Journal of Physiology (1994), 481.3 Characterization of the hyperpolarization-activated current, If, in ventricular myocytes isolated from hypertensive rats Elisabetta Cerbai, Mario Barbieri and Alessandro Mugelli * Institute of Pharmacology, University of Ferrara, and *Department of Pharmacology, University of Firenze, Italy 1. Left ventricular myocytes isolated from the heart of young (2-month-old) and old (18- to 20-month-old) spontaneously hypertensive rats (SHRs) were studied in the whole-cell configuration. Since multicellular preparations from old SHRs show a diastolic depolarization phase, we performed experiments to test whether it was associated with the presence of a hyperpolarization-activated If-like current. 2. In control Tyrode solution, a time-dependent increasing inward current activated by hyperpolarization was recorded in myocytes from old SHRs showing a diastolic depolarization phase. A barium-insensitive, caesium-sensitive, time-dependent inward current was recorded in a minority (4 of 33) of cells from young SHRs (membrane capacitance, 160 + 7 pF) but in 93% (25 of 27, P < 0 01) of myocytes from old SHRs (membrane capacitance, 355 + 19 pF, P < 0 01). 3. The current was fully activated at -120 mV and voltage of half-maximal activation was -88-1 + 1-5 mV; it was blocked by extracellular CsCl (4 mM) in a voltage-dependent manner. Reducing [K+]O from 25 to 5-4 mm caused a shift of the reversal potential from -17-3 + 3-8 to -25-7 + 2-7 mV and a 60% decrease of current conductance. 4. These findings suggest that an If-like current is present in rat ventricular myocytes from old SHRs, where it might favour the occurrence of spontaneous action potentials. Pacemaker activity is a property of specialized myocytes that are able to generate spontaneous action potentials (DiFrancesco, 1993). If is a non-selective cation current activated on hyperpolarization which is thought to contribute to pacemaker activity in both primary (sinus node) and secondary (Purkinje fibres) pacemakers (DiFrancesco, 1981a, b, 1991). If provides an inward current which might play a role in determining the diastolic depolarization phase (DiFrancesco, 1985; Irisawa, Brown & Giles, 1993). It has been reported recently (Yu, Chang & Cohen, 1993) that If is present in ventricular myocytes from guinea-pig and canine hearts. However, the current activates at such negative potentials (more negative than -120 mV) that it could not have any physiological role, i.e. it does not cause diastolic depolarization. We have reported recently that left papillary muscles isolated from the heart of 18-month-old spontaneously hypertensive rats (old SHRs) consistently show a prolongation of action potential duration and a marked diastolic depolarization phase (Barbieri et al. 1994). This diastolic depolarization was not present in 2-month-old SHRs (young SHRs) (Barbieri et al. 1994). Patch-clamped left ventricular myocytes from these hearts maintained the action potential prolongation, which appeared to be due to a selective reduction in transient outward current density (Cerbai, Barbieri, Li & Mugelli, 1994a). We hypothesized that the diastolic depolarization phase observed in papillary muscles of old SHRs could be associated with the presence of an If-like current. Here we report that a current activated on hyperpolarization was present in the majority of myocytes from old SHRs and in a minority of those from young SHRs. We provide evidence that this current has the electrophysiological characteristics of If. Part of this work has been published in abstract form (Cerbai, Barbieri, Porciatti & Mugelli, 1994b). METHODS The investigation conforms to the rules for the care and use of laboratory animals of the European Community (86/609/ CEE). Male SHRs were obtained from Charles River (Como, Italy). They were maintained in our animal facility until killed. Systolic blood pressure, measured using a tail-cuff system (BP Recorder, Basile, Varese, Italy), was 183 + 3 mmHg in 2-month-old SHRs (young SHRs) (n= 7) and 208 + 14 mmHg This manuscript was accepted as a Short Paper for rapid publication. MS 3804, pp. 585-591 585 ) by guest on July 14, 2011 jp.physoc.org Downloaded from J Physiol (

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Physiology (1994), 481.3
Characterization of the hyperpolarization-activated current,If, in ventricular myocytes isolated from hypertensive rats
Elisabetta Cerbai, Mario Barbieri and Alessandro Mugelli *
Institute of Pharmacology, University of Ferrara, and *Department of Pharmacology,University of Firenze, Italy
1. Left ventricular myocytes isolated from the heart of young (2-month-old) and old (18- to20-month-old) spontaneously hypertensive rats (SHRs) were studied in the whole-cellconfiguration. Since multicellular preparations from old SHRs show a diastolicdepolarization phase, we performed experiments to test whether it was associated withthe presence of a hyperpolarization-activated If-like current.
2. In control Tyrode solution, a time-dependent increasing inward current activated byhyperpolarization was recorded in myocytes from old SHRs showing a diastolicdepolarization phase. A barium-insensitive, caesium-sensitive, time-dependent inwardcurrent was recorded in a minority (4 of 33) of cells from young SHRs (membranecapacitance, 160 + 7 pF) but in 93% (25 of 27, P < 0 01) of myocytes from old SHRs(membrane capacitance, 355 + 19 pF, P < 0 01).
3. The current was fully activated at -120 mV and voltage of half-maximal activation was-88-1 + 1-5 mV; it was blocked by extracellular CsCl (4 mM) in a voltage-dependentmanner. Reducing [K+]O from 25 to 5-4 mm caused a shift of the reversal potential from-17-3 + 3-8 to -25-7 + 2-7 mV and a 60% decrease of current conductance.
4. These findings suggest that an If-like current is present in rat ventricular myocytesfrom old SHRs, where it might favour the occurrence of spontaneous action potentials.
Pacemaker activity is a property of specialized myocytesthat are able to generate spontaneous action potentials(DiFrancesco, 1993). If is a non-selective cation currentactivated on hyperpolarization which is thought to contributeto pacemaker activity in both primary (sinus node) andsecondary (Purkinje fibres) pacemakers (DiFrancesco,1981a, b, 1991). If provides an inward current which mightplay a role in determining the diastolic depolarizationphase (DiFrancesco, 1985; Irisawa, Brown & Giles, 1993).
It has been reported recently (Yu, Chang & Cohen, 1993)that If is present in ventricular myocytes from guinea-pigand canine hearts. However, the current activates at suchnegative potentials (more negative than -120 mV) that itcould not have any physiological role, i.e. it does not causediastolic depolarization.We have reported recently that left papillary muscles
isolated from the heart of 18-month-old spontaneouslyhypertensive rats (old SHRs) consistently show aprolongation of action potential duration and a markeddiastolic depolarization phase (Barbieri et al. 1994). Thisdiastolic depolarization was not present in 2-month-oldSHRs (young SHRs) (Barbieri et al. 1994). Patch-clampedleft ventricular myocytes from these hearts maintained
the action potential prolongation, which appeared to bedue to a selective reduction in transient outward currentdensity (Cerbai, Barbieri, Li & Mugelli, 1994a). Wehypothesized that the diastolic depolarization phaseobserved in papillary muscles of old SHRs could beassociated with the presence of an If-like current. Here wereport that a current activated on hyperpolarization waspresent in the majority of myocytes from old SHRs and ina minority of those from young SHRs. We provideevidence that this current has the electrophysiologicalcharacteristics of If. Part of this work has been publishedin abstract form (Cerbai, Barbieri, Porciatti & Mugelli,1994b).
METHODSThe investigation conforms to the rules for the care and use oflaboratory animals of the European Community (86/609/CEE).
Male SHRs were obtained from Charles River (Como,Italy). They were maintained in our animal facility untilkilled. Systolic blood pressure, measured using a tail-cuff system(BP Recorder, Basile, Varese, Italy), was 183 + 3 mmHg in2-month-old SHRs (young SHRs) (n= 7) and 208 + 14 mmHg
This manuscript was accepted as a Short Paper for rapid publication.
MS3804, pp.585-591 585
) by guest on July 14, 2011jp.physoc.orgDownloaded from J Physiol (

E. Cerbai, M. Barbieri and A. Mugelli
in 18- to 20-month-old SHRs (old SHRs) (n = 6, notsignificant). The old SHRs were observed daily for signs ofheart failure. All animals used were free of these signs.
Before killing, the animals were injected with 500 i.u.heparin i.P. (Liquemin; Roche, Basle, Switzerland) andanaesthetized with ether. The heart was rapidly excised,rinsed in cool nominally calcium-free solution (LCS) andweighed.
Cell isolationSingle left ventricular myocytes were isolated from young orold SHRs using a protocol based on previously describedprocedures (Barbieri et al. 1994; Cerbai et al. 1994a). Afterdigestion of the heart, the left ventricle, including the septum,was cut off, chopped into small pieces and gently stirred inLCS containing (mM): NaCl, 120; KCl, 10; KH2PO4, 1P2; MgCl2,1P2; glucose, 20; taurine, 10; and pyruvate, 5; the pH wasadjusted to 7T20 with Hepes-NaOH. Cardiomyocytes thatappeared in the supernatant were purified by gravitysedimentation, collected and stored in LCS supplemented withalbumin (1 mg ml-1; Boehringer Mannheim), penicillin(50 i.u. ml-'; Gibco Live Technologies, UK), streptomycin(50 jug ml-'; Gibco Laboratories) and 1 mm CaCl2. Cells werestored at room temperature (20-22 °C) and used within 10 h oftheir isolation.
SolutionsA drop of cell suspension was placed in the experimentalchamber (0'2 ml) and superfused by means of a peristalticpump (Masterflex, model 7524/05, Cole-Parmer InstrumentCompany) at a flow rate of 1P8 ml min-'; a three-line systemcontrolled by electronic valves allowed solutions to be changedrapidly. The recording chamber was mounted on the stage ofan inverted microscope (TMS, Nikon). The control solutionwas a Tyrode solution containing (mM): NaCl, 137; KCl, 5-4;CaCl2, 1P5; MgCl2,12; Hepes, 5; glucose, 10; pH was adjustedto 7-35 with NaOH; the temperature was kept at 36-0 + 0 5 'C.The internal solution in the patch pipettes contained (mM):potassium aspartate, 130; MgCl2, 2; Na2ATP, 5; CaCl2, 5;ethyleneglycol-bis-(f8-aminoethylether)N,N,N',N'-tetraaceticacid (EGTA), 11; Hepes, 10; adjusted to pH 7-20 with KOH. Nocorrection was made for liquid junction potential betweeninternal pipette solution and bath solutions. To study thehyperpolarization-activated current, the extracellular controlsolution was modified to reduce the interference ofcomponents other than If (Yu et al. 1993), by adding (mM):BaCl2, 8; MnCl2, 2; CdCl2, 0-2; 4-aminopyridine, 0 5; and theconcentration of KCl was increased to 25 mm (unlessindicated). When necessary, CsCl from a 1 M stock solution wasadded to the Tyrode solution to reach the final concentrationof 4 mm. A stock solution of (-)-isoprenaline (1 mM) containing1 g F1 ascorbic acid was prepared on the day of the experimentand diluted in Tyrode solution to reach a final concentration of0-1 /M. (-)-Isoprenaline HCl and 4-aminopyridine wereobtained from Sigma Chemical Company (Italy). All reagentswere of analytical grade.
Experimental protocolsThe experimental set-up was similar to that describedpreviously (Cerbai, Ambrosio, Porciatti, Chiariello, Giotti &Mugelli, 1991). The whole-cell configuration of the patch-clamptechnique was used to record action potential and membranecurrents. The electrical signal was recorded by a patchamplifier (Axopatch 1D, Axon Instrument Inc.), digitized(Labmaster TL-1 DMA, Scientific Solutions), and displayed on
the monitor of a 386 personal computer and a digitaloscilloscope (Nicolet 310, Nicolet Instrumentation Company).The cut-off frequency was 20 kHz. Current and voltageprotocol generation, data acquisition, and analysis wereperformed using the pCLAMP software (version 5.5.1, AxonInstruments, Inc.). MicroCal Origin (MicroCal Software Inc.,Northampton, MA, USA) was used for further analysis.
After gaining intracellular access, whole-cell membranecapacitance was measured by applying ± 10 mV voltage stepsfrom a holding potential of -70 mV and calculated accordingto the equation:
Cm = (Io- I)/A Vm,where Cm is the membrane capacitance, r, is the time constantof the membrane capacitance, Io is the initial current value, IOOis the amplitude of steady-state current and A Vm is theamplitude of voltage steps. Series resistance (R8), calculated asR8= A Vm/Io (see above), ranged from 2 to 7 MQ and wascompensated in all cells by about 80%. No capacitancecorrection was used.
Recording was started after 5 min dialysis of the cell.Action potentials were elicited at a rate of 0-2 Hz and sampledat 2 kHz. In voltage-clamp experiments, If was elicited byhyperpolarizing steps (varying from -60 to -170 mV) from aholding potential of -40 or -50 mV. Steps were applied at lowfrequency (maximum rate, 0-1 Hz) and sampled at 0 5-1 kHz.
Data analysisThe occurrence of If, expressed as the ratio between thenumber of cells showing caesium-sensitive, barium-insensitive, time-dependent increasing inward current elicitedby hyperpolarizing steps, and the total number of cellsstudied, was compared in different groups of animals bymeans of the x2 test (2 x 2 contingency table). All the otherdata are presented as the means + standard error of the mean(S.E.M.). Comparison between two groups was performed bymeans of Student's t test for paired data. A P value of lessthan 0 05 was considered significant.
RESULTS
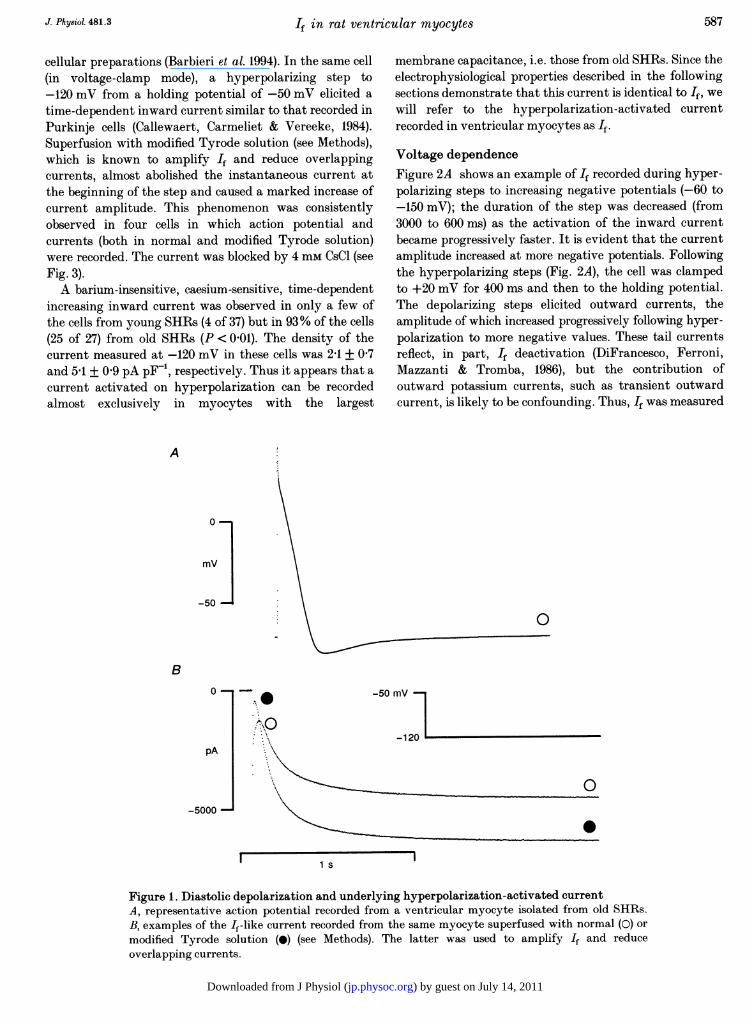
Membrane capacitanceMembrane capacitance was systematically measured in allcells. Young SHRs had a cell capacitance of 160 + 7 pF(n = 37); as expected (Cerbai et al. 1994a), membranecapacitance was dramatically increased in old SHRs(355 + 19 pF, n = 27, P < 0 01 versus young SHRs).Diastolic depolarization and If-like current inleft ventricular myocytes from old SHRsFigure 1A shows a representative action potential recordedfrom left ventricular myocytes of old SHRs. The actionpotential characteristics of these cells have been previouslyreported in detail (Cerbai et al. 1994a). Maximum diastolicpotential of young and old SHRs was -72-7 + 1P0 (n = 14)and -71P9 + 1-4 mV (n = 13, P > 0 05), respectively. Thepresence of a marked diastolic depolarization phase, whichwas absent in the young rats (not shown), is apparent.This phenomenon was previously observed in multi-
586 J. Physiol. 481.3
) by guest on July 14, 2011jp.physoc.orgDownloaded from J Physiol (
If in rat ventricular myocytes
cellular preparations (Barbieri et al. 1994). In the same cell(in voltage-clamp mode), a hyperpolarizing step to-120 mV from a holding potential of -50 mV elicited atime-dependent inward current similar to that recorded inPurkinje cells (Callewaert, Carmeliet & Vereeke, 1984).Superfusion with modified Tyrode solution (see Methods),which is known to amplify If and reduce overlappingcurrents, almost abolished the instantaneous current atthe beginning of the step and caused a marked increase ofcurrent amplitude. This phenomenon was consistentlyobserved in four cells in which action potential andcurrents (both in normal and modified Tyrode solution)were recorded. The current was blocked by 4 mm CsCl (seeFig. 3).A barium-insensitive, caesium-sensitive, time-dependent
increasing inward current was observed in only a few ofthe cells from young SHRs (4 of 37) but in 93% of the cells(25 of 27) from old SHRs (P < 0 01). The density of thecurrent measured at -120 mV in these cells was 2-1 + 0 7and 5-1 + 0 9 pA pF-t, respectively. Thus it appears that acurrent activated on hyperpolarization can be recordedalmost exclusively in myocytes with the largest
membrane capacitance, i.e. those from old SHRs. Since theelectrophysiological properties described in the followingsections demonstrate that this current is identical to If, wewill refer to the hyperpolarization-activated currentrecorded in ventricular myocytes as If.
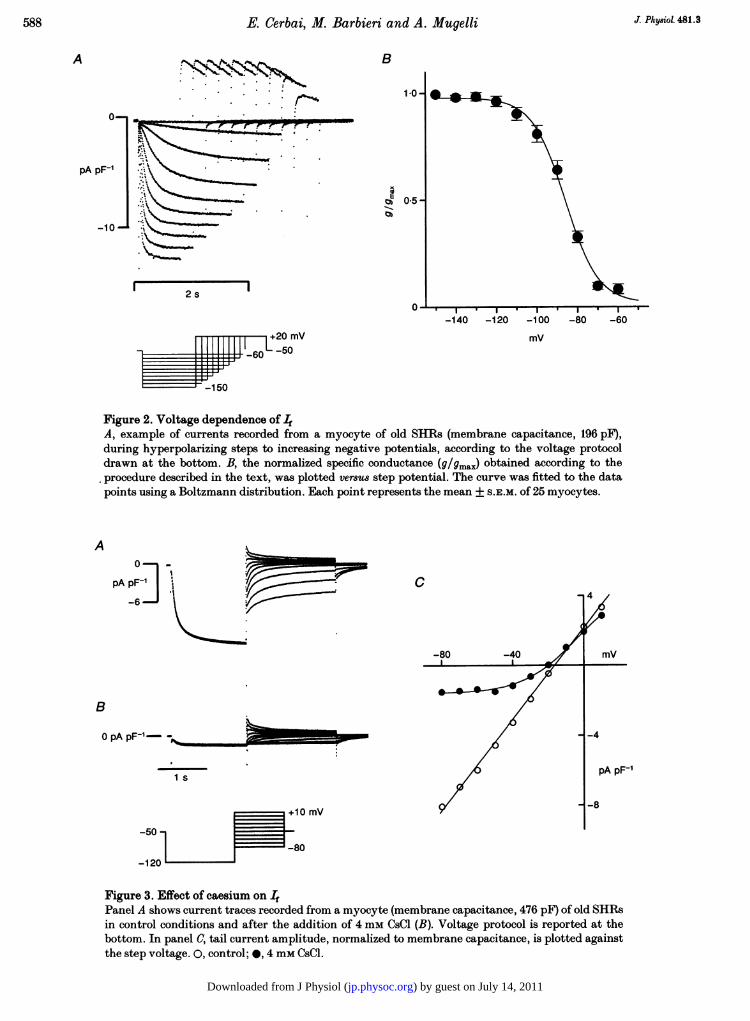
Voltage dependenceFigure 2A shows an example of If recorded during hyper-polarizing steps to increasing negative potentials (-60 to-150 mV); the duration of the step was decreased (from3000 to 600 ms) as the activation of the inward currentbecame progressively faster. It is evident that the currentamplitude increased at more negative potentials. Followingthe hyperpolarizing steps (Fig. 2A), the cell was clampedto +20 mV for 400 ms and then to the holding potential.The depolarizing steps elicited outward currents, theamplitude of which increased progressively following hyper-polarization to more negative values. These tail currentsreflect, in part, If deactivation (DiFrancesco, Ferroni,Mazzanti & Tromba, 1986), but the contribution ofoutward potassium currents, such as transient outwardcurrent, is likely to be confounding. Thus, If was measured
A
mV]
0
B0-
pA
-5000-
-50 mV7
.0j
0
.
1 s
Figure 1. Diastolic depolarization and underlying hyperpolarization-activated currentA, representative action potential recorded from a ventricular myocyte isolated from old SHRs.B, examples of the If-like current recorded from the same myocyte superfused with normal (0) ormodified Tyrode solution (0) (see Methods). The latter was used to amplify If and reduceoverlapping currents.
587J. Physiol. 481.3
l
) by guest on July 14, 2011jp.physoc.orgDownloaded from J Physiol (
E. Cerbai, M. Barbieri and A. Mugelli
B
1*0-
0m 05-0)
I.
I I
1-140 -120 -100
mV
Figure 2. Voltage dependence of IfA, example of currents recorded from a myocyte of old SHIRs (membrane capacitance, 196 pF),during hyperpolarizing steps to increasing negative potentials, according to the voltage protocoldrawn at the bottom. B, the normalized specific conductance (9/9max) obtained according to theprocedure described in the text, was plotted versus step potential. The curve was fitted to the datapoints using a Boltzmann distribution. Each point represents the mean + S.E.M. of 25 myocytes.
A t
pA PF I
B
pA pF-' -,__
1 s
+10 mV
-50
80
-120
C
-80
Figure 3. Effect of caesium on 4
Panel A shows current traces recorded from a myocyte (membrane capacitance, 476 pF) of old SHRsin control conditions and after the addition of 4 mm CsCl (B). Voltage protocol is reported at thebottom. In panel C, tail current amplitude, normalized to membrane capacitance, is plotted againstthe step voltage. 0, control; 0, 4 mm CsCl.
A
0-
pA pF-1
-10-
2 sI .
-80 -60
-
J. Physiol. 481.35-88
N
I~ I
20 mvT 1- [:-5060
F--- -150
) by guest on July 14, 2011jp.physoc.orgDownloaded from J Physiol (
If in rat ventricular myocytes
as the difference between the instantaneous current at thebeginning of the hyperpolarizing step and the steady-statecurrent recorded at the end of the step; If was normalizedwith respect to membrane capacitance. Specific conductancewas determined as a function of membrane potentialaccording to the following equation:
g = I/( Vm -Vrev),where g is the conductance calculated at membrane potentialVm, I the current amplitude and Vrev the reversalpotential of the fully activated current derived from theanalysis of tail currents of Fig. 4. The maximum specificconductance (gmax) was obtained by using a computer-calculated Boltzmann fit according to the equation:
g = gmrax/{1 + exp[( V½-Vm)/k]},where g is the conductance calculated at membrane potentialVm, Vt½ is the potential of half-maximal activation and k isthe slope factor describing the steepness of the activationcurve. Using the value Of gmax obtained from thisequation, we plotted the normalized specific conductance(g/gmax) versus the membrane potential. The results ofsuch an analysis, performed in twenty-five left ventricularcells from old SHRs, are summarized in Fig. 2B. Thecurrent was fully activated at -120 mV, Vt was-88X1 + 1X5 mV and k was -7-6 + 1-3 mV; the value for
gmax was calculated to be 54-4 + 1P7 pS pF-'. The voltagedependence of If has been studied in the cells from youngSHRs in which If occurred; a complete activation curvewas obtained in four cells. The mean V1 was -87 + 1 mV,a value which is not different from that obtained in cellsfrom old SHRs. A similar voltage protocol was applied tothirteen SHRs cells not showing evidence of If duringhyperpolarization to -120 mV. In no case could If bedemonstrated even at the most negative potentials(-160 mV). The addition of (-)-isoprenaline (1 mM) (n = 3)failed to induce the occurrence of If (data not shown).
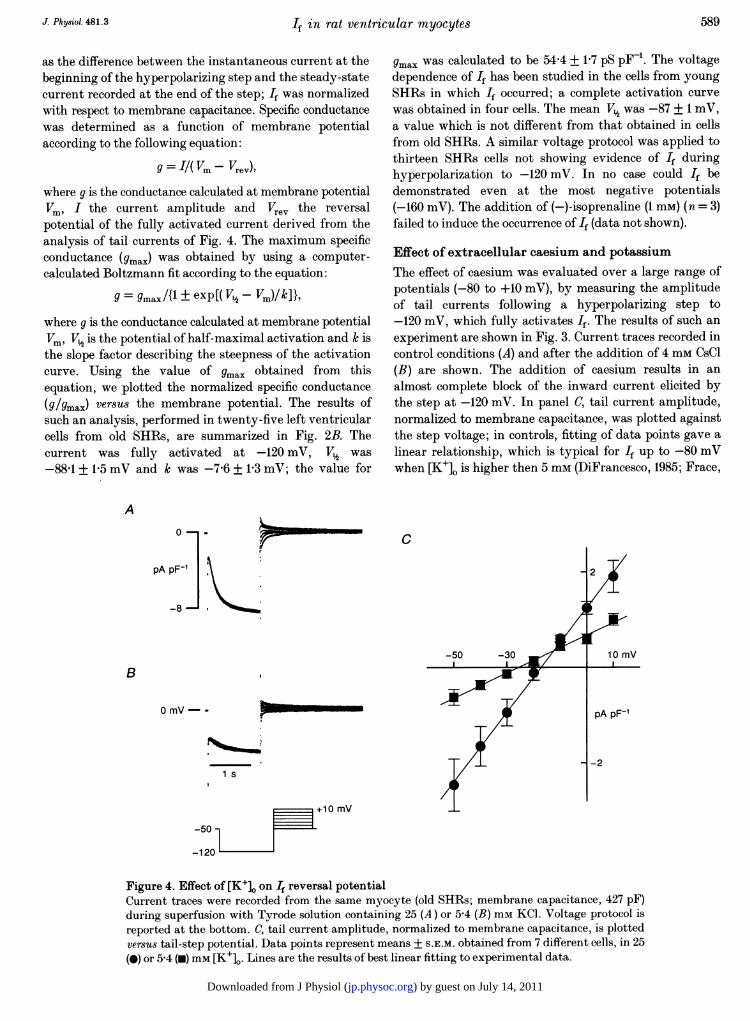
Effect of extracellular caesium and potassiumThe effect of caesium was evaluated over a large range ofpotentials (-80 to +10 mV), by measuring the amplitudeof tail currents following a hyperpolarizing step to-120 mV, which fully activates If. The results of such anexperiment are shown in Fig. 3. Current traces recorded incontrol conditions (A) and after the addition of 4 mm CsCl(B) are shown. The addition of caesium results in analmost complete block of the inward current elicited bythe step at -120 mV. In panel C, tail current amplitude,normalized to membrane capacitance, was plotted againstthe step voltage; in controls, fitting of data points gave alinear relationship, which is typical for If up to -80 mVwhen [K+]. is higher then 5 mm (DiFrancesco, 1985; Frace,
A
C
pA pF-1
-50 -30
B
0 mV- -
1 s
L+10 mV-50 -
-120
Figure 4. Effect of [K+]o on If reversal potentialCurrent traces were recorded from the same myocyte (old SHRs; membrane capacitance, 427 pF)during superfusion with Tyrode solution containing 25 (A) or 5-4 (B) mm KCI. Voltage protocol isreported at the bottom. C, tail current amplitude, normalized to membrane capacitance, is plottedversus tail-step potential. Data points represent means + S.E.M. obtained from 7 different cells, in 25(0) or 5-4 (-) mm [K+]0. Lines are the results of best linear fitting to experimental data.
589J. Physiol. 481.3
) by guest on July 14, 2011jp.physoc.orgDownloaded from J Physiol (

590 E. Cerbai, M. Barbieri and A. Mugelli J. PhysioL 481.3
Maruoka & Noma, 1992). In the presence of caesium, thenegative region of the I-V relationship was characterizedby a progressive reduction of the current, showing a clear-cut voltage-dependent blockade. The effect of caesium onthe hyperpolarization-activated current in left ventricularcells ofSHRs was qualitatively similar to that reported forthe pacemaker current recorded from Purkinje fibres(DiFrancesco, 1982) and Purkinje cells (Callewaert et al.1984). The reversal potential in the presence of caesiumwas -17 0 + 0 3 mV (n = 7), a value not different fromthat measured in control conditions (see below).The measurement of tail current amplitude was used to
evaluate the reversal potential of If at different extracellularpotassium concentrations ([K+]0.) Tail currents, followinga hyperpolarizing step to -120 mV which maximallyactivates If, were elicited by steps in the range of -50 to+10 mV. Current traces in Fig. 4 were obtained in thesame cell superfused with Tyrode solution containingeither 25 (A) or 5 4 mm (B) [K+]. The most evident effectcaused by the reduction in [K+]0 was a marked decrease inIf amplitude. Tail current amplitudes, normalized tomembrane capacitance, obtained from seven different cellssuperfused with 25 mm [K+]0 are plotted as a function oftail step potential in the I-V relationship of Fig. 4C. Bestfit through data points gave a linear relationship, with aslope of 73 0 + 1P3 pS pF-1, which intersected the X-axis(reversal potential of If) at -17-3 + 3-8 mV. With 5-4 mM[K+]0, the reversal potential was -25-7 + 2-7 mV and slopewas 26 4 + 1 0 pS pF_' (n = 7) (Fig. 4). Such a shift of thereversal potential was comparable to that reported for Ifin Purkinje fibres (DiFrancesco, 1982) and sino-atrial nodecells (DiFrancesco et at. 1986) exposed to a similarreduction in extracellular KV. The decrease in conductanceas a consequence of decreasing [K+]o is also typical for If inboth primary (DiFrancesco et al. 1986; Frace et al. 1992)and subsidiary pacemaker cells (DiFrancesco, 1982; Zhou &Lipsius, 1992).
DISCUSSIONThe present results demonstrate for the first time thatIf exists in myocytes of old SHRs where it might play afunctional role, i.e. contributing to the diastolicdepolarization phase. The current has the electro-physiological characteristics of If in pacemaker tissue(DiFrancesco, 1993) in that it is typically activated uponhyperpolarization, being fully activated at -120 mV; it iscaesium and barium insensitive, and has a reversalpotential consistent with selectivity for Na+ and K+(DiFrancesco, 1981b; DiFrancesco et at. 1986; Ho, Brown &Noble, 1994). The permeability ratio PN./PK, calculatedfrom the reversal potential values, was 0 34 in both 5*4and 25 mm [K+]O. The current was recorded also in normalTyrode solution from cells showing a diastolicdepolarization phase; its amplitude was increased bysuperfusing the cell with a Tyrode solution modified in
order to amplify If. Thus it is unlikely that occurrence ofthe hyperpolarization-activated current represents anartifact due to our experimental conditions.
Another relevant consideration from our study is thatIf is practically only expressed by severely hypertrophiedmyocytes, and studies are in progress to determine therelative importance of hypertrophy and aging on theoccurrence of If in ventricular myocytes. If is present, infact, in more than 90% of left ventricular myocytes of oldSHRs but only in about 10% myocytes from young SHRs.This finding confirms and extends our previous observationsin papillary muscles (Barbieri et al. 1994) where a diastolicdepolarization phase was present in 18-month-old SHRsbut not in young SHRs. Our preliminary results suggestthat an If-like current may be present in 18-month-oldnormotensive Wistar Kyoto rats (Cerbai et at. 1994b). Ifoccurred in only four of thirty-seven cells from youngSHRs; thus a reliable analysis of its properties could not beperformed. However, in these cells the voltage of half-maximal activation is similar to that observed in cellsfrom old SHRs. If If is not present at -120 mV, it cannotbe activated at more negative voltages, nor can it beinduced by isoprenaline. Thus, in the rat the situationseems to be quite different from that reported by Yu et al.(1993) in guinea-pig and canine myocytes. In theirexperiments (the majority of which were carried out in thepresence of cyclic AMP in the pipette or of externalisoprenaline), an If-like current is apparently present inall the cells provided that a sufficiently negative hyper-polarizing step is given. These authors hypothesized thatshifting If to very negative (and not physiological)potentials could be the mechanism for avoidingpacemaking in ventricular cells. In the rat, a less 'wastefulapproach' appears to operate: the If gene seems to beturned off and the If channel is not synthesized. In the fewcells of young SHRs where an If-like current is detected, itactivates at a similar voltage but its density is lower thanin old SHRs, and possibly not sufficient to play afunctional role.At variance with the results obtained in normal guinea-
pig and canine ventricular myocytes, the activation of Ifin hypertrophied rat myocytes occurs at voltages near thephysiological diastolic potential of these myocytes. Undercertain circumstances it may play a major role: it might bespeculated that it contributes to an increased propensityof the hypertrophied heart for arrhythmias, as observedexperimentally (Belichard, Pruneau & Rochette, 1988;Pahor et al. 1989; Barbieri et at. 1994) and clinically(McLenachan, Henderson, Morris & Dargie, 1987).
REFERENCESBARBIERI, M., VARANI, K., CERBAI, E., GUERRA, L., Li, Q.,
BOREA, P. A. & MUGELLI, A. (1994). Electrophysiological basisfor the enhanced cardiac arrhythmogenic effect of isoprenalinein aged spontaneously hypertensive rats. Journal of Molecularand Cellular Cardiology 26, 849-860.
) by guest on July 14, 2011jp.physoc.orgDownloaded from J Physiol (

J. Ph8ioL. 481.3 If in rat ventricular myocytes 591
BELICHARD, P., PRUNEAU, D. & ROCHETTE, L. (1988). Influence ofspontaneous hypertension and cardiac hypertrophy on theseverity of ischemic arrhythmias in the rat. Basic Research inCardiology 83, 560-566.
CALLEWAERT, G., CARMELIET, E. & VEREEKE, J. (1984). Singlecardiac Purkinje cells: general electrophysiology and voltage-clamp analysis of the pace-maker current. Journal ofPhysiology 349,643-661.
CERBAI, E., AMBROSIO, G., PORCIATTI, F., CHIARIELLO, M.,GIOTTI, A. & MUGELLI, A. (1991). Cellular electrophysiologicalbasis for oxygen radical-induced arrhythmias: a patch-clampstudy in guinea-pig ventricular myocytes. Circulation 84,1773-1782.
CERBAI, E., BARBIERI, M., Li, Q. & MUGELLI, A. (1994a). Ionicbasis of action potential prolongation of hypertrophied cardiacmyocytes isolated from hypertensive rats of different ages.Cardiovascular Research 28,1180-1187.
CERBAI, E., BARBIERI, M., PORCIATTI, F. & MUGELLI, A. (1994b).The hyperpolarization-activated current If in rat leftventricular myocytes is modulated by hypertrophy. Journal ofMolecular and Cellular Cardiology 26, 89 (abstract).
DIFRANCESCO, D. (1981a). A new interpretation of the pace-makercurrent in calf Purkinje fibres. Journal of Physiology 314,359-376.
DIFRANCESCO, D. (1981b). A study of the ionic nature of the pace-maker current of the pacemaker current in calf Purkinje fibres.Journal of Physiology 314, 377-393.
DIFRANCESCO, D. (1982). Block and activation of the pace-makerchannel in calf Purkinje fibres: effects of potassium, caesiumand rubidium. Journal of Physiology 329, 485-507.
DIFRANCESCO, D. (1985). The cardiac hyperpolarizing-activatedcurrent, if. Origins and development. Progress in Biophysicsand Molecular Biology 46, 163-183.
DIFRANcESco, D. (1991). The contribution of the 'pacemaker'current (if) to generation of spontaneous activity in rabbit sino-atrial node myocytes. Journal of Physiology 434, 23-40.
DIFRANcEScO, D. (1993). Pacemaker mechanisms in cardiac tissue.Annual Review of Physiology 55, 455-472.
DIFRANCESCO, D., FERRONI, A., MAZZANTI, M. & TROMBA, C.(1986). Properties of the hyperpolarizing-activated current (if)in cells isolated from the rabbit sino-atrial node. Journal ofPhysiology 377, 61-88.
FRACE, A. M., MARUOKA, F. & NOMA, A. (1992). External K+increases Na+ conductance of the hyperpolarization-activatedcurrent in rabbit cardiac pacemaker cells. Pfluigers Archiv 421,97-99.
Ho, W. -K., BROWN, H. F. & NOBLE, D. (1994). High selectivity ofthe if channel to Na+ and K+ in rabbit sinoatrial node cells.Pfluigers Archiv 426, 68-74.
IRISAWA, H., BROWN, H. F. & GILES, W. (1993). Cardiac pace-making in the sinoatrial node. Physiological Reviews 73,197-227.
MCLENACHAN, J. M., HENDERSON, E., MORRIS, K. I. & DARGIE, H. J.(1987). Ventricular arrhythmias in patients with hypertensiveleft ventricular hypertrophy. New England Journal ofMedicine 317, 787-792.
PAHOR, M., LO GIUDICE, P., BERNABEI, R., Di GENNARO, M.,PACIFICI, L., RAMACCI, M. T. & CARBONIN, P. U. (1989). Age-related increase in the incidence of ventricular arrhythmias inisolated hearts from spontaneously hypertensive rats.Cardiovascular Drugs and Therapy 3,163-169.
Yu, H., CHANG, F. & COHEN, I. S. (1993). Pacemaker current existsin ventricular myocytes. Circulation Research 72, 232-236.
ZHOU, Z. & LiPsius, S. (1992). Properties of the pacemaker current(If) in latent pacemaker cells isolated from cat right atrium.Journal of Physiology 453, 503-523.
AcknowledgementsWe wish to thank Dr Francesco Porciatti for helpfuldiscussion. This work was partly supported by grants fromTelethon (Project No. 526) and Ministero dell'Universit'a edella Ricerca Scientifica e Tecnologica 60% (University ofFirenze).
Received 14 September 1994; accepted 14 October 1994.
) by guest on July 14, 2011jp.physoc.orgDownloaded from J Physiol (
Related Documents











