RESEARCH LETTER Characterization of swarming motility in Citrobacter freundii Yanguang Cong 1 , Jing Wang 1 , Zhijin Chen 1 , Kun Xiong 1 , Qiwang Xu 2 & Fuquan Hu 1 1 Department of Microbiology, Third Military Medical University, Chongqing, China; and 2 Center of Biological Wave, Third Military Medical University, Chongqing, China Correspondence: Fuquan Hu, Department of Microbiology, Third Military Medical University, Chongqing 400038, China. Tel./fax: 186 23 6875 2834; e-mail: [email protected] Received 23 September 2010; revised 30 December 2010; accepted 18 January 2011. Final version published online 1 March 2011. DOI:10.1111/j.1574-6968.2011.02225.x Editor: Reggie Lo Keywords Citrobacter freundii; swarming; motility. Abstract Bacterial swarming motility is a flagella-dependent translocation on the surface environment. It has received extensive attention as a population behavior invol- ving numerous genes. Here, we report that Citrobacter freundii, an opportunistic pathogen, exhibits swarming movement on a solid medium surface with appro- priate agar concentration. The swarming behavior of C. freundii was described in detail. Insertional mutagenesis with transposon Mini-Tn5 was carried out to discover genetic determinants related to the swarming of C. freundii. A number of swarming genes were identified, among which flhD, motA, motB, wzx, rfaL, rfaJ, rfbX, rfaG, rcsD, rcsC, gshB, fabF , dam, pgi, and rssB have been characterized previously in other species. In mutants related to lipopolysaccharide synthesis and RcsCDB signal system, a propensity to form poorly motile bacterial aggregates on the agar surface was observed. The aggregates hampered bacterial surface migra- tion. In several mutants, the insertion sites were identified to be in the ORF of yqhC, yeeZ, CKO_03941, glgC, and ttrA, which have never been shown to be involved in swarming. Our results revealed several novel characteristics of swarming motility in C. freundii which are worthy of further study. Introduction Bacterial swarming is a flagella-dependent surface transloca- tion exhibited by a wide variety of flagellated bacteria (for a review, see Allison & Hughes, 1991; Fraser & Hughes, 1999; Harshey, 2003; Kaiser, 2007; Kearns, 2010). This form of locomotion, which was identified as forming a typical swarm colony on a solid media surface characterized by circular symmetry and regularly spaced concentric terraces, was first described by Hauser (1885) in Proteus mirabilis over a century ago. Both swimming and swarming motilities depend on bacterial flagella, but they differ in many ways. The most noticeable distinction is that swimming is an individual behavior, whereas swarming is a movement of bacterial populations. Moreover, the cells exhibit differen- tiation during swarming; they are usually elongated and hyperflagellated compared with the vegetative cells grown in liquid media (Allison & Hughes, 1991; Harshey, 2003; Rather, 2005). Swarming also shares features with other surface phenomena, such as biofilm formation and host invasion, and is associated with pathogenesis in some organisms. For example, swarming of P. mirabilis facilitates ascending colonization of the urinary tract and is conducive to biofilm formation on catheters (Allison et al., 1994; Stickler et al., 1998). Expression of flagella and virulence factors are coordinated in P. mirabilis and Serratia liquefa- ciens (Allison et al., 1992; Givskov et al., 1995). The flagellar export apparatus of Yersinia enterocolitica also functions as a secretion system for the transport of a virulence-associated phospholipase (Young et al., 1999). In many species, swarm- ing bacteria exhibit adaptive resistance to multiple antibio- tics (Butler et al., 2010). In recent years, system-screening studies in various species have revealed numerous swarming-related genes. These genes are involved in flagellar assembly, synthesis of polysaccharides, chemosensors, signal regulation, and metabolic pathways, whereas others are hypothetical genes with unknown functions (Kearns et al., 2004; Inoue et al., 2007; Overhage et al., 2007). However, the genetic determinants for this special process vary among species, indicating different swarming patterns in various swarming bacteria. Therefore, the study of swarming motility in various bacteria would facilitate a thorough understanding of this special bacterial motion. Considering that many types of genes are related to swarming motility, such a study also provides a tractable model to study the function of FEMS Microbiol Lett 317 (2011) 160–171 c 2011 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved MICROBIOLOGY LETTERS Downloaded from https://academic.oup.com/femsle/article/317/2/160/623831 by guest on 22 January 2022

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

R E S E A R C H L E T T E R
Characterizationof swarmingmotility inCitrobacter freundiiYanguang Cong1, Jing Wang1, Zhijin Chen1, Kun Xiong1, Qiwang Xu2 & Fuquan Hu1
1Department of Microbiology, Third Military Medical University, Chongqing, China; and 2Center of Biological Wave, Third Military Medical University,
Chongqing, China
Correspondence: Fuquan Hu, Department
of Microbiology, Third Military Medical
University, Chongqing 400038, China.
Tel./fax: 186 23 6875 2834; e-mail:
Received 23 September 2010; revised 30
December 2010; accepted 18 January 2011.
Final version published online 1 March 2011.
DOI:10.1111/j.1574-6968.2011.02225.x
Editor: Reggie Lo
Keywords
Citrobacter freundii; swarming; motility.
Abstract
Bacterial swarming motility is a flagella-dependent translocation on the surface
environment. It has received extensive attention as a population behavior invol-
ving numerous genes. Here, we report that Citrobacter freundii, an opportunistic
pathogen, exhibits swarming movement on a solid medium surface with appro-
priate agar concentration. The swarming behavior of C. freundii was described in
detail. Insertional mutagenesis with transposon Mini-Tn5 was carried out to
discover genetic determinants related to the swarming of C. freundii. A number of
swarming genes were identified, among which flhD, motA, motB, wzx, rfaL, rfaJ,
rfbX, rfaG, rcsD, rcsC, gshB, fabF, dam, pgi, and rssB have been characterized
previously in other species. In mutants related to lipopolysaccharide synthesis and
RcsCDB signal system, a propensity to form poorly motile bacterial aggregates on
the agar surface was observed. The aggregates hampered bacterial surface migra-
tion. In several mutants, the insertion sites were identified to be in the ORF of
yqhC, yeeZ, CKO_03941, glgC, and ttrA, which have never been shown to be
involved in swarming. Our results revealed several novel characteristics of
swarming motility in C. freundii which are worthy of further study.
Introduction
Bacterial swarming is a flagella-dependent surface transloca-
tion exhibited by a wide variety of flagellated bacteria (for a
review, see Allison & Hughes, 1991; Fraser & Hughes, 1999;
Harshey, 2003; Kaiser, 2007; Kearns, 2010). This form of
locomotion, which was identified as forming a typical
swarm colony on a solid media surface characterized by
circular symmetry and regularly spaced concentric terraces,
was first described by Hauser (1885) in Proteus mirabilis
over a century ago. Both swimming and swarming motilities
depend on bacterial flagella, but they differ in many ways.
The most noticeable distinction is that swimming is an
individual behavior, whereas swarming is a movement of
bacterial populations. Moreover, the cells exhibit differen-
tiation during swarming; they are usually elongated and
hyperflagellated compared with the vegetative cells grown in
liquid media (Allison & Hughes, 1991; Harshey, 2003;
Rather, 2005). Swarming also shares features with other
surface phenomena, such as biofilm formation and host
invasion, and is associated with pathogenesis in some
organisms. For example, swarming of P. mirabilis facilitates
ascending colonization of the urinary tract and is conducive
to biofilm formation on catheters (Allison et al., 1994;
Stickler et al., 1998). Expression of flagella and virulence
factors are coordinated in P. mirabilis and Serratia liquefa-
ciens (Allison et al., 1992; Givskov et al., 1995). The flagellar
export apparatus of Yersinia enterocolitica also functions as a
secretion system for the transport of a virulence-associated
phospholipase (Young et al., 1999). In many species, swarm-
ing bacteria exhibit adaptive resistance to multiple antibio-
tics (Butler et al., 2010).
In recent years, system-screening studies in various
species have revealed numerous swarming-related genes.
These genes are involved in flagellar assembly, synthesis
of polysaccharides, chemosensors, signal regulation, and
metabolic pathways, whereas others are hypothetical
genes with unknown functions (Kearns et al., 2004; Inoue
et al., 2007; Overhage et al., 2007). However, the genetic
determinants for this special process vary among species,
indicating different swarming patterns in various swarming
bacteria. Therefore, the study of swarming motility in
various bacteria would facilitate a thorough understanding
of this special bacterial motion. Considering that many
types of genes are related to swarming motility, such a study
also provides a tractable model to study the function of
FEMS Microbiol Lett 317 (2011) 160–171c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
MIC
ROBI
OLO
GY
LET
TER
SD
ownloaded from
https://academic.oup.com
/femsle/article/317/2/160/623831 by guest on 22 January 2022

genes involved in bacterial differentiation, multicellularity,
and pathogenesis.
Citrobacter freundii is a motile gram-negative bacterium
living in soil and aqueous environments; it is often isolated
in clinical specimens as an opportunistic pathogen. In this
study, we demonstrated that swarming motility could be
induced in C. freundii. It was examined in detail because
little is known about this motility in C. freundii. To discover
the genetic determinants that affect swarming, the mini-Tn5
transposon mutation was used to screen swarming-
associated genes by impairing bacterial swarming ability.
Our results showed that a number of genes are related to the
swarming of C. freundii, among which several have been
newly identified.
Materials and methods
Bacterial strains and media
The following strains were used in this study: C. freundii
ATCC8090 was a gift from Dr Tomofusa Tsuchiya of
Okayama University, Japan; P. mirabilis CMCC49003 was
purchased from the China Medical Culture Collection
Center; and Escherichia coli S17-1(lpir)/pUT mini-Tn5-Km
was a gift from Dr Victor de Lorenzo of the Centro Nacional
de Biotecnologia CSIC, Spain.
M9 salts medium supplemented with 0.5% glucose was
used as the minimal medium. The swarm medium con-
tained 10 g tryptone, 10 g NaCl, and 5 g of glucose L�1 the
final agar concentration was 0.5%. The swim medium
contained the same constituents solidified with 0.3% agar.
To better visualize swarming colonies, a vital dye, triphenyl
tetrazolium chloride (TTC), was added to achieve a final
concentration of 0.05% when required. Both swim and
swarm plates were allowed to dry overnight at room temper-
ature before use. Antibiotics were added, when appropriate,
at the following concentrations: kanamycin at 100 mg mL�1
and rifampicin at 100 mg mL�1.
Light microscopy
To observe swarming motility, 1 mL culture incubated for
10 h in lysogeny broth (LB) (adjusted to 0.5 OD600 nm) was
inoculated onto a thin layer of solid swarm media in a Petri
dish (6 mL media per plate). The plates were directly
observed at � 400 magnification under an Olympus in-
verted microscope IX71 in a room heated to 30 1C. Sterile
slides were occasionally used instead of Petri dishes to
achieve better visualization. The slides were submerged in
swarm media, which was solidified with 0.5% agar, to obtain
a thin layer of media on the surface and dried at 37 1C briefly
before use. After inoculation, the bacteria on the surface of
the media were observed under the inverted microscope.
Images were recorded using a video camera.
Electron microscopy
For negative staining, formvar-coated TEM grids (copper,
75 mesh) were floated on a drop of bacterial cells suspended
in phosphate-buffered saline (PBS, pH 7.4) for 5 min to
allow the adhesion of bacterial cells. The grids were stained
for 5 min using 2% phosphotungstate. After staining, these
were rinsed with water and then air dried. For ultrathin
sectioning, bacteria were washed and suspended in PBS,
fixed in 0.2% v/v glutaraldehyde, and embedded in Spurr
resin. The specimens were examined with a transmission
electron microscope (Philips Tecnai 10).
Mutagenesis with mini-Tn5 and isolation ofmutants defective in swarming
Mutagenesis was performed according to the method de-
scribed by de Lorenzo et al. (1990). Citrobacter freundii and
E. coli S17-1 (lpir)/pUT mini-Tn5-Km were grown over-
night in LB media with rifampicin and kanamycin, respec-
tively. A 100-mL aliquot of each culture was mixed in 5 mL of
10 mM MgSO4 and filtered through a 0.45-mm cellulose
membrane filter. The filter was then placed on the surface of
an LB plate and incubated at 37 1C for 10 h. The bacteria on
the filter surface were washed and suspended in 2 mL of
10 mM MgSO4. About 100 mL of the resulting bacterial
suspensions was spread onto LB plates containing kanamy-
cin and rifampicin and incubated at 37 1C for �36 h. The
antibiotic-resistant bacteria were then transferred to swarm
agar plates and incubated at 37 1C for 12 h. All swarming-
defective colonies were selected.
Analysis of swarming defects
All swarming-defective strains were screened on swim media
to identify the mutants possessing functional flagella but
unable to swarm. Defects in flagellar function were identi-
fied by the absence of outward migration on the media; this
was then confirmed by direct observation under a light
microscope.
Identification of mutant genes
An inverse PCR method was used to amplify the sequence
flanking the inserted mini-Tn5 transposon in the chromo-
some of the swarming-defective mutants. Genomic DNA from
each mutant was isolated according to the cetyltrimethylam-
monium bromide protocol and completely digested with TaqI.
The DNA fragments were self-ligated with T4 DNA ligase and
then used as templates for inverse PCR with the primers P904
(50-GGAGAGGCTATTCGGCTATG-30) and P194c (50-GTAA
GGTGATCCGGTGGATG-30), which were designed according
to the motile sequence of Mini-Tn5-Km plasmid. The PCR
products were separated by agarose gel electrophoresis
and then purified using a gel extraction kit (Watson
FEMS Microbiol Lett 317 (2011) 160–171 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
161Swarming motility in C. freundii
Dow
nloaded from https://academ
ic.oup.com/fem
sle/article/317/2/160/623831 by guest on 22 January 2022

Biotechnologies Inc.). The PCR products were directly
sequenced at Shanghai GeneCore BioTechnologies. If sequen-
cing failed, the PCR products were ligated to PMD18-T vector
(Takara Co. Ltd, Dalian, China) and the sequencing was
attempted again. To identify the mutant genes, nucleotide
sequence databases were searched with the BLASTN and BLASTX
programs developed by the National Center for Biotechnology
Information (NCBI).
Isolation of flagellin
Flagellin was isolated from bacterial cells according to the
method described by DePamphilis & Adler (1971). The
bacterial cells were suspended in 0.1 M Tris-HCl buffer
(pH 7.5). Flagellar filaments were sheared with a tissue
homogenizer at maximum speed for 30 s. The bacteria were
observed microscopically to ascertain loss of motility. Cell
debris was removed from the flagella by centrifugation at
15 000 g for 15 min, and flagellar filaments were then pelleted
from the supernate by ultracentrifugation and then suspended
in 0.1 M Tris-HCl buffer (pH 7.5). Sodium dodecyl sulfate-
polyacrylamide gel electrophoresis (SDS-PAGE) was used to
analyze the purity of samples.
Preparation of flagellin antiserum
Flagellin protein dissolved in Tris-HCl buffer was emulsified
in Freund’s incomplete adjuvant (1 : 3). One rabbit was
immunized three times at intervals of 2 weeks. Serum was
collected 1 week after the final injection and stored at � 20 1C.
Analysis of flagellin production byWestern blotting
Bacteria were suspended in Tris-HCl buffer and then adjusted
to 1 OD600 nm. All the cell lysates were subjected to SDS-PAGE
electrophoresis and then transferred onto a nitrocellulose
membrane. The flagellin was visualized via Western blotting
with enhanced chemiluminescence detection (Pierce). Rabbit
polyclonal antiflagellin serum was used as the primary anti-
body. The secondary antibody was a goat anti-rabbit immuno-
globulin G conjugated with horseradish peroxidase. Detection
was performed according to the protocol of the supplier.
Hydrophilicity measurement
The surface hydrophilicity of bacterial cells was quantified
using bacterial adherence to hydrocarbon (BATH) test, ori-
ginally described by Rosenberg et al. (1980). Bacterial cells
suspended in a BATH buffer (adjusted to 1 OD, 4 mL) were
vortexed with xylene (1 mL) for 60 s. After the phases were
allowed to separate, the aqueous phase was carefully removed
and the A600 nm was measured. The results were expressed as
the percentage in OD of the aqueous phase compared with the
OD of the cell suspension without xylene.
Fluorescence staining with acridine orange
Bacterial smears were fixed with methanol and then stained
using 0.01% acridine orange in 0.05 M PBS (pH 4.8) for
5 min. The samples were viewed at � 1000 magnification
with an Olympus BX51 microscope.
Results and discussion
Differentiation from vegetativecells to swarming cells
When grown in liquid media, C. freundii cells were 0.5–2.0-
mm-long rods (mean value is 1.74� 0.18; 10 cells were
observed) with one to two polar or lateral flagella (mean
value is 1.6� 0.5; 10 cells were observed). When inoculated
onto a solid media surface, usually after 3–4 h bacterial cells
underwent a change in both shape and flagellar production.
They became hyperflagellated (mean value is 13.7� 3.5,
Po 0.05; 10 cells were observed) and slightly elongated (mean
value is 4.55� 0.79, Po 0.05; 10 cells were observed) (Fig. 1a
and b). They also displayed a special form of translocation, i.e.
swarming, on the media with appropriate agar concentration.
Citrobacter freundii cells exhibited swarming motility optimally
on 0.5–0.7% agar and not on agar with concentrations over
1%. On these high concentration agars, the decreased water
content inhibited the bacterial motility. When inoculated on
0.5% agar surface, after 3–4 h of stationary phase, bacterial cells
differentiated into swarming cells and then moved rapidly and
colonized the entire surface in 6–8 h with an expansion rate of
0.44–0.58 cm h�1 (Fig. 1c). The flagellin of C. freundii isolated
from swarming cells grown on swarming media and from
vegetative cells grown in liquid media possess the same
molecular mass (�47.5 kDa) based on their respective migra-
tion distances in SDS-PAGE electrophoresis (Fig. 2a).
Effect of nutrient composition on theswarming motility of C. freundii
Besides agar concentration, nutrient composition in the
medium served as another critical factor affecting swarming
motility. Citrobacter freundii cells were unable to swarm on
the M9 minimal media, although they had grown well and
displayed normal swimming motility in M9 liquid media.
Swarming requires the presence of certain inducers in the
swarm agar plates. Usually, casamino acids satisfy the
requirement for swarming. Proteus mirabilis and Pseudomo-
nas aeruginosa have been shown to respond to single amino
acids as inducers of swarming motility (Allison et al., 1993;
Kohler et al., 2000). However, in this study, C. freundii did
not swarm on the minimal media M9 supplemented with
either each of 20 amino acids or a mixture of amino acids
(casamino acids) until tryptone or peptone was added into
FEMS Microbiol Lett 317 (2011) 160–171c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
162 Y. Cong et al.
Dow
nloaded from https://academ
ic.oup.com/fem
sle/article/317/2/160/623831 by guest on 22 January 2022
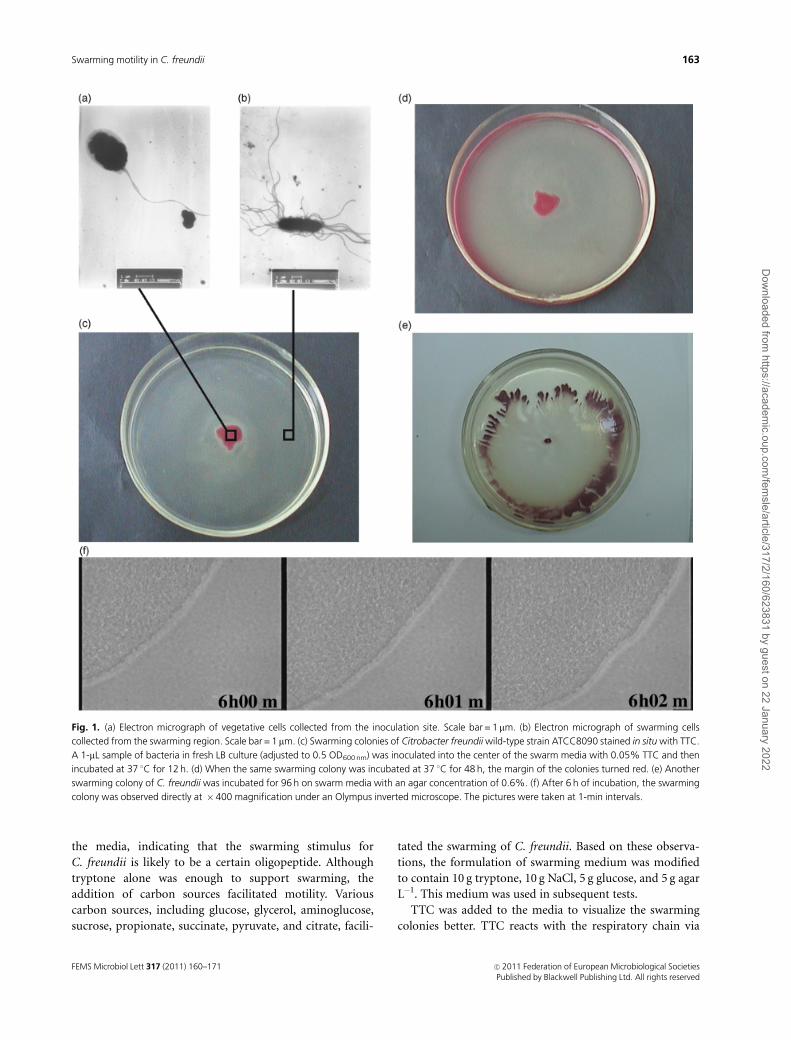
the media, indicating that the swarming stimulus for
C. freundii is likely to be a certain oligopeptide. Although
tryptone alone was enough to support swarming, the
addition of carbon sources facilitated motility. Various
carbon sources, including glucose, glycerol, aminoglucose,
sucrose, propionate, succinate, pyruvate, and citrate, facili-
tated the swarming of C. freundii. Based on these observa-
tions, the formulation of swarming medium was modified
to contain 10 g tryptone, 10 g NaCl, 5 g glucose, and 5 g agar
L�1. This medium was used in subsequent tests.
TTC was added to the media to visualize the swarming
colonies better. TTC reacts with the respiratory chain via
Fig. 1. (a) Electron micrograph of vegetative cells collected from the inoculation site. Scale bar = 1mm. (b) Electron micrograph of swarming cells
collected from the swarming region. Scale bar = 1 mm. (c) Swarming colonies of Citrobacter freundii wild-type strain ATCC8090 stained in situ with TTC.
A 1-mL sample of bacteria in fresh LB culture (adjusted to 0.5 OD600 nm) was inoculated into the center of the swarm media with 0.05% TTC and then
incubated at 37 1C for 12 h. (d) When the same swarming colony was incubated at 37 1C for 48 h, the margin of the colonies turned red. (e) Another
swarming colony of C. freundii was incubated for 96 h on swarm media with an agar concentration of 0.6%. (f) After 6 h of incubation, the swarming
colony was observed directly at �400 magnification under an Olympus inverted microscope. The pictures were taken at 1-min intervals.
FEMS Microbiol Lett 317 (2011) 160–171 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
163Swarming motility in C. freundii
Dow
nloaded from https://academ
ic.oup.com/fem
sle/article/317/2/160/623831 by guest on 22 January 2022

cytochrome oxidase and is reduced to formazan, an insolu-
ble red pigment, in the cells (Boker-Schmitt et al., 1982).
Because it stains cells in situ, TTC is commonly used to aid
in the examination of bacterial colonies (Parrington et al.,
1993; Semmler et al., 1999). Figure 1c–e shows that TTC
stains swarming colonies in situ and discriminates the
different regions composed of swarming and vegetative cells,
as reported in a previous study (Falkinham & Hoffman,
1984). Bacteria in the red zones (inoculation sites) consisted
of vegetative cells with normal morphologies and rare
flagella (Fig. 1a), whereas those in the lightly staining zones
consisted of swarming cells with elongated bodies and dense
flagella (Fig. 1b). A similar phenomenon was observed in P.
mirabilis and alternate color cycles of red (consolidation)
and lightly stained (swarming) areas were visible in the
stained colonies (Supporting Information, Fig. S5).
It is evident from the color alteration of the bacterial
colonies that the vegetative cells in red zones had a high
aerobic respiration rate and might have obtained energy
mainly from the tricarboxylic acid (TCA) cycle. In compar-
ison, the swarming cells in the light zones had a low aerobic
respiration rate and perhaps primarily obtained their energy
from sugar fermentation. This assumption is supported by a
previous study. In E. coli, three components of the TCA cycle
and aerobic respiration, sdhCDAB (succinate dehydrogen-
ase), cyoABCDE (cytochrome o ubiquinol oxidase), and gltA
(citrate synthase), were demonstrated to be downregulated
by the transcriptional regulatory complex FlhD/FlhC, a
global regulator involved in many cellular processes (Pruss
et al., 2003). High-level FlhD/FlhC is induced in swarming
colonies and then apparently results in deduced expression
of the abovementioned enzymes as well as inhibition of TCA
cycle and respiratory process. The repressed aerobic respira-
tion in swarming cells could explain the staining character-
istics of the swarming colonies observed in this study. Our
results indicated that TTC is a suitable dye for staining
swarming colonies that are difficult to distinguish.
Microscopic features of swarmingmotility in C. freundii
As observed under the inverted microscope, the swarming
colonies of C. freundii consisted of one tier of cells on the agar
surface (Fig. 1f and Video S1). Swarming cells seemed to form a
wet environment on the agar surface, which likely provided
enough space for the bacteria to rotate their flagella. Single
bacteria were not found moving on the surface of the media,
although these bacteria certainly possessed the same functional
flagella. The moist milieu of swarming colonies could be due to
bacterial polysaccharide structures that extract and contain
water from the media. The significant role of the polysacchar-
ide structure on swarming has been revealed in previous studies
(Toguchi et al., 2000; Inoue et al., 2007). To support this point,
swarming cells of C. freundii were observed to be more
hydrophilic compared with vegetative cells (0.961 for swarming
cells; 0.814 for vegetative cells, Po 0.05; Fig. S2) in this work.
In swarming colonies, C. freundii cells moved actively.
However, the cells at the periphery of colonies were less active
due to the decreased moisture capacity in areas where the cells
moved out and back occasionally, or were pushed by the
actively moving cells in the central region. As a result, the edge
of colonies expanded outward continuously. Citrobacter
freundii did not display alternating cycles of swarming and
consolidation during the development of swarming colonies
as in P. mirabilis. Once differentiation occurred at the
inoculation site, swarming cells spread continuously, until
they occupied the entire agar surface. Even when inoculated
onto a large plate with a diameter over 20 cm, alternating
cycles of swarming and consolidation were not observed.
Transposon mutagenesis and screening ofmutants with defective swarming motility
Transposon mutagenesis involving the use of Mini-Tn5 on a
suicide plasmid pUT was carried out to further understand
the genetic determinants of swarming motility in C. freundii.
Fig. 2. (a) SDS-PAGE of purified flagellin samples
isolated from Citrobacter freundii wild-type strain
ATCC8090 incubated in two different media. 1,
Incubated in broth; 2, incubated in swarm medium
with 0.5% agar; M, molecular size marker. (b)
Flagellin expression of different mutants incubated
in swarm medium with 0.5% agar. Aliquots
of OD600nm-equalized cells were analyzed by
Western blotting using antiflagellin antibodies
as described in Materials and methods.
FEMS Microbiol Lett 317 (2011) 160–171c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
164 Y. Cong et al.
Dow
nloaded from https://academ
ic.oup.com/fem
sle/article/317/2/160/623831 by guest on 22 January 2022

A total of 85 swarming-defective mutants were screened
from approximately 6000 transconjugants; of the 85 mu-
tants, 53 were defective in both swimming and swarming.
The remaining 32 mutants were defective in swarming but
not swimming. The mutants with normal swimming pat-
tern were subjected to further sequence analysis to deter-
mine the insertionally mutated gene. Given that swarming is
dependent on functional flagella, as demonstrated in pre-
vious studies, of the 53 swimming-defective mutants, only
five randomly selected mutants were further subjected to
sequence analysis. As a whole, sequences produced valid
results with only four exceptions (CF407, CF415, CF701,
and CF711). In most cases, the most similar genes obtained
through the homology searches usually belonged to Citro-
bacter koseri ATCC BAA-895, a species of Citrobacter with
complete genome sequence information. The results of the
homology searches are listed in Tables 1 and 2 and are also
described in the following two sections on genes that have
been previously characterized in other species and those first
identified in this study.
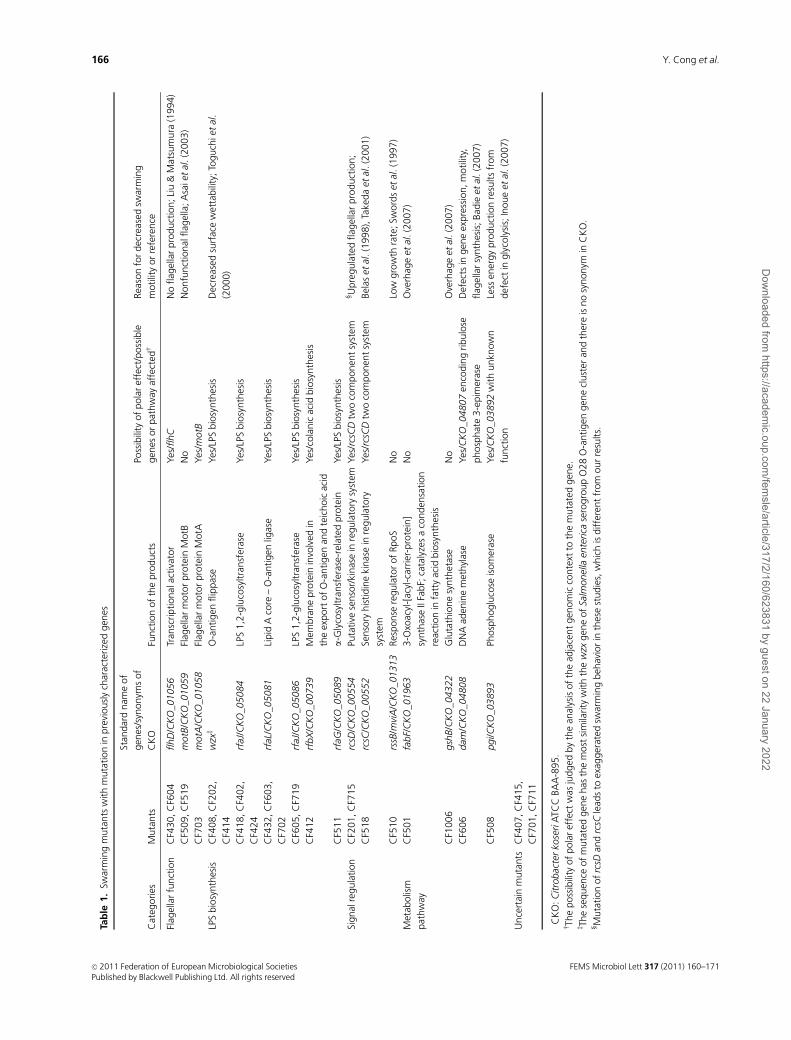
Previously characterized genes
As many as 16 swarming-related genes identified in our
study have already been characterized previously in other
species. The underlying causes for the defective swarming
motility of the mutants are listed in Table 1. However, some
of them are worthy of further discussion.
Swarming motility of C. freundii is dependenton flagella
As expected, flhD, motA, and motB mutants were identified
among the five mutants found to be defective in both
swarming and swimming motilities. Because the flhDC
operon controls flagellar biosynthesis (Liu & Matsumura,
1994), no flagellum was produced in the flhD mutants (Fig.
2b). On the other hand, although the motA and motB
mutants produced flagella, they were still unable to move
because MotA and MotB formed a proton channel that
transferred proton-motive force to drive the flagella (Asai
et al., 2003); either motA or motB gene mutations resulted in
the production of nonfunctional flagella (Figs 2b and 3c).
These data demonstrate that the swarming of C. freundii is
dependent on functional flagella, as in other swarming
bacteria (Kearns, 2010).
Genes related to lipopolysaccharide synthesis
The largest gene cluster identified in our study is involved in
the synthesis of lipopolysaccharide. Altogether, 13 mutants
were isolated, of which six mutated genes – wzx, rfaL, rfbX,
rfaJ/CKO_05084, rfaJ/CKO_05086, and rfaG – were identi-
fied. The swarming ability of these mutants was dramatically
decreased (two of them are shown in Fig. 3g and h as
examples). As observed directly under inverted microscope,
only a few bacterial cells were actively motile in the swarm-
ing colonies of these mutants and these were mainly
distributed at the edges. In the central region, most cells
formed aggregates that scarcely moved (Videos S2 and S3).
In contrast, in wild-type colonies, all swarming cells were
actively motile (cells in the edge of colonies were less active)
and no aggregation was observed (Video S1).
The hydrophilicity of these mutants was decreased com-
pared with the wild type (Fig. S2), which could have led to
the aggregation. In a previous study, many transposon
swarming mutants isolated in Salmonella enterica serovar
Typhimurium have been shown to have mutations in the
lipopolysaccharide biosynthetic pathway (Toguchi et al.,
2000). The authors suggested that the O antigen directly or
indirectly improved the surface wettability required for
swarm colony expansion. Our observation showed that the
polysaccharide structure on the cell surface had important
role not only in overcoming friction between bacterial cells
and media surface, but also in reducing intercellular inter-
action. The poorly motile aggregates formed with bacteria
on the agar surface because of the O antigen defects could
account for the defective swarming in addition to the
decreased wettability of the agar surface.
The rcsC-rcsD-rcsB system
rcsC and rcsD mutants were identified in this study, and both
mutants displayed defective swarming behavior (Fig. 3a and
b). The products of rcsC and rcsD, together with RcsB,
constitute the regulator of the capsule synthesis (Rcs)
phosphorelay system. The regulator RcsB is activated by the
transfer of a phosphate group from its cognate sensor, RcsC,
through a histidine-containing phosphotransmitter (Hpt)
domain intermediate called RcsD (previously called YojN;
Takeda et al., 2001). The Rcs system has been implicated in
the regulation of bacterial responses to osmotic and other
kinds of membrane stress, growth at low temperatures in the
presence of glucose and zinc, and growth on solid surfaces
(Carballes et al., 1999; Majdalani & Gottesman, 2005; Wang
et al., 2007). Mutations in rcsC and rcsD affect the temporal
regulation of swarming motility and result in precocious
behavior in E. coli and P. mirabilis (Belas et al., 1998; Takeda
et al., 2001). Francez-Charlot et al. (2003) have shown that
the RcsCDB system negatively regulates the flhDC operon in
E. coli and that the exaggerated swarming behavior of the
rcsC and rcsD (yojN) mutants is probably the consequence of
the higher basal expression of the flhDC operon in the rcs
mutants, leading to a higher expression of genes, including
those required for the synthesis of flagellin. In contrast, in
our study, the colonies of C. freundii rcsC and rcsD mutants
FEMS Microbiol Lett 317 (2011) 160–171 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
165Swarming motility in C. freundii
Dow
nloaded from https://academ
ic.oup.com/fem
sle/article/317/2/160/623831 by guest on 22 January 2022
Tab
le1.
Swar
min
gm
uta
nts
with
muta
tion
inpre
viousl
ych
arac
terize
dgen
es
Cat
egories
Muta
nts
Stan
dar
dnam
eof
gen
es/s
ynonym
sof
CK
O�
Funct
ion
of
the
pro
duct
s
Poss
ibili
tyof
pola
ref
fect
/poss
ible
gen
esor
pat
hw
ayaf
fect
edw
Rea
son
for
dec
reas
edsw
arm
ing
motilit
yor
refe
rence
Flag
ella
rfu
nct
ion
CF4
30,C
F604
flhD
/CK
O_0
1056
Tran
scriptional
activa
tor
Yes
/flhC
No
flag
ella
rpro
duct
ion;Li
u&
Mat
sum
ura
(1994)
CF5
09,C
F519
motB
/CK
O_0
1059
Flag
ella
rm
oto
rpro
tein
MotB
No
Nonfu
nct
ional
flag
ella
;A
saie
tal
.(2
003)
CF7
03
motA
/CK
O_0
1058
Flag
ella
rm
oto
rpro
tein
MotA
Yes
/motB
LPS
bio
synth
esis
CF4
08,C
F202,
CF4
14
wzxz
O-a
ntigen
flip
pas
eYes
/LPS
bio
synth
esis
Dec
reas
edsu
rfac
ew
etta
bili
ty;To
guch
iet
al.
(2000)
CF4
18,C
F402,
CF4
24
rfaJ
/CK
O_0
5084
LPS
1,2
-glu
cosy
ltra
nsf
eras
eYes
/LPS
bio
synth
esis
CF4
32,C
F603,
CF7
02
rfaL
/CK
O_0
5081
Lipid
Aco
re–
O-a
ntigen
ligas
eYes
/LPS
bio
synth
esis
CF6
05,C
F719
rfaJ
/CK
O_0
5086
LPS
1,2
-glu
cosy
ltra
nsf
eras
eYes
/LPS
bio
synth
esis
CF4
12
rfbX
/CK
O_0
0739
Mem
bra
ne
pro
tein
invo
lved
in
the
export
of
O-a
ntigen
and
teic
hoic
acid
Yes
/cola
nic
acid
bio
synth
esis
CF5
11
rfaG
/CK
O_0
5089
a-G
lyco
syltra
nsf
eras
e-re
late
dpro
tein
Yes
/LPS
bio
synth
esis
Signal
regula
tion
CF2
01,C
F715
rcsD
/CK
O_0
0554
Puta
tive
senso
r/ki
nas
ein
regula
tory
syst
emYes
/rcs
CD
two
com
ponen
tsy
stem
‰U
pre
gula
ted
flag
ella
rpro
duct
ion;
Bel
aset
al.(1
998),
Take
da
etal
.(2
001)
CF5
18
rcsC
/CK
O_0
0552
Senso
ryhis
tidin
eki
nas
ein
regula
tory
syst
em
Yes
/rcs
CD
two
com
ponen
tsy
stem
CF5
10
rssB
/mvi
A/C
KO
_01313
Res
ponse
regula
tor
of
RpoS
No
Low
gro
wth
rate
;Sw
ord
set
al.(1
997)
Met
abolis
m
pat
hw
ay
CF5
01
fabF/
CK
O_0
1963
3-O
xoac
yl-[
acyl
-car
rier
-pro
tein
]
synth
ase
IIFa
bF;
cata
lyze
sa
conden
sation
reac
tion
infa
tty
acid
bio
synth
esis
No
Ove
rhag
eet
al.(2
007)
CF1
006
gsh
B/C
KO
_04322
Glu
tath
ione
synth
etas
eN
oO
verh
age
etal
.(2
007)
CF6
06
dam
/CK
O_0
4808
DN
Aad
enin
em
ethyl
ase
Yes
/CK
O_0
4807
enco
din
gribulo
se
phosp
hat
e3-e
pim
eras
e
Def
ects
ingen
eex
pre
ssio
n,m
otilit
y,
flag
ella
rsy
nth
esis
;Bad
ieet
al.(2
007)
CF5
08
pgi/C
KO
_03893
Phosp
hoglu
cose
isom
eras
eYes
/CK
O_0
3892
with
unkn
ow
n
funct
ion
Less
ener
gy
pro
duct
ion
resu
lts
from
def
ect
ingly
coly
sis;
Inoue
etal
.(2
007)
Unce
rtai
nm
uta
nts
CF4
07,C
F415,
CF7
01,C
F711
� CK
O:C
itro
bac
ter
kose
riA
TCC
BA
A-8
95.
w The
poss
ibili
tyof
pola
ref
fect
was
judged
by
the
anal
ysis
of
the
adja
cent
gen
om
icco
nte
xtto
the
muta
ted
gen
e.z T
he
sequen
ceof
muta
ted
gen
ehas
the
most
sim
ilarity
with
the
wzx
gen
eof
Salm
onel
laen
terica
sero
gro
up
O28
O-a
ntigen
gen
ecl
ust
eran
dth
ere
isno
synonym
inC
KO
.‰M
uta
tion
of
rcsD
and
rcsC
lead
sto
exag
ger
ated
swar
min
gbeh
avio
rin
thes
est
udie
s,w
hic
his
diffe
rent
from
our
resu
lts.
FEMS Microbiol Lett 317 (2011) 160–171c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
166 Y. Cong et al.
Dow
nloaded from https://academ
ic.oup.com/fem
sle/article/317/2/160/623831 by guest on 22 January 2022

were not precocious (Fig. 3a and b). As observed directly
under the inverted microscope, similar to those lipopoly-
saccharide mutants, mutants of rcsD and rcsC formed
aggregates in the swarming colonies (Video S4). As the
regulator of capsule synthesis, mutations in RcsD and RcsC
certainly lead to a decrease in bacterial surface hydrophili-
city, which was supported by our results of BATH measure-
ment (Fig. S2).
New swarming genes identified in our study
Aside from the previously characterized genes, several new
swarming-related genes were identified in the present study.
A novel yqhC gene
Four mutants were identified as having yqhC gene muta-
tions that formed small colonies on the swarm plate (Fig.
3d). The product of the yqhC gene is a putative AraC-family
transcriptional regulatory protein, as annotated in the
NCBI. Most members of the AraC-XylS proteins are positive
transcriptional regulators involved in the control of many
important processes related to carbon metabolism, stress
responses, and pathogenesis (Egan, 2002). The flagellar
production of yqhC mutant was comparable to that of the
wild type (Fig. 2b), suggesting that the decrease of swarm
ability of yqhC mutant was not due to a disruption of
flagellar synthesis. However, the high output of yqhC
mutants in our study indicates a close relationship between
swarming motility and the function of the yqhC gene. In a
recent study, yqhC gene in E. coli has been shown to regulate
the transcription of the adjacent genes, yqhD and dkgA, that
encode NADPH-dependent oxidoreductases with broad-
substrate ranges that include furfural and methylglyoxal
(Perez et al., 2008; Turner et al., 2010). As 0.5% glucose was
added into the swarm media, the concentration of aldehydes
was inevitably increased in bacterial cells of the yqhC mutant
due to the lack of expression of the yqhD and dkgA genes.
The high concentration of aldehydes was harmful to the
bacterial cells and may have interrupted swarming in some
unknown ways.
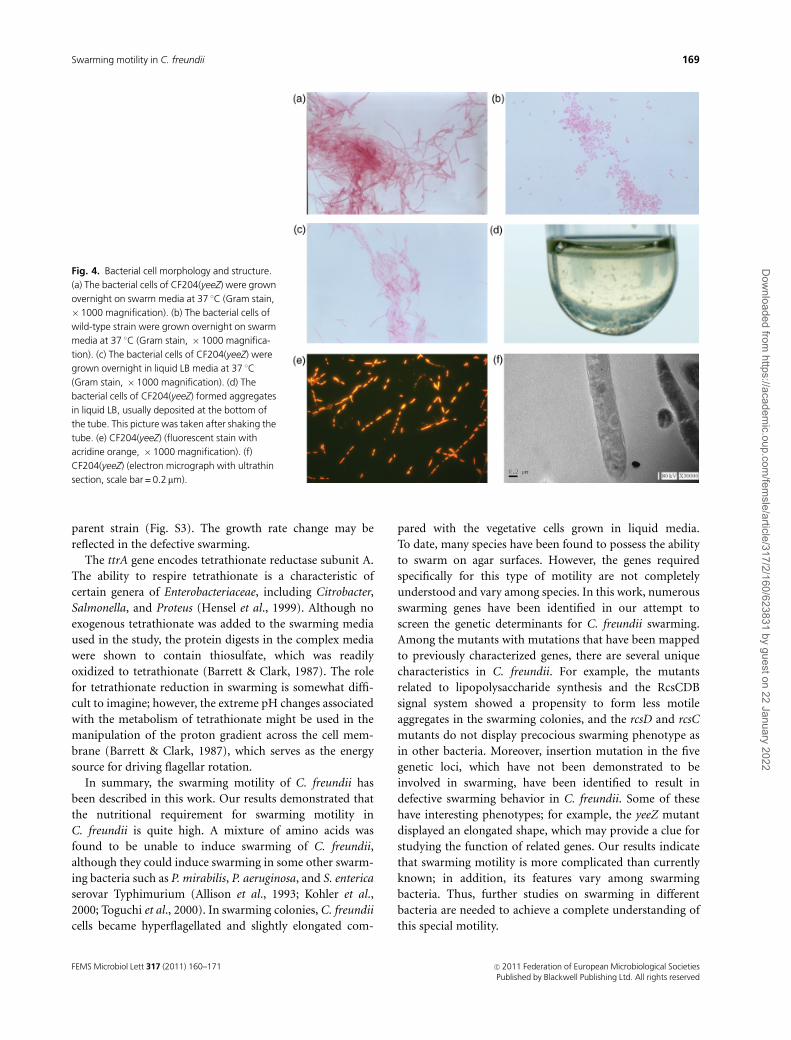
A novel yeeZ gene
Among the mutants, the yeeZ mutant was notable because it
displayed an elongated shape whether grown in liquid media
or on the surface of the solid plate (Fig. 4a–c). In liquid
media, elongated bacteria formed aggregates that were even
deposited at the bottom of the tube (Fig. 4d). The product
of yeeZ is a putative nucleoside-diphosphate-sugar epimer-
ase that has a possible role in polysaccharide synthesis.
However, no direct functional studies of yeeZ have been
undertaken until now. Based on the results of the fluores-
cence staining with acridine orange, the bacterial cells of
yeeZ mutant were multinucleate (Fig. 4e), and the bacterial
cell walls were intact, as revealed by electron microscopy
(Fig. 4f). Hydrophilicity of the mutant decreased compared
with the wild type (Fig. S2) and the insertional mutation in
yeeZ gene also resulted in dramatic low growth rate (Fig.
S3). These features suggest that the function of yeeZ gene
may be associated with bacterial cell division. However, the
downstream gene, CKO_00769, which encodes a putative
LysR-type transcriptional regulator, overlaps in sequence
with the yeeZ gene. So the possibility that the novel features
of CF204 may be due to the polar effect of transposon on the
expression of the CKO_00769 must be considered.
In liquid media, the mutant bacteria were motile but less
active than the wild type even though the flagellin level of
the yeeZ mutant was comparable to that of the wild type
(Fig. 2b and Video S5). Cell elongation has been previously
Table 2. Swarming mutants with mutation in previously uncharacterized genes
Mutants
Standard name of genes/
synonyms of CKO� Function of the products
Possibility of polar effect/possible
genes or pathway affectedw
CF426, CF1001,
CF405, CF717
yqhC/CKO_04404 Transcriptional regulator, AraC family K00567 Yes/CKO_04405, CKO_04406(yqhD)
and dkgA
CF204 yeeZ/CKO_00768 Hypothetical protein, nucleoside-diphosphate
sugar epimerases
Yes/CKO_00769
CF520 CKO_03941z Hypothetical protein, oligoketide
cyclase/lipid transport protein
Yes/CKO_03940 with unknown
function
CF1002 ttrA‰ Tetrathionate reductase subunit A No
CF101 glgC/CKO_04849 Glucose-1-phosphate adenylyltransferase;
ADP-glucose pyrophosphorylase
Yes/glycogen synthesis
�CKO: Citrobacter koseri ATCC BAA-895.wThe possibility of polar effect was judged by the analysis of the adjacent genomic context to the mutated gene.zThere is no standard gene name.‰The sequence of mutated gene has the most similarity with the ttrA gene of Salmonella enterica ssp. enterica serovar Agona str. SL483 and there is no
synonym in CKO.
FEMS Microbiol Lett 317 (2011) 160–171 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
167Swarming motility in C. freundii
Dow
nloaded from https://academ
ic.oup.com/fem
sle/article/317/2/160/623831 by guest on 22 January 2022
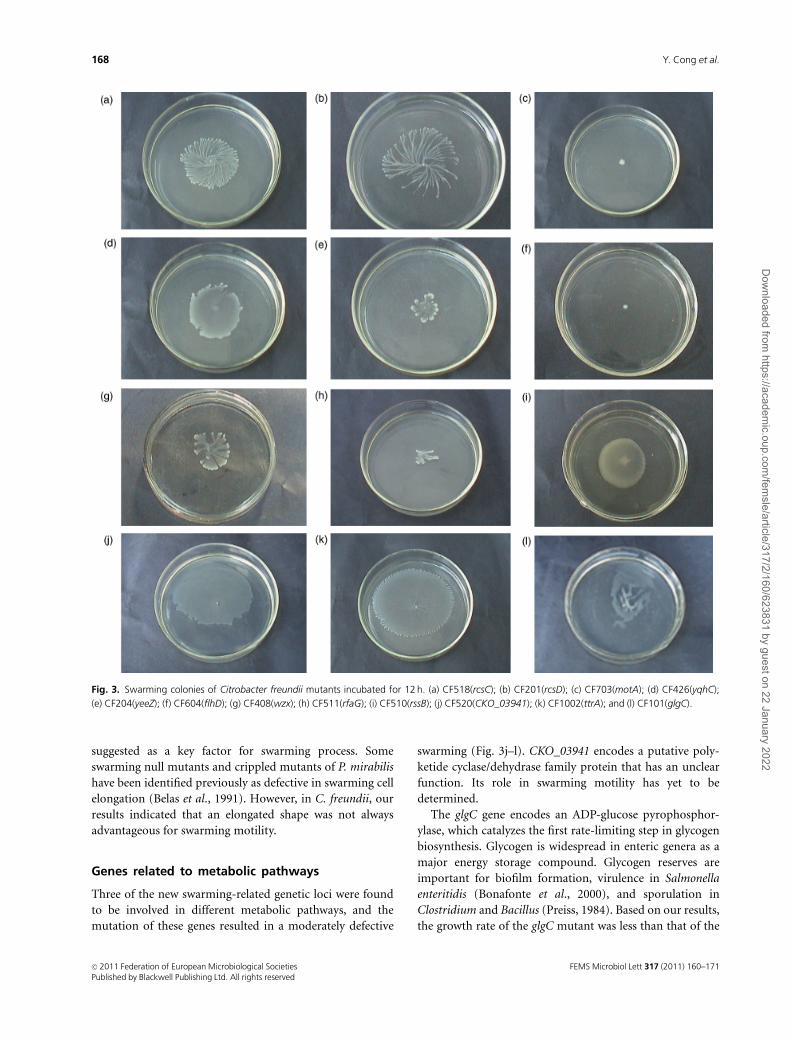
suggested as a key factor for swarming process. Some
swarming null mutants and crippled mutants of P. mirabilis
have been identified previously as defective in swarming cell
elongation (Belas et al., 1991). However, in C. freundii, our
results indicated that an elongated shape was not always
advantageous for swarming motility.
Genes related to metabolic pathways
Three of the new swarming-related genetic loci were found
to be involved in different metabolic pathways, and the
mutation of these genes resulted in a moderately defective
swarming (Fig. 3j–l). CKO_03941 encodes a putative poly-
ketide cyclase/dehydrase family protein that has an unclear
function. Its role in swarming motility has yet to be
determined.
The glgC gene encodes an ADP-glucose pyrophosphor-
ylase, which catalyzes the first rate-limiting step in glycogen
biosynthesis. Glycogen is widespread in enteric genera as a
major energy storage compound. Glycogen reserves are
important for biofilm formation, virulence in Salmonella
enteritidis (Bonafonte et al., 2000), and sporulation in
Clostridium and Bacillus (Preiss, 1984). Based on our results,
the growth rate of the glgC mutant was less than that of the
Fig. 3. Swarming colonies of Citrobacter freundii mutants incubated for 12 h. (a) CF518(rcsC); (b) CF201(rcsD); (c) CF703(motA); (d) CF426(yqhC);
(e) CF204(yeeZ); (f) CF604(flhD); (g) CF408(wzx); (h) CF511(rfaG); (i) CF510(rssB); (j) CF520(CKO_03941); (k) CF1002(ttrA); and (l) CF101(glgC).
FEMS Microbiol Lett 317 (2011) 160–171c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
168 Y. Cong et al.
Dow
nloaded from https://academ
ic.oup.com/fem
sle/article/317/2/160/623831 by guest on 22 January 2022
parent strain (Fig. S3). The growth rate change may be
reflected in the defective swarming.
The ttrA gene encodes tetrathionate reductase subunit A.
The ability to respire tetrathionate is a characteristic of
certain genera of Enterobacteriaceae, including Citrobacter,
Salmonella, and Proteus (Hensel et al., 1999). Although no
exogenous tetrathionate was added to the swarming media
used in the study, the protein digests in the complex media
were shown to contain thiosulfate, which was readily
oxidized to tetrathionate (Barrett & Clark, 1987). The role
for tetrathionate reduction in swarming is somewhat diffi-
cult to imagine; however, the extreme pH changes associated
with the metabolism of tetrathionate might be used in the
manipulation of the proton gradient across the cell mem-
brane (Barrett & Clark, 1987), which serves as the energy
source for driving flagellar rotation.
In summary, the swarming motility of C. freundii has
been described in this work. Our results demonstrated that
the nutritional requirement for swarming motility in
C. freundii is quite high. A mixture of amino acids was
found to be unable to induce swarming of C. freundii,
although they could induce swarming in some other swarm-
ing bacteria such as P. mirabilis, P. aeruginosa, and S. enterica
serovar Typhimurium (Allison et al., 1993; Kohler et al.,
2000; Toguchi et al., 2000). In swarming colonies, C. freundii
cells became hyperflagellated and slightly elongated com-
pared with the vegetative cells grown in liquid media.
To date, many species have been found to possess the ability
to swarm on agar surfaces. However, the genes required
specifically for this type of motility are not completely
understood and vary among species. In this work, numerous
swarming genes have been identified in our attempt to
screen the genetic determinants for C. freundii swarming.
Among the mutants with mutations that have been mapped
to previously characterized genes, there are several unique
characteristics in C. freundii. For example, the mutants
related to lipopolysaccharide synthesis and the RcsCDB
signal system showed a propensity to form less motile
aggregates in the swarming colonies, and the rcsD and rcsC
mutants do not display precocious swarming phenotype as
in other bacteria. Moreover, insertion mutation in the five
genetic loci, which have not been demonstrated to be
involved in swarming, have been identified to result in
defective swarming behavior in C. freundii. Some of these
have interesting phenotypes; for example, the yeeZ mutant
displayed an elongated shape, which may provide a clue for
studying the function of related genes. Our results indicate
that swarming motility is more complicated than currently
known; in addition, its features vary among swarming
bacteria. Thus, further studies on swarming in different
bacteria are needed to achieve a complete understanding of
this special motility.
Fig. 4. Bacterial cell morphology and structure.
(a) The bacterial cells of CF204(yeeZ) were grown
overnight on swarm media at 37 1C (Gram stain,
� 1000 magnification). (b) The bacterial cells of
wild-type strain were grown overnight on swarm
media at 37 1C (Gram stain, � 1000 magnifica-
tion). (c) The bacterial cells of CF204(yeeZ) were
grown overnight in liquid LB media at 37 1C
(Gram stain, � 1000 magnification). (d) The
bacterial cells of CF204(yeeZ) formed aggregates
in liquid LB, usually deposited at the bottom of
the tube. This picture was taken after shaking the
tube. (e) CF204(yeeZ) (fluorescent stain with
acridine orange, �1000 magnification). (f)
CF204(yeeZ) (electron micrograph with ultrathin
section, scale bar = 0.2mm).
FEMS Microbiol Lett 317 (2011) 160–171 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
169Swarming motility in C. freundii
Dow
nloaded from https://academ
ic.oup.com/fem
sle/article/317/2/160/623831 by guest on 22 January 2022

Acknowledgements
We thank Tomofusa Tsuchiya of Okayama University, Japan,
for providing strain C. freundii. We also gratefully acknowl-
edge Victor de Lorenzo of Centro Nacional de Biotecnologia
CSIC, Spain, for providing Mini-Tn5 transposon.
References
Allison C & Hughes C (1991) Bacterial swarming: an example of
prokaryotic differentiation and multicellular behaviour. Sci
Prog 75: 403–422.
Allison C, Lai HC & Hughes C (1992) Co-ordinate expression of
virulence genes during swarm-cell differentiation and
population migration of Proteus mirabilis. Mol Microbiol 6:
1583–1591.
Allison C, Lai HC, Gygi D & Hughes C (1993) Cell differentiation
of Proteus mirabilis is initiated by glutamine, a specific
chemoattractant for swarming cells. Mol Microbiol 8: 53–60.
Allison C, Emody L, Coleman N & Hughes C (1994) The role of
swarm cell differentiation and multicellular migration in the
uropathogenicity of Proteus mirabilis. J Infect Dis 169:
1155–1158.
Asai Y, Yakushi T & Kawagishi I (2003) Ion-coupling
determinants of Na1-driven and H1-driven flagellar motors.
J Mol Biol 327: 453–463.
Badie G, Heithoff DM, Sinsheimer RL & Mahan MJ (2007)
Altered levels of Salmonella DNA adenine methylase are
associated with defects in gene expression, motility, flagellar
synthesis, and bile resistance in the pathogenic strain 14028
but not in the laboratory strain LT2. J Bacteriol 189:
1556–1564.
Barrett EL & Clark MA (1987) Tetrathionate reduction and
production of hydrogen sulfide from thiosulfate. Microbiol Rev
51: 192–205.
Belas R, Erskine D & Flaherty D (1991) Proteus mirabilis mutants
defective in swarmer cell differentiation and multicellular
behavior. J Bacteriol 173: 6279–6288.
Belas R, Schneider R & Melch M (1998) Characterization of
Proteus mirabilis precocious swarming mutants: identification
of rsbA, encoding a regulator of swarming behavior. J Bacteriol
180: 6126–6139.
Boker-Schmitt E, Francisci S & Schweyen RJ (1982) Mutations
releasing mitochondrial biogenesis from glucose repression in
Saccharomyces cerevisiae. J Bacteriol 151: 303–310.
Bonafonte MA, Solano C, Sesma B, Alvarez M, Montuenga L,
Garcia-Ros D & Gamazo C (2000) The relationship between
glycogen synthesis, biofilm formation and virulence in
Salmonella enteritidis. FEMS Microbiol Lett 191: 31–36.
Butler MT, Wang Q & Harshey RM (2010) Cell density and
mobility protect swarming bacteria against antibiotics. P Natl
Acad Sci USA 107: 3776–3781.
Carballes F, Bertrand C, Bouche JP & Cam K (1999) Regulation of
Escherichia coli cell division genes ftsA and ftsZ by the two-
component system rcsC-rcsB. Mol Microbiol 34: 442–450.
de Lorenzo V, Herrero M, Jakubzik U & Timmis KN (1990) Mini-
Tn5 transposon derivatives for insertion mutagenesis
promoter probing, and chromosomal insertion of cloned
DNA in gram-negative eubacteria. J Bacteriol 172: 6568–6572.
DePamphilis ML & Adler J (1971) Fine structure and isolation of
the hook-basal body complex of flagella from Escherichia coli
and Bacillus subtilis. J Bacteriol 105: 384–395.
Egan SM (2002) Growing repertoire of AraC/XylS activators.
J Bacteriol 184: 5529–5532.
Falkinham JO III & Hoffman PS (1984) Unique developmental
characteristics of the swarm and short cells of Proteus vulgaris
and Proteus mirabilis. J Bacteriol 158: 1037–1040.
Francez-Charlot A, Laugel B, Van Gemert A, Dubarry N,
Wiorowski F, Castanie-Cornet MP, Gutierrez C & Cam K
(2003) RcsCDB His-Asp phosphorelay system negatively
regulates the flhDC operon in Escherichia coli. Mol Microbiol
49: 823–832.
Fraser GM & Hughes C (1999) Swarming motility. Curr Opin
Microbiol 2: 630–635.
Givskov M, Eberl L, Christiansen G, Bendik MJ & Molin S (1995)
Induction of phospholipase and flagellar synthesis in Serratia
liquefaciens is controlled by expression of the master operon
flhD. Mol Microbiol 15: 445–454.
Harshey RM (2003) Bacterial motility on a surface: many ways to
a common goal. Annu Rev Microbiol 57: 249–273.
Hauser G (1885) Uber Faulnisbakterien und deren Beziehung zur
Septicamie. F.G.W. Vogel, Leipzig, Germany.
Hensel M, Hinsley AP, Nikolaus T, Sawers G & Berks BC (1999)
The genetic basis of tetrathionate respiration in Salmonella
typhimurium. Mol Microbiol 32: 275–287.
Inoue T, Shingaki R, Hirose S, Waki K, Mori H & Fukui K (2007)
Genome-wide screening of genes required for swarming
motility in Escherichia coli K-12. J Bacteriol 189: 950–957.
Kaiser D (2007) Bacterial swarming: a re-examination of cell-
movement patterns. Curr Biol 17: 561–570.
Kearns DB (2010) A field guide to bacterial swarming motility.
Nat Rev Microbiol 8: 634–644.
Kearns DB, Chu F, Rudner R & Losick R (2004) Genes governing
swarming in Bacillus subtilis and evidence for a phase variation
mechanism controlling surface motility. Mol Microbiol 52:
357–369.
Kohler T, Curty LK & Barja F (2000) Swarming of Pseudomonas
aeruginosa is dependent on cell-to-cell signaling and requires
flagella and pili. J Bacteriol 182: 5990–5996.
Liu X & Matsumura P (1994) The FlhD/FlhC complex, a
transcriptional activator of the Escherichia coli flagellar class II
operons. J Bacteriol 176: 7345–7351.
Majdalani N & Gottesman S (2005) The Rcs phosphorelay: a
complex signal transduction system. Annu Rev Microbiol 59:
379–405.
Overhage J, Lewenza S, Marr AK & Hancock REW (2007)
Identification of genes involved in swarming motility using a
Pseudomonas aeruginosa PAO1 Mini-Tn5-lux mutant library. J
Bacteriol 189: 2164–2169.
FEMS Microbiol Lett 317 (2011) 160–171c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
170 Y. Cong et al.
Dow
nloaded from https://academ
ic.oup.com/fem
sle/article/317/2/160/623831 by guest on 22 January 2022

Parrington LJ, Sharpe AN & Peterkin PI (1993) Improved aerobic
colony count technique for hydrophobic grid membrane
filters. Appl Environ Microb 59: 2784–2789.
Perez JM, Arenas FA, Pradenas GA, Sandoval JM & Vasquez CC
(2008) Escherichia coli YqhD exhibits aldehyde reductase
activity and protects from the harmful effect of lipid
peroxidation-derived aldehydes. J Biol Chem 283: 7346–7353.
Preiss J (1984) Bacterial glycogen synthesis and its regulation.
Annu Rev Microbiol 38: 419–458.
Pruss BM, Campbell JW, Van Dyk TK, Zhu C, Kogan Y &
Matsumura P (2003) FlhD/FlhC is a regulator of anaerobic
respiration and the Entner–Doudoroff pathway through
induction of the methyl-accepting chemotaxis protein. J
Bacteriol 185: 534–543.
Rather PN (2005) Swarmer cell differentiation in Proteus
mirabilis. Environ Microbiol 7: 1065–1073.
Rosenberg M, Gutnick D & Rosenberg E (1980) Adherence of
bacteria to hydrocarbons: a simple method for measuring cell-
surface hydrophobicity. FEMS Microbiol Lett 9: 29–33.
Semmler ABT, Whitchurch CB & Mattick JS (1999) A re-
examination of twitching motility in Pseudomonas aeruginosa.
Microbiology 145: 2863–2873.
Stickler D, Morris N, Moreno MC & Sabbuba N (1998) Studies
on the formation of crystalline bacterial biofilms on urethral
catheters. Eur J Clin Microbiol 17: 649–652.
Swords WE, Giddings A & Benjamin WH Jr (1997) Bacterial
phenotypes mediated by mviA and their relationship to the
mouse virulence of Salmonella typhimurium. Microb
Pathogenesis 22: 353–362.
Takeda SI, Fujisawa Y, Matsubara M, Aiba H & Mizuno T (2001)
A novel feature of the multistep phosphorelay in Escherichia
coli: a revised model of the RcsC ! YojN ! RcsB signalling
pathway implicated in capsular synthesis and swarming
behaviour. Mol Microbiol 40: 440–450.
Toguchi A, Siano M, Burkart M & Harshey RM (2000) Genetics
of swarming motility in Salmonella enterica serovar
Typhimurium: critical role for lipopolysaccharide. J Bacteriol
182: 6308–6321.
Turner PC, Miller EN, Jarboe LR, Baggett CL, Shanmugam KT &
Ingram LO (2011) YqhC regulates transcription of the adjacent
Escherichia coli genes yqhD and dkgA that are involved in
furfural tolerance. J Ind Microbiol Biot 38: 431–439.
Wang Q, Zhao Y, McClelland M & Harshey RM (2007) The
RcsCDB signaling system and swarming motility in Salmonella
enterica serovar typhimurium: dual regulation of flagellar and
SPI-2 virulence genes. J Bacteriol 189: 8447–8457.
Young GM, Schmiel DH & Miller VL (1999) A new pathway for
the secretion of virulence factors by bacteria: the flagellar
export apparatus functions as a protein secretion system. P
Natl Acad Sci USA 96: 6456–6461.
Supporting Information
Additional Supporting Information may be found in the
online version of this article:
Fig. S1. Electron micrograph of bacterial cell collected from
LB plate with 1.5% agar; scale bar = 2mm.
Fig. S2. Bacterial surface hydrophilicities measured by
BATH method, as described in the Materials and methods.
Fig. S3. Growth curves of the mutant and wild-type strains.
Fig. S4. SDS-PAGE of lipopolysaccharide profiles.
Fig. S5. Swarming colonies of Proteus mirabilis CMCC49003
stained in situ with TTC.
Video S1. Movement of wild type cells on swarm media.
Video S2. Movement of wzx mutant cells on swarm media
(episode 1).
Video S3. Movement of wzx mutant cells on swarm media
(episode 2).
Video S4. Movement of rcsD mutant cells on swarm media.
Video S5. Movement of yeeZ mutant cells in liquid LB
media.
Please note: Wiley-Blackwell is not responsible for the
content or functionality of any supporting materials sup-
plied by the authors. Any queries (other than missing
material) should be directed to the corresponding author
for the article.
FEMS Microbiol Lett 317 (2011) 160–171 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
171Swarming motility in C. freundii
Dow
nloaded from https://academ
ic.oup.com/fem
sle/article/317/2/160/623831 by guest on 22 January 2022
Related Documents











