Characterization of the DEG-3/DES-2 Receptor: A Nicotinic Acetylcholine Receptor That Mutates to Cause Neuronal Degeneration Lina Yassin,* Boaz Gillo,* Tamar Kahan, ² Sarah Halevi,* Margalit Eshel,* and Millet Treinin* ,1 *Department of Physiology, Hadassah Medical School, Jerusalem 91120, Israel; and ² Computer Unit, Hadassah Medical School, Jerusalem 91120, Israel The nicotinic acetylcholine receptor family (nAChR) is a large family of acetylcholine-gated cation channels. Here we characterize the Caenorhabditis elegans DEG-3/ DES-2 nAChR, a receptor identified due to its involvement in neuronal degeneration. Pharmacological analysis of a DEG-3/DES-2 receptor expressed in Xenopus oocytes shows that this receptor is preferentially activated by cho- line. This choline sensitivity of the DEG-3/DES-2 channel can explain its role in neuronal degeneration, as shown by the toxic effects of choline on oocytes expressing the mutant DEG-3/DES-2 channel. We also show that in C. elegans the DEG-3/DES-2 receptor is localized to nonsyn- aptic regions, including the sensory endings of chemo- sensory neurons. This localization is in agreement with a role for this receptor in chemosensation of choline, as inferred from a defect in chemotaxis for choline seen in deg-3 mutants. Thus, this work also provides evidence for the diversity of nonsynaptic activities associated with nAChRs. INTRODUCTION Ligand-gated ion channels (LGIC) are a large group consisting of the nicotinic acetylcholine receptors (nAChR), 5-HT receptors, GABA receptors, and glycine receptors. Members of this gene family play a central role in fast synaptic transmission. The best-character- ized of this family are the nAChRs, which form a large and structurally heterogeneous group (Steinbach and Ifune, 1989). In Caenorhabditis elegans there are 42 differ- ent nAChR subunits (Bargmann, 1999) including two groups that are unique to C. elegans. Of these, the deg-3 group constitutes a very early branch in the nAChR family (Mongan et al., 1998; Le Novre and Changeux, 1995–2000). deg-3 codes for an a subunit of a C. elegans nAChR. This gene was identified due to a dominant allele, deg-3(u662), that results in neuronal degeneration (DEG stands for DEGeneration of certain neurons) (Treinin and Chalfie, 1995). des-2, a second gene in this group, also codes for an a subunit of a nAChR and was shown to be necessary for deg-3-dependent channel ac- tivity in vivo and when expressed in Xenopus oocytes (Treinin et al., 1998) (DES stands for DEgeneration Sup- pressor, as mutations in this gene suppress the degen- erations resulting from the deg-3(u662) mutation). deg-3 and des-2 are expressed in a small number of neurons. Their role in the activity of these cells was not known, however, as mutations eliminating the activity of these two genes are not associated with a visible phenotype. Here we present an analysis of the DEG-3/DES-2 channel. Activity of this channel was characterized fol- lowing heterologous expression in Xenopus oocytes. This analysis shows that the DEG-3/DES-2 channel has high calcium permeability and a pharmacological pro- file that shows some similarities to other divergent branches of the nAChR family (a27 and a29), with one interesting difference: preferential activation by choline relative to ACh. We also found evidence for a nonsyn- aptic role of this channel in vivo, as revealed by a chemotaxis defect observed in deg-3 mutants. In these mutants chemotaxis to choline is reduced, a result sug- gesting a role for the DEG-3/DES-2 receptor as a cho- line sensor in chemosensory neurons. Indeed this recep- 1 To whom correspondence should be addressed at Department of Physiology, Hadassah Medical School, P.O. Box 12272, Jerusalem 91120, Israel. Fax: 972-2-6439736. E-mail: [email protected]. Molecular and Cellular Neuroscience 17, 589 –599 (2001) doi:10.1006/mcne.2000.0944, available online at http://www.idealibrary.com on MCN 1044-7431/01 $35.00 Copyright © 2001 by Academic Press All rights of reproduction in any form reserved. 589

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

DiDslctmeasridtn
P9
Molecular and Cellular Neuroscience 17, 589–599 (2001)
doi:10.1006/mcne.2000.0944, available online at http://www.idealibrary.com on MCN
A
Characterization of the DEG-3/DES-2 Receptor: ANicotinic Acetylcholine Receptor That Mutates toCause Neuronal Degeneration
Lina Yassin,* Boaz Gillo,* Tamar Kahan,† Sarah Halevi,*Margalit Eshel,* and Millet Treinin* ,1
*Department of Physiology, Hadassah Medical School, Jerusalem 91120, Israel; and†Computer Unit, Hadassah Medical School, Jerusalem 91120, Israel
gf1
t
aTht
clThfib
m
The nicotinic acetylcholine receptor family (nAChR) is alarge family of acetylcholine-gated cation channels. Herewe characterize the Caenorhabditis elegans DEG-3/
ES-2 nAChR, a receptor identified due to its involvementn neuronal degeneration. Pharmacological analysis of aEG-3/DES-2 receptor expressed in Xenopus oocyteshows that this receptor is preferentially activated by cho-
ine. This choline sensitivity of the DEG-3/DES-2 channelan explain its role in neuronal degeneration, as shown byhe toxic effects of choline on oocytes expressing theutant DEG-3/DES-2 channel. We also show that in C.
legans the DEG-3/DES-2 receptor is localized to nonsyn-ptic regions, including the sensory endings of chemo-ensory neurons. This localization is in agreement with aole for this receptor in chemosensation of choline, asnferred from a defect in chemotaxis for choline seen ineg-3 mutants. Thus, this work also provides evidence forhe diversity of nonsynaptic activities associated withAChRs.
INTRODUCTION
Ligand-gated ion channels (LGIC) are a large groupconsisting of the nicotinic acetylcholine receptors(nAChR), 5-HT receptors, GABA receptors, and glycinereceptors. Members of this gene family play a centralrole in fast synaptic transmission. The best-character-ized of this family are the nAChRs, which form a largeand structurally heterogeneous group (Steinbach andIfune, 1989). In Caenorhabditis elegans there are 42 differ-
1 To whom correspondence should be addressed at Department of
glhysiology, Hadassah Medical School, P.O. Box 12272, Jerusalem1120, Israel. Fax: 972-2-6439736. E-mail: [email protected].
1044-7431/01 $35.00Copyright © 2001 by Academic Press
ll rights of reproduction in any form reserved.
ent nAChR subunits (Bargmann, 1999) including twogroups that are unique to C. elegans. Of these, the deg-3
roup constitutes a very early branch in the nAChRamily (Mongan et al., 1998; Le Novre and Changeux,995–2000). deg-3 codes for an a subunit of a C. elegans
nAChR. This gene was identified due to a dominantallele, deg-3(u662), that results in neuronal degeneration(DEG stands for DEGeneration of certain neurons)(Treinin and Chalfie, 1995). des-2, a second gene in thisgroup, also codes for an a subunit of a nAChR and wasshown to be necessary for deg-3-dependent channel ac-ivity in vivo and when expressed in Xenopus oocytes
(Treinin et al., 1998) (DES stands for DEgeneration Sup-pressor, as mutations in this gene suppress the degen-erations resulting from the deg-3(u662) mutation). deg-3nd des-2 are expressed in a small number of neurons.heir role in the activity of these cells was not known,owever, as mutations eliminating the activity of these
wo genes are not associated with a visible phenotype.Here we present an analysis of the DEG-3/DES-2
hannel. Activity of this channel was characterized fol-owing heterologous expression in Xenopus oocytes.his analysis shows that the DEG-3/DES-2 channel hasigh calcium permeability and a pharmacological pro-le that shows some similarities to other divergentranches of the nAChR family (a27 and a29), with one
interesting difference: preferential activation by cholinerelative to ACh. We also found evidence for a nonsyn-aptic role of this channel in vivo, as revealed by achemotaxis defect observed in deg-3 mutants. In these
utants chemotaxis to choline is reduced, a result sug-
esting a role for the DEG-3/DES-2 receptor as a cho-ine sensor in chemosensory neurons. Indeed this recep-
589
tcwti
aiebd
att
wrCmcsa
1iCat
Ddwf
N
590 Yassin et al.
tor is localized to sensory endings of chemosensoryneurons as seen following antibody staining.
We have previously shown that a mutant DEG-3(u662)/DES-2 channel has altered kinetics, specificallyshowing no decay of channel activity in the presence ofits agonist (Treinin et al., 1998). However, it was notclear how these changes in the activity of the mutantchannel contribute to the degeneration process. Herewe show that a low concentration of choline, a concen-tration found in normal serum (Zhou et al., 1999), isoxic to oocytes expressing the mutant DEG-3/DES-2hannel much more than to oocytes expressing theild-type channel. This toxicity of choline may underlie
he degeneration of neurons seen in nematodes carry-ng the deg-3(u662) mutation.
RESULTS
Pharmacological Analysis of theDEG-3/DES-2 Channel
Sequence analysis of deg-3 and des-2 show that bothre highly divergent members of the nAChR family, asdentity with other members of this family does notxceed 35%. These limited homologies to otherranches of the nAChR family make assignment ofeg-3 and des-2 to any of the known nAChR branches
impossible. Indeed, phylogenetic analysis done byMongan et al. (1998) and Le Novre and Changeux(1995–2000) shows that deg-3 and des-2 are members of
yet uncharacterized group. This group emerged fromhe nAChR tree early in evolution and is so far uniqueo C. elegans.
In order to characterize the DEG-3/DES-2 receptore used pharmacological analysis of the DEG-3/DES-2
eceptor expressed in Xenopus oocytes (Treinin andhalfie, 1995). For this purpose we recorded the maxi-al current amplitude of the responses to bath appli-
ations of agonists and antagonists. Previously we havehown that ACh, the classical agonist of nAChRs, is angonist of the DEG-3/DES-2 channel (Treinin et al.,
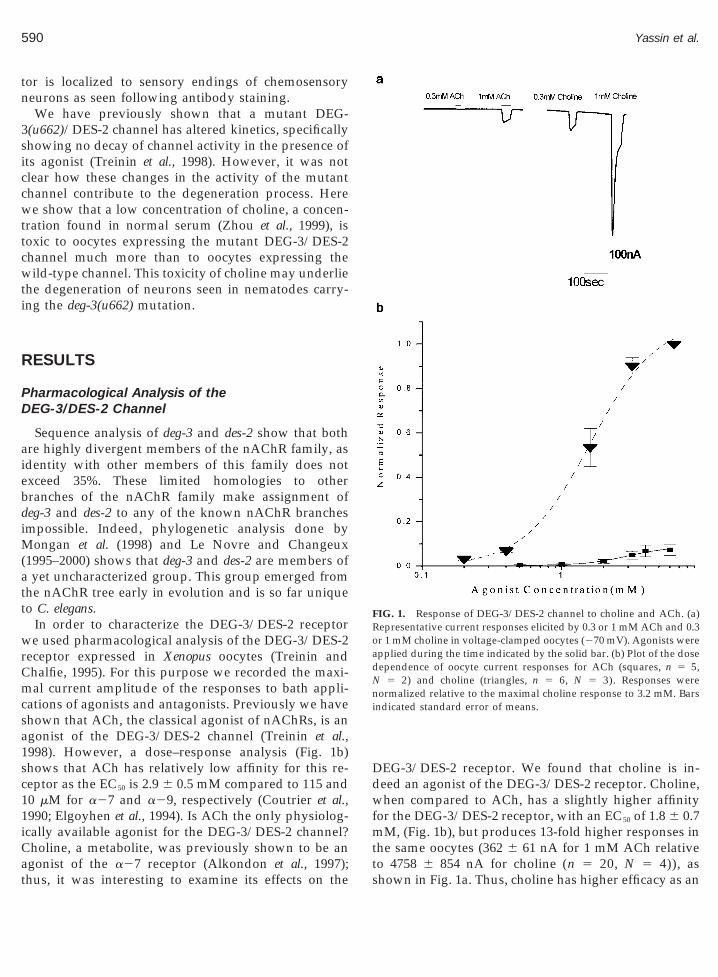
1998). However, a dose–response analysis (Fig. 1b)shows that ACh has relatively low affinity for this re-ceptor as the EC50 is 2.9 6 0.5 mM compared to 115 and10 mM for a27 and a29, respectively (Coutrier et al.,990; Elgoyhen et al., 1994). Is ACh the only physiolog-cally available agonist for the DEG-3/DES-2 channel?holine, a metabolite, was previously shown to be an
gonist of the a27 receptor (Alkondon et al., 1997);hus, it was interesting to examine its effects on the sEG-3/DES-2 receptor. We found that choline is in-eed an agonist of the DEG-3/DES-2 receptor. Choline,hen compared to ACh, has a slightly higher affinity
or the DEG-3/DES-2 receptor, with an EC50 of 1.8 6 0.7mM, (Fig. 1b), but produces 13-fold higher responses inthe same oocytes (362 6 61 nA for 1 mM ACh relative
FIG. 1. Response of DEG-3/DES-2 channel to choline and ACh. (a)Representative current responses elicited by 0.3 or 1 mM ACh and 0.3or 1 mM choline in voltage-clamped oocytes (270 mV). Agonists wereapplied during the time indicated by the solid bar. (b) Plot of the dosedependence of oocyte current responses for ACh (squares, n 5 5,
5 2) and choline (triangles, n 5 6, N 5 3). Responses werenormalized relative to the maximal choline response to 3.2 mM. Barsindicated standard error of means.
to 4758 6 854 nA for choline (n 5 20, N 5 4)), ashown in Fig. 1a. Thus, choline has higher efficacy as an
aITi
ept
CO
c
1iD(gsirliiamiicedc
3c
591Characterization of the DEG-3/DES-2 nAChR
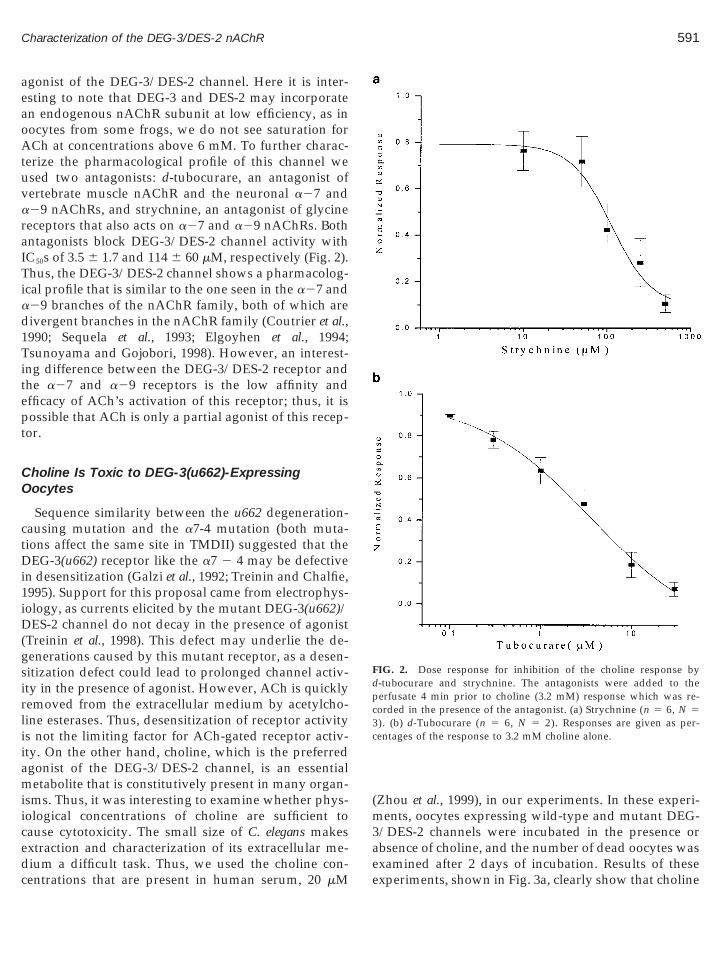
agonist of the DEG-3/DES-2 channel. Here it is inter-esting to note that DEG-3 and DES-2 may incorporatean endogenous nAChR subunit at low efficiency, as inoocytes from some frogs, we do not see saturation forACh at concentrations above 6 mM. To further charac-terize the pharmacological profile of this channel weused two antagonists: d-tubocurare, an antagonist ofvertebrate muscle nAChR and the neuronal a27 anda29 nAChRs, and strychnine, an antagonist of glycinereceptors that also acts on a27 and a29 nAChRs. Bothntagonists block DEG-3/DES-2 channel activity withC50s of 3.5 6 1.7 and 114 6 60 mM, respectively (Fig. 2).hus, the DEG-3/DES-2 channel shows a pharmacolog-
cal profile that is similar to the one seen in the a27 anda29 branches of the nAChR family, both of which aredivergent branches in the nAChR family (Coutrier et al.,1990; Sequela et al., 1993; Elgoyhen et al., 1994;Tsunoyama and Gojobori, 1998). However, an interest-ing difference between the DEG-3/DES-2 receptor andthe a27 and a29 receptors is the low affinity andfficacy of ACh’s activation of this receptor; thus, it isossible that ACh is only a partial agonist of this recep-
or.
holine Is Toxic to DEG-3(u662)-Expressingocytes
Sequence similarity between the u662 degeneration-ausing mutation and the a7-4 mutation (both muta-
tions affect the same site in TMDII) suggested that theDEG-3(u662) receptor like the a7 2 4 may be defectivein desensitization (Galzi et al., 1992; Treinin and Chalfie,995). Support for this proposal came from electrophys-ology, as currents elicited by the mutant DEG-3(u662)/ES-2 channel do not decay in the presence of agonist
Treinin et al., 1998). This defect may underlie the de-enerations caused by this mutant receptor, as a desen-itization defect could lead to prolonged channel activ-ty in the presence of agonist. However, ACh is quicklyemoved from the extracellular medium by acetylcho-ine esterases. Thus, desensitization of receptor activitys not the limiting factor for ACh-gated receptor activ-ty. On the other hand, choline, which is the preferredgonist of the DEG-3/DES-2 channel, is an essentialetabolite that is constitutively present in many organ-
sms. Thus, it was interesting to examine whether phys-ological concentrations of choline are sufficient toause cytotoxicity. The small size of C. elegans makesxtraction and characterization of its extracellular me-
ium a difficult task. Thus, we used the choline con-entrations that are present in human serum, 20 mM(Zhou et al., 1999), in our experiments. In these experi-ments, oocytes expressing wild-type and mutant DEG-3/DES-2 channels were incubated in the presence orabsence of choline, and the number of dead oocytes was
FIG. 2. Dose response for inhibition of the choline response byd-tubocurare and strychnine. The antagonists were added to theperfusate 4 min prior to choline (3.2 mM) response which was re-corded in the presence of the antagonist. (a) Strychnine (n 5 6, N 5
). (b) d-Tubocurare (n 5 6, N 5 2). Responses are given as per-entages of the response to 3.2 mM choline alone.
examined after 2 days of incubation. Results of theseexperiments, shown in Fig. 3a, clearly show that choline
tefrdet
592 Yassin et al.
concentrations as low as 20 mM are significantly moretoxic to oocytes expressing the mutant DEG-3(u662)/
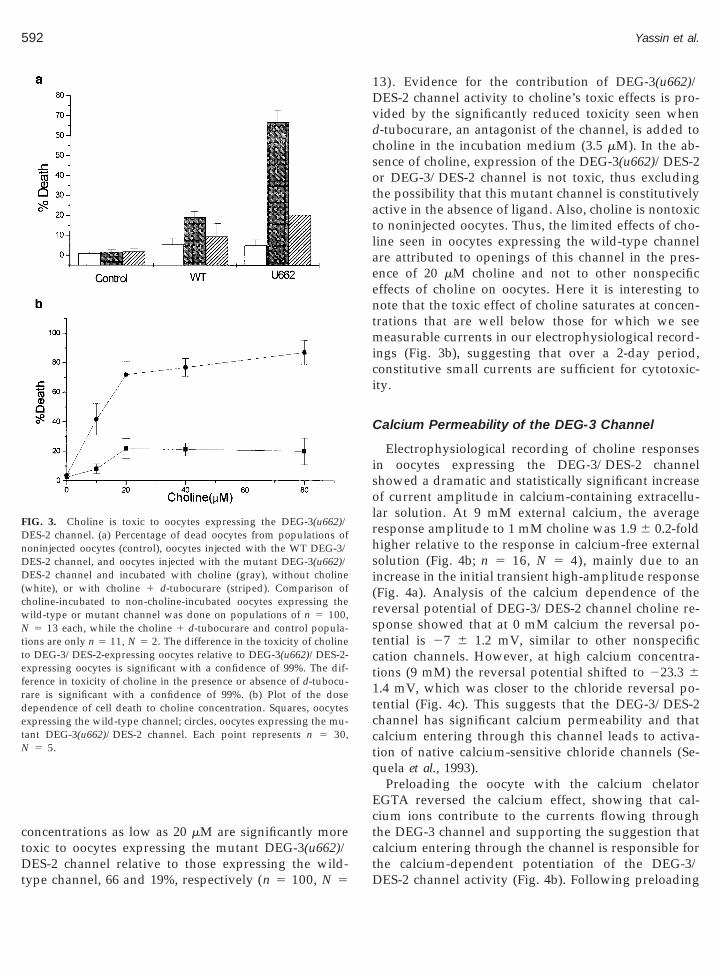
FIG. 3. Choline is toxic to oocytes expressing the DEG-3(u662)/DES-2 channel. (a) Percentage of dead oocytes from populations ofnoninjected oocytes (control), oocytes injected with the WT DEG-3/DES-2 channel, and oocytes injected with the mutant DEG-3(u662)/DES-2 channel and incubated with choline (gray), without choline(white), or with choline 1 d-tubocurare (striped). Comparison ofcholine-incubated to non-choline-incubated oocytes expressing thewild-type or mutant channel was done on populations of n 5 100,N 5 13 each, while the choline 1 d-tubocurare and control popula-tions are only n 5 11, N 5 2. The difference in the toxicity of cholineo DEG-3/DES-2-expressing oocytes relative to DEG-3(u662)/DES-2-xpressing oocytes is significant with a confidence of 99%. The dif-erence in toxicity of choline in the presence or absence of d-tubocu-are is significant with a confidence of 99%. (b) Plot of the doseependence of cell death to choline concentration. Squares, oocytesxpressing the wild-type channel; circles, oocytes expressing the mu-ant DEG-3(u662)/DES-2 channel. Each point represents n 5 30,
N 5 5.
DES-2 channel relative to those expressing the wild-type channel, 66 and 19%, respectively (n 5 100, N 5
13). Evidence for the contribution of DEG-3(u662)/DES-2 channel activity to choline’s toxic effects is pro-vided by the significantly reduced toxicity seen whend-tubocurare, an antagonist of the channel, is added tocholine in the incubation medium (3.5 mM). In the ab-sence of choline, expression of the DEG-3(u662)/DES-2or DEG-3/DES-2 channel is not toxic, thus excludingthe possibility that this mutant channel is constitutivelyactive in the absence of ligand. Also, choline is nontoxicto noninjected oocytes. Thus, the limited effects of cho-line seen in oocytes expressing the wild-type channelare attributed to openings of this channel in the pres-ence of 20 mM choline and not to other nonspecificeffects of choline on oocytes. Here it is interesting tonote that the toxic effect of choline saturates at concen-trations that are well below those for which we seemeasurable currents in our electrophysiological record-ings (Fig. 3b), suggesting that over a 2-day period,constitutive small currents are sufficient for cytotoxic-ity.
Calcium Permeability of the DEG-3 Channel
Electrophysiological recording of choline responsesin oocytes expressing the DEG-3/DES-2 channelshowed a dramatic and statistically significant increaseof current amplitude in calcium-containing extracellu-lar solution. At 9 mM external calcium, the averageresponse amplitude to 1 mM choline was 1.9 6 0.2-foldhigher relative to the response in calcium-free externalsolution (Fig. 4b; n 5 16, N 5 4), mainly due to anincrease in the initial transient high-amplitude response(Fig. 4a). Analysis of the calcium dependence of thereversal potential of DEG-3/DES-2 channel choline re-sponse showed that at 0 mM calcium the reversal po-tential is 27 6 1.2 mV, similar to other nonspecificcation channels. However, at high calcium concentra-tions (9 mM) the reversal potential shifted to 223.3 61.4 mV, which was closer to the chloride reversal po-tential (Fig. 4c). This suggests that the DEG-3/DES-2channel has significant calcium permeability and thatcalcium entering through this channel leads to activa-tion of native calcium-sensitive chloride channels (Se-quela et al., 1993).
Preloading the oocyte with the calcium chelatorEGTA reversed the calcium effect, showing that cal-cium ions contribute to the currents flowing throughthe DEG-3 channel and supporting the suggestion thatcalcium entering through the channel is responsible for
the calcium-dependent potentiation of the DEG-3/DES-2 channel activity (Fig. 4b). Following preloading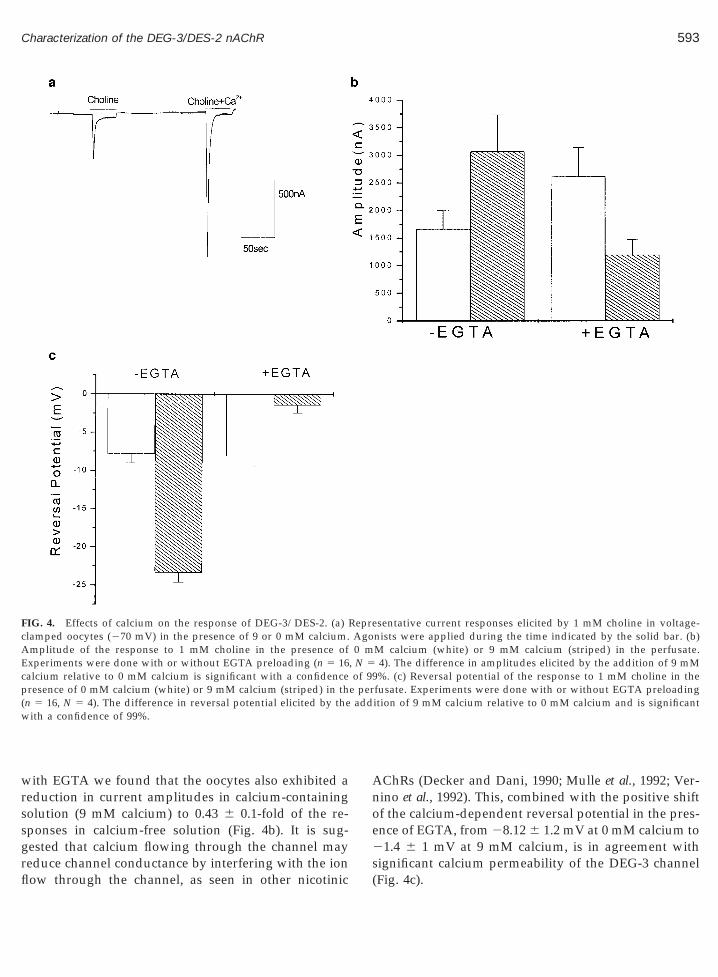
sgrfl
Anoe2
AE
add
593Characterization of the DEG-3/DES-2 nAChR
with EGTA we found that the oocytes also exhibited areduction in current amplitudes in calcium-containingsolution (9 mM calcium) to 0.43 6 0.1-fold of the re-ponses in calcium-free solution (Fig. 4b). It is sug-ested that calcium flowing through the channel may
FIG. 4. Effects of calcium on the response of DEG-3/DES-2. (a)clamped oocytes (270 mV) in the presence of 9 or 0 mM calcium.
mplitude of the response to 1 mM choline in the presence ofxperiments were done with or without EGTA preloading (n 5 16
calcium relative to 0 mM calcium is significant with a confidencepresence of 0 mM calcium (white) or 9 mM calcium (striped) in th(n 5 16, N 5 4). The difference in reversal potential elicited by thewith a confidence of 99%.
educe channel conductance by interfering with the ionow through the channel, as seen in other nicotinic
s(
ChRs (Decker and Dani, 1990; Mulle et al., 1992; Ver-ino et al., 1992). This, combined with the positive shiftf the calcium-dependent reversal potential in the pres-nce of EGTA, from 28.12 6 1.2 mV at 0 mM calcium to1.4 6 1 mV at 9 mM calcium, is in agreement with
sentative current responses elicited by 1 mM choline in voltage-nists were applied during the time indicated by the solid bar. (b)M calcium (white) or 9 mM calcium (striped) in the perfusate.4). The difference in amplitudes elicited by the addition of 9 mM%. (c) Reversal potential of the response to 1 mM choline in the
fusate. Experiments were done with or without EGTA preloadingition of 9 mM calcium relative to 0 mM calcium and is significant
RepreAgo0 m
, N 5of 99
e per
ignificant calcium permeability of the DEG-3 channelFig. 4c).
arpaaiccrntiitD
tsfis3t5c5pnpmctst
e sec
594 Yassin et al.
The DEG-3/DES-2 Channel Does Not ShowSynaptic Localization
Previous analysis using reporter genes showed ex-pression of deg-3 and des-2 in the sensory neurons PVDnd IL2s (Treinin et al., 1998), neurons that are known toeceive limited synaptic inputs, and specifically no in-uts from cholinergic neurons (White et al., 1986; Randnd Nonet, 1997). This observation suggested a nonsyn-ptic role for the DEG-3/DES-2 receptor. However, asnformation on synaptic connections outside the ventralord or nerve ring is incomplete, no final conclusionsould be made from these observations. The cholineesponse of the DEG-3/DES-2 channel also suggested aonsynaptic role for this receptor, as choline is unlikely
o be specifically released in synapses. To further exam-ne this possibility we characterized the cellular local-zation of the DEG-3/DES-2 receptor using antibodies
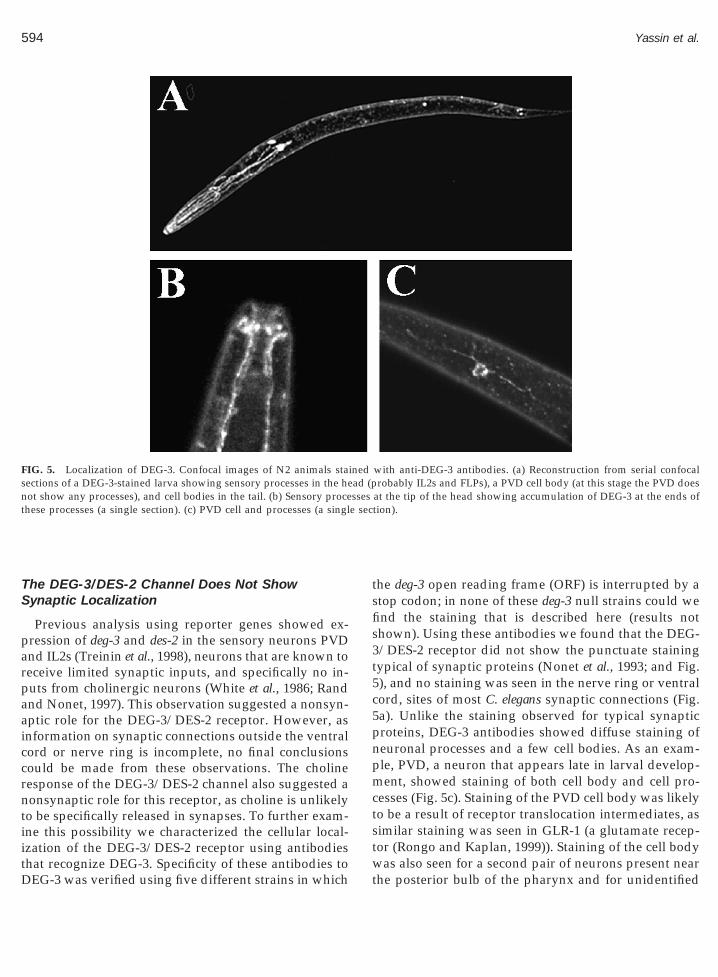
FIG. 5. Localization of DEG-3. Confocal images of N2 animals stasections of a DEG-3-stained larva showing sensory processes in the henot show any processes), and cell bodies in the tail. (b) Sensory procethese processes (a single section). (c) PVD cell and processes (a singl
hat recognize DEG-3. Specificity of these antibodies toEG-3 was verified using five different strains in which
wt
he deg-3 open reading frame (ORF) is interrupted by atop codon; in none of these deg-3 null strains could wend the staining that is described here (results nothown). Using these antibodies we found that the DEG-/DES-2 receptor did not show the punctuate stainingypical of synaptic proteins (Nonet et al., 1993; and Fig.), and no staining was seen in the nerve ring or ventralord, sites of most C. elegans synaptic connections (Fig.a). Unlike the staining observed for typical synapticroteins, DEG-3 antibodies showed diffuse staining ofeuronal processes and a few cell bodies. As an exam-le, PVD, a neuron that appears late in larval develop-ent, showed staining of both cell body and cell pro-
esses (Fig. 5c). Staining of the PVD cell body was likelyo be a result of receptor translocation intermediates, asimilar staining was seen in GLR-1 (a glutamate recep-or (Rongo and Kaplan, 1999)). Staining of the cell body
with anti-DEG-3 antibodies. (a) Reconstruction from serial confocalrobably IL2s and FLPs), a PVD cell body (at this stage the PVD does
at the tip of the head showing accumulation of DEG-3 at the ends oftion).
inedad (psses
as also seen for a second pair of neurons present nearhe posterior bulb of the pharynx and for unidentified

(ocr
i
rafwc1wmmdaT
itanmmi(coae
d3aArnoDnt2otttfilcse
595Characterization of the DEG-3/DES-2 nAChR
cells in the tail. In addition, we usually observed agroup of neuronal processes leading to the tip of thehead (Fig. 5a). These processes are likely to belong tothe FLP sensory neurons and the IL-2 chemosensoryneurons. Interestingly, in these processes we see stron-ger staining in the sensory endings, localization that isindicative for a role in sensory transduction (Fig. 5b).
Chemotaxis Defects in deg-3 Mutants
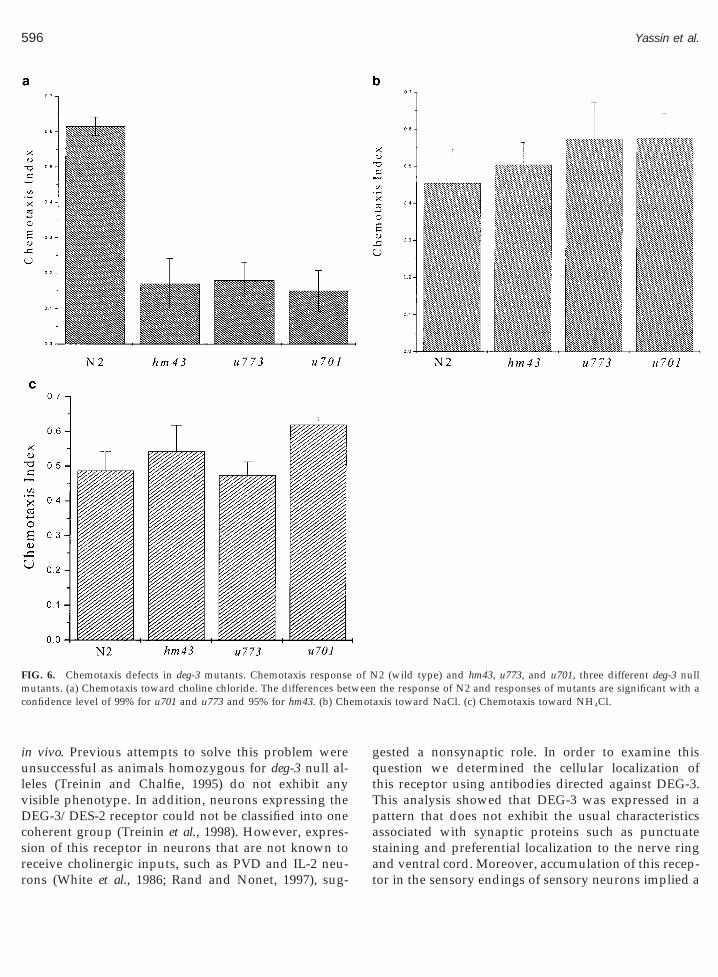
The DEG-3/DES-2 receptor is activated by cholineand DEG-3 is localized to the sensory endings of che-mosensory neurons. These results suggested a role forthis receptor in chemosensation of choline. In order toexamine this possibility it was necessary to assay C.elegans for its ability to chemotax toward choline. In-deed wild-type N2 animals show robust chemotaxistoward choline chloride, and this chemotaxis is signif-icantly reduced in mutant strains lacking the deg-3 geneFig. 6a). Similar chemotaxis in wild type and reductionf chemotaxis in deg-3 mutants is observed when usingholine bicarbonate (results not shown). The decreasedesponse to choline seen in deg-3 mutants cannot be
explained by a general reduction in chemotaxis to wa-ter-soluble attractants since response to NaCl or NH4Cls not reduced (Figs. 6b and 6c). NH4Cl specifically has
been used to examine the chemotactic response to chlo-ride ions (Bargmann and Horvitz, 1991). Thus, theseresults also show that reduction in the response tochloride is not the cause of the decreased response ofdeg-3 mutants to choline chloride. Also, we did not seea decrease in the response to the volatile odorant ben-zaldehyde (results not shown), again demonstrating thecholine specificity of this chemotactic defect. Thus, it islikely that the choline-activated DEG-3/DES-2 channelis specifically needed for the chemotactic response tocholine, implying a direct role in sensing this metabo-lite. This suggestion is in agreement with the localiza-tion of this receptor to sensory processes at the tip of thehead, processes that probably belong to the IL-2 che-mosensory neurons.
DISCUSSION
DEG-3 and DES-2 are members of a divergent branchof the nAChR family. Using pharmacological analysison a DEG-3/DES-2 receptor expressed in Xenopus oo-cytes, we show that this receptor is activated by ACh
and choline and antagonized by d-tubocurare andstrychnine. This pharmacological profile is similar to Dthe one known for a27 (Sequela et al., 1993; Alkondonet al., 1997). However, unlike a27, the DEG-3/DES-2eceptor is preferentially activated by choline, while theffinity for ACh is 10- to 100-fold lower than the affinityound in other nAChRs. Another feature that is shared
ith other branches of the nAChR family is high cal-ium permeability (Sequela et al., 1993; Elgoyhen et al.,994). Such a feature provides these receptors with aide range of signaling properties. Specifically for theutant DEG-3(u662)/DES-2 receptor, high calcium per-eability is likely to play an important role in the
egeneration process that results from the deregulatedctivity of this receptor (Treinin and Chalfie, 1995;reinin et al., 1998).The DEG-3 nAChR subunit was identified due to its
nvolvement in neuronal degeneration. The degenera-ion-causing mutation deg-3(u662) produces receptorctivity that does not decay in the presence of its ago-ist (Treinin et al., 1998). It is postulated that thisutation, like other C. elegans degeneration-causingutations, leads to cell death through a mechan-
sm that depends on increased channel activityGarcia-Anoveros et al., 1998). The cause for this in-reased channel activity is probably lack of inactivationf the mutant channel in the presence of agonist (prob-bly a desensitization defect), as other possible differ-nces such as decreased EC50 or increased whole cell
currents were not seen. However, in vivo, ACh-depen-ent activation of the degeneration-causing DEG-(u662)/DES-2 channel is unlikely to cause prolongedctivation, since acetylcholine esterases quickly removeCh from the extracellular medium, thus terminating
eceptor activity. Such a backup termination mecha-ism is not available for choline-dependent activationf this receptor. Thus, the unique pharmacology of theEG-3/DES-2 receptor may underlie its involvement ineuronal degeneration. Indeed choline at a concentra-
ion of 20 mM is much more toxic to DEG-3(u662)/DES--expressing oocytes than to DEG-3/DES-2-expressingocytes. This concentration of choline is found consti-utively in human serum (Zhou et al., 1999). Althoughhe small size of C. elegans makes it technically difficulto extract and characterize the extracellular mediumrom this organism, we know that in C. elegans cholines an essential metabolite (Vanfleteren, 1980). Thus, it isikely that the extracellular medium of C. elegans willontain similar low constitutive levels of choline, levelsufficient to exert toxic effects on DEG-3(u662)/DES-2-xpressing neurons.
In order to understand the activity of the DEG-3/ES-2 receptor, it is necessary to know what role it has
ulvDc
m weenc emot
596 Yassin et al.
in vivo. Previous attempts to solve this problem werensuccessful as animals homozygous for deg-3 null al-
eles (Treinin and Chalfie, 1995) do not exhibit anyisible phenotype. In addition, neurons expressing theEG-3/DES-2 receptor could not be classified into one
oherent group (Treinin et al., 1998). However, expres-sion of this receptor in neurons that are not known to
FIG. 6. Chemotaxis defects in deg-3 mutants. Chemotaxis responseutants. (a) Chemotaxis toward choline chloride. The differences bet
onfidence level of 99% for u701 and u773 and 95% for hm43. (b) Ch
receive cholinergic inputs, such as PVD and IL-2 neu-rons (White et al., 1986; Rand and Nonet, 1997), sug-
gested a nonsynaptic role. In order to examine thisquestion we determined the cellular localization ofthis receptor using antibodies directed against DEG-3.This analysis showed that DEG-3 was expressed in apattern that does not exhibit the usual characteristicsassociated with synaptic proteins such as punctuatestaining and preferential localization to the nerve ring
2 (wild type) and hm43, u773, and u701, three different deg-3 nullthe response of N2 and responses of mutants are significant with a
axis toward NaCl. (c) Chemotaxis toward NH4Cl.
of N
and ventral cord. Moreover, accumulation of this recep-tor in the sensory endings of sensory neurons implied a

Wt
(tcdn
owitoDa
597Characterization of the DEG-3/DES-2 nAChR
role in sensory transduction. Indeed, animals homozy-gous for deg-3 mutations have a defect in chemotaxisthat is specific to choline. This finding, aside from iden-tifying a function associated with the DEG-3/DES-2receptor, supports an in vivo role for choline activationof the DEG-3/DES-2 channel. The specificity of thechemotaxis defect and the localization of the receptor tosensory endings are in agreement with a role of theDEG-3/DES-2 receptor as a sensor for choline, directingchemotaxis toward this important metabolite (Van-fleteren, 1980).
The presence of constitutive levels of choline in theextracellular fluid could potentially interfere with che-mosensation toward exogenous choline. However, thisis unlikely for two reasons: First, prolonged incubationin low (20 mM) choline does not lead to receptor desen-sitization (L. Yassin, unpublished results), and second,sensory endings of chemosensory neurons are open tothe outside environment and are unlikely to be affectedby the internal choline concentrations.
A role for an nAChR in chemotaxis toward a water-soluble chemical (the equivalent of taste reception) is anovel finding, as nAChRs have not been previouslyimplicated in such a function. However, nAChRs wereshown to function in chemotaxis of single cells, a27was implicated in neuronal migration toward ACh(Zheng et al., 1994), and an nAChR-dependent chemo-taxis toward acetylcholine was also shown for keratin-ocytes (Grando, 1997). Thus, involvement of nAChRs inchemotaxis is not completely novel.
One striking result of the analysis of the DEG-3/DES-2 receptor is its preference for choline vs ACh.Previously it was shown that a27 is also activated bycholine although at lower affinity relative to ACh. Didthis choline sensitivity appear independently in thesetwo separate branches of the nAChR family tree? Anapproach toward answering this question is suggestedby two facts: First, molecular clock analysis suggeststhat the LGIC family emerged at least 2500 millionyears ago, before the first eukaryotes (Ortells and Lunt,1995), suggesting a nonneuronal function for the ances-tor of this family. Second, choline, like glycine, seroto-nin, and glutamate, and unlike ACh, is a metabolite.Thus, the first ligand-gated channels could not havehad a role in neuronal activity; instead, a role in che-mosensation of metabolites that are the ligands of thesereceptors is a distinct possibility. Support for this sug-gestion is provided by the known chemotactic re-sponses of protozoans to a number of neurotransmit-
ters, including choline (Hauser et al., 1975). Althoughthe molecules mediating these activities were not char-Sp
acterized, ancestors of the ligand-gated ion channels arelikely candidates. Here it is important to note that thehigh calcium permeability found in some nAChRs pro-vides a mechanism that could link between ligand bind-ing and rearrangements of the cytoskeleton that arenecessary for chemotaxis of the responding cell (re-viewed in Mooseker et al., 1986). A role for ligand-gatedchannels in chemosensation of metabolites may alsoexplain the presence of a glutamate-gated potassiumchannel in bacteria (Chen et al., 1999). We also speculatethat high sensitivity to ACh, which is not a metabolite,appeared late in evolution in order to enable fast ter-mination of synaptic signaling by acetylcholine ester-ases. Fast termination of synaptic signaling is essentialfor both information coding and as a safeguard againstneuronal degeneration.
EXPERIMENTAL METHODS
Heterologous Expression and Electrophysiology
des-2 and deg-3 cRNAs (5 ng each) were injected intoXenopus oocytes and the resulting channel activity wasanalyzed as previously described (Treinin et al., 1998).
hen not otherwise indicated, oocytes were main-ained in ND96 with 5 mM MgCl2 and 0 mM CaCl2
(Gillo et al., 1987). Reversal potential was examinedusing a 1.2-s voltage ramp from 270 to 130 mV. ForEGTA preloading experiments, 50 nl of 25 mM EGTAwas injected into oocytes. Curves were fitted to thelogarithmic equation ( A 1 2 A 2)/(1 1 (X/X 0)
P) 1 A 2,where A 1 is the initial normalized response, A 2 is thefinal normalized response, X 0 is EC50, and P is powerHill coefficient). Hill coefficient values provided byhis analysis are 2, 3, 1, and 2 for ACh, choline, tubo-urare, and strychnine, respectively. This analysis wasone using the Origin 5.0 program. N indicates theumber of frogs and n the number of oocytes assayed.Choline toxicity assays were performed as follows:
ocytes were injected with des-2 cRNA combined withild-type or mutant deg-3 cRNA. One day following
njection, choline or choline 1 d-tubocurare was addedo some of these oocytes, and the number of deadocytes was examined after 2 days of such incubation.ead oocytes have severe loss of pigmentation from the
nimal hemisphere or the appearance of spots or holes.
ignificance of the results was examined using a two-opulation t test (Sigma plot).
C
Bgb
B
C
C
G
G
G
G
H
L
M
M
M
N
N
O
R
R
598 Yassin et al.
Immunohistochemistry
Rabbit polyclonal antibodies were generated by in-jection of a KLH-conjugated synthetic peptide identicalto the C-terminus of DEG-3 (AQYSYDHPDDRFFNV, alysine was added at the N-terminal of this peptide tofacilitate conjugation to KLH). Prior to staining, theserum was affinity purified on Affigel 15. The secondantibody is goat anti-rabbit, Alexa 488, purchased fromMolecular Probes, at a dilution of 1:150. Staining wasperformed using a picric acid fixation protocol asadapted to C. elegans in Nonet et al. (1997).
hemotaxis Assays
Chemotaxis assays were performed as described inargmann et al. (1993), except for the generation of theradient. The gradient was produced by agar plugs cuty a 1-ml blue pipet tip that were soaked in 20 ml
choline chloride (1 M) or double-distilled water for thecholine experiments or 40 ml for the NaCl (1 M) orNH4Cl (1 M) for the chloride control experiment. Plugswere positioned on the assay plates for 6 h prior to theexperiment in order to build the gradient. Significanceof the results was examined using a two-population ttest. In each experiment 100–200 adults were tested.
ACKNOWLEDGMENTS
We thank B. Minke for his generosity in providing us with frogsand electrophysiological facilities and Y. Stern-Bach for critical read-ing of the manuscript. This work was supported by U.S.–Israel Bina-tional Science Foundation Grant 95-0133/2 and by the Israel ScienceFoundation founded by the Israel Academy of Sciences and Humani-ties–DOROT Science Fellowship Foundation.
REFERENCES
Alkondon, M., Pereira, E. F. R., Cortes, W. S., Maelicke, A., andAlbuquerque, E. X. (1997). Choline is a selective agonist of a7nicotinic acetylcholine receptors in the rat brain neurons. Eur.J. Neurosci. 9: 2734–2742.
argmann, C. I. (1998). Neurobiology of the Caenorhabditis elegansgenome. Science 282: 2028–2033.
Bargmann, C. I., Hartwieg, E., and Horvitz, H. R. (1993). Odorant-selective genes and neurons mediate olfaction in C. elegans. Cell 74:515–527.
Bargmann, C. I., and Horvitz, H. R. (1991). Chemosensory neuronswith overlapping functions direct chemotaxis to multiple chemicals
in C. elegans. Neuron 7: 729–742.hen, G-Q., Cui, C., Mayer M. L., and Gouaux, E. (1999). Functional S
characterization of a potassium-selective prokaryotic glutamate re-ceptor. Nature 402: 817–821.
outrier, S., Bertrand, D., Matter, J. M., Hernandez, M.-C., Bertrand,S., Millar, N., Valera, S., Barkas, T., and Ballivet, M. (1990). Aneuronal nicotinic acetylcholine receptor subunit (a27) is develop-mentally regulated and forms a homo-oligomeric channel blockedby alpha-BTX. Neuron 5: 847–856.
Decker, E. R., and Dani, J. A. (1990). Calcium permeability of thenicotinic acetylcholine receptor: The single-channel calcium influxis significant. J. Neurosci. 10: 3413–3420.
Elgoyhen, A. B., Johnson, D. S., Boulter, J., Vetter, D. E., and Heine-mann, S. (1994). a29: An acetylcholine receptor with novel phar-macological properties expressed in rat chochlear hair cells. Cell 79:705–715.alzi, J-L., Devillers-Thiery, A., Hussy, N., Bertrand, S., Changeux,J-P., and Bertrand, D. (1992). Mutations in the channel domain of aneuronal nicotinic receptor convert ion selectivity from cationic toanionic. Nature 359: 500–505.arcia-Anoveros, J., Garcia, J. A., Liu, J-D., and Corey, D. P. (1998).The nematode degenerin UNC-105 forms ion channels that areactivated by degeneration or hypercontraction-causing mutations.Neuron 20: 1231–1241.illo, B., Lass, Y., Nadler, E., and Oron, Y. (1987). The involvement ofinositol 1,4,5-triphosphate and calcium in the two-component re-sponse to acetylcholine in Xenopus leavis oocytes. J. Physiol. London392: 349–361.rando, S. A. (1997). Biological functions of keratinocyte cholinergicreceptors. J. Invest. Dermatol. Symp. Proc. 2: 41–48.auser, D. C. R., Levandowsky, M., and Glassgold, J. M. (1975).Ultrasensitive chemosensory responses by a protozoan to epineph-rine and other neurochemicals. Science 190: 285–286.
e Novre, N., and Changeux, J.-P. (1995–2000). Phylogeny of cationicreceptor subunits. http://www.pasteur.fr/recherche/banques/LGIC/catphylogen.html.ongan, N. P., Baylis, H. A., Adcock, C., Smith, G. R., Sansom,M. S. P., and Sattelle, D. B. (1998). An extensive and diverse genefamily of nicotinic acetylcholine receptor alpha subunits in Caeno-rhabditis elegans. Receptors Channels 6: 213–228.ooseker, M. S., Coleman, T. R., and Conzelman, K. A. (1986). Cal-cium and the regulation of cytoskeletal assembly, structure andcontractility. Ciba Found Symp. 122: 232–249.ulle, C., Choquet, D., Korn, H., and Changeux, J-P. (1992). Calciuminflux through nicotinic receptor in rat central neurons: Its rele-vance to cellular regulation. Neuron 8: 135–143.onet, M. L., Grundahl, K., Meyer, B. J., and Rand, J. B. (1993).Synaptic function is impaired but not eliminated in C. elegansmutants lacking synaptotagmin. Cell 73: 1291–1305.onet, M. L., Staunton, J. E., Kilgard, M. P., Fergestad, T., Hartwieg,E., Horvitz, H. R., Jorgensen, E. M., and Meyer, B. J. (1997). Caeno-rhabditis elegans rab-3 mutant synapses exhibit impaired functionand are partially depleted of vesicles. J. Neurosci. 17: 8061–8073.rtells, M. O., and Lunt, G. G. (1995). Evolutionary history of theligand-gated ion-channel superfamily of receptors. Trends Neurosci.18: 121–127.
and, J. B., and Nonet, M. L. (1997). Neurotransmitter assignments forspecific neurons. In C. elegans II (D. L. Riddle, T. Blumenthal, B. J.Meyer, and J. R. Priess, Eds.), pp. 1049–1052. Cold Spring HarborLaboratory Press, Cold Spring Harbor, NY.
ongo, C., and Kaplan, J. M. (1999). CamKII regulates the density of
central glutamatergic synapses in vivo. Nature 402: 195–199.equela, P., Wadiche, J., Dineley-Miller, K., Dani, J. A., and Patrick,

S
T
599Characterization of the DEG-3/DES-2 nAChR
J. W. (1993). Molecular cloning, functional properties, and distribu-tion of rat brain a7: A nicotinic cation channel highly permeable tocalcium. J. Neurosci. 13: 596–604.
teinbach, J. H., and Ifune, C. (1989). How many kinds of nicotinicacetylcholine receptor are there? Trends Neurosci. 12: 3–6.
reinin, M., and Chalfie, M. (1995). A mutated acetylcholine receptorsubunit causes neuronal degeneration in C. elegans. Neuron 14:871–877.
Treinin, M., Gillo, B., Liebman, L., and Chalfie, M. (1998). Two func-tionally dependent acetylcholine subunits are encoded in a singleCaenorhabiditis elegans operon. Proc. Natl. Acad. Sci. USA 95:15492–15495.
Tsunoyama, K., and Gojobori, T. (1998). Evolution of nicotinic acetyl-choline receptors. Mol. Biol. Evol. 15: 518–527.
Vanfleteren, J. R. (1980). Nematodes as nutritional models. In Nema-todes as Biological Models. Vol. 2. Aging and Other Models Systems
(B. M. Zuckerman, Ed.), 1st ed., pp. 47–79. Academic Press, NewYork.
Vernino, S., Amador, M., Luetje, C. W., Patrick, J., and Dani, J. A.(1992). Calcium modulation and high calcium permeability of neu-ronal nicotinic acetylcholine receptors. Neuron 8: 127–134.
White, J. G., Southgate, E., Thomson, J. N., and Brenner, S. (1986). Thestructure of the nervous system of the nematode Caenorhabditiselegans. Philos. Trans. R. Soc. London Ser. B 314: 1–340.
Zheng, J. Q., Felder, M., Connor, J. A., and Poo, M. M. (1994). Turningof nerve growth cones induced by neurotransmitters. Nature 368:140–144.
Zhou, M., Engel, A. G., and Auerbach, A. (1999). Serum cholineactivates mutant acetylcholine receptors that cause slow chanel
congenital myasthenic syndromes. Proc. Natl. Acad. Sci. USA 96:10466–10471.Received August 17, 2000Revised November 20, 2000
Accepted November 30, 2000
Related Documents

![Study of Estrogen Receptor, Progesterone Receptor, …...[CANCER RESEARCH 49,4298-4304, August 1. 1989] Study of Estrogen Receptor, Progesterone Receptor, and the Estrogen-regulated](https://static.cupdf.com/doc/110x72/5f95792bbdbd5e0915333803/study-of-estrogen-receptor-progesterone-receptor-cancer-research-494298-4304.jpg)









![RECEPTOR THEORY AND PRACTICE - University of North ... receptor... · RECEPTOR THEORY AND PRACTICE ... Fractional occupancy [Ligand Receptor] ... as long as one could separate the](https://static.cupdf.com/doc/110x72/5ab205a37f8b9abc2f8d6c3c/receptor-theory-and-practice-university-of-north-receptorreceptor-theory.jpg)