Mycol. Res. 96 (7), 555-562 (1992) Printed in Great Britain Characterization of morphological mutants generated spontaneously in glucose-limited, continuous flow cultures of Fusarium graminearum A3/5 MARIL YN G. WIEBE, GEOFFREY D. ROBSON, ANTHONY P. J. TRINeI Microbiology Research Group, Department of Cell and Structural Biology, School of Biological Sciences, Stopford Building, University of Manchester, Manchester M13 9PT, u.K. STEPHEN G. OLIVER Manchester Biotechnology Centre, UMIST. p.o. Box 88, Manchester M60 lQD 555 An analysis was made of the morphology (mycelial branching and hyphal diameter) and growth (specific growth rate and colony radial growth rate) of 20 strains of morphological mutants which arose spontaneously in prolonged, glucose-limited, continuous flow cultures of Fusan"um graminearum A 3/5. All the morphological mutants formed mycelia which were more highly branched than the wild type, and produced colonies which expanded in radius more slowly than the wild type. All the mutants were recessive to the wild type and were assigned to three complementation groups. Analysis of the morphology of colonies produced by macroconidia harvested from heterokaryons formed between pairs of morphological mutants indicated that the ratio of the mutant nuclei present in the heterokaryons varied between wide limits, and did not generally reflect the initial ratio of the two types of macroconidia in the inoculum from which the heterokaryons arose. The reason why some mutants had, compared to the parental strain, a selective advantage in glucose-limited, continuous flow culture is not known, but no evidence was obtained that this advantage was correlated with increased specific growth rate. Filamentous micro-organisms, such as fungi and actino- mycetes, exhibit a high degree of genetic instability which may compromise industrial-scale processes, such as antibiotic production (Righelato, 1976; Hardy & Oliver, 1985). It is unlikely that these organisms have a higher intrinsic level of genetic, and therefore morphological, variation than other microbes, rather it is probable that a large number of genes determine morphology and that mutations in anyone gene may have a profound effect on the fitness of the organism, especially in suspension culture. Gamjobst & Tatum (1967) screened 90 morphological mutants of Neurospora crassa and assigned them to 58 loci in seven linkage groups. These mutants were isolated mainly after uv irradiation of conidia and would thus be expected to form a representative cross- section of the morphological mutations potentially available. The study demonstrated that a large number of genes were involved in determining mycelial morphology and showed that they were sometimes clustered in specific regions of the chromosomes. Many morphological mutants of N. crassa are described as , colonial' because their mycelia are more branched than those of the wild type and because they form colonies which expand in radius at a much slower rate than wild-type colonies (Gamjobst & Tatum, 1967; Trinci, 1973a, b). By mixing the spores of a colonial mutant with the wild-type spores in various ratios, Barratt & Gamjobst (1949) found that the mutation was recessive and the growth rate of the heterokaryon was dependent on the ratio of nuclear types contributed by the two parental strains. Dodge (1942) demonstrated that complementation occurred between two colonial mutants of N. crassa, with the heterokaryon having the wild-type phenotype. However, heterokaryons between complementing biochemical mutants of N. crassa did not always form colonies with wild-type extension rates (measured in growth tubes), and the extension rate of the heterokaryon was influenced by the ratio of the mutant nuclei present (Pittenger & Atwood, 1954). Selection of morphological mutants has been observed during continuous flow cultures of fungi (Trinci et aI., 1990). These mutants increase rapidly in the population (i.e. they have a selective advantage), eventually replacing the parental strain (Wiebe el al., 1991). As there are a large number of genes that affect mycelial morphology (Gamjobst & Tatum, 1967), there is a relatively high probability that random mutations occurring during prolonged continuous flow culture will affect morphology, and under non-selective conditions these morphological mutants will be easier to recognize than biochemical mutations. However, although the existence of a large number of genes affecting mycelial morphology explains why morphological mutants frequently arise in chemos tat cultures of fungi (Trinci el al., 1990), it does not explain why some mutants have a selective advantage over their parental strains. Morphological mutants isolated from continuous-flow cultures of Penicillium chrysogenum (Righelato, 1976),

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Mycol. Res. 96 (7), 555-562 (1992) Printed in Great Britain
Characterization of morphological mutants generatedspontaneously in glucose-limited, continuous flow cultures ofFusarium graminearum A3/5
MARIL YN G. WIEBE, GEOFFREY D. ROBSON, ANTHONY P. J. TRINeI
Microbiology Research Group, Department of Cell and Structural Biology, School of Biological Sciences, Stopford Building, University ofManchester, Manchester M13 9PT, u.K.
STEPHEN G. OLIVER
Manchester Biotechnology Centre, UMIST. p.o. Box 88, Manchester M60 lQD
555
An analysis was made of the morphology (mycelial branching and hyphal diameter) and growth (specific growth rate and colonyradial growth rate) of 20 strains of morphological mutants which arose spontaneously in prolonged, glucose-limited, continuous flowcultures of Fusan"um graminearum A 3/5. All the morphological mutants formed mycelia which were more highly branched than thewild type, and produced colonies which expanded in radius more slowly than the wild type. All the mutants were recessive to thewild type and were assigned to three complementation groups. Analysis of the morphology of colonies produced by macroconidiaharvested from heterokaryons formed between pairs of morphological mutants indicated that the ratio of the mutant nuclei presentin the heterokaryons varied between wide limits, and did not generally reflect the initial ratio of the two types of macroconidia inthe inoculum from which the heterokaryons arose. The reason why some mutants had, compared to the parental strain, a selectiveadvantage in glucose-limited, continuous flow culture is not known, but no evidence was obtained that this advantage was correlatedwith increased specific growth rate.
Filamentous micro-organisms, such as fungi and actinomycetes, exhibit a high degree of genetic instability whichmay compromise industrial-scale processes, such as antibioticproduction (Righelato, 1976; Hardy & Oliver, 1985). It isunlikely that these organisms have a higher intrinsic level ofgenetic, and therefore morphological, variation than othermicrobes, rather it is probable that a large number of genesdetermine morphology and that mutations in anyone genemay have a profound effect on the fitness of the organism,especially in suspension culture. Gamjobst & Tatum (1967)screened 90 morphological mutants of Neurospora crassa andassigned them to 58 loci in seven linkage groups. Thesemutants were isolated mainly after uv irradiation of conidiaand would thus be expected to form a representative crosssection of the morphological mutations potentially available.The study demonstrated that a large number of genes wereinvolved in determining mycelial morphology and showedthat they were sometimes clustered in specific regions of thechromosomes.
Many morphological mutants of N. crassa are described as,colonial' because their mycelia are more branched than thoseof the wild type and because they form colonies which expandin radius at a much slower rate than wild-type colonies(Gamjobst & Tatum, 1967; Trinci, 1973a, b). By mixing thespores of a colonial mutant with the wild-type spores invarious ratios, Barratt & Gamjobst (1949) found that themutation was recessive and the growth rate of the
heterokaryon was dependent on the ratio of nuclear typescontributed by the two parental strains. Dodge (1942)demonstrated that complementation occurred between twocolonial mutants of N. crassa, with the heterokaryon havingthe wild-type phenotype. However, heterokaryons betweencomplementing biochemical mutants of N. crassa did notalways form colonies with wild-type extension rates (measuredin growth tubes), and the extension rate of the heterokaryonwas influenced by the ratio of the mutant nuclei present(Pittenger & Atwood, 1954).
Selection of morphological mutants has been observedduring continuous flow cultures of fungi (Trinci et aI., 1990).These mutants increase rapidly in the population (i.e. theyhave a selective advantage), eventually replacing the parentalstrain (Wiebe el al., 1991). As there are a large number ofgenes that affect mycelial morphology (Gamjobst & Tatum,1967), there is a relatively high probability that randommutations occurring during prolonged continuous flow culturewill affect morphology, and under non-selective conditionsthese morphological mutants will be easier to recognize thanbiochemical mutations. However, although the existence of alarge number of genes affecting mycelial morphology explainswhy morphological mutants frequently arise in chemostatcultures of fungi (Trinci el al., 1990), it does not explain whysome mutants have a selective advantage over their parentalstrains. Morphological mutants isolated from continuous-flowcultures of Penicillium chrysogenum (Righelato, 1976),

Morphological mutants of Fusarium
Paecilomyces puntonii, P. variotii, Gliocladium virens, Trichodermaviride and Byssochlamys nivea (Forss ef al., 1974) had specificgrowth rates which were greater than their parental strain, andthis explains their selective advantage. However, the selectiveadvantage of morphological mutants of Fusarium graminearumA 3/5 (Trinci et al., 1990; Wiebe et al., 1991) over the wildtype is not known.
Wiebe et a!. (1991) reported the isolation of 18 morphological mutants generated spontaneously in glucoselimited continuous flow cultures of F. graminearum A 3/5.These mutants were distinguished from each other on thebasis of differences in colony morphology and colony radialgrowth rate. As they were isolated from the same highlyselective culture condition (glucose limitation), it was expectedthat only a small proportion of the morphology-determininggenes might be represented; these might be mutants in whichthe changed morphology was linked to an increased specificgrowth rate, to a reduced saturation constant (Ks ) value forglucose, or to increased fragmentation (Wiebe ef al., 1991).
Marlow Foods grows F. graminearum in continuous flowculture to produce QUORN'i\) mycoprotein for humanconsumption (Trinci et al., 1990; Trinci, 1992). The appearanceof morphological mutants during the course of the fermentation precipitates the premature termination of theculture and subsequent loss in production. Thus an understanding of the nature of these mutations, their selectiveadvantage and genetic variation could yield information ofvalue for the control of the industrial fermentation of F.graminearum. In this paper 20 morphological mutants,generated spontaneously during long-term glucose-limited,continuous flow cultures, are described, and assigned to threecomplementation groups.
MATERIALS AND METHODS
Strains and media
Fusarium graminearum Schwabe A 3/5 was obtained from MrT. W. Naylor, Marlow Foods, Billingham, U.K. The colonialstrains were isolated from prolonged, glucose-limited, continuous flow cultures of F. graminearum A 3/5 (Wiebe et a!',1991). Strains were isolated between 220 and 600 h fromonset of steady state in three separate chemostat experiments(designated CCl, CC2 and MC3). Strain numbers indicate theexperiment (e.g. CCl) and order of isolation (e.g. CCl-l, CCI2, CCI-3, etc.), which was not necessarily the order in whichmutants appeared in the population. Macroconidia of allstrains were stored at -70°C in 20% (vIv) glycerol solution.
The defined medium of Vogel (1956) was used withglucose (10 g 1- 1
) as the carbon source instead of sucrose.Vogel's mineral salts solution was prepared at x 50 finalconcentration, sterilized by membrane (0'2 >Am diam.) filtrationand added to the sterile glucose solution (autoclaved at 121°for 15 min). In some experiments, glutamine, NaNOa,(NH4)2S04 (all at 2 g 1-1), or glutamine plus NaNOa (eachat 1 g 1-1) was substituted for the nitrogen source(2 g NH4NOa l-1
) in Vogel's medium, and sometimes themedium was solidified with agar (Davis Gelatine; 15 g I-I,final concentration).
556
Culture conditions
Batch liquid cultures of F. graminearum were grown in 50 mlvolumes of liquid medium in 250 ml Ehrlenmeyer flasks. Eachflask was inoculated with ca 1 X 106 spores suspended indistilled water. Cultures were incubated on a rotary shaker(with a throw of 2'5 cm) at 200 rpm.
Colonies were grown in 9 em diam. Petri dishes containing20 ml agar medium, which were inoculated with either a smalldrop of spore suspension (homokaryons) or 4 mm diam. agarplugs (homokaryons and heterokaryons) taken from thegrowing colony margin. All cultures were incubated at 25°.
Measurements of fungal growth and morphology
Growth of liquid cultures was measured in batch culture byincrease in optical density at 420 nm using an SP 8-100ultraviolet spectrophotometer (12'5 x 45 mm cell, PyeUnicam, Cambridge). Measurements of colony diameters weremade with a rule at x 10 magnification using a Shadowmaster(Baty & Co.).
Measurements of hyphal growth unit length (a measure ofmycelial branching; Trinci, 1973 b) and hyphal diameter weremade using a MeasureMouse computer system (AnalyticalMeasuring Systems, Pampisford, Cambridge) as described byWiebe & Trinci (1991), on mycelia which had been incubatedfor ca 20 h in liquid medium in batch culture. For hyphalgrowth unit lengths, 25 mycelia with 5 or more hyphal tipswere measured at final magnifications of x 140, x 350 orx 1440, as appropriate. A minimum of 50 hyphal diameters
(measured at x 1440 magnification) were measured for eachstrain. Tests for significant differences were made usingScheffe's multiple-range test.
Heterokaryon formation
As wild-type colonies expand in radius at a faster rate thanthose of colonial mutants, a simple complementation test wasdevised, based on the fact that the heterokaryons ofcomplementing colonial mutants which had wild-type or nearwild-type phenotype could be distinguished from the twoparents as one or more fast-growing sectors (Fig. 1). In thecomplementation test, a small drop of spore suspension fromone colonial mutant was inoculated onto agar mediumimmediately adjacent to, but not touching, a drop of similarsize from another colonial mutant. The drops were allowed todry and the plates were incubated for 5-7 days at 25°. As themycelia grew, heterokaryons developed along the interfacebetween the two mycelia and emerged as fast-groWing sectorson either side of the two parental colonies (Fig. 1) or as adistinct tuft along the interface between the two mutants.Each parental colony was also inoculated adjacent to itself asa control. Up to three crosses were made using one Petri dish(Fig. 1). All crosses were tested on glutamine-, sodium nitrate-,and/or glutamine plus sodium nitrate-containing media, andapparent heterokaryons were subcultured on to Vogel'smedium containing ammonium nitrate and allowed to growfor 6-8 days. For measurements of colony radial growth rates(Kr ) of heterokaryons, a 4 mm diam. cork borer was used tocut plugs from the margin of the heterokaryon and transferred
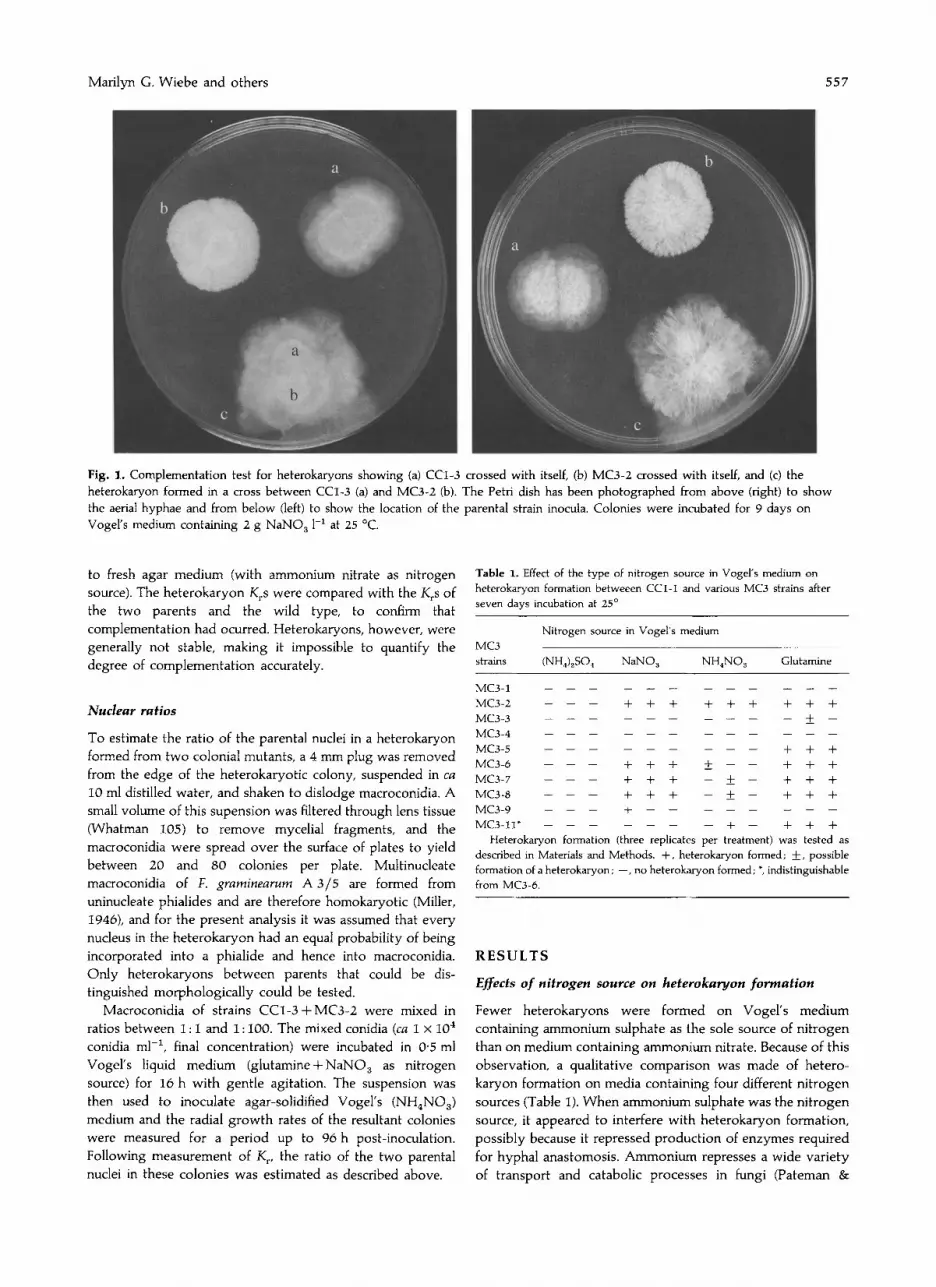
Marilyn G. Wiebe and others 557
Fig. 1. Complementation test for heterokaryons showing (a) CCl-3 crossed with itself, (b) MC3-2 crossed with itself, and (c) theheterokaryon formed in a cross between CCl-3 (a) and MC3-2 (b). The Petri dish has been photographed from above (right) to showthe aerial hyphae and from below (left) to show the location of the parental strain inocula. Colonies were incubated for 9 days onVogel's medium containing 2 g NaN03 1-1 at 25°C.
Nitrogen source in Vogel's medium
Table 1. Effect of the type of nitrogen source in Vogel's medium onheterokaryon formation betweeen CCl·l and various MC3 strains afterseven days incubation at 25°
to fresh agar medium (with ammonium nitrate as nitrogensource). The heterokaryon Krs were compared with the Krs ofthe two parents and the wild type, to confirm thatcomplementation had ocurred. Heterokaryons, however, weregenerally not stable, making it impossible to quantify thedegree of complementation accurately.
MC3strains Glutamine
RESULTS
++±
+
+ + ++ + ++ + ++ + +
±±
±
+ + +++ +
Effects of nitrogen source on heterokaryon formation
Fewer heterokaryons were formed on Vogel's mediumcontaining ammonium sulphate as the sole source of nitrogenthan on medium containing ammonium nitrate. Because of thisobservation, a qualitative comparison was made of heterokaryon formation on media containing four different nitrogensources (Table I). When ammonium sulphate was the nitrogensource, it appeared to interfere with heterokaryon formation,possibly because it repressed production of enzymes requiredfor hyphaI anastomosis. Ammonium represses a wide varietyof transport and catabolic processes in fungi (Pateman &
MO·lMO-2MC3-3MC3-4MC3-5MC3-6 + + +MO-7 + + +MO-8 + + +MC3-9 +MO·U' + + + +
Heterokaryon formation (three replicates per treatment) was tested asdescribed in Materials and Methods. +. heterokaryon formed: ±, possibleformation of a heterokaryon; -, no heterokaryon formed; " indistinguishablefrom MC3-6.
To estimate the ratio of the parental nuclei in a heterokaryonformed from two colonial mutants, a 4 mm plug was removedfrom the edge of the heterokaryotic colony, suspended in ca
10 ml distilled water, and shaken to dislodge macroconidia. Asmall volume of this supension was filtered through lens tissue(Whatman 105) to remove mycelial fragments, and themacroconidia were spread over the surface of plates to yieldbetween 20 and 80 colonies per plate. Multinucleatemacroconidia of F. graminearum A 3/5 are formed fromuninucleate phialides and are therefore homokaryotic (Miller,1946), and for the present analysis it was assumed that everynucleus in the heterokaryon had an equal probability of beingincorporated into a phialide and hence into macroconidia.Only heterokaryons between parents that could be distinguished morphologically could be tested.
Macroconidia of strains CCI-3 +MC3-2 were mixed inratios between I: I and I: 100. The mixed conidia (ca I X 104
conidia ml-\ final concentration) were incubated in 0'5 mlVogel's liqUid medium (glutamine +NaN0 3 as nitrogensource) for 16 h with gentle agitation. The suspension wasthen used to inoculate agar-solidified Vogel's (NH4N03 )
medium and the radial growth rates of the resultant colonieswere measured for a period up to 96 h post-inoculation.Following measurement of Kr, the ratio of the two parentalnuclei in these colonies was estimated as described above.
Nuclear ratios
Morphological mutants of Fusarium 558
Table 2. Hyphal growth unit length (G), hyphal diameter, hyphal growth unil volume (G v,") and specific growth rate (11) of morphological mutants ofFusarium graminearum A 3/5 grown at 25° in shake flask culture on Vogel's medium containing 2 g NH,NO, 1-1
Morphologytype Strain
Specificgrowth rate(11, h-1)'
Hyphalgrowth unitlength(G,l1m)'
Hypha!diameter(11m)'
Hyphalgrowth unitvolume(G,.Ol' 11m')
S90
167156226363
161194530
23227S
2692674182763.2S
411.276385
661436.284819
l'6±0-OS
2-2±O'll3-0±O'OIS
l'7±O'08*1-8±0-OS*
1'9 ± 0-09*1-8±0-07*2-2±0-102-0±O'092-1 ±O-1O2-2±0-102'0±0-12*2-4±0-1O
1'8 ± 0'083-9±0-293'O±O'143-7±0-174'3±0-lS
2-2±0-212'l±O'17*1-7±0-1l*2-8±0-12
Wild typeA
c
B
D
A 3/5 0'23±0'008 232±ll'tMO-! 0'18±0'007 14±1"MO-3 0'22±0'019* 22±PCCl-I 0'23±0'01l* 21± l'
CC2-2 0'19±0'019 25±P
MO-2 0'23±0'008* 80±3h
MO-7 0'22±0'007* Sl±3b
CCl-2 0'23±0'012* 7S±2b
MC3-4 0'23 ± O'OlM 102 ± 4d «-)
MC3-5 0'23±0'01M 108±3''''')MO-6 0'22±0'010* 9S±3'
MC3-8 0-24 ± 0'012* 1OS±2d")MC3-9 0-24±0'009* 1l0±3d
MO-ll§ 0-23±0-01l* 88±3'CC1-S 0-23±0-008* 94±4"d)CCl-9 0-24±0-01l* 108±3d
CC2-1 0-23±0'007* 88±J<CC2-3 0-25 ± 0-012* 8S ± 3'"
CCl-3 0-24±0-012* 174±13'CCI-4 0-22±0-OO7* 126±seCCl-7 0-2S±0-OO2* 12S±6'
CCl-8 0'21±0'013* 133+5'• Mean (± S_E.) of 4, 2S and so replicates for 11, G and hyphal diam_ respectively.t Values in a column with the same superscript (a-f) are not significantly different (P < O-OS, Scheffe's multiple-range test).* Not significantly different (P < O-OS) from A 3/S.
§ Indistinguishable from MC3-6.
Kinghorn, 1976) including the production of extracellularproteases (Cohen, 1972), It was concluded that glutamine wasthe best nitrogen source for heterokaryon formation in F.graminearum and that crosses which did not complement onglutamine-containing medium should also be tested on sodiumnitrate-containing medium_ Consequently, all crosses weretested on glutamine-, sodium nitrate-, and/or glutamine plussodium nitrate-containing media_
Growth and morphology of colonial mutants
Table 2 gives the hyphal growth unit length (G), diameter,hyphal growth unit volume (GyO]' Trinci, 1984), and specificgrowth rate (1-1) of 2I isolates of morphological mutants of F.graminearum A 3/5; these represented 20 different strains_
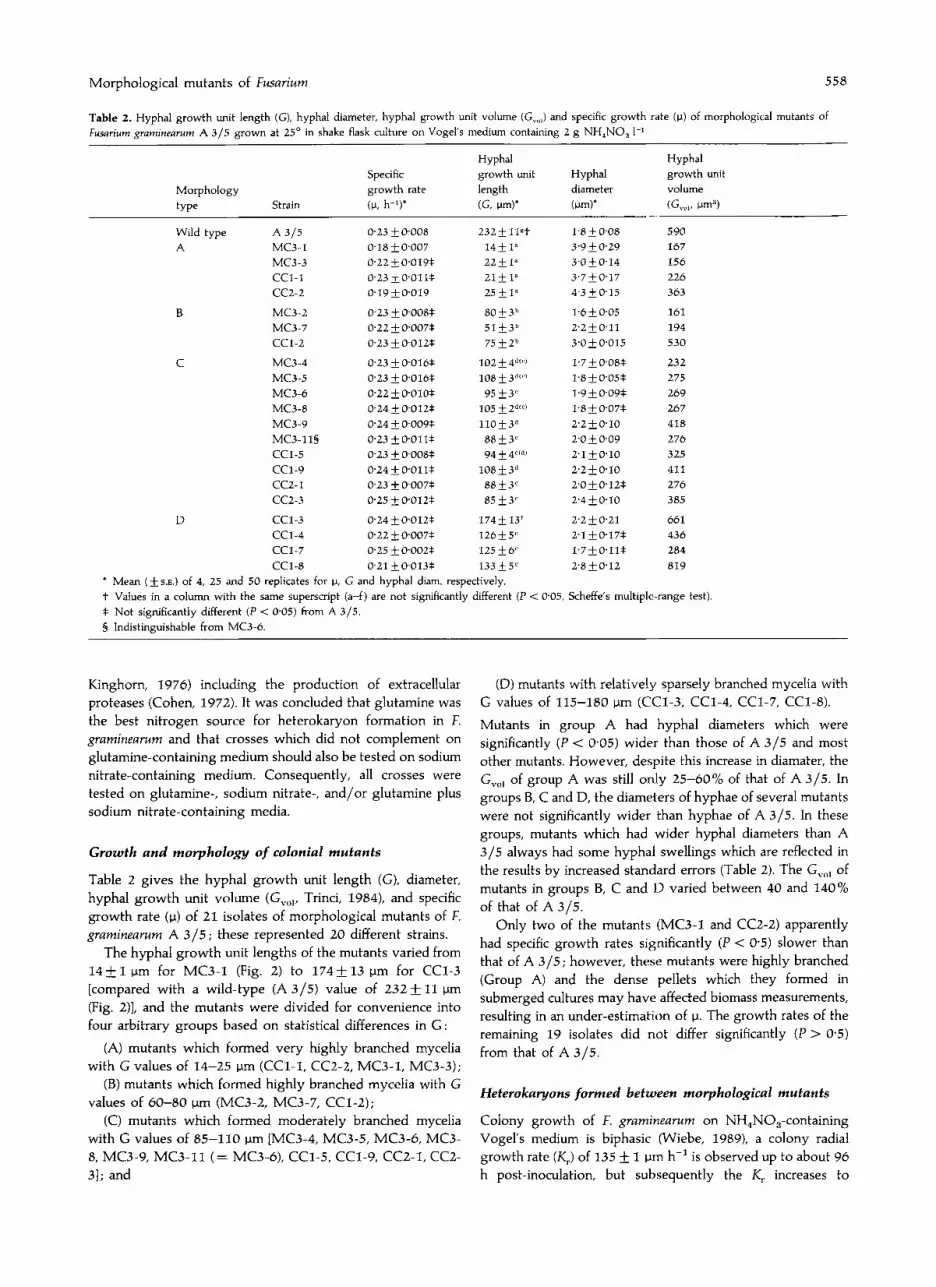
The hyphal growth unit lengths of the mutants varied from14 ±1 I-Im for MC3-1 (Fig, 2) to 174 ±13 I-Im for CCI-3[compared with a wild-type (A 3/5) value of 232 ± 11 I-Im(Fig_ 2)], and the mutants were divided for convenience intofour arbitrary groups based on statistical differences in G:
(A) mutants which formed very highly branched myceliawith G values of 14-25 I-Im (CCI-I, CC2-2, MC3-I, MD-3);
(B) mutants which formed highly branched mycelia with Gvalues of 60-80 I-Im (MC3-2, MC3-7, CCI-2);
(C) mutants which formed moderately branched myceliawith G values of 8S-IIO I-Im [MC3-4, MC3-5, MC3-6, MC38, MD-9, MO-II (= MO-6), CCI-5, CCI-9, CCl-l, CCl3]; and
(D) mutants with relatively sparsely branched mycelia withG values of II5-180 I-Im (CCI-3, CCI-4, CCI-7, CCl-8)_
Mutants in group A had hyphal diameters which weresignificantly (P < 0'05) wider than those of A 3/5 and mostother mutants. However, despite this increase in diamater, theGY01 of group A was still only 25-60% of that of A 3/5_ Ingroups B, C and D, the diameters of hyphae of several mutantswere not significantly wider than hyphae of A 3/5. In thesegroups, mutants which had wider hyphaI diameters than A3/5 always had some hyphaI swellings which are reflected inthe results by increased standard errors (Table 2)_ The G"O) ofmutants in groups B, C and D varied between 40 and 140 %of that of A 3/5.
Only two of the mutants (MC3-1 and CC2-2) apparentlyhad speCific growth rates significantly (P < 0'5) slower thanthat of A 3/5; however, these mutants were highly branched(Group A) and the dense pellets which they formed insubmerged cultures may have affected biomass measurements,resulting in an under-estimation of 1-1. The growth rates of theremaining 19 isolates did not differ significantly (P> 0'5)from that of A 3/5.
Heterokaryons formed between morphological mutants
Colony growth of F. graminearum on NH4N03-containingVogel's medium is biphasic (Wiebe, 1989), a colony radialgrowth rate (Kr ) of 135 ± 1 I-Im h- 1 is observed up to about 96h post-inoculation, but subsequently the Kr increases to
Marilyn G. Wiebe and others 559
CCI-2
MC3-9
MC3-?0----------<
MC3-8
CC2-2CCI-S MC3-SI I"""""""
MC3-6 (MC3-ll) CC 1-9I I t--------t
CC2-1
CCI-8
Fig. 3. Complementation map (based on the data in Table 3) for 21isolates of morphological mutants of F. gmminearumA 3/5 generatedspontaneously in glucose-limited continuous flow culture.
mentation data is given in Fig. 3. The mutants were dividedinto 17 groups, several of which were overlapping, suggestingthat allelic complementation occurred. Thus, as shown in Fig. 3,it appears that mutations in only three complementationgroups (I to III) accounted for the genetic variation observedbetween the 21 isolates. Complementation groups I and II wereeach represented by only one mutant (MC3-9 and CCl - 2respectively), and these mutants complemented all otherstrains. Complementation group III contained 19 isolates andwas non-linear. Of the 19 isolates in complementation GroupIII, two (MC3-6 and MC3-11) isolated from the sameexperiment showed identical complementation patterns and,as they were also morphologically very similar and had thesame growth kinetics (Table 2), it was concluded that the samemutant had been isolated twice; subsequently, both MC3-6and MC3-11 are referred to as MC3-6. Two other mutants
Complementation group II
Complementation group III
Complementation group I
50~m
Fig, 2, Tracings of mycelial fragments of CCI-I, MC3-1 and A3/5.Cultures were grown in liquid Vogel's medium for ca 20 h at 25°.
205 ± 3 ~m h-1. The reason for this change in Kr is notknown, but because of it, Kr measurements in the presentstudy were made from early (up to fa 96 h post-inoculation)colony growth.
The complementation pattern of the 21 isolates is shown inTable 3. Colony radial growth rates of parent and heterokaryotic strains are indicated where data are available. The Krsof the mutants varied from 18 ± 2 ~m h-1 (CC1-l) to 105 ±2 ~m h-1 (CCl-8). The map prepared from the comple-
Table 3. Complementation pattern for 21 morphological mutants of F. graminearumA 3/5 and colony radial growth rates (at 25° on Vogel's mediumcontaining 2 g NH.N03 1-1 of the heterokaryons/parental strains, where data are available; K, for A3/5 = 135 ± 1~ h-1 (e = complements, 0 =does not complement, • = indistinguishable from MC3-6)
MO-l MO-2 MC3-3 MO-4 MO-5 MO-6 MO-7 MO-8 MO-9 MO-11' CCI-1 CO-2 CO-3 CCl-4 CO-5 CO-7 CCl-8 CCl-9 CO-l CO-2 CO-3
MO-l 20 • 0 0 0 • • • • • 0 • 0 0 • 0 0 0 0 0 •MO-2 107 41 • • • • • • • • • • • • • • • • • 0 •MO-3 125 76 • • • • • • • 0 • 0 0 • 0 • • • • •MO-4 135 103 63 • • • • • • • • • • • • • • • 0 •MO-5 124 110 138 69 • • • • • • • • • • • 0 0 0 • •MO-6 142 183 111 133 95 58 • • • 0 • • • • 0 • 0 • • • •MO-7 111 103 133 131 131 133 81 0 • • • • • • • 0 • • • • •MO-8 82 133 104 133 143 115 48 • • • • • • • 0 • • • • •MO-9 151 107 142 153 168 195 138 16S 83 • • • • • • • • • • • •MO-11' 102 129 127 139 132 132 95 170 59 • • • • 0 • 0 • • • •CO-l 117 136 108 180 101 137 135 94 18 • 0 0 • 0 • • • • •CCl-2 123 129 101 126 122 131 130 135 183 129 88 • • • • • • • • •CCI-3 124 103 139 122 138 86 114 129 12S 56 0 • 0 • • • • •CCl-4 114 112 105 144 139 126 120 12S 131 68 • 0 • • • • •CO-5 92 93 117 120 120 lOS 157 124 112 134 79 119 61 • 0 0 0 • •CO-7 112 150 91 122 115 111 122 106 73 • • • 0 •CCl-8 138 145 154 117 161 138 139 129 139 127 105 0 0 0 •CCl-9 130 136 128 123 145 163 129 123 122 128 122 139 111 87 0 • •CC2-1 123 128 138 154 117 140 135 108 137 137 128 140 79 0 •CO-2 189 114 137 145 180 136 151 110 113 82 153 161 124 26 0CO-3 116 96 116 120 98 134 134 108 186 125 116 157 114 118 118 199 101 99 44

Morphological mutants of Fusarium 560
Table 4. Ratio or colonies produced or parental strains A and B. whichdevelop from the conidia collected from A/B heterokaryons
Ratio of parental macroconidia in inoculumfrom which heterokaryon «) has formed
Fig. 4. Radial growth rates (±S.E.) of: CCl-3 (0) and MC3-2 (e)
and heterokaryons «)) formed when macToconidia of both strainswere mixed in the inoculum in the ratio of CCl-3 to MC3-2 indicatedalong the x axis. Ratios above the heterokaryon colony radial growthrates indicate the ratio of macroconidia of each produced from 7-dayold heterokaryons. Colonies were grown for 96 h on modifiedVogel's medium containing 2 g NH4NOs 1-1 at 25° and there were6 replicates for each Kr .
1 :2
1:510: 1
Ratio of coloniesA:B fromheterokaryons
CCJ-2CCJ-9CC2-2
Strain B
150
CCJ-lCCJ-lCCl-l
Strain A
'";'c<1
125U:::E ,~
<? ..c:E
U 6 100U '"'- '"·20 0en -03 u 751: u
..<:: .~
~ ~2 ..><ell e
~til '".,::a ..c:
MC3-2'" -0.... c.... '" 25Q0
"0u
0 II-----'a:O 100:1 10:1 1:1 1:10 1:100 O:a
a wild-type phenotype or only a partly wild-type phenotype.Barratt & Garnjobst (1949) confirmed that in heterokaryonsformed between N. crassa wild-type and a colonial mutant (Y8743), where the colonial strain contributed more nuclei thanthe wild type, the growth rate (hyphaI extension in a growthtube) of the resultant heterokaryon was slower than that of thewild type and was dependent on the ratio of the wild type andmutant conidia in the inoculum. Using biochemical mutants ofN. crassa, Pittenger & Atwood (1954) found that the nuclearratio in a heterokaryon often remained constant, even thoughits extension rate in growth tubes was below the maximumpossible for heterokaryons formed between the particularmutants.
Assuming that each nuclear type has an equal probability ofentering a phialide and thus producing macroconidia, theratio of parental colony types arising fom macroconidiacollected from a heterokaryon will reflect the ratio of parentalnuclei in the heterokaryon. Collecting macroconidia fromheterokaryons and germinating them on agar mediumdemonstrated that the ratios of the parental nuclei present inthe heterokaryons varied within wide limits (Table 4). Evengreater variation was observed in the nuclear ratios of
DISCUSSION
The data in Table 3 demonstrate that for each of the 21morphological mutants isolated (20 strains), the mutant allelewas recessive to the wild-type allele, as each of the mutantsformed a wild-type heterokaryon with at least one othermutant. Further, where tested, heterokaryons formed betweencolonial mutants and A 3/5 had wild-type phenotypes (M. G.Wiebe, unpublished results). However, not all heterokaryonsformed between pairs of morphological mutants (Table 3)exhibited a full wild-type growth rate (Kr ) over the incubationperiod tested. Beadle & Coonradt (1944) suggested that theratio of nuclei contributed to a heterokaryon from eachmutant would determine whether the heterokaryon exhibited
(MC3-3 and CCl-l) had the same phenotype; both formedvery dense mycelia with no aerial hyphae, and both showeda high degree of reversion to less dense mycelia, with tuftedaerial hyphae similar to those of CCI-3, CCI-4 and CCI-7.Therefore, these two mutants, isolated from separate experiments, are probably also genotypically the same. CCI-3, CCl4 and CCI-7 differed only in the relative sparseness of theirbranching and the abundance of aerial hyphae they producedon agar-solidified medium. In liquid culture they all hadhyphal growth unit lengths greater than 120 I-Im (Group D).No allelic complementation was observed between these fivestrains.
With the exception of MC3-8, which did not complementCCI-7, mutants with wild-type aerial hyphae were clusteredtogether at one end of the map. Most of these mutants weremoderately branched in liquid culture (Group C).
Nuclear ratios in heterokaryons formed betweenmorphological mutants
Macroconidia were taken from three heterokaryons each ofwhich contained CCl-l as one parent, because it is a strainwhich can easily be distinguished from most other morphological strains of F. graminearum. The colonies whichdeveloped from these macroconidia indicated that, in theheterokaryons, the nuclear ratios of the parental strains varied(approximately) from 1:2 to 10: 1 (Table 4).
Figure 4 shows the colony radial growth rates observed forheterokaryotic colonies formed from mixed spore inocula ofCCI-3 +MC3-2. The initial ratio of the two parentalmacroconidia in the mixed inoculum is indicated on the x-axis,and the final ratio of the two parents in macroconidiaharvested from the heterokaryon is indicated above eachpoint on the graph. The final ratio of CCI-3 to MC3-2 in theheterokaryon which had the fastest colony radial growth ratewas greater than 1: 800; no CCI-3 colonies were formed frommacroconidia harvested from the fastest (highest Kr)-growingheterokaryons; however, it was demonstrated that the MC32 macroconidia and hyphal fragments collected from thesecolonies were not revertants as their Kr (55 ± 2 I-Im h-1
) wasthe same as that of the original strain (54 ± 5 I-Im h-1; Kr
measured from non-conidial inocula). For crosses betweenCCl-1 + MC3-5 (results not shown), a final ratio in theheterokaryon of 3 :1 for the parental macroconidia respectivelygave the fastest colony radial growth rate (174 ± 2 I-Im h-1
).

Marilyn G. Wiebe and others
macroconidia collected from heterokaryons started fromdifferent ratios of parental macroconidia, and the final nuclearratio observed from macroconidia collected from heterokaryons did not generally reflect the initial ratio of theparental macroconidia in the inoculum (Fig. 4). Shifts innuclear ratio have sometimes been observed in heterokaryonsof N. crassa, and generally these changes were in a directionwhich resulted in the fastest extension rates (Pittenger &
Atwood, 1954). In crosses between F.graminearum CCl-l andMC3-5 (results not shown) or CCl-3 and MC3-2, shifts in thenuclear ratio in the heterokaryons did not always result inoptimal extension rates. However, if observations for CCl-3plus MC3-2 heterokaryons had been continued beyond 96 h(the time at which the final K measurement was made) theymight eventually have reached optimal nuclear ratios, andhence maximal or wild-type K values (Fig. 3). In otherheterokaryons tested, a separation of the parental nucleartypes with a breakdown of the heterokaryon occurred. Theseobservations suggest that heterokaryons with Krs less thanthat of the wild type (Table 3) are as likely to reflectsuboptimal nuclear ratios as allelic complementation or partialdominance. However, evidence for allelic complementationwas also observed.
Allelic complementation has frequently been reported in N.crassa (Fincham & Pateman, 1957; Woodward et al., 1958;Fincham & Coddington, 1963). Allelic complementation isgenerally observed where the mutant enzyme is a dimer oroligomer and a hybrid enzyme can be formed between thetwo defective parental enzymes, although complementation isalso possible for monomeric enzymes (Fincham, 1985).Woodward et al. (1958) were able to measure adenylosuccinaseactivity in heterokaryons of N. crassa ad-4 mutants (impairedin adenylosuccinase activity) and found enzyme activitiesranging between 0 and 25 % of the wild-type activity, whileextension rates ranged from 5 to 100 % of the control.Fincham & Coddington (1963) observed similar levels ofenzyme activity in allelic complementation of N. crassa ammutants (impaired in glutamate dehydrogenase activity) bothin vivo and in vitro. The enzyme(s) which determines thephenotype of the morphological mutants described here isunknown and thus could not be measured. As in N. crassa,some allelic mutants of F. graminearum complemented toproduce heterokaryons with wild-type morphology and otherswere only partly restored (Table 3). Nuclear ratios may also beimportant in determining the degree of allelic complementationobserved (Fincham & Coddington, 1963).
That only three complementation groups accounted for thevariety of morphological mutants isolated from glucoselimited, continuous flow cultures of F. graminearum A 3/5suggests that mutations in only a small proportion ofmorphology-determining genes were selectively advatageousfor the cultural conditions (glucose limitation) employed. Nosignificantly increased specific growth rates were observedamong these mutants (Table 2). However, a small increase in1-1, which might account for a selective advantage, would notnecessarily be detected by the method used here to measuregrowth rate (e.g. see Read et al., 1986). Similarly, the substratesaturation constant (Ks) of F. graminearum for glucose is verylow (2'5 I..lM; M. G. Wiebe, unpublished result) and small
36
561
differences in Ks) between the mutants and A 3/5 would bevery difficult to detect experimentally. Although no differencein the Michaelis constant for transport (Km , measured byuptake of radioactive glucose) was observed between A 3/5and CCl-l, the mutant with the greatest selective advantageunder glucose limitation (M. G. Wiebe, unpublished result),residual glucose concentrations in glucose-limited, continuousflow cultures at a dilution rate of 0.19 h-1 were lower formixed cultures of A 3/5 containing morphological mutantsthan for pure A 3/5 cultures (G. D. Robson, unpublishedresult), suggesting that some alteration in glucose metabolismmay confer a selective advantage to the mutants. Alternatively,the altered morphology may itself be selectively advantageousin some way. However, a highly branched mutant (CI06) ofA 3/5 isolated from a non-limited continuous flow culture(turbidostat) did not have a selective advantage in glucoselimited conditions (Wiebe, 1989).
We thank the Science and Engineering Research Council andMarlow Foods for supporting this work.
REFERENCES
Barratt, R. W. & Gamjobst, L. (1949). Genetics of a colonial microconidiatingmutant strain of Neurospora crassa. Genetics 34, 351-369.
Beadle, G. W. & Coonradt, V. L. (1944). Heterocaryosis in Neurospora crassa.Genetics 29, 291-308.
Cohen, B. L. (1972). Ammonium repression of extracellular protease inAspergillus nidulans. Journal of General Microbiology 71, 293-299.
Dodge, B. O. (1942). Heterocaryotic vigor in Neurospora. Bulletin of theTorrey Botanical Club 69, 75-9I.
Fincham, j. R. S. (1985). From auxotrophic mutants to DNA sequences. InGene Manipulations in Fungi (ed. j. W. Bennett & L. L. Lasure), pp. 3-34.Academic Press: London.
Fincham, j. R. S. & Coddington, A. (1963). Complementation at the am locusof Neurospora crassa: a reaction between different mutant forms ofglutamate dehydrogenase. Journal of Molecular Biology, 6, 361-373.
Fincham, ). R. S. & Pateman, ). A. (1957). Formation of an enzyme throughcomplementary action of mutant ' alleles' in separate nuclei of aheterokaryon. Nature 179, 741-742.
Forss, K. G.. Gadd, G. 0., Lundell, R. O. & Williamson, H. W. (1974). Processfor the manufacture of protein-containing substances for fodder, foodstuffsand technical application. U.S. Patent Office, patent no. 3,809,614.
Gamjobst, L. & Tatum, E. L. (1967). A survey of new morphological mutantsin Neurospora crassa. Genetics 57, 579--604.
Hardy, K. G. & Oliver, S. G. (1985). Genetics and biotechnology. InBiotechnology: Principles and Applications (ed. l. j. Higgins, D.). Best & J.James), pp. 257-282. Blackwell, Oxford, UK
Miller, j. j. (1946). Cultural and taxonomic studies on certain fusaria. l.Mutations in culture. Canadian Journal of Research, 24 C. 188-212.
Pateman, j. A. & Kinghorn. ). K. (1976). Nitrogen metabolism. In TheFilamentous Fungi, vol. 2 (ed. j. E. Smith & D. R. Berry), pp. 159-237.Edward Arnold: London.
Pittinger, T. H. & Atwood, K. C. (1954). The relation of growth rate tonuclear ratio in Neurospora heterokaryons. Genetics 39, 987-988.
Read, D. L Gardner, D. C. J. & Oliver, S. G. (1986). The yeast and plasmidstrategies for the survival of a selfish DNA. Molecular & General Genetics205,417-42I.
Righelato, R. C. (1976). Selection of strains of Penicillium chrysogenum withreduced penicillin yields in continuous cultures. Journal of Applied Chemistry& Biotechnology, 26, 153-159.
Trinci, A. P.). (1973a). Growth of wild type and spreading colonial mutantsof Neurospora crassa in batch culture and on agar medium. Archiv furMilcrobiologie 91, 113-126.
Trinci, A. P. J. (1973 b). The hyphal growth unit of wild type and spreadingcolonial mutants of Neurospora crassa. Archiv fUr Milcrobiologie 91, 127-136.
MYC96

c. W. H. & Giles, N. H. (1958).locus in Neurospora crassa. Genetics 44,
Morphological mutants of Fusarium
Trind, A. P. J. (1984). Regulation of hyphal branching and hyphal orientation.In Ecology and Physiology of the Fungal Mycelium, Eighth Symposium of theBritish Mycological Society (ed. D. H. Jennings & A. D. M. Rayner), pp.23-52. Cambridge University Press: Cambridge, U.K.
Trind, A. P. J. (1992). Presidential address 1991. Myco-protein: a twenty-yearovernight success story. Mycological Research 96, 1-13.
Trind, A. P. L Robson, G. D., Wiebe, M. G., Cunliffe, B. & Naylor, T. W.(1990). Growth and morphology of Fusarium graminearum and other fungiin batch and continuous culture. In Microbial Growth Dynamics (ed. R. K.Poole, M. J. Bazin & C. W. Keevil), pp. 17-38. lRL Press: Oxford, U.K.
Vogel. H. J. (1956). A convenient growth medium for Neurospora (MediumN). Microbial Genetics Bulletin 13, 42-44.
(Accepted 13 December 1991)
562
Wiebe, M. G. (1989). The morphology of Fusarium graminearum in batch andcontinuous culture. Ph.D. Thesis, University of Manchester.
Wiebe, M. G. & Trind, A. P. J. (1991). Dilution rate as a determinant ofmycelial morphology in continuous culture. Biotechnology and Bioengineering38, 75-81.
Wiebe, M. G., Trind, A. P. L Cunliffe, B., Robson, G. D. & Oliver, S. G.(1991). Appearance of morphological (colonial) mutants in glucose-limited,continuous flow cultures of Fusarium graminearum. Mycological Research 95,
1284-1288.Woodward, D. 0., Partridge,
Complementation at the Ad-41237-1244.
Related Documents











