Characterization of Pseudomonas syringae ssp. savastanoi strains isolated from ash N. S. Iacobellis a , A. Caponero b * and A. Evidente c a Dipartimento di Biologia, Difesa e Biotecnologie Agro Forestali, Universita ` degli Studi della Basilicata, Via N. Sauro, 85, 85100 Potenza, Italy; b Dipartimento di Patologia vegetale, Universita ` degli Studi di Bari, Via Amendola 165/A, 70124 Bari, Italy; and c Dipartimento di Scienze Chimico-Agrarie, Universita ` di Napoli ‘Federico II’, Via Universita ` 100, 80055 Portici, Italy Isolates of Pseudomonas syringae ssp. savastanoi from ash were examined for their ability to produce phytohormones in culture and for pathogenicity, in comparison with isolates from olive and oleander. Nineteen out of 20 ash isolates produced low levels of indole-3-acetic acid and its methyl ester but no cytokinins. In contrast, the remaining isolate, NCPPB3474, accumulated high levels of auxins and cytokinins in culture, comparable to those of olive and oleander strains. Hybridization of DNA preparations with tryptophan mono-oxygenase (iaaM) and isopentenyl transferase (ipt) gene-containing probes showed sequences of DNA homologous to both probes only in isolate NCPPB3474, and in which the iaaM and ipt genes were located on the chromosome and on a plasmid of about 80 kb, respectively. When assayed for pathogenicity on ash, olive and oleander, 19 of the 20 ash isolates caused disease only on ash but NCPPB3474 caused knots on both ash and olive. Oleander isolates infected all three hosts whereas those from olive caused symptoms only on olive and ash. All the cultures were able to multiply in host plant tissues, but the growth rates and final population densities were correlated to the plant species inoculated and the host origin of the isolates. In particular, the highest population densities were reached by isolates capable of causing symptoms on the inoculated host. The phytohormone production shown by ash, olive and oleander isolates of P. syringae ssp. savastanoi was in accordance with the type of symptoms: cankers accompanied by wart-like excrescences on ash and knots on olive and oleander. Furthermore, the pathogenic features of these isolates and, in particular, their growth patterns in the different host tissues, support previous evidence on the existence of three distinct pathovars in P. syringae ssp. savastanoi. Introduction Pseudomonas syringae ssp. savastanoi infects oleander (Nerium oleander) in the Apocynaceae, and several hosts in the Oleaceae, of which the most commonly attacked are olive (Olea europaea), ash (Fraxinus excelsior), privet (Ligustrum japonicum) and species of Jasminum, Forsythia and Phyllyirea. On all host species the pathogen induces an overgrowth of the infected tissues, causing knots to form on oleander, olive and several species of Oleaceae and cankers accompanied by wart- like excrescences on ash. Strains of the pathogen isolated from the different host plants were named differently in the past (Surico & Iacobellis, 1992). More recently, the pathogen was classified as Pseudomonas syringae pv. savastanoi with- out distinguishing its host origin (Young et al., 1978). Janse (1981) re-examined isolates from olive, oleander and ash and found that their biochemical, serological and pathological features differed from those of other members of P. syringae. Isolates from oleander, olive and ash were distinguishable by their pathogenicity on host plants. Consequently, Janse (1982a) proposed that the bacterium should be classified as a new subspecies, P. syringae ssp. savastanoi, and that three pathovars (pv. fraxinii, pv. oleae and pv. nerii) should be distinguished on the basis of their pathological features. This proposal was partly rejected by the International Society for Plant Pathology (ISPP) Subcommittee on the Taxonomy of Plant Pathogenic Bacteria only because the pathotype strains had not been proposed by the author. In fact, they lumped the three pathovars together under the designation Pseudomonas syringae ssp. savastanoi (Young et al., 1991). More recently, Gardan et al. (1992a) suggested that P.s. ssp. savastanoi should be elevated to the species level Pseudomonas savastanoi sp.nov. in view of its physio- logical and biochemical characters and on the basis of DNA–DNA hybridization data. Pseudomonas savas- tanoi thus includes pv. savastanoi, pv. glycinea and pv. phaseolicola, strains from oleander, with the Oleaceae being grouped in pv. savastanoi. Plant Pathology (1998) 47, 73–83 Q 1998 BSPP 73 *Present address: Ente Sviluppo Agricolo Basilicata, Via Passarelli, 7, Matera, Italy. Accepted 29 July 1997.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Characterization of Pseudomonas syringae ssp.savastanoi strains isolated from ash
N. S. Iacobellisa, A. Caponerob* and A. Evidentec
aDipartimento di Biologia, Difesa e Biotecnologie Agro Forestali, Universita degli Studi della Basilicata, Via N. Sauro, 85,85100 Potenza, Italy; bDipartimento di Patologia vegetale, Universita degli Studi di Bari, Via Amendola 165/A, 70124 Bari, Italy; andcDipartimento di Scienze Chimico-Agrarie, Universita di Napoli ‘Federico II’, Via Universita 100, 80055 Portici, Italy
Isolates of Pseudomonas syringae ssp. savastanoi from ash were examined for their ability to produce phytohormonesin culture and for pathogenicity, in comparison with isolates from olive and oleander. Nineteen out of 20 ash isolatesproduced low levels of indole-3-acetic acid and its methyl ester but no cytokinins. In contrast, the remaining isolate,NCPPB3474, accumulated high levels of auxins and cytokinins in culture, comparable to those of olive and oleanderstrains. Hybridization of DNA preparations with tryptophan mono-oxygenase (iaaM) and isopentenyl transferase(ipt) gene-containing probes showed sequences of DNA homologous to both probes only in isolate NCPPB3474, andin which the iaaM and ipt genes were located on the chromosome and on a plasmid of about 80 kb, respectively. Whenassayed for pathogenicity on ash, olive and oleander, 19 of the 20 ash isolates caused disease only on ash butNCPPB3474 caused knots on both ash and olive. Oleander isolates infected all three hosts whereas those from olivecaused symptoms only on olive and ash. All the cultures were able to multiply in host plant tissues, but the growthrates and final population densities were correlated to the plant species inoculated and the host origin of the isolates. Inparticular, the highest population densities were reached by isolates capable of causing symptoms on the inoculatedhost. The phytohormone production shown by ash, olive and oleander isolates of P. syringae ssp. savastanoi was inaccordance with the type of symptoms: cankers accompanied by wart-like excrescences on ash and knots on olive andoleander. Furthermore, the pathogenic features of these isolates and, in particular, their growth patterns in the differenthost tissues, support previous evidence on the existence of three distinct pathovars in P. syringae ssp. savastanoi.
Introduction
Pseudomonas syringae ssp. savastanoi infects oleander(Nerium oleander) in the Apocynaceae, and several hostsin the Oleaceae, of which the most commonly attackedare olive (Olea europaea), ash (Fraxinus excelsior),privet (Ligustrum japonicum) and species of Jasminum,Forsythia and Phyllyirea. On all host species thepathogen induces an overgrowth of the infected tissues,causing knots to form on oleander, olive and severalspecies of Oleaceae and cankers accompanied by wart-like excrescences on ash.
Strains of the pathogen isolated from the differenthost plants were named differently in the past (Surico &Iacobellis, 1992). More recently, the pathogen wasclassified as Pseudomonas syringae pv. savastanoi with-out distinguishing its host origin (Young et al., 1978).Janse (1981) re-examined isolates from olive, oleanderand ash and found that their biochemical, serological
and pathological features differed from those of othermembers of P. syringae. Isolates from oleander, olive andash were distinguishable by their pathogenicity on hostplants. Consequently, Janse (1982a) proposed that thebacterium should be classified as a new subspecies, P.syringae ssp. savastanoi, and that three pathovars (pv.fraxinii, pv. oleae and pv. nerii) should be distinguishedon the basis of their pathological features.
This proposal was partly rejected by the InternationalSociety for Plant Pathology (ISPP) Subcommittee on theTaxonomy of Plant Pathogenic Bacteria only because thepathotype strains had not been proposed by the author.In fact, they lumped the three pathovars together underthe designation Pseudomonas syringae ssp. savastanoi(Young et al., 1991).
More recently, Gardan et al. (1992a) suggested thatP.s. ssp. savastanoi should be elevated to the species levelPseudomonas savastanoi sp.nov. in view of its physio-logical and biochemical characters and on the basis ofDNA–DNA hybridization data. Pseudomonas savas-tanoi thus includes pv. savastanoi, pv. glycinea and pv.phaseolicola, strains from oleander, with the Oleaceaebeing grouped in pv. savastanoi.
Plant Pathology (1998) 47, 73–83
Q 1998 BSPP 73
*Present address: Ente Sviluppo Agricolo Basilicata,Via Passarelli, 7, Matera, Italy.
Accepted 29 July 1997.
Studies have shown that isolates of the bacteriumfrom the different host species may be differentiated onseveral characteristics, including pathogenicity andproduction of phytohormones (Surico & Iacobellis,1992). In particular, it is well established that olive andoleander strains produce high amounts of indole-3-acetic acid (IAA) and of cytokinins in culture. Further-more, knot formation on olive and oleander depends onthe ability of P. syringae ssp. savastanoi to producephytohormones (Surico & Iacobellis, 1992; Iacobellis etal., 1994).
In olive and oleander strains, IAA biosynthesis fromL-tryptophan, is catalysed by the enzymes tryptophanmono-oxygenase and indolacetamide hydrolase,encoded by the genes iaaM and iaaH, respectively(Comai & Kosuge, 1982; Yamada et al., 1985). Thebiosynthesis of cytokinins is catalysed by an isopentenyltransferase enzyme whose genetic determinant, ipt, hasbeen cloned and sequenced (Powell & Morris, 1986).
In contrast, the ability of ash strains to producephytohormones has not yet been defined and conflictingfindings have been reported (Janse, 1981; Gardan et al.,1992b). In fact, Janse (1981) reported that, in culture,ash strains produce small amounts of substances withthe same chromatographic behaviour as IAA. These datahave not been confirmed by Gardan et al. (1992b) andno substances with cytokinin-like activity have beendetected.
In the present paper the production of phytohormonesby ash strains and their pathogenetic features on ash,olive and oleander are reported.
Materials and methods
Bacterial cultures
The Pseudomonas syringae ssp. savastanoi cultures usedare listed in Table 1. Bacteria were maintained at ¹708Cin presence of 15% (v/v) glycerol. Bacterial subcultureswere obtained on nutrient agar containing 2% glycerol(NGA) and slants incubated at 268C, unless otherwisestated.
Before use, the cultures were tested for characters ofthe LOPAT scheme (Lelliott & Stead, 1987).
Production of plant growth substances
Bacteriological vials containing 3 mL of liquid KB medium(King et al., 1954), with or without 0.5 mM L-trypto-phan, were inoculated with a loopful of bacterial cells atlog phase. After 5 days incubation under shaking(200 r.p.m.) at 268C, cultures were centrifuged and theindole content, expressed as mg L¹1 IAA equivalents,was determined colorimetrically by Salkowski’s reagent(Gordon & Weber, 1951). Three vials were used for eachisolate. The experiment was repeated twice and theresults analysed for variance and by Duncan’s multiplerange test.
To determine cytokinin production, bacteria were
grown under shaking at 268C in 1 L Erlenmeyer flaskscontaining 200 mL of MM (minimal medium) (5 gglucose, 3 g K2HPO4, 1 g NH4Cl, 0.3 g MgSO4.7H2O,0.5 g sodium citrate per litre of distilled water). After 5days incubation, cells were removed by centrifugation(7000 g, 10 min) and filtration (Millipore 0.45 mm).Filtrates were extracted four times with an equalvolume of ethyl acetate at pH 8.5. The extracts weredried over anhydrous Na2SO4, concentrated underreduced pressure and the organic residues dissolved ina small volume of methanol. The extracts were tested forcytokinin activity, using the etiolated cucumber cotyle-don bioassay (Fletcher et al., 1982). The content ofcytokinin-like substances was expressed as mg L¹1
equivalents of t-zeatin (t-Z).
Characterization of phytohormones fromrepresentative cultures
To characterize the phytohormones produced, three P.syringae ssp. savastanoi isolates were selected. Two
N. S. Iacobellis et al.74
Q 1998 BSPP Plant Pathology (1998) 47, 73–83
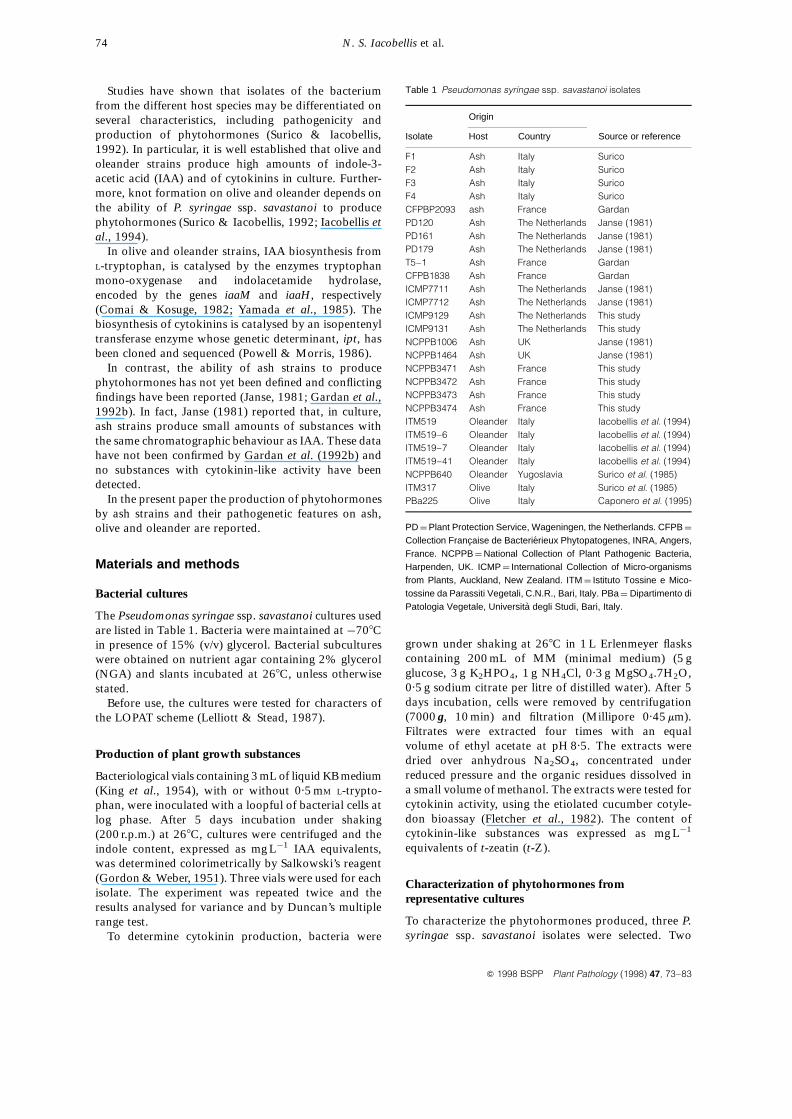
Table 1 Pseudomonas syringae ssp. savastanoi isolates
Origin
Isolate Host Country Source or reference
F1 Ash Italy SuricoF2 Ash Italy SuricoF3 Ash Italy SuricoF4 Ash Italy SuricoCFPBP2093 ash France GardanPD120 Ash The Netherlands Janse (1981)PD161 Ash The Netherlands Janse (1981)PD179 Ash The Netherlands Janse (1981)T5–1 Ash France GardanCFPB1838 Ash France GardanICMP7711 Ash The Netherlands Janse (1981)ICMP7712 Ash The Netherlands Janse (1981)ICMP9129 Ash The Netherlands This studyICMP9131 Ash The Netherlands This studyNCPPB1006 Ash UK Janse (1981)NCPPB1464 Ash UK Janse (1981)NCPPB3471 Ash France This studyNCPPB3472 Ash France This studyNCPPB3473 Ash France This studyNCPPB3474 Ash France This studyITM519 Oleander Italy Iacobellis et al. (1994)ITM519–6 Oleander Italy Iacobellis et al. (1994)ITM519–7 Oleander Italy Iacobellis et al. (1994)ITM519–41 Oleander Italy Iacobellis et al. (1994)NCPPB640 Oleander Yugoslavia Surico et al. (1985)ITM317 Olive Italy Surico et al. (1985)PBa225 Olive Italy Caponero et al. (1995)
PD ¼ Plant Protection Service, Wageningen, the Netherlands. CFPB ¼
Collection Francaise de Bacterierieux Phytopatogenes, INRA, Angers,France. NCPPB ¼ National Collection of Plant Pathogenic Bacteria,Harpenden, UK. ICMP ¼ International Collection of Micro-organismsfrom Plants, Auckland, New Zealand. ITM ¼ Istituto Tossine e Mico-tossine da Parassiti Vegetali, C.N.R., Bari, Italy. PBa ¼ Dipartimento diPatologia Vegetale, Universita degli Studi, Bari, Italy.

(PD179 and F4) showed the typical features of most ashisolates; the other (NCPPB3474) was atypical. Bacteriawere grown under shaking as above in 1 L Erlenmeyerflasks containing 200 mL of MM medium with 0.5 mM
L-tryptophan. After 5 days incubation, cells were removedby centrifugation (7000 g, 10 min) and filtration (Milli-pore 0.45 mm). Filtrates were extracted in darkness fourtimes with an equal volume of ethyl acetate at pH 2.8and subsequently at pH 8.5. The extracts were driedover anhydrous Na2SO4 and concentrated underreduced pressure. Organic residues were dissolved in asmall volume of methanol and analysed for indole andcytokinin content by thin layer chromatography (TLC),applying the methods described in Table 2.
For auxin quantification, aliquots of the acidicextracts were fractionated by TLC (silica gel, solventA). Regions from the developed chromatograms corre-sponding to authentic IAA and its methyl ester (IAAMe)were scraped off, eluted from silica gel with methanol,and the organic phases were analysed by high-perfor-mance liquid chromatography (HPLC) under the condi-tions reported in Table 2. IAA and IAAMe quantificationwas determined from peak areas using standard curvesmade with authentic samples of IAA and IAAMe.
Alkaline extracts were analysed by TLC (silica gel,solvent C). Authentic samples of the known cytokininsdihydrozeatin (diHZ), 9-b-D-dihydrozeatin riboside(diHZR), trans-zeatin riboside (t-ZR) and 10 0-methylzea-tin riboside (10 0MeZR), were used as standards. UV-absorbing bands from alkaline extracts that co-migratedwith authentic substances were recovered and fraction-ated on reverse phase plates (solvent D) and by HPLC asindicated in Table 2. Cytokinin quantification wasdetermined from peak areas using standard curves
made with authentic samples of diHZ, diHZR, t-ZRand 10 0MeZR.
The identity of the substances co-purifying withauthentic IAA, IAAMe, diHZ, diHZR, t-ZR and100MeZR was confirmed by spectroscopic analysis, asreported previously (Evidente et al., 1995).
DNA analysis
Pseudomonas syringae ssp. savastanoi isolates weregrown under orbital shaking (200 r.p.m.) in KB brothat 268C. After 24 h incubation, 1 mL cultures werecentrifuged (7000 g, 10 min) and pellets were washedtwice with 0.85% NaCl, suspended in a minimal volumeof this solution and stored overnight at ¹208C. PlasmidDNA was extracted by the alkaline lysis procedure(Maniatis et al., 1982). Plasmid DNA preparations wereanalysed by horizontal gel electrophoresis in 0.5%agarose tris-borate buffer. Plasmids from P. syringaessp. savastanoi strain ITM519 were used as molecularweight standards (Iacobellis et al., 1994).
Genomic DNA was extracted as reported by Glass &Kosuge (1988).
DNA was transferred from agarose gel to nitro-cellulose filters (BA85; Schleicher and Schuell, Germany)according to the manufacturer’s instructions, or spotteddirectly on the filters and hybridized with DNA probesradioactively labelled using the multiprime DNA label-ling system (Amersham, Amersham, UK). A 2.8-kbEcoRI fragment of the tryptophan methyl-transferasegene, iaaM (Palm et al., 1989), and a 0.6-kb RsaIinternal fragment of the isopentenyl transferase gene, ipt(Powell & Morris, 1986), both isolated from P. syringaessp. savastanoi, were used as probes.
Pseudomonas syringae ssp. savastanoi from ash 75
Q 1998 BSPP Plant Pathology (1998) 47, 73–83
Table 2 Chromatographic methods for the analysis of indoles and cytokinins produced by Pseudomonas syringae ssp. savastanoi ash isolates
HPLCTLC Reverse-phase TLC (Hypersil ODS 5 mm ‘Altech’,(Merk, Kieselgel (Whatmann, Stratochrom, 150 ×4.6 mm)60F254, 0.25 mm) SIF254 KC18, 0.02 mm)
Flow rate UV-detectorCompounda Solvent Detected by Solvent Detected by Solvent mL min¹1 nm
IAA, (A) CHCl3- UV light, MeCN- 1 280IAAMe EtOAc- Salkowki’s reagent H2O,
MeOH, 7 : 3, v/v2/2/1, v/v/v
(B) CHCl3- UV light,MeOH, Salkowski’s reagent6 : 4, v/v
t-ZR, (C) BuOH- UV light (D) H2O- UV light 0–30% 2.5 262diHZ, AcOH-H2O, EtOH, MeCNb,10 0MeZR, 60 :15 : 25, 6 : 4, v/v in 20 mindiHZR v/v/v
a IAA ¼ indole-3-acetic acid; IAAMe ¼ IAA methyl ester; t-ZR ¼ trans-zeatin riboside; diHZ ¼ dihydrozeatin; diHZR ¼ dihydro-zeatin riboside;10 0MeZR ¼ 10 0-methyl zeatin riboside.b This is an aqueous solution in which pH is brought to 7 with triethylammonium carbonate (TEAB), obtained by CO2 saturation of a 2.5 M solution oftriethylamine in H2O.
Pathogenicity tests
Pseudomonas syringae ssp. savastanoi isolates wereinoculated into 1-year-old ash, olive and oleanderstems. Wounds about 1 cm long were made in the barkwith a sterile scalpel dipped in bacterial suspensionscontaining about 109 CFU mL¹1. Three wounds on threeplants per host species were inoculated with each isolate.Sterile distilled water (SDW) was used as control. Theinoculated wounds were then covered with parafilm for96 h. The inoculated plants, maintained in a glasshouseat 25 6 28C and 80–85% humidity, were observed forsymptom development.
For some selected cultures, bacterial populationdensities in the infected tissues were determined 1month after inoculation. The inoculated stems wererinsed with SDW and tissues at infection sites wereremoved and homogenized in SDW in a mortar. The
resulting suspensions were serially diluted and plated onnutrient agar containing 1% sucrose (NSA) for colonycounts. Population density was expressed as CFU perinoculum site. For each isolate, three inoculum sites wereused. The experiment was repeated twice and resultswere analysed for variance and by Duncan’s multiplerange test.
Bacterial growth in plant leaves
Selected isolates of P. syringae ssp. savastanoi were used.Hypodermic syringes were used to inject bacterialsuspensions containing about 106 CFU mL¹1 into theabaxial surface of ash, oleander and olive leaves. Leavestreated with SDW were used as controls. The treated leafareas appeared watersoaked. The plants were main-tained in a glasshouse as above.
N. S. Iacobellis et al.76
Q 1998 BSPP Plant Pathology (1998) 47, 73–83
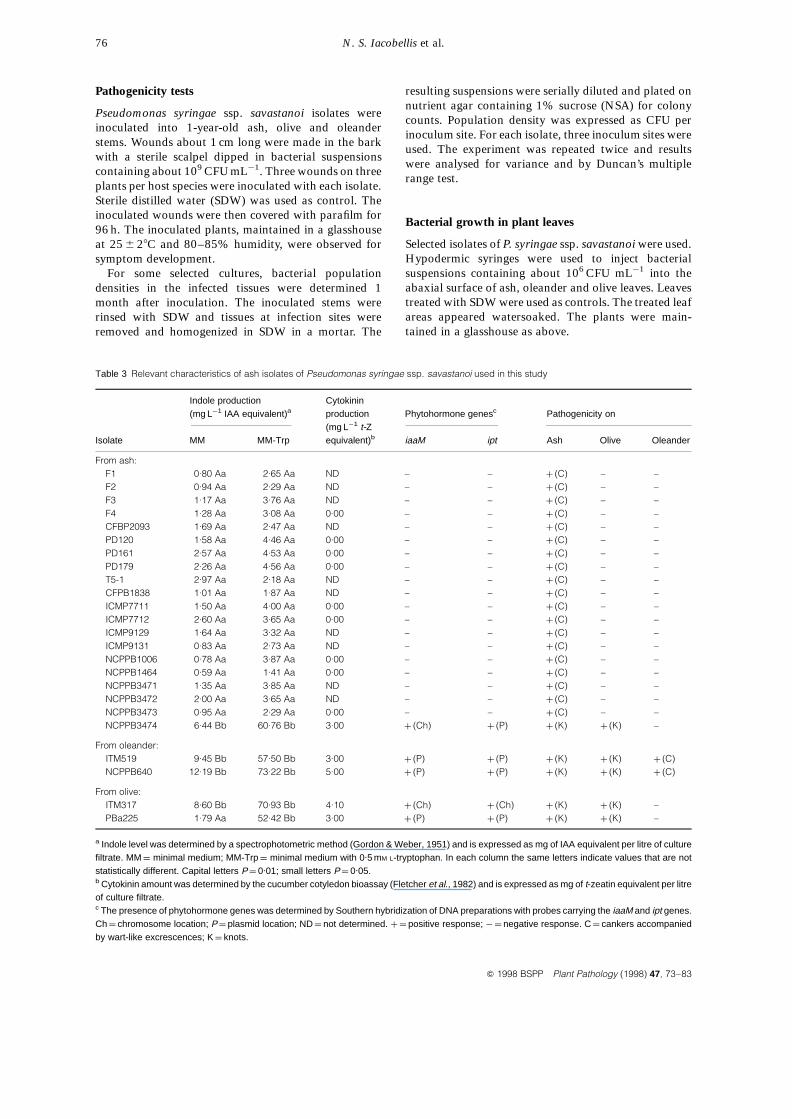
Table 3 Relevant characteristics of ash isolates of Pseudomonas syringae ssp. savastanoi used in this study
Indole production Cytokinin(mg L¹1 IAA equivalent)a production Phytohormone genesc Pathogenicity on
(mg L¹1 t-ZIsolate MM MM-Trp equivalent)b iaaM ipt Ash Olive Oleander
From ash:F1 0.80 Aa 2.65 Aa ND – – þ (C) – –F2 0.94 Aa 2.29 Aa ND – – þ (C) – –F3 1.17 Aa 3.76 Aa ND – – þ (C) – –F4 1.28 Aa 3.08 Aa 0.00 – – þ (C) – –CFBP2093 1.69 Aa 2.47 Aa ND – – þ (C) – –PD120 1.58 Aa 4.46 Aa 0.00 – – þ (C) – –PD161 2.57 Aa 4.53 Aa 0.00 – – þ (C) – –PD179 2.26 Aa 4.56 Aa 0.00 – – þ (C) – –T5-1 2.97 Aa 2.18 Aa ND – – þ (C) – –CFPB1838 1.01 Aa 1.87 Aa ND – – þ (C) – –ICMP7711 1.50 Aa 4.00 Aa 0.00 – – þ (C) – –ICMP7712 2.60 Aa 3.65 Aa 0.00 – – þ (C) – –ICMP9129 1.64 Aa 3.32 Aa ND – – þ (C) – –ICMP9131 0.83 Aa 2.73 Aa ND – – þ (C) – –NCPPB1006 0.78 Aa 3.87 Aa 0.00 – – þ (C) – –NCPPB1464 0.59 Aa 1.41 Aa 0.00 – – þ (C) – –NCPPB3471 1.35 Aa 3.85 Aa ND – – þ (C) – –NCPPB3472 2.00 Aa 3.65 Aa ND – – þ (C) – –NCPPB3473 0.95 Aa 2.29 Aa 0.00 – – þ (C) – –NCPPB3474 6.44 Bb 60.76 Bb 3.00 þ (Ch) þ (P) þ (K) þ (K) –
From oleander:ITM519 9.45 Bb 57.50 Bb 3.00 þ (P) þ (P) þ (K) þ (K) þ (C)NCPPB640 12.19 Bb 73.22 Bb 5.00 þ (P) þ (P) þ (K) þ (K) þ (C)
From olive:ITM317 8.60 Bb 70.93 Bb 4.10 þ (Ch) þ (Ch) þ (K) þ (K) –PBa225 1.79 Aa 52.42 Bb 3.00 þ (P) þ (P) þ (K) þ (K) –
a Indole level was determined by a spectrophotometric method (Gordon & Weber, 1951) and is expressed as mg of IAA equivalent per litre of culturefiltrate. MM ¼ minimal medium; MM-Trp ¼ minimal medium with 0.5 mM L-tryptophan. In each column the same letters indicate values that are notstatistically different. Capital letters P ¼ 0.01; small letters P ¼ 0.05.b Cytokinin amount was determined by the cucumber cotyledon bioassay (Fletcher et al., 1982) and is expressed as mg of t-zeatin equivalent per litreof culture filtrate.c The presence of phytohormone genes was determined by Southern hybridization of DNA preparations with probes carrying the iaaM and ipt genes.Ch ¼ chromosome location; P ¼ plasmid location; ND ¼ not determined. þ ¼ positive response; ¹¼ negative response. C ¼ cankers accompaniedby wart-like excrescences; K ¼ knots.
At 0, 1, 2, 3 and 5 days after inoculation, three leavesper isolate were rinsed with SDW, the surfaces cleanedwith 60% ethanol, dried, and sections of inoculatedtissues were weighed and homogenized in a mortar inSDW. The resulting suspensions were serially diluted andplated on NSA for colony counts. Population densitywas expressed as CFU per 100 mg of fresh tissue. Theexperiment was repeated twice and the results wereanalysed for variance and by Duncan’s multiple rangetest.
Results
Biochemical and physiological characterization
All the isolates showed the characters of the LOPAT Ibgroup of fluorescent pseudomonads to which P. syringaessp. savastanoi belongs (Lelliot & Stead, 1987). Theonly variable character was the morphology of thecolony when bacteria were grown on NSA: the ashisolates, namely PD120, PD161, PD179, ICMP9131,NCPPB3471 and NCPPB3473, produced levaniformcolonies, whereas the others did not. Isolate PD120 didnot produce any fluorescent pigments when grown onKB medium.
Indole production
All the P. syringae ssp. savastanoi ash isolates producedindoles when grown in minimal medium MM (Table 3).In particular, 19 out of 20 isolates accumulated amountsof indoles ranging from 0.59 to 2.97 mg L¹1 IAAequivalents, which were not significantly different.Olive and oleander strains produced higher levels ofindoles, different (P ¼ 0.01) from those of most of theash isolates, the only exception being olive strainPBa225, which accumulated levels similar to those ofthe ash isolates.
When ash isolates were grown in the presence oftryptophan, they produced higher amounts of indoles,ranging from 1.41 to 4.56 mg L¹1 IAA equivalents(Table 3), but still much lower than those of the oliveand oleander isolates which, under the same growthconditions, accumulated indoles ranging from 52.42 to73.22 mg L¹1 IAA equivalents. The olive strain PBa225also produced 52.42 mg L¹1 IAA equivalents in thepresence of the precursor.
The remaining ash isolate NCPPB3474 behaveddifferently. It produced 6.44 and 60.76 mg L¹1 IAAequivalents when grown with or without tryptophan,respectively (Table 3). These levels of indoles werehigher (P ¼ 0.01) than those produced by the other ashisolates, and nearer to those of the olive and oleanderisolates.
Cytokinin production
The alkaline extracts from nine of 10 ash cultures tested
in the cucumber cotyledon bioassay showed nocytokinin-like activities (Table 3). Only the extractfrom NCPPB3474 showed a strong cytokinin-likeactivity, corresponding to about 3 mg L¹1 t-Z equiva-lents. Levels of cytokinin-like activities ranging from 3to 5 mg L¹1 t-Z equivalents were also observed inthe alkaline extracts of the olive and oleander strains(Table 3).
Characterization of phytohormones fromrepresentative cultures
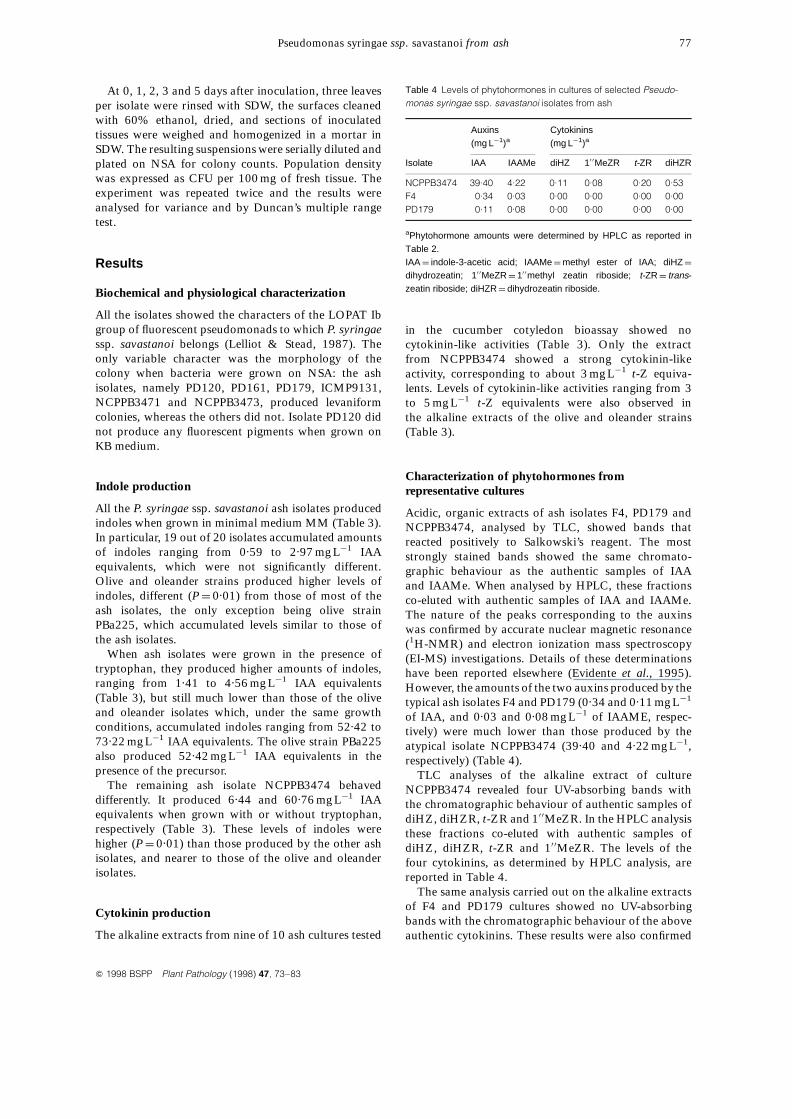
Acidic, organic extracts of ash isolates F4, PD179 andNCPPB3474, analysed by TLC, showed bands thatreacted positively to Salkowski’s reagent. The moststrongly stained bands showed the same chromato-graphic behaviour as the authentic samples of IAAand IAAMe. When analysed by HPLC, these fractionsco-eluted with authentic samples of IAA and IAAMe.The nature of the peaks corresponding to the auxinswas confirmed by accurate nuclear magnetic resonance(1H-NMR) and electron ionization mass spectroscopy(EI-MS) investigations. Details of these determinationshave been reported elsewhere (Evidente et al., 1995).However, the amounts of the two auxins produced by thetypical ash isolates F4 and PD179 (0.34 and 0.11 mg L¹1
of IAA, and 0.03 and 0.08 mg L¹1 of IAAME, respec-tively) were much lower than those produced by theatypical isolate NCPPB3474 (39.40 and 4.22 mg L¹1,respectively) (Table 4).
TLC analyses of the alkaline extract of cultureNCPPB3474 revealed four UV-absorbing bands withthe chromatographic behaviour of authentic samples ofdiHZ, diHZR, t-ZR and 10 0MeZR. In the HPLC analysisthese fractions co-eluted with authentic samples ofdiHZ, diHZR, t-ZR and 10 0MeZR. The levels of thefour cytokinins, as determined by HPLC analysis, arereported in Table 4.
The same analysis carried out on the alkaline extractsof F4 and PD179 cultures showed no UV-absorbingbands with the chromatographic behaviour of the aboveauthentic cytokinins. These results were also confirmed
Pseudomonas syringae ssp. savastanoi from ash 77
Q 1998 BSPP Plant Pathology (1998) 47, 73–83
Table 4 Levels of phytohormones in cultures of selected Pseudo-monas syringae ssp. savastanoi isolates from ash
Auxins Cytokinins(mg L¹1)a (mg L¹1)a
Isolate IAA IAAMe diHZ 10 0MeZR t-ZR diHZR
NCPPB3474 39.40 4.22 0.11 0.08 0.20 0.53F4 0.34 0.03 0.00 0.00 0.00 0.00PD179 0.11 0.08 0.00 0.00 0.00 0.00
aPhytohormone amounts were determined by HPLC as reported inTable 2.IAA ¼ indole-3-acetic acid; IAAMe ¼ methyl ester of IAA; diHZ ¼
dihydrozeatin; 10 0MeZR ¼ 10 0methyl zeatin riboside; t-ZR ¼ trans-zeatin riboside; diHZR ¼ dihydrozeatin riboside.
by HPLC analysis. The nature of the peaks correspond-ing to the cytokinins was confirmed by accurate 1H- and13C-NMR and EI-MS investigations. Details of theabove determinations have been reported elsewhere(Evidente et al., 1995).
Plasmid content
All the ash isolates contained two to six plasmidsranging in size from 10 to 113 kb. Although plasmidswith similar sizes appeared to be present in someisolates, no correlation between the plasmid contentand the geographic origin of the cultures was observed.The only exceptions were cultures F1, F2, F3 and F4,isolated from ash plants in a limited area in northernItaly. These showed four plasmids (F1) and five plasmids(F2, F3, F4) of apparently similar size.
Phytohormone gene location
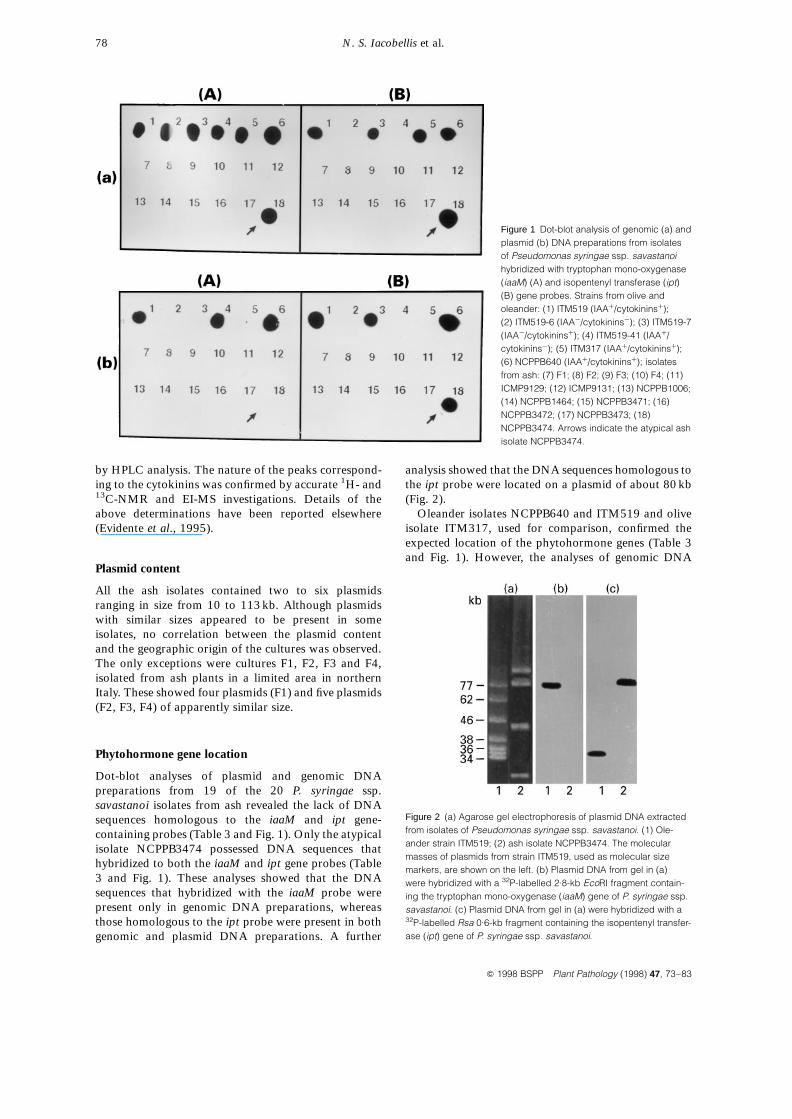
Dot-blot analyses of plasmid and genomic DNApreparations from 19 of the 20 P. syringae ssp.savastanoi isolates from ash revealed the lack of DNAsequences homologous to the iaaM and ipt gene-containing probes (Table 3 and Fig. 1). Only the atypicalisolate NCPPB3474 possessed DNA sequences thathybridized to both the iaaM and ipt gene probes (Table3 and Fig. 1). These analyses showed that the DNAsequences that hybridized with the iaaM probe werepresent only in genomic DNA preparations, whereasthose homologous to the ipt probe were present in bothgenomic and plasmid DNA preparations. A further
analysis showed that the DNA sequences homologous tothe ipt probe were located on a plasmid of about 80 kb(Fig. 2).
Oleander isolates NCPPB640 and ITM519 and oliveisolate ITM317, used for comparison, confirmed theexpected location of the phytohormone genes (Table 3and Fig. 1). However, the analyses of genomic DNA
N. S. Iacobellis et al.78
Q 1998 BSPP Plant Pathology (1998) 47, 73–83
Figure 1 Dot-blot analysis of genomic (a) andplasmid (b) DNA preparations from isolatesof Pseudomonas syringae ssp. savastanoihybridized with tryptophan mono-oxygenase(iaaM) (A) and isopentenyl transferase (ipt)(B) gene probes. Strains from olive andoleander: (1) ITM519 (IAAþ/cytokininsþ);(2) ITM519-6 (IAA¹/cytokinins¹); (3) ITM519-7(IAA¹/cytokininsþ); (4) ITM519-41 (IAAþ/cytokinins¹); (5) ITM317 (IAAþ/cytokininsþ);(6) NCPPB640 (IAAþ/cytokininsþ); isolatesfrom ash: (7) F1; (8) F2; (9) F3; (10) F4; (11)ICMP9129; (12) ICMP9131; (13) NCPPB1006;(14) NCPPB1464; (15) NCPPB3471; (16)NCPPB3472; (17) NCPPB3473; (18)NCPPB3474. Arrows indicate the atypical ashisolate NCPPB3474.
Figure 2 (a) Agarose gel electrophoresis of plasmid DNA extractedfrom isolates of Pseudomonas syringae ssp. savastanoi. (1) Ole-ander strain ITM519; (2) ash isolate NCPPB3474. The molecularmasses of plasmids from strain ITM519, used as molecular sizemarkers, are shown on the left. (b) Plasmid DNA from gel in (a)were hybridized with a 32P-labelled 2.8-kb EcoRI fragment contain-ing the tryptophan mono-oxygenase (iaaM) gene of P. syringae ssp.savastanoi. (c) Plasmid DNA from gel in (a) were hybridized with a32P-labelled Rsa 0.6-kb fragment containing the isopentenyl transfer-ase (ipt) gene of P. syringae ssp. savastanoi.

from isolates ITM519-6 and ITM519-7, two IAA¹
derivatives of isolate ITM519 lacking the IAA operonlocated on the plasmid of 77 kb (Iacobellis et al., 1994),showed signals of hybridization with the iaaM geneprobe (Fig. 1). As expected, no such signals wereobserved when the plasmid DNA preparations of theseisolates were used (Fig. 1).
Pathogenicity tests
All the P. syringae ssp. savastanoi isolates from ashcaused symptoms when inoculated on ash (Table 3). For
the majority of cultures (19 out of 20), the first symp-toms were observed 7–10 days after inoculation. At thattime, the edges of the inoculated wounds appearedwatersoaked and swollen; during the following days, amoderate tissue proliferation was observed between theedges of the wound. Thirty days after inoculation, theinfected tissues turned brown and appeared as opencankers accompanied by wart-like excrescences (Table 3and Fig. 3).
Isolate NCPPB3474 was virulent on ash but both thedevelopment of the symptoms and their final appearancewere different from those caused by the other ashisolates. At 5–7 days after inoculation, the infectedtissues were watersoaked and their overgrowth wasapparent; thereafter they evolved into typical knots(Table 3 and Fig. 3). Similar symptom development wasobserved when olive and oleander strains were inocu-lated on ash; again the final symptoms were typical knots(Table 3 and Fig. 3).
On olive plants, 19 of the 20 ash isolates caused nosymptoms, although in some cases the inoculated tissuesbecame swollen. In contrast, isolate NCPPB3474 causedknots similar to those caused by olive and oleanderisolates (Table 3 and Fig. 3). On oleander plants, nosymptoms were caused by ash and olive isolates, butoleander isolates induced typical knots (Table 3 andFig. 3). No symptoms were observed on plants treatedwith sterile distilled water (Fig. 3).
Bacterial growth in leaves
All the P. syringae ssp. savastanoi isolates were able tomultiply in ash and oleander leaves but not in oliveleaves (Fig. 4). In ash leaves, the highest growth ratewas observed up to 2 days after inoculation (3 daysin the case of isolate PD120). At that time the popula-tion densities ranged from 3.2 ×103 to 7.0 ×104 CFUper 100 mg fresh weight tissue. Thereafter, bacteriacontinued to multiply but at a reduced growth rate. Fivedays after inoculation they reached population densitiesranging from 1.2 ×104 to 3.2 ×106 CFU per 100 mgfresh weight tissue. At that time, isolate PD120 reachedthe highest population density (3.2 ×106 CFU per 100 mgfresh weight tissue), significantly different (P ¼ 0.01) fromthose of all the other isolates. No significant differenceswere observed among the population densities of isolatesICMP7712, NCPPB3474 (both from ash), ITM519(from oleander) and ITM317 (from olive). The popula-tion density of ash isolate PD161 differed (P ¼ 0.01)from that of ash isolate PD120 and the other ash, oliveand oleander isolates. No apparent correlation betweenthe host origin of the isolates and their growth capabilitywas observed. Seven days after inoculation, the zones ofthe ash leaves infiltrated with bacteria turned chloroticand subsequently necrotic. No symptoms were observedon leaves treated with SDW.
Two days after inoculation into oleander leaves, theoleander isolate ITM519 reached a population densityof 6.4 ×106 CFU per 100 mg fresh weight tissue (Fig. 4).
Pseudomonas syringae ssp. savastanoi from ash 79
Q 1998 BSPP Plant Pathology (1998) 47, 73–83
Figure 3 Symptoms on ash (a), olive (b) and oleander (c) stems 30days after inoculation with strains of Pseudomonas syringae ssp.savastanoi from ash, olive and oleander. (1) ICMP7712 (from ash);(2) NCPPB3474 (from ash); (3) ITM317 (from olive); (4) ITM519 (fromoleander); (5) treated with sterile distilled water.

This population level was higher (P ¼ 0.01) than those ofthe olive and ash strains, which reached densitiesranging from 7.9 ×104 to 8.0 ×105 CFU per 100 mgfresh weight tissue. Thereafter, bacteria continued tomultiply but at a reduced growth rate (Fig. 4). Five daysafter inoculation, oleander isolate ITM519 showed thehighest population density (3.9 ×107 CFU per 100 mgfresh weight tissue), which differed (P ¼ 0.01) fromthose of the olive and ash strain. No significantdifferences were observed among the population den-sities of isolates PD161, ICMP7712, NCPPB3474 andITM317, which ranged from 1.7 ×105 to 3.4 ×106 CFUper 100 mg fresh weight tissue. The population densityof ash isolate PD120 reached 3.4 × 106 CFU per 100 mgfresh weight tissue and differed (P ¼ 0.01) from those ofthe above isolates.
About 10 days after inoculation with the oleanderisolate ITM519, the inoculation sites showed out-growths that subsequently evolved into knots. Nosymptoms were observed on leaves inoculated witholive and ash isolates or treated with SDW.
No isolates were able to multiply in olive leaves(Fig. 4). In fact, bacterial populations rapidly decreasedand 2 days after inoculation no living cells wererecovered.
Olive leaves treated with SDW or inoculated withbacteria showed no symptoms and remained apparentlyhealthy for several months.
Bacterial growth in bark tissues
The different growth capability of P. syringae ssp.savastanoi isolates in host plant tissues was confirmedwhen selected isolates were inoculated into ash, oliveand oleander stems (Table 5).
In ash bark tissues, isolates PD179 and F4 from ash,which caused cankers accompanied by wart-like excres-cences, reached similar population densities of 1.7 ×108
and 1.9 ×108 CFU per inoculum site, respectively, 1
month after inoculation. After the same period, oleanderand olive isolates ITM519 and ITM317, which causedknots, produced population densities of 7.3 ×107 and3.1 ×107 CFU per inoculum site, respectively, whichdiffered significantly (P ¼ 0.01) and were also signifi-cantly different from those of the ash isolates.
In olive bark tissues, isolates PD179 and F4 from ash,which caused no symptoms on this host, gave populationdensities of 1.6 ×105 and 6.6 ×105 CFU per inoculumsite, respectively. These population levels were lower(P ¼ 0.01) than those produced by olive and oleanderisolates ITM317 and ITM519, which caused knots onolive and produced population densities of 6 ×107 and3 ×107 CFU per inoculum site, respectively.
On oleander, ash isolates PD179, F4 and olive isolate
N. S. Iacobellis et al.80
Q 1998 BSPP Plant Pathology (1998) 47, 73–83
Figure 4 Growth curves of Pseudomonassyringae ssp. savastanoi strains from ash(PD120, PD161, ICMP7712, NCPPB3474),olive (ITM317) and oleander (ITM519) in leaftissues of ash (a), oleander (b) and olive (c).
Table 5 Population densities of Pseudomonas syringae ssp. savas-tanoi isolates from ash (PD179 and F4), olive (ITM317) and oleander(ITM519) 30 days after inoculation in host plant stems
Population densityIsolate Host plant (CFU inoculum site)a Symptoms
PD179 Ash 1.7 ×108 Aa CF4 Ash 1.9 ×108 Aa CITM317 Ash 3.1 ×107 Cc KITM519 Ash 7.3 ×107 Bb K
PD179 Olive 6.6 ×105 Cc –F4 Olive 1.6 ×105 Cc –ITM317 Olive 6.0 ×107 Aa KITM519 Olive 3.0 ×107 Bb K
PD179 Oleander 2.1 ×105 Bb –F4 Oleander 6.6 ×104 Bb –ITM317 Oleander 2.4 ×105 Bb –ITM519 Oleander 2.1 ×107 Aa K
aIn each column the same letters indicate values that are notstatistically different. Capital letters P ¼ 0.01; small letters P ¼ 0.05.C ¼ cankers accompanied by wart-like excrescences; K ¼ knots; ¹ ¼
no symptoms.

ITM317, which did not cause any symptoms, producedpopulation levels ranging from 1.2 ×105 to2.1 ×105 CFU per inoculum site, which were notsignificantly different (P ¼ 0.05). The oleander isolateITM519, which caused symptoms (knots) on this hostplant, reached the highest population density of2.1 ×107 CFU per inoculum site, which was significantlyhigher (P ¼ 0.01) than those shown by either ash or oliveisolates.
Discussion
Isolates of P. syringae ssp. savastanoi from ash, olive andoleander are indistinguishable on the basis of the bio-chemical and physiological characters of the LOPATscheme (Lelliot & Stead, 1987), although variationsamong the isolates in the production of levan andfluorescent pigments have been observed (Janse, 1981;Iacobellis et al., 1993).
Despite the apparent homogenicity of biochemicaland physiological characters of the P. syringae ssp.savastanoi isolates from ash, olive and oleander, as wellas their high DNA–DNA homology (Janse, 1981;Gardan et al., 1992a), the findings of this study confirmthat they differ in their production of phytohormonesand in their pathogenicity.
Ash isolates did not produce cytokinins, in contrast tothose from olive and oleander which typically producethem in high amounts (Surico & Iacobellis, 1992).Moreover, they did not possess sequences of DNAhomologous to the ipt gene, which plays a key role in thecytokinin production of oleander and olive isolates of P.syringae ssp. savastanoi (Powell & Morris, 1986). Thesefindings confirm the previous results of Janse (1981),who found no substances with cytokinin-like activities inthe organic extracts of ash isolate cultures. Furthermore,they indicate that cytokinins have no role in the bacterialcanker of ash, as opposed to their activity in olive andoleander knot diseases (Surico & Iacobellis, 1992;Iacobellis et al., 1994).
Ash isolates typically produced auxins but their levelsin culture were much lower than those produced by oliveand oleander isolates (Surico & Iacobellis, 1992; Gardanet al., 1992b; Iacobellis et al., 1994). These resultsresolve previous conflicting reports on the ability of ashstrains to produce auxins (Janse, 1981; Gardan et al.,1992b).
Noteworthy is the fact that, in culture, ash strainsproduced not only IAA but also its methyl ester. Thismetabolite of IAA has been isolated from the culturesof some olive isolates of P. syringae ssp. savastanoi(Iacobellis et al., 1993) and of P. amygdali (Evidente etal., 1993), the agent of hyperplastic bacterial cankers ofalmond. No evidence is yet available for the presenceof this metabolite in oleander isolates. The fact thatthe methyl ester of IAA, thought to be a storage form ofIAA in higher plants (Cohen & Bandurski, 1982), is acommon metabolite in some overgrowth-inducingphytopathogenic bacteria, suggests that this compound
may be important. It is possible that IAAMe, as well asIAA–lysine conjugates (Hutzinger & Kosuge, 1968;Evidente et al., 1986; Glass & Kosuge, 1988), may playa role in the regulation of the IAA pool size in oleanderisolates of P. syringae ssp. savastanoi and may possiblyaffect their virulence.
The absence of sequences homologous to iaaM in theDNA of ash isolates, which is in accord with the findingsof Gardan et al. (1992b), indicates that, in thesebacteria, indole-3-acetamide is not an intermediate inthe IAA biosynthetic pathway. Since an IAA subsidiarypathway, with indole-3-pyruvic acid as an intermediate,has been shown to function in P. syringae ssp. savastanoi(Surico & Iacobellis, 1992) and in Erwinia herbicola pv.gypsophylae, (Manulis et al., 1991; Clark et al., 1993),as well as in a number of non-gall-forming plant patho-genic pseudomonads and xanthomonads (Fett et al.,1987), it cannot be ruled out that the alternativepathway might be involved in IAA biosynthesis in ashisolates.
At present, the role of auxins in symptom develop-ment on ash is still unknown but, as already suggested(Surico & Iacobellis, 1992), they might be responsiblefor the formation of the wart-like excrescences. Theextensive periderm formation in diseased ash tissues alsosuggests this effect (Janse, 1982b). The availability ofmutants either lacking the ability to produce auxins orwhose auxin-producing ability had changed might allowthe role of auxin to be confirmed.
The cross-inoculation tests carried out in the presentstudy confirm that isolates from ash, olive and oleandercause different symptoms and have different host ranges.The different ability of ash, olive and oleander strains tocause symptoms on host plants was positively correlatedwith the population densities, which were higher forvirulent than for avirulent isolates. For example,oleander isolates produced population densities at least100 times higher in oleander than those reached after thesame period of time by isolates from ash and olive,which were avirulent on oleander.
These findings were confirmed when isolates of P.syringae ssp. savastanoi were cross-inoculated into hostplant leaf tissues, similarly to previous tests in whicholive strains were inoculated into oleander leaves(Mugnai et al., 1993; Iacobellis et al., 1994). The onlyexception was when bacteria were injected into oliveleaves and, surprisingly, none of the isolates was able tosurvive in them. Since under other assay conditions oliveand oleander isolates could multiply in olive leaf tissues(Surico, 1993), it is not clear why no living cells wererecovered just a few days after inoculation. Possibly,under these assay conditions, plant factors such aspolyphenols and particularly antimicrobial derivativesof the glucoside oleuropein may have been important(Fleming et al., 1973; Capasso et al., 1997).
Among the 20 ash isolates used in this study only(NCPPB3474) behaved atypically in both phytohor-mone production and pathogenicity, in which it wasidentical to typical olive isolates (Surico & Iacobellis,
Pseudomonas syringae ssp. savastanoi from ash 81
Q 1998 BSPP Plant Pathology (1998) 47, 73–83

1992). Two other isolates from ash showing thephytohormone phenotype of olive strains have beenreported (Gardan et al., 1992a); however, these isolateswere avirulent on olive and induced typical cankers onash (Gardan et al., 1992a).
The host specificity of ash, olive and oleander strainsof P. syringae ssp. savastanoi should also be taken intoaccount in the classification of this plant pathogenicbacterium. Also important in this respect is the apparentlack of cross-infections under natural conditions. Thiswas recently supported when oleander and olive plants,both showing knot disease, were growing in closeproximity or with physical contact, but no oleanderstrains were isolated from knots on olive (Caponeroet al., 1995). To date, there is only one reported isolationof a typical oleander strain from a knot on olive (Wilson& Magie, 1963; Surico et al., 1984).
During preparation of this paper, the ISPP Subcom-mitee on Taxonomy of Plant Pathogenic Bacteria at leastpartly validated new combinations of the namesproposed by Janse (1982a) for the different strains ofthe pathogen. In fact, oleander, olive and ash strains havebeen classified as Pseudomonas savastanoi pv. nerii,P. savastanoi pv. savastanoi and P. savastanoi pv. fraxini,respectively (Young et al., 1996). The above classificationreflects accurately the differences among the oleander,olive and ash strains of the pathogen in pathogenicity,phytohormone production, sugar metabolism (Wilsonet al., 1972), fatty acid profiles (Varvaro & Sasser, 1987;Janse, 1991; Wells et al., 1991), DNA fingerprints(Mugnai et al., 1994) and bacteriocin production(Iacobellis et al., 1995).
In conclusion, the findings of this study indicate thatash strains typically produce auxins, although at muchlower levels than olive and oleander strains; ash strainsproduce no cytokinins. The phytohormone productioncapability of the different strains of P. syringae ssp.savastanoi was correlated with the type of symptomscaused on host plants, i.e. canker accompanied by wart-like excrescences on ash, and knots on olive and oleander.
Acknowledgements
This work was supported by grants of the NationalResearch Council of Italy (CNR), Special Project RAISA,and CNR project NOSOG number 0664.
References
Capasso R, Cristinzio G, Evidente A, Visco C, Iannini C,1995. Oleuropein and other polyphenols from olive (Oleaeuropea) for protecting the plant against Pseudomonassyringae subsp. savastanoi. In: Rudolph K. et al. eds.Pseudomonas syringae Pathovars and Related Pathogens.Dordrecht, the Netherlands: Kluwer, 133–137.
Caponero A, Contesini AM, Iacobellis NS, 1995. Populationdiversity of Pseudomonas syringae subsp. savastanoi onolive and oleander. Plant Pathology 44, 848–55.
Clark E, Manulis S, Ophir Y, Barash I, Gafni Y, 1993.
Cloning and characterization of iaaM and iaaH fromErwinia herbicola pathovar gypsophilae. Phytopathology83, 234–40.
Cohen JD, Bandursky RS, 1982. Chemistry and physiology ofthe bound auxins. Annual Review of Plant Physiology 33,403–30.
Comai L, Kosuge T, 1982. Cloning and characterization ofiaaM, a virulence determinant of Pseudomonas savastanoi.Journal of Bacteriology 148, 40–6.
Evidente A, Di Maio E, Caponero A, Iacobellis NS, 1995.Plant growth regulators from ash strains of Pseudomonassyringae subsp. savastanoi. Experientia 51, 990–3.
Evidente A, Iacobellis NS, Sisto A, 1993. Isolation of indole-3-acetic acid methyl ester, a metabolite of indole-3-aceticacid from Pseudomonas amygdali. Experientia 49, 182–3.
Evidente A, Surico G, Iacobellis NS, Randazzo G, 1986. a-N-acetyl-indole-3-acetyl-e-lysine: a metabolite of indole-3-acetic acid from Pseudomonas syringae pv. savastanoi. Phy-tochemistry 25, 125–8.
Fett WF, Osman SF, Dunn MF, 1987. Auxin production byplant-pathogenic pseudomonads and xanthomonads.Applied and Environmental Microbiology 53, 1839–45.
Fleming HP, Walter WM, Etchells JL, 1973. Antimicrobialproperties of oleuropein and products of its hydrolysis fromgreen olives. Applied Microbiology 26, 777–82.
Fletcher RA, Kallidumbil V, Steele P, 1982. An improvedbioassay for cytokinins using cucumber cotyledons. PlantPhysiology 69, 675–7.
Gardan L, Bollet C, Abu-Ghorrah M, Grimont F, GrimontPAD, 1992a. DNA relatedness among the pathovar strainsof Pseudomonas syringae subsp. savastanoi Janse (1982)and proposal of Pseudomonas savastanoi sp.nov. Inter-national Journal of Systematic Bacteriology 42, 606–12.
Gardan L, David C, Morel M, Glickman E, Abu-Ghorrah M,Petit A, Dessaux Y, 1992b. Evidence for a correlationbetween auxin production and host plant species amongstrains of Pseudomonas syringae subsp. savastanoi. Appliedand Environmental Microbiology 58, 1780–3.
Glass NL, Kosuge T, 1988. Role of indoleacetic acid-lysinesynthetase in regulation of indoleacetic acid pool size andvirulence of Pseudomonas syringae subsp. savastanoi.Journal of Bacteriology 170, 2367–73.
Gordon SA, Weber RP, 1951. Colorimetric estimation of indo-leacetic acid. Plant Physiology 26, 192–5.
Hutzinger O, Kosuge T, 1968. 3-Indoleacetyl-L-lysine, a newconjugate of 3-indoleacetic acid produced by Pseudomonassavastanoi. In: Wightman F, Setterfield G, eds. Biochemistryand Physiology of Plant Growth Substances. Ottawa,Canada: The Runge Press.
Iacobellis NS, Contesini AM, Surico G, 1995. Bacteriocin pro-duction by Pseudomonas syringae subsp. savastanoi. Phyto-pathologia Mediterranea 34, 15–22.
Iacobellis NS, Sisto A, Surico G, 1993. Occurrence of unusualstrains of Pseudomonas syringae subsp. savastanoi on olivein central Italy. EPPO Bulletin 23, 429–35.
Iacobellis NS, Sisto A, Surico G, Evidente A, Di Maio E,1994. Pathogenicity of Pseudomonas syringae subsp.savastanoi mutants defective in phytohormone production.Journal of Phytopathology 140, 238–48.
Janse JD, 1981. The bacterial disease of ash (Fraxinusexcelsior), caused by Pseudomonas syringae subsp.
N. S. Iacobellis et al.82
Q 1998 BSPP Plant Pathology (1998) 47, 73–83

savastanoi pv. fraxini. II. Etiology and taxonomic consid-erations. European Journal of Forest Pathology 11, 425–38.
Janse JD, 1982a. Pseudomonas syringae subsp. savastanoi(ex Smith) subsp. nov., nom. rev., the bacterium causingexcrescences on Oleaceae and Nerium oleander L.International Journal of Systematic Bacteriology 32, 166–9.
Janse JD, 1982b. The bacterial disease of ash (Fraxinusexcelsior), caused by Pseudomonas syringae subsp.savastanoi pv. fraxini. III. Pathogenesis. European Journalof Forest Pathology 12, 218–31.
Janse JD, 1991. Pathovars discrimination within Pseudomo-nas syringae subsp. savastanoi using whole cell fatty acidsand pathogenicity as criteria. Systematic and AppliedMicrobiology 13, 79–84.
King EO, Ward MK, Raney DE, 1954. Two simple media forthe demonstration of pyocyanin and fluorescein. Journal ofLaboratory and Clinical Medicine 44, 301–7.
Lelliott RA, Stead DE, 1987. Methods for the diagnosis ofbacterial diseases of plants. In: Preece TF, ed. Methods inPlant Pathology, Vol. 2. Oxford, UK: Blackwell ScientificPublications.
Maniatis T, Fritsh EF, Sambrook J, 1982. Molecular Cloning:A Laboratory Manual. Cold Spring Harbor, New York:Cold Spring Harbor Laboratory.
Manulis S, Gafni Y, Clark E, Zutra D, Ophir Y, 1991.Indole-3-acetic acid biosynthetic pathways in Erwiniaherbicola in relation to pathogenicity on Gypsophila pani-culata. Physiological and Molecular Plant Pathology 39,161–71.
Mugnai L, Giovannetti L, Ventura S, Surico G, 1994. Thegrouping of strains of Pseudomonas syringae subsp.savastanoi by DNA restriction fingerprinting. Journal ofPhytopathology 142, 209–18.
Mugnai L, Surico G, Iacobellis NS, 1993. Responseof explants and cultured cells of oleander to inoculationwith strains of Pseudomonas syringae subsp. savastanoi.Petria 3, 27–39.
Palm CJ, Gaffney T, Kosuge T, 1989. Cotranscription of genesencoding indoleacetic acid production in Pseudomonas syr-ingae subsp. savastanoi. Journal of Bacteriology 171,1002–9.
Powell GK, Morris RO, 1986. Nucleotide sequence andexpression of a Pseudomonas savastanoi cytokinin bio-synthetic gene: homology with Agrobacterium tumefacienstmr and tzs loci. Nucleic Acids Research 6, 225–35.
Surico G, 1993. Symptom development in olive and oleanderleaves inoculated with Pseudomonas syringae subsp.
savastanoi and scanning electron microscopy of infections.Petria 3, 117–27.
Surico G, Comai L, Kosuge T, 1984. Pathogenicity of strainsof Pseudomonas syringae pv. savastanoi and theirindoleacetic acid-deficient mutants on olive and oleander.Phytopathology 74, 490–3.
Surico G, Iacobellis NS, 1992. Phytohormones and olive knotdisease. In: Verma DPS, ed. Molecular Signals in Plant–Microbe Communications. Boca Raton: CRC Press,209–27.
Surico G, Iacobellis NS, Sisto A, 1985. Studies on the role ofindole-3-acetic acid and cytokinins in the formation ofknots on olive and oleander plants by Pseudomonassyringae pv. savastanoi. Physiological Plant Pathology 26,309–20.
Varvaro L, Sasser M, 1987. Fatty acid profiles of Pseudo-monas syringae pv. savastanoi. In: Proceedings of the 3rdInternational Working Group on Pseudomonas syringaepathovars, 1987. Portugal.
Wells JM, Casano FJ, Surico G, 1991. Fatty acid compositionof Pseudomonas syringae pv. savastanoi. Journal of Phyto-pathology 133, 152–62.
Wilson EE, Heskett MG, Johnson L, Kosuge T, 1972. Meta-bolic behavior of Pseudomonas savastanoi isolates fromolive and oleander on certain carbohydrate and aminosubstrates. Phytopathology 62, 349–55.
Wilson EE, Magie AR, 1963. Physiological, serological, andpathological evidence that Pseudomonas tonelliana isidentical with Pseudomonas savastanoi. Phytopathology53, 653–9.
Yamada T, Palm CJ, Brooks B, Kosuge T, 1985. Nucleotidesequence of the Pseudomonas savastanoi indoleacetic acidgenes and homology with Agrobacterium tumefaciensT-DNA. Proceedings of the National Academy of Sciences-USA 82, 6522–6.
Young JM, Bradbury JF, Davis RE, Dickey RS, Ercolani GL,Hayward AC, Vidaver AK, 1991. Nomenclatural revisionsof plant pathogenic bacteria and list of names 1980–1988.ISPP Subcommittee on Taxonomy of PhytopathogenicBacteria. Review of Plant Pathology 70, 211–21.
Young JM, Dye DW, Bradbury JF, Panagopoulos CG, RobbsCF, 1978. A proposed nomenclature and classification forplant pathogenic bacteria. New Zealand Journal ofAgricultural Research 21, 153–77.
Young JM, Saddler GS, Takikawa Y, Deboer SH, Vauterin L,Gardan L, Gvozdyak RI, Stead DE, 1996. Names of plantpathogenic bacteria 1864–1995. ISPP Subcommittee onTaxonomy of Phytopathogenic Bacteria. Review of PlantPathology 75, 721–63.
Pseudomonas syringae ssp. savastanoi from ash 83
Q 1998 BSPP Plant Pathology (1998) 47, 73–83
Related Documents











