Chemical Defense Against Fouling in the Solitary Ascidian Phallusia nigra BOAZ MAYZEL*, MARKUS HABER, AND MICHA ILAN Department of Zoology, George S. Wise Faculty of Life Sciences, Tel Aviv University, Tel Aviv 6997801, Israel Abstract. The solitary ascidian Phallusia nigra is rarely fouled by epibionts. Here, we tested the antifouling activity of its crude extracts in laboratory and field assays. P. nigra extracts inhibited the growth of all eight tested environmen- tal bacteria and two of four laboratory bacteria. Extracts of the sympatric, but fouled solitary ascidian Herdmania mo- mus inhibited only one test bacterium. Scanning electron microscopy confirmed that the tunic surface of P. nigra is largely bacteria-free. Both ascidian extracts significantly inhibited the larval metamorphosis of the bryozoan Bugula neritina at the tested concentration range of 0.05–2 mg ml 1 . Both crude extracts were toxic to larvae of the brine shrimp Artemia salina at natural volumetric whole-tissue concentrations, but only P. nigra showed activity at 2 mg ml 1 and below (LC 50 1.11 mg ml 1 ). P. nigra crude extracts also significantly reduced the settlement of barna- cles, polychaetes, and algae in Mediterranean field assays and barnacle settlement in Red Sea trials. Comparisons between control experiments and pH values monitored in all experiments indicate that the observed effects were not due to acidity of the organic extracts. Our results show that P. nigra secondary metabolites have antifouling activities, which may act in synergy with previously proposed physi- ological antifouling mechanisms. Introduction In marine benthic environments, any exposed, unde- fended, long-lived surface becomes fouled (Wahl, 1989). The fouling process consists of multiple steps starting with the development of an organic film by adhesion of macro- molecules, followed by the settlement of microorganisms and the development of a microbial biofilm. The settlement of unicellular and multicellular eukaryotes establishes the mature fouling community composed of prokaryotes, fungi, diatoms, algae, protists, and invertebrates (Krug, 2006). Fouling also occurs on the surfaces of living organisms, and sessile marine invertebrates are particularly susceptible to fouling (Wahl, 1989). Although fouling organisms can be beneficial to their hosts, for example by offering camouflage and by providing protection from predators by their second- ary metabolites (Laudien and Wahl, 1999), the interaction is largely considered harmful. Among the negative effects are increased hydrodynamic drag on the host, which can lead to dislodgment of sessile hosts; and disruption of the host’s feeding, either by direct competition for food particles or by changing the water flow to the host, which is especially important for filter-feeding organisms (Stoecker, 1978; Wahl, 1989; Krug, 2006). Ascidians (phylum: Chordata, subphylum: Tunicata) are soft-bodied, sessile, filter-feeding organisms. They occur in all oceans in colonial and solitary forms. Fouling on ascid- ians can lead to the obstruction of the inhalant and exhalant siphons as well as to the clogging of the branchial basket, important to both respiration and feeding (Lambert, 1968; Koplovitz et al., 2011). Both naturally fouled and non- fouled ascidian species have been reported from various geographical regions (e.g., Stoecker, 1980; Uriz et al., 1991; Davis and White, 1994). Fouled ascidian species might use behavioral responses (e.g., actively closing the siphons or squirting water and particles from the branchial basket through apertures [Hoyle, 1953]) and physical responses (e.g., regular tissue sloughing [Goodbody, 1962] and mucus secretion [Wahl et al., 1998]) to overcome fouling. Short- lived ascidians might be able to grow and reproduce before becoming fouled (Stoecker, 1980). Among the taxa reported as epibionts on ascidians are bacteria, algae, bryozoans, Received 25 September 2013; accepted 5 September 2014. * To whom correspondence should be addressed. E-mail: bmayzel@ gmail.com Reference: Biol. Bull. 227: 232–241. (December 2014) © 2014 Marine Biological Laboratory 232

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Chemical Defense Against Fouling in the SolitaryAscidian Phallusia nigra
BOAZ MAYZEL*, MARKUS HABER, AND MICHA ILAN
Department of Zoology, George S. Wise Faculty of Life Sciences, Tel Aviv University,Tel Aviv 6997801, Israel
Abstract. The solitary ascidian Phallusia nigra is rarelyfouled by epibionts. Here, we tested the antifouling activityof its crude extracts in laboratory and field assays. P. nigraextracts inhibited the growth of all eight tested environmen-tal bacteria and two of four laboratory bacteria. Extracts ofthe sympatric, but fouled solitary ascidian Herdmania mo-mus inhibited only one test bacterium. Scanning electronmicroscopy confirmed that the tunic surface of P. nigra islargely bacteria-free. Both ascidian extracts significantlyinhibited the larval metamorphosis of the bryozoan Bugulaneritina at the tested concentration range of 0.05–2 mgml�1. Both crude extracts were toxic to larvae of the brineshrimp Artemia salina at natural volumetric whole-tissueconcentrations, but only P. nigra showed activity at 2 mgml�1 and below (LC50 � 1.11 mg ml�1). P. nigra crudeextracts also significantly reduced the settlement of barna-cles, polychaetes, and algae in Mediterranean field assaysand barnacle settlement in Red Sea trials. Comparisonsbetween control experiments and pH values monitored in allexperiments indicate that the observed effects were not dueto acidity of the organic extracts. Our results show that P.nigra secondary metabolites have antifouling activities,which may act in synergy with previously proposed physi-ological antifouling mechanisms.
Introduction
In marine benthic environments, any exposed, unde-fended, long-lived surface becomes fouled (Wahl, 1989).The fouling process consists of multiple steps starting withthe development of an organic film by adhesion of macro-molecules, followed by the settlement of microorganisms
and the development of a microbial biofilm. The settlementof unicellular and multicellular eukaryotes establishes themature fouling community composed of prokaryotes, fungi,diatoms, algae, protists, and invertebrates (Krug, 2006).Fouling also occurs on the surfaces of living organisms, andsessile marine invertebrates are particularly susceptible tofouling (Wahl, 1989). Although fouling organisms can bebeneficial to their hosts, for example by offering camouflageand by providing protection from predators by their second-ary metabolites (Laudien and Wahl, 1999), the interaction islargely considered harmful. Among the negative effects areincreased hydrodynamic drag on the host, which can lead todislodgment of sessile hosts; and disruption of the host’sfeeding, either by direct competition for food particles or bychanging the water flow to the host, which is especiallyimportant for filter-feeding organisms (Stoecker, 1978;Wahl, 1989; Krug, 2006).
Ascidians (phylum: Chordata, subphylum: Tunicata) aresoft-bodied, sessile, filter-feeding organisms. They occur inall oceans in colonial and solitary forms. Fouling on ascid-ians can lead to the obstruction of the inhalant and exhalantsiphons as well as to the clogging of the branchial basket,important to both respiration and feeding (Lambert, 1968;Koplovitz et al., 2011). Both naturally fouled and non-fouled ascidian species have been reported from variousgeographical regions (e.g., Stoecker, 1980; Uriz et al., 1991;Davis and White, 1994). Fouled ascidian species might usebehavioral responses (e.g., actively closing the siphons orsquirting water and particles from the branchial basketthrough apertures [Hoyle, 1953]) and physical responses(e.g., regular tissue sloughing [Goodbody, 1962] and mucussecretion [Wahl et al., 1998]) to overcome fouling. Short-lived ascidians might be able to grow and reproduce beforebecoming fouled (Stoecker, 1980). Among the taxa reportedas epibionts on ascidians are bacteria, algae, bryozoans,
Received 25 September 2013; accepted 5 September 2014.* To whom correspondence should be addressed. E-mail: bmayzel@
gmail.com
Reference: Biol. Bull. 227: 232–241. (December 2014)© 2014 Marine Biological Laboratory
232

hydroids, soft corals, and sponges (Stoecker, 1980; Davisand White, 1994; Wahl et al., 1994).
Chemical defense against fouling has been found in sev-eral ascidians and against various fouling organisms. As-cidian crude extracts have been shown to inhibit fouling bybarnacles and bryozoans (Davis and Wright, 1989; Bryan etal., 2003) as well as the growth and settlement of environ-mental bacteria (Wahl et al., 1994; Bryan et al., 2003) anddiatoms (McClintock et al., 2004; Koplovitz et al., 2011).Furthermore, antifouling and antibacterial compounds havebeen isolated from ascidians (Davis and Wright, 1990;Tsukamoto et al., 1994; Davis and Bremner, 1999). Tunicsurface-associated bacteria from ascidians have been shownto produce antibacterial and antifouling compounds in lab-oratory cultures (Olguin-Uribe et al., 1997; Holmstrom etal., 1998, 2002), and symbiotic bacteria have been assumedto be the real producers of some known bioactive com-pounds originally isolated from their ascidian hosts (Sim-mons et al., 2008).
In the present study, we investigated the antifoulingchemical defense potential of the solitary tunicate Phallusianigra (�Ascidia nigra) (Savigny, 1816). P. nigra is a cos-mopolitan species occurring in the West Atlantic from Flor-ida throughout the Caribbean (Bermuda, Jamaica) to Brazil,throughout the Mediterranean Sea, and in the Red Sea fromYemen to the northern tip of the Gulf of Aqaba. Along theIsraeli Mediterranean Sea, it is commonly found at depthsbetween 1 and 30 m as part of the fouling community onhard substrates such as rocks or man-made structures (Shen-kar and Loya, 2009). Even though P. nigra lives up to 2years (Goodbody, 1962) and in environments with highfouling pressure (Goodbody, 1965), it has frequently beendescribed as free of epibionts (Goodbody, 1962; Stoecker,1978, 1980; authors’ own observations (Fig. 1A); see alsofigure 2b in Shenkar and Loya, 2009). Early studies sug-gested that P. nigra regularly sloughs off its external layersand thus cleans itself from epibionts (Hecht, 1918; Good-body, 1962). High vanadium concentrations and low tunicsurface pH (�2) were also suggested as defenses againstfouling organisms (Stoecker, 1978). These assumptionswere challenged by later studies as ascidian species con-taining high vanadium concentrations, comparable to thosein P. nigra, were found to be fouled (Stoecker, 1980). Tunicsurface acidity as an antifouling mechanism in ascidianswas also later questioned due to the presence of heavilyfouled ascidians with low tunic surface pH (Parry, 1984;Davis and Wright, 1989).
The role of secondary metabolites in the defense of P.nigra, especially against fouling, is poorly understood. Re-sults from predator feeding-deterrence assays with chemicalextracts indicated that while whole-tissue extracts did notdeter fish feeding, active (i.e., deterrent) compounds werepresent in the extracts from gonads and the combined tissueof the branchial basket and gut (Pisut and Pawlik, 2002).
Furthermore, crude extracts of P. nigra were active againstvarious standard laboratory bacteria (Jaffarali et al., 2008;Amutha et al., 2010), but not against four known pathogensof marine invertebrates (Odate and Pawlik, 2006).
Given the unclear role of secondary metabolites in thefouling defense of P. nigra, its clean surface, and the goodtrack record of ascidians as a source of antifouling com-pounds, we investigated the antifouling activity of the sec-ondary metabolites of P. nigra in laboratory and field as-says. For comparison, we assayed organic extracts from thesolitary ascidian Herdmania momus (Savigny, 1816) in thelaboratory assays. This latter species is usually fouled innature (Davis and White, 1994; Shenkar and Loya, 2009;authors’ own observation (Fig. 1D)) and is sympatric withP. nigra along the Israeli Mediterranean and Red Sea coasts.Since every step in the fouling process affects the subse-quent ones (Krug, 2006; Wahl, 2008), we first tested theantibacterial activity of extracts against a panel of sympatricand laboratory bacteria, and used scanning electron micros-copy to determine natural bacterial fouling on the tunicsurface of P. nigra and H. momus. The ability of extracts toinhibit metamorphosis of a representative macro-foulinginvertebrate was examined against larvae of the sympatricbryozoan Bugula neritina (Linnaeus, 1758). General toxic-ity of crude extracts was tested against larvae of the brineshrimp Artemia salina (Linnaeus, 1758). Finally, extracts ofP. nigra from the Red Sea and the Mediterranean Sea wereexposed to natural fouling communities in field assays intheir respective locations. In order to decouple the antifoul-ing effect of low pH from that of secondary metabolites, thepH of the extracts was monitored in all assays, and controlexperiments were performed for all laboratory assays.
Materials and Methods
Sample collection and extraction
The ascidians Phallusia nigra and Herdmania momuswere collected by scuba from various sites along the IsraeliMediterranean coast at depths from 5 to 30 m. P. nigrasamples used in Red Sea experiments were collected inEilat, northern Red Sea, Israel. After each dive, the volumeof the collected ascidians was determined by seawater dis-placement. Samples were then stored at �20 °C until ex-traction took place.
Frozen ascidians of known (wet) volume were cut into1-cm3 cubes and covered with a mixture of 1:1 dichloro-methane and methanol in a flask. The flask was kept at 4 °Cfor 24 h, during which it was periodically shaken to ensurethat the ascidian tissue was equally exposed and extractedby both solvents. The resulting phases methanol:water anddichloromethane were separated, and both phases were fil-tered (Whatman, 8-�m pore size). Each phase was evapo-rated to dryness using a rotary evaporator (Buchi RotavaporR-114) with a water bath at a temperature below 40 °C.
233B. MAYZEL ET AL.

Extracts of both phases were combined into the same glassbulb. The extraction was repeated a second time, followedby a final extraction with methanol at 4 °C for 6 h. Allphases were combined in the same glass vial, resulting inthe crude extract, which was weighed and stored at �20 °Cuntil use in the assays. Natural concentrations were calcu-lated by dividing the crude extract weight by the wet ascid-ian volume.
Scanning electron microscopy
Collected ascidians were rinsed with 0.22-�m-filteredseawater, fixed in 2.5% glutaraldehyde in seawater, and keptat 4 °C until further preparation. From three individuals of
each species, three 1 � 1-cm samples were taken andwashed with PBS, dehydrated in an EtOH gradient followedby critical-point drying with liquid CO2, and coated withgold-palladium. The tunic surfaces of the samples werescanned for bacteria at magnifications ranging from 100�to 7000� using a Jeol SEM JSM 840A.
Antibacterial assay
Crude organic extracts from P. nigra and H. momus weretested for antibacterial activity against eight environmentalbacteria and four standard laboratory bacteria (Table 1).Isolation and identification of the environmental bacteriafrom the Israeli Mediterranean coast are described in Haber
Figure 1. Comparative images of the sympatric ascidians Phallusia nigra and Herdmania momus. (A) P.nigra free of fouling in natural habitat. SEM images of P. nigra at 4000� magnification (B) and at 7000�magnification (C), tunic is free of bacteria. (D) H. momus fouled by epibionts in natural environment. SEMimages of H. momus at 4000� magnification (E) and at 7000� magnification (F), bacteria marked with arrowsare evident on tunic.
234 ANTIFOULING DEFENSE IN P. NIGRA

et al. (2011). The test protocol was adapted from Kelman etal. (2001). Test bacteria were cultured overnight in liquidmedium (Marine Broth 2216 and LB for environmental andlaboratory bacteria, respectively). Culture densities weremeasured at 620 nm using a Turner SP-830 spectrophotom-eter. The cultures were adjusted to an optical density of 0.04and diluted 1:100. From the diluted cultures, 100 �l wasadded to a 96-well plate containing 100 �l of appropriateliquid medium, DMSO (final concentration 0.5% v/v), anda series of extract concentrations (ranging from 0.625 to 20mg ml�1, which represents 3% to 100% and 6% to 200% ofnatural volumetric concentration of P. nigra and H. momus,
respectively, and including a solvent control with 0 mgml�1). Each extract concentration and control was assayedin triplicate. Initial optical density was measured at 620 nmon a GENESIS Workstation 200 (Tecan) using SPECTRA-Fluor Plus (Tecan), after 5 s of shaking and 2 s of settling.Plates were incubated at 30 °C with shaking (80 rpm).Optical density measurements were repeated after 24 and48 h. Minimal inhibitory concentrations were determined asthe smallest amount of extract needed to inhibit growth bymore than 85% compared to growth in the controls withoutextract.
Bugula neritina larvae assay
Colonies of B. neritina were collected from marinas inTel Aviv and Herzelia, Israel (Mediterranean Sea). Aftertransfer to the laboratory, colonies were maintained in anaquarium. To promote larval release, colonies were kept inthe dark for 24 h and then exposed to light (Dahms et al.,2007). The released larvae were collected during the next 30min by using a glass pipette and were transferred into0.22-�m-filtered seawater. Larvae were then immediatelypipetted into the assay wells. The experiment was conductedin 24-well tissue-culture plates (Corning). Extracts weredissolved in methanol and diluted to the required concen-trations (see below). An aliquot of 2.7 ml of dissolvedextract was pipetted into each well and evaporated to dry-ness. Methanol only was used for control wells. Three tofour larvae were pipetted into each well with 2.7 ml ofseawater. Larval vitality state was recorded as alive-swim-ming, alive-metamorphosed, and dead after 0, 24, and 48 h.The effect of P. nigra and H. momus extracts at concentra-tions between 0.05 and 2 mg ml�1 was determined by
Figure 2. Bugula neritina larval metamorphosis inhibition assay byextracts of tunicates (Phallusia nigra and Herdmania momus) after 24 h.Extracts of both species significantly inhibited the metamorphosis at allconcentrations compared to controls without extracts. Kruskal-Wallis test,P � 0.01 at all concentrations; letters indicate groupings based on multiplecomparison tests. Gray, control; white, H. momus extract; black, P. nigraextract.
Table 1
Minimal inhibitory concentrations (in mg ml�1) of tunicate chemical extracts in antibacterial assay after 24 h
Phylum; (Class) Test bacterium GenBank accn. no. Origin P. nigra H. momus
Gram positiveFirmicutes Bacillus subtilis Lab 20 —Firmicutes Staphylococcus aureus Lab 20 —Actinobacteria Kocuria sp. ESY10 GU479627 Sed 5 —Firmicutes Lysinibacillus sp. ESY09 GU059941 Sed 10 —Firmicutes Sporosarcina sp. NB90 GU479626 Sed 5 —
Gram negativeProteobacteria; (�-Proteobacteria) Pseudomonas aerigunosa PA01 Lab — —Proteobacteria; (�-Proteobacteria) Escherichia coli GM1655 Lab — —Proteobacteria; (�-Proteobacteria) Pseudomonas sp. NB86 GU479630 Seaw 10 —Proteobacteria; (�-Proteobacteria Proteobacteria) Sulfitobacter sp. ESY17 GU479629 Seaw 10 —Proteobacteria; (�-Proteobacteria) Vibrio sp. ESY12 GU479628 Sed.w 10 —Proteobacteria; (�-Proteobacteria Proteobacteria) Erythrobacter sp. ESY19 Seaw 5 —Proteobacteria; (�-Proteobacteria Proteobacteria) Loktanella sp. ESY23 Seaw 5 10
The natural volumetric tissue concentrations of Herdmania momus and Phallusia nigra were 9 and 20 mg ml�1, respectively.Origin of test bacteria is indicated by: Lab, Standard laboratory test bacterium; Sed, Sediment; Sed.w, Sediment water; Seaw, Seawater.—, no inhibition.
235B. MAYZEL ET AL.

comparison with the larval development in the controlwells. At least 12 wells were analyzed for every testedconcentration and controls, and the percentage of metamor-phosed larvae was calculated for each well. As obtaineddata were not normally distributed, significance of observeddifferences between wells containing P. nigra and H. mo-mus extracts at the various concentrations and controls wasdetermined using a Kruskal-Wallis test, followed by multi-ple comparison tests once significant differences were found.All statistical analyses were done using Statistica 9 software.
Brine shrimp assay
Toxicity of P. nigra and H. momus crude extracts wastested against nauplii of the brine shrimp Artemia salina.Brine shrimp were hatched from eggs in an aquarium andcollected using a glass pipette into 0.22-�m-filtered artifi-cial seawater (Instant Ocean). Extracts were dissolved inDMSO and diluted to the required concentrations (finalDMSO concentration in the assay 1% v/v). Ten to fifteenbrine shrimp larvae were transferred to a 2-ml Eppendorftube with a minimum amount of artificial seawater. Thevolume was adjusted to 1.2 ml with artificial seawater, and0.3 ml of extract was added. A DMSO control withoutextract was prepared accordingly. Each extract concentra-tion and the control were tested in triplicate. The numberand vitality state—alive (any movement of the larvae) ordead)—of the brine shrimp larvae in each tube were re-corded after addition of extract or DMSO control at 0, 24,and 48 h. The extracts of both ascidian species were testedat natural whole-tissue volumetric concentration and 2 mgml�1 (� 10% and 20% of natural whole-tissue volumetricconcentration for P. nigra and H. momus, respectively). Inaddition, a range of concentrations (0.01–5 mg ml�1) wastested for the P. nigra crude extract to determine the LC50,which was calculated by trimmed Spearman-Kaber estima-tion using the TSK program ver. 1.5 as provided by theecological exposure research division of the U.S. Environ-mental Protection Agency (www.epa.gov/eerd/stats2.htm asaccessed 7 May, 2012).
Field experiments
Agar gel surfaces containing the crude extract of P. nigrawere used as substrate for the settlement of fouling organ-isms in natural settings. The experimental protocol wasadapted from reports using similar methods (Slattery et al.,1995; Henrikson and Pawlik, 1995). Agar solution (5% w/vDifco, granulated agar) was autoclaved and kept at 50 °Cuntil extract addition. The dry extract obtained from 50 mlof ascidian tissue was dissolved in 3 ml of methanol and mixedinto 50 ml of agar solution, resulting in natural volumetricextract concentration in the gel. Control gels contained only3 ml of methanol. To prepare the settlement assay, 15 ml ofagar solution was poured into each well of a six-well tissue-
culture plate. In each plate three wells contained agar withextract, and three wells were filled only with control agar.The order of the control and treatment wells was random-ized. The plates were secured to plastic meshes, which wereattached with cable ties to piers at a depth of 0.5–1 m, nextto naturally fouled surfaces at the Interuniversity Institute inEilat (Red Sea) and Herzelia Marina (Mediterranean Sea).In each experiment the plates were deployed and monitoredweekly for the occurrence of damages and disturbances tothe plates for a period of 4 weeks to allow for settlement anddevelopment of fouling organisms. After retrieval, plateswere examined for macrofouling under a stereoscope, andthe number of epibionts in each well was recorded. Foulingorganisms were divided into three main categories: mac-roalgae, barnacles, and sessile polychaetes (Serpulidae). Totest for seasonal variation of fouling organisms, this exper-iment was conducted eight times throughout the year in theMediterranean and twice in the Red Sea (dates of theexperiments are given in the figures).
The three groups of settled epibionts (macroalgae, barna-cles, and polychaetes) were analyzed separately. Since thenumber of settled epibionts in each well was too low forstatistical analysis, the data from all wells of the sametreatment in each plate were combined. In each plate thedistribution of settled epibionts on wells containing extractwas compared to their distribution in control wells by areplicated goodness of fit test (Sokal and Rolf, 1995). Foreach group of epibionts, only experiments in which settle-ment occurred were included in the analysis.
During the deployment of the agar settlement surfaces,the extract slowly diffused out of the agar. To determine theconcentration of extract in the agar from the initial deploy-ment until the end of the experiment, six tissue-culturemulti-well plates with agar gel containing extract and con-trols were prepared and deployed. The first plate was testedbefore deployment and the others after 1, 7, 14, 21, and 28days. From each plate the three agar gels containing theextract were removed from the wells, combined, and thenre-extracted. We used the same protocol as in the initialextraction, except that a butanol/water separation phase wasadded to the original extraction protocol to remove saltsfrom the extracts. The same procedure was applied to thematching control wells. Once salts were removed, theweight of the extract from control wells was subtracted fromthe weight of extract in the matching treatment wells. Theresulting “net” extract weights indicated the change in theamount of P. nigra extract due to leakage from the agar gelsthroughout the experiment. This experiment was performedtwice, once in May and once in October during the researchduration. Thin-layer chromatography (TLC) was used toverify that the same chemical compounds were found in theextract throughout the settlement experiments. Silica plates(Kieselgel 60 F254, Merck) were loaded with samples fromtreatment and control wells in chronological order of their
236 ANTIFOULING DEFENSE IN P. NIGRA

deployment. A toluene/dichloromethane 1:1 mixture wasused as the mobile phase. TLC profiles were compared aftervisualization under UV light (254 nm) and after developingthe TLC plate with vanillin stain (6 g vanillin in 1.5 mlconcentrated sulfuric acid and 95 ml 96% ethanol).
Measurement of pH and pH control experiments
The pH in the assay wells was recorded at the end of alllaboratory assays using pH indicator paper (Merck 0–14and Merck 5.0–10.0). In the field assay, the pH of the agarmixtures was measured during plate preparation, and thesurface pH of the plates was measured again after 24 h ofdeployment. For laboratory assays, control experimentswere performed with media adjusted to pH 5.5–8.5 (anti-bacterial assay) and seawater adjusted to pH 6.0–8.0 (B.neritina larvae assay, brine shrimp assay) in 0.5-unit incre-ments according to the range of pH measured at the end ofthe assays. These measurements and experiments were per-formed to ensure the obtained results were not influenced byextract acidity.
Results
Natural bacterial fouling on ascidian tunics andantibacterial assay
Tunic samples of Herdmania momus were heavily over-grown by epibionts. As this did not allow a quantitativecomparison between both tunicate species, only a qualita-tive screen of the samples was performed. Scanning electronmicroscopy examination showed that the tunic of Phallusianigra was largely bacteria-free, with the exception of a fewclusters of coccoid-shaped bacteria. In contrast, variousbacteria were clearly evident on the tunics of the sympatricH. momus (Fig. 1).
Chemical extract of the two ascidian species showed verydifferent activity profiles in the antibacterial assay (Table 1).The P. nigra extract inhibited the growth of all environ-mental test bacteria at 5–10 mg ml�1 (25%–50% of naturalwhole-tissue volumetric concentration) and the two gram-positive laboratory test bacteria at 20 mg ml�1. The H.momus extract inhibited only the gram-negative environ-mental alpha-proteobacterium Loktanella sp. ESY23 at 10mg ml�1, a concentration slightly above natural whole-tissue volumetric concentration.
The pH in tests with laboratory bacteria conducted in LBmedium was 8.5 for all extract concentrations of both as-cidian species and the controls. Growth of laboratory bac-teria was not inhibited in LB medium adjusted to pH 5.5–8.5. In tests with environmental bacteria cultured in MarineBroth 2216, the pH was 8.5 in the controls and all H. momusconcentrations. The pH of extracts of P. nigra in MarineBroth 2216 at 20, 10, and 5 mg ml�1 was 5.5, 6.5, and 7.5,respectively. Marine Broth 2216 adjusted to pH 5.5 to 8.5did not inhibit the growth of half of the environmental
bacteria. The growth of the remaining four bacteria strainsESY10, ESY12, ESY17, and ESY23 was only inhibited atpH 5.5, but not at pH 6.0 to 8.5. The pH at the minimalinhibitory concentrations for these bacteria was 6.5 (ESY12,ESY17) and 7.5 (ESY10, ESY23), respectively.
Bugula neritina assay
Significant differences in the metamorphosis of B. ner-itina larvae between treatments were found at all concen-trations tested, which were well below natural whole-tissueconcentrations (Kruskal-Wallis test H2 � 20, n � 48, P �0.005 for all concentrations). These differences were due tothe lower number of metamorphosed larvae in wells withthe ascidian extracts compared to the controls (Fig. 2). Theaverage number of metamorphosed larvae was always lowerin tests with P. nigra crude extracts than in the correspond-ing tests with H. momus crude extracts. However, thisdifference was only significant at concentrations of 0.1 and0.25 mg ml�1 (Kruskal-Wallis test, H2 � 25.718, P � 0.025and Kruskal-Wallis test, H2 � 27.561, P � 0.030, respec-tively). At the lowest tested concentration of 0.05 mg ml�1
more than 50% of the B. neritina larvae metamorphosed inwells with both extracts (52% and 62% in wells with P.nigra and H. momus extracts, respectively).
Throughout the experiment pH 7 was recorded in allwells containing P. nigra extracts, pH 7–7.5 in wells con-taining H. momus extract, and pH 8 in control wells. Controlexperiments performed in seawater adjusted to pH 6.0 to 8.0showed larval metamorphosis and settlement was signifi-cantly inhibited only at pH 6, but not at pH 6.5–8.0(Kruskal-Wallis test, H4 � 24.78, P � 0.0001, multiplecomparisons test P � 0.03 to �0.001).
Brine shrimp assay
H. momus and P. nigra crude extracts killed all brineshrimp larvae at their respective natural whole-tissue volu-metric concentration of 9 and 20 mg ml�1. At 2 mg ml�1,however, only the P. nigra crude extract was active andkilled on average 82.2% (� 10.2% STD), whereas mortalityin the H. momus crude extract was almost the same as in thecontrol (Table 2). Based on the tested range of concentra-tions (Table 2) the LC50 of the P. nigra crude extract wasfound to be 1.11 mg ml�1 with a 95% confidence of0.94–1.32 mg ml�1 (trimmed Spearman-Kaber estimation).The seawater pH after addition of the P. nigra crude extractswas between 6 and 7. Survival of brine shrimp was notaffected in seawater adjusted to pH 6.0–8.0.
Field assay results
P. nigra crude extract was tested at natural concentrationin the field against various epibionts present in the ascid-ians’ natural environment. During the 28 days of the Med-iterranean field experiment, as much as 52% of the original
237B. MAYZEL ET AL.

extract weight leaked out from the gel surfaces (Fig. 3).Qualitative TLC analysis of the P. nigra extract content ofthese agar gels during the settlement trials showed that thesame chemical pattern was present before the gels wereimmersed in seawater and throughout the the experiment.The TLC analysis of the control agar gels showed nocomparable spots. Therefore all active secondary metabo-lites originally found in P. nigra extract were still present inthe settlement gels at the end of the field experiments.During gel preparation, the pH for agar gels with H. momusextract was recorded as 6.1, agar with P. nigra extract waspH 4, and the pH of agar control gels was 5.5. However, thepH for all gels after a 24-h exposure to seawater wasrecorded as 7.5.
In the Mediterranean settlement trials, barnacle settle-
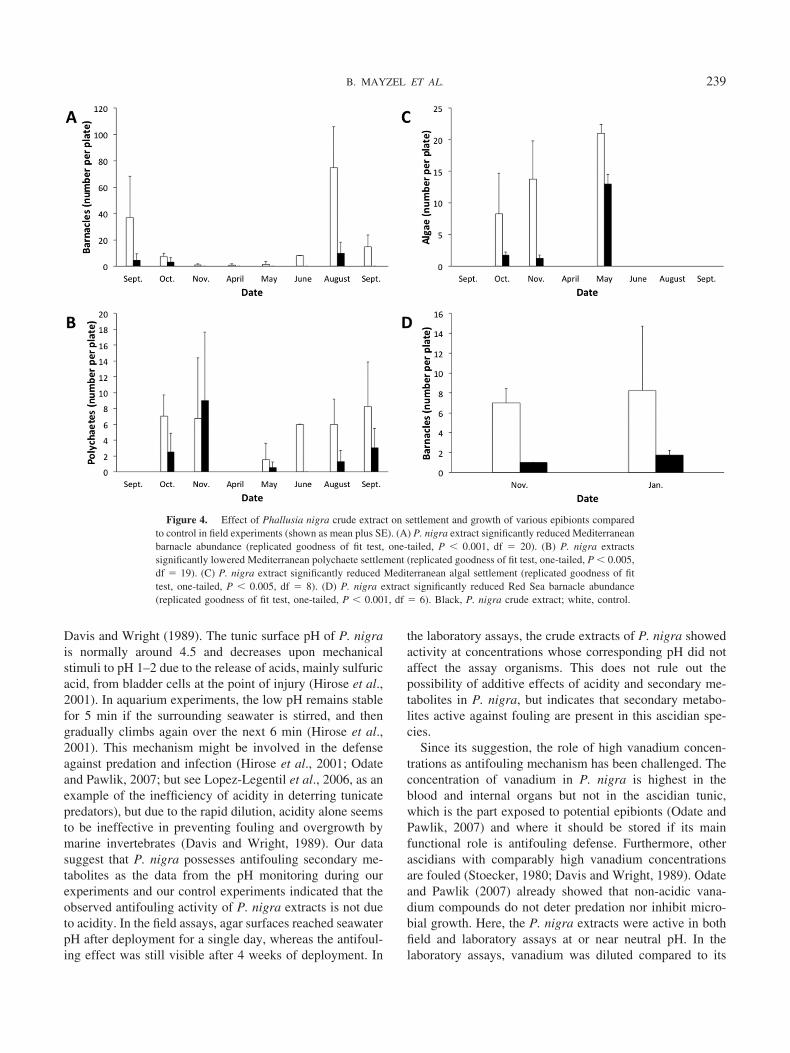
ment was most abundant from June to October, while verylittle settlement occurred from October till March (Fig. 4A).Barnacle settlement on P. nigra extract wells was signifi-cantly lower than on control gels throughout the experiment(replicated goodness of fit test, one-tailed, P � 0.001, df �20). Settlement of polychaetes was most abundant betweenMay and November, and none occurred from December tillApril (Fig. 4B). Statistical analysis of the results showed asignificant difference in polychaete settlement betweenwells containing P. nigra extract and control wells (repli-cated goodness of fit test, one-tailed, P � 0.005, df � 19).Analysis of all statistic parameters of this test showed thatthis was due to a much lower polychaete settlement on gelscontaining the P. nigra extract, in all but one of the exper-iments. Algal settlement was noted from October to May(Fig. 4C). A significant difference was found between theamount of algae on P. nigra extract wells and on controlwells (replicated goodness of fit test, one-tailed, P � 0.001,df � 8) due to a much lower algal settlement on gelscontaining the P. nigra extract, throughout all the experi-ments.
Barnacle settlement was observed in both Red Sea trials(Fig. 4D). Barnacles settled significantly less on gels with P.nigra extract than on control gels, throughout all the exper-iments (replicated goodness of fit test, one-tailed, P �0.001, df � 6).
Discussion
Previous studies on the ascidian Phallusia nigra sug-gested that it keeps its surface free of fouling organisms bytissue sloughing (Hecht, 1918; Goodbody, 1962), high va-nadium concentrations (Stoecker, 1978), and the acidic tu-nic surface (Stoecker, 1978; 1980; Odate and Pawlik, 2007).After confirming that tunics of Mediterranean P. nigra arelargely free of fouling organisms, we examined the role ofsecondary metabolites in antifouling defense of P. nigra.Our results demonstrated that P. nigra crude extracts inhib-ited the growth of environmental bacteria, inhibited themetamorphosis of the bryozoan Bugula neritina, and re-duced settlement by polychaetes, algae, and barnacles infield experiments. These activities were not due to low pHof the extracts, as shown by pH monitoring and controlexperiments. Therefore, secondary metabolites defend P.nigra against the settlement and growth of fouling organ-isms on its surface. A similar role for secondary metaboliteshas previously been shown in sponges, corals, bryozoans,and other ascidian species (Bryan et al., 2003; Haber et al.,2011; Fusetani 2012).
The idea that acids act as an antifouling mechanism inascidians has already been challenged by Parry (1984) fortwo reasons: (i) the rapid dilution of acids by seawater and(ii) the presence of heavily fouled ascidian species with lowtunic acidity (pH � 2), which was also later reported by
Table 2
Evaluation of crude extract toxicity on brine shrimp
Concentration(mg ml�1)
P. nigraAverage mortality
(%) (� STD)
H. momusAverage mortality
(%) (� STD) pH
0 (Seawater Control) 3.95 (� 0.87) 80 (DMSO Control) 4.5 (� 0.07) 7.50.01 0.00 (� 0.0) n.d. 7.50.05 0.00 (� 0.0) n.d. 7.50.10 2.22 (� 3.9) n.d. 7.00.25 3.13 (� 4.4) n.d. 7.00.50 3.33 (� 5.8) n.d. 7.51.00 34.95 (� 22.1) n.d. 7.02.00 82.22 (� 10.2) 9.17 (�4.6) n.d.5.00 100.00 (� 0.0) n.d. 6.5Natural (volumetric) 100.00 (� 0.0) 100.00 (� 0.0) n.d.
pH after addition of Phallusia nigra extract as measured by pH paper.The natural volumetric tissue concentrations of Herdmania momus andPhallusia nigra were 9 and 20 mg ml�1, respectively. n.d., not determined.
Figure 3. Changes in Phallusia nigra extract concentration in agarsettlement plates during 28 days deployment in the field. Results are meanpercentage (�SE) of extract mass after removal of salt content, calculatedfrom two field experiments. Only a single point is shown for day 21 due toloss of a plate.
238 ANTIFOULING DEFENSE IN P. NIGRA
Davis and Wright (1989). The tunic surface pH of P. nigrais normally around 4.5 and decreases upon mechanicalstimuli to pH 1–2 due to the release of acids, mainly sulfuricacid, from bladder cells at the point of injury (Hirose et al.,2001). In aquarium experiments, the low pH remains stablefor 5 min if the surrounding seawater is stirred, and thengradually climbs again over the next 6 min (Hirose et al.,2001). This mechanism might be involved in the defenseagainst predation and infection (Hirose et al., 2001; Odateand Pawlik, 2007; but see Lopez-Legentil et al., 2006, as anexample of the inefficiency of acidity in deterring tunicatepredators), but due to the rapid dilution, acidity alone seemsto be ineffective in preventing fouling and overgrowth bymarine invertebrates (Davis and Wright, 1989). Our datasuggest that P. nigra possesses antifouling secondary me-tabolites as the data from the pH monitoring during ourexperiments and our control experiments indicated that theobserved antifouling activity of P. nigra extracts is not dueto acidity. In the field assays, agar surfaces reached seawaterpH after deployment for a single day, whereas the antifoul-ing effect was still visible after 4 weeks of deployment. In
the laboratory assays, the crude extracts of P. nigra showedactivity at concentrations whose corresponding pH did notaffect the assay organisms. This does not rule out thepossibility of additive effects of acidity and secondary me-tabolites in P. nigra, but indicates that secondary metabo-lites active against fouling are present in this ascidian spe-cies.
Since its suggestion, the role of high vanadium concen-trations as antifouling mechanism has been challenged. Theconcentration of vanadium in P. nigra is highest in theblood and internal organs but not in the ascidian tunic,which is the part exposed to potential epibionts (Odate andPawlik, 2007) and where it should be stored if its mainfunctional role is antifouling defense. Furthermore, otherascidians with comparably high vanadium concentrationsare fouled (Stoecker, 1980; Davis and Wright, 1989). Odateand Pawlik (2007) already showed that non-acidic vana-dium compounds do not deter predation nor inhibit micro-bial growth. Here, the P. nigra extracts were active in bothfield and laboratory assays at or near neutral pH. In thelaboratory assays, vanadium was diluted compared to its
Figure 4. Effect of Phallusia nigra crude extract on settlement and growth of various epibionts comparedto control in field experiments (shown as mean plus SE). (A) P. nigra extract significantly reduced Mediterraneanbarnacle abundance (replicated goodness of fit test, one-tailed, P � 0.001, df � 20). (B) P. nigra extractssignificantly lowered Mediterranean polychaete settlement (replicated goodness of fit test, one-tailed, P � 0.005,df � 19). (C) P. nigra extract significantly reduced Mediterranean algal settlement (replicated goodness of fittest, one-tailed, P � 0.005, df � 8). (D) P. nigra extract significantly reduced Red Sea barnacle abundance(replicated goodness of fit test, one-tailed, P � 0.001, df � 6). Black, P. nigra crude extract; white, control.
239B. MAYZEL ET AL.

natural volumetric concentration (e.g., up to 400 times in theB. neritina assay), while activity was still observed and pHwas non-acidic. Therefore our results suggest that secondarymetabolites present in the crude extracts are involved in thedefense of P. nigra against fouling by bacteria, bryozoans,polychaetes, and barnacles.
Testing ecologically relevant concentrations is a majorchallenge in chemical ecology. We used whole-tissue vol-umetric concentrations, which probably underestimate theconcentrations experienced by fouling organisms becausecompounds might be accumulated at the tunic surface. Yeteven in the antibacterial assay, where activity was found atrelatively high concentrations, it nonetheless was belownatural (whole-tissue) volumetric concentration. The ex-tracts examined here contained a large amount of salts,probably from seawater in the ascidians’ tissues, whichincreased the extract weight. The salts are not responsiblefor the observed activities in the antibacterial assay since thesympatric H. momus did not produce positive results at thesame concentrations as P. nigra. Although we tested thedefense against bacterial fouling in a growth inhibitionassay, bacterial fouling can also be prevented by compoundsthat are non- or weakly growth-inhibiting, as shown byprevention of bacterial attachment to the surface in otherascidians (Wahl et al., 1994), as well as for several spongeextracts and compounds (Kelly et al., 2003). A similarmechanism might also exist in the ascidian P. nigra givenits bacteria-free tunic surface (Fig. 1), and could worktogether with physical defenses such as regular shedding oftunic tissue (Hecht, 1918; Goodbody, 1962).
The effect of P. nigra extract on Bugula neritina settle-ment may be a result of its direct effect on the larvae or aresult of settlement cues from bacteria affected by theextract. Although the larvae were rinsed in filtered seawaterand the assays were performed in filter-sterilized seawater,bacteria introduced with the larvae might have influencedsettlement. However, B. neritina larvae are also known tosettle rapidly and effectively on polystyrene plates withouta biofilm (Maki et al., 1989; Dahms et al., 2004). Therefore,while we cannot rule out the influence of the antibacterialactivity of the P. nigra extracts, we view these results as adirect inhibition of B. neritina settlement. The inhibitoryeffect of H. momus extracts on B. neritina larval settlementwas somewhat surprising since this ascidian is normallycovered by fouling organisms. In surveys conducted atsoutheastern Australian rock reefs, this ascidian was alwaysfouled, although bryozoans were not among the reportedfouling organisms (Davis and White, 1994). The observedactivity was not due to a general toxicity of H. momusextract, as nauplii of the brine shrimp Artemia salina werenot killed at 2 mg m�1, the highest tested concentration inthe B. neritina assay. A possible explanation is that H.momus extracts might rather be selectively active againstvarious epibionts.
The P. nigra extract activities in the laboratory assayswith larvae of the bryozoan B. neritina and the brine shrimpA. salina were found at relatively low concentrations, giventhe high salt portion in the extracts, indicating the potentialfor highly effective compound(s). The active compound (orcompounds) could be an interesting candidate for potentialcommercial exploitation and deserve further investigation.The results of the field assays indicate that these compoundsare likely lipophilic. Hydrophilic substances would haverapidly leaked out of the gel, together with acids and salts,as indicated by the rapid initial drop in extract weight in gelsre-extracted after 24 h of deployment (Fig. 3). The presenceof antifouling activity in the gel even after 4 weeks ofdeployment in the field points therefore to lipophilic activecompounds. This suggestion is in line with theoretical ar-guments that antifouling compounds should be lipophilic inorder to remain on an organism’s surface and avoid rapidloss to the environment (Steinberg and de Nys, 2002).
In conclusion, the results reported here from field exper-iments, laboratory assays, and pH monitoring and controlexperiments show that secondary metabolites have a keyrole in keeping P. nigra surface free of epibionts. As inother ascidians (Wahl and Banaigs, 1991), this chemicalantifouling defense may act alongside physiological andphysical defenses suggested long ago (Hecht, 1918; Good-body, 1962), but whose efficiency in P. nigra still need to bedetermined. The different activities observed in the extractsmight also work additively—for example, the inhibition ofepibiotic bacteria can lead to the absence of settlement cuesnecessary for macrofouling organisms. These fouling or-ganisms would be affected directly by inhibitory secondarymetabolites and indirectly by the absence of settlement cues.To evaluate this suggestion, further studies are needed andthese would benefit from the isolation of the active second-ary metabolites, which could be of biotechnological interest.
Acknowledgments
We thank Olga Bunis for valuable help with lab experi-ments.
Literature Cited
Amutha, K. B., V. K. Meenakshi, and S. Senthamarai. 2010. Evalu-ation of the antibacterial activity and antimitotic activities of biofoulingmarine ascidian extracts of Tuticorin coast. Int. J. Pharma. Sci. 2:750–758.
Bryan, P. J., J. B. McClintock, M. Slattery, and D. P. Rittschof. 2003.A comparative study of the non-acidic chemically mediated antifoulantproperties of three sympatric species of ascidians associated withseagrass habitats. Biofouling 19: 235–245.
Dahms, H.-U., S. Dobretsov, and P.-Y. Qian. 2004. The effect ofbacterial and diatom biofilms on the settlement of the bryozoan Bugulaneritina. J. Exp. Mar. Biol. Ecol. 313: 191–209.
Dahms, H. U., Q. F. Gao, and J. S. Hwang. 2007. Optimized mainte-nance and larval production of the bryozoan Bugula neritina (Buguli-dae: Gymnolaemata) in the laboratory. Aquaculture 265: 169–175.
240 ANTIFOULING DEFENSE IN P. NIGRA

Davis, A. R,, and J. B. Bremner. 1999. Potential antifouling naturalproducts from ascidians: A review. Pp. 259–308 in Recent Advances inMarine Biotechnology, Vol 3, M. Fingerman, R. Nagabhushanam, andM.-F. Thompson, eds. CRC Press, Boca Raton, FL.
Davis, A. R., and G. A. White. 1994. Epibiosis in a guild of sessilesubtidal invertebrates in south-eastern Australia: a quantitative survey.J. Exp. Mar. Biol. Ecol. 177: 1–14.
Davis, A. R., and A. E. Wright. 1989. Interspecific differences in thefouling of two congeneric ascidians (Eudistoma olivaceum & E. cap-sulatum): Is surface acidity an effective defence? Mar. Biol. 102:491–497.
Davis, A. R., and A. E. Wright. 1990. Inhibition of larval settlement bynatural products from the ascidian, Eudistoma olivaceum (Van Name).J. Chem. Ecol. 16: 1349–1357.
Fusetani, N. 2012. Marine natural products. Pp. 31–64 in Natural Prod-ucts in Chemical Biology, N. Civjan, ed. John Wiley, Hoboken, NJ.
Goodbody, I. 1962. The biology of Ascidia nigra (Savigny). I. Survivaland mortality in an adult population. Biol. Bull. 122: 40–51.
Goodbody, I. 1965. The biology of Ascidia nigra (Savigny). III. Theannual pattern of colonization. Biol. Bull. 129: 128–133.
Haber, M., M. Carbone, E. Mollo, M. Gavagnin, and M. Ilan. 2011.Chemical defense against predators and bacterial fouling in the Med-iterranean sponges Axinella polypoides and A. verrucosa. Mar. Ecol.Prog. Ser. 422: 113–122.
Hecht, S. 1918. The physiology of Ascidia atra Lesueur. I. Generalphysiology. J. Exp. Zool. 25: 229–259.
Henrikson, A. A., and J. R. Pawlik. 1995. A new antifouling assaymethod: results from field experiments using extracts of four marineorganisms. J. Exp. Mar. Biol. Ecol. 194: 157–165.
Hirose, E., H. Yamashiro, and Y. Mori. 2001. Properties of tunic acidin the ascidian Phallusia nigra (Ascidiidae, Phlebobranchia). Zoolog.Sci. 18: 309–314.
Holmstrom, C., S. James, B. A. Neilan, D. C. White, and S. Kjelleberg.1998. Pseudoalteromonas tunicata sp. nov., a bacterium that pro-duces antifouling agents. Int. J. Syst. Bacteriol. 48: 1205–1212.
Holmstrom, C., S. Egan, A. Franks, S. McCloy, and S. Kjelleberg.2002. Antifouling activities expressed by marine surface associatedPseudoalteromonas species. FEMS Microbiol. Ecol. 41: 47–58.
Hoyle, G. 1953. Spontaneous squirting of an ascidian, Phallusia mam-millata Cuvier. J. Mar. Biol. Assoc. UK 31: 541–562.
Jaffarali, H. A., M. Tamilselvi, and V. Sivakumar. 2008. Antibacterialactivity of the marine ascidians Phallusia nigra and Herdmania pallidafrom the Tuticorin coast, India. J. Biol. Res. Thessalon. 10: 171–179.
Kelly, S. R., P. R. Jensen, T. P. Henkel, W. Fenical, and J. R. Pawlik.2003. Effects of Caribbean sponge extracts on bacterial attachment.Aquat. Microb. Ecol. 31: 175–182.
Kelman, D., Y. Kashman, E. Rosenberg, M. Ilan, I. Ifrach, and Y.Loya. 2001. Antimicrobial activity of the reef sponge Amphimedonviridis from the Red Sea: evidence for selective toxicity. Aquat. Mi-crob. Ecol. 24: 9–16.
Koplovitz, G., J. B. McClintock, C. D. Amsler, and B. J. Baker. 2011.A comprehensive evaluation of the potential chemical defenses ofAntarctic ascidians against sympatric fouling microorganisms. Mar.Biol. 158: 2661–2671.
Krug, P. J. 2006. Defense of benthic invertebrates against surface col-onization by larvae: a chemical arms race. Prog. Mol. Subcell. Biol. 42:1–53.
Lambert, G. 1968. The general ecology and growth of a solitary ascid-ian, Corella willmeriana. Biol. Bull. 135: 296–307.
Laudien, J., and M. Wahl. 1999. Indirect effects of epibiosis on hostmortality: Seastar predation on differently fouled mussels. Mar. Ecol.20: 35–47.
Lopez-Legentil, S., X. Turon, and P. Schupp. 2006. Chemical andphysical defenses against predators in Cystodytes (Ascidiaceae). J. Exp.Mar. Biol. Ecol. 332: 27–36.
Maki, J. S., D. Rittschof, A. R. Schmidt, A. G. Snyder, and R. Mitchell.1989. Factors controlling attachment of bryozoan larvae: a compari-son of bacterial films and unfilmed surfaces. Biol. Bull. 177: 295–302.
McClintock, J. B., M. O. Amsler, C. D. Amsler, K. J. Southworth, C.Petrie, and J. B. Baker. 2004. Biochemical composition, energycontent and chemical antifeedant and antifoulant defenses of the colo-nial Antarctic ascidian Distaplia cylindrica. Mar. Biol. 145: 885–894.
Odate, S., and J. R. Pawlik. 2007. The role of vanadium in the chemicaldefense of the solitary tunicate, Phallusia nigra. J. Chem. Ecol. 33:643–654.
Olguin-Uribe, G., E. Abou-Mansour, A. Boulander, H. Debard, C.Francisco, and G. Combaut. 1997. 6-Bromoindole-3-carbaldehyde,from an Acinetobacter sp. bacterium associated with the ascidianStomozoa murrayi. J. Chem. Ecol. 23: 2507–2521.
Parry, D. L. 1984. Chemical properties of the test of ascidians in relationto predation. Mar. Ecol. Prog. Ser. 17: 279–282.
Pisut, D. P., and J. R. Pawlik. 2002. Anti-predatory chemical defensesof ascidians: secondary metabolites or inorganic acids? J. Exp. Mar.Biol. Ecol. 270: 203–214.
Shenkar, N., and Y. Loya. 2009. Non-indigenous ascidians (Chordata:Tunicata) along the Mediterranean coast of Israel. Mar. Biodivers. Rec.2: e166, doi:10.1017/S1755267209990753.
Simmons, T. J., R. C. Coates, B. R. Clark, N. Engene, D. Gonzalez, E.Esquenazi, P. C. Dorrestein, and W. H. Gerwick. 2008. Biosyn-thetic origin of natural products isolated from marine microorganism-invertebrate assemblages. Proc. Natl. Acad. Sci. USA 105: 4587–4594.
Slattery, M., J. B. McClintock, and J. N. Heine. 1995. Chemicaldefenses in Antarctic soft corals: evidence for antifouling compounds.J. Exp. Mar. Biol. Ecol. 190: 61–77.
Sokal, R. R., and F. J. Rohlf. 1995. Analysis of frequencies. Pp.685–733 in Biometry: the Principles and Practice of Statistics inBiological Research, 3rd ed., R. R. Sokal, and F. J. Rohlf, eds. W. H.Freeman, San Fransisco.
Steinberg, P. D., and R. de Nys. 2002. Chemical mediation of coloni-zation of seaweed surfaces. J. Phycol. 38: 621–629.
Stoecker, D. 1978. Resistance of a tunicate to fouling. Biol. Bull. 155:615–626.
Stoecker, D. 1980. Relationships between chemical defense and ecologyin benthic ascidians. Mar. Ecol. Prog. Ser. 3: 257–265.
Tsukamoto, S., H. Kato, H. Hirota, and N. Fusetani. 1994. Antibac-terial and antifungal sulfated alkane and alkenes from the hepato-pancreas of the ascidian Halocynthia roretzi. J. Nat. Prod. 57: 1606–1609.
Uriz, M., D. Martin, X. Turon, E. Ballesteros, R. Hughes, and C.Acebal. 1991. An approach to the ecological significance of thechemically mediated bioactivity in Mediterranean benthic communi-ties. Mar. Ecol. Prog. Ser. 70: 175–188.
Wahl, M. 1989. Marine epibiosis. I. Fouling and antifouling: some basicaspects. Mar. Ecol. Prog. Ser. 58: 175–189.
Wahl, M. 2008. Ecological lever and interface ecology: epibiosis mod-ulates the interaction between host and environment. Biofouling 24:427–438.
Wahl, M., and B. Banaigs. 1991. Marine epibiosis. 3. Possible antifoul-ing defense adaptations in Polysyncraton lacazei (Giard) (Didemnidae,Ascidiacea). J. Exp. Mar. Biol. Ecol. 145: 49–63.
Wahl, M., P. R. Jensen, and W. Fenical. 1994. Chemical control ofbacterial epibiosis on ascidians. Mar. Ecol. Prog. Ser. 110: 45–57.
Wahl, M., K. Kroger, and M. Lenz. 1998. Non-toxic protection againstepibiosis. Biofouling 12(1-3): 205–226.
241B. MAYZEL ET AL.
Related Documents











