Hepatology Research 15 (1999) 10 – 21 Chemically induced liver regeneration is characterized by specific IL-6 gene expression Adriana Salazar-Montes, Ana Rosa Rinco ´ n, Arturo Panduro, Juan Armenda ´riz-Borunda * Institute of Molecular Biology in Medicine, CUCS, Uni6ersity of Guadalajara, Apdo. Postal 2 -500, 44281 Guadalajara, Jal, Mexico Received 30 March 1998; received in revised form 1 June 1998; accepted 15 June 1998 Abstract IL-6 and TNFa have been shown as important cytokines in liver regeneration induced in partially hepatectomized rats. Nonetheless, IL-6 gene expression in the course of chemically induced hepatic regeneration, resembling what occurs in humans infected by hepatitis virus in acute manner, remains obscure. To gain an insight on this matter we determined IL-6 and TNFa gene expression in Wistar rats after acute and chronic liver intoxication with CCl 4 and/or turpentine. We used a semiquantitative method of RT-PCR to analyze cytokine gene expression and, quite interestingly, IL-6 was detected only at 24 h after acute damage as opposed to TNFa expression that was increased from 6 to 48 h after acute injury. A single intradermally-injected dose of turpentine did not suffice to induce IL-6 gene expression at any studied times, but, as expected, turpentine-treated animals showed decreased albumin mRNA transcripts as compared with the control rats. Furthermore, livers from cirrhotic animals did not show detectable IL-6 gene transcripts, but, IL-6 expression re-appeared in cirrhotic rats treated with an additional acute CCl 4 dose, indicating that IL-6 is a critical component of the brisk regenerative response induced by CCl 4 . © 1999 Elsevier Science Ireland Ltd. All rights reserved. Keywords: Gene expression-regulation; RT-PCR; IL-6; Liver regeneration/inflammation * Corresponding author. Tel.: +52 3 6174159; fax: +52 3 6174159. E-mail address : [email protected] (J. Armenda ´riz-Borunda) 1386-6346/99/$ - see front matter © 1999 Elsevier Science Ireland Ltd. All rights reserved. PII S1386-6346(98)00056-4

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Hepatology Research 15 (1999) 10–21
Chemically induced liver regeneration ischaracterized by specific IL-6 gene expression
Adriana Salazar-Montes, Ana Rosa Rincon, Arturo Panduro,Juan Armendariz-Borunda *
Institute of Molecular Biology in Medicine, CUCS, Uni6ersity of Guadalajara, Apdo. Postal 2-500,44281 Guadalajara, Jal, Mexico
Received 30 March 1998; received in revised form 1 June 1998; accepted 15 June 1998
Abstract
IL-6 and TNFa have been shown as important cytokines in liver regeneration induced inpartially hepatectomized rats. Nonetheless, IL-6 gene expression in the course of chemicallyinduced hepatic regeneration, resembling what occurs in humans infected by hepatitis virusin acute manner, remains obscure. To gain an insight on this matter we determined IL-6 andTNFa gene expression in Wistar rats after acute and chronic liver intoxication with CCl4and/or turpentine. We used a semiquantitative method of RT-PCR to analyze cytokine geneexpression and, quite interestingly, IL-6 was detected only at 24 h after acute damage asopposed to TNFa expression that was increased from 6 to 48 h after acute injury. A singleintradermally-injected dose of turpentine did not suffice to induce IL-6 gene expression atany studied times, but, as expected, turpentine-treated animals showed decreased albuminmRNA transcripts as compared with the control rats. Furthermore, livers from cirrhoticanimals did not show detectable IL-6 gene transcripts, but, IL-6 expression re-appeared incirrhotic rats treated with an additional acute CCl4 dose, indicating that IL-6 is a criticalcomponent of the brisk regenerative response induced by CCl4. © 1999 Elsevier ScienceIreland Ltd. All rights reserved.
Keywords: Gene expression-regulation; RT-PCR; IL-6; Liver regeneration/inflammation
* Corresponding author. Tel.: +52 3 6174159; fax: +52 3 6174159.E-mail address : [email protected] (J. Armendariz-Borunda)
1386-6346/99/$ - see front matter © 1999 Elsevier Science Ireland Ltd. All rights reserved.
PII S1386-6346(98)00056-4

A. Salazar-Montes et al. / Hepatology Research 15 (1999) 10–21 11
1. Introduction
Interleukin-6 (IL-6) is a 26-kDa pleiotropic cytokine exerting multiple biologicactivities on different types of target cells and mediates many of the hepaticaspects of the acute phase response, as well as the induction of acute phaseproteins by cultured hepatocytes or cells lines derived from hepatocytes [1,2].
At the same time, expression of IL-6 can be induced in various cell typesincluding fibroblasts, monocytes/macrophages, T cells, B cells, endothelial, epider-mal, synovial, and diverse tumor cells [3]. In spite of such evidence, the role andgene expression of IL-6 on the course of several hepatic injuries such as alcoholicliver disease (ALD), induced either in humans or rats, remains a matter ofcontroversy. It is known that patients with ALD display several of the manifesta-tions of the acute phase response [4–6] and blood levels of IL-1b, TNFa andIL-6 are increased [7–10]. Upon recovery of an acute episode of alcoholic hepati-tis, blood IL-6 levels return to normal values, whereas TNFa and IL-1 remainelevated for several weeks [9]. Another interesting finding shows IL-6 expressionin 100% of the patients with primary biliary cirrhosis [11]. In this context,Kupffer cells and hepatic sinusoidal cells are thought to be the major sources ofthese inflammatory cytokines [12].
In experimental animal models, IL-6 has been shown to be an important factorin the course of liver regeneration induced after partial hepatectomy in rats andsignificant increases in serum concentrations of IL-6 have been reported 24–48 hafter the surgical procedure in rats [13,14]. Nonetheless, a more conclusive studyhas unveiled IL-6 gene expression in liver regeneration; with the availability ofmice harboring a targeted disruption of the gene encoding IL-6, Cressman et al.[15] demonstrated that these mice had impaired liver regeneration characterizedby liver necrosis and failure. There was a blunted DNA synthetic response inhepatocytes of these mice but not in nonparenchymal cells. Furthermore, theyfound important abnormalities of transcriptional factors including absence ofSTAT-3 (signal transducer and activator of transcription protein 3) activation,and depressed AP-1, Myc and cyclin DI gene expression. The treatment ofIL-6-deficient transgenic mice with a single preoperative dose of IL-6 returnedSTAT-3 binding, gene expression and hepatocyte proliferation to near normaland prevented liver damage. Taken together, these data suggests that IL-6 is acritical component of the regenerative response induced in hepatectomized ani-mals [15]. Furthermore, compelling evidence has accumulated showing IL-6 as anessential and irreplaceable cytokine in order for hepatic regeneration proceed tocompletion, but the IL-6 role on triggering the initiating response on this physio-logical phenomena has yet to be proven [16].
On the other hand, Greenwel et al. [17] have shown that IL-6 plays a majorrole in mediating the expression of acute phase proteins and the increase in a1 (I)procollagen mRNA after turpentine administration. However, studies performedby Northermann et al. [18] showed no IL-6 mRNA expression in rat liversundergoing an acute-phase response following turpentine injection, suggesting that

A. Salazar-Montes et al. / Hepatology Research 15 (1999) 10–2112
Kupffer cells were not especially activated in the acute-phase response. An interest-ing fact is that the same study showed no IL-6 gene expression by normal livers[18]. The role of TNFa has also been analyzed in hepatic regeneration induced bypartial hepatectomy. Yamada et al. [19] showed that TNFa signaling through TNFreceptor (TNFR-I) can initiate liver regeneration and acts by activating an IL-6dependent pathway that involves the STAT-3 transcription factor.
The aim of this investigation was to determine the role and timing of IL-6 geneexpression on the regenerative response induced by acute chemical intoxication withCCl4. To gain insight on this matter, we investigated IL-6 gene expression in liverof rats intoxicated acute and chronically with CCl4 or turpentine. Our results pointto IL-6 as an important component in the brisk regenerative response induced byCCl4.
2. Materials and methods
2.1. Reagents and equipment
CCl4 was purchased from Merck (Darmstadt, Germany). Mineral oil and agarosewere from Sigma (St. Louis, MO). Trizol, M-MLV reverse transcriptase, dNTPsRNAases inhibitor, Taq DNA polymerase, random primers and PCR primers wereobtained from Gibco BRL (Grand Island, NY). Polytron homogenizer was fromBrinkmann (Switzerland). Thermocycler was purchased from Perkin-Elmer (Nor-walk, CT).
2.2. Animals
Rats used in this study were obtained from Charles Rivers Wistar strainweighting 200 g and housed according to the Animal Care protocol established byUniversity of Guadalajara Animal Committee. For the acute intoxication experi-ments, three rats were given a single dose of 0.5 ml per 100 g weight of a 1:1CCl4/mineral oil mix intragastrically; the animals were sacrificed and the livers wereremoved at the indicated time-points. Three control animals were administratedonly the vehicle. For the experiments of acute phase response induction, two animalgroups (three rats per group) were used: one group was given a single dose of 0.5ml per 200 g of turpentine intradermally; the animals were sacrificed and the liverswere removed at the indicated time points. Another group of rats were first madecirrhotic by chronic administration of CCl4 for 8 weeks [20]. Briefly, 80 g rats weretreated i.p. with 200 m l of 1:6 CCl4/mineral oil mix for the first week, three timesa week. The animals were then injected with 1:5, 1:4 and 1:3 mix for the 2nd, 3rdand 4–8th weeks, respectively. Then, these animals were subjected to a furtheridentical treatment with turpentine as mentioned above. Finally, last group of rats(n=3) were given a chronic dose of CCl4 as described [20] and sacrificed after 8weeks of treatment.

A. Salazar-Montes et al. / Hepatology Research 15 (1999) 10–21 13
2.3. Extraction and quantitation of RNA
Isolation of total RNA from rat livers was carried out according tothe method described by Chomczynski and Sacchi [21]. Briefly, liver tissuewas homogenized using a polytron system in the presence of Trizol™,chloroform was added, the aqueous phase was obtained and the RNAprecipitated with isopropanol at 4°C overnight. The quantity and intactness ofRNA were routinely tested by determining A260/280 and ethidium bromidefluorescence of RNA electrophoresed in 1% formaldehyde-containing agarosegels.
2.4. Determination of cytokines and albumin gene expression by RT-PCR
Three reverse transcriptase reactions per animal were carried out in order toobtain the corresponding cDNAs. Reverse transcriptase-polymerase chain reac-tion (RT-PCR) was performed according to previously described methodology[22]. To achieve this, we standardized a semiquantitative PCR method based inthe co-amplification of the target gene (IL-6) and a constitutive gene (b-actin) inthe same reaction tube. Briefly, RNA from liver samples was isolated withTrizol™ and 2 mg of total RNA were reverse transcribed in 0.05 M Tris–HClpH 8.3, 40 mM KCl, 7 mM MgCl2 buffer containing 0.05 mg m l−1 of randomhexamers, 1 mM dNTPs mix, 0.05 U m l−1 RNase inhibitor and 200 U m l−1
murine leukemia virus reverse transcriptase M-MLV. Samples were incubated for10 min at 70°C and then 60 min at 37.5°C. Reverse transcriptase was furtherinactivated by heating the sample tubes at 95°C for 10 min. cDNAs obtainedthis way, were used immediately for reaction or were stored at −20°C until use.PCR reactions were performed according to our previous observations where wedetermine the optimal conditions to pick up IL-6 gene expression in the contextof the co-determination of the expression of a constitutive gene like b-actin.Gene products amplification was performed in a PCR buffer of 50 mM Tris–HCl, pH 9.0, and 50 mM NaCl containing a mix of 100 mM dNTPs and 1 U ofTaq DNA polymerase. Final primer concentrations were as follows: 0.3 mM forIL-6 sense (5%-CTTCCAGCCAGTTGCCTTCT-3%) and antisense (5%-GACAG-CATTGGAAGTTGGGG-3%) [23] oligonucleotides and 0.03 mM for b-actinsense (5%-TCCTGTGGCATCCACGAAACT-3%) and antisense (5%-TAGCAG-GTGGCGTTTACGAAG-3%) oligonucleotides [24]. Final TNFa primer concen-tration was: sense (5%-CGAGTGACAAGCCCGTAGCC-3%) 0.3 mM andantisense (5%-GGATGAACACGCCAGTCGCC-3%) [23] oligonucleotide 0.3 mM.Albumin primers used were: sense (5%-TTGCCAAGTACATGTGTGGTGAG-3%)0.03 mM and antisense (5%-GGTTCTTCTACAAGAGGCTG-3%) [25]. Amplifica-tion reactions were overlaid with light mineral oil and held at 95°C for ‘hot-start’ PCR for 5 min and run in an automated thermal cycler for 40 cycles eachof 95°C for 1 min, 60°C for 1 min and 72°C for 1.5 min and a final cycle ofextension for 5 min at 72°C.

A. Salazar-Montes et al. / Hepatology Research 15 (1999) 10–2114
2.5. Photographic and densitometric scanning
The densitometric analysis of the PCR products were performed in a DU seriesBeckman spectrophotometer using a gel scan area program. Samples prepared byelectrophoresis were photographed with 665 Polaroid film while being exposed toUV light and the negative of the film containing the dark bands was scanned by theinstrument. After the scan was complete, the area corresponding to each bandrepresenting the corresponding amplified PCR products, was automatically calcu-lated and normalized against the area represented by the expression of the constitu-tive gene. The results were then expressed as arbitrary absorbance units.
3. Results
According to our previous results, liver histology was characterized by anextensive hemorrhage and massive infiltration of inflammatory cells around centralveins and portal tracts mainly at 48 h after acute CCl4 injury, being resolvedthereafter [26]. On the other hand, cirrhosis in the rats treated chronically with CCl4was histologically demonstrated by parenchymal nodules of irregular size, sur-rounded by bundles of extracellular matrix components and by the presence ofnecrotized hepatocytes (data not shown).
Because the cytokine mRNAs exist at low levels in rat liver [27], we extractedtotal RNA from liver of rats subjected to several chemical injuries and performedRT-PCR using cytokine-specific primers to compare IL-6 expression between thedifferent groups. Three rats were studied in each group and they were treated asdescribed in Section 2. Moreover, three RT-PCR reactions were carried out foreach animal. The differences between the experimental and the control groups werenot due to variations in the RNA isolation procedure or the efficiency of the reversetranscription reaction, since equivalent amounts of b-actin mRNA (expression of aconstitutive gene) were expressed by rat livers from the same group.
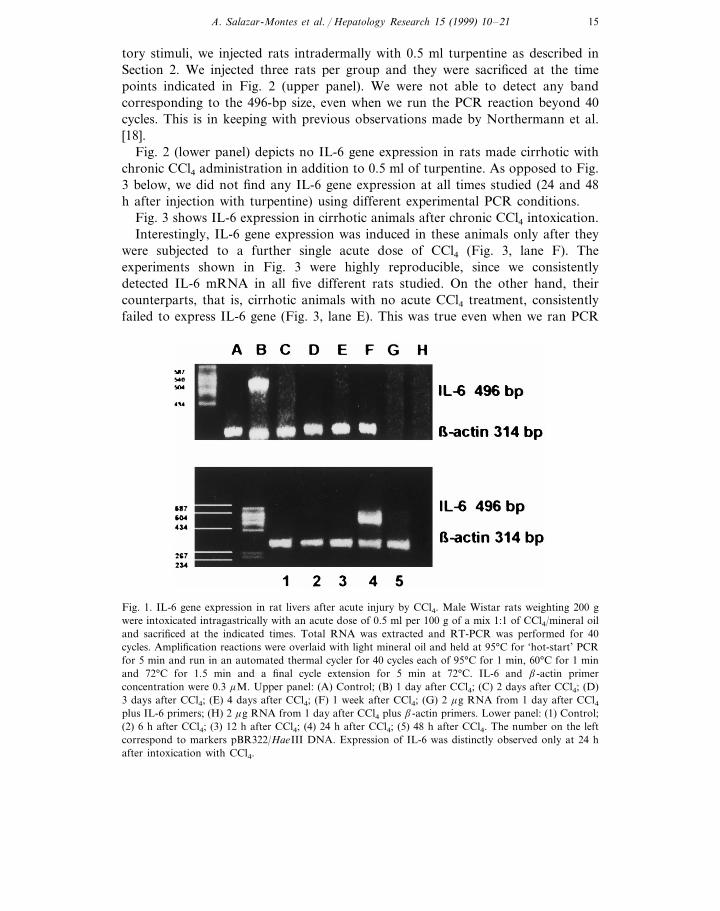
Fig. 1 depicts a ethidium bromide-stained agarose gel showing two differentseries of experiments carried out to detect the amplified RT-PCR products forb-actin (314 bp) and IL-6 (496 bp) mRNAs. It is clear that IL-6 gene expressionwas strictly detected only at 24 h after acute CCl4 intoxication. We did not detectIL-6 mRNA neither before (Fig. 1, lower panel) nor after (Fig. 1, upper panel) 24h post-CCl4 acute intoxication. It is also important to note that IL-6 correspondingband was detected only after a rigorous and exhaustive series of experiments carriedon at different experimental conditions, i.e. different number of cycles and temper-atures of denaturalization and annealing (data not shown). We could only detectIL-6 after 40 cycles of amplification. This fact did not introduce experimentalartifacts as is shown by the clear presence of the bands corresponding to IL-6 andb-actin transcripts. We also performed experiments to rule out genomic DNAcontamination of RNA samples (see Fig. 1G and H, upper panel).
To compare the specific gene expression of IL-6 in a different model ofinflammation where an acute phase response is induced by nonspecific inflamma-
A. Salazar-Montes et al. / Hepatology Research 15 (1999) 10–21 15
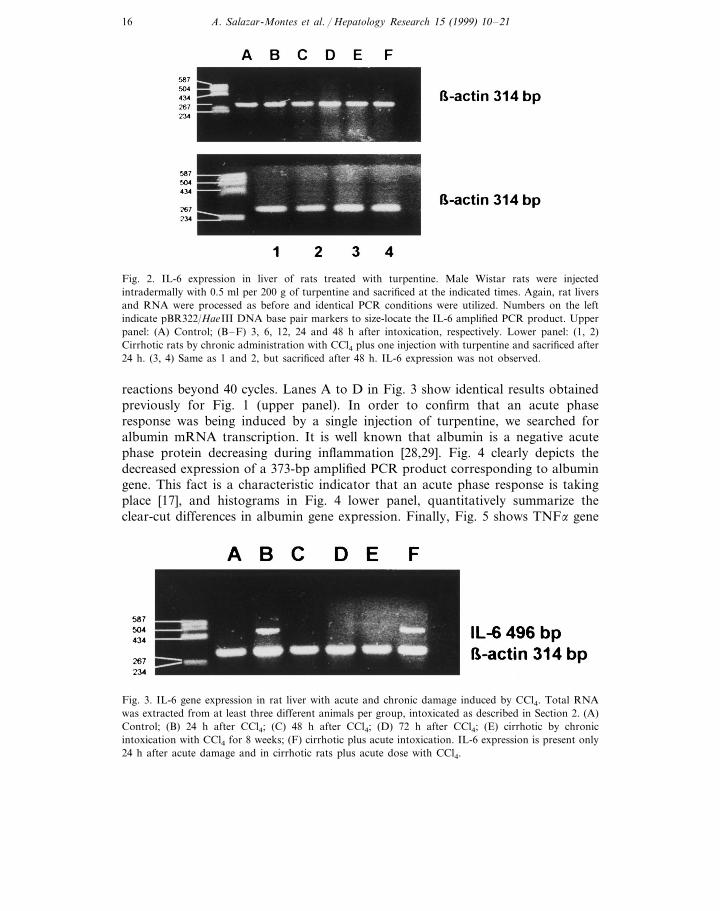
tory stimuli, we injected rats intradermally with 0.5 ml turpentine as described inSection 2. We injected three rats per group and they were sacrificed at the timepoints indicated in Fig. 2 (upper panel). We were not able to detect any bandcorresponding to the 496-bp size, even when we run the PCR reaction beyond 40cycles. This is in keeping with previous observations made by Northermann et al.[18].
Fig. 2 (lower panel) depicts no IL-6 gene expression in rats made cirrhotic withchronic CCl4 administration in addition to 0.5 ml of turpentine. As opposed to Fig.3 below, we did not find any IL-6 gene expression at all times studied (24 and 48h after injection with turpentine) using different experimental PCR conditions.
Fig. 3 shows IL-6 expression in cirrhotic animals after chronic CCl4 intoxication.Interestingly, IL-6 gene expression was induced in these animals only after they
were subjected to a further single acute dose of CCl4 (Fig. 3, lane F). Theexperiments shown in Fig. 3 were highly reproducible, since we consistentlydetected IL-6 mRNA in all five different rats studied. On the other hand, theircounterparts, that is, cirrhotic animals with no acute CCl4 treatment, consistentlyfailed to express IL-6 gene (Fig. 3, lane E). This was true even when we ran PCR
Fig. 1. IL-6 gene expression in rat livers after acute injury by CCl4. Male Wistar rats weighting 200 gwere intoxicated intragastrically with an acute dose of 0.5 ml per 100 g of a mix 1:1 of CCl4/mineral oiland sacrificed at the indicated times. Total RNA was extracted and RT-PCR was performed for 40cycles. Amplification reactions were overlaid with light mineral oil and held at 95°C for ‘hot-start’ PCRfor 5 min and run in an automated thermal cycler for 40 cycles each of 95°C for 1 min, 60°C for 1 minand 72°C for 1.5 min and a final cycle extension for 5 min at 72°C. IL-6 and b-actin primerconcentration were 0.3 mM. Upper panel: (A) Control; (B) 1 day after CCl4; (C) 2 days after CCl4; (D)3 days after CCl4; (E) 4 days after CCl4; (F) 1 week after CCl4; (G) 2 mg RNA from 1 day after CCl4plus IL-6 primers; (H) 2 mg RNA from 1 day after CCl4 plus b-actin primers. Lower panel: (1) Control;(2) 6 h after CCl4; (3) 12 h after CCl4; (4) 24 h after CCl4; (5) 48 h after CCl4. The number on the leftcorrespond to markers pBR322/HaeIII DNA. Expression of IL-6 was distinctly observed only at 24 hafter intoxication with CCl4.
A. Salazar-Montes et al. / Hepatology Research 15 (1999) 10–2116
Fig. 2. IL-6 expression in liver of rats treated with turpentine. Male Wistar rats were injectedintradermally with 0.5 ml per 200 g of turpentine and sacrificed at the indicated times. Again, rat liversand RNA were processed as before and identical PCR conditions were utilized. Numbers on the leftindicate pBR322/HaeIII DNA base pair markers to size-locate the IL-6 amplified PCR product. Upperpanel: (A) Control; (B–F) 3, 6, 12, 24 and 48 h after intoxication, respectively. Lower panel: (1, 2)Cirrhotic rats by chronic administration with CCl4 plus one injection with turpentine and sacrificed after24 h. (3, 4) Same as 1 and 2, but sacrificed after 48 h. IL-6 expression was not observed.
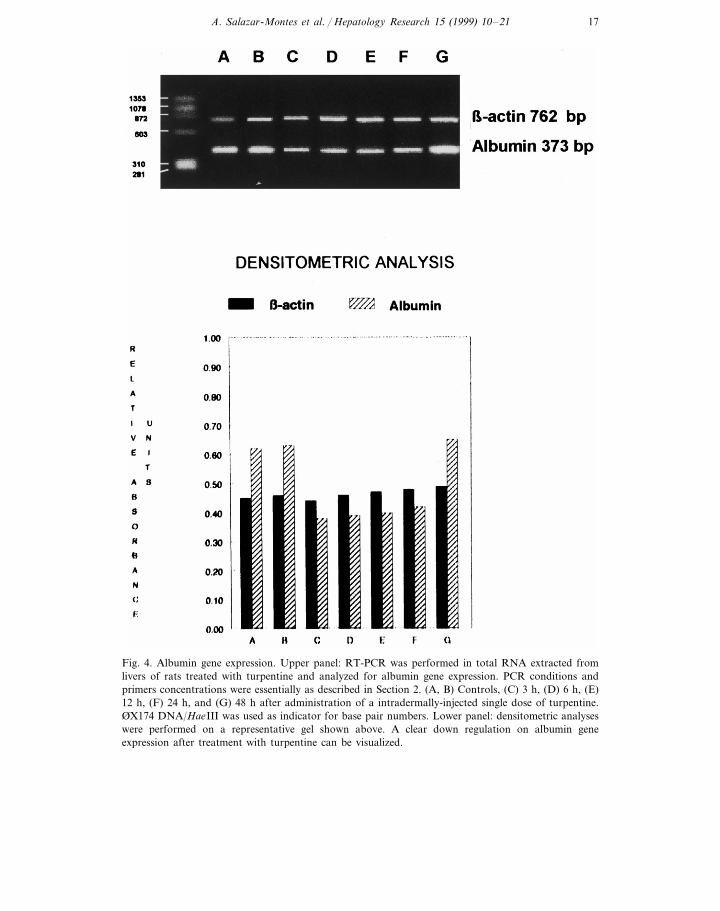
reactions beyond 40 cycles. Lanes A to D in Fig. 3 show identical results obtainedpreviously for Fig. 1 (upper panel). In order to confirm that an acute phaseresponse was being induced by a single injection of turpentine, we searched foralbumin mRNA transcription. It is well known that albumin is a negative acutephase protein decreasing during inflammation [28,29]. Fig. 4 clearly depicts thedecreased expression of a 373-bp amplified PCR product corresponding to albumingene. This fact is a characteristic indicator that an acute phase response is takingplace [17], and histograms in Fig. 4 lower panel, quantitatively summarize theclear-cut differences in albumin gene expression. Finally, Fig. 5 shows TNFa gene
Fig. 3. IL-6 gene expression in rat liver with acute and chronic damage induced by CCl4. Total RNAwas extracted from at least three different animals per group, intoxicated as described in Section 2. (A)Control; (B) 24 h after CCl4; (C) 48 h after CCl4; (D) 72 h after CCl4; (E) cirrhotic by chronicintoxication with CCl4 for 8 weeks; (F) cirrhotic plus acute intoxication. IL-6 expression is present only24 h after acute damage and in cirrhotic rats plus acute dose with CCl4.
A. Salazar-Montes et al. / Hepatology Research 15 (1999) 10–21 17
Fig. 4. Albumin gene expression. Upper panel: RT-PCR was performed in total RNA extracted fromlivers of rats treated with turpentine and analyzed for albumin gene expression. PCR conditions andprimers concentrations were essentially as described in Section 2. (A, B) Controls, (C) 3 h, (D) 6 h, (E)12 h, (F) 24 h, and (G) 48 h after administration of a intradermally-injected single dose of turpentine.ØX174 DNA/HaeIII was used as indicator for base pair numbers. Lower panel: densitometric analyseswere performed on a representative gel shown above. A clear down regulation on albumin geneexpression after treatment with turpentine can be visualized.

A. Salazar-Montes et al. / Hepatology Research 15 (1999) 10–2118
Fig. 5. TNFa gene expression. RT-PCR was performed in total RNA extracted from livers of ratsintoxicated with an acute dose of CCl4 to induce a brisk liver regeneration, and TNFa gene expressionwas measured. PCR conditions and primers concentrations were essentially as described before. (A)Control, (B) 6 h after CCl4 intoxication, (C) 12 h after CCl4 intoxication, (D) 48 h after intoxication withCCl4.
expression increased as early as 6 h and peaking at 48 h post-acute CCl4intoxication.
4. Discussion
Data obtained from several experimental models of inflammation has demon-strated that the mechanisms of inflammation are tightly regulated by the timelyexpression of IL-6 and TNFa [16]. Thus, a rapid release of secreted IL-Ia, TNFa
and IL-6 occurs in response to in vitro or in vivo LPS challenge in the mouse liver.This increase is associated with increased cytokine-specific mRNA transcripts [12].The increase of several transcriptional factors secondary to an induction of TNFa
mRNA occurring very early during the regenerative response, has clearly implicatedTNFa as a major effector of signal pathways of liver regeneration. Hence, TNFa
has a role to play in the early signaling pathways of liver regeneration induced inpartially hepatectomized animals [16]. Several lines of evidence invoke the TNFa
role in regulating secretion of IL-6 through a previous induction of NF-kb
transcription factors [16]. Since our data show TNFa gene expression inductiontakes place early after acute liver, TNFa mRNA expression might be a previousprerequisite for IL-6 gene transcription. We believe that in our experimental system,TNFa induces the activity of specific transcriptional factors that activate IL-6 genetranscription. We are in the process of investigating whether the nuclear proteinsactivity reflects the phosphorylation of already pre-existent proteins or if a ‘denovo’ proteins synthesis is involved.
Although we found that IL-6 mRNA is predominantly produced by Kupffer cellsin the CCl4-intoxicated animals (data not shown), other investigators have pro-posed that other non-parenchymal cells may contribute to the liver cytokine pool.Luster et al. [12] addressed this question in selectively depleted Kupffer cells usingDMDP-encapsulated (dichloromethylene diphosphonate) liposomes. They showedthat IL-Ia and TNFa secretion was markedly reduced in Kupffer cells depletedlivers, whereas secretion of IL-6 was reduced by 60%, suggesting that IL-6 secretion

A. Salazar-Montes et al. / Hepatology Research 15 (1999) 10–21 19
involves additional cell types. This is consistent with earlier immunohistochemicalstudies in which TNFa and IL-1a, but not IL-6, were detected in Kupffer cells afterLPS administration [27].
On the other hand, evidence has been produced where plasma IL-6 concentra-tions increased after partial hepatectomy in rats, peaking by 24 h, suggesting thatIL-6 mRNA induction should reach its highest peak 0–12 h post-hepatectomy. Thisin keeping with our results since we detected IL-6 mRNA peaking at 24 h, a findingthat could be explained by the fact that in CCl4-induced regeneration, hepatic celldamage precedes the proper establishment of the DNA synthesis initiation andconsequently hepatocyte proliferation. Furthermore, the physiological relevance ofthe dramatic decrease in IL-6 mRNA expression at 48 h remains to be elucidated.Here, we also demonstrated that IL-6 gene is not expressed in livers from CCl4cirrhotic animals, but strikingly, cirrhotic animals receiving an additional acutedose of CCl4 expressed high levels of hepatic IL-6 gene supporting our belief thatthis cytokine is a major element in the cytokine cascade induced by chemicalhepatectomy. Even when the mechanisms of regeneration (surgically or chemicallyinduced) are multifaceted, we believe they are different when compared with thoseinduced by a non-specific inflammatory stimuli. On this regard we did not find IL-6gene expression in the livers of animals treated with a single injection of turpentineto elicit an acute phase response. This finding is interesting since we corroboratedthe actual induction of the acute phase response by RT-PCR determination of thetypical drop in the levels of albumin mRNA. Finally, and in spite of the redun-dancy in the mechanisms the liver displays in regeneration, either after partialhepatectomy by surgery or chemically-induced, there is general consensus thatTNFa and IL-6 are important components of the pathways leading to regeneration.
Acknowledgements
These studies were supported in part by grants from CONACyT no. 4947-M9409and no. 0513P-M9506 to JAB.
References
[1] Baumann H, Muller-Eberhard U. Synthesis of hemopexin and custein protease inhibitor iscoordinately regulated by HSF-II and interferon B2 in rats hepatoma cells. Biochem Biophys ResCommun 1987;146:1218–28.
[2] Perimutter DH, May LT, Sehgal PB. Interferon b2/interleukin-6 modulates synthesis of alpha 1antitrypsin in human mononuclear phagocytes and in human hepatoma cells. J Clin Invest1989;84:138–44.
[3] Le J, Vilcek J. Interleukin 6: a multifunctional cytokine regulating immune reactions and the acutephase protein response. Lab Invest 1989;61(6):588–601.
[4] Nolan JP. Intestinal endotoxins as mediators of hepatic injury: an idea whose time has come again.Hepatology 1989;10:887–91.
[5] Sweeting J. Tumor necrosis factor in alcoholic hepatitis. Gastroenterology 1989;97:1056–7.

A. Salazar-Montes et al. / Hepatology Research 15 (1999) 10–2120
[6] Thiele DL. Tumor necrosis factor: the acute phase response and the pathogenesis of alcoholic liverdisease. Hepatology 1989;9:497–9.
[7] Anastassakos C, Alexander GJ, Wolstencroft RA, Avery JA, Portmann BC, Panayi GS, DumondeDC. Interleukin-1 and interleukin-2 activity in chronic hepatitis B virus infection. Gastroenterology1988;94:999–1005.
[8] Deviere J, Content J, Denys P, Vandernbussche P, Schandene L, Wybran J. High interleukin-6serum levels and increased production by leukocytes in alcoholic liver cirrhosis: correlation withIgA serum levels and lymphokine production. Clin Exp Immunol 1989;77:221–5.
[9] Khoruts A, Stahnke L, McClain CJ, Logan G, Allen JI. Circulating tumor necrosis factor,interleukin-1 and interleukin-6 concentrations in chronic alcoholic patients. Hepatology1991;13:267–76.
[10] Yoshioka K, Kakumu S, Arao M, Tautsumi Y, Inove M. Tumor necrosis factor alpha productionby peripheral blood mononuclear cells of patients with chronic liver disease. Hepatology1989;10:769–73.
[11] Martinez OM, Villanueva JC, Gershwin ME, Krams SM. Cytokine patterns and cytotoxicmediators in primary biliary cirrhosis. Hepatology 1995;21(1):113–9.
[12] Luster MI, Germolec DR, Yoshida T, Kayama F, Thompson M. Endotoxin-induced cytokine geneexpression and excretion in the liver. Hepatology 1994;19(2):480–8.
[13] Diehl AM, Clain CM. Ethanol decreases hepatic sensitivity to the hepatotrophic effects ofinterleukin-6 (IL-6) [Abstract]. Gastroenterology 1990;98:A582.
[14] Enayati P, Murray F, Brennan MD, Fong Y. Systemic and liver cytokine activation. Arch Surg1994;129:1159–64.
[15] Cressman DE, Greenbaum LE, DeAngelis RA, Ciliberto G, Furth EE, Poli V, Taub R. Liverfailureand defective hepatocyte regeneration in Interleukin 6-deficient mice. Science 1996;274:1379–83.
[16] Michaelopoulos GK, De Frances MC. Liver regeneration. Science 1997;276:60–6.[17] Greenwel P, Iraburo MJ, Reyes-Romero M, Meraz-Cruz N, Casado E, Solis-Herruzo JA, Rojkind
M. Induction of an acute phase response in rats stimulates the expression of a1(I) procollagenmessenger ribonucleic acid in their liver. Possible role of interleukin-6. Lab Invest 1995;72(1):83–91.
[18] Northermann W, Hattori M, Baffet G, Braciak TA, Fletcher GR, Abraham LJ, Gauldie J,Baumann M, Fey GH. Mol Biol Med 1990;7:273–85.
[19] Yamada Y, Kirillova I, Peschon JJ, Fausto N. Initiation of liver growth by tumor necrosis factor:deficient liver regeneration in mice lacking type I tumor necrosis factor receptor. Proc Natl Acad SciUSA 1997;94:1441–6.
[20] Armendariz-Borunda J, Seyer JM, Kang AH, Raghow R. Regulation of TGF-b gene expression inrat liver intoxicated with carbon tetrachloride. FASEB J 1990;4:215–21.
[21] Chomczynsky P, Sacchi N. Single-step method of RNA isolation by acid guanidium thiocyanate-phenol-chloroform extraction. Anal Biochem 1987;162:156–9.
[22] Armendariz-Borunda J, LeGros L Jr., Campollo O, Panduro A, Rincon AR. Antisense s-oligodeoxynucleotides down-regulate TGF-b production by Kupffer cells from CCl4 injured ratlivers. Biochim. Biophys. Acta 1997;1353:241–52.
[23] Siegling A, Lehmann M, Platzer C, Emmrich F, Volk HD. A novel multispecific competitorfragment for quantitative PCR analysis of cytokine gene expression in rats. J Immunol Methods1994;177:23–8.
[24] Coligan JE, Kruisbeek AM, Margulies DH, Shevach EM, Strober W. Curr Protoc Immunol1994;10:23–5.
[25] Ogawa K, Ohta T, Inagaki M, Nagase S. Identification of F344 rat hepatocytes transplanted withinthe liver of congenic analbuminemic rats by the polymerase chain reaction. Transplantation1993;56(1):9–15.
[26] Armendariz-Borunda J, Katai H, Jones CM, Seyer JM, Kang AH, Raghow R. Transforminggrowth factor b gene expression is transiently enhanced at a critical stage during liver regenerationafter CCl4 treatment. Lab Invest 1993;69:283–94.
[27] Chensue SW, Terbuh PD, Remick DG, Scales WE, Kunkel SL. In vivo biologic and immunohisto-chemical analysis of interleukin-1 alpha, beta and tumor necrosis factor during experimentalendotoxemia: kinetics kupffer cell expression and glucocorticoid effects. Am J Pathol 1991;138:395–402.

A. Salazar-Montes et al. / Hepatology Research 15 (1999) 10–21 21
[28] Lampreave F, Gonzalez Ramon N, Martinez-Ayensa S, Hernandez MA, Lorenzo HK, Garcia-GilA, Pineiro A. Characterization on the acute phase serum protein response in pigs. Electrophoresis1994;15(5):672–6.
[29] Yiangou M, Paraskeva E, Hsieh CC, Markou E, Victoratos P, Scouras Z, Papaconstantinou J.Induction of a subgoup of acute phase protein genes in mouse liver by hyperthermia. BiochimBiophys Acta 1998;1396(2):191–206.
.
Related Documents











