et al. (1995) observed increased Cho/NAA in the cerebellar vermisover 3–4 weeks of abstinence from alcohol. Bendszus et al. (2001) reportedincreases in both frontal and cerebellar lobar NAA/Cr and cerebellar lobar Cho/Crratios after approximately 5 weeks of abstinence. After that interval, a higherfrontal NAA/Cr ratio was related to better auditory-verbal memory while increasedcerebellar vermis NAA/Cr ratio positively correlated with attention/concentration.Parks et al. (2002) observed that vermis NAA levels increased over 3 months ofabstinence from alcohol, which was also related to improved auditory-verballearning. In contrast to the group’s earlier study (Martin et al. 1995), vermian Cholevels did not recover after 3 months, and the authors suggested this might indicatecontinued compromise of cerebellar vermis tissue, consistent with neuropathologicfindings (Harper 1998). Higher mI was observed in the anterior cingulate gyrus,thalamus, frontal and parietal WM of 1 month-abstinent alcoholics but not in6 year-abstinent alcoholics (Schweinsburg et al. 2001, 2000), suggestingreversible membrane breakdown or osmolytic changes with abstinence fromalcohol. Bartsch et al. (2007) reported significant increases of cerebellar Cho andmesial frontal NAA over approximately 1 month of abstinence. Increasing mesialfrontal NAA was positively related to improving attention. Of note, the authorsincluded only smoking alcoholics who consumed less than 10 cigarettes per day.In a longitudinal multi-volume 1H MRSI study, Ende et al. (2005) observeddecreased Cho concentrations in the frontal WM, dorsolateral prefrontal cortex,superior frontal gyrus and cerebellar GM and vermis in individuals 1–4 weeksafter detoxification and a significant Cho recovery over the following 3 months ofabstinence. Decreased NAA was observed to be relatively persistent in the frontalWM, as no metabolite recovery (besides Cho) was observed after 3 months and nofurther recovery of any of the metabolite levels were apparent between 3 and6 months of abstinence.
Ende et al. (2006), (2010) also found that higher alcohol consumption in non-abstinent individuals with AUD and in light-drinking controls without AUD wasassociated with higher frontal Cho levels. While associations between higher Choand more acute alcohol consumption have been corroborated in rodent studies (Leeet al. 2003b; Weber-Fahr et al. 2010; Zahr et al. 2010; Zahr and Sullivan 2008),human studies in recently detoxified and 1 month-abstinent alcoholics have foundunchanged or reduced Cho levels (Schweinsburg et al. 2001, 2000; Bendszus et al.2001; Durazzo et al. 2004; Ende et al. 2005; Parks et al. 2002). Additionally, Chowas reported to show signficant increases or normalize with continued abstinence(Durazzo et al. 2006a; Ende et al. 2005). Whether this difference is a consequenceof chronic and excessive alcohol consumption or a consequence of detoxification
D. J. Meyerhoff et al.

per se requires further investigation. Zahr et al. (2010) suggested that low Cho inhuman studies can be explained by undetected or subclinical pathologies,including thiamine deficiency or liver cirrhosis. Lee et al. (2003a) found an initialCho increase in rats after 16 weeks of alcohol exposure followed by a significantdecrease after 60 weeks, which supports the hypothesis that Cho levels reverse anddecrease below normal with duration of chronic alcohol abuse. More recent rodentstudies, however, failed to detect a significant Cho decrease with prolongedexposure or during withdrawal (Weber-Fahr et al. 2010; Zahr et al. 2010, 2009).Therefore, the time course of Cho concentration changes in alcohol dependenceand during abstinence do not appear to be consistent across studies, and the sig-nificance of the temporal Cho level dynamics as a function of drinking severity areas of yet unclear. They are likely complicated by the fact that Cho levels reflectdifferent biological processes, including cellular membrane turnover and density aswell as myelin anabolism and catabolism.
To date, there is no fully quantitative AUD study that reported significantreductions in Cr concentrations relative to light-drinking controls and only onestudy (Durazzo et al. 2006a) that reported a significant Cr increase with abstinence.In contrast, recent rodent studies of active alcohol exposure support a Cr decrease(Zahr et al. 2010; Weber-Fahr et al. 2010) that quickly normalized after alcoholwithdrawal. Furthermore, mI, the putative astrocyte marker and osmolyte,decreased in rodents during chronic ethanol exposure and recovered to normallevels after alcohol withdrawal (Braunova et al. 2000; Weber-Fahr et al. 2010).Studies in human alcoholics reported no mI changes, except for one study showingelevated mI in some brain regions after detoxification (Schweinsburg et al. 2000).Although not consistent across species, the mI findings overall suggest somealcohol-related alterations of cell membrane metabolism and gliotic or osmoticchanges.
Modulations and adaptations of reciprocal glutamatergic and GABAergicprojections from frontal brain regions, basal forebrain and midbrain likely con-tribute to the neural basis of substance dependence (Kalivas and Volkow 2005).Chronic alcohol-induced adaptations within the glutamatergic systems, a hyper-glutamatergic state, contribute to the induction and maintenance of alcoholdependence (Kalivas et al. 2009; Spanagel 2009). Those adaptations then maycause a hyper-excitability of the central nervous system when alcohol is removed(withdrawal) and represent a mechanism involved in early relapse behavior (Lit-tleton 1995). This concept receives support from the application of glutamatemodulators or functional glutamate antagonists, such as acamprosate, for thereduction of alcohol consumption. Such pharmacotherapies have becomeincreasingly important in treating both AUD and other substance use disorders,centering on medications modulating common neurotransmitters such as serotonin,dopamine, Glu and GABA. Thus, a better understanding of the specific effects ofAUD on brain GABA and Glu concentrations and flux may further advancethe development and efficacy of pharmacological treatment. Basal cerebral con-centrations of specific neurotransmitters have been linked to behavior. In rats,frontal Glu transmission has been associated with drug seeking (Kalivas and
Chronic Alcohol Consumption, Abstinence and Relapse

Volkow 2005; McFarland et al. 2003), and in humans, low striatal Glu concen-tration measured by 1H MRS has been linked to a decline in neurocognitive testperformance with normal ageing (Zahr et al. 2008). Glu concentrations in theprefrontal medial cortex were associated negatively with sensation seeking andpositively with measures of impulsivity (Gallinat et al. 2007a; Hoerst et al. 2010).Modulation of the inhibitory GABA system by alcohol is implicated in thedevelopment of alcohol tolerance, dependence and withdrawal and in emotionalprocessing/judgment. In humans, some studies report decreased plasma and CSFGABA at 1 month of abstinence from alcohol and normal GABA levels by6 months of sobriety (Adinoff et al. 1995; Coffman and Petty 1985).
Although the 1H MRS detectable amino acid levels represent the metabolicallyavailable brain pools (which are much larger than the respective neurotransmitterpools), they are in tight equilibrium with synaptic levels (Rothman et al. 2003).Therefore, MRS-derived GABA and Glu concentrations provide valuable infor-mation on the role and functional significance of these neurotransmitter systems.Consistent with plasma and CSF GABA levels, tissue GABA levels measured by1H MRS in occipital cortex of a small sample of alcoholics at about 1 month ofabstinence were approximately 25% lower than in non-alcoholic controls (Beharet al. 1999). A later study, however, showed that GABA levels were elevated in1 week abstinent non-smoking alcoholics and normalized after 4 weeks (Masonet al. 2006). GABA levels in smoking alcoholics were normal and did not changeover time. Glu is an endogenous agonist of N-methyl-D-aspartate receptors andtheir increased activity (postsynaptic receptors) may produce neurotoxicity pre-sumably through dysregulation of Ca2 ? influx (see Bleich et al. 2004; De Witte2004). Glu levels are increased during alcohol withdrawal in animal models(see Bleich et al. 2004 for review) and in anterior cingulate cortex of humans(Frischknecht et al. 2010). At 6 days of abstinence, anterior cingulate Glu wasdecreased below control levels and then increased significantly into the controlrange within the following 24 days of sobriety (Mon et al. 2010). An early in vivo1H MRS study suggests that Glx (the sum of Glu and glutamine) in healthycontrols is lower relative to placebo 20 min after infusion of acamprosate (whichhas been shown to decrease alcohol consumption) (Bolo et al. 1998), consistentwith microdialysis results in alcohol-dependent rats treated with acamprosate(Dahchour et al. 2005). In a small placebo controlled clinical trial, Glu in anteriorcingulate (relative to Cr levels) decreased in recently abstinent alcohol-dependentindividuals over 4 weeks of treatment with acamprosate (Umhau et al. 2010). Inrats, Zahr et al. (2009) found increased Glx after 24 weeks of ethanol vaporexposure when the blood alcohol level was still high, while Weber-Fahr et al.(2010) detected increasing Glu levels during acute withdrawal that were still highafter 3 days of abstinence. Glu levels normalized within 3 weeks of abstinencewhen withdrawal symptoms subsided. Together, these dynamic changes observedin both rodent and human studies are consistent with a hyperglutamatergic stateduring withdrawal and normalization of central Glu levels within a few weeks ofsustained abstinence.
D. J. Meyerhoff et al.

5 Treatment-Seeking Versus Treatment Naïve Alcoholicsand Common Comorbidities
The vast majority of research investigating brain changes in AUD has been con-ducted with individuals in substance abuse treatment. Although readily accessiblefor scientific research, these treatment-seeking individuals are a minority amongpersons with AUD, with the majority being treatment-naïve (Fein and Landman2005; Hasin et al. 2007). Treatment-seeking cohorts also generally present with ahigher severity of medical, psychiatric and substance use comorbidities that mayaffect MR outcome measures. These common comorbidities may independentlyinfluence brain biochemistry, structure and function. Furthermore, treatment-seeking alcoholics usually have more severe alcohol consumption (up to 50%higher consumption over lifetime) and more periods of abstinence than theirtreatment-naïve counterparts (Fein and Landman 2005). Due to the prevalenceand/or magnitude of comorbid conditions and alcohol consumption, it may bereasonably expected that the treatment–seeking population has generally greaterbrain injury than the treatment-naïve population and that both do not simplyrepresent a continuum of AUD on a progressive scale. It is widely recognized thatthese comorbid characteristics may promote considerable variability in the patternand magnitude of neurobiological and neurocognitive abnormalities demonstratedin AUD, during detoxification and sustained abstinence. Our research has focusedon studying the unique impact of common comorbidities on the brain in personswith AUD to try to better understand the brain changes in this complex population(reviewed in (Durazzo and Meyerhoff 2007).
In a 1H MRSI study we observed that, relative to light-drinking controls,metabolite abnormalities in community-dwelling heavy drinkers (the vast majoritywere alcohol-dependent) are less pronounced and demonstrate a different patternof metabolite abnormalities than reported in recently abstinent treated alcoholics(Gazdzinski et al. 2008a; Meyerhoff et al. 2004). Nevertheless, compared to light/non-drinking controls, treatment-naïve heavy drinkers had lower NAA concen-trations in frontal WM and parietal GM, suggesting greater neuronal injury, andparietal GM Cr was elevated. Although small, the frontal NAA reduction wasfunctionally significant as it was associated with poorer performances on measuresof executive skills and working memory as well as lower frontal P300b ampli-tudes. Furthermore, age, sex, family history of alcohol problems and drinkingpattern (binge vs. non-binge) modulated brain metabolite abnormalities. We alsocompared these treatment-naïve heavy drinkers to 1 week-abstinent treatment-seeking alcoholics (Gazdzinski et al. 2008a). In conjunction with smaller lobarGM volumes and thalami in treatment-seeking versus treatment-naïve individuals,NAA, Cho and mI concentrations were lower in multiple brain regions. Whilelower WM NAA was completely explained by average number of drinks permonth over 1 year prior to study, the other metabolite group differences were notexplained by alcohol consumption levels, demographic, and clinical variables orby psychiatric comorbidities.
Chronic Alcohol Consumption, Abstinence and Relapse

Taken together, the brain structural, metabolic and functional differencesbetween treatment-seeking and treatment-naïve alcoholic populations suggest thatthe neurobiological abnormalities observed in clinical convenience samples ofalcoholics in treatment cannot be generalized to the much larger treatment-naïvepopulation with AUD. Instead, the comorbid occurrence of neuropsychiatric andsubstance use factors need to be considered specifically when examining neuro-biological and neurocognitive consequences in AUD.
The most prevalent comorbidity in AUD is chronic cigarette smoking. It isestimated that 60–80% of AUDs are chronic smokers (Durazzo et al. 2007;Romberger and Grant 2004). A growing body of research suggests that chronicsmoking, independent of AUD, is associated with abnormalities in brain morphol-ogy, cerebral blood flow, neurochemistry and neurocognition that are similar tothose reported in AUD (Durazzo and Meyerhoff 2007). We have investigated theeffects of concurrent chronic cigarette smoking on regional brain morphology(Gazdzinski et al. 2005), blood flow (Gazdzinski et al. 2006; Mon et al. 2009) andmetabolite concentrations (Durazzo et al. 2004) in 1 week-abstinent, treatment-seeking individuals with AUD as well as longitudinal brain metabolite changesduring short-term abstinence from alcohol (Durazzo et al. 2006a). Both chronicalcohol consumption and chronic smoking independently are associated with sig-nificant neocortical GM loss. We observed that 1 week-abstinent, treatment-seekingsmoking alcoholics compared to their non-smoking counterparts demonstratedlower NAA concentrations in frontal WM, parietal GM, and lenticular nuclei as wellas lower NAA and Cho in the midbrain (Durazzo et al. 2004). Alcohol dependence,independent of smoking, was associated with lower Cho concentrations in thethalamic and parietal lobes and lower frontal lobe NAA and Cho, the latter con-sistent with other reports (see above). Neither alcohol dependence nor chronicsmoking was associated with significant alterations of Cr and mI concentrations inany of the lobar regions analyzed. Among smoking alcoholics, greater nicotinedependence and a higher number of cigarettes per day were negatively correlatedwith absolute NAA concentrations in thalamic and lenticular nuclei. Lower cere-bellar vermis NAA was associated with poorer visuomotor scanning speed (smok-ers) and poorer visuospatial learning and memory (non-smokers). These in vivo 1HMRSI findings suggest that chronic smoking compounds alcohol-induced neuronalinjury and cell membrane injury in the frontal lobes of persons with AUD and hasindependent adverse effects on neuronal viability and cell membrane turnover/synthesis in the vermis and midbrain. Findings are largely consistent with metabolicchanges described in non-alcoholic chronic smokers (Gallinat et al. 2007b). 1H MRSstudies also showed that brain GABA concentrations in animals and humans aremodulated by nicotine and/or cigarette smoking (Epperson et al. 2005; Zhu andChiappinelli 1999), so that alterations of these metabolites in smoking alcoholicswould not be a surprise. Such smoking-induced metabolic brain abnormalities arelikely of clinical significance as they are accompanied by lower performance oncognitive tests that require fast and flexible processing, such as set-shifting, pro-cessing speed and cognitive efficiency (see e.g., Durazzo et al. 2006b; Friend et al.2005; Glass et al. 2006 and references cited therein).
D. J. Meyerhoff et al.

In longitudinal 1H MRSI studies of treatment-seeking alcoholics, afterapproximately 1 month of abstinence from alcohol, we found significant increasesof NAA and Cho concentrations in WM and GM of the frontal and parietal lobes(Durazzo et al. 2006a), consistent with the literature described above (Bendszuset al. 2001; Ende et al. 2005). Despite not being significantly reduced cross-sectionally, mI and Cr levels increased over time in the frontal WM only. Whensmoking status was considered, non-smokers showed widespread metaboliteincreases, whereas increases in smokers were much less pronounced and seen infewer brain regions. In non-smokers, metabolite gains were related to improve-ments in visuospatial learning, visuospatial memory and working memory, visu-omotor scanning speed and incidental learning, while smoking alcoholics showedsignificantly fewer of such relationships. Furthermore, in abstinent smokers, longersmoking duration was related to smaller longitudinal increases in frontal WMNAA, frontal WM Cho, and thalamic Cho. Similarly, NAA and Cho levelsincreased significantly in medial temporal lobe (including hippocampal tissue) ofnon-smokers abstinent for 1 month, but the corresponding concentrations insmokers remained depressed relative to non-smoking light-drinking controls(Gazdzinski et al. 2008b). mI tended to increase in non-smoking alcoholics. In thecombined alcoholic cohort (i.e., smokers plus non-smokers), increasing Cho, Crand mI were associated with improving visuospatial memory.
Occipital GM GABA concentrations during recovery from AUD are alsomodulated by smoking status (Mason et al. 2006). At 1 week of abstinence,cortical GABA levels were higher in alcohol-dependent non-smokers thansmokers. After approximately 3 weeks of abstinence, GABA levels were lowerthan at 1 week and similar between alcoholic non-smokers and smokers. HigherGABA during early withdrawal may reflect compensation for reduced corticalbenzodiazepine-GABAA receptor function thought to contribute to alcohol toler-ance and withdrawal. The subsequent decline may reflect ‘‘normalization’’ ofGABAA receptor function with sobriety.
6 The Neurobiological Correlates of Relapse in Alcohol UseDisorders (AUD)
More than 60% of individuals who seek treatment for AUD will return to haz-ardous levels of alcohol consumption, with the majority relapsing within 6 monthsfollowing their last treatment (see Durazzo et al. 2010a, 2010b). While muchresearch has addressed the potential neuropsychological, psychiatric, sociodemo-graphic and behavioral factors associated with relapse in AUD, the neurobiologicalfactors associated with sustained sobriety and/or increased risk for relapse aftertreatment for AUD are not well understood. A greater understanding of theseobjective factors can provide a better understanding of the mechanisms driving therelapse/remit cycle and maintenance of long-term abstinence.
Chronic Alcohol Consumption, Abstinence and Relapse

In a longitudinal 1H MRS study of treatment-seeking alcohol-dependent indi-viduals, Parks and colleagues (Parks et al. 2002) observed that those who relapsedwithin 3 weeks of study demonstrated lower cerebellar NAA and Cho at 3–5 daysof abstinence relative to controls. However, cerebellar or frontal metabolite con-centration reductions were not observed between controls and individuals whorelapsed after 3 weeks of abstinence. A more recent study combined 1H MRSImeasurement of absolute regional metabolite concentrations with assessment ofmajor psychiatric disorders, and comprehensive neurocognitive testing in treat-ment-seeking participants at approximately 1 month of abstinence from alcohol(Durazzo et al. 2008). Participants were followed for 6–12 months after treatment,and were retrospectively classified as abstainers (no alcohol consumption) andresumers (any alcohol consumption) and then contrasted on outcome measures at1 month of abstinence. Temporal GM NAA, frontal WM NAA, frontal GMCho, processing speed and comorbid unipolar mood disorder were independentpredictors of resumption of drinking. In a companion 1H MRSI study of a largelysimilar population, we analyzed metabolite levels at 1 week of abstinence inregions of the extended brain reward system as a function of relapse status (Durazzoet al. 2010b). Resumers demonstrated significantly lower baseline NAA concen-trations than non-smoking non/light-drinkers and abstainers in dorsolateral pre-frontal cortex, anterior cingulate cortex, insula, superior corona radiata andcerebellar vermis. Resumers also exhibited lower Cr concentrations than abstainersin the dorsolateral prefrontal cortex, superior corona radiata and cerebellar vermis.Importantly, abstainers did not differ significantly from the controls on baselinemetabolite concentrations in any region. In resumers, moderate to strong rela-tionships were apparent between regional baseline NAA levels and several mea-sures of post-treatment alcohol consumption, strongly suggesting that lowerneuronal integrity in regions of the extended brain reward system at baseline pre-dicts greater relapse severity.
7 Conclusions and Perspectives
This review describes the association of AUD with adverse neurobiological con-sequences as measured by 1H MRS in treatment-seeking and non-treatment-seeking cohorts and animal models. The most consistent finding isreduced NAA in both AUD and chronically alcohol exposed rats and its partial(human) to full (rat) reversibility with sustained abstinence. In general, alcohol-induced changes in animal brain metabolites are seen during the intoxicationphase, with little if any long-lasting dependence-related changes in brain metab-olites. Most human studies, however, not only detect neurobiological changes afteryears of chronic alcohol consumption, but also observe partial recovery of NAAover several months of abstinence, while Cho ultimately recovers to control levelswithin a shorter time frame. The potential of 1H MRS to monitor dynamic changesof metabolite levels as a function of duration of sobriety and as a consequence ofpharmacological treatment has been demonstrated in both animals and humans.
D. J. Meyerhoff et al.

A review of the human AUD literature suggests that examining individuals withAUD as a homogeneous group without consideration of common comorbidities,such as other substance use (including chronic cigarette smoking) and psychiatricdisorders (even if subclinical) may confound our understanding of the factorscontributing to the neurobiological and neurocognitive dysfunction observed inAUD. Additionally, failure to consider the potential influence of commoncomorbidities in AUD may obscure the identification of the neurobiologicalfactors associated with neurobiological recovery during abstinence as well as thoserelated to relapse. MR-based spectroscopy studies of animal models have theadvantage of investigating the nature and pattern of brain injury that are specifi-cally related to chronic alcohol exposure, without the potential of confoundingeffects from comorbidities common in human AUD. To date, there are no 1H MRSstudies in animals that involved exposure to alcohol and other substances, forexample the combined effects of alcohol and cigarette smoke/nicotine exposure onbrain metabolite. In AUD, the comorbid misuse of other substances such asmarijuana, cocaine and cigarettes is exceedingly common (polysubstance use) andthe clinical reality facing most treatment providers. Therefore, it may be advisablethat future research with animal models incorporate other substances that areprevalent in AUD to facilitate increased generalizability of research findings tohumans.
Human studies reveal that chronic smoking in treatment-seeking and treatment-naïve cohorts with AUD compounds regional neurobiological abnormalities.Furthermore, chronic smoking in AUD is associated with diminished, perhapsdelayed, recuperation of regional biochemical markers of neuronal viability andcell membrane synthesis/turnover during abstinence from alcohol. If chroniccigarette smoking is confirmed to modulate brain neurobiology and neurocognitionin additional human studies and in animal models, we may have to entertain thepossibility that smoking and non-smoking individuals with AUD differ in thenature or extent of their response to pharmacological and/or behavioral interven-tions designed to promote abstinence from alcohol. Even without theseconfirmatory studies, the reviewed literature, in conjunction with the knownmortality and morbidity associated with chronic smoking, lends support to thegrowing clinical initiative that encourages chronic smokers entering treatment forAUD to participate also in a smoking cessation program.
Given the high prevalence of medical, psychiatric and substance misusecomorbidities in AUD, it is important to understand to what extent these factorscan contribute to the neurobiological and neurocognitive abnormalities observed inAUD. Examining AUD samples under particular consideration of these commoncomorbidities increases the clinical relevance and generalizability of the data, assuch cohorts are more representative of the typical treatment-naïve and treatment-seeking populations. Additional prospective research, with larger groups of femaleparticipants is required to evaluate for sex effects, particularly since it is unclear ifmales and females manifest the same degree or pattern of alcohol-inducedneurobiological and neurocognitive abnormalities at equivalent drinking severitylevels (Mann et al. 1992; Parsons and Nixon 1998; Sullivan et al. 2004).
Chronic Alcohol Consumption, Abstinence and Relapse

Moderate to strong relationships between 1H MRS measures and variousmeasures of neurocognition in cross-sectional and longitudinal studies indicatethat MRS-derived neurobiological measures are robust and relevant predictors ofbrain function and therefore drug use behavior. Hence, the application of MRS inthe study of neurobiological factors associated with abstinence and relapse high-light the clinical usefulness of this approach for the prediction of relapse, inconjunction with the more conventional neurocognitive and psychiatric factors.Recent 1H MRS data support the potential of MR-derived measures to assist in theidentification of objective factors associated with increased risk for resumption ofhazardous drinking following treatment for AUD. Neuroimaging-based investi-gations of the factors associated with the chronic relapse/remit cycle in AUD mayalso facilitate the development of more efficacious pharmacological and behavioralinterventions for AUD.
Finally, in vivo 1H MRS, as part of the emerging field of ‘‘imaging genetics’’may provide readily accessible, objective, functionally significant and region-specific neurobiological measures (endophenotypes) that successfully link specificgenotypes to neurocognition and psychiatric symptomatology in relatively smallpatient cohorts (for review see Meyerhoff and Durazzo 2008). If early evidence ofgenetic effects on MRS-detectable metabolite measures are confirmed, MRSgenetics research will not only offer clues to the functional significance of geneticdifferences in AUD, but MRS can potentially influence the future of clinicalmanagement of AUD via monitoring the efficacy of pharmacological and behav-ioral interventions as a function of genotype.
Acknowledgements This review was supported by NIH R01 AA10788 (DJM), K01 DA24136(TCD), and grants from the Deutsche Forschungsgemeinschaft (SFB636) (GE) and the Bun-desministerium für Bildung und Forschung (BMBF) NGFN-plus (GE).
References
Adinoff B, Kramer GL, Petty F (1995) Levels of gamma-aminobutyric acid in cerebrospinal fluidand plasma during alcohol withdrawal. Psychiatry Res 59:137–144
Babikian T, Freier MC, Ashwal S, Riggs ML, Burley T, Holshouser BA (2006) MR spectroscopy:predicting long-term neuropsychological outcome following pediatric TBI. J Magn ResonImaging 24:801–811
Bartsch AJ, Homola G, Biller A, Smith SM, Weijers HG, Wiesbeck GA, Jenkinson M, DeStefano N, Solymosi L, Bendszus M (2007) Manifestations of early brain recovery associatedwith abstinence from alcoholism. Brain 130:36–47
Baslow MH (2003) N-acetylaspartate in the vertebrate brain: metabolism and function.Neurochem Res 28:941–953
Baslow MH, Guilfoyle DN (2007) Using proton magnetic resonance imaging and spectroscopy tounderstand brain ‘‘activation’’. Brain Lang 102:153–164
Behar K et al (1999) Preliminary evidenceof low cortical GABA levels in localized 1H-MRspectra of alcohol-dependent and hepatic encephalopathy patients. Am J Psychiatry 156:952–954
D. J. Meyerhoff et al.

Benarroch EE (2008) N-acetylaspartate and N-acetylaspartylglutamate: neurobiology and clinicalsignificance. Neurology 70:1353–1357
Bendszus M, Weijers HG, Wiesbeck G, Warmuth-Metz M, Bartsch AJ, Engels S, Boning J,Solymosi L (2001) Sequential MR imaging and proton MR spectroscopy in patients whounderwent recent detoxification for chronic alcoholism: correlation with clinical andneuropsychological data. Am J Neuroradiol 22:1926–1932
Bertolino A, Sciota D, Brudaglio F, Altamura M, Blasi G, Bellomo A, Antonucci N, Callicott JH,Goldberg TE, Scarabino T, Weinberger DR, Nardini M (2003) Working memory deficits andlevels of N-acetylaspartate in patients with schizophreniform disorder. Am J Psychiatry160:483–489
Bleich S, Degner D, Sperling W, Bonsch D, Thurauf N, Kornhuber J (2004) Homocysteine as aneurotoxin in chronic alcoholism. Prog Neuropsychopharmacol Biol Psychiatry 28:453–464
Bolo N, Nedelec JF, Muzet M, De Witte P, Dahchour A, Durbin P, Macher JP (1998) Centraleffects of acamprosate: part 2. Acamprosate modifies the brain in vivo proton magneticresonance spectrum in healthy young male volunteers. Psychiatry Res 82:115–127
Boulanger Y, Labelle M, Khiat A (2000) Role of phospholipase A(2) on the variations of thecholine signal intensity observed by 1H magnetic resonance spectroscopy in brain diseases.Brain Res Brain Res Rev 33:380–389
Brand A, Richter-Landsberg C, Leibfritz D (1993) Multinuclear NMR studies on the energymetabolism of glial and neuronal cells. Dev Neurosci 15:289–298
Braunova Z, Kasparova S, Mlynarik V, Mierisova S, Liptaj T, Tkac I, Gvozdjakova A (2000)Metabolic changes in rat brain after prolonged ethanol consumption measured by 1H and 31PMRS experiments. Cell Mol Neurobiol 20:703–715
Brenner RE, Munro PM, Williams SC, Bell JD, Barker GJ, Hawkins CP, Landon DN, McDonaldWI (1993) The proton NMR spectrum in acute EAE: the significance of the change in theCho:Cr ratio. Magn Reson Med 29:737–745
Choi C, Bhardwaj PP, Kalra S, Casault CA, Yasmin US, Allen PS, Coupland NJ (2007)Measurement of GABA and contaminants in gray and white matter in human brain in vivo.Magn Reson Med 58:27–33
Coffman JA, Petty F (1985) Plasma GABA levels in chronic alcoholics. Am J Psychiatry142:1204–1205
Crews FT, Nixon K (2009) Mechanisms of neurodegeneration and regeneration in alcoholism.Alcohol Alcohol 44:115–127
Crews FT, Collins MA, Dlugos C, Littleton J, Wilkins L, Neafsey EJ, Pentney R, Snell LD,Tabakoff B, Zou J, Noronha A (2004) Alcohol-induced neurodegeneration: when, where andwhy? Alcohol Clin Exp Res 28:350–364
Daeppen JB, Smith TL, Danko GP, Gordon L, Landi NA, Nurnberger JI Jr, Bucholz KK, RaimoE, Schuckit MA (2000) Clinical correlates of cigarette smoking and nicotine dependence inalcohol-dependent men and women. The collaborative study group on the genetics ofalcoholism. Alcohol Alcohol 35:171–175
Dahchour A, Lallemand F, Ward RJ, De Witte P (2005) Production of reactive oxygen speciesfollowing acute ethanol or acetaldehyde and its reduction by acamprosate in chronicallyalcoholized rats. Eur J Pharmacol 520:51–58
Dawson DA, Goldstein RB, Grant BF (2007) Rates and correlates of relapse among individuals inremission from DSM-IV alcohol dependence: a 3 year follow-up. Alcohol Clin Exp Res31:2036–2045
De Stefano N, Matthews PM, Arnold DL (1995) Reversible decreases in N-acetylaspartate afteracute brain injury. Magn Reson Med 34:721–727
De Witte P (2004) Imbalance between neuroexcitatory and neuroinhibitory amino acids causescraving for ethanol. Addict Behav 29:1325–1339
Dlugos CA, Pentney RJ (1997) Morphometric evidence that the total number of synapses onPurkinje neurons of old F344 rats is reduced after long-term ethanol treatment and restored tocontrol levels after recovery. Alcohol and Alcoholism 32:161–172
Chronic Alcohol Consumption, Abstinence and Relapse

Durazzo TC, Meyerhoff DJ (2007) Neurobiological and neurocognitive effects of chroniccigarette smoking and alcoholism. Front Biosci 12:4079–4100
Durazzo TC, Gazdzinski S, Banys P, Meyerhoff DJ (2004) Cigarette smoking exacerbateschronic alcohol-induced brain damage: a preliminary metabolite imaging study. Alcohol ClinExp Res 28:1849–1860
Durazzo TC, Gazdzinski S, Banys P, Meyerhoff DJ (2006a) Brain metabolite concentrations andneurocognition during short-term recovery from alcohol dependence: Preliminary evidence ofthe effects of concurrent chronic cigarette smoking. Alc Clin Exp Research 30:539–551
Durazzo TC, Rothlind JC, Gazdzinski S, Banys P, Meyerhoff DJ (2006b) A comparison ofneurocognitive function in nonsmoking and chronically smoking short-term abstinentalcoholics. Alcohol 39:1–11
Durazzo TC, Gazdzinski S, Meyerhoff DJ (2007) The neurobiological and neurocognitiveconsequences of chronic cigarette smoking in alcohol use disorders. Alcohol Alcohol 42:174–185
Durazzo TC, Gazdzinski S, Yeh PH, Meyerhoff DJ (2008) Combined neuroimaging, neurocog-nitive and psychiatric factors to predict alcohol consumption following treatment for alcoholdependence. Alcohol Alcoholi 43:683–691
Durazzo T, Gazdzinski S, Mon A, Meyerhoff D (2010a) Cortical perfusion in alcohol-dependentindividuals during short-term abstinence: relationships to resumption of hazardous drinkingafter treatment. Alcohol 44:201–210
Durazzo TC, Pathak V, Gazdzinski S, Mon A, Meyerhoff DJ (2010b) Metabolite levels in thebrain reward pathway discriminate those who remain abstinent from those who resumehazardous alcohol consumption after treatment for alcohol dependence. J Stud Alcohol Drugs71:278–289
Ende G, Braus DF, Walter S, Weber-Fahr W, Soher B, Maudsley AA, Henn FA (2000) Effects ofage, medication, and illness duration on the N-acetyl aspartate signal of the anterior cingulateregion in schizophrenia. Schizophr Res 41:389–395
Ende G, Welzel H, Walter S, Weber-Fahr W, Diehl A, Hermann D, Heinz A, Mann K (2005)Monitoring the Effects of Chronic Alcohol Consumption and Abstinence on BrainMetabolism: A Longitudinal Proton Magnetic Resonance Spectroscopy Study. Biol Psychi-atry 58:974–980
Ende G, Walter S, Welzel H, Demirakca T, Wokrina T, Ruf M, Ulrich M, Diehl A, Henn FA,Mann K (2006) Alcohol consumption significantly influences the MR signal of frontalcholine-containing compounds. Neuroimage 32:740–746
Ende G, Hermann D, Hoerst M, Tunc-Skarka N, Oberthuer G, Wichert S, Rabinstein J, Weber-Fahr W, Mann K, Vollstaedt-Kein S (2010) Frontal white matter choline-containingcompounds increase with alcohol consumption and glutamate decreases with increasingaddiction criteria. ISMRM, Stockholm, Sweden, pp Prog #: 2160
Epperson CN, O’Malley S, Czarkowski KA, Gueorguieva R, Jatlow P, Sanacora G, Rothman DL,Krystal JH, Mason GF (2005) Sex, GABA, and nicotine: the impact of smoking on corticalGABA levels across the menstrual cycle as measured with proton magnetic resonancespectroscopy. Biol Psychiatry 57:44–48
Fabricius K, Pakkenberg H, Pakkenberg B (2007) No changes in neocortical cell volumes or glialcell numbers in chronic alcoholic subjects compared to control subjects. Alcohol Alcohol42:400–406
Fein G, Landman B (2005) Treated and treatment-naïve alcoholics come from different popu-lations. Alcohol 35:19–26
Fein G, Meyerhoff DJ, Di Sclafani V, Ezekiel F, Poole N, MacKay S, Dillon WP, Constans J-M,Weiner MW (1994) 1H magnetic resonance spectroscopic imaging separates neuronal fromGlial changes in alcohol-related brain atrophy. Chapter in NIAAA Research Monograph No.27/Alcohol and Glial Cells: 227–241
Fein G, Di Sclafani V, Cardenas VA, Goldmann H, Tolou-Shams M, Meyerhoff DJ (2002)Cortical gray matter loss in treatment–naïve alcohol dependent individuals. Alcohol Clin ExpRes 26:558–564
D. J. Meyerhoff et al.

Ferguson KJ, MacLullich AM, Marshall I, Deary IJ, Starr JM, Seckl JR, Wardlaw JM (2002)Magnetic resonance spectroscopy and cognitive function in healthy elderly men. Brain125:2743–2749
Fisher SK, Novak JE, Agranoff BW (2002) Inositol and higher inositol phosphates in neuraltissues: homeostasis, metabolism and functional significance. J Neurochem. 82:736–754
Friend KB, Malloy PF, Sindelar HA (2005) The effects of chronic nicotine and alcohol use onneurocognitive function. Addict Behav 30:193–202
Frishknecht U, Hermann D, Hoerst N, Tunc-Skarka N, Kiefer F, Ende G, Mann KF (2010)Increased ACC glutamate levels in patients during alcohol withdrawal return to normal aftertwo weeks of abstinence. Alcohol Clin Exp Res 34(Suppl.):132A
Gallinat J, Kunz D, Lang UE, Neu P, Kassim N, Kienast T, Seifert F, Schubert F, Bajbouj M(2007a) Association between cerebral glutamate and human behaviour: the sensation seekingpersonality trait. Neuroimage 34:671–678
Gallinat J, Lang UE, Jacobsen LK, Bajbouj M, Kalus P, von Haebler D, Seifert F, Schubert F(2007b) Abnormal hippocampal neurochemistry in smokers: evidence from proton magneticresonance spectroscopy at 3 T. J Clin Psychopharmacol 27:80–84
Gazdzinski S, Durazzo TC, Studholme C, Song E, Banys P, Meyerhoff DJ (2005) Quantitativebrain MRI in alcohol dependence: preliminary evidence for effects of concurrent chroniccigarette smoking on regional brain volumes. Alcohol Clin Exp Res 29:1484–1495
Gazdzinski S, Durazzo T, Jahng GH, Ezekiel F, Banys P, Meyerhoff D (2006) Effects of chronicalcohol dependence and chronic cigarette smoking on cerebral perfusion: a preliminarymagnetic resonance study. Alcohol Clin Exp Res 30:947–958
Gazdzinski S, Durazzo TC, Weiner MW, Meyerhoff DJ (2008a) Are treated alcoholicsrepresentative of the entire population with alcohol use disorders? A magnetic resonancestudy of brain injury. Alcohol 42:67–76
Gazdzinski S, Durazzo TC, Yeh PH, Hardin D, Banys P, Meyerhoff DJ (2008b) Chronic cigarettesmoking modulates injury and short-term recovery of the medial temporal lobe in alcoholics.Psychiatry Res 162:133–145
Glass JM, Adams KM, Nigg JT, Wong MM, Puttler LI, Buu A, Jester JM, Fitzgerald HE, ZuckerRA (2006) Smoking is associated with neurocognitive deficits in alcoholism. Drug AlcoholDepend 82:119–126
Hansson AC, Nixon K, Rimondini R, Damadzic R, Sommer WH, Eskay R, Crews FT, Heilig M(2010) Long-term suppression of forebrain neurogenesis and loss of neuronal progenitor cellsfollowing prolonged alcohol dependence in rats. Int J Neuropsychopharmacol 13:583–593
Harding AJ, Wong A, Svoboda M, Kril JJ, Halliday GM (1997) Chronic alcohol consumptiondoes not cause hippocampal neuron loss in humans. Hippocampus 7:78–87
Harper C (1998) The neuropathology of alcohol-specific brain damage, or does alcohol damagethe brain? Journal of Neuropathology and Experimental Neurology 57:101–110
Harper C (2009) The neuropathology of alcohol-related brain damage. Alcohol Alcohol 44:136–140
Harper C, Kril J (1989) Patterns of neuronal loss in the cerebral cortex in chronic alcoholicpatients. J.Neurol.Sci. 92:81–89
Harper C, Dixon G, Sheedy D, Garrick T (2003) Neuropathological alterations in alcoholicbrains. Studies arising from the New South Wales Tissue Resource Centre. Progress in Neuro-Psychopharmacology and Biological Psychiatry 27:951–961
Harris BR, Gibson DA, Prendergast MA, Blanchard JA, Holley RC, Hart SR, Scotland RL, FosterTC, Pedigo NW, Littleton JM (2003) The neurotoxicity induced by ethanol withdrawal inmature organotypic hippocampal slices might involve cross-talk between metabotropicglutamate type 5 receptors and N-methyl-D-aspartate receptors. Alcohol Clin Exp Res27:1724–1735
Hasin DS, Stinson FS, Ogburn E, Grant BF (2007) Prevalence, correlates, disability, andcomorbidity of DSM-IV alcohol abuse and dependence in the United States: results from theNational Epidemiologic Survey on Alcohol and Related Conditions. Arch Gen Psychiatry64:830–842
Chronic Alcohol Consumption, Abstinence and Relapse

Hazell AS, Butterworth RF (1999) Hepatic encephalopathy: An update of pathophysiologicmechanisms. Proc Soc Exp Biol Med 222:99–112
He J, Overstreet DH, Crews FT (2009) Abstinence from moderate alcohol self-administrationalters progenitor cell proliferation and differentiation in multiple brain regions of male andfemale P rats. Alcohol Clin Exp Res 33:129–138
Hetherington HP, Newcomer BR, Pan JW (1998) Measurements of human cerebral GABA at 4.1T using numerically optimized editing pulses. Magn Reson Med 39:6–10
Hirakawa K, Uekusa K, Sato S, Nihira M (1994) MRI and MRS studies on acute effects ofethanol in the rat brain. Japanese Journal of Legal Medicine 48:63–74
Hoerst M, Weber-Fahr W, Tunc-Skarka N, Ruf M, Bohus M, Schmahl C, Ende G (2010)Glutamate levels in the anterior cingulate cortex correlate with self-reported impulsivity inpatients with borderline personality disorder and healthy controls. Arch Gen Psychiatry InPress
Hugg JW, Kuzniecky RI, Gilliam FG, Morawetz RB, Faught RE, Hetherington HP (1996)Normalization of contralateral metabolic function following temporal lobectomy demon-strated by h-1 magnetic resonance spectroscopic imaging. Ann.Neurol. V40:236–239
Jagannathan NR, Desai NG, Raghunathan P (1996) Brain metabolite changes in alcoholism: Anin vivo proton magnetic resonance spectroscopy (MRS) study. Magnetic Resonance Imaging14:553–557
Jansen JF, Backes WH, Nicolay K, Kooi ME (2006) 1H MR spectroscopy of the brain: absolutequantification of metabolites. Radiology 240:318–332
Jensen GB, Pakkenberg B (1993) Do alcoholics drink their neurons away? Lancet 342:1201–1204
John U, Meyer C, Rumpf HJ, Schumann A, Thyrian JR, Hapke U (2003) Strength of therelationship between tobacco smoking, nicotine dependence and the severity of alcoholdependence syndrome criteria in a population-based sample. Alcohol Alcohol 38:606–612
Kalivas PW, O’Brien C (2008) Drug addiction as a pathology of staged neuroplasticity.Neuropsychopharmacology 33:166–180
Kalivas PW, Volkow ND (2005) The neural basis of addiction: a pathology of motivation andchoice. Am J Psychiatry 162:1403–1413
Kalivas PW, Lalumiere RT, Knackstedt L, Shen H (2009) Glutamate transmission in addiction.Neuropharmacology 56(Suppl 1):169–173
Ke Y, Cohen BM, Lowen S, Hirashima F, Nassar L, Renshaw PF (2002) Biexponential transverserelaxation (T(2)) of the proton MRS creatine resonance in human brain. Magn Reson Med47:232–238
Korbo L (1999) Glial cell loss in the hippocampus of alcoholics. Alcohol Clin Exp Res 23:164–168
Kril JJ, Halliday GM (1999) Brain shrinkage in alcoholics: a decade on and what have welearned? Prog Neurobiol 58:381–387
Kril JJ, Harding AJ, Wong A, Svoboda MD, Halliday GM (1996) Is the hippocampus damaged inalcoholics with memory loss? Alcohol: Clin Exp Res 20:76A
Lee H, Holburn GH, Price RR (2003a) Proton MR spectroscopic studies of chronic alcoholexposure on the rat brain. J Magn Reson Imaging 18:147–151
Lee PL, Yiannoutsos CT, Ernst T, Chang L, Marra CM, Jarvik JG, Richards TL, Kwok EW,Kolson DL, Simpson D, Tang CY, Schifitto G, Ketonen LM, Meyerhoff DJ, Lenkinski RE,Gonzalez RG, Navia BA (2003b) A multi-center 1H MRS study of the AIDS dementiacomplex: Validation and preliminary analysis. J Magn Reson Imaging 17:625–633
Li BS, Babb JS, Soher BJ, Maudsley AA, Gonen O (2002) Reproducibility of 3D protonspectroscopy in the human brain. Magn Reson Med 47:439–446
Littleton J (1995) Acamprosate in alcohol dependence: how does it work? Addiction 90:1179–1188
Maisto SA, Connors GJ (2006) Relapse in the addictive behaviors: integration and futuredirections. Clin Psychol Rev 26:229–231
D. J. Meyerhoff et al.

Mann K, Batra A, Gunthner A, Schroth G (1992) Do women develop alcoholic brain damagemore readily than men? Alcohol Clin Exp Res, pp 1052–1056
Mansell D, Penk W, Hankin CS, Lee A, Spiro A 3rd, Skinner KM, Hsieh J, Kazis LE (2006) Theillness burden of alcohol-related disorders among VA patients: the veterans health study.J Ambul Care Manage 29:61–70
Martin PR, Gibbs SJ, Nimmerrichter AA, Riddle WR, Welch LW, Willcott MR (1995) Brainproton magnetic resonance spectroscopy studies in recently abstinent alcoholics. Alcohol ClinExp Res 19:1078–1082
Martinez-Bisbal MC, Arana E, Marti-Bonmati L, Martinez-Granados B, Celda B (2006)Cognitive impairment: classification by proton magnetic resonance spectroscopy and thecontributions of conventional magnetic resonance imaging]. Radiologia 48:301–307
Mason GF, Petrakis IL, de Graaf RA, Gueorguieva R, Guidone E, Coric V, Epperson CN,Rothman DL, Krystal JH (2006) Cortical gamma-aminobutyric acid levels and the recoveryfrom ethanol dependence: preliminary evidence of modification by cigarette smoking. BiolPsychiatry 59:85–93
Maudsley AA, Domenig C, Govind V, Darkazanli A, Studholme C, Arheart K, Bloomer C (2009)Mapping of brain metabolite distributions by volumetric proton MR spectroscopic imaging(MRSI). Magn Reson Med 61:548–559
McFarland K, Lapish CC, Kalivas PW (2003) Prefrontal glutamate release into the core of thenucleus accumbens mediates cocaine-induced reinstatement of drug-seeking behavior.J Neurosci 23:3531–3537
Mertens JR, Lu YW, Parthasarathy S, Moore C, Weisner CM (2003) Medical and psychiatricconditions of alcohol and drug treatment patients in an HMO: comparison with matchedcontrols. Arch Intern Med 163:2511–2517
Meyerhoff DJ, Durazzo TC (2008) Proton magnetic resonance spectroscopy in alcohol usedisorders: a potential new endophenotype? Alcohol Clin Exp Res 32:1146–1158
Meyerhoff D, Blumenfeld R, Truran D, Lindgren J, Flenniken D, Cardenas V, Chao LL, RothlindJ, Studholme C, Weiner H (2004) Effects of heavy drinking, binge drinking, and familyhistory of alcoholism on regional brain metabolites. Alcohol Clin Exp Res 28:650–661
Miguel-Hidalgo JJ, Wei J, Andrew M, Overholser JC, Jurjus G, Stockmeier CA, Rajkowska G(2002) Glia pathology in the prefrontal cortex in alcohol dependence with and withoutdepressive symptoms. Biol Psychiatry 52:1121–1133
Miguel-Hidalgo JJ, Overholser JC, Meltzer HY, Stockmeier CA, Rajkowska G (2006) Reducedglial and neuronal packing density in the orbitofrontal cortex in alcohol dependence and itsrelationship with suicide and duration of alcohol dependence. Alcohol Clin Exp Res 30:1845–1855
Miller BL, Chang L, Booth R, Ernst T, Cornford M, Nikas D, McBride D, Jenden DJ (1996) Invivo 1H MRS choline: correlation with in vitro chemistry/histology. Life Sciences 58:1929–1935
Modi S, Bhattacharya M, Kumar P, Deshpande SN, Tripathi RP, Khushu S (2009) Brainmetabolite changes in alcoholism: Localized proton magnetic resonance spectroscopy studyof the occipital lobe. Eur J Radiol
Moffett JR, Namboodiri MA, Cangro CB, Neale JH (1991) Immunohistochemical localization ofN-acetylaspartate in rat brain. Neuroreport 2:131–134
Mon A, Durazzo TC, Gazdzinski S, Meyerhoff DJ (2009) The Impact of Chronic CigaretteSmoking on Recovery From Cortical Gray Matter Perfusion Deficits in Alcohol Dependence:Longitudinal Arterial Spin Labeling MRI. Alcohol Clin Exp Res
Mon A, Durazzo TC, Meyerhoff DJ (2010) Metabolite levels by magnetic resonancespectroscopy during abstinence from alcohol. Alcohol Clin Exp Res 34(supplement):81A
Moss HB, Chen CM, Yi HY (2007) Subtypes of alcohol dependence in a nationallyrepresentative sample. Drug Alcohol Depend 91:149–158
Nicholas PC, Kim D, Crews FT, Macdonald JM (2008) 1H NMR-based metabolomic analysisof liver, serum, and brain following ethanol administration in rats. Chem Res Toxicol 21:408–420
Chronic Alcohol Consumption, Abstinence and Relapse

Ohrmann P, Kugel H, Bauer J, Siegmund A, Kolkebeck K, Suslow T, Wiedl KH, RothermundtM, Arolt V, Pedersen A (2008) Learning potential on the WCST in schizophrenia is related tothe neuronal integrity of the anterior cingulate cortex as measured by proton magneticresonance spectroscopy. Schizophr Res 106:156–163
Oscar-Berman M (2000) NIAAA Research Monograph No. 34: Neuropsychological vulnerabi-lites in chronic alcoholism Review of NIAAA’s Neuroscience and Behavioral ResearchPortfolio. NIAAA, Bethesda, MD, pp 437–472
Pan JW, Takahashi K (2005) Interdependence of N-acetyl aspartate and high-energy phosphatesin healthy human brain. Ann Neurol 57:92–97
Parks MH, Dawant BM, Riddle WR, Hartmann SL, Dietrich MS, Nickel MK, Price RR, MartinPR (2002) Longitudinal brain metabolic characterization of chronic alcoholics with protonmagnetic resonance spectroscopy. Alcohol Clin Exp Res 26:1368–1380
Parsons OA, Nixon SJ (1998) Cognitive functioning in sober social drinkers: a review of theresearch since 1986. J Stud Alcohol 59:180–190
Pettegrew JW, Kopp SJ, Minshew NJ, Glonek T, Feliksik JM, Tow JP, Cohen MM (1987) 31Pnuclear magnetic resonance studies of phosphoglyceride metabolism in developing anddegenerating brain: preliminary observations. J Neuropathol.Exp.Neurol 46:419–430
Pettegrew JW, Panchalingam K, Withers G, McKeag D, Strychor S (1990) Changes in brainenergy and phospholipid metabolism during development and aging in the Fischer 344 rat.J Neuropathol.Exp.Neurol 49:237–249
Pfefferbaum A, Sullivan EV, Mathalon DH, Shear PK, Rosenbloom MJ, Lim KO (1995)Longitudinal changes in magnetic resonance imaging brain volumes in abstinent and relapsedalcoholics. Alcohol Clin Exp Res 19:1177–1191
Pfefferbaum A, Adalsteinsson E, Bell RL, Sullivan EV (2007) Development and resolution ofbrain lesions caused by pyrithiamine- and dietary-induced thiamine deficiency and alcoholexposure in the alcohol-preferring rat: a longitudinal magnetic resonance imaging andspectroscopy study. Neuropsychopharmacology 32:1159–1177
Pouwels PJ, Frahm J (1998) Regional metabolite concentrations in human brain as determined byquantitative localized proton MRS. Magn Reson Med 39:53–60
Prendergast MA, Harris BR, Mayer S, Littleton JM (2000) Chronic, but not acute, nicotineexposure attenuates ethanol withdrawal-induced hippocampal damage in vitro. Alcohol ClinExp Res 24:1583–1592
Rajkowska G, Selemon LD, Goldman-Rakic PS (1998) Neuronal and glial somal size in theprefrontal cortex: a postmortem morphometric study of schizophrenia and Huntington disease.Arch Gen Psychiatry 55:215–224
Riederer F, Bittsansky M, Lehner-Baumgartner E, Baumgartner C, Mlynarik V, Gruber S, MoserE, Kaya M, Serles W (2007) Decrease of NAA with aging outside the seizure focus in mesialtemporal lobe epilepsy–a proton-MRS study at 3 Tesla. Brain Res 1179:131–139
Romberger DJ, Grant K (2004) Alcohol consumption and smoking status: the role of smokingcessation. Biomed Pharmacother 58:77–83
Room R (2004) Smoking and drinking as complementary behaviours. Biomed Pharmacother58:111–115
Rosen Y, Lenkinski RE (2007) Recent advances in magnetic resonance neurospectroscopy.Neurotherapeutics 4:330–345
Ross B, Bluml S (2001) Magnetic resonance spectroscopy of the human brain. Anat Rec 265:54–84
Ross B, Michaelis T (1994) Clinical applications of magnetic resonance spectroscopy.Magn.Reson.Q. 10:191–247
Ross AJ, Sachdev PS (2004) Magnetic resonance spectroscopy in cognitive research. Brain ResBrain Res Rev 44:83–102
Rothman DL, Petroff OA, Behar KL, Mattson RH (1993a) Localized 1H NMR measurements ofgamma-aminobutyric acid in human brain in vivo. Proceedings of the National Academy ofScience, U.S.A. 90:5662–5666
D. J. Meyerhoff et al.

Rothman DL, Petroff OAC, Behar KL, Matson RH (1993b) Localized 1H NMR measurements ofgamma-aminobutyric acid in human brain in vivo. Proc Natl Acad Sci USA 90:5662–5666
Rothman DL, Behar KL, Hyder F, Shulman RG (2003) In vivo NMR studies of the glutamateneurotransmitter flux and neuroenergetics: implications for brain function. Annu Rev Physiol65:401–427
Sartorius A, Lugenbiel P, Mahlstedt MM, Ende G, Schloss P, Vollmayr B (2008) Protonmagnetic resonance spectroscopic creatine correlates with creatine transporter protein densityin rat brain. J Neurosci Methods 172:215–219
Schuff N, Ezekiel F, Gamst AC, Amend DL, Capizzano AA, Maudsley AA, Weiner MW (2001)Region and tissue differences of metabolites in normally aged brain using multislice 1Hmagnetic resonance spectroscopic imaging. Magn Reson Med 45:899–907
Schuff N, Meyerhoff DJ, Mueller S, Chao L, Sacrey DT, Laxer K, Weiner MW (2006) N-acetylaspartate as a marker of neuronal injury in neurodegenerative disease. Adv Exp MedBiol 576:241–262 discussion 361-3
Schwab ME, Bartholdi D (1996) Degeneration and regeneration of axons in the lesioned spinalcord. Physiol Rev 76:319–370
Schweinsburg BC, Taylor MJ, Videen JS, Alhassoon OM, Patterson TL, Grant I (2000) Elevatedmyo-inositol in gray matter of recently detoxified but not long-term alcoholics: A preliminaryMR spectroscopy study. Alc Clin Exp Res. 24:699–770
Schweinsburg BC, Taylor MJ, Alhassoon OM, Videen JS, Brown GG, Patterson TL, Berger F,Grant I (2001) Chemical pathology in brain white matter of recently detoxified alcoholics: a1H magnetic resonance spectroscopy investigation of alcohol-associated frontal lobe injury.Alcohol Clin Exp Res 25:924–934
Seitz D, Widmann U, Seeger U, Nagele T, Klose U, Mann K, Grodd W (1999) Localized protonmagnetic resonance spectroscopy of the cerebellum in detoxifing alcoholics. Alcohol ClinExp Res 23:158–163
Selemon LD, Goldman-Rakic PS (1999) The reduced neuropil hypothesis: a circuit based modelof schizophrenia. Biol Psychiatry 45:17–25
Simmons ML, Frondoza CG, Coyle JT (1991) Immunocytochemical localization of N-acetyl-aspartate with monoclonal antibodies. Neuroscience 45:37–45
Spanagel R (2009) Alcoholism: a systems approach from molecular physiology to addictivebehavior. Physiol Rev 89(2):649–705
Steen RG, Hamer RM, Lieberman JA (2005) Measurement of brain metabolites by 1H magneticresonance spectroscopy in patients with schizophrenia: a systematic review and meta-analysis.Neuropsychopharmacology 30:1949–1962
Stinson FS, Grant BF, Dawson DA, Ruan WJ, Huang B, Saha T (2005) Comorbidity betweenDSM-IV alcohol and specific drug use disorders in the United States: results from theNational Epidemiologic Survey on Alcohol and Related Conditions. Drug Alcohol Depend80:105–116
Sullivan EV (2000) NIAAA Research Monograph No. 34: Human brain vulnerability toalcoholism: Evidence from neuroimaging studies. In: Noronha A, Eckardt M, Warren K (eds)Review of NIAAA’s neuroscience and behavioral research portfolio. National Institute onAlcohol Abuse and Alcoholism, Bethesda, MD, pp 473-508
Sullivan EV, Pfefferbaum A (2005) Neurocircuitry in alcoholism: a substrate of disruption andrepair. Psychopharmacology (Berl) 180:583–594
Sullivan EV, Rosenbloom M, Serventi KL, Pfefferbaum A (2004) Effects of age and sex onvolumes of the thalamus, pons, and cortex. Neurobiol Aging 25:185–192
Thomson AD, Marshall EJ (2006) The natural history and pathophysiology of Wernicke’sEncephalopathy and Korsakoff’s Psychosis. Alcohol Alcohol 41:151–158
Umhau JC, Momenan R, Schwandt ML, Singley E, Lifshitz M, Doty L, Adams LJ, Vengeliene V,Spanagel R, Zhang Y, Shen, George DT, Hommer D, Heilig M (2010) Effect of Acamprosateon Magnetic Resonance Spectroscopy Measures of Central Glutamate in Detoxified Alcohol-Dependent Individuals: A Randomized Controlled Experimental Medicine Study. Arch GenPsychiatry 67:1069-1077
Chronic Alcohol Consumption, Abstinence and Relapse

Vion-Dury J, Meyerhoff DJ, Cozzone PJ, Weiner MW (1994) What might be the impact onneurology of the analysis of brain metabolism by in vivo magnetic resonance spectroscopy?[editorial]. Journal of Neurology 241:354–371
Weber-Fahr W, Ende G, Sartorius A, Spanagel R, Falfan-Melgoza C, Cleppien D, Sommer W(2010) 1H MRS profiling at 9.4T in prefrontal cortex and hippocampus of ethanol dependentrats during intoxication, withdrawal and protracted abstinence ISMRM, Stockholm, Sweden,pp Prog #: 2403
Xin L, Mlynarik V, Lei H, Gruetter R (2010) Influence of regional macromolecule baseline on thequantification of neurochemical profile in rat brain ISMRM, Stockholm, Sweden, pp Prog #:321
Yildiz-Yesiloglu A, Ankerst DP (2006a) Neurochemical alterations of the brain in bipolardisorder and their implications for pathophysiology: a systematic review of the in vivo protonmagnetic resonance spectroscopy findings. Prog Neuropsychopharmacol Biol Psychiatry30:969–995
Yildiz-Yesiloglu A, Ankerst DP (2006b) Review of 1H magnetic resonance spectroscopy findingsin major depressive disorder: a meta-analysis. Psychiatry Res 147:1–25
Zahr NM, Sullivan EV (2008) Translational studies of alcoholism: bridging the gap. Alcohol ResHealth 31:215–230
Zahr NM, Mayer D, Pfefferbaum A, Sullivan EV (2008) Low striatal glutamate levels underliecognitive decline in the elderly: evidence from in vivo molecular spectroscopy. Cereb Cortex18:2241–2250
Zahr NM, Mayer D, Vinco S, Orduna J, Luong R, Sullivan EV, Pfefferbaum A (2009) In vivoevidence for alcohol-induced neurochemical changes in rat brain without protractedwithdrawal, pronounced thiamine deficiency, or severe liver damage. Neuropsychopharma-cology 34:1427–1442
Zahr NM, Mayer D, Rohlfing T, Hasak MP, Hsu O, Vinco S, Orduna J, Luong R, Sullivan EV,Pfefferbaum A (2010) Brain injury and recovery following binge ethanol: evidence from invivo magnetic resonance spectroscopy. Biol Psychiatry 67:846–854
Zhu PJ, Chiappinelli VA (1999) Nicotine modulates evoked GABAergic transmission in thebrain. J Neurophysiol 82:3041–3045
Zywiak WH, Stout RL, Trefry WB, Glasser I, Connors GJ, Maisto SA, Westerberg VS (2006)Alcohol relapse repetition, gender, and predictive validity. J Subst Abuse Treat 30:349–353
D. J. Meyerhoff et al.
Related Documents











