Molecular Microbiology (2004) 52(3), 873–893 doi:10.1111/j.1365-2958.2004.04026.x © 2004 Blackwell Publishing Ltd Blackwell Science, LtdOxford, UKMMIMolecular Microbiology0950-382XBlackwell Publishing Ltd, 2004 ? 2004523873893Original ArticleChemosensory control of twitching motility in P. aeruginosaC. B. Whitchurch et al. Accepted 9 January, 2004. *For correspondence. E-mail [email protected]; Tel. (+61) 7 3346 2110; Fax (+61) 7 3346 2111. Characterization of a complex chemosensory signal transduction system which controls twitching motility in Pseudomonas aeruginosa Cynthia B. Whitchurch, 1,4 Andrew J. Leech, 1 , 2 Michael D. Young, 1,2 Derek Kennedy, 1,3 Jennifer L. Sargent, 1,2 Jacob J. Bertrand, 6 Annalese B. T. Semmler, 1,2 Albert S. Mellick, 1,2 Paul R. Martin, 1 Richard A. Alm, 1 Matthew Hobbs, 1 Scott A. Beatson, 1,2 Bixing Huang, 1 Lam Nguyen, 4 James C. Commolli, 4 Joanne N. Engel, 4,5 Aldis Darzins, 7 and John S. Mattick 1,2 * 1 ARC Special Research Centre for Functional and Applied Genomics, Institute for Molecular Bioscience, University of Queensland, Brisbane, QLD 4072, Australia. 2 School of Molecular and Microbial Sciences, University of Queensland, Brisbane, QLD 4072, Australia. 3 School of Biomolecular and Biomedical Sciences, Griffith University, Nathan QLD 4111, Australia. 4 Department of Medicine, University of California, San Francisco, San Francisco, California, 94143-0654, USA. 5 Microbiology and Immunology, University of California, San Francisco, California, 94143-0654, USA. 6 Biochemistry and Biophysics, University of California, San Francisco, California, 94143-0654, USA. 7 Promega Corporation, 2800 Woods Hollow Road, Madison, WI 53711-5399, USA. Summary Virulence of the opportunistic pathogen Pseudomo- nas aeruginosa involves the coordinate expression of a wide range of virulence factors including type IV pili which are required for colonization of host tissues and are associated with a form of surface transloca- tion termed twitching motility. Twitching motility in P. aeruginosa is controlled by a complex signal trans- duction pathway which shares many modules in com- mon with chemosensory systems controlling flagella rotation in bacteria and which is composed, in part, of the previously described proteins PilG, PilH, PilI, PilJ and PilK. Here we describe another three compo- nents of this pathway: ChpA, ChpB and ChpC, as well as two downstream genes, ChpD and ChpE, which may also be involved. The central component of the pathway, ChpA, possesses nine potential sites of phosphorylation: six histidine-containing phospho- transfer (HPt) domains, two novel serine- and threonine-containing phosphotransfer (SPt, TPt) domains and a CheY-like receiver domain at its C- terminus, and as such represents one of the most complex signalling proteins yet described in nature. We show that the Chp chemosensory system controls twitching motility and type IV pili biogenesis through control of pili assembly and/or retraction as well as expression of the pilin subunit gene pilA . The Chp system is also required for full virulence in a mouse model of acute pneumonia. Introduction Pseudomonas aeruginosa is a ubiquitous Gram-negative bacterium found throughout the environment, and is also an opportunistic pathogen of a wide variety of eukaryotes including mice, insects, nematodes, and plants (Jander et al ., 2000; Rahme et al ., 2000; Tan and Ausubel, 2000). It also causes serious and often life-threatening infections in immunocompromised humans such as those who are suffering from severe burns, cystic fibrosis or AIDS, who are undergoing cancer chemotherapy, or who recovering from major surgery (Giamarellou, 2000). The most critical threat to the health and survival of individuals with cystic fibrosis is the persistent injury to the lungs and airways caused by chronic infection with P. aeruginosa. Pseudomo- nas aeruginosa is equipped with a large arsenal of secreted and cell associated virulence factors, including adhesins, exopolysaccharides, proteases, lipases, phos- pholipases, siderophores, biosurfactants and exotoxins, which provide nutrients for bacterial growth, enhance inva- sive potential, and/or directly damage host tissue. The major adhesins of P. aeruginosa which facilitate attachment of the bacterium to host epithelial cells are type IV pili, which are filamentous structures located at the poles of the bacterial cell. Type IV pili are retractile and mediate a mode of surface translocation termed twitching motility. They also act as receptors for certain bacteriophages (for recent review see Mattick, 2002). Twitching motility occurs in a wide variety of pathogenic bacteria, including Neisseria gonorrhea , and is mediated by pili extension and retraction (Merz et al ., 2000; Skerker

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Molecular Microbiology (2004)
52
(3), 873–893 doi:10.1111/j.1365-2958.2004.04026.x
© 2004 Blackwell Publishing Ltd
Blackwell Science, LtdOxford, UKMMIMolecular Microbiology0950-382XBlackwell Publishing Ltd, 2004
? 2004
52
3873893
Original Article
Chemosensory control of twitching motility in P. aeruginosaC. B. Whitchurch
et al.
Accepted 9 January, 2004. *For correspondence. [email protected]; Tel. (+61) 7 3346 2110; Fax (+61) 7 33462111.
Characterization of a complex chemosensory signal transduction system which controls twitching motility in
Pseudomonas aeruginosa
Cynthia B. Whitchurch,
1,4
Andrew J. Leech,
1
,
2
Michael D. Young,
1,2
Derek Kennedy,
1,3
Jennifer L. Sargent,
1,2
Jacob J. Bertrand,
6
Annalese B. T. Semmler,
1,2
Albert S. Mellick,
1,2
Paul R. Martin,
1
Richard A. Alm,
1
Matthew Hobbs,
1
Scott A. Beatson,
1,2
Bixing Huang,
1
Lam Nguyen,
4
James C. Commolli,
4
Joanne N. Engel,
4,5
Aldis Darzins,
7
and John S. Mattick
1,2
*
1
ARC Special Research Centre for Functional and Applied Genomics, Institute for Molecular Bioscience, University of Queensland, Brisbane, QLD 4072, Australia.
2
School of Molecular and Microbial Sciences, University of Queensland, Brisbane, QLD 4072, Australia.
3
School of Biomolecular and Biomedical Sciences, Griffith University, Nathan QLD 4111, Australia.
4
Department of Medicine, University of California, San Francisco, San Francisco, California, 94143-0654, USA.
5
Microbiology and Immunology, University of California, San Francisco, California, 94143-0654, USA.
6
Biochemistry and Biophysics, University of California, San Francisco, California, 94143-0654, USA.
7
Promega Corporation, 2800 Woods Hollow Road, Madison, WI 53711-5399, USA.
Summary
Virulence of the opportunistic pathogen
Pseudomo-nas aeruginosa
involves the coordinate expression ofa wide range of virulence factors including type IV piliwhich are required for colonization of host tissuesand are associated with a form of surface transloca-tion termed twitching motility. Twitching motility in
P.aeruginosa
is controlled by a complex signal trans-duction pathway which shares many modules in com-mon with chemosensory systems controlling flagellarotation in bacteria and which is composed, in part,of the previously described proteins PilG, PilH, PilI,PilJ and PilK. Here we describe another three compo-nents of this pathway: ChpA, ChpB and ChpC, as wellas two downstream genes, ChpD and ChpE, whichmay also be involved. The central component of the
pathway, ChpA, possesses nine potential sites ofphosphorylation: six histidine-containing phospho-transfer (HPt) domains, two novel serine- andthreonine-containing phosphotransfer (SPt, TPt)domains and a CheY-like receiver domain at its C-terminus, and as such represents one of the mostcomplex signalling proteins yet described in nature.We show that the Chp chemosensory system controlstwitching motility and type IV pili biogenesis throughcontrol of pili assembly and/or retraction as well asexpression of the pilin subunit gene
pilA
. The Chpsystem is also required for full virulence in a mousemodel of acute pneumonia.
Introduction
Pseudomonas aeruginosa
is a ubiquitous Gram-negativebacterium found throughout the environment, and is alsoan opportunistic pathogen of a wide variety of eukaryotesincluding mice, insects, nematodes, and plants (Jander
et al
., 2000; Rahme
et al
., 2000; Tan and Ausubel, 2000).It also causes serious and often life-threatening infectionsin immunocompromised humans such as those who aresuffering from severe burns, cystic fibrosis or AIDS, whoare undergoing cancer chemotherapy, or who recoveringfrom major surgery (Giamarellou, 2000). The most criticalthreat to the health and survival of individuals with cysticfibrosis is the persistent injury to the lungs and airwayscaused by chronic infection with
P. aeruginosa. Pseudomo-nas aeruginosa
is equipped with a large arsenal ofsecreted and cell associated virulence factors, includingadhesins, exopolysaccharides, proteases, lipases, phos-pholipases, siderophores, biosurfactants and exotoxins,which provide nutrients for bacterial growth, enhance inva-sive potential, and/or directly damage host tissue.
The major adhesins of
P. aeruginosa
which facilitateattachment of the bacterium to host epithelial cells aretype IV pili, which are filamentous structures located atthe poles of the bacterial cell. Type IV pili are retractileand mediate a mode of surface translocation termedtwitching motility. They also act as receptors for certainbacteriophages (for recent review see Mattick, 2002).Twitching motility occurs in a wide variety of pathogenicbacteria, including
Neisseria gonorrhea
, and is mediatedby pili extension and retraction (Merz
et al
., 2000; Skerker

874
C. B. Whitchurch
et al.
© 2004 Blackwell Publishing Ltd,
Molecular Microbiology
,
52
, 873–893
and Berg, 2001; Mattick, 2002). Twitching motility, at leastin
P. aeruginosa
, is also involved in biofilm development(O’Toole and Kolter, 1998; Chiang and Burrows, 2003;Klausen
et al
., 2003a,b).
Pseudomonas aeruginosa
has become in recent yearsone of the main model organisms for the characterizationof the molecular genetics and biology of type IV pili andtwitching motility. Thus far more than 35 genes have beenidentified to be involved in the biogenesis, function andregulation of type IV pili in
P. aeruginosa
(Alm and Mattick,1997; Mattick, 2002). Many of these have homology toother gene/protein sets involved in protein secretion andDNA uptake in various bacteria (Whitchurch
et al
., 1991;Hobbs and Mattick, 1993; Mattick and Alm, 1995). Thebiogenesis and function of type IV pili in
P. aeruginosa
iscontrolled by multiple signal transduction systems, includ-ing the two-component sensor regulator systems
pilSR
(Hobbs
et al
., 1993; Strom and Lory, 1993) and
algR/fimS
(Whitchurch
et al
., 1996), as well as the global carbonmetabolism regulator Crc (O’Toole
et al
., 2000), and thevirulence factor regulator Vfr (Beatson
et al
., 2002).Twitching motility in
P. aeruginosa
is also controlled bya chemosensory system which is encoded in part by thepreviously characterized gene cluster
pilG, pilH, pilI, pilJ
and
pilK
(Darzins, 1993; 1994; 1995). In this study wehave identified another three members of this system(
chpA, chpB, chpC)
and another two genes (
chpD
,
chpE
)which might also be involved. This gene cluster is locatedimmediately downstream of
pilK
. Together the proteinsencoded by these genes appear to comprise a complexchemosensory signal transduction pathway which sharesmany modules in common with chemosensory systemswhich control flagella rotation in bacteria.
The most extensively studied of these chemosensorysystems are those that control swimming motility in theenteric species
Escherichia coli
and
Salmonella enterica
serovar
Typhimurium
. The biochemistry of signalling reac-tions in these systems are well characterized and atomicresolution structures for most of the domains and compo-nents of these systems have been solved (for reviews seeFalke
et al
., 1997; Djordjevic and Stock, 1998; Armitage,1999; Bourret and Stock, 2002). The enteric chemosen-sory systems are composed of membrane boundchemoreceptors referred to as methyl accepting chemot-axis proteins (MCPs) which are coupled via the adaptorprotein CheW to the multidomain histidine protein kinaseCheA. The MCPs function to modulate the kinase activityof CheA in response to chemical stimuli. CheA possessesin its C-terminus conserved motifs required for nucleotidebinding. Autophosphorylation occurs at a conserved his-tidine in the N-terminus of the protein in a domain termedP1. In recent years it has become evident that the P1domain of CheA is structurally related to the histidinecontaining phosphotransfer (HPt) domains found in inter-
mediates of multistep phosphorelay reactions such as theC-terminal HPt domain of ArcB (Kato
et al
., 1997) and theyeast protein YpD1 (Song
et al
., 1999; Xu and West,1999). The high-energy phosphoryl group is then trans-ferred from the histidine of the CheA P1/HPt domain to aconserved aspartate in the response regulator CheY,which then interacts directly with the flagellar motor toswitch the direction of flagellar rotation. The rate of auto-catalytic dephosphorylation of CheY-P is enhancedthrough interaction with CheZ. Sensory adaptation whichallows temporal control of swimming motility occursthrough methylation of specific glutamate residues on theMCP to reset it into a non-signalling state. The methylationstatus of the MCP is adjusted via competing activities ofthe methyl transferase CheR and the methyl esteraseCheB. CheB also possesses a response regulator moduleand is competitively phosphorylated by CheA (Falke
et al
.,1997; Djordjevic and Stock, 1998; Armitage, 1999; Bour-ret and Stock, 2002).
Over recent years more complex chemosensory sys-tems have been described in other bacteria and havebeen found to control, in addition to swimming, other bac-terial motilities including swarming, gliding and twitching(Burkart
et al
., 1998; Armitage, 1999; Bourret and Stock,2002; Mattick, 2002; Winther-Larsen and Koomey, 2002).In fact the
Pseudomonas aeruginosa
PAO1 genome hasfour gene clusters encoding chemotaxis-like phosphorelaysignal transduction systems (Croft
et al
., 2000; Stover
et al
., 2000). Two of these systems are involved in chemo-tactic control of swimming motility (Masduki
et al
., 1995;Ferrandez
et al
., 2002), the Wsp gene cluster controlssurface properties that lead to autoaggregation (D
¢
Arge-nio
et al
., 2002) and the
pilG-K
,
chpA-C
gene cluster isinvolved in the control of twitching motility (Darzins, 1993;1994; 1995) which is the focus of this study.
Results
Sequence analysis of the region downstream of
pilG-K
We have previously generated a library of twitching motil-ity mutants through transposon (Tn
5-
B21) mutagenesis ofthe
P. aeruginosa
strain K genome (Hobbs
et al
., 1993).Southern blot and sequence analysis of the points oftransposon insertions in this library identified four mutants(S43, S115, S154 and S247) which contain transposoninsertions in the previously characterized gene
pilJ
(Fig. 1A)
.
We also identified another nine mutants (S10,S23, S33, S45, S52, S129, S164, S209 and S349) whichhave transposon insertions in the region immediatelydownstream of
pilK
(Fig. 1A).The 12 kb region immediately downstream of
pilK
wascloned from the PAO1 cosmid pMO01539 and sequencedusing a combination of
Exo
III deletions, subcloning and

Chemosensory control of twitching motility in
P. aeruginosa 875
© 2004 Blackwell Publishing Ltd,
Molecular Microbiology
,
52
, 873–893
primer walking (GenBank Accession Number U79580).Analysis of the sequence of this region identifiedfive open reading frames (ORFs) which we have de-signated
chpA
,
chpB
,
chpC
,
chpD
and
chpE
(Fig. 1A).These ORFS are also referred to as PA0413 – PA0417 inthe completed
P. aeruginosa
genome sequence(http://www.pseudomonas.com; Stover
et al
., 2000). Thepredicted gene products of these ORFs were character-
ized using a combination of
BLAST
searches of GenBank(http://www.ncbi.nlm.nih.gov/
BLAST
/; Altschul
et al.
, 1997)with
PFAM
(http://www.sanger.ac.uk/Software/Pfam/;Bateman
et al.
, 2002) and
SMART
(http://smart.embl-heidelberg.de/; Schultz
et al
., 1998; Letunic
et al
., 2002)analyses. Together with the upstream genes
pilG
,
pilH
,
pilI
,
pilJ
and
pilK
the predicted gene products of
chpA
,
chpB
,
chpC
,
chpD
and
chpE
appear to constitute a com-
Fig. 1.
A. Organization of the
pilGHIJK
and
chpABCDE
gene cluster. Arrows represent the direction of transcription of the genes. The location of transposon insertions are indicated by filled triangles. The sites of insertion of the TcR cassette for generation of allelic exchange mutants are indicated by open triangles/boxes. Relevant restriction enzyme sites are also shown. The class of proteins to which each gene product belongs is shown above the boxes.B. Schematic of the ChpA protein. Relevant domains and features of the ChpA protein are indicated. ‘HPt’, ‘TPt’ and ‘SPt’ indicate the predicted histidine, threonine or serine containing phosphotransfer domains, respectively; ‘FimL-like’ denotes the region of ChpA which shares homology with FimL and encompasses both HPt1 and TPt; ‘HisKc-like’ denotes the autocatalytic histidine kinase domain which incorporates the CheY docking domain (P2) and the N, G1, F, and G2 boxes which are required for ATP binding; ‘CheW’ indicates the CheW-like domain which is involved in mediating CheA interactions with CheW; and ‘CheY’ denotes the CheY-like response regulator domain.C. Alignment of the putative phosphotransfer domains of ChpA with the P 1/HPt domain of
E. coli
CheA. Similar residues are boxed, identical residues are indicated in bold and are shaded. *denotes the position of the predicted site of phosphorylation in each domain. The alignment was generated using the
CLUSTALW
feature in MacVector (Oxford Molecular Group).

876
C. B. Whitchurch et al.
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 873–893
plex signal transduction pathway analogous to the bacte-rial chemotaxis systems which control flagella rotation inresponse to chemical attractants and repellents. In lightof this we have designated this putative chemosensorypathway the Chp (chemosensory pili) system.
The predicted ATG start codon of the first ORF of thisregion (chpA, PA0413) overlaps the TGA stop codon ofpilK. This 7431 bp ORF is predicted to encode a 2477 aa(269 Kda) multidomain protein which belongs to the familyof CheA-like histidine kinases. The central portion of ChpAresembles other CheA-like proteins and contains a con-served CheY docking domain (P2); an autocatalytic histi-dine kinase domain including the N, G1, F, and G2 boxeswhich are required for ATP binding; and a CheW-likedomain which is involved in mediating CheA interactionswith CheW (Falke et al., 1997) (Fig. 1B). At the C-terminusof ChpA is located a response regulator module. ChpAthen is a hybrid of homologues of the histidine kinaseCheA and the response regulator CheY. According tothe SMART database (http://smart.embl-heidelberg.de/;Schultz et al., 1998; Letunic et al., 2002) there have beenmore than 20 CheA/CheY hybrid proteins reported to date.
The N-terminal two-thirds of ChpA contains eight pre-dicted phosphotransfer domains, six of which are conven-tional histidine-containing HPt (or P1) domains and twoothers which have threonine and serine in place of histi-dine at their active site (Figs 1B, C). BLAST searches ofGenBank and SMART analyses indicate that most CheA-like histidine kinases possess only one HPt domain in theirN-termini. However BLAST and SMART analyses indicatethat each of the following bacteria encode a CheA-likehistidine kinase that possesses more than one HPtdomain: Synechocystis PCC6803 (NP_442714; 2 HPt),Nitrosomonas europaea ATCC 19718 (NP_841302.1;3 HPt), Xylella fastidiosa strains Ann-1 (ZP_00041851.1;4 HPt), Dixon (ZP_00039906.1; 4 HPt), 9a5c(NP_299234.1; 4 HPt), Temeula 1 (NP_779067.1; 4 HPt),Ralstonia metallidurans (ZP_00021550.1; 4HPt), Ral-stonia solanacearum (NP_518793.1; 4 HPt), Xantho-monas campestris pv. campestris str. ATCC 33913(NP_638269.1; 4 HPt), Xanthomonas axonopodis pv. citristr. 306 (NP_643407.1; 4 HPt), P. putida KT2440(NP_747090.1; 4 HPt), P. syringae pv. tomato str DC3000(NP_794763.1; 5 HPt), P. syringae pv. syringae B728a(ZP_00125157.1; 5 HPt) P. fluorescens PfO-1(ZP_00085475.1; 5 HPt) and Microbulbifer degredans 2–40 (ZP_00067212.1; 5HPt). Each of these multi-HPt con-taining CheA-like histidine kinases also possesses aCheY domain at its C-terminus proteins. In fact theseproteins appear to be bona fide homologues of the P.aeruginosa ChpA (see Discussion).
In addition to the six HPt domains, P. aeruginosa ChpAalso possesses another two domains which are homolo-gous to HPt domains, except that they contain a threonine
or a serine residue (T476, S1226) in place of the normalhistidine at the phosphorylation site (Fig. 1B and C).Three-dimensional homology modelling suggests thatthese predictions are correct (see below). We havetermed the potential threonine-containing phosphotrans-fer domain and the potential serine-containing phospho-transfer domain TPt and SPt domains respectively.Potential Tpt/SPt domains are also found in the ChpAhomologues of N. europaea, X. fastidiosa, M. degredans,P. putida, P. fluorescens and P. syringae. Over the pastfew years it has become evident that serine/threonineprotein kinases are widespread in bacteria (Kennelly,2002), but ChpA and its homologues are the first exam-ples of putative serine/threonine phosphorylation in thecontext where a bacterial protein belonging to the classof CheA-like protein kinases appears to have the poten-tial to be phosphorylated at serine/threonine residues.Whether phosphorylation at these residues occurs byautophosphorylation or by phosphorylation by a serine/threonine kinase encoded elsewhere in the genomeremains to be determined. In total then ChpA possessesnine potential sites of phosphorylation, including six his-tidines, a threonine and a serine in conserved HPt/TPt/SPt domains in the N-terminal two-thirds of the protein,and the conserved aspartate of the CheY domain at theC-terminus, thus making it the most complex member ofthis class of CheA-like histidine kinases reported andquite possibly the most complex signal transduction pro-tein yet described in nature.
The N-terminal 562 aa of ChpA which encompass thefirst HPt and TPt domains are also homologous (33%identity, 51% similarity) to FimL, a protein encoded byPA1822 that we have identified to be required for normaltwitching motility in P. aeruginosa (C. B. Whitchurch, S. A.Beatson, T. Jakobsen, J. C. Commolli, J. L. Sargent, J. J.Bertrand, P. J. Kang, J. S. Mattick and J. N. Engel, manu-script in preparation). Interestingly, whereas FimL ishomologous to the N-terminal region of ChpA, it does notcontain either of the predicted phosphorylatable histidineor threonine residues although the surrounding se-quences remain highly conserved.
We had originally concluded that this ‘FimL-like’ regionwas encoded by a separate gene, which we had desig-nated pilL (Alm and Mattick, 1997). However, our originalsequence submitted to GenBank contained a frameshifterror. To further confirm the size of ChpA we performedWestern blot analysis of FLAG epitope-tagged ChpAexpressed in P. aeruginosa. No bands were detected inWestern blots of whole cell samples of P. aeruginosatransformants containing the vector (pUCPSK) alone orthe plasmid containing the chpA coding sequence lackingthe FLAG-tag (pUCPChpA) whereas a single band at~280 kDa was detected in cells expressing FLAG-taggedChpA (pUCPChpA-FLAG) (Fig. 2). A single unique band

Chemosensory control of twitching motility in P. aeruginosa 877
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 873–893
of the same molecular weight was observed with transfor-mants of both pUCPChpA and pUCPChpA-FLAG (but notpUCPSK) when SDS–PAGE gels of whole cell sampleswere Coomassie stained (Fig. 2). The predicted molecularweight for the shorter version of ChpA (which lacks the‘FimL-like’ region) is only 178 KDa whereas the observedsize for the ChpA (~280 KDa) is in broad agreement withthe predicted value of 269 kDa for the gene product of thelonger chpA coding sequence.
The predicted ATG start codon of the second ORF(PA0414) in this region, which we have termed chpB,overlaps the final seven residues of chpA. chpB encodesa 343 aa protein homologous to the CheB methyl-esterases which in the chemotaxis systems function todemethylate the sensory methyl accepting chemotaxisproteins (MCPs). We predict by analogy therefore thatChpB serves to demethylate (at least) the predicted MCPPilJ as part of a sensory adaptation feedback circuit. TheTGA stop codon of chpB overlaps the predicted ATG startcodon of the third ORF (PA0415) of this region which hasbeen designated chpC and which encodes a 168 aa pro-tein predicted by SMART analysis (http://smart.embl-heidelberg.de/ Schultz et al., 1998; Letunic et al., 2002)to be a CheW-like protein. The Chp system therefore hastwo CheW homologues (PilI and ChpC) which we predictby analogy to the well characterized Che systems of otherbacteria to function to couple PilJ and perhaps otherMCPs to ChpA. P. aeruginosa contains 26 genes pre-
dicted to encode MCPs which are scattered around thegenome (Croft et al., 2000).
Located eight residues downstream of the stop codonof chpC is the predicted ATG start codon of the fourthORF (PA0416) of the region which has been termed chpD,as it appears likely to be part of the chp cluster given theclose spacing between it and chpC. This ORF is predictedto encode a 264 aa transcriptional regulator proteinbelonging to the AraC family (Gallegos et al., 1997). Thisclass of regulator is not a member of the two-componenttransmitter-receiver phosphorelay systems but insteadbinds effector molecules via the N-terminal and centralregions of the protein and activates transcription of targetgenes through a conserved helix–turn–helix motif locatedin the C-terminus of the protein. The effector moleculebound by ChpD and the target genes of this regulator areunknown.
The final ORF of this region (PA0417), which we havetentatively termed chpE, is predicted to start 78 residuesdownstream of chpD. chpE encodes a 203 aa proteinwhich is predicted to have a signal sequence and sixtransmembrane domains and appears therefore to be asmall integral inner membrane protein. Database analy-ses predict that ChpE is a member of the LysE family ofamino acid efflux proteins (Aleshin et al., 1999; Vrljicet al., 1999). Closer analysis of the ChpE primarysequence indicates that it is a member of the RhtB sub-family (Aleshin et al., 1999). RhtB is an E. coli proteinwhich is responsible for the efflux of homoserine andhomoserine lactone (Zakataeva et al., 1999). The solutetransported by P. aeruginosa ChpE is unknown. Locatedabout 100 nucleotides past the stop codon of chpE is apotential rho-independent transcriptional terminator.
3D-structural modelling of putative phosphotransfer domains of ChpA
As described above, we have identified eight motifs inChpA that show significant primary sequence homologyto the histidine containing phosphotransfer (P1/Hpt)domains found in CheA-like histidine kinases. Compari-son of the solved structures of the histidine containingphosphotransfer domains of CheA-like histidine kinases(Zhou and Dahlquist, 1997; Mourey et al., 2001) with thestructures of HPt domains found in intermediates of mul-tistep phosphorelay reactions such as the C-terminal HPtdomain of ArcB (Kato et al., 1997) and the yeast proteinYpD1 (Song et al., 1999; Xu and West, 1999) has dem-onstrated that they each contain a conserved four-helixbundle core which is required for phosphotransfer (Fig. 3)(Song et al., 1999; Xu and West, 1999; Mourey et al.,2001). We used 3D-structural modelling to examine thelikely three-dimensional structures of the predicted phos-photransfer domains Hpt1, Hpt4 and Spt of ChpA (Fig. 3;
Fig. 2. Determination of ChpA molecular weight. P. aeruginosa car-rying the cloning vector pUCPSK (lane 1); pUCPchpA expressing untagged ChpA (lane 2); or pUCPChpA-FLAG expressing FLAG-tagged ChpA (lane 3). The left panel is a Coomassie stained 4–20% SDS–PAGE gradient gel of whole cell samples. The right panel is an anti-FLAG M2 immunoblot of whole cell samples. These results are typical of triplicate experiments. Identical results were obtained with both PAK and PAO1 transformants.

878 C. B. Whitchurch et al.
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 873–893
data not shown), as a means of obtaining additional evi-dence that these domains are capable of functioning asphosphotransfer domains. The comparison of the solvedstructures of ArcB, CheA and YPD1, and the modelled
structures of these putative phosphotransfer domains ofChpA predict that the phosphotransfer domains of ChpAalso have a common core structure consisting of an anti-parallel four-helix core (data not shown; see Fig. 3 for the

Chemosensory control of twitching motility in P. aeruginosa 879
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 873–893
predicted structure of the ChpA SPt domain). In ArcB andCheA, these core structures are further stabilized by a fifthhelix at the N-terminal and C-terminal regions of thesedomains, respectively (Kato et al., 1997; Mourey et al.,2001). Our modelling suggests that the ChpA phospho-transfer domains share the same antiparallel four-helixcore but the position of a fifth helix cannot be accuratelypredicted. These proteins may use more complex struc-tures to stabilize the overall fold as is observed in YPD1(Song et al., 1999; Xu and West, 1999).
Of particular interest were the predicted Tpt/Spt phos-photransfer domains in which the active histidine isreplaced with a threonine or serine (Figs 1 and 3). Closercomparison of the phosphotransfer domains of eitherArcB or CheA and the putative serine phosphotransfer(SPt) domain of ChpA show that the environment sur-rounding Ser1231 contains compensatory amino acidsubstitutions which allow Ser1231 access to interactivedonor proteins (Fig. 3B). The positioning of Arg1235 onthe border of a flexible loop proximal to Ser1231 is com-patible with a model in which Arg1235 is free to be dis-placed during interactions with donor molecule. Uponphosphorylation of Ser1231 we propose that Arg1235would rotate back and neutralize the charged group onSer1231. This would be consistent with previous reportsthat suggest positively charged residues such as argininemay neutralize the negative charge of phosphoryl groups(Kato et al., 1997). Potential conformational motions assuggested here would be consistent with data reportedfrom other phosphorylation events (Cox et al., 1994) aswell as having a role in stabilizing the helical structuresas has been observed in the HPr protein of Bacillus sub-tilis (Pullen et al., 1995). Other compensatory changesinclude the replacement of Glu70 in CheA by Leu1253 inSpt of ChpA. This substitution compensates for thechange in the active phosphorylation site of His48 inCheA, to Ser1231 in ChpA. These substitutions ensurethat the interactive side chain of Ser1231 is orientated sothat it may be exposed to the solvent interface and allowit to be phosphorylated. Arg1235 of ChpA is shown inFig. 3B in an orientation consistent with the proposed
activated state of Ser1231. In this conformation thecharged side chain of Arg1235 would neutralize that of aphosphorylated serine at position 1231.
These analyses predict that at least the HPt1, HPt4 andthe SPt domains of ChpA have three-dimensional struc-tures which are consistent with a role in phosphotransfer.Given the high degree of sequence conservation amongstthe ChpA phosphotransfer domains we predict that eachwould have conserved three-dimensional structures. Weare currently working toward experimentally demonstrat-ing that each of the eight predicted HPt/TPt and SPtdomains of ChpA are capable of functioning as phospho-transfer domains. This in silico study also predicts that thesubstitution of serine for histidine in the SPt domain ofChpA is accompanied by compensatory changes in thesurrounding residues.
Phenotypic characterization of PAK pilJ, chpA-E mutants
Sequence analysis of the chp region revealed that ourcollection of PAK Tn5-B21 mutants contained 4 insertionsin pilJ (S43, S115, S154 and S247) and 9 insertions inchpA (S10, S23, S45, S33, S52, S129, S164, S209 andS349) (Fig. 1). Because we had not identified transposoninsertion mutants of chpB, chpC, chpD and chpE we gen-erated allelic exchange mutants of these genes in P.aeruginosa strain PAK (Fig. 1) to assess their phenotypes.
We assayed the pilJ, and chpA-E mutants for sensitivityto the type IV pilus-specific bacteriophage PO4 (Bradley,1973), as described previously (Whitchurch et al., 1991).All mutants demonstrated wild-type titres to this phage(data not shown). The ability of mutants of chpA, chpB,chpC, chpD and chpE to swim through 0.3% agar wasthen tested to determine if any of these components arealso involved in the control of flagella-mediated swimmingmotility. Consistent with observations previously reportedfor mutants of pilG-K, the mutants of chpA-E alsodemonstrate wild-type swimming motility via this assay(data not shown). These observations indicate that theChp chemosensory pathway is not intersecting with theswimming chemotaxis system of P. aeruginosa.
Fig. 3. Homology modelling of the phosphotransfer domains of ChpA.A. Sequence alignments of phosphotransfer domains within ArcB, Ypd1 and CheA used for structural modelling. The phosphorylated histidine of CheA, ArcB and YPD1 and the putative phosphorylated serine in SPt of ChpA are boxed. These alignments were used to assign secondary structures to the ChpA homology model. Alpha helices (a-A to a-E) derived from the ArcB structure are shown as solid lines above the alignment whereas alpha helices derived from the CheA structure (a-A¢ to a-E¢) are shown below the alignment. The boxed amino acids show the conserved position of the active amino acids; His717 in ArcB, His64 in Ypd1, His48 in CheA and the proposed active serine (Ser 1231) in ChpA. Numbers indicate amino acid positions within the indicated protein.B. Ribbon structures of the antiparallel helix core of the HPt (histidine phosphotransfer) domains in ArcB, CheA and Ypd1 and the proposed model for the Spt (serine phosphotransfer) domain of ChpA. Only the amino acid backbone has been shown with the active histidines shown as solid rendered in ball and stick (His717 in ArcB, His64 in Ypd1 and His 48 in CheA). The proposed active serine (Ser 1231 in ChpA) is also rendered in the proposed model of ChpA. Below the ribbon structures is shown the amino acid backbone of the local environment surrounding the active histidines (solid rendered) of ArcB, CheA and Ypd1 as well as the proposed active serine (solid rendered) in ChpA. Specific amino acids surrounding the active residue have also been solid rendered and labelled for reference. The appropriate alpha helices are also labelled.

880 C. B. Whitchurch et al.
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 873–893
The twitching motility phenotypes of the PAK mutantsof pilJ, chpA-E were examined via the subsurface stabassay. In this assay the strain to be examined is stabinoculated through a 1% agar plate to the underlying Petridish. Normal twitching motility results in rapid colonyexpansion at the agar/Petri dish interface, whereas non-twitching mutants such as PAKpilA::TcR (pilin subunitgene mutant) produce no such zone of expansion(Semmler et al., 1999) (Fig. 4). Strains PAKchpB::TcR,PAKchpC::TcR, PAKchpD::TcR and PAKchpE::TcR
exhibit wild-type twitching motility (data not shown). Inter-estingly, the twitching stab assays demonstrated that allof the PAK mutants of pilJ show different twitching motilityphenotypes to that previously described for PAO1 mutantsof this gene. In the PAO1 genetic background, mutants ofpilJ demonstrate no twitching motility via the subsurfacestab assay (Darzins, 1994). However, in the PAK back-ground the pilJ mutants clearly demonstrate some twitch-ing motility though it is highly aberrant (Fig. 4A).
Different effects on twitching motility were observed fordifferent mutant alleles of chpA. Aberrant twitching motilitysimilar to pilJ mutants was observed in chpA mutants S52,S129, S164, S209, which carry transposon insertionswithin the FimL-like domain of ChpA as well as in mutantsS10 and S23 which have insertions between the HPt3 andSpt domains (Fig. 4A). Strain S349 which carries thetransposon insertion between Hpt5 and HPt6 and strainsS45 and S33 which have insertions within the histidinekinase domain of ChpA behave similarly to the non-piliated pilA mutant in this assay and appear to be com-pletely devoid of twitching motility (Fig. 4A). It is interest-ing that more severe twitching motility phenotypes occurin mutants with more C-terminal Tn insertions within chpA.The reason for this is unclear but one possibility is thatthere exists additional chpA transcripts which are initiatedbeyond the transposon insertions sites of S52, S164,S209, S129, S10 and S23, although we only have evi-dence for one isoform of ChpA (Fig. 2). However, as theepitope-tagged ChpA used to confirm the size of full-length ChpA was expressed from a multicopy plasmidunder Plac control, this study does not rule out the pos-sibility that ChpA may also exist in shorter isoforms whoseexpression was undetectable in this study. Alternatively,there may be some readthrough from the transposon or(given the large size of ChpA) some cryptic internal pro-moters in the chpA gene which can produce partiallyfunctional ChpA containing the C-terminal CheY domain,which has shown to be absolutely required for twitchingmotility by site-directed mutagenesis (A. Leech and J. S.Mattick, unpubl. obs.).
We attempted to complement the various chpA mutantsof PAK with the clone pUCPChpA without success. How-ever, this clone also completely abolished twitching motil-ity in wild-type PAK whereas the vector pUCPSK showedno inhibition of twitching motility (data not shown). Whilstwe believe it unlikely that the defects in twitching motilityin all nine of the Tn5-B21 chpA mutants could be due tosecondary mutations elsewhere in the genome, it remainspossible that the defects are due to polar effects on neigh-bouring genes. We addressed these issues by construct-ing an in-frame deletion mutant of chpA in strain PAO1.As was observed for the PAK transposon insertionmutants S349, S33 and S45, this mutant, PAO1DchpAwas completely devoid of twitching motility (data not
Fig. 4. A. Subsurface twitching motility assay of P. aeruginosa strains PAK; PAKpilA::TcR; S247 (pilJ ), S164 (chpA); S23 (chpA); and S33 (chpA); Strains were incubated at 37∞C for 24 h post inoculation (PAK) or for 48 h (PAKpilA::TcR; S247, S164, S23, S33) before staining. The bar represents 1 cm.B. Light microscopy of twitching zones of P. aeruginosa strains PAK and S164. Micrographs of all pilJ and chpA mutants were indistin-guishable from that shown here for S164. Micrographs of chpB, chpC, chpD and chpE mutants were indistinguishable from that of PAK. The bar represents 50 mm.

Chemosensory control of twitching motility in P. aeruginosa 881
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 873–893
shown), confirming that chpA is required for twitchingmotility in P. aeruginosa.
As had been observed with chpA mutants of PAK,attempts to complement PAO1DchpA with pUCPChpAwere unsuccessful as this clone also abolished twitchingmotility in wild-type PAO1 whereas the vector showed noinhibition of this phenotype. The clone pUCPChpA con-tains chpA inserted downstream of the Plac promoter(which is constitutively active in P. aeruginosa). Theseobservations suggest that overexpression of chpA inter-feres with normal twitching motility in wild-type P. aerugi-nosa. In light of this, we cloned chpA into pMMB206, alow copy number vector that carries the repressor lacIand has a weakly expressed PtaclacUV5 promoter(Morales et al., 1991), to generate plasmid pMMBChpA.The presence of pMMBChpA in PAO1 caused only avery mild reduction in the diameters of the twitchingmotility zones relative to the vector control and pMMB-ChpA was able to restore twitching motility toPAO1DchpA to that of PAO1 containing pMMBChpA(data not shown). These observations indicate that highlevels of ChpA expression, as presumably occurs withpUCPChpA, represses twitching motility in wild-type P.aeruginosa. Cloning chpA behind the weakly expressedPtaclacUV promoter in pMMB206 alleviated the repres-sion problem and allowed complementation of twitchingmotility to PAO1DchpA. Interestingly, pMMBChpA wasstill unable to complement twitching motility to the chpAmutants of strain PAK. Both the chpA clones used in thisstudy were subcloned from cosmid clones of PAO1genome sequences, and thus is it possible thatsequence differences between strains PAK and PAO1might explain the inability of these clones to complementPAK mutants. The observation that PAK mutants of pilJshow different twitching motility phenotypes to that ofPAO1 mutants (see above) also suggests that there aresome strain differences in this system.
Time-lapse video microscopy
Time-lapse video microscopy may be used to examine thedynamics of twitching motility at a cellular level (Semmleret al., 1999). Such studies have revealed that twitchingmotility is a complex social process in which the bacteriamove in a highly coordinated fashion, initially forming raftsof cells which move away from the colony edge, behindwhich an intricate lattice-like network of cells is formed(see Fig. 4B). Cells within this network follow each otherclosely (via cell-cell contact/pili extension and retraction)and demonstrate frequent reversals of direction. Cellswithin the outgoing rafts also demonstrate some cell rever-sal but do so less frequently than those in the network.Non-twitching mutants demonstrate no differentiation ofthe colony edge (Semmler et al., 1999).
Using this time-lapse video microscopy technique,we examined the twitching motility of selected mutants ofpilJ, chpA-E. As expected, we found that those mutantswhich produce normal twitching zones via the subsur-face stab assay (i.e. PAKchpB::TcR, PAKchpC::TcR,PAKchpD::TcR and PAKchpE::TcR) also appear tobehave normally when examined microscopically (datanot shown). The pilJ mutant S247 and chpA mutants S164and S23 which demonstrate similar aberrant twitchingmotility via the stab assay also showed similar phenotypeswhen examined microscopically (Fig. 4B). These mutantsappear capable of the early stages of twitching motilityforming large, exaggerated rafts of cells which move awayfrom the colony edge but lack the characteristic latticeformation. These rafts move at rates of about 0.2 mmmin-1 which is about 20 times slower than the rate at whichwild-type leading edge rafts move, and only about twiceas fast as the rate of colony expansion as the result of celldivision as calculated from the non-motile PAKpilA::TcRmutant (Semmler et al., 1999). Assuming that the overallrate of translocation of the leading edge rafts is governedby both the rate of individual cell motilities as well as thefrequency of cellular reversals, it is therefore conceivablethat the reduced rate of leading edge translocation couldbe due to either or both a reduction in individual cellmotilities and/or increased rates of reversal frequencies.Although it is difficult with our time-lapse video microscopytechnique to track individual cells in the large, slow mov-ing, densely packed, leading edge rafts, it does seemclear that the rate of individual cell motilities is significantlydecreased relative to wild type and that there is no evi-dence for increased reversal frequencies. Indeed, unlikethe wild-type situation where cells within the leading edgerafts can be readily seen to reverse direction and moveagainst the overall direction of the raft, S247 (pilJ ), S23(chpA) and S164 (chpA) show no detectable cellularreversals. These observations suggest that the Chp sys-tem controls at least the rate of cellular movements andpossibly also the frequency of cell reversal during theprocess of twitching motility.
Interestingly, the mutants of chpA (S349, S45 and S33),which show no twitching motility macroscopically, alsoform large rafts of cells (Fig. 4B). These rafts move at thesame rate as that of colony expansion measured in thenon-twitching pilA mutant. Unlike PAKpilA::TcR whichforms an even colony edge (Semmler et al., 1999), thecells of the chpA mutants (S349, S45 and S33) are ableto assemble into the raft-like projections which are seenat the leading edge of twitching zones. However, the cellswithin these assemblies appear to be non-motile andshow no detectable cellular reversals. Thus it appears thatthe defects in colony expansion seen in the various pilJand chpA mutants are likely accounted for by decreasedrates of cellular motility. This is consistent with our hypoth-

882 C. B. Whitchurch et al.
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 873–893
esis that the Chp chemosensory system influences therate of expeditionary radiation of the colony through con-trol of twitching motility in response to environmental cues.
Pilin production
The PAK mutants of pilJ, chpA, chpB, chpC, chpD andchpE can be categorized based on their twitching motilityphenotypes. Pili production by the chpC, chpD and chpEmutants which demonstrate normal twitching motility was
assessed by ELISA of whole cells using antisera againstthe major pilin subunit PilA (Fig. 5A). These analyses indi-cated that the PAKchpB::TcR mutant is hyperpiliated rel-ative to wild-type PAK, which is not surprising if the normalfunction of ChpB, by analogy to CheB in E. coli, is tomediate a feedback circuit for sensory adaptation.The PAKchpC::TcR, PAKchpD::TcR and PAKchpE::TcRmutants produce less surface pili than wild-type with thechpC mutant demonstrating the most significant decreasein surface piliation. Western analysis of whole cells using
Fig. 5. Pilin expression in wild type and P. aeruginosa mutant strains.A. ELISA of whole cells from the Pseudomonas strains PAK ( ); PAKpilA::TcR ( ); S247 (pilJ ) ( ); S164 (chpA) ( ); S23 (chpA) ( ); S33 (chpA) ( ); PAKchpB::TcR ( ); PAKchpC::TcR ( ); PAKchpD::TcR ( ) and PAKchpE::TcR ( ). Pilin was detected with anti-PilA serum and is indicative of the levels of surface pili in these strains.B. Immunoblot of pilin found in whole cell (WC) preparations after surface pili have been sheared (top panel) of strains PAK (lane 1); PAKpilA::TcR (lane 2); S247 (pilJ, lane 3); S164 (chpA, lane 4); S23 (chpA, lane 5); S33 (chpA, lane 6); PAKchpB::TcR (lane 7); PAKchpC::TcR (lane 8); PAKchpD::TcR (lane 9) and PAKchpE::TcR (lane 10). Immunoblot of pilin found in sheared surface pili preparations (bottom panel) of strains PAK (lane 1); PAKpilA::TcR (lane3); R306 (pilV, lane 4); S247 (pilJ, lane 5) S164 (chpA, lane 6); and S23 (chpA, lane 7). The pilV mutant strain which is defective in assembly of pili (Alm and Mattick, 1995) was included in these assays to control for the contribution of pilin to the surface pili samples as a result of cell lysis.C. PpilA::xylE transcriptional reporter assays. Catechol dioxygenase activity was determined for wild-type PAK, pilJ, chpA, chpB, chpC, chpD and chpE mutants carrying the PpilA::xylE transcriptional fusion reporter plasmid pMIC66. The pilR mutant (R94) carrying pMIC66 was included as a negative control as the transcriptional activator PilR is required for pilA transcription (Hobbs et al., 1993). Mean activities from three independent experiments are shown and error bars (where visible) represent the standard deviation of the mean.D. ELISA of whole cells from the Pseudomonas strains PAK + pUCP19 (�); PAK + pAW103 ( ); S33 (chpA) + pUCP19 ( ); S33 (chpA) + pAW103 ( ). Pilin was detected with anti-pilin serum and is indicative of the levels of surface pili in these strains. The ELISAs and Westerns shown here are representative of results obtained from multiple assays.

Chemosensory control of twitching motility in P. aeruginosa 883
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 873–893
anti-PilA antisera show that these mutants produce wild-type levels of cell-associated pilin (Fig. 5B). Northernanalysis (data not shown) and PpilA::xylE transcriptionalreporter assays (Fig. 5C) indicate that these mutants alsoshow wild-type levels of pilA transcription. These obser-vations suggest that the hyperpiliated phenotype of thechpB mutant and the reduced levels of surface piliationobserved in the chpC, chpD, chpE mutants is not occur-ring at the level of pilin production but is probably due toaltered rates of pilus assembly and/or retraction. However,these alterations in levels of surface piliation do not trans-late into obvious defects in twitching motility under theconditions of our assay.
The PAK mutants of pilJ (represented here by S247)and N-terminal chpA (represented by S164 and S23)which have aberrant twitching motilities, show little to noproduction of surface pili by the whole cell ELISA(Fig. 5A). However, Western blots of sheared surface piliusing anti-PilA antiserum demonstrate that these pilJ andchpA mutants do have a small but detectable amount ofsurface piliation (Fig. 5B) which is presumably sufficientfor PO4 phage sensitivity and for mediating the smalldegree of aberrant twitching motility seen in thesemutants. Whole cell Western analysis, using an anti-PilAantiserum, shows that these strains produce normal levelsof cell-associated pilin (Fig. 5B). Northern analysis (datanot shown) and transcriptional PpilA::xylE promoterreporter assays (Fig. 5C) demonstrate that these mutantsshow wild-type levels of pilA expression. These resultsindicate that the alterations in levels of surface pili seenin these mutants are not due to defects in pilin productionwhich suggests that the defects are occurring at the pointof pilus extension and/or retraction.
The non-twitching chpA mutants S349, S45 and S33(represented here by S33) showed little to no detectablesurface pili by ELISA (Fig. 5A). These mutants also nor-mally showed almost undetectable levels of pilin in whole-cell samples (Fig. 5B; see below). Northern analysis (datanot shown) and PpilA::xylE promoter reporter assays(Fig. 5C) indicated that the defect in pilin production in thisclass of chpA mutant is due to reduced pilA transcription,although this was variable (see below). However extendedexposure of both the whole cell Western and Northernanalyses (data not shown) indicated that a small amountof pilin was produced by these chpA mutants, which ispresumably sufficient for mediating sensitivity to the pili-specific bacteriophage PO4. Constitutive expression ofthe pilin gene pilA from the plasmid pAW103 restoresproduction of surface pili to S33 as shown by ELISA(Fig. 5D). This result indicates that ChpA is not requiredfor the expression of other components of the pilus struc-ture or assembly machinery. However, in trans expressionof pilA with pAW103, which restores twitching motility topilA mutants (Watson et al., 1996b), does not complement
twitching motility to chpA mutants (data not shown) indi-cating that ChpA is also required for other aspects oftwitching motility.
It should be noted, however, that during our studies wefound that the chpA mutants S349, S45 and S33 at timesproduced wild-type levels of cell-associated PilA as shownby both PpilA::xylE reporter and whole cell Western (datanot shown). In these instances however, surface piliationremained low and these mutants remained completelynon-twitching, in contrast to the aberrant twitching motilityphenotypes observed with the pilJ (S247) and N-terminalchpA mutants (S164, S23). We have not been able toaccount for the variable levels of pilA expression in theseC-terminal chpA mutants. We can only speculate thatsome variability in media batches or other environmentalcondition in different experiments was responsible for thealtered pilA expression phenotypes.
The Chp system is required for cytotoxicity of epithelial monolayers in vitro
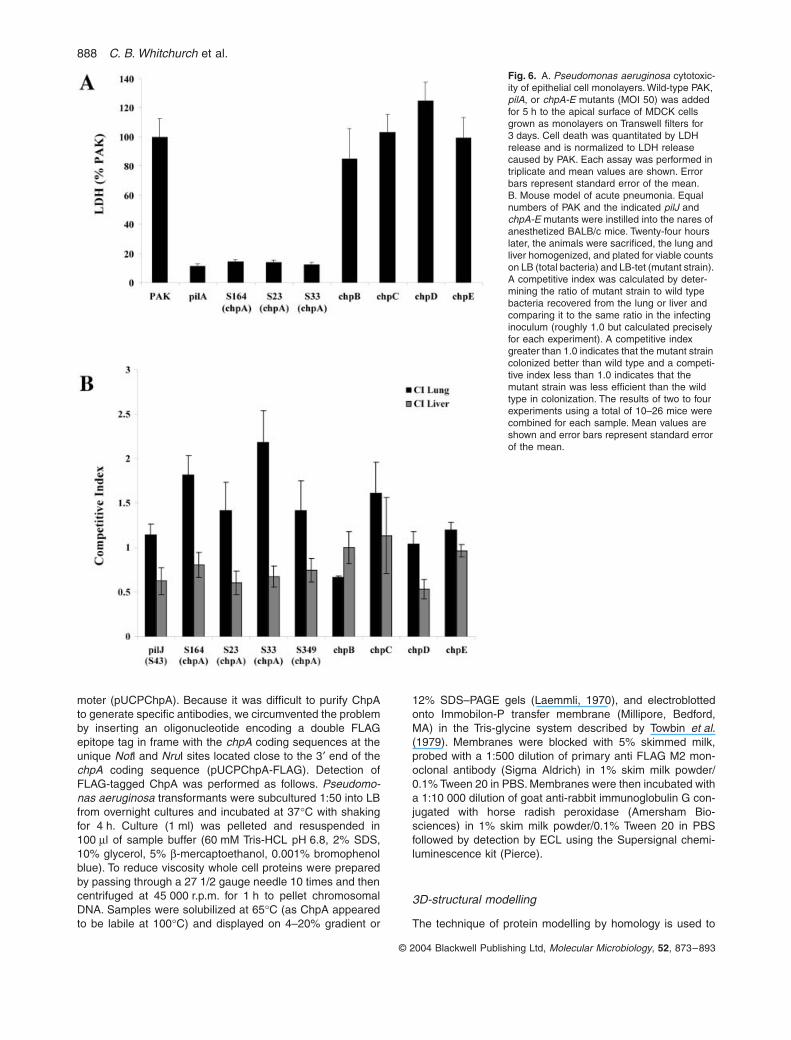
Previous studies have indicated a role for functional typeIV pili in P. aeruginosa interactions with cultured cell linesas well as for virulence in animal models of disease(Comolli et al., 1999; Kang et al., 1997). Given the involve-ment of the chp cluster gene products in twitching motility,we tested whether representative mutants of the chp clus-ter affected the ability of PAK to damage epithelial cells invitro. Transposon insertion mutants of chpA (S164 whichcontains an insertion into the FimL-like domain; S23 whichcontains an insertion between HPt3 and SPt, and S33which contains a C-terminal insertion in the histidinekinase domain) as well as allelic exchange mutants ofchpB, chpC, chpD and chpE were added to the apicalsurface of MDCK cells grown as polarized monolayers onTranswell filters. Compared to PAK, the cytotoxicity of thechpB, C, D, and E mutants, which have normal twitchingmotility, was not significantly different from wild type(Fig. 6A). All of the chpA mutants (S164, S23, and S33)showed greatly reduced cytotoxicity, with the magnitudeof the decrease being similar to that observed for the pilAmutant (Fig. 6A). Together with our previous observationsthat piliated but non-twitching strains of P. aeruginosademonstrate similarly reduced cytotoxicity (Comolli et al.,1999), these results confirm that cytotoxicity of epithelialcells by P. aeruginosa requires fully functional type IV pili.
The Chp system contributes to full virulence in a mouse model of acute pneumonia
The role of chp genes in virulence was assessed in amouse model of acute pneumonia by two approaches.First, the mortality of mice intranasally inoculated withPAK, pilA, or the pilJ (S43) and the chpA (S164, S23, S33

884 C. B. Whitchurch et al.
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 873–893
and S349) mutants was followed over a 7-day period.While none of the mice inoculated with the pilA strain died,no difference in mortality between wild-type PAK and thepilJ and chpA mutants was observed (data not shown).
Using colonization as a more sensitive measure of vir-ulence, we performed competition assays which allowedus to compare the efficiency of colonization of the lung orliver of the pilJ and chpA-E mutant strains to that of thewild type (Fig. 6B). A competitive index was calculated bygenerating the ratio of mutant strain to wild type bacteriarecovered from the lung or liver and comparing it to thesame ratio in the infecting inoculum (roughly 1.0 but cal-culated precisely for each experiment). A competitiveindex greater than 1.0 indicates that the mutant straincolonized better than wild type and a competitive indexless than 1.0 indicates that the mutant strain was lessefficient than the wild type in colonization. Interestingly,the chpA mutants were more effective in colonizing thelung compared to wild type (C.I. > 1.5). However, theywere less able to spread to and/or colonize the liver. Thiswas best appreciated when the ratio of the lung C.I. toliver C.I. was compared, which for the pilJ and chpAmutants was <0.6 (Fig. 6B and data not shown). Interest-ingly the chpD mutant also showed a defect in spread tothe liver though this strain shows wild-type twitching motil-ity. This suggests that chpD, which is predicted to encodea protein belonging to the AraC class of transcriptionalactivators, plays a role in the spread of P. aeruginosa tothe liver in vivo.
Discussion
The proteins PilG-K, ChpA-E appear to constitute a com-plex chemosensory system (Chp) which is analogous tobacterial chemotaxis systems which control swimmingmotility in response to environmental stimuli (Falke et al.,1997; Djordjevic and Stock, 1998; Armitage, 1999; Bour-ret and Stock, 2002). The P. aeruginosa Chp system con-sists of a putative methyl accepting chemoreceptor PilJ;a methyl transferase CheR-like protein PilK, 2 CheWhomologues (PilI and ChpC) and a methyl esterase CheBhomologue ChpB. The Chp system also has three CheYresponse regulators modules: PilG and PilH and a domainlocated at the C-terminus of the CheA-like histidine kinaseof this pathway, ChpA. ChpA is a complex protein withnine putative sites of phosphorylation: six histidines, athreonine and a serine, each located in conserved HPt-like phosphotransfer domains, and an aspartate in a C-terminal CheY receiver domain. It is likely that ChpA isresponsible for phosphotransfer reactions with PilG andPilH as well as an intramolecular phosphotransfer to itsCheY domain. By analogy with the well-characterized Chesystems in which CheY interacts with the flagella motorto regulate swimming motility, either or both of the CheY
homologues PilG and PilH may interact with the putativetype IV pilus motor to control twitching motility. The Chpsystem, like many other bacterial chemotaxis systems,lacks a homologue of the enteric CheZ which serves toaccelerate dephosphorylation of phosphorylated CheY.Other chemotaxis systems such as that of Rhodobactersphaeroides utilize multiple CheY proteins to act as phos-phate sinks to add a similar layer of regulation to thesystem (Armitage, 1999). It is possible that either of PilGor PilH and/or the C-terminal CheY domain of ChpAserves a similar function in this system.
We predict that the Chp chemosensory pathway con-trols twitching motility in response to environmental cuesin a manner analogous to the chemotactic control of swim-ming motility. The P. aeruginosa genome encodes a totalof 26 putative methyl accepting chemotaxis proteins (Croftet al., 2000; Stover et al., 2000). As the Chp system pos-sesses two CheW homologues (PilI and ChpC), it is pos-sible that these serve to complex a number of MCPs tothe histidine kinase ChpA. This model provides for theinput of multiple environmental signals which are inte-grated by the Chp system to control twitching motility.Although much is known about the molecular genetics oftwitching motility in P. aeruginosa, little is known about theenvironmental cues that control this motility. Our observa-tions in combination with previous studies (Darzins, 1993,1994, 1995) have demonstrated that pilI, pilJ, and chpAbut not chpC are required for normal twitching motility. Byanalogy with the well-characterized chemotaxis systemsof enteric bacteria, these observations suggest that PilImay be the adaptor protein which couples at least theMCP PilJ to ChpA and that this complex is essential forthe control of twitching motility. It is possible that thesecond CheW homologue of this system, ChpC, servesto complex other MCPs to ChpA to facilitate the input ofmultiple environmental cues into this chemosensory path-way. However, these complexes are not essential for thecontrol of twitching motility at least under the conditionsassayed in this study.
It is also now clear, both morphologically and geneti-cally, that twitching motility and the social aspect of glidingmotility, such as occurs in Myxococcus xanthus, areessentially the same process (Semmler et al., 1999; Mat-tick, 2002). The Chp system we have described hereresembles in many aspects the Frz system of M. xanthuswhich also has multiple CheY response regulators mod-ules found in FrzZ (which is a fusion of two CheY domains)and the C-terminus of the CheA/CheY hybrid FrzE (Wardand Zusman, 1999). FrzZ and FrzE then are probablyanalogous in function to PilG, PilH and ChpA. The Frzsystem also has homologues of PilJ (FrzCD), PilK (FrzF),ChpB (FrzG) and PilI and ChpC (FrzA, FrzB) (Ward andZusman, 1999). FrzB has been previously thought to bea protein unique to the Frz system but our analyses indi-

Chemosensory control of twitching motility in P. aeruginosa 885
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 873–893
cate that this protein is likely to be a second CheW (PilI/ChpC) homologue of this system. However, the M. xan-thus Frz system differs from the Chp system in that thehistidine kinase of the system FrzA possesses only asingle site of autophosphorylation in contrast to the eightpredicted phosphotransfer domains identified in ChpA.The Frz system and chemotaxis systems of other bacteriaalso lack homologues of ChpD and ChpE. The function (ifany) of these two proteins within the Chp chemosensorypathway is currently unclear.
The M. xanthus Frz system controls the reversal fre-quency of cells during gliding motility. Cellular reversalsare thought to be due to switching of the sites of piliextrusion from one pole of the bacterium to the other (Sunet al., 2000). We predict that the Chp system would playa similar role in the control of twitching motility in P. aerug-inosa. Indeed, time-lapse video microscopy suggests thatthe Chp system controls both the rate of cellular move-ments and the frequency of cellular reversals duringtwitching motility.
Cellular movements during twitching motility in P. aerug-inosa and N. gonorrhoeae have recently been shown tobe mediated by pilus extension and retraction (Merz et al.,2000; Skerker and Berg, 2001). It possible that the Chpchemosensory system exerts its influence on twitchingmotility through control of pilus assembly and/or retrac-tion. Our studies have demonstrated that Chp systemmutants show altered levels of surface piliation. The hyp-erpiliated phenotype of chpB mutants could conceivablybe caused by either a reduced rate of pilus retraction oran increased rate of pilus assembly. Conversely mutantsof pilJ, chpA and chpC are significantly reduced in theirlevels of surface piliation which may be accounted for byincreased retraction or reduced assembly rates. We pro-pose that in a manner analogous to CheY interaction withthe flagella motor to control swimming motility in bacteria,that the CheY homologues of this system PilG and/or PilHinteract with the putative pilus motor to control pilus exten-sion and retraction. This could occur through direct inter-action with the nucleotide binding proteins PilB, PilT andPilU which are thought to control pilus extension (PilB)and retraction (PilT/U) (Mattick, 2002). In addition to directinteractions with the pili motor we have found that the Chpsystem (via ChpA) also influences type IV pili biogenesisthrough control of pilA expression.
Motility in the cyanobacterium Synechocystis PCC6803has recently been shown to be mediated by type IV pili(Bhaya et al., 2000). Phototactic control of this motility ismediated by two chemosensory pathways referred to asTax1 and Tax3 (Bhaya et al., 2001). Like the Chp and Frzsystems both the Tax1 and Tax3 pathways possess ahybrid CheA/CheY histidine kinase (TaxAY1 and TaxAY3).Neither of these proteins is as complex as ChpA althoughTaxAY1 does possess two HPt domains. Each of these
systems also have 3 CheY domains represented by TaxY1and TaxY3, the C-terminal domain of TaxAY1 and TaxAY3and a domain located at the C-termini of TaxP1 andTaxP3. Like the Frz and Chp systems the Tax1 systempossesses two CheW homologues: TaxW1 and the pre-dicted product of the gene with Cyanobase designationsll0044 (which had not been recognized as a CheWhomologue and is located directly downstream of taxAY1).Interestingly, searches of the completed SynechocystisPCC6803 genome indicates that genes encoding homo-logues of CheB or CheR are not present in the Syn-echocystis PCC6803 genome. Mutants of the Tax1 andTax3 systems show defects in the biogenesis of type IVpili (Bhaya et al., 2001). It therefore appears that the Tax1and Tax3 chemosensory systems of SynechocystisPCC6803 intersect to control type IV pili biogenesis andtwitching motility in a similar manner to the Chp systemof P. aeruginosa.
Our database analyses have indicated that the genomesequences of N. europaea, X. fastidiosa, R. metallidurans,R. solanacearum, X. campestris, X. axonopodsi, M.degredans, P. putida, P. syringae and P. fluorescens eachencode a multi-Hpt domain containing CheA/CheY hybrid.Analysis of the preliminary genome sequences of Diche-lobacter nodosus (kindly provided by TIGR) indicates thatthis organism also encodes a ChpA homologue whichcontains at least three HPt domains and an SPT domain.These proteins are highly similar to one another andto P. aeruginosa ChpA and appear to represent a distinctsubclass of CheA-like histidine kinases. All of the ChpAhomologues (except those found in Ralstonia and Xanth-omonas species) also contain putative TPt and/or SPtdomains. This suggests that the presence of a potentiallyphosphorylatable serine or threonine in this type of phos-photransfer domain may be an as yet unrecognized con-served feature of some CheA-like bacterial kinases.
Where sufficiently assembled sequences are availableit is evident that the similarity between the ChpAhomologues extends to gene organization. The codingsequence for the ChpA homologue of M. degredansresides in a gene cluster that encodes homologues ofeach of PilG, PilH, PilI, PilJ, PilK, ChpB and ChpC in thesame organization as the P. aeruginosa Chp cluster (seeFig. 1). ChpA homologues of D. nodosus, P. fluorescens,P. putida, and P. syringae are also encoded in a conservedgene clusters with conserved gene order except thatthese systems are missing homologues of both PilK andChpB. The Chp clusters of X. fastidiosa, X. axonopodisand X. campestris are missing PilK homologues but doencode a distantly related homologue of ChpB. X. fastid-iosa is also missing a PilH homologue. The R. solan-acearum cluster is missing homologues of PilK, ChpB andChpC. Except for the D. nodosus sequences (which arepreliminary and may contain frame-shift errors) and the

886 C. B. Whitchurch et al.
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 873–893
ChpB homologues X. fastidiosa, X. axonopodis and X.campestris, each orthologous protein in these clusters arehighly similar (80–90%) across their entire length. Inter-estingly, none of these other Chp clusters encode homo-logues of the P. aeruginosa proteins ChpD and ChpEwhich belong to the AraC family of transcriptional regula-tors and the LysE family of amino acid efflux proteins,respectively. This suggests either that the role of theseproteins is specific to the P. aeruginosa system or thatthey are not bona fide components of this pathway. OurP. aeruginosa chpD and chpE mutants show no apparentdefects in twitching motility and only mild perturbation inpili biogenesis and thus the role of these proteins in thischemosensory pathway is unclear.
Further genome analysis predicts that all of these bac-teria possess type IV pili so it is likely that the Chp systemin each is involved in the regulation of twitching motilitysimilarly to that described here for P. aeruginosa. How-ever, our database analyses indicate that the majority ofbacteria (with available genome sequences) that possessTfp including Neisseria gonorrhoeae, N. meningitidis,Legionella pneumophila, Shewanella putrefaciens, S.oneidensis MR-1, Vibrio cholerae, V. parahaemolyticusand V. vulnificus do not encode an orthologous Chp sys-tem. Interestingly, the bacteria that do have identifiableChp systems are found in soil, water and/or plants, withthe possible exception of D. nodosus which is consideredan obligate pathogen albeit in an agricultural context.
Our studies have also indicated that the Chp system isrequired for cytotoxicity to epithelial cell monolayers andfor full virulence in a mouse model of acute pneumonia.We had previously identified a transposon insertion mu-tant of chpA in a screen for P. aerugionosa PA103 mutantsdefective in epithelial cell injury (Kang et al., 1997). Herewe show that P. aeruginosa PAK mutants of pilJ, chpA andchpD are less able to spread to and/or colonize the liverand that chpA mutants are more effective in colonizing thelung compared to wild type. It is possible that the observedreduction in colonization of the liver and enhanced colo-nization of the lungs seen in these mutants is related tochemotactic control of twitching motility by this system.Interestingly, in a recent screen for in vivo regulators ofVibrio cholerae virulence it was found that motile (swim-ming) but non-chemotactic mutants of V. cholerae showedenhanced colonization of the mouse intestine althoughwith aberrant distribution (Lee et al., 2001). Our observa-tions also suggest that the putative transcriptional regula-tor ChpD plays some role in the ability of P. aeruginosa tospread to and/or colonize the liver.
Interestingly, components of the Chp system wererecently identified in a genetic screen for P. aeruginosamutants that were defective in killing of the fruit fly Droso-phila melanogaster (D¢Argenio et al., 2001). This studydemonstrated that the requirement for the Chp system in
Drosophila killing was separate from its involvement intwitching motility. These investigators proposed that theChp system may control the expression of other P. aerug-inosa virulence factors. We are currently investigating themechanisms via which the Chp signal transduction sys-tem controls twitching motility and how this chemosensorysystem coordinates the expression of other virulencedeterminants of P. aeruginosa. However, it is clear that P.aeruginosa has evolved a highly complex chemosensorysystem to control twitching motility in response to environ-mental signals and which, either directly or indirectly, coor-dinates twitching motility with the expression of manyother virulence determinants of this pathogen.
Experimental procedures
Bacterial strains, plasmids and media
The E. coli strain DH5a (recA endA1 gyrA96 hsdR17 thi-1supE44 relA1 f80 dlacZDM15) was used in all genetic manip-ulations and in the preparation of DNA sequencing templates.Escherichia coli strain S17-1 was used as the donor strain inthe bacterial conjugation. The P. aeruginosa strains usedwere PAO1 strain ATCC 15692 (American Type Culture Col-lection); PAK (D. Bradley, Memorial University of Newfound-lands, St John’s, Canada), Tn5-B21 mutants of this strain(Hobbs et al., 1993), including the previously characterizedpilR mutant R94 (Hobbs et al., 1993); pilV mutant R306 (Almand Mattick, 1995); and PAKpilA::TcR (previously referred toas AWK; Watson et al., 1996b). A PAO1 cosmid library (kindlyprovided by B. Holloway, Monash University, Clayton, Austra-lia; Ratnaningsih et al., 1990; Huang et al., 2000) was usedin the subcloning and sequence analysis of the chp region.The preparation of E. coli competent cells and transformationprotocols were followed according to Sambrook et al. (1989).Preparation of P. aeruginosa competent cells and transfor-mations were performed as described previously (Matticket al., 1987). Plasmids used in this study are listed in Table 1.Escherichia coli and P. aeruginosa liquid cultures were main-tained in Luria–Bertani (LB) broth and solid media was pre-pared by adding 1–1.5% agar. The media used in the lightmicroscopy contained 4 g l-1 tryptone, 2 g l-1 yeast extract,2 g l-1 NaCl, 1 g l-1 MgSO4·7H2O and 8 g l-1 GelGro (ICN) asa solidifying agent. The following antibiotic concentrationswere used for the selection of E. coli: tetracycline–12.5 mg ml-1 for plasmid selection and 40 mg ml-1 forcosmid selection; ampicillin 100 mg ml-1, chloramphenicol25 mg ml-1 and kanamycin 50 mg ml-1. The concentration ofantibiotics for the selection of P. aeruginosa were carbeni-cillin 250 mg ml-1, chloramphenicol 250 mg ml-1, rifampicin20 mg ml-1 and tetracycline 200 mg ml-1.
Recombinant DNA techniques and sequence analysis
The preparation of plasmid DNA, restriction endonucleasedigestion (New England Biolabs), ligation reactions, Southernblotting, and radiolabelling of probe were carried out usingstandard protocols (Sambrook et al., 1989). Sequence tem-plates were generated by a combination of subcloning and

Chemosensory control of twitching motility in P. aeruginosa 887
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 873–893
shotgun cloning strategies. The dsDNA was prepared forsequencing using a modified alkaline lysis method involvingPEG precipitation (Applied Biosystems). The sequencing wasperformed using the dideoxy chain termination Applied Bio-systems PRISM on a 373A automated sequencer. BLAST
searches of the GenBank databases were performed atNCBI.
Identification of transposon insertions
Genomic sequences flanking the site of insertion of the trans-poson in mutant S45 was cloned using a procedure termed‘marker rescue’ which involved digestion of chromosomalDNA from the S45 mutant with BglII (which does not cutwithin the transposon) and HindIII (which cuts once within thetransposon beyond the tetracycline resistance marker); liga-tion into pBluescriptII (Stratagene) and recovery of tetracy-cline resistant E. coli colonies. The cloned genomic DNAflanking the site of transposon insertion was used to screena reference PAO1 genomic cosmid library identifying fourcosmids (pMO02116, pMO02218, pMO01109, pMO01539)which contained wild-type DNA of this region.
Southern analysis of KpnI chromosomal digests of the Tn5-B21 library of twitching motility mutants using the insert ofcosmid pMO02218 as a probe identified another 12 mutants(S10, S23, S33, S52, S154, S247, S43, S115, S129, S164,S209 and S349) with transposon insertions within this region.Marker rescue clones of the DNA flanking the site of trans-poson insertion in each of these 12 mutants was cloned bydigestion with EcoRI or HindIII. The DNA adjacent to thetransposon insertion in each of the mutants were thensequenced using a primer which is specific for the invertedrepeats of Tn5-B21. Four of the mutants (S43, S115, S154,and S247) had transposon insertions in the previouslydescribed gene pilJ whereas the remainder had inserted intonovel sequences. Further mapping via Southern analysisrevealed that the mutants S10, S23, S33, S45, S52, S129,S164, S209, and S349 contained transposon insertionsdownstream of pilK.
Expression and immunoblotting of FLAG-tagged ChpA
To confirm the size of ChpA, the coding sequence for chpAwas cloned into pUCPSK under the control of the Plac pro-
Table 1. Plasmids used in this study.
Plasmid Relevant characteristics Reference
pSM-TET Source of TcR cassette Mongkolsuk et al. (1993)pRIC380 P. aeruginosa suicide vector Alm and Mattick (1996)pJEN34 pRIC380 with EZ::TN < TET-1 > in b lactamase gene; TcR This studypUK21, pOK12 E. coli cloning vectors, KmR Vieira and Messing (1991)pUC21 E. coli cloning vectors, ApR Vieira and Messing (1991)pUCPSK/KS P. aeruginosa – E. coli shuttle vectors, ApR Watson et al. (1996a)pBluescript II SK/KS E. coli cloning vector, ApR StratagenepGEM-T E. coli cloning vector, ApR PromegapUCP19 P. aeruginosa – E. coli shuttle vector, ApR Schweizer (1991)pMMB206 Broad host range vector with inducible PtalacUV promoter, CmR Morales et al. (1991)pAW103 1.2 kb HindIII fragment containing PAK pilA in pUCP19 Watson et al., (1996b)X1918 Source of promoterless xylE reporter gene cassette Schweizer (1993)pMO02116, pMO02218,
pMO01109, pMO01539pLA2917 containing partial Sau3A PAO1 chromosomal DNA fragments.
These cosmids contain regions of the PAO1 genome that span the pilG-K, chpA-E cluster.
Ratnaningsih et al. (1990)
pAW153 1.5 kb KpnI/NotI fragment containing chpB in pOK12 This studypMIC9719 4 kb BglII/SacI fragment from pMO01539 containing region
downstream of chpB cloned into pUK21This study
pMIC86 pAW153 with TcR cassette cloned in ClaI site of chpB This studypMIC9722 pMIC9719 with TcR cassette cloned into KpnI site of chpC This studypMIC69 pMIC9719 with TcR cassette cloned into XhoI site of chpD This studypMIC34 pMIC9719 with TcR cassette cloned into ApaI site sof chpE This studypMIC87 pRIC380 carrying chpB::TcR from pMIC86 on SpeI fragment This studypMIC9723 pRIC380 carrying chpC::TcR from pMIC9722 on SpeI fragment This studypMIC79 pRIC380 carrying chpD::TcR from pMIC69 on SpeI fragment This studypMIC35 pRIC380 carrying chpE::TcR from pMIC34 on SpeI fragment This studypRIC221 470 bp HindIII/BspHI fragment covering the pilA promoter cloned into
HindIII/NcoI sites of pUC21This study
pMIC63 PpilA::xylE trancriptional reporter; xylE from X1918 cloned as a BamHI fragment into the BglII site (downstream of pilA promoter) in pRIC221
This study
pMIC66 SpeI fragment from pMIC63 cloned into pUCPKS This studypUCPChpA 8 kb SacI/ClaI fragment from pMO01539 containing chpA in pUCPSK This studypUCPChpA-FLAG pUCPChpA- + double FLAG tag inserted into NotI/NruI sites This studypMMBChpA 8 kb SacI/ClaI fragment from pMO01539 containing chpA in pMMB206 This studypJB2 pGEM-T containing 1 KB region 5¢ of chpA This studypJB3 pGEM-T containing 1 KB region 3¢of chpA This studypJB4 EcoRI/HindIII fragment from pJB2 and HindIII/XbaI fragment from pJB3
concatamerized and cloned into EcoRI/XbaI sites of pOK12This study
pJB5 pJEN34 carrying DchpA from pJB4 on SpeI fragment This study
Antibiotic resistance abbreviations: Tc, tetracycline; Km, kanamycin; Ap, ampicillin; Cm, chloramphenicol.
888 C. B. Whitchurch et al.
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 873–893
moter (pUCPChpA). Because it was difficult to purify ChpAto generate specific antibodies, we circumvented the problemby inserting an oligonucleotide encoding a double FLAGepitope tag in frame with the chpA coding sequences at theunique NotI and NruI sites located close to the 3¢ end of thechpA coding sequence (pUCPChpA-FLAG). Detection ofFLAG-tagged ChpA was performed as follows. Pseudomo-nas aeruginosa transformants were subcultured 1:50 into LBfrom overnight cultures and incubated at 37∞C with shakingfor 4 h. Culture (1 ml) was pelleted and resuspended in100 ml of sample buffer (60 mM Tris-HCL pH 6.8, 2% SDS,10% glycerol, 5% b-mercaptoethanol, 0.001% bromophenolblue). To reduce viscosity whole cell proteins were preparedby passing through a 27 1/2 gauge needle 10 times and thencentrifuged at 45 000 r.p.m. for 1 h to pellet chromosomalDNA. Samples were solubilized at 65∞C (as ChpA appearedto be labile at 100∞C) and displayed on 4–20% gradient or
12% SDS–PAGE gels (Laemmli, 1970), and electroblottedonto Immobilon-P transfer membrane (Millipore, Bedford,MA) in the Tris-glycine system described by Towbin et al.(1979). Membranes were blocked with 5% skimmed milk,probed with a 1:500 dilution of primary anti FLAG M2 mon-oclonal antibody (Sigma Aldrich) in 1% skim milk powder/0.1% Tween 20 in PBS. Membranes were then incubated witha 1:10 000 dilution of goat anti-rabbit immunoglobulin G con-jugated with horse radish peroxidase (Amersham Bio-sciences) in 1% skim milk powder/0.1% Tween 20 in PBSfollowed by detection by ECL using the Supersignal chemi-luminescence kit (Pierce).
3D-structural modelling
The technique of protein modelling by homology is used to
Fig. 6. A. Pseudomonas aeruginosa cytotoxic-ity of epithelial cell monolayers. Wild-type PAK, pilA, or chpA-E mutants (MOI 50) was added for 5 h to the apical surface of MDCK cells grown as monolayers on Transwell filters for 3 days. Cell death was quantitated by LDH release and is normalized to LDH release caused by PAK. Each assay was performed in triplicate and mean values are shown. Error bars represent standard error of the mean.B. Mouse model of acute pneumonia. Equal numbers of PAK and the indicated pilJ and chpA-E mutants were instilled into the nares of anesthetized BALB/c mice. Twenty-four hours later, the animals were sacrificed, the lung and liver homogenized, and plated for viable counts on LB (total bacteria) and LB-tet (mutant strain). A competitive index was calculated by deter-mining the ratio of mutant strain to wild type bacteria recovered from the lung or liver and comparing it to the same ratio in the infecting inoculum (roughly 1.0 but calculated precisely for each experiment). A competitive index greater than 1.0 indicates that the mutant strain colonized better than wild type and a competi-tive index less than 1.0 indicates that the mutant strain was less efficient than the wild type in colonization. The results of two to four experiments using a total of 10–26 mice were combined for each sample. Mean values are shown and error bars represent standard error of the mean.

Chemosensory control of twitching motility in P. aeruginosa 889
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 873–893
assign 3D-structure to a protein of unknown structure basedon structural information from members of the same family(Blundell et al., 1987). The crystal structure of the C-terminalHPt domain of ArcB (1AOB.pdb; Kato et al., 1997) and theNMR structure of the phosphotransfer site of CheA (coordi-nates for CheA were kindly provided by F. W. Dahlquist,University of Oregon) were used as templates to assignstructures to the HPt1, SPt and HPt4 domains of ChpA.
Coordinates were assigned to the conserved secondarystructural regions of the ArcB and CheA according to thealignment shown in Fig. 3 using Molecular Simulations.InsightII package on a Silicon Graphics Octane work station.All suitable loop structures were obtained by searching theBrookhaven Protein Databank for proteins containing loopsof the same length joining similar secondary structural units.The resultant crude model was then refined by energy mini-mization using Biosym’s CVFF forcefield to a convergencecriterion of less than 5.0kcal/molA (maximum derivative) asreported elsewhere (Urquhart et al., 2000).
Based on studies of all coordinate entries in theBrookhaven Databank derived by X-ray, NMR and computa-tional methods, along with a number of deliberately misfoldedproteins, Eisenberg et al. (1989) suggested that a cut-off of<0.45 ¥ Scalc is useful for identifying grossly misfolded struc-tures. Models with scores between 0.45 ¥ Scalc and Scalcare considered correct, although it is still possible that locallymisfolded regions occur. The examination of the profile scorein a moving window scan of 10 residues can be used toidentify locally misfolded regions.
Construction of isogenic mutants
Allelic exchange mutants of the genes chpB, chpC, chpD andchpE were constructed using the sucrose selection systemdescribed previously (Schweizer, 1992; Alm and Mattick,1996), as follows. The tetracycline resistance cassette (TcR)from pSM-TET was inserted into the PstI site of chpB; theKpnI site of chpC; the XhoI site of chpD; or the internal ApaIsites of chpE. Suicide clones for allelic exchange were con-structed as described in Table 1. These clones each containthe genes sacB/sacR which promote sensitivity to sucroseand oriT which enables conjugal transfer. The constructswere then transformed into the E. coli donor strain S17-1 inpreparation for mating into PAK. Following conjugation,transconjugates were selected on 5% sucrose media con-taining tetracycline. This allows selection of colonies in whichthe plasmid sequences have been excised while leaving thehomologously recombined mutated gene in the chromosome.All mutants were genotypically confirmed by Southern blot-ting. An in frame deletion of chpA in PAO1 was constructedas follows. An approximatley 1 kb region 5¢ of chpA andincluding the first 32 bp of chpA was amplified with primerschpA6 (5¢-GAATTCAAGATGGCGAGCGAGATGCG-3¢) andchpA7 (5¢-AAGCTTCGTGCCGGTCACCCATAGCC-3¢). Anapproximately 1 kb region 3¢of chpA and encompassing thefinal 214 bp of chpA was amplified with primers chpA8(5¢-AAGCTTTCGAGATGCCGCGCATGGAC-3¢ and chpA9(5¢-TCTAGACAGTCGGGGCCGAACTGCTG). The PCR pro-ducts were cloned into pGEM-T and sequence confirmed.The cloned PCR products were excised and the fragmentsconcatamerized and cloned into pOK12. The chpA in frame
deletion construct was then shuttled into the suicide vectorpJEN34. pJEN34 is pRIC380 carrying the EZ::TN < TET-1 >transposon (EpiCentre) in the b-lactamase gene. The result-ant chpA deletion construct pJB5 was transformed into S17-1 and mated into PAO1. Following sucrose selection, tetra-cycline sensitive colonies were PCR amplified with chpA6and chpA7 to identify colonies in which the wild-type chpAsequences had been replaced with the in frame deletion ofchpA. These mutants were genotypically confirmed by South-ern blotting.
Motility assays
To assay subsurface twitching motility, the P. aeruginosa col-ony to be tested was stab inoculated through a 1% agar plate.Following incubation at 37∞C, a zone of motility could bevisualized between the agar and Petri dish interface. To aidvisualization, the agar was compressed and then stainedusing a 0.05% Coomassie brilliant blue R250 stain (40%methanol, 10% acetic acid) as described previously (Alm andMattick, 1995). Swimming motility was assayed by stab inoc-ulation of the colony to be tested into a 0.3% agar platefollowed by incubation overnight at 37∞C.
Microscopy
Light microscopy was performed as previously described(Semmler et al., 1999). Sterile microscope slides were sub-merged in molten GelGro media to obtain a thin layer ofmedia coating the slide. The slides were allowed to set in ahorizontal position and air dried briefly before use. The slideswere then inoculated with a small loopful of bacteria takenfrom an overnight plate culture. A sterile glass coverslip wasplaced over the point of inoculation and the slide transferredto a large Petri dish containing a moist tissue and sealed withNescofilm (BANDO Chemical Industries) to maintain humidconditions. Incubation times ranged from 2 to 6 h at 37∞C.
Slide cultures were examined using a Zeiss microscopeAxioskop 50 with Nomarski facilities at 200–400 ¥ magnifica-tion. Video microscopy was performed in a room heated to30∞C. Video images were recorded over a period of 2–4 h atspeeds of either 1 field/3.22 s; 1 field/0.66 s; or real time(1 field per 1/50 second) using a JVC TK-CI38IEG videocamera connected to a Sanyo TLS-S2500P time-lapse videorecorder.
PilA immunoblotting
Detection of cell-associated pilin was performed as follows.Bacteria were grown for 24 h on LB agar, scraped from theagar surface, and resuspended to OD600 = 1.0 in 50 mM Nacarbonate buffer (pH 9.6). One ml of the sample was centri-fuged and the cell pellet resuspended in 20 ml sample buffer(60 mM Tris-HCL pH 6.8, 2% SDS, 10% glycerol, 5% b-mer-captoethanol, 0.001% bromophenol blue). To reduce viscos-ity, the samples were sheared by 20 passages through a 271/2 gauge needle and heated to 100∞C for 5 min. Surface piliwere isolated by harvesting cells from 24 h agar plate cul-tures into 2 ml of phosphate-buffered saline, and vortexingfor 2 min. The suspension was centrifuged at low-speed

890 C. B. Whitchurch et al.
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 873–893
(2300 g for 5 min) to remove whole cells, after which thesupernatant was collected and subjected to a high-speedcentrifugation (15 000 g for 20 min) to remove cell debris.The resulting supernatant was incubated overnight at 4∞C inthe presence of 100 mM MgCl2 to precipitate pili, as describedpreviously (Alm and Mattick, 1995). The precipitate was col-lected by centrifugation (15 000 g for 20 min) and resus-pended in gel loading buffer. Samples were then displayedwith 15% SDS–PAGE gels (Laemmli, 1970), and electroblot-ted onto Immobilon-P transfer membrane (Millipore, Bedford,MA) in the Tris-glycine system described by Towbin et al.(1979). Membranes were blocked with 5% skimmed milk,probed with a 1:5000 dilution of primary anti-PilA antibody in1% skim milk powder in PBS. Membranes were then incu-bated with a 1:10 000 dilution of goat anti-rabbit immunoglo-bulin G conjugated with horse radish peroxidase (BoehringerMannheim) in 1% skim milk powder in PBS followed by detec-tion by ECL using the Supersignal chemiluminescence kit(Pierce).
ELISA
Enzyme-linked immunosorbent assays (ELISAs) to deter-mine the amount of surface pili were performed as follows.Bacteria were grown for 16–24 h on LB agar, scraped fromthe agar surface, and resuspended to OD600 = 1.0 in 50 mMNa carbonate buffer (pH 9.6). Two-hundred microlitres of thecell suspension were loaded to wells of a 96-well ELISA plateand incubated overnight at 4∞C. Wells were then washed inPBS containing 0.5% Tween-20, and blocked for 1 h at 37∞Cwith 3% BSA. Wells were washed again, and twofold serialdilutions of primary antibody (anti-PilA antibody at a startingdilution of 1:500) added for 1 h at 37∞C. Wells were thenwashed, incubated at 37∞C for 1 h with secondary antibody(1:5000 dilution of goat anti-rabbit immunoglobulan G conju-gated with alkaline phosphatase (Boehringer), washed andcolour developed using 4 mg ml-1 p-nitrophenyl phosphate(Sigma) in 1 M Tris-Cl buffer (pH 8.0). Alkaline phosphataseactivity was determined by absorbance at 405 nm using anELISA reader (Bio-Rad).
Assays of pilA expression
RNA preparation and Northern blots of pilA mRNA levels wasperformed as previously described (Whitchurch and Mattick,1994). PpilA::xylE transcriptional reporter assays were per-formed as follows. pMIC66 was transformed in to the strainsto be assayed. Dilutions (1:100) of overnight cultures wereincubated with shaking at 37∞C for 16 h. One hundred micro-litres of each culture was transferred to wells of a 96-welldish and culture OD measurements at 595 nm recorded in aBio-Rad ELISA plate reader. Acetone (20 ml) and 80 ml assaybuffer (50 mM K2HPO4, pH 7.5) was added to each sample.Twenty microlitres of the cells/acetone/assay buffer solutiondiluted in 160 ml assay buffer was assayed after addition of20 ml of the catechol 2,3-dioxygenase substrate, catechol(1.1 mg ml-1). The enzyme catalyses the conversion of cate-chol to b-muconic e-semialdehyde, a coloured product whichabsorbs light at 380 nm. Thus the rate of conversion of sub-strate to product can be followed by the absorbance at
380 nm. The rate of reaction over 2 min was recorded bytaking A380 readings at 15 second time intervals in a Bench-Mark Microplate Reader (Bio-Rad). Initial rates of reactionwere determined with Microplate Manager 3.11 v1.57software.
Cytotoxicity assays
Cytotoxicity assays were carried out in 3-day-old MDCKconfluent monolayers grown on Transwells as previouslydescribed (Comolli et al., 1999) with the following modifica-tions. Bacteria were used at an MOI of approximately 50.Lactate dehydrogenase assays were performed accordingto the manufacturer’s specifications (Sigma) following co-cultivation of the bacteria with the filter-grown MDCK cells forthe indicated times.
Mouse model for acute pneumonia
Mouse mortality and competitive index assays were carriedout as previously described (Comolli et al., 1999). For themortality studies, groups of five lightly anesthetized 6–8-week-old Balb/C female mice were inoculated directly in thenares with 50 ml of each strain (A600 of 0.5; approximately 108
bacteria). Mice were monitored over the subsequent sevendays in compliance with the guidelines of Animal Care Com-mittee of the University of California, San Francisco. Theinoculum dose for the wild type and mutant strains for thecompetitive index experiments was adjusted to an A600 of 0.75(approximately 108 bacteria) and the mice were sacrificedafter 24 h.
Acknowledgements
We would like to thank Professor F. W. Dahlquist for kindlyproviding the NMR structure of CheA which was used forstructural modelling and J. Dyason for his assistance with the3D-structural modelling. This work was supported by theNational Health and Medical Research Council of Australiaand the Australian Research Council. J.N.E. was supportedby grants from the NIH (AI42806), the Cystic Fibrosis Foun-dation, and the American Lung Association. During a portionof this work, J.N.E. was a Career Investigator of the AmericanLung Association. J.C.C. was supported by the Bank ofAmerica-Gianinni Foundation.
References
Aleshin, V.V., Zakataeva, N.P., and Livshits, V.A. (1999) Anew family of amino-acid-efflux proteins. Trends BiochemSci 24: 133–135.
Alm, R.A., and Mattick, J.S. (1995) Identification of a gene,pilV, required for type 4 fimbrial biogenesis in Pseudomo-nas aeruginosa whose product possesses a prepilin-likeleader sequence. Mol Microbiol 16: 485–496.
Alm, R.A., and Mattick, J.S. (1996) Identification of two geneswith prepilin-like leader sequences required for type 4 fim-brial biogenesis in Pseudomonas aeruginosa. J Bacteriol178: 3809–3817.

Chemosensory control of twitching motility in P. aeruginosa 891
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 873–893
Alm, R.A., and Mattick, J.S. (1997) Genes involved in thebiogenesis and function of type-4 fimbriae in Pseudomo-nas aeruginosa. Gene 192: 89–98.
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J.,Zhang, Z., Miller, W., and Lipman, D.J. (1997) GappedBLAST and PSI-BLAST: a new generation of protein databasesearch programs. Nucleic Acids Res 25: 3389–3402.
Armitage, J.P. (1999) Bacterial tactic responses. Adv MicrobPhysiol 41: 229–289.
Bateman, A., Birney, E., Cerruti, L., Durbin, R., Etwiller, L.,Eddy, S.R., et al. (2002) The Pfam protein families data-base. Nucleic Acids Res 30: 276–280.
Beatson, S.A., Whitchurch, C.B., Sargent, J.L., Levesque,R.C., and Mattick, J.S. (2002) Differential regulation oftwitching motility and elastase production by Vfr inPseudomonas aeruginosa. J Bacteriol 184: 3605–3613.
Bhaya, D., Bianco, N.R., Bryant, D., and Grossman, A.(2000) Type IV pilus biogenesis and motility in the cyano-bacterium Synechocystis sp. PCC6803. Mol Microbiol 37:941–951.
Bhaya, D., Takahashi, A., and Grossman, A.R. (2001) Lightregulation of type IV pilus-dependent motility bychemosensor-like elements in Synechocystis PCC6803.Proc Natl Acad Sci USA 98: 7540–7545.
Blundell, T.L., Sibanda, B.L., Sternberg, M.J., and Thornton,J.M. (1987) Knowledge-based prediction of protein struc-tures and the design of novel molecules. Nature 326: 347–352.
Bourret, R.B., and Stock, A.M. (2002) Molecular informationprocessing: lessons from bacterial chemotaxis. J BiolChem 277: 9625–9628.
Bradley, D.E. (1973) A pilus dependent Pseudomonas aerug-inosa bacteriophage with a long non-retractile tail. Virology51: 489–492.
Burkart, M., Toguchi, A., and Harshey, R.M. (1998) Thechemotaxis system, but not chemotaxis, is essential forswarming motility in Escherichia coli. Proc Natl Acad SciUSA 95: 2568–2573.
Chiang, P., and Burrows, L.L. (2003) Biofilm formation byhyperpiliated mutants of Pseudomonas aeruginosa. J Bac-teriol 185: 2374–2378.
Comolli, J.C., Hauser, A.R., Waite, L., Whitchurch, C.B.,Mattick, J.S., and Engel, J.N. (1999) Pseudomonasaeruginosa gene products PilT and PilU are required forcytotoxicity in vitro and virulence in a mouse model of acutepneumonia. Infect Immun 67: 3625–3630.
Cox, S., Radzio-Andzelm, E., and Taylor, S.S. (1994) Domainmovements in protein kinases. Curr Opin Struct Biol 4:893–901.
Croft, L., Beatson, S.A., Whitchurch, C.B., Huang, B., Blake-ley, R.L., and Mattick, J.S. (2000) An interactive web-basedPseudomonas aeruginosa genome database: discovery ofnew genes, pathways and structures. Microbiol 146: 2351–2364.
D’Argenio, D.A., Gallagher, L.A., Berg, C.A., and Manoil, C.(2001) Drosophila as a model host for Pseudomonasaeruginosa infection. J Bacteriol 183: 1466–1471.
D’Argenio, D.A., Calfee, M.W., Rainey, P.B., and Pesci, E.C.(2002) Autolysis and autoaggregation in Pseudomonasaeruginosa colony morphology mutants. J Bacteriol 184:6481–6489.
Darzins, A. (1993) The pilG gene product, required forPseudomonas aeruginosa pilus production and twitchingmotility, is homologous to the enteric, single-domainresponse regulator CheY. J Bacteriol 175: 5934–5944.
Darzins, A. (1994) Characterization of a Pseudomonasaeruginosa gene cluster involved in pilus biosythesis andtwitching motility: sequence similarity to the chemotaxisproteins of enterics and the gliding bacterium Myxococcusxanthus. Mol Microbiol 11: 137–153.
Darzins, A. (1995) The Pseudomonas aeruginosa pilK geneencodes a chemotactic methyltransferase (CheR) homo-logue that is translationally regulated. Mol Microbiol 15:703–717.
Djordjevic, S., and Stock, A.M. (1998) Structural analysis ofbacterial chemotaxis proteins: components of a dynamicsignaling system. J Struct Biol 124: 189–200.
Eisenberg, D., Wesson, M., and Yamashita, M. (1989) Inter-pretation of protein folding and binding with atomic solva-tion parameters. Chem Scrip 29A: 217–221.
Falke, J.J., Bass, R.B., Butler, S.L., Chervitz, S.A., andDanielson, M.A. (1997) The two-component signaling path-way of bacterial chemotaxis: a molecular view of signaltransduction by receptors, kinases, and adaptationenzymes. Annu Rev Cell Dev Biol 13: 457–512.
Ferrandez, A., Hawkins, A.C., Summerfield, D.T., and Har-wood, C.S. (2002) Cluster II che genes from Pseudomonasaeruginosa are required for an optimal chemotacticresponse. J Bacteriol 184: 4374–4383.
Gallegos, M.T., Schleif, R., Bairoch, A., Hofmann, K., andRamos, J.L. (1997) Arac/XylS family of transcriptional reg-ulators. Microbiol Mol Biol Rev 61: 393–410.
Giamarellou, H. (2000) Therapeutic guidelines forPseudomonas aeruginosa infections. Int J AntimicrobAgents 16: 103–106.
Hobbs, M., and Mattick, J.S. (1993) Common components inthe assembly of type 4 fimbriae, DNA transfer systems,filamentous phage and protein-secretion apparatus: a gen-eral system for the formation of surface-associated proteincomplexes. Mol Microbiol 10: 233–243.
Hobbs, M., Collie, E.S., Free, P.D., Livingston, S.P., andMattick, J.S. (1993) PilS and PilR, a two-component tran-scriptional regulatory system controlling expression of type4 fimbriae in Pseudomonas aeruginosa. Mol Microbiol 7:669–682.
Huang, B., Whitchurch, C.B., Croft, L., Beatson, S.A., andMattick, J.S. (2000) A minimal tiling path cosmid library forfunctional analysis of the Pseudomonas aeruginosa PAO1genome. Microb Comp Genomics 5: 189–203.
Jander, G., Rahme, L.G., and Ausubel, F.M. (2000) Positivecorrelation between virulence of Pseudomonas aerugi-nosa mutants in mice and insects. J Bacteriol 182: 3843–3845.
Kang, P.J., Hauser, A.R., Apodaca, G., Fleiszig, S.M.,Wiener-Kronish, J., Mostov, K., and Engel, J.N. (1997)Identification of Pseudomonas aeruginosa genes requiredfor epithelial cell injury. Mol Microbiol 24: 1249–1262.
Kato, M., Mizuno, T., Shimizu, T., and Hakoshima, T. (1997)Insights into multistep phosphorelay from the crystal struc-ture of the C-terminal HPt domain of ArcB. Cell 88: 717–723.
Kennelly, P.J. (2002) Protein kinases and protein phos-

892 C. B. Whitchurch et al.
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 873–893
phatases in prokaryotes: a genomic perspective. FEMSMicrobiol Lett 206: 1–8.
Klausen, M., Aaes-Jorgensen, A., Molin, S., and Tolker-Nielsen, T. (2003a) Involvement of bacterial migration inthe development of complex multicellular structures in Pseu-domonas aeruginosa biofilms. Mol Microbiol 50: 61–68.
Klausen, M., Heydorn, A., Ragas, P., Lambertsen, L., Aaes-Jorgensen, A., Molin, S., and Tolker-Nielsen, T. (2003b)Biofilm formation by Pseudomonas aeruginosa wild type,flagella and type IV pili mutants. Mol Microbiol 48: 1511–1524.
Laemmli, U.K. (1970) Cleavage of structural proteins duringthe assembly of the head of bacteriophage T4. Nature 227:680–685.
Lee, S.H., Butler, S.M., and Camilli, A. (2001) Selection forin vivo regulators of bacterial virulence. Proc Natl Acad SciUSA 98: 6889–6894.
Letunic, I., Goodstadt, L., Dickens, N.J., Doerks, T., Schultz,J., Mott, R., et al. (2002) Recent improvements to theSMART domain-based sequence annotation resource.Nucleic Acids Res 30: 242–244.
Masduki, A., Nakamura, J., Ohga, T., Umezaki, R., Kato, J.,and Ohtake, H. (1995) Isolation and characterization ofchemotaxis mutants and genes of Pseudomonas aerugi-nosa. J Bacteriol 177: 948–952.
Mattick, J.S. (2002) Type IV pili and twitching motility. AnnuRev Microbiol 56: 289–314.
Mattick, J.S., and Alm, R.A. (1995) Common architecture oftype 4 fimbriae and complexes involved in macromoleculartraffic. Trends Microbiol 3: 411–413.
Mattick, J.S., Bills, M.M., Anderson, B.J., Dalrymple, B., Mott,M.R., and Egerton, J.R. (1987) Morphogenetic expressionof Bacteroides nodosus fimbriae in Pseudomonas aerugi-nosa. J Bacteriol 169: 33–41.
Merz, A.J., So, M., and Sheetz, M.P. (2000) Pilus retractionpowers bacterial twitching motility. Nature 407: 98–102.
Mongkolsuk, S., Vattanaviboon, P., Rabibhadana, S., andKiatpapan, P. (1993) Versatile gene cassette plasmids tofacilitate the construction of generalized and specializedcloning vectors. Gene 124: 131–132.
Morales, V.M., Backman, A., and Bagdasarian, M. (1991) Aseries of wide-host-range low-copy-number vectors thatallow direct screening for recombinants. Gene 97: 39–47.
Mourey, L., Da Re, S., Pedelacq, J.D., Tolstykh, T., Faurie,C., Guillet, V., et al. (2001) Crystal structure of the CheAhistidine phosphotransfer domain that mediates responseregulator phosphorylation in bacterial chemotaxis. J BiolChem 276: 31074–31082.
O’Toole, G.A., and Kolter, R. (1998) Flagellar and twitchingmotility are necessary for Pseudomonas aeruginosa bio-film development. Mol Microbiol 30: 295–304.
O’Toole, G.A., Gibbs, K.A., Hager, P.W., Phibbs, P.V. Jr andKolter, R. (2000) The global carbon metabolism regulatorCrc is a component of a signal transduction pathwayrequired for biofilm development by Pseudomonas aerug-inosa. J Bacteriol 182: 425–431.
Pullen, K., Rajagopal, P., Branchini, B.R., Huffine, M.E.,Reizer, J., Saier, M.H. Jr et al. (1995) Phosphorylation ofserine-46 in HPr, a key regulatory protein in bacteria,results in stabilization of its solution structure. Protein Sci4: 2478–2486.
Rahme, L.G., Ausubel, F.M., Cao, H., Drenkard, E., Goum-nerov, B.C., Lau, G.W., et al. (2000) Plants and animalsshare functionally common bacterial virulence factors.Proc Natl Acad Sci USA 97: 8815–8821.
Ratnaningsih, E., Dharmsthiti, S., Krishnapillai, V., Morgan,A., Sinclair, M., and Holloway, B.W. (1990) A combinedphysical and genetic map of Pseudomonas aeruginosaPAO. J Gen Microbiol 136: 2351–2357.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989) Molecu-lar Cloning: a Laboratory Manual. Cold Spring Harbour,New York: Cold Spring Harbor Laboratory Press.
Schultz, J., Milpetz, F., Bork, P., and Ponting, C.P. (1998)SMART, a simple modular architecture research tool: iden-tification of signaling domains. Proc Natl Acad Sci USA 95:5857–5864.
Schweizer, H.P. (1991) Escherichia-Pseudomonas shuttlevectors derived from pUC18/19. Gene 97: 109–112.
Schweizer, H.P. (1992) Allelic exchange in Pseudomonasaeruginosa using novel ColE1-type vectors and a family ofcassettes containing a portable oriT and the counter-selectable Bacillus subtilis sacB marker. Mol Microbiol 6:1195–1204.
Schweizer, H.P. (1993) Two plasmids, X1918 and Z1918, foreasy recovery of the xylE and lacZ reporter genes. Gene134: 89–91.
Semmler, A.B.T., Whitchuch, C.B., and Mattick, J.S. (1999)A re-examination of twitching motility in Pseudomonasaeruginosa. Microbiol 145: 2863–2873.
Skerker, J.M., and Berg, H.C. (2001) Direct observation ofextension and retraction of type IV pili. Proc Natl Acad SciUSA 98: 6901–6904.
Song, H.K., Lee, J.Y., Lee, M.G., Moon, J., Min, K., Yang,J.K., and Suh, S.W. (1999) Insights into eukaryotic multi-step phosphorelay signal transduction revealed by thecrystal structure of Ypd1p from Saccharomyces cerevisiae.J Mol Biol 293: 753–761.
Stover, C.K., Pham, X.Q., Erwin, A.L., Mizoguchi, S.D., War-rener, P., Hickey, M.J., et al. (2000) Complete genomesequence of Pseudomonas aeruginosa PA01, an opportu-nistic pathogen. Nature 406: 959–964.
Strom, M.S., and Lory, S. (1993) Structure-function and bio-genesis of the type IV pili. Annu Rev Microbiol 47: 565–596.
Sun, H., Zusman, D.R., and Shi, W. (2000) Type IV pilusof Myxococcus xanthus is a motility apparatus controlledby the frz chemosensory system. Curr Biol 10: 1143–1146.
Tan, M.W., and Ausubel, F.M. (2000) Caenorhabditis ele-gans: a model genetic host to study Pseudomonas aerug-inosa pathogenesis. Curr Opin Microbiol 3: 29–34.
Towbin, H., Staehelin, T., and Gordon, J. (1979) Electro-phoretic transfer of proteins from polyacrylamide gels tonitrocellulose sheets: procedure and some applications.Proc Natl Acad Sci USA 76: 4350–4354.
Urquhart, A.J., Kennedy, D., Gould, S.J., and Crane, D.I.(2000) Interaction of Pex5p, the type 1 peroxisome target-ing signal receptor, with the peroxisomal membrane pro-teins Pex14p and Pex13p. J Biol Chem 275: 4127–4136.
Vieira, J., and Messing, J. (1991) New pUC-derived cloningvectors with different selectable markers and DNA replica-tion origins. Gene 100: 189–194.

Chemosensory control of twitching motility in P. aeruginosa 893
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 873–893
Vrljic, M., Garg, J., Bellmann, A., Wachi, S., Freudl, R.,Malecki, M.J., et al. (1999) The LysE superfamily: topologyof the lysine exporter LysE of Corynebacterium glutamicum,a paradyme for a novel superfamily of transmembranesolute translocators. J Mol Microbiol Biotechnol 1: 327–336.
Ward, M.J., and Zusman, D.R. (1999) Motility in Myxococcusxanthus and its role in developmental aggregation. CurrOpin Microbiol 2: 624–629.
Watson, A.A., Alm, R.A., and Mattick, J.S. (1996a) Construc-tion of improved vectors for protein production inPseudomonas aeruginosa. Gene 172: 163–164.
Watson, A.A., Mattick, J.S., and Alm, R.A. (1996b) Functionalexpression of heterologous type 4 fimbriae in Pseudomo-nas aeruginosa. Gene 175: 143–150.
Whitchurch, C.B., and Mattick, J.S. (1994) Characterisationof a gene, pilU, required for twitching motility but not phagesensitivity in Pseudomonas aeruginosa. Mol Microbiol 13:1079–1091.
Whitchurch, C.B., Hobbs, M., Livingston, S.P., Krishnapillai,V., and Mattick, J.S. (1991) Characterisation of aPseudomonas aeruginosa twitching motility gene and evi-
dence for a specialised protein export system widespreadin eubacteria. Gene 101: 33–44.
Whitchurch, C.B., Alm, R.A., and Mattick, J.S. (1996) Thealginate regulator AlgR and an associated sensor FimS arerequired for twitching motility in Pseudomonas aeruginosa.Proc Natl Acad Sci USA 93: 9839–9843.
Winther-Larsen, H.C., and Koomey, M. (2002) Transcrip-tional, chemosensory and cell-contact-dependent regula-tion of type IV pilus expression. Curr Opin Microbiol 5:173–178.
Xu, Q., and West, A.H. (1999) Conservation of structure andfunction among histidine-containing phosphotransfer (HPt)domains as revealed by the crystal structure of YPD1. JMol Biol 292: 1039–1050.
Zakataeva, N.P., Aleshin, V.V., Tokmakova, I.L., Troshin,P.V., and Livshits, V.A. (1999) The novel transmembraneEscherichia coli proteins involved in the amino acid efflux.FEBS Lett 452: 228–232.
Zhou, H., and Dahlquist, F.W. (1997) Phosphotransfer site ofthe chemotaxis-specific protein kinase CheA as revealedby NMR. Biochemistry 36: 699–710.
Related Documents





![Fimbrias - [DePa] Departamento de Programas …depa.fquim.unam.mx/amyd/archivero/U2g_FimbriasPiliF_19172.pdf · Twitching motility Modelo de motilidad para Neisseria gonorrhoeae.Esta](https://static.cupdf.com/doc/110x72/5bb197c209d3f246678df3b4/fimbrias-depa-departamento-de-programas-depafquimunammxamydarchiverou2gfimbriaspilif19172pdf.jpg)





