Chiral front propagation in liquid-crystalline materials: Formation of the planar monodomain twisted plywood architecture of biological fibrous composites Gino De Luca and Alejandro D. Rey* Department of Chemical Engineering, McGill University, 3610 University Street, Montreal, Quebec, Canada H3A 2B2 ~Received 6 November 2002; published 30 January 2004! Biological fibrous composites commonly exhibit an architecture known as twisted plywood, which is similar to that of the cholesteric liquid-crystalline mesophases. The explanation for the structural similarity is that biological fibrous composites adopt a lyotropic cholesteric liquid-crystalline phase during their formation process. In this work, a mathematical model based on the Landau–de Gennes theory of liquid crystals has been developed to reproduce the process by which long chiral fibrous molecules form the twisted plywood structures observed in biological composites. The dynamics of the process was then further investigated by analytically solving a simplified version of the governing equations. Results obtained from the model are in good qualita- tive agreement with the theory of Neville @Biology of Fibrous Composites ~Cambridge University Press, Cambridge, England, 1993!# who hypothesized the necessity of a constraining layer to lock the direction of the helical axis of the plywood in order to create a monodomain structure. Computational results indicate that the plywood architecture is obtained by a chiral front propagation process with a fully relaxed wake. The effects of chirality and concentration on the formation process kinetics are characterized. DOI: 10.1103/PhysRevE.69.011706 PACS number~s!: 61.30.2v, 83.80.Xz, 61.30.Dk, 61.30.St I. INTRODUCTION Skeletal and protective extracellular tissues of biological systems are highly organized composite materials @1–7#. They exhibit remarkable mechanical properties but are built from relatively simple constituents such as cellulose, chitin, and collagen @1,7#. In addition, these composite materials are biodegradable and assembled at ambient temperature and pressure in an aqueous environment. Biological composites have therefore undeniable advantages over synthetic materi- als and are of a growing interest in applied material science. A major challenge in the field is to produce synthetic equiva- lents of these composite materials. However, to develop a synthetic route, the structural formation process of biological composites needs to be precisely described and understood @6#. Strategies developed by material engineers to solve me- chanical problems are often similar to the ones employed by living organisms and even the lower ones @8#. Natural com- posites exhibit laminated architectures called plywoods @1#. The laminated architecture, the most widely found in nature, is the twisted plywood, also referred as helicoidal plywood @1–7#. The plywood architecture, found in nearly all regular extracellular assemblies of living systems, is well docu- mented in the literature @1–7#. But how are fibrous macromolecules precisely manipu- lated into the extracellular matrix so as to form the twisted plywood assemblies found in biological composites? The most probable answer is that the extracellular matrix, which surrounds the fibrous water-insoluble molecules, passes through a mobile phase during its development and self- assembly @1–7#. In thermodynamic terms, self-assembly is a free energy minimization process driven by entropy. The free energy of the system is lowered as the excluded volume be- tween the molecules is reduced @8#. Because most naturally occurring biological polymers are chiral—i.e., they are not superimposable on their mirror image—the excluded volume is favorably reduced by twisted/helical packing. Knowing that natural composites are usually compacted, partially ordered, and required to pass via a more or less fluid state, it is likely that a lyotropic liquid-crystalline phase is involved during their supramolecular self-assembly. Lyotro- pic liquid-crystalline phases form three-dimensional assem- blies, which combine the fluidity of liquids with the long- range orientational order of crystals, above a certain critical concentration of molecules in the solution @9#. Among the different types of liquid-crystalline phases found in nature, cholesteric mesophases demonstrate the greatest structural similarities with the architecture of biological composites. Cholesteric mesophases are three-dimensional assemblies whose molecules lie on a series of equidistant pseudoplanes that are slightly rotated with respect to one another @9#. These mesophases, which are made up of optically active molecules, are characterized by a length scale known as the pitch, p 0 , corresponding to the distance required by the mol- ecules to accomplish a 2p radians rotation of their long axis. Figure 1 shows a schematic of the plywood architecture adopted by biological composites ~adapted from @3#!. The fiberlike constituents display the classical cholesteric or chi- ral nematic spatial organization defined, in a rectangular ( x , y , z ) coordinate system, by n x 5cos u ~ y ! , n y 50, n z 52sin u ~ y ! , ~1a! with u ~ y ! 5 2 p p 0 y , N5j, ~1b! *Corresponding author. FAX: ~514! 398-6678. Electronic address: [email protected] PHYSICAL REVIEW E 69, 011706 ~2004! 1063-651X/2004/69~1!/011706~13!/$22.50 ©2004 The American Physical Society 69 011706-1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

PHYSICAL REVIEW E 69, 011706 ~2004!
Chiral front propagation in liquid-crystalline materials: Formation of the planar monodomaintwisted plywood architecture of biological fibrous composites
Gino De Luca and Alejandro D. Rey*Department of Chemical Engineering, McGill University, 3610 University Street, Montreal, Quebec, Canada H3A 2B2
~Received 6 November 2002; published 30 January 2004!
Biological fibrous composites commonly exhibit an architecture known as twisted plywood, which is similarto that of the cholesteric liquid-crystalline mesophases. The explanation for the structural similarity is thatbiological fibrous composites adopt a lyotropic cholesteric liquid-crystalline phase during their formationprocess. In this work, a mathematical model based on the Landau–de Gennes theory of liquid crystals has beendeveloped to reproduce the process by which long chiral fibrous molecules form the twisted plywood structuresobserved in biological composites. The dynamics of the process was then further investigated by analyticallysolving a simplified version of the governing equations. Results obtained from the model are in good qualita-tive agreement with the theory of Neville@Biology of Fibrous Composites~Cambridge University Press,Cambridge, England, 1993!# who hypothesized the necessity of a constraining layer to lock the direction of thehelical axis of the plywood in order to create a monodomain structure. Computational results indicate that theplywood architecture is obtained by a chiral front propagation process with a fully relaxed wake. The effectsof chirality and concentration on the formation process kinetics are characterized.
DOI: 10.1103/PhysRevE.69.011706 PACS number~s!: 61.30.2v, 83.80.Xz, 61.30.Dk, 61.30.St
ca
utinrea
ittec
vapcato
mb
reodaru
puedhicsela
reebe-
otme
ed,uidisro-m--ical
re,cturals.bliesnes
tivethe
ol-.ure
hi-lar
:
I. INTRODUCTION
Skeletal and protective extracellular tissues of biologisystems are highly organized composite materials@1–7#.They exhibit remarkable mechanical properties but are bfrom relatively simple constituents such as cellulose, chiand collagen@1,7#. In addition, these composite materials abiodegradable and assembled at ambient temperaturepressure in an aqueous environment. Biological composhave therefore undeniable advantages over synthetic maals and are of a growing interest in applied material scienA major challenge in the field is to produce synthetic equilents of these composite materials. However, to develosynthetic route, the structural formation process of biologicomposites needs to be precisely described and unders@6#.
Strategies developed by material engineers to solvechanical problems are often similar to the ones employedliving organisms and even the lower ones@8#. Natural com-posites exhibit laminated architectures called plywoods@1#.The laminated architecture, the most widely found in natuis the twisted plywood, also referred as helicoidal plywo@1–7#. The plywood architecture, found in nearly all regulextracellular assemblies of living systems, is well docmented in the literature@1–7#.
But how are fibrous macromolecules precisely manilated into the extracellular matrix so as to form the twistplywood assemblies found in biological composites? Tmost probable answer is that the extracellular matrix, whsurrounds the fibrous water-insoluble molecules, pasthrough a mobile phase during its development and sassembly@1–7#. In thermodynamic terms, self-assembly is
*Corresponding author. FAX:~514! 398-6678. Electronic [email protected]
1063-651X/2004/69~1!/011706~13!/$22.50 69 0117
l
ilt,
ndesri-
e.-alod
e-y
,
-
-
ehesf-
free energy minimization process driven by entropy. The fenergy of the system is lowered as the excluded volumetween the molecules is reduced@8#. Because most naturallyoccurring biological polymers are chiral—i.e., they are nsuperimposable on their mirror image—the excluded voluis favorably reduced by twisted/helical packing.
Knowing that natural composites are usually compactpartially ordered, and required to pass via a more or less flstate, it is likely that a lyotropic liquid-crystalline phaseinvolved during their supramolecular self-assembly. Lyotpic liquid-crystalline phases form three-dimensional asseblies, which combine the fluidity of liquids with the longrange orientational order of crystals, above a certain critconcentration of molecules in the solution@9#. Among thedifferent types of liquid-crystalline phases found in natucholesteric mesophases demonstrate the greatest strusimilarities with the architecture of biological compositeCholesteric mesophases are three-dimensional assemwhose molecules lie on a series of equidistant pseudoplathat are slightly rotated with respect to one another@9#.These mesophases, which are made up of optically acmolecules, are characterized by a length scale known aspitch, p0 , corresponding to the distance required by the mecules to accomplish a 2p radians rotation of their long axisFigure 1 shows a schematic of the plywood architectadopted by biological composites~adapted from@3#!. Thefiberlike constituents display the classical cholesteric or cral nematic spatial organization defined, in a rectangu(x,y,z) coordinate system, by
nx5cosu~y!, ny50, nz52sinu~y!, ~1a!
with
u~y!52p
p0y, N5 j , ~1b!
©2004 The American Physical Society06-1

ors
yice
to
idsex
eeostethtaoalhi
tteve
e-
l
sye
reic
s oftoid-
eed,ssict
e-nes of
odalthehi-clu-
nel or-ta-m-
do-am-
es
es of
eter
e of
hlle
on
nt;df
G. De LUCA AND A. D. REY PHYSICAL REVIEW E69, 011706 ~2004!
wheren is the unit vector~director! of the fibers,u is theangle between the director and thex axis, p0 is the pitch,Nis the unit helix vector, andj is the unit vector in theydirection. Figure 1~b! presents a schematic of the directfield along a plane containingN and shows that the fiberrotate continuously along thez direction. Figure 1~c! showsthat an oblique cut of the plywood architecture displaarced patterns. The arced patterns are common in biologcomposites and have been thoroughly characterized by Nille @1#. It is well known that such arced patterns are likelyarise from a cholestericlike organization@1–7#.
Despite their similar architectures, cholesteric liqucrystalline phases and biological fibrous composites prean essential difference. While liquid-crystalline materials eist as a distinct state of matter which is intermediate betwa liquid and a solid state, mature biological fibrous compites are in a solid state. The liquid-crystalline characwhich is hypothesized to exist during the first steps ofpolymer secretion by the cells, is transformed in a solid sby molecular cross-links. However, the imprint of the chlesteric liquid-crystalline phase in the mineralized materiis testified to by the properties of the twisted plywood arctecture in electron and polarized-light microscopy@1–5#.Liquid-crystalline theories developed in condensed-maphysics are consequently helpful to advance structural deopment knowledge for biological fibrous composites.
The mechanical reliability of biological composites rsides in their highly hierarchical structure~e.g., molecules,macromolecules, microfibrils, fibrils, fibers, etc.!, but also intheir twisted plywood architecture@1,4,6,7#. One essentiafeature of these structures is that they are monodomains@1#.The quasiabsence of defects found in most compositebelieved@1# to be due to the presence of a constraining lathat fixes the rotation axis~N! of the plywood architecturealong the surface unit normal~j !. Thus knowledge of theconditions under which monodomain defect-free structuare generated is essential to the understanding of biologcomposites.
FIG. 1. Schematic representation of the twisted plywood arctecture.~a! Parallel and equidistant layers. On each layer, paralines indicate the orientation of the fibers. The fibrillar directirotates by a small constant angle from one layer to the next.~b!Transverse section plane; the fibers appear as dots or segmedifferent length, all parallel to each other.~c! Oblique section planethe fibers appear as superposed series of nested arcs. The perioseen in these various planes corresponds to a 180° rotation ofibers. Adapted from@3#.
01170
salv-
-nt-n-
r,ete-s-
rl-
isr
sal
Based on these considerations, the specific objectivethis paper are~i! to simulate the ordering process leadingthe twisted plywood architecture found in cholesteric liqucrystalline phases and biological fibrous composites,~ii ! toidentify the conditions under which the stable defect-frmonodomain helicoidal plywood architecture is obtaineand~iii ! to characterize the kinetics of the formation proceof a monodomain cholesteric liquid crystal and thus predthose of a twisted biological fibrous composite.
This paper is organized as follows. Section II briefly prsents the invoked theoretical framework on liquid-crystallimesophases and the derivation of the governing equationthe model describing the formation of the twisted plywoarchitecture. Section III is devoted to the computationmethods used to solve the model. Section IV presentscomputational and analytical results on the plywood arctecture formation process. Section V presents the consions.
II. THEORY AND GOVERNING EQUATIONS
A. Description of the long-range orientational order
As mentioned above, the structure of liquid-crystallimesophases is characterized by a long-range orientationader of their constituent molecules. This long-range orientional order is commonly described by a second-order symetric traceless tensorQ corresponding to the seconmoment of the orientation distribution function. This macrscopic quantity, usually referred as the tensor order pareter, reads@9#
Q5SS nn2d
3D11
3P~mm2 l"l!, ~2a!
where the following restrictions apply:
Q5QT, ~2b!
tr~Q!50, ~2c!
21
2<S<1, ~2d!
23
2<P<
3
2, ~2e!
n"n5m"m5 ll51, ~2f!
nn1mm1 ll5d. ~2g!
The unit vectorsn, m, andl presented in Eqs.~2a!, ~2f!, and~2g! form an orthogonal director triad which characterizthe orientation of the phase. The unit vectorn is known asthe uniaxial director, andm and l as biaxial directors. ThequantitiesS and P, which are, respectively, known as thuniaxial and biaxial scalar order parameters, are measurethe molecular alignment. The uniaxial scalar order paramgives the degree of alignment along the uniaxial directorn,while the biaxial scalar order parameter gives the degre
i-l
s of
icitythe
6-2

eouinb
t
eiee-rnn
tete
sernriah
tlng
therthe
atin-
-ter
e
tion
ss
ngeref-
nds
pla-ine
reents-
lta-(0
CHIRAL FRONT PROPAGATION IN LIQUID- . . . PHYSICAL REVIEW E 69, 011706 ~2004!
alignment along the first biaxial directorm. The scalar orderparameters and the tensor order parameterQ ~main frame!are given by
S53
2~n"Q"n!, ~3a!
P53
2~m"Q"m2 l"Q"l!, ~3b!
Q5F 21
3~S2P! 0 0
0 21
3~S1P! 0
0 02
3S
G . ~3c!
The tensor order parameterQ characterizes thoroughly thmicrostructure of the phase by combining information aborientation and alignment. Its quadrupolar symmetry retathe usual head-tail invariance that implies equivalencetween the director fieldn(r ) and 2n(r ). According to Eq.~3! the correspondences between phase and alignmenisotropic (S50,P50), uniaxial (SÞ0,P50), and biaxial(SÞ0,PÞ0).
B. Landau–de Gennes theory of liquid-crystalline materials
The Landau–de Gennes theory expresses the free endensity of the liquid-crystalline material as a power serexpansion of scalar invariants of the tensor order paramQ and its gradients¹Q @9# representing short- and longrange elastic effects, respectively. In the absence of extefields, the total free energy density of the mesophase cagiven in the following dimensionless form@9,10#:
f 5 f is1 f sr1 f lr , ~4a!
f sr51
2 S 12U
3 D tr~Q2!2U
3tr~Q3!1
U
4@ tr~Q2!#2, ~4b!
f lr51
2 H F S j
h0D ~¹3Q!14pS j
p0DQG2
1yF S j
h0D ~¹•Q!G2J ,
~4c!
where f is is the free energy density of the isotropic stawhich depends on conventional thermodynamic paramesuch as temperature, pressure, and concentration.f sr and f lrare, respectively, the short- and long-range contributionthe total free energy densityf . The dimensionless parametU is a thermodynamic potential proportional to the dimesionless concentration of rodlike molecules in the matewhich drives the isotropic-cholesteric phase transition. Tthermodynamic potentialU is related to the concentrationCby the relationU53C/C* , whereC* is the concentration athe phase transition. The parameterj is a coherence/internalength that gives the distance over which variations of lo
01170
ts
e-
are
rgyster
albe
rs,
to
-le
-
range orientational order can occur. This length furnishesorder of magnitude for the size of a defect. The parameteh0is an external/geometric length that gives the thickness ofmaterial. The parameterp0 is the pitch of the cholestericliquid-crystalline material. It is essential to recognize ththis model is therefore of a mesoscopic nature since itcludes a molecular~j! length scale and a macroscopic (h0)length scale. The remaining parametery represents a measure of the elastic anisotropy of the material. This parameis constrained to be greater than21/2 in order to ensure thepositivity and thermodynamic stability of the long-rangcontribution to the free energy.
The time evolution of the tensor order parameterQ isgiven by a standard dimensionless gradient flow equa@11,12#:
2g~Q!]Q
]t5
d f
dQ5F ] f
]Q2¹•
] f
]¹QG [s]
, ~5a!
g~Q!5S 123
2tr~Q2! D 2
, ~5b!
where the superscript@s# denotes symmetric and traceletensors, and whereg~Q! is a rotational viscosity. SubstitutingEqs. ~5a! and ~5b! into Eqs.~4a!, ~4b!, and ~4c! yields thedimensionless governing equationQ5Q(r ,t) for the timeevolution of the tensor order parameter:
2g~Q!]Q
]t5SR1LR, ~6a!
SR5S 12U
3 DQ2U~Q"Q! [s]1U~Q:Q!Q, ~6b!
LR52S j
h0D 2
$¹2Q2@¹•~¹Q!T# [s]1y@¹•~¹•Q!# [s]%
1S j
h0D S j
p0D @28p~¹3Q!# [s]1S j
p0D 2
@216p2Q#.
~6c!
The tensors SR and LR represent the short- and long-racontributions to the dynamics ofQ. The coupling parametej/h0 controls the balance between short- and long-rangefects while j/p0 controls the balance between chiral aachiral effects. In the limitj/p0→0, the material describean achiral ordinary nematic liquid-crystalline material.
III. COMPUTATIONAL MODELING
In order to understand the process which leads to thenar monodomain twisted plywood architecture observedthe broad majority of biological fibrous composites, wsimulate the time evolution of the tensor order parameteQin a lyotropic cholesteric liquid-crystalline material. Sincthe tensor order parameter has five independent compon~symmetric and traceless!, five coupled time-dependent nonlinear partial differential equations need to be solve simuneously. The computational domain is the unit square<x<1,0<y<1), and henceQ5Q(x,y,t). We emphasize
6-3

d
omd
pe
fa
-lere
e
tao
aousioele
ee
in
icaltheofndge-etialed
thea-n
me-in
c-ityicof
theof
cap-ions
ca-
g
theso-
d
el-
sys-m-
G. De LUCA AND A. D. REY PHYSICAL REVIEW E69, 011706 ~2004!
that, despite the fact that the simulation domain is twomensional~2D!, Q conserves its five degrees of freedom.
A. Boundary conditions
In order to restrict the influence of physical boundariesthe material bulk, periodic boundary conditions are eployed on thex direction. The boundary conditions adopteon the remaining bounding surfaces are of the Dirichlet tyThe upper boundary of the computational domain (y51)describes an isotropic state in order to represent the surof a secreting cell, as described in@1#, andQ is defined byEq. ~2a!. The lower boundary (y50) corresponds to a crystallized ordered layer. Along this boundary, the molecudescribe a strong planar anchoring and accordingly the ditor triad is given by
n~y50!5~1 0 0!, ~7a!
m~y50!5~0 1 0!, ~7b!
l~y50!5~0 0 1!. ~7c!
The alignment of the molecules along these particular dirtions is given by the equilibrium scalar order parameterSeqand Peq. These values are determined by the steady-ssolutions of the following autonomous nonlinear systemcoupled differential equations:
]S
]t5
3
2 S n•]Q
]t•nD
5S 22
9P2S2
1
9P21
1
3S22
2
3S31
1
3SD
3U2S24p2S j
p0D 2
~S2P!, ~8a!
]P
]t5
3
2 S m•
]Q
]t•m2 l•
]Q
]t• lD
5S 22
3S2P2
2
3PS2
2
9P31
1
3PD
3U2P112p2S j
p0D 2
~S2P!. ~8b!
Note that we have]n/]t5]m/]t5] l/]t50.
B. Initial conditions
Initially, the liquid-crystalline material is taken to be instable isotropic state. This disordered state corresponds tphysical situation of a low concentration of chiral fibromolecules in the surrounding extracellular matrix of the blogical composite. The initial tensor order parameter fiQ05Q(t50) characterizing the initial microstructure of thmaterial is defined in the following way:
Q05S0S e1e12d
3D11
3P0~e2e22e3e3!, ~9!
01170
i-
n-
.
ce
sc-
c-
tef
the
-d
whereS0 and P0 are random numbers between21310210
and 1310210 representing thermal fluctuations in the degrof order. The random orthogonal unit vectorse1 , e2 , ande3
describe the initial isotropic orientation of the moleculesthe material.
C. Numerical methods
The governing equations are solved using the classnumerical method of lines which consists of transformingset of governing partial differential equations into a setordinary differential equations by spatial discretization asubsequently integrating with respect to time. Since theometry of the computational domain is trivial, we ussecond-order finite difference approximations for the spaderivatives. The mesh refinement technique was performto confirm the invariance of the results with respect togrid spacing. Time integration was performed with a RungKutta-Chebyshev algorithm. This explicit method, with aextended domain of stability, possesses an adaptive tistepping scheme that captures the physics containedabrupt changes of the tensor order parameter@13#.
IV. RESULTS AND DISCUSSIONS
A. Phase-plane analysis of cholesteric order
Ordinary nematic liquid-crystalline materials are charaterized by their uniaxiality, whose amplitude and intensare directly related to the value of the thermodynampotential—i.e., temperature or concentration. In the casecholesteric mesophases~chiral nematics!, the situation issomewhat more complicated since the symmetry ofstructure is a function of the helical pitch. The importancethis length scale on the symmetry of the mesophase istured by a phase-plane analysis of the system of equat~8a! and ~8b!.
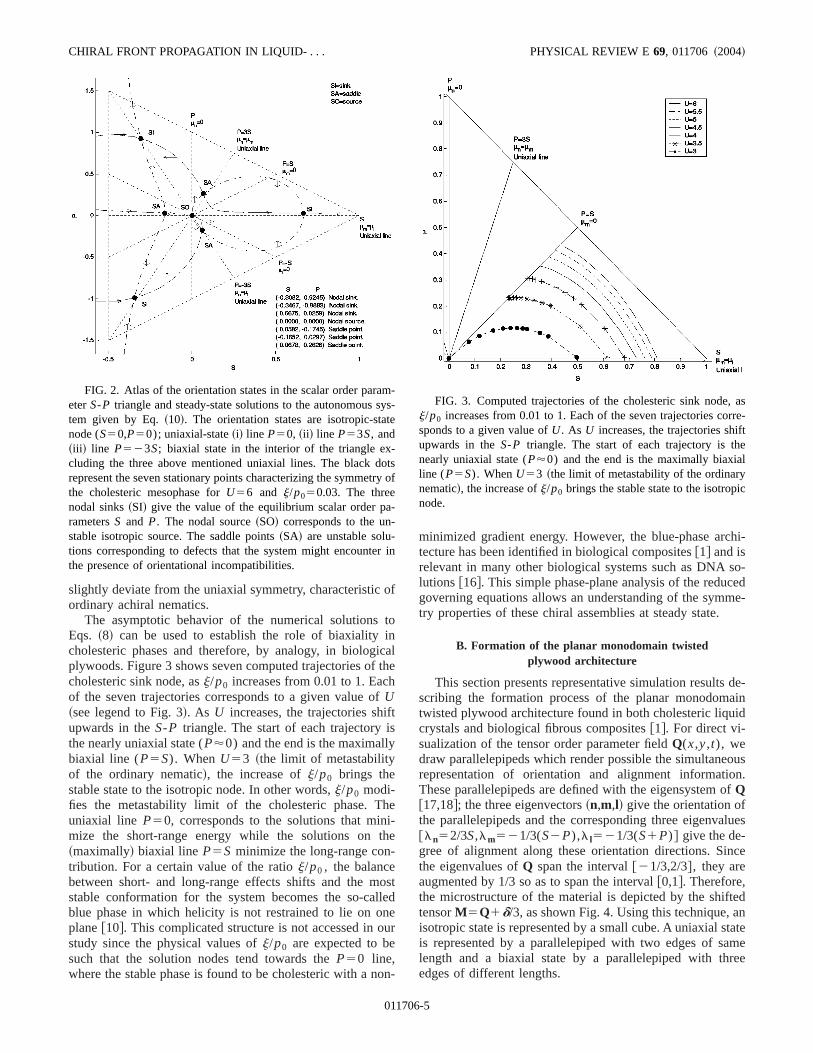
Figure 2 exhibits all possible orientation states in the slar order parameterS-P triangle for a given parametricchoice. The states described are as follows@14#: ~a! isotro-pic state~the three eigenvalues ofQ are identical!: S50 andP50; ~b! uniaxial state~two eigenvalues ofQ are identical!:~i! line P50, ~ii ! line P53S, and~iii ! line P523S; and~c!biaxial state~the three eigenvalues ofQ are distinct!: SÞ0andPÞ0, interior of the order parameter triangle excludinthe three uniaxial lines.
In addition to this atlas of ordering states, Fig. 2 givesseven stationary points corresponding to the steady-statelutions for the autonomous system given by Eqs.~8a! and~8b! for U56 andj/p050.03 ~the pitchp0 is 33.33 timeslonger than the coherence length!. The dashed lines indicatenullclines. The three nodal sinks~SI! give the value of theequilibrium scalar order parametersS andP. In simulations,the system always picks up the attractor located on thS.0 part of the plane, since uniaxiality prevails over biaxiaity in this system. The nodal source~SO! corresponds to theunstable isotropic source. The remaining saddle points~SA!are unstable solutions corresponding to defects that thetem might encounter in the presence of orientational incopatibilities @15#. The chirality (j/p0) forces the system to
6-4
o
toinicthhfiftisy
hi-th-
olleneou
o
chi-
so-ed
me-
de-ainid
ousn.f
lues
nce
tedantateme
ree
amsyte
-o
try
a--
r
asrre-ftel
ic
CHIRAL FRONT PROPAGATION IN LIQUID- . . . PHYSICAL REVIEW E 69, 011706 ~2004!
slightly deviate from the uniaxial symmetry, characteristicordinary achiral nematics.
The asymptotic behavior of the numerical solutionsEqs. ~8! can be used to establish the role of biaxialitycholesteric phases and therefore, by analogy, in biologplywoods. Figure 3 shows seven computed trajectories ofcholesteric sink node, asj/p0 increases from 0.01 to 1. Eacof the seven trajectories corresponds to a given value oU~see legend to Fig. 3!. As U increases, the trajectories shupwards in theS-P triangle. The start of each trajectorythe nearly uniaxial state (P'0) and the end is the maximallbiaxial line (P5S). WhenU53 ~the limit of metastabilityof the ordinary nematic!, the increase ofj/p0 brings thestable state to the isotropic node. In other words,j/p0 modi-fies the metastability limit of the cholesteric phase. Tuniaxial line P50, corresponds to the solutions that minmize the short-range energy while the solutions on~maximally! biaxial line P5S minimize the long-range contribution. For a certain value of the ratioj/p0 , the balancebetween short- and long-range effects shifts and the mstable conformation for the system becomes the so-cablue phase in which helicity is not restrained to lie on oplane@10#. This complicated structure is not accessed instudy since the physical values ofj/p0 are expected to besuch that the solution nodes tend towards theP50 line,where the stable phase is found to be cholesteric with a n
FIG. 2. Atlas of the orientation states in the scalar order pareterS-P triangle and steady-state solutions to the autonomoustem given by Eq.~10!. The orientation states are isotropic-stanode (S50,P50); uniaxial-state~i! line P50, ~ii ! line P53S, and~iii ! line P523S; biaxial state in the interior of the triangle excluding the three above mentioned uniaxial lines. The black drepresent the seven stationary points characterizing the symmethe cholesteric mesophase forU56 and j/p050.03. The threenodal sinks~SI! give the value of the equilibrium scalar order prametersS and P. The nodal source~SO! corresponds to the unstable isotropic source. The saddle points~SA! are unstable solu-tions corresponding to defects that the system might encountethe presence of orientational incompatibilities.
01170
f
ale
e
e
std
r
n-
minimized gradient energy. However, the blue-phase artecture has been identified in biological composites@1# and isrelevant in many other biological systems such as DNAlutions @16#. This simple phase-plane analysis of the reducgoverning equations allows an understanding of the symtry properties of these chiral assemblies at steady state.
B. Formation of the planar monodomain twistedplywood architecture
This section presents representative simulation resultsscribing the formation process of the planar monodomtwisted plywood architecture found in both cholesteric liqucrystals and biological fibrous composites@1#. For direct vi-sualization of the tensor order parameter fieldQ(x,y,t), wedraw parallelepipeds which render possible the simultanerepresentation of orientation and alignment informatioThese parallelepipeds are defined with the eigensystem oQ@17,18#; the three eigenvectors~n,m,l! give the orientation ofthe parallelepipeds and the corresponding three eigenva@ln52/3S,lm521/3(S2P),l l521/3(S1P)# give the de-gree of alignment along these orientation directions. Sithe eigenvalues ofQ span the interval@21/3,2/3#, they areaugmented by 1/3 so as to span the interval@0,1#. Therefore,the microstructure of the material is depicted by the shiftensorM5Q1d/3, as shown Fig. 4. Using this technique,isotropic state is represented by a small cube. A uniaxial sis represented by a parallelepiped with two edges of salength and a biaxial state by a parallelepiped with thedges of different lengths.
-s-
tsof
in
FIG. 3. Computed trajectories of the cholesteric sink node,j/p0 increases from 0.01 to 1. Each of the seven trajectories cosponds to a given value ofU. As U increases, the trajectories shiupwards in theS-P triangle. The start of each trajectory is thnearly uniaxial state (P'0) and the end is the maximally biaxialine (P5S). WhenU53 ~the limit of metastability of the ordinarynematic!, the increase ofj/p0 brings the stable state to the isotropnode.
6-5

hi
tiou
to
el-
ocalientsn-attoin
nto
ate
soy
-ubsdg
G. De LUCA AND A. D. REY PHYSICAL REVIEW E69, 011706 ~2004!
Since the effect of elastic anisotropy is out of scope in tstudy, we use the one-constant approximationy51 that ren-ders the splay, the twist, and the bend modes of deforma@6# indistinguishable and simplifies the long-range contrib
FIG. 4. Representation of microstructure by the shifted tenM5Q1d/3. The parallelepipeds are constructed with the eigenstem of Q; the three eigenvectors~n,m,l! giving the orientation andthe three eigenvalues (ln ,lm ,l l) augmented by 1/3 giving the degree of alignment. An isotropic state is represented by a small cA uniaxial state is represented by a parallelepiped with two edgesame length and a biaxial state by a parallelepiped with three eof different lengths.
01170
s
n-
tion of the governing equation as follows:
LR52S j
h0D 2
@¹2•Q#1S j
h0D S j
p0D @28p~¹3Q!# [s]
1S j
p0D 2
@216p2Q#. ~10!
Figure 5 shows a time series of the tensor fieldM (x,y,t)describing the propagation of order in the material, forU56, j/p050.03, andj/h050.015. ~For this representativesimulation the dimensionless pitch is therefore equal1/2.! The computational grid has 301 nodes in thex andydirections. When the thermodynamic potentialU ~propor-tional to concentration of fibrous molecules in the extraclular matrix! increases fromU,8/3 to U56, the homog-enous isotropic state becomes unstable and a nonlspontaneous phase-ordering process starts. Strong gradof order (S andP) are created between the initial constraiing layer aty50 and the isotropic material bulk. Recall that y50, (S,P) are the stable biaxial steady-state solutionsEqs. ~10!. As a result, order and orientation are inducedthe unstable isotropic bulk of the material and converted ia stable cholesteric phase. Figure 5~a! shows a field of ran-domly oriented cuboids representing the initial isotropic stof the material. Figures 5~b! and 5~c! depict the time evolu-
rs-
e.ofes
t-
-
-red-
t-
FIG. 5. Time evolution of themicrostructure.~a! Field of ran-domly oriented cuboids represening the initial isotropic state of thematerial. ~b!,~c! Order progressesas a moving front from the stableinitial constraining layer andleaves behind a fully relaxed helical microstructure. ~d! Steady-state microstructure of the material corresponding to the planamonodomain cholesteric structuranalogous to the twisted plywooarchitecture. The absence of topological defects is due to the facthat there is unique helical direction for the whole material. Thespecific times are 0~a!, 10 ~b!, 20~c!, 30 ~d!. Boxes are of unitlength.
6-6

inesed
n
thie
gsicreawbyn
ifi
theeno
ics--y
td
exonmte
thth
la
tylehten
.prsideooye
es
r-ed
ch.pa-
so,lityalled
td
g-ply-this
uid
y
ces,c-
s,e-
em
s.al
onsofa
-o-
ndg to
e
ve
CHIRAL FRONT PROPAGATION IN LIQUID- . . . PHYSICAL REVIEW E 69, 011706 ~2004!
tion of the microstructure. Order progresses as a movfront from the stable initial constraining layer and leavbehind it a fully relaxed helical microstructure. The ordermaterial exhibits a single helical direction~N! that is perpen-dicular to the moving front~and the surface of structure iformation!. The material remains homogeneous in thex di-rection perpendicular to the front. Once established,defect-free cholesteric order is stable and no further reortation mechanisms occur in its wake. Figure 5~d! gives thesteady-state microstructure of the material correspondinthe planar monodomain cholesteric structure analogouthe twisted plywood architecture. The absence of topologdefects is due to the fact that there is a unique helical dition ~N! for the whole material and therefore no orientationincompatibilities between neighboring regions exist. Knoing that the microstructure of the bulk is greatly affectedthe limiting surfaces and therefore this ‘‘ideal’’ arrangemeof the molecules within the material is due to the specanchoring of the molecules on the physical boundaries.
Periodic boundary conditions have been employed onx direction in order to remove surface effects from the sidThe upper boundary of the computational domain represthe surface of the secreting cell along which the fibrous mecules are isotropically ordered. Hence, the only physboundary that can act on the material bulk is the previousolidified ordered layer~lower boundary of the computational domain!. Along this initial constraining layer, the molecules describe a smooth~the director orientation is perfectlregular! planar~molecules parallel to the surface! state, thesystem naturally adopts its helical direction~N! perpendicu-larly to the restraining surface. It is understood here thathe domain were to be expanded—i.e., as would be expanin a biological system—this boundary would be orderedactly like its predecessor and serve as the new initial cstraining layer in the expansion cycle of the biological coposite. Therefore, by analogy, the monodomain twisarchitecture of biological fibrous composites is assumedarise through an ordering process qualitatively similar toone described above, in which a planar anchoring ofchiral molecules at the surface is indispensable.
Figure 6 shows the time evolution of the uniaxial scaorder parameterS ~degree of alignment! and the out-of-planecomponentunzu of the director field using gray scale intensiplots, for the same parametric values and dimensiontimes used in the simulations results shown in Fig. 5. Tfields go from white to black as the scalar order paramegoes from its equilibrium value to zero, and the out-of-placomponentunzu goes from 0 to61. From Fig. 6, it is clearthat both order (S) and orientationunzu propagate as frontsBoth fronts have the same speed, but the director frontcedes the scalar order front parameter. This is not surprias an increase in alignment—i.e., the scalar orparameter—requires a well-established average directorentation. In other words, the distance that separates the frof the two processes corresponds to a preoriented laTherefore the phase-ordering process consists first of thetablishment of orientation and then an increase of alignmalong the direction adopted by the system. The thicknesthe preorientation layer corresponds~roughly! to the half-
01170
g
en-
totoalc-l-
tc
es.tsl-ally
ifed--
-dtoee
r
sser
e
e-ngrri-ntser.s-
ntof
cholesteric pitch. In addition to the lag, Figs. 6~b!, 6~d!, and6~f! illustrate the periodic extinction of light that characteizes the twisted plywood architecture observed in polarizlight microscopy and that arises every half-cholesteric pit
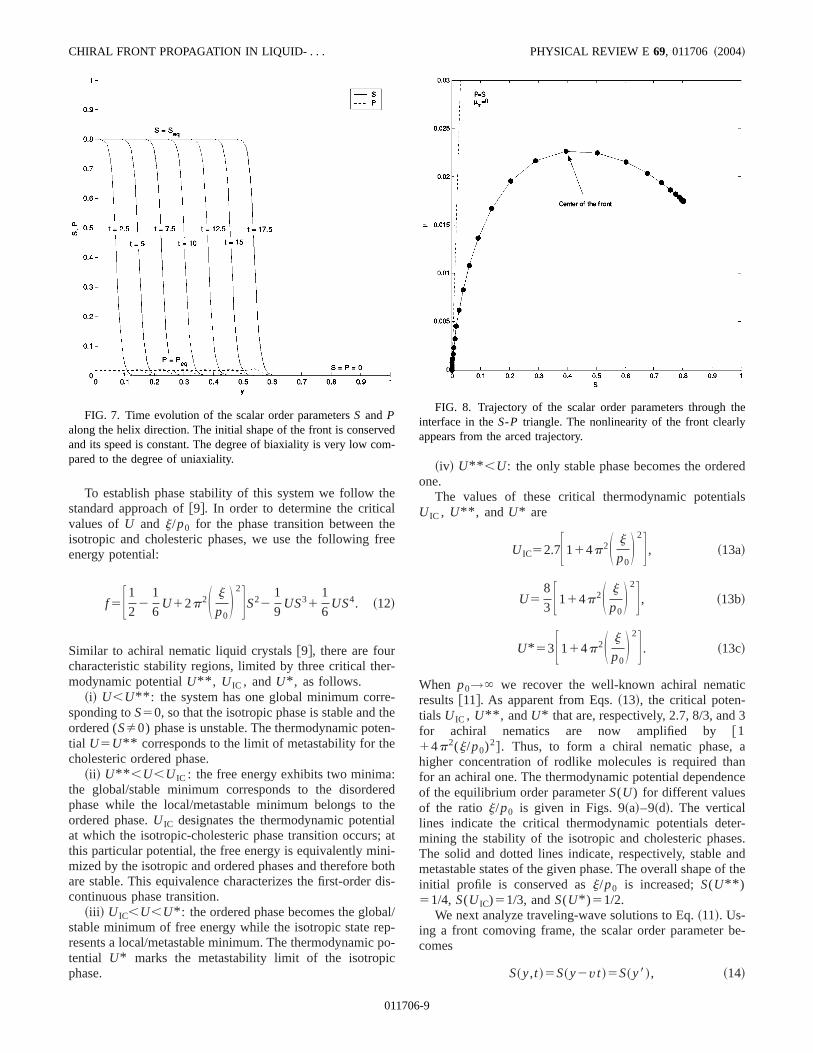
Figure 7 shows the time evolution of the scalar orderrametersS and P along the helix direction~N!. The pictureshows that the initial shape of the front is conserved. Althe speed of the front is constant and the degree of biaxiais very low compared to the degree of uniaxiality. The smovershoot in the biaxial order parameter profiles is explainby the nonlinearity of the front. SinceP is coupled toS andits gradient, the highest value ofP appears at the highesvalue in the gradient ofS. This is easily seen by the arcetrajectory made by the front in theS-P triangle ~Fig. 8!.
C. Kinetics of the phase-ordering process
In this section, we focus on the kinetics of the orderinprocess that leads to the planar monodomain twistedwood. The objective is to get a sense of the speed ofprocess with respect to the thermodynamic potentialU andthe length scale ratiosj/p0 andj/h0 ~material properties!.
In order to do this, we assume that the cholesteric liqcrystal is uniaxial~this is justified by the strong uniaxialitythat prevails over biaxiality in the cholesteric geometr!.Also, we assume that the directorn is fixed in time and thatit describes an ideal helix along the normal to the interfa~this is justified by the observation that, in the simulationthe kinetics of the moving front are unaffected by the diretor triad orientation!. Given these simplifying assumptionthe original problem reduces to a one-dimensional timdependent Ginzburg-Landau~TDGL! type of equation@19–21#.
The TDGL equation associated with the present problreads
]S
]t2S j
h0D 2 ]2S
]y2 52] f
]S5F211
1
3U24p2S j
p0D 2GS
11
3US22
2
3US3. ~11!
Equation~11! accepts two types of topological solutionThe first type of solutions interpolating between a locmaximum and a minimum of the free energy potentialf (S)are called relaxation modes. These traveling-wave solutihave a multitude of possible velocities. A second typetraveling-wave solutions, interpolating between two minimof the free energy potentialf (S), are called domain wallsolutions~or interface layers!. These solutions have a velocity uniquely determined by the form of the free energy ptential f (S) and by the boundary conditions@22#. We aretherefore interested in the latter solution, which are time- aspace-dependent traveling-wave solutions correspondinthe roots of the free energy potentialf (S). Although solu-tions to Eq.~12! are well known for the case of an infinitpitch p0 ~i.e., achiral nematics! @19–21#, the presence ofchirality (p0Þ0) introduces new significant effects that hanot been established or explained.
6-7

zero and
c
G. De LUCA AND A. D. REY PHYSICAL REVIEW E69, 011706 ~2004!
FIG. 6. Time evolution of the uniaxial scalar order parameterS ~left! and the out-of-plane componentunzu of the director field~right!using gray scale intensity plots. The fields go from white to black as the scalar order parameter goes from its equilibrium value tothe out-of-plane componentunzu goes from 0 to61. The specific time are 5~a!,~b!; 12.5~c!,~d!; 25 ~e!,~f!. Alignment (S) and orientationunzupropagate as fronts of equal speeds; however, the scalar order front parameter lags the director front, because to growS needs a well-established average orientation. The thickness of the preorientation layer corresponds~roughly! to the half-cholesteric pitch. The periodiextinction of light arises every half-cholesteric pitch. Boxes are of unit length.
011706-8
helere
r
e-thene
:rethia;nibod
aepp
c
red
als
ic
3
aannce
er-es.andf the
be-
the
em
CHIRAL FRONT PROPAGATION IN LIQUID- . . . PHYSICAL REVIEW E 69, 011706 ~2004!
To establish phase stability of this system we follow tstandard approach of@9#. In order to determine the criticavalues ofU and j/p0 for the phase transition between thisotropic and cholesteric phases, we use the following fenergy potential:
f 5F1
22
1
6U12p2S j
p0D 2GS22
1
9US31
1
6US4. ~12!
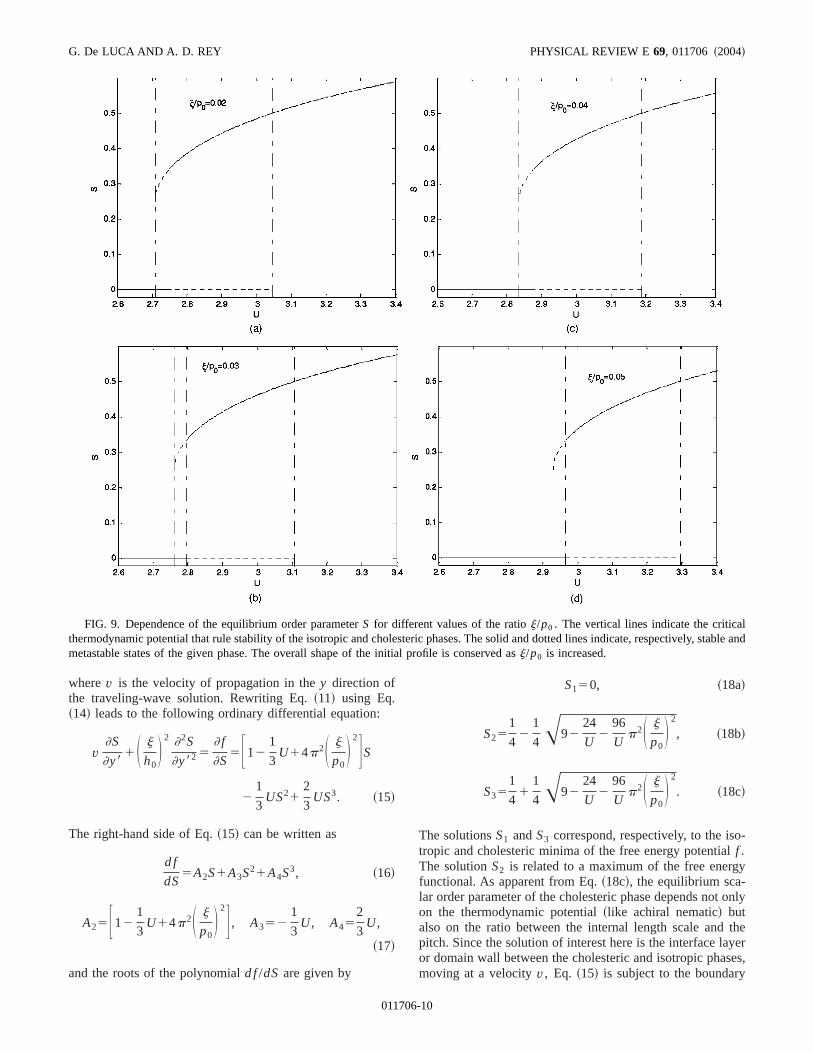
Similar to achiral nematic liquid crystals@9#, there are fourcharacteristic stability regions, limited by three critical themodynamic potentialU** , U IC , andU* , as follows.
~i! U,U** : the system has one global minimum corrsponding toS50, so that the isotropic phase is stable andordered (SÞ0) phase is unstable. The thermodynamic pottial U5U** corresponds to the limit of metastability for thcholesteric ordered phase.
~ii ! U** ,U,U IC : the free energy exhibits two minimathe global/stable minimum corresponds to the disordephase while the local/metastable minimum belongs toordered phase.U IC designates the thermodynamic potentat which the isotropic-cholesteric phase transition occursthis particular potential, the free energy is equivalently mimized by the isotropic and ordered phases and thereforeare stable. This equivalence characterizes the first-ordercontinuous phase transition.
~iii ! U IC,U,U* : the ordered phase becomes the globstable minimum of free energy while the isotropic state rresents a local/metastable minimum. The thermodynamictential U* marks the metastability limit of the isotropiphase.
FIG. 7. Time evolution of the scalar order parametersS and Palong the helix direction. The initial shape of the front is conservand its speed is constant. The degree of biaxiality is very low copared to the degree of uniaxiality.
01170
e
-
e-
delat-th
is-
l/-o-
~iv! U** ,U: the only stable phase becomes the ordeone.
The values of these critical thermodynamic potentiU IC , U** , andU* are
U IC52.7F114p2S j
p0D 2G , ~13a!
U58
3 F114p2S j
p0D 2G , ~13b!
U* 53F114p2S j
p0D 2G . ~13c!
When p0→` we recover the well-known achiral nematresults@11#. As apparent from Eqs.~13!, the critical poten-tials U IC , U** , andU* that are, respectively, 2.7, 8/3, andfor achiral nematics are now amplified by@114p2(j/p0)2#. Thus, to form a chiral nematic phase,higher concentration of rodlike molecules is required thfor an achiral one. The thermodynamic potential dependeof the equilibrium order parameterS(U) for different valuesof the ratio j/p0 is given in Figs. 9~a!–9~d!. The verticallines indicate the critical thermodynamic potentials detmining the stability of the isotropic and cholesteric phasThe solid and dotted lines indicate, respectively, stablemetastable states of the given phase. The overall shape oinitial profile is conserved asj/p0 is increased;S(U** )51/4, S(U IC)51/3, andS(U* )51/2.
We next analyze traveling-wave solutions to Eq.~11!. Us-ing a front comoving frame, the scalar order parametercomes
S~y,t !5S~y2vt !5S~y8!, ~14!
FIG. 8. Trajectory of the scalar order parameters throughinterface in theS-P triangle. The nonlinearity of the front clearlyappears from the arced trajectory.d-
6-9
ltable and
G. De LUCA AND A. D. REY PHYSICAL REVIEW E69, 011706 ~2004!
FIG. 9. Dependence of the equilibrium order parameterS for different values of the ratioj/p0 . The vertical lines indicate the criticathermodynamic potential that rule stability of the isotropic and cholesteric phases. The solid and dotted lines indicate, respectively, smetastable states of the given phase. The overall shape of the initial profile is conserved asj/p0 is increased.
-ly
only
theyerses,
wherev is the velocity of propagation in they direction ofthe traveling-wave solution. Rewriting Eq.~11! using Eq.~14! leads to the following ordinary differential equation:
v]S
]y81S j
h0D 2 ]2S
]y82 5] f
]S5F12
1
3U14p2S j
p0D 2GS
21
3US21
2
3US3. ~15!
The right-hand side of Eq.~15! can be written as
d f
dS5A2S1A3S21A4S3, ~16!
A25F121
3U14p2S j
p0D 2G , A352
1
3U, A45
2
3U,
~17!
and the roots of the polynomiald f /dS are given by
01170
S150, ~18a!
S251
42
1
4A92
24
U2
96
Up2S j
p0D 2
, ~18b!
S351
41
1
4A92
24
U2
96
Up2S j
p0D 2
. ~18c!
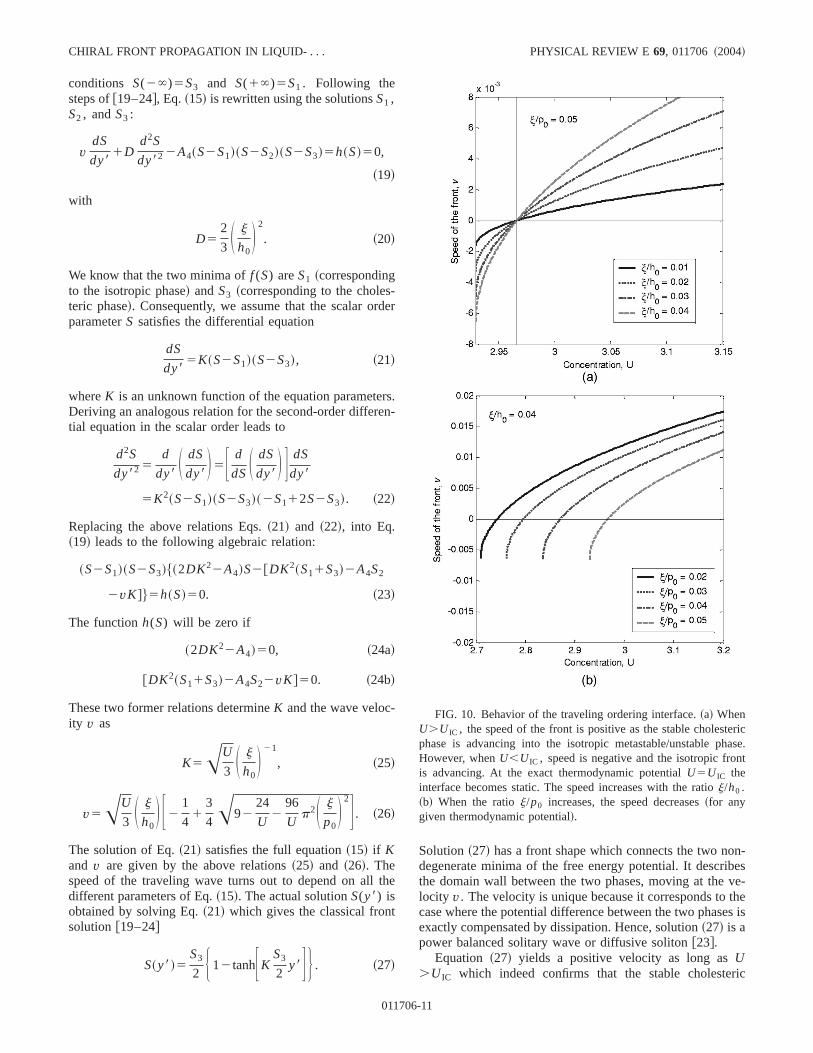
The solutionsS1 andS3 correspond, respectively, to the isotropic and cholesteric minima of the free energy potentiaf .The solutionS2 is related to a maximum of the free energfunctional. As apparent from Eq.~18c!, the equilibrium sca-lar order parameter of the cholesteric phase depends noton the thermodynamic potential~like achiral nematic! butalso on the ratio between the internal length scale andpitch. Since the solution of interest here is the interface laor domain wall between the cholesteric and isotropic phamoving at a velocityv, Eq. ~15! is subject to the boundary
6-10
-d
rsen
th
t
n-besve-thees is
ric
ericase.
nt
CHIRAL FRONT PROPAGATION IN LIQUID- . . . PHYSICAL REVIEW E 69, 011706 ~2004!
conditions S(2`)5S3 and S(1`)5S1 . Following thesteps of@19–24#, Eq.~15! is rewritten using the solutionsS1 ,S2 , andS3 :
vdS
dy81D
d2S
dy82 2A4~S2S1!~S2S2!~S2S3!5h~S!50,
~19!
with
D52
3 S j
h0D 2
. ~20!
We know that the two minima off (S) areS1 ~correspondingto the isotropic phase! andS3 ~corresponding to the cholesteric phase!. Consequently, we assume that the scalar orparameterS satisfies the differential equation
dS
dy85K~S2S1!~S2S3!, ~21!
whereK is an unknown function of the equation parameteDeriving an analogous relation for the second-order differtial equation in the scalar order leads to
d2S
dy82 5d
dy8 S dS
dy8D5F d
dSS dS
dy8D G dS
dy8
5K2~S2S1!~S2S3!~2S112S2S3!. ~22!
Replacing the above relations Eqs.~21! and ~22!, into Eq.~19! leads to the following algebraic relation:
~S2S1!~S2S3!$~2DK22A4!S2@DK2~S11S3!2A4S2
2vK#%5h~S!50. ~23!
The functionh(S) will be zero if
~2DK22A4!50, ~24a!
@DK2~S11S3!2A4S22vK#50. ~24b!
These two former relations determineK and the wave veloc-ity v as
K5AU
3 S j
h0D 21
, ~25!
v5AU
3 S j
h0D F2
1
41
3
4A92
24
U2
96
Up2S j
p0D 2G . ~26!
The solution of Eq.~21! satisfies the full equation~15! if Kand v are given by the above relations~25! and ~26!. Thespeed of the traveling wave turns out to depend on alldifferent parameters of Eq.~15!. The actual solutionS(y8) isobtained by solving Eq.~21! which gives the classical fronsolution @19–24#
S~y8!5S3
2 H 12tanhFKS3
2y8G J . ~27!
01170
er
.-
e
Solution~27! has a front shape which connects the two nodegenerate minima of the free energy potential. It descrithe domain wall between the two phases, moving at thelocity v. The velocity is unique because it corresponds tocase where the potential difference between the two phasexactly compensated by dissipation. Hence, solution~27! is apower balanced solitary wave or diffusive soliton@23#.
Equation ~27! yields a positive velocity as long asU.U IC which indeed confirms that the stable choleste
FIG. 10. Behavior of the traveling ordering interface.~a! WhenU.U IC , the speed of the front is positive as the stable cholestphase is advancing into the isotropic metastable/unstable phHowever, whenU,U IC , speed is negative and the isotropic frois advancing. At the exact thermodynamic potentialU5U IC theinterface becomes static. The speed increases with the ratioj/h0 .~b! When the ratioj/p0 increases, the speed decreases~for anygiven thermodynamic potential!.
6-11
ableetialthe
ver,the
ae italy-
theisred
er-
erase
onick-
ing
ame
pa-
erthat
ndxialm-
m-mesestlythee-l toionof a
beena-essinro-tex-stedc-
The
r a
-o
G. De LUCA AND A. D. REY PHYSICAL REVIEW E69, 011706 ~2004!
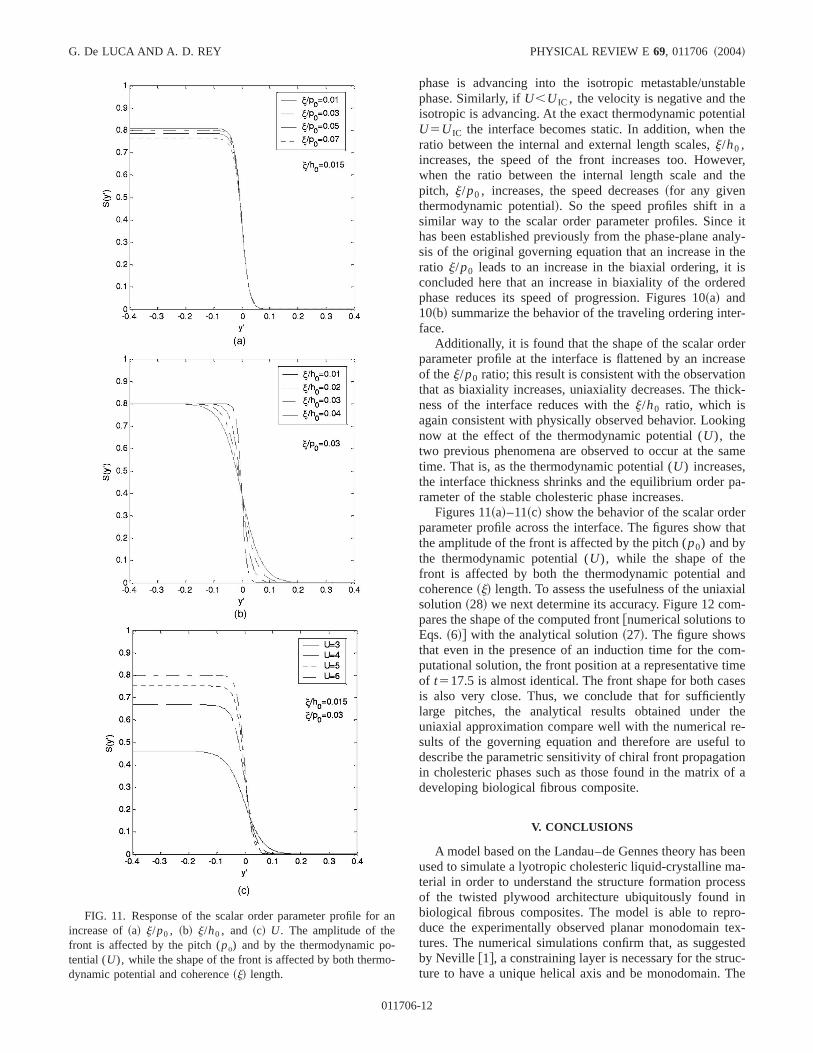
FIG. 11. Response of the scalar order parameter profile foincrease of~a! j/p0 , ~b! j/h0 , and ~c! U. The amplitude of thefront is affected by the pitch (po) and by the thermodynamic potential (U), while the shape of the front is affected by both thermdynamic potential and coherence~j! length.
01170
phase is advancing into the isotropic metastable/unstphase. Similarly, ifU,U IC , the velocity is negative and thisotropic is advancing. At the exact thermodynamic potenU5U IC the interface becomes static. In addition, whenratio between the internal and external length scales,j/h0 ,increases, the speed of the front increases too. Howewhen the ratio between the internal length scale andpitch, j/p0 , increases, the speed decreases~for any giventhermodynamic potential!. So the speed profiles shift insimilar way to the scalar order parameter profiles. Sinchas been established previously from the phase-plane ansis of the original governing equation that an increase inratio j/p0 leads to an increase in the biaxial ordering, itconcluded here that an increase in biaxiality of the ordephase reduces its speed of progression. Figures 10~a! and10~b! summarize the behavior of the traveling ordering intface.
Additionally, it is found that the shape of the scalar ordparameter profile at the interface is flattened by an increof thej/p0 ratio; this result is consistent with the observatithat as biaxiality increases, uniaxiality decreases. The thness of the interface reduces with thej/h0 ratio, which isagain consistent with physically observed behavior. Looknow at the effect of the thermodynamic potential (U), thetwo previous phenomena are observed to occur at the stime. That is, as the thermodynamic potential (U) increases,the interface thickness shrinks and the equilibrium orderrameter of the stable cholesteric phase increases.
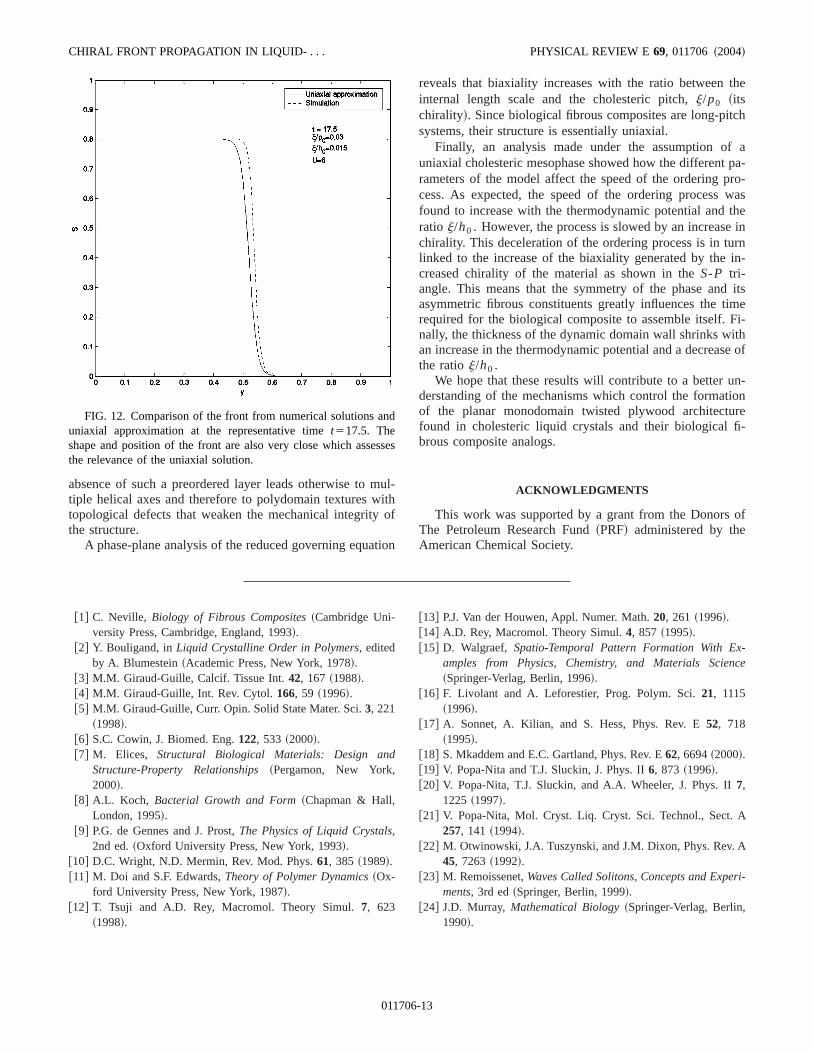
Figures 11~a!–11~c! show the behavior of the scalar ordparameter profile across the interface. The figures showthe amplitude of the front is affected by the pitch (p0) and bythe thermodynamic potential (U), while the shape of thefront is affected by both the thermodynamic potential acoherence~j! length. To assess the usefulness of the uniasolution~28! we next determine its accuracy. Figure 12 copares the shape of the computed front@numerical solutions toEqs.~6!# with the analytical solution~27!. The figure showsthat even in the presence of an induction time for the coputational solution, the front position at a representative tiof t517.5 is almost identical. The front shape for both cais also very close. Thus, we conclude that for sufficienlarge pitches, the analytical results obtained underuniaxial approximation compare well with the numerical rsults of the governing equation and therefore are usefudescribe the parametric sensitivity of chiral front propagatin cholesteric phases such as those found in the matrixdeveloping biological fibrous composite.
V. CONCLUSIONS
A model based on the Landau–de Gennes theory hasused to simulate a lyotropic cholesteric liquid-crystalline mterial in order to understand the structure formation procof the twisted plywood architecture ubiquitously foundbiological fibrous composites. The model is able to repduce the experimentally observed planar monodomaintures. The numerical simulations confirm that, as suggeby Neville @1#, a constraining layer is necessary for the struture to have a unique helical axis and be monodomain.
n
-
6-12
mito
tio
the
ch
f apa-ro-was
thein
rnin-
itsimeFi-ithe of
un-ionre
fi-
of
nd
ss
CHIRAL FRONT PROPAGATION IN LIQUID- . . . PHYSICAL REVIEW E 69, 011706 ~2004!
absence of such a preordered layer leads otherwise totiple helical axes and therefore to polydomain textures wtopological defects that weaken the mechanical integritythe structure.
A phase-plane analysis of the reduced governing equa
FIG. 12. Comparison of the front from numerical solutions auniaxial approximation at the representative timet517.5. Theshape and position of the front are also very close which assethe relevance of the uniaxial solution.
01170
ul-hf
n
reveals that biaxiality increases with the ratio betweeninternal length scale and the cholesteric pitch,j/p0 ~itschirality!. Since biological fibrous composites are long-pitsystems, their structure is essentially uniaxial.
Finally, an analysis made under the assumption ouniaxial cholesteric mesophase showed how the differentrameters of the model affect the speed of the ordering pcess. As expected, the speed of the ordering processfound to increase with the thermodynamic potential andratio j/h0 . However, the process is slowed by an increasechirality. This deceleration of the ordering process is in tulinked to the increase of the biaxiality generated by thecreased chirality of the material as shown in theS-P tri-angle. This means that the symmetry of the phase andasymmetric fibrous constituents greatly influences the trequired for the biological composite to assemble itself.nally, the thickness of the dynamic domain wall shrinks wan increase in the thermodynamic potential and a decreasthe ratioj/h0 .
We hope that these results will contribute to a betterderstanding of the mechanisms which control the formatof the planar monodomain twisted plywood architectufound in cholesteric liquid crystals and their biologicalbrous composite analogs.
ACKNOWLEDGMENTS
This work was supported by a grant from the DonorsThe Petroleum Research Fund~PRF! administered by theAmerican Chemical Society.
es
-ce
A
A
ri-
@1# C. Neville, Biology of Fibrous Composites~Cambridge Uni-versity Press, Cambridge, England, 1993!.
@2# Y. Bouligand, inLiquid Crystalline Order in Polymers, editedby A. Blumestein~Academic Press, New York, 1978!.
@3# M.M. Giraud-Guille, Calcif. Tissue Int.42, 167 ~1988!.@4# M.M. Giraud-Guille, Int. Rev. Cytol.166, 59 ~1996!.@5# M.M. Giraud-Guille, Curr. Opin. Solid State Mater. Sci.3, 221
~1998!.@6# S.C. Cowin, J. Biomed. Eng.122, 533 ~2000!.@7# M. Elices, Structural Biological Materials: Design and
Structure-Property Relationships~Pergamon, New York,2000!.
@8# A.L. Koch, Bacterial Growth and Form~Chapman & Hall,London, 1995!.
@9# P.G. de Gennes and J. Prost,The Physics of Liquid Crystals,2nd ed.~Oxford University Press, New York, 1993!.
@10# D.C. Wright, N.D. Mermin, Rev. Mod. Phys.61, 385 ~1989!.@11# M. Doi and S.F. Edwards,Theory of Polymer Dynamics~Ox-
ford University Press, New York, 1987!.@12# T. Tsuji and A.D. Rey, Macromol. Theory Simul.7, 623
~1998!.
@13# P.J. Van der Houwen, Appl. Numer. Math.20, 261 ~1996!.@14# A.D. Rey, Macromol. Theory Simul.4, 857 ~1995!.@15# D. Walgraef, Spatio-Temporal Pattern Formation With Ex
amples from Physics, Chemistry, and Materials Scien~Springer-Verlag, Berlin, 1996!.
@16# F. Livolant and A. Leforestier, Prog. Polym. Sci.21, 1115~1996!.
@17# A. Sonnet, A. Kilian, and S. Hess, Phys. Rev. E52, 718~1995!.
@18# S. Mkaddem and E.C. Gartland, Phys. Rev. E62, 6694~2000!.@19# V. Popa-Nita and T.J. Sluckin, J. Phys. II6, 873 ~1996!.@20# V. Popa-Nita, T.J. Sluckin, and A.A. Wheeler, J. Phys. II7,
1225 ~1997!.@21# V. Popa-Nita, Mol. Cryst. Liq. Cryst. Sci. Technol., Sect.
257, 141 ~1994!.@22# M. Otwinowski, J.A. Tuszynski, and J.M. Dixon, Phys. Rev.
45, 7263~1992!.@23# M. Remoissenet,Waves Called Solitons, Concepts and Expe
ments, 3rd ed~Springer, Berlin, 1999!.@24# J.D. Murray,Mathematical Biology~Springer-Verlag, Berlin,
1990!.
6-13
Related Documents











