Abstract A 28.7-kb DNA region containing the gdhA gene of Aspergillus awamori was cloned from a genomic DNA library. A fragment of 2570 nucleotides was se- quenced that contained ORF1, of 1380 bp, encoding a pro- tein of 460 amino acids (Mr 49.4 kDa). The encoded pro- tein showed high similarity to the NADP-dependent glu- tamate dehydrogenases of different organisms. The cloned gene was functional since it complemented two different Aspergillus nidulans gdhA mutants, restoring high levels of NADP-dependent glutamate dehydrogenase to the trans- formants. The A. awamori gdhA gene was located by pulsed-field gel electrophoresis in a 5.5-Mb band (corre- sponding to a doublet of chromosomes II and III), and was transcribed as a monocistronic transcript of 1.7 kb. Tran- script levels of the gdhA gene were very high during the rapid growth phase and decreased drastically after 48 h of cultivation. Very high expression levels of the gdhA gene were observed in media with ammonium or asparagine as the nitrogen source, whereas glutamic acid repressed tran- scription of the gdhA gene. These results indicate that ex- pression of the gdhA gene is subject to a strong nitrogen regulation at the transcriptional level. Key words Glutamate dehydrogenase · Nitrogen regulation · Aspergillus awamori · Chromosome resolution · Promoter Introduction Aspergillus awamori is widely used for the secretion of ex- tracellular proteins, including homologous proteins [e.g. glucoamylase (Finkelstein et al. 1989), α-amylase (Kor- man et al. 1990), 1.4-beta-endoxylanase (Hessing et al. 1994)] and a variety of heterologous polypeptides (Ver- does et al. 1995; Gouka et al. 1996). The basic molecular genetics of A. awamori is largely unknown. This species is amenable to efficient transformation (Gouka et al. 1995) but only a few A. awamori genes have as yet been cloned and characterized (Berka et al. 1990; Thompson 1990; Adams and Royer 1990; Piddington et al. 1993; Hijarru- bia et al. 1997). Nitrogen regulation of metabolism in filamentous fungi (Marzluf 1997) is particularly interesting in the context of gene expression for protein secretion. The proteins to be secreted are targeted to the endoplasmic reticulum and pro- ceed via the Golgi system to the cytoplasmic membrane, where they are released. During protein secretion the poly- peptides to be secreted are subject to “quality controls” by several mechanisms (Hammond and Helenius 1995) and those that do not fold adequately for transport through the different secretory compartments are degraded by pro- teases. Proteolytic processing plays an important role in protein secretion. In addition to their involvement in pro- tein degradation, specific peptidases (e.g. the KEX pro- tease cleaving specifically at Arg-Lys sites in polypeptides; Calmels et al. 1991) are used to release polypeptides of interest attached to carrier extracellular proteins (Gouka et al. 1997). These proteolytic systems are subject to nitro- gen regulation (Cohen 1973). NADP-dependent glutamate dehydrogenase (GDH) (encoded by gdhA) has been shown to play an important role in nitrogen metabolism and regulation in Aspergillus nidulans (Kinghorn and Pateman 1973a). The gdhA mu- tants lack repression by ammonium of a number of en- zymes and uptake systems, including extracellular pro- teases, nitrate reductase, xanthine dehydrogenase and glu- tamate uptake (Kinghorn and Pateman 1973b). Curr Genet (1998) 34: 50–59 © Springer-Verlag 1998 Received: 13 January / 12 May 1998 Rosa-Elena Cardoza · Francisco-José Moralejo Santiago Gutiérrez · Javier Casqueiro Francisco Fierro · Juan F. Martín Characterization and nitrogen-source regulation at the transcriptional level of the gdhA gene of Aspergillus awamori encoding an NADP-dependent glutamate dehydrogenase R.-E. Cardoza · F.-J. Moralejo · S. Gutiérrez · J. Casqueiro F. Fierro · J. F. Martín Instituto de Biotecnología INBIOTEC, Parque Científico de León, Avda. del Real, 1, E-24006 León, Spain J. F. Martín (‰) Universidad de León, Facultad de Biología, Area de Microbiología, E-24071 León, Spain Tel.: +34-87-291505 Fax: +34-87-291506 e-mail: [email protected] Communicated by K. Esser ORIGINAL PAPER

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Abstract A 28.7-kb DNA region containing the gdhAgene of Aspergillus awamori was cloned from a genomicDNA library. A fragment of 2570 nucleotides was se-quenced that contained ORF1, of 1380 bp, encoding a pro-tein of 460 amino acids (Mr 49.4 kDa). The encoded pro-tein showed high similarity to the NADP-dependent glu-tamate dehydrogenases of different organisms. The clonedgene was functional since it complemented two differentAspergillus nidulans gdhA mutants, restoring high levelsof NADP-dependent glutamate dehydrogenase to the trans-formants. The A. awamori gdhA gene was located bypulsed-field gel electrophoresis in a 5.5-Mb band (corre-sponding to a doublet of chromosomes II and III), and wastranscribed as a monocistronic transcript of 1.7 kb. Tran-script levels of the gdhA gene were very high during therapid growth phase and decreased drastically after 48 h ofcultivation. Very high expression levels of the gdhA genewere observed in media with ammonium or asparagine asthe nitrogen source, whereas glutamic acid repressed tran-scription of the gdhA gene. These results indicate that ex-pression of the gdhA gene is subject to a strong nitrogenregulation at the transcriptional level.
Key words Glutamate dehydrogenase · Nitrogen regulation · Aspergillus awamori · Chromosome resolution · Promoter
Introduction
Aspergillus awamori is widely used for the secretion of ex-tracellular proteins, including homologous proteins [e.g.glucoamylase (Finkelstein et al. 1989), α-amylase (Kor-man et al. 1990), 1.4-beta-endoxylanase (Hessing et al.1994)] and a variety of heterologous polypeptides (Ver-does et al. 1995; Gouka et al. 1996). The basic moleculargenetics of A. awamori is largely unknown. This speciesis amenable to efficient transformation (Gouka et al. 1995)but only a few A. awamori genes have as yet been clonedand characterized (Berka et al. 1990; Thompson 1990; Adams and Royer 1990; Piddington et al. 1993; Hijarru-bia et al. 1997).
Nitrogen regulation of metabolism in filamentous fungi(Marzluf 1997) is particularly interesting in the context ofgene expression for protein secretion. The proteins to besecreted are targeted to the endoplasmic reticulum and pro-ceed via the Golgi system to the cytoplasmic membrane,where they are released. During protein secretion the poly-peptides to be secreted are subject to “quality controls” byseveral mechanisms (Hammond and Helenius 1995) andthose that do not fold adequately for transport through thedifferent secretory compartments are degraded by pro-teases. Proteolytic processing plays an important role inprotein secretion. In addition to their involvement in pro-tein degradation, specific peptidases (e.g. the KEX pro-tease cleaving specifically at Arg-Lys sites in polypeptides;Calmels et al. 1991) are used to release polypeptides ofinterest attached to carrier extracellular proteins (Gouka etal. 1997). These proteolytic systems are subject to nitro-gen regulation (Cohen 1973).
NADP-dependent glutamate dehydrogenase (GDH)(encoded by gdhA) has been shown to play an importantrole in nitrogen metabolism and regulation in Aspergillusnidulans (Kinghorn and Pateman 1973a). The gdhA mu-tants lack repression by ammonium of a number of en-zymes and uptake systems, including extracellular pro-teases, nitrate reductase, xanthine dehydrogenase and glu-tamate uptake (Kinghorn and Pateman 1973b).
Curr Genet (1998) 34: 50–59 © Springer-Verlag 1998
Received: 13 January / 12 May 1998
Rosa-Elena Cardoza · Francisco-José MoralejoSantiago Gutiérrez · Javier CasqueiroFrancisco Fierro · Juan F. Martín
Characterization and nitrogen-source regulation at the transcriptional levelof the gdhA gene of Aspergillus awamori encoding an NADP-dependent glutamate dehydrogenase
R.-E. Cardoza · F.-J. Moralejo · S. Gutiérrez · J. CasqueiroF. Fierro · J. F. MartínInstituto de Biotecnología INBIOTEC, Parque Científico de León, Avda. del Real, 1, E-24006 León, Spain
J. F. Martín (½)Universidad de León, Facultad de Biología,Area de Microbiología, E-24071 León, SpainTel.: +34-87-291505Fax: +34-87-291506e-mail: [email protected]
Communicated by K. Esser
ORIGINAL PAPER

It was, therefore, of great interest to clone and charac-terize the gdhA gene of A. awamori as a tool to study ni-trogen metabolism and its regulation in this fungus.
In this article we report the characterization of the A. awamori gdhA gene encoding NADP-dependent gluta-mate dehydrogenase, the complementation of gdhA mu-tants, and transcriptional studies with different nitrogensources.
Materials and methods
Microorganisms and cloning vectors. A. awamori ATCC 22342 wasused as the source of DNA and RNA. A. nidulans mutants A686 (gdhA1, yA2, methH2, galA1) and A699 (gdhA1, biA1) (Kinghornand Pateman 1973a) were obtained from the Fungal Genetics StockCenter, and used for complementation studies with the gdhA genefrom A. awamori. The partial glutamate auxotrophy of these twostrains was confirmed by growth on media with glutamic acid, orhigh ammonium sulphate concentrations (100 mM), as a nitrogensource. Both gdhA mutants grow very poorly on high ammoniumsulphate concentration but showed normal growth on glutamic acid.A. nidulans ATCC 28901 (Ditchburn et al. 1974) was used as a con-trol in the complementation studies of the gdhA mutants. Escheri-chia coli DH5α served as the recipient strain for high-frequency plas-mid transformation. E. coli WK6 was used as a host for obtainingsingle-stranded DNA from pBluescript plasmids, and E. coli NM539served as a host for Lambda GEM12 (Promega Co., Wis.) phage de-rivatives.
Media and culture conditions. Filamentous fungi were routinelymaintained on solid-Power sporulation medium (Fierro et al. 1996)at 30°C for 3 days. A. awamori and A. nidulans seed cultures in CMmedium (containing 20 g/l malt extract; 5 g/l yeast extract; 5 g/l glu-cose) were inoculated with 106 spores/ml and grown at 28°C in a ro-tary G10 incubator (New Brunswick Scientific, New Brunswick,N. J.) for 48 h. For gdhA transcript isolation and characterizationstudies, A. awamori cultures in MDFA medium (Shen et al. 1984)were incubated with a 15% seed culture and grown at 30°C for48–72 h in a rotary shaker as described above.
A. awamori genomic library. A genomic library of total DNA of A.awamori ATCC 22342 was constructed in a Lambda GEM12 phagevector. Total DNA was extracted as described by Specht et al. (1982)and partially digested with Sau3AI to obtain DNA fragments ofbetween 17 and 23 kb. This DNA was purified by sucrose-gradientcentrifugation, ligated to Lambda GEM12 phage arms, and packagedin vitro using a Gigapack III Gold packaging system (Stratagene),resulting in a total of 8×104 recombinant phages.
Transformation of A. nidulans gdhA mutants. A. nidulans A686 andA699 were transformed by the method of Yelton et al. (1984) withthe 7.1-kb plasmid pGDHaw that contains the A. awamori gdhA genein a 2570-bp XbaI fragment. This fragment also contains an upstreampromoter region of 740-bp and a 322-bp region downstream fromORF1 (gdhA gene). The 2570-bp XbaI fragment was inserted intothe XbaI site of the fungal vector pIBRC43 that contains the phle-omycin resistance marker (under the control of the A. awamori gdhpromoter), giving rise to plasmid pGDHaw.
Southern blotting and hybridization. DNA from positive phages andtotal DNA from A. awamori was digested with restriction endonu-cleases, electrophoresed in 0.7% agarose and blotted by standardtechniques (Sambrook et al. 1989). Probes were labelled by nicktranslation with [32P]dCTP and hybridized by standard methods(Sambrook et al. 1989).
Isolation of RNA, Northern hybridization and slot blotting. TotalRNA of A. awamori was obtained by the phenol-SDS method (Au-
subel et al. 1987) from mycelia grown for 12, 24, 48, 60 or 72 h inMDFA medium (Shen et al. 1984) with 55.5 mM of glucose and10 mM of ammonium sulphate as carbon and nitrogen sources re-spectively. For nitrogen-regulation studies, the MDFA base medi-um (without ammonium sulphate) was supplemented with differ-ent nitrogen sources including glutamic acid, L-glutamine, sodiumnitrite, sodium nitrate and L-asparagine at a 10 mM final concen-tration. For Northern analysis, total RNA (5 µg) was run on a 1.2%agarose-formaldehyde gel. The gel was blotted onto a nylon filter(Nytran 0.45; Schleicher and Schuell) by standard methods. TheRNA was fixed by UV irradiation using a UV-Stratalinker 2400lamp (Stratagene, La Jolla, Calif.). For slot blotting, the RNA (5 µg)was loaded on a filter (Nytran 0.45), by vacuum, in a Bio-Dot SFMicrofiltration apparatus (Slot Blotting, Bio-Rad). The RNA wasfixed by UV irradiation as above. The filters were pre-hybridizedfor 3 h at 42°C in 50% formamide, 5× Denhardt’s, 5×SSPE, 0.1%SDS, 500 µg of denatured salmon-sperm DNA per ml, and hybri-dized in the same buffer containing 100 µg of denatured salmon-sperm DNA per ml at 42°C for 18 h, using as a probe an internalfragment (0.694-kb PvuII) of the A. awamori gdhA gene. The fil-ter was washed once in 2×SSC, 0.1% SDS at 42°C for 15 min, oncein 0.1×SSC, 0.1% SDS at 42°C for 15 min, and once more in 0.1×SSC, 0.1% SDS at 55°C for 20 min and then autoradiographed withAmersham X-ray film. The mRNA was purified from total RNAby using the Poly(A) Quick mRNA-isolation kit (Stratagene, LaJolla, Calif.).
cDNA synthesis and PCR amplification. cDNA was obtained fromtotal RNA extracted as described above from mycelia grown for 48 hin MDFA medium. The first and second cDNA strands were synthe-tized using a cDNA synthesis kit from Stratagene (La Jolla, Calif.).This cDNA was used for PCR-amplification of the fragments con-taining the exon-exon junctions by the following program: 1 cycleat 94°C for 5 min, 50°C for 1 min, 72°C for 1 min followed by 30cycles at 94°C for 1 min, 50°C for 1 min, 72°C for 1 min and final-ly one cycle at 72°C for 8 min.
Primer extension. Identification of the transcription start point wasperformed with 2 µg of mRNA obtained from mycelia grown inMDFA for 48 h using the procedure described by Ausubel et al.(1987). Extension was primed with the oligonucleotide “Pe” 5′-GGGGTTCTTCTGGAAGAGGGT-3′ (see Results).
DNA sequencing. DNA fragments containing sequences that hy-bridized with gdhA were subcloned into pBluescript SK+ in bothorientations, and sequenced by generating ordered sets of dele-tions with the Erase-a-base system (Promega Co., Wis.) by diges-tion with exonuclease III from appropriate ends, followed by re-moval of single-stranded DNA with S1 exonuclease. Sequencingof fragments of the gdhA gene was performed by the dideoxynu-cleotide chain-termination method (Sanger et al. 1977). For se-quencing the cDNA clones containing the exon-exon junctions, re-actions were performed with 90 ng of dsDNA using the GeneAmpPCR 2400 system coupled to the ABI-PRISM 310 automatic se-quencer (Perkin Elmer). Computer analysis of nucleotide and ami-no-acid sequences was made with the DNASTAR Programs (DNA-STAR, Inc., UK).
Enzyme assays. Nicotinamide adenine dinucleotide phosphate (NADP)-specific glutamate dehydrogenase (NADP-GDH) was as-sayed by following the reductive amination of α-ketoglutarate in thepresence of ammonia and NADPH (Kinghorn and Pateman 1973a)and expressed as units of enzyme activity per mg of protein. The in-itial reaction velocity was estimated from the change in optical den-sity at 340 nm in a Hitachi U-2001 spectrophotometer. One unit ofglutamate dehydrogenase was defined as the activity that catalyzesthe oxidation of 1 nmol of NADPH per min.
Accession number. The nucleotide sequence of the A. awamori gdhAgene has been deposited in the EMBL GeneBank under the acces-sion number Y15784.
51
Oligonucleotides. The following oligonucleotides were used as prim-ers for introns I and II:
IA 5′ ATG TCT AAC CTT CCT CAC 3′IB 5′ ACC CTT ACC ACC ACC CAT 3′IIA 5′ CGC TTC TGT GTT TCC TTC 3′IIB 5′ GTA CTT GAA CTT GTT GGC 3′
Chromosome resolution by pulsed-field gel electrophoresis. Proto-plasts of A. awamori were prepared by the method of Yelton et al.(1984). High-molecular-weight DNA was obtained by lysing the pro-toplasts in agarose blocks as described previously (Fierro et al. 1993).Resolution of the chromosomes was performed in 0.6% agarose(FastLane, FMC, Rockland, Me., USA) in 0.5× modified TBE (buf-fer 100 mM Tris, 100 mM boric acid; 0.2 mM NaEDTA) in a CHEFDRII apparatus (BioRad). The buffer was kept at 10°C in the elec-trophoresis chamber and changed every 48 h. Electrophoresis con-ditions and pulse times were as indicated in the legend to Fig. 4. Thegels were blotted onto nylon membranes by capillarity, and hybri-dized with a 0.694-kb PvuII probe, internal to the gdhA gene, la-belled by nick translation (Gutiérrez et al. 1991). Pre-hybridizationand hybridization were made in standard buffer with 40% formamide(Sambrook et al. 1989) at 42°C.
Results
Cloning of a DNA fragment of A. awamori containingthe gdhA gene
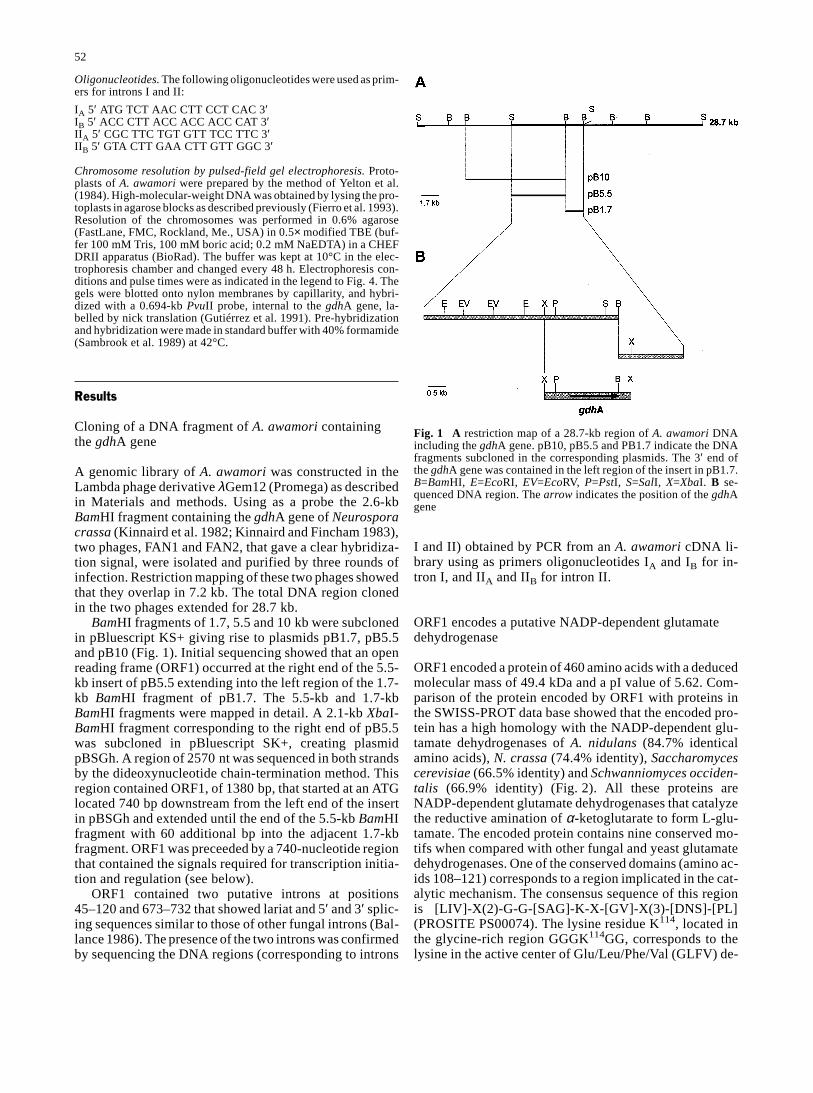
A genomic library of A. awamori was constructed in theLambda phage derivative λGem12 (Promega) as describedin Materials and methods. Using as a probe the 2.6-kbBamHI fragment containing the gdhA gene of Neurosporacrassa (Kinnaird et al. 1982; Kinnaird and Fincham 1983),two phages, FAN1 and FAN2, that gave a clear hybridiza-tion signal, were isolated and purified by three rounds ofinfection. Restriction mapping of these two phages showedthat they overlap in 7.2 kb. The total DNA region clonedin the two phages extended for 28.7 kb.
BamHI fragments of 1.7, 5.5 and 10 kb were subclonedin pBluescript KS+ giving rise to plasmids pB1.7, pB5.5and pB10 (Fig. 1). Initial sequencing showed that an openreading frame (ORF1) occurred at the right end of the 5.5-kb insert of pB5.5 extending into the left region of the 1.7-kb BamHI fragment of pB1.7. The 5.5-kb and 1.7-kbBamHI fragments were mapped in detail. A 2.1-kb XbaI-BamHI fragment corresponding to the right end of pB5.5was subcloned in pBluescript SK+, creating plasmidpBSGh. A region of 2570 nt was sequenced in both strandsby the dideoxynucleotide chain-termination method. Thisregion contained ORF1, of 1380 bp, that started at an ATGlocated 740 bp downstream from the left end of the insertin pBSGh and extended until the end of the 5.5-kb BamHIfragment with 60 additional bp into the adjacent 1.7-kbfragment. ORF1 was preceeded by a 740-nucleotide regionthat contained the signals required for transcription initia-tion and regulation (see below).
ORF1 contained two putative introns at positions45–120 and 673–732 that showed lariat and 5′ and 3′ splic-ing sequences similar to those of other fungal introns (Bal-lance 1986). The presence of the two introns was confirmedby sequencing the DNA regions (corresponding to introns
I and II) obtained by PCR from an A. awamori cDNA li-brary using as primers oligonucleotides IA and IB for in-tron I, and IIA and IIB for intron II.
ORF1 encodes a putative NADP-dependent glutamatedehydrogenase
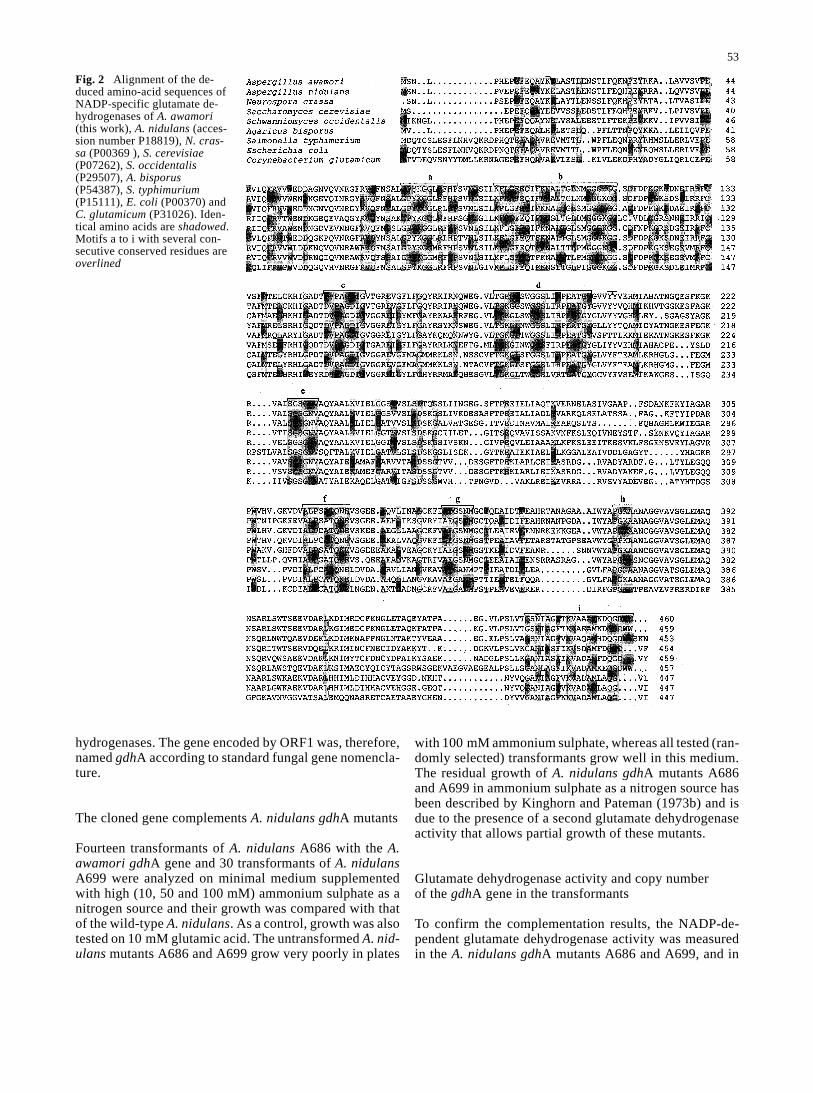
ORF1 encoded a protein of 460 amino acids with a deducedmolecular mass of 49.4 kDa and a pI value of 5.62. Com-parison of the protein encoded by ORF1 with proteins inthe SWISS-PROT data base showed that the encoded pro-tein has a high homology with the NADP-dependent glu-tamate dehydrogenases of A. nidulans (84.7% identicalamino acids), N. crassa (74.4% identity), Saccharomycescerevisiae (66.5% identity) and Schwanniomyces occiden-talis (66.9% identity) (Fig. 2). All these proteins areNADP-dependent glutamate dehydrogenases that catalyzethe reductive amination of α-ketoglutarate to form L-glu-tamate. The encoded protein contains nine conserved mo-tifs when compared with other fungal and yeast glutamatedehydrogenases. One of the conserved domains (amino ac-ids 108–121) corresponds to a region implicated in the cat-alytic mechanism. The consensus sequence of this regionis [LIV]-X(2)-G-G-[SAG]-K-X-[GV]-X(3)-[DNS]-[PL](PROSITE PS00074). The lysine residue K114, located inthe glycine-rich region GGGK114GG, corresponds to thelysine in the active center of Glu/Leu/Phe/Val (GLFV) de-
52
Fig. 1 A restriction map of a 28.7-kb region of A. awamori DNAincluding the gdhA gene. pB10, pB5.5 and PB1.7 indicate the DNAfragments subcloned in the corresponding plasmids. The 3′ end ofthe gdhA gene was contained in the left region of the insert in pB1.7.B=BamHI, E=EcoRI, EV=EcoRV, P=PstI, S=SalI, X=XbaI. B se-quenced DNA region. The arrow indicates the position of the gdhAgene
hydrogenases. The gene encoded by ORF1 was, therefore,named gdhA according to standard fungal gene nomencla-ture.
The cloned gene complements A. nidulans gdhA mutants
Fourteen transformants of A. nidulans A686 with the A.awamori gdhA gene and 30 transformants of A. nidulansA699 were analyzed on minimal medium supplementedwith high (10, 50 and 100 mM) ammonium sulphate as anitrogen source and their growth was compared with thatof the wild-type A. nidulans. As a control, growth was alsotested on 10 mM glutamic acid. The untransformed A. nid-ulans mutants A686 and A699 grow very poorly in plates
with 100 mM ammonium sulphate, whereas all tested (ran-domly selected) transformants grow well in this medium.The residual growth of A. nidulans gdhA mutants A686and A699 in ammonium sulphate as a nitrogen source hasbeen described by Kinghorn and Pateman (1973b) and isdue to the presence of a second glutamate dehydrogenaseactivity that allows partial growth of these mutants.
Glutamate dehydrogenase activity and copy numberof the gdhA gene in the transformants
To confirm the complementation results, the NADP-de-pendent glutamate dehydrogenase activity was measuredin the A. nidulans gdhA mutants A686 and A699, and in
53
Fig. 2 Alignment of the de-duced amino-acid sequences ofNADP-specific glutamate de-hydrogenases of A. awamori(this work), A. nidulans (acces-sion number P18819), N. cras-sa (P00369 ), S. cerevisiae(P07262), S. occidentalis(P29507), A. bisporus(P54387), S. typhimurium(P15111), E. coli (P00370) andC. glutamicum (P31026). Iden-tical amino acids are shadowed.Motifs a to i with several con-secutive conserved residues areoverlined
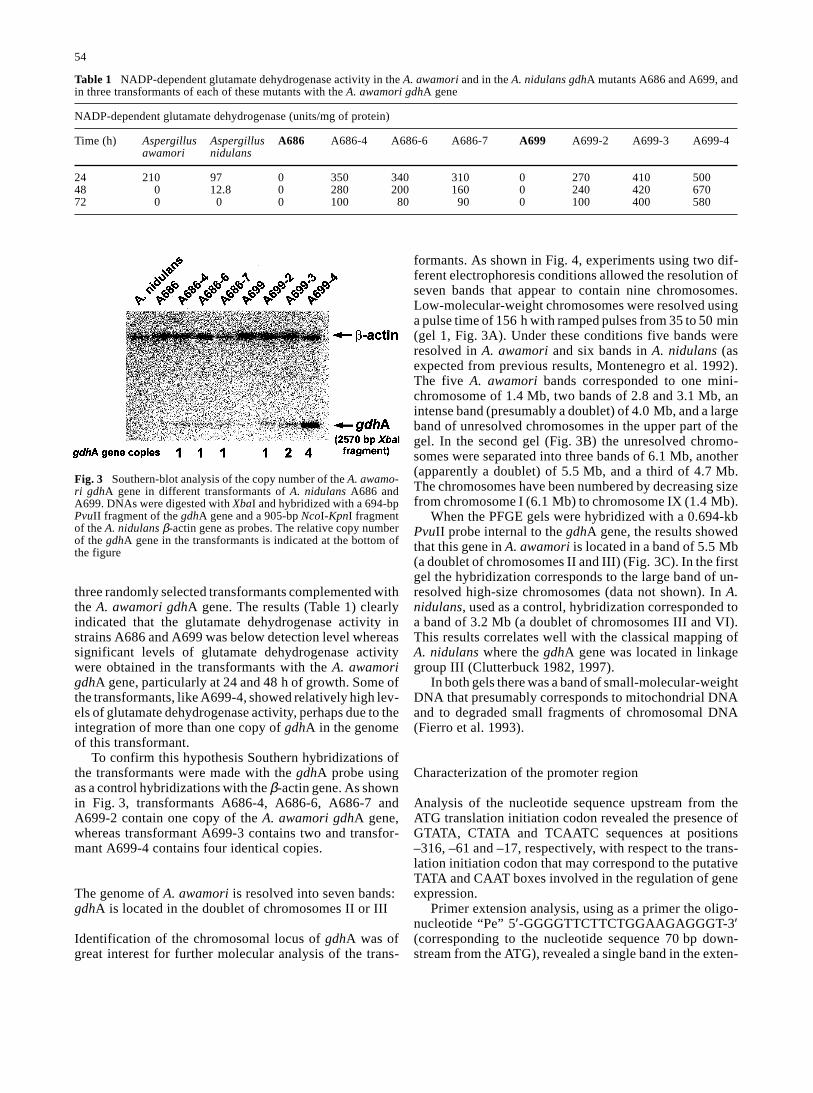
three randomly selected transformants complemented withthe A. awamori gdhA gene. The results (Table 1) clearlyindicated that the glutamate dehydrogenase activity instrains A686 and A699 was below detection level whereassignificant levels of glutamate dehydrogenase activitywere obtained in the transformants with the A. awamorigdhA gene, particularly at 24 and 48 h of growth. Some ofthe transformants, like A699-4, showed relatively high lev-els of glutamate dehydrogenase activity, perhaps due to theintegration of more than one copy of gdhA in the genomeof this transformant.
To confirm this hypothesis Southern hybridizations ofthe transformants were made with the gdhA probe usingas a control hybridizations with the β-actin gene. As shownin Fig. 3, transformants A686-4, A686-6, A686-7 andA699-2 contain one copy of the A. awamori gdhA gene,whereas transformant A699-3 contains two and transfor-mant A699-4 contains four identical copies.
The genome of A. awamori is resolved into seven bands:gdhA is located in the doublet of chromosomes II or III
Identification of the chromosomal locus of gdhA was ofgreat interest for further molecular analysis of the trans-
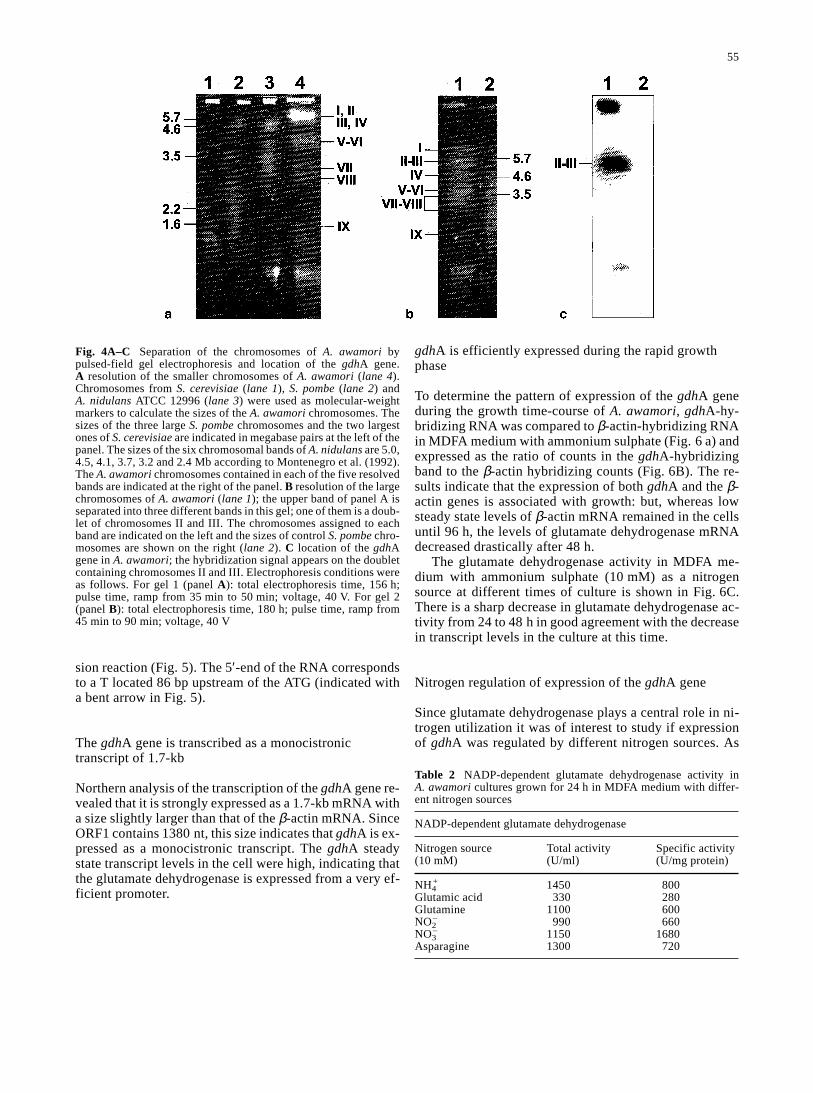
formants. As shown in Fig. 4, experiments using two dif-ferent electrophoresis conditions allowed the resolution ofseven bands that appear to contain nine chromosomes.Low-molecular-weight chromosomes were resolved usinga pulse time of 156 h with ramped pulses from 35 to 50 min(gel 1, Fig. 3A). Under these conditions five bands wereresolved in A. awamori and six bands in A. nidulans (asexpected from previous results, Montenegro et al. 1992).The five A. awamori bands corresponded to one mini-chromosome of 1.4 Mb, two bands of 2.8 and 3.1 Mb, anintense band (presumably a doublet) of 4.0 Mb, and a largeband of unresolved chromosomes in the upper part of thegel. In the second gel (Fig. 3B) the unresolved chromo-somes were separated into three bands of 6.1 Mb, another(apparently a doublet) of 5.5 Mb, and a third of 4.7 Mb.The chromosomes have been numbered by decreasing sizefrom chromosome I (6.1 Mb) to chromosome IX (1.4 Mb).
When the PFGE gels were hybridized with a 0.694-kbPvuII probe internal to the gdhA gene, the results showedthat this gene in A. awamori is located in a band of 5.5 Mb(a doublet of chromosomes II and III) (Fig. 3C). In the firstgel the hybridization corresponds to the large band of un-resolved high-size chromosomes (data not shown). In A.nidulans, used as a control, hybridization corresponded toa band of 3.2 Mb (a doublet of chromosomes III and VI).This results correlates well with the classical mapping ofA. nidulans where the gdhA gene was located in linkagegroup III (Clutterbuck 1982, 1997).
In both gels there was a band of small-molecular-weightDNA that presumably corresponds to mitochondrial DNAand to degraded small fragments of chromosomal DNA (Fierro et al. 1993).
Characterization of the promoter region
Analysis of the nucleotide sequence upstream from theATG translation initiation codon revealed the presence ofGTATA, CTATA and TCAATC sequences at positions–316, –61 and –17, respectively, with respect to the trans-lation initiation codon that may correspond to the putativeTATA and CAAT boxes involved in the regulation of geneexpression.
Primer extension analysis, using as a primer the oligo-nucleotide “Pe” 5′-GGGGTTCTTCTGGAAGAGGGT-3′(corresponding to the nucleotide sequence 70 bp down-stream from the ATG), revealed a single band in the exten-
54
Table 1 NADP-dependent glutamate dehydrogenase activity in the A. awamori and in the A. nidulans gdhA mutants A686 and A699, andin three transformants of each of these mutants with the A. awamori gdhA gene
NADP-dependent glutamate dehydrogenase (units/mg of protein)
Time (h) Aspergillus Aspergillus A686 A686-4 A686-6 A686-7 A699 A699-2 A699-3 A699-4awamori nidulans
24 210 97 0 350 340 310 0 270 410 50048 0 12.8 0 280 200 160 0 240 420 67072 0 0 0 100 80 90 0 100 400 580
Fig. 3 Southern-blot analysis of the copy number of the A. awamo-ri gdhA gene in different transformants of A. nidulans A686 andA699. DNAs were digested with XbaI and hybridized with a 694-bpPvuII fragment of the gdhA gene and a 905-bp NcoI-KpnI fragmentof the A. nidulans β-actin gene as probes. The relative copy numberof the gdhA gene in the transformants is indicated at the bottom ofthe figure
sion reaction (Fig. 5). The 5′-end of the RNA correspondsto a T located 86 bp upstream of the ATG (indicated witha bent arrow in Fig. 5).
The gdhA gene is transcribed as a monocistronic transcript of 1.7-kb
Northern analysis of the transcription of the gdhA gene re-vealed that it is strongly expressed as a 1.7-kb mRNA witha size slightly larger than that of the β-actin mRNA. SinceORF1 contains 1380 nt, this size indicates that gdhA is ex-pressed as a monocistronic transcript. The gdhA steadystate transcript levels in the cell were high, indicating thatthe glutamate dehydrogenase is expressed from a very ef-ficient promoter.
gdhA is efficiently expressed during the rapid growthphase
To determine the pattern of expression of the gdhA geneduring the growth time-course of A. awamori, gdhA-hy-bridizing RNA was compared to β-actin-hybridizing RNAin MDFA medium with ammonium sulphate (Fig. 6 a) andexpressed as the ratio of counts in the gdhA-hybridizingband to the β-actin hybridizing counts (Fig. 6B). The re-sults indicate that the expression of both gdhA and the β-actin genes is associated with growth: but, whereas lowsteady state levels of β-actin mRNA remained in the cellsuntil 96 h, the levels of glutamate dehydrogenase mRNAdecreased drastically after 48 h.
The glutamate dehydrogenase activity in MDFA me-dium with ammonium sulphate (10 mM) as a nitrogensource at different times of culture is shown in Fig. 6C.There is a sharp decrease in glutamate dehydrogenase ac-tivity from 24 to 48 h in good agreement with the decreasein transcript levels in the culture at this time.
Nitrogen regulation of expression of the gdhA gene
Since glutamate dehydrogenase plays a central role in ni-trogen utilization it was of interest to study if expressionof gdhA was regulated by different nitrogen sources. As
55
Fig. 4A–C Separation of the chromosomes of A. awamori bypulsed-field gel electrophoresis and location of the gdhA gene. A resolution of the smaller chromosomes of A. awamori (lane 4).Chromosomes from S. cerevisiae (lane 1), S. pombe (lane 2) and A. nidulans ATCC 12996 (lane 3) were used as molecular-weightmarkers to calculate the sizes of the A. awamori chromosomes. Thesizes of the three large S. pombe chromosomes and the two largestones of S. cerevisiae are indicated in megabase pairs at the left of thepanel. The sizes of the six chromosomal bands of A. nidulans are 5.0,4.5, 4.1, 3.7, 3.2 and 2.4 Mb according to Montenegro et al. (1992).The A. awamori chromosomes contained in each of the five resolvedbands are indicated at the right of the panel. B resolution of the largechromosomes of A. awamori (lane 1); the upper band of panel A isseparated into three different bands in this gel; one of them is a doub-let of chromosomes II and III. The chromosomes assigned to eachband are indicated on the left and the sizes of control S. pombe chro-mosomes are shown on the right (lane 2). C location of the gdhAgene in A. awamori; the hybridization signal appears on the doubletcontaining chromosomes II and III. Electrophoresis conditions wereas follows. For gel 1 (panel A): total electrophoresis time, 156 h;pulse time, ramp from 35 min to 50 min; voltage, 40 V. For gel 2(panel B): total electrophoresis time, 180 h; pulse time, ramp from45 min to 90 min; voltage, 40 V
Table 2 NADP-dependent glutamate dehydrogenase activity in A. awamori cultures grown for 24 h in MDFA medium with differ-ent nitrogen sources
NADP-dependent glutamate dehydrogenase
Nitrogen source Total activity Specific activity(10 mM) (U/ml) (U/mg protein)
NH4+ 1450 800
Glutamic acid 330 280Glutamine 1100 600NO2
– 990 660NO3
– 1150 1680Asparagine 1300 720

shown in Fig. 7, very high gdhA transcript levels were ob-tained in 24-h cultures in MDFA medium containing NH4
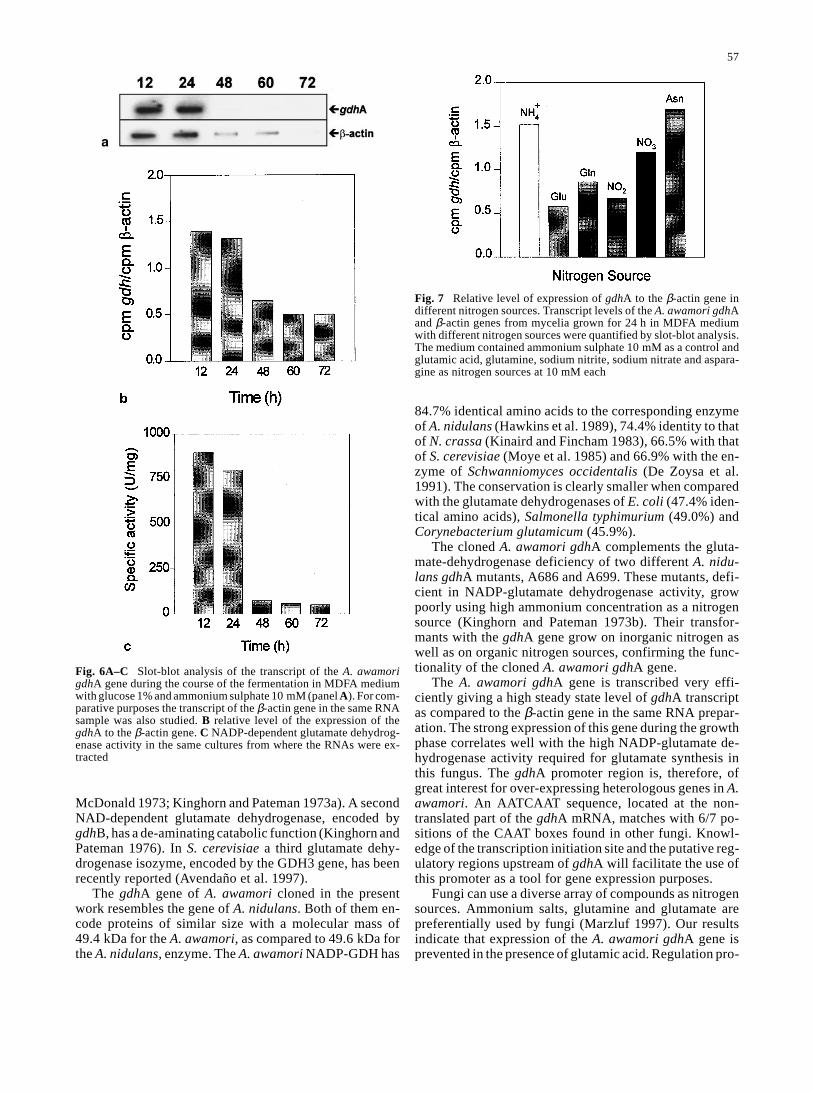
+,or asparagine, as sole nitrogen sources. Glutamic acid re-pressed the transcription of the gdhA gene whereas inter-mediate levels of expression (normalized with respect tothe β-actin) were observed in media with nitrate, glutamineor nitrite. These results indicate that the NADP-dependentglutamate dehydrogenase is subject to strong nitrogen reg-ulation at the transcriptional level.
The glutamate dehydrogenase in 24-h cultures grownin the different nitrogen sources (10 mM) is shown in Table 2. The highest activity per ml of culture was observedin cultures with NH4
+ or asparagine as nitrogen sources.These two nitrogen sources favored a strong growth. Whenexpressed per mg of protein in the cell extracts, the high-est specific activity was observed in medium with nitratedue to the low growth in MDFA medium with nitrate asthe sole nitrogen source. The lowest activity was observedin medium with glutamate as the nitrogen source, confirm-ing the results observed at the mRNA level.
Discussion
L-glutamate dehydrogenases which use either NAD orNADPH as coenzymes occur in different organisms (Brit-
56
Fig. 5A, B Primer extension identification of the 5′ end of the gdhAtranscript. The primer extension reaction was performed as indicat-ed in Materials and methods. A one protected band (arrow) is ob-served in the extension reaction (lane Pe). The G, A, T, and C lanescorrespond to the sequencing reactions of M13 phage from the –40primer. B promoter region of the gdhA gene of A. awamori (0.74-kbfragment). The GTATA, CTATA and CAAT boxes are underlined inbold face and the initiation of the transcription is indicated by a bentarrow at the T of position –86
ton et al. 1992; Benachenhou-Lahfa et al. 1993). In A. nid-ulans there are two distinct glutamate dehydrogenases en-coded by different structural genes. An NADP-dependentglutamate dehydrogenase (E. C.1.4.1.4), encoded by thegdhA gene (Gurr et al. 1986; Hawkins et al. 1989), cata-lyzes the reductive amination of α-ketoglutarate to gluta-mate with ammonium as the nitrogen source (Arst and
McDonald 1973; Kinghorn and Pateman 1973a). A secondNAD-dependent glutamate dehydrogenase, encoded bygdhB, has a de-aminating catabolic function (Kinghorn andPateman 1976). In S. cerevisiae a third glutamate dehy-drogenase isozyme, encoded by the GDH3 gene, has beenrecently reported (Avendaño et al. 1997).
The gdhA gene of A. awamori cloned in the presentwork resembles the gene of A. nidulans. Both of them en-code proteins of similar size with a molecular mass of49.4 kDa for the A. awamori, as compared to 49.6 kDa forthe A. nidulans, enzyme. The A. awamori NADP-GDH has
84.7% identical amino acids to the corresponding enzymeof A. nidulans (Hawkins et al. 1989), 74.4% identity to thatof N. crassa (Kinaird and Fincham 1983), 66.5% with thatof S. cerevisiae (Moye et al. 1985) and 66.9% with the en-zyme of Schwanniomyces occidentalis (De Zoysa et al.1991). The conservation is clearly smaller when comparedwith the glutamate dehydrogenases of E. coli (47.4% iden-tical amino acids), Salmonella typhimurium (49.0%) andCorynebacterium glutamicum (45.9%).
The cloned A. awamori gdhA complements the gluta-mate-dehydrogenase deficiency of two different A. nidu-lans gdhA mutants, A686 and A699. These mutants, defi-cient in NADP-glutamate dehydrogenase activity, growpoorly using high ammonium concentration as a nitrogensource (Kinghorn and Pateman 1973b). Their transfor-mants with the gdhA gene grow on inorganic nitrogen aswell as on organic nitrogen sources, confirming the func-tionality of the cloned A. awamori gdhA gene.
The A. awamori gdhA gene is transcribed very effi-ciently giving a high steady state level of gdhA transcriptas compared to the β-actin gene in the same RNA prepar-ation. The strong expression of this gene during the growthphase correlates well with the high NADP-glutamate de-hydrogenase activity required for glutamate synthesis inthis fungus. The gdhA promoter region is, therefore, ofgreat interest for over-expressing heterologous genes in A.awamori. An AATCAAT sequence, located at the non-translated part of the gdhA mRNA, matches with 6/7 po-sitions of the CAAT boxes found in other fungi. Knowl-edge of the transcription initiation site and the putative reg-ulatory regions upstream of gdhA will facilitate the use ofthis promoter as a tool for gene expression purposes.
Fungi can use a diverse array of compounds as nitrogensources. Ammonium salts, glutamine and glutamate arepreferentially used by fungi (Marzluf 1997). Our resultsindicate that expression of the A. awamori gdhA gene isprevented in the presence of glutamic acid. Regulation pro-
57
Fig. 6A–C Slot-blot analysis of the transcript of the A. awamorigdhA gene during the course of the fermentation in MDFA mediumwith glucose 1% and ammonium sulphate 10 mM (panel A). For com-parative purposes the transcript of the β-actin gene in the same RNAsample was also studied. B relative level of the expression of thegdhA to the β-actin gene. C NADP-dependent glutamate dehydrog-enase activity in the same cultures from where the RNAs were ex-tracted
Fig. 7 Relative level of expression of gdhA to the β-actin gene indifferent nitrogen sources. Transcript levels of the A. awamori gdhAand β-actin genes from mycelia grown for 24 h in MDFA mediumwith different nitrogen sources were quantified by slot-blot analysis.The medium contained ammonium sulphate 10 mM as a control andglutamic acid, glutamine, sodium nitrite, sodium nitrate and aspara-gine as nitrogen sources at 10 mM each

ceeds at the transcriptional level as shown by the low levelof gdhA-hybridizing RNA in glutamate-grown cultures.Similar results were observed in A. nidulans (Hawkins etal. 1989). Nitrogen control of catabolic enzymes and per-meases in fungi actually involves activation of the struc-tural genes, which is prevented in the presence of preferrednitrogen souces. Positively acting global regulatory genes,such as areA, have been isolated in A. nidulans (Kudla etal. 1990; Caddick 1992), but their relationship to the tran-scription of gdhA has not been established.
In A. nidulans the gdhA (but not the gdhB) mutants arede-repressed for a number of enzyme and uptake systemswhich are regulated by ammonium. Kinghorn and Pateman(1973b) proposed that the NADP-dependent glutamate de-hydrogenase protein plays a role in nitrogen regulation, aspart of a complex regulatory system. These authors pro-posed that the NADP-dependent glutamate dehydrogenaseis a multi-functional protein which has catalytic activityand also plays a role in the regulation of a number of otheractivities involved in nitrogen metabolism. The results ofthe Northern analysis fit well with the general model of therole of the NADP-dependent glutamate dehydrogenase innitrogen metabolism. The high expression levels of gdhAin a medium with ammonium as a nitrogen source suggestthat the increased levels of the glutamate dehydrogenaseformed may be responsible (together with other transcrip-tional factors) for the control of ammonium-regulatedgenes. The availability of the A. awamori gdhA gene willallow us to study the role of this enzyme in the nitrogenregulation of this industrially important strain.
Acknowledgements This work was supported by a grant of UR-QUIMA, S. A. (Barcelona, Spain). We thank Francisco J. Fernándezand Ana T. Marcos for helpful discussions, and B. Martín and M.Corrales for excellent technical assistance.
References
Adams RA, Royer T (1990) Complete genomic sequence encodingtrpC from Aspergillus niger var. awamori. Nucleic Acids Res18:4931
Arst HNJr, MacDonald, K ( 1973) A mutant of Aspergillus nidulanslacking NADP-linked glutamate dehydrogenase. Mol Gen Gen-et 128:111–141
Ausubel FM, Brent R, Kingston RE, Moore D, Smith JA, SeidmanJG, Struhl K (1987) Current protocols in molecular biology. JohnWiley and Sons, New York
Avendaño A, Deluna A, Olivera H, Valenzuela L, González A (1997)GHD3 encodes a glutamate dehydrogenase isozyme, a previous-ly unrecognized route for glutamate biosynthesis in Saccharo-myces cerevisiae. J Bacteriol 179:5594–5597
Ballance DJ (1986) Sequences important for gene expression in fil-amentous fungi. Yeast 2:229–236
Benachenhou-Lahfa N, Forterre P, Labedan B (1993) Evolution ofglutamate dehydrogenase genes: evidence for two paralogousprotein families and unusual branching patterns of the archaebac-teria in the universal tree of life. J Mol Evol 36:335–346
Berka RM, Ward M, Wilson LJ, Hayenga KJ, Kodama KH, Carlo-magno LP, Thompson SA (1990) Molecular cloning and deletionof the gene encoding aspergillopepsin A from Aspergillus awa-mori. Gene 86:153–162
Britton KL, Baker PJ, Rice DW, Stillman TJ (1992) Structural rela-tionship between the hexameric and tetrameric family of gluta-mate dehydrogenases. Eur J Biochem 209:851–859
Caddick MX (1992) Characterization of a major Aspergillus regula-tory gene, areA. In: Stahl U, Tudzynski P (eds) Molecular biol-ogy of filamentous fungi. VCH Press, Weinheim, Germany, pp 141–152
Calmels TPG, Martin F, Durand H, Tiraby G (1991) Proteolyticevents in the processing of secreted proteins in fungi. J Biotech-nol 17:51–66
Clutterbuck AJ (1982) Aspergillus nidulans (nuclear genes). In:O’Brien SJ (ed) Genetic maps, vol. 2. National Cancer Institute,Maryland, USA, pp 208–217
Clutterbuck AJ (1997) The validity of the Aspergillus nidulans link-age map. Fungal Genet Biol 21:267–277
Cohen BL (1973) The neutral and alkaline proteases of Aspergillusnidulans. J Gen Microbiol 77:521–528
De Zoysa PA, Connerton IF, Watson DC, Johnston JR (1991) Clon-ing, sequencing and expression of the Schwanniomyces occiden-talis NADP-dependent glutamate dehydrogenase gene. CurrGenet 20:219–224
Ditchburn P, Giddings B, MacDonald KD (1974) Rapid screeningfor the isolation of mutants of Aspergillus nidulans with increasedpenicillin yields. J Appl Bacteriol 37:515–523
Fierro F, Gutiérrez S, Díez B, Martín JF (1993) Resolution of fourchromosomes in penicillin-producing filamentous fungi: the pen-icillin gene cluster is located on chromosome II (9.6 Mb) in Pen-icillium notatum and chromosome I (10.4 Mb) in Penicilliumchrysogenum. Mol Gen Genet 241:573–578
Fierro F, Montenegro E, Gutiérrez S, Martín JF (1996) Mutantsblocked in penicillin biosynthesis show a deletion of the entirepenicillin gene cluster at a specific site within a conserved hex-anucleotide sequence. Appl Microbiol Biotechnol 43:597–604
Finkelstein DB, Rambosek J, Crewford MS, Soliday CL, McAda PC,Leach J (1989) Protein secretion in Aspergillus niger. In: Her-shaberger CL, Queener SW, Hegeman G (eds) Genetics and mo-lecular biology of industrial microorganisms. American Societyof Microbiology. Washington, D. C., pp 295–300
Gouka RJ, Hessing JG, Stam H, Musters W, van den Hondel CA(1995) A novel strategy for the isolation of defined pyrG mutantsand the development of a site-specific intergration system for As-pergillus awamori. Curr Genet 27:536–540
Gouka RJ, Punt PJ, Hessing JG, van den Hondel CA (1996) Analy-sis of heterologous protein production in defined recombinantAspergillus awamori strains. Appl Environ Microbiol 62:1951–1957
Gouka RJ, Punt PJ, van den Hondel CAMJJ (1997) Efficient produc-tion of secreted proteins by Aspergillus: progress, limitations andprospects. Appl Microbiol Biotechnol 47:1–11
Gurr SJ, Hawkins AR, Drainas C, Kinghorn JR (1986) Isolation andidentification of the Aspergillus nidulans gdhA gene encodingNADP-linked glutamate dehydrogenase. Curr Genet 10:761–766
Gutiérrez S, Díez B, Alvarez E, Barredo JL, Martín JF (1991) Ex-pression of the penDE gene of Penicillium chrysogenum encod-ing isopenicillin N acyltransferase in Cephalosporium acremo-nium: production of benzylpenicillin by the transformants. MolGen Genet 225:56–64
Hammond C, Helenius A (1995) Quality control in the secretorypathway. Curr Opin Cell Biol 7:523–529
Hawkins AR, Gutt SJ, Montague P, Kinghorn JR (1989) Nucleotidesequence and regulation of expression of the Aspergillus nidu-lans gdhA gene encoding NADP-dependent glutamate dehydrog-enase. Mol Gen Genet 218:105–111
Hessing JG, van Rotterdam C, Verbakel JM, Roza M, Maat J, vanGorcom RF, van den Hondel CA (1994) Isolation and character-ization of a 1,4-beta-endoxylanase gene of A. awamori. Curr Gen-et 26:228–232
Hijarrubia MJ, Casqueiro J, Gutiérrez S, Fernández FJ, Martín JF(1997) Characterization of the bip gene of Aspergillus awamoriencoding a protein with an HDEL retention signal homologousto the mammalian BiP involved in polypeptide secretion. CurrGenet 32:139–146
58

Kinghorn JR, Pateman JA (1973a) NAD and NADP L-glutamate de-hydrogenase activity and ammonium regulation in Aspergillusnidulans. J Gen Microbiol 78:39–46
Kinghorn JR, Pateman JA (1973b) A new class of mutants affectingammonium regulation and methyl-ammonium resistance in As-pergillus nidulans. Heredity 31:427
Kinghorn JR, Pateman JA (1976) Mutants of Aspergillus nidulanslacking NAD-specific glutamate dehydrogenase. J Bacteriol 125:42–47
Kinnaird JH, Fincham JRS (1983) The complete nucleotide sequenceof the Neurospora crassa am (NADPH-specific glutamate dehy-drogenase) gene. Gene 26:253–260
Kinnaird JH, Keighren MA, Kinsey JA, Eaton M, Fincham JRS(1982) Cloning of the am (glutamate dehydrogenase) gene ofNeurospora crassa through the use of a synthetic DNA probe.Gene 20:387–396
Korman DR, Baylis FT, Barnett CC, Carmona CL, Kodama KH, Roy-er TJ, Thompson SA, Ward M, Wilson LJ, Berka RM (1990) Clon-ing, characterization, and expression of two alpha-amylase genesfrom Aspergillus niger var. awamori. Curr Genet 17:203–212
Kudla B, Caddick MX, Langdon T, Martínez-Rossi NM, Bennett CF,Sibley S, Davis RW, Arst HN (1990) The regulatory gene areAmediating nitrogen metabolite repression in Aspergillus nidulans.Mutations affecting specificity of gene activation alter a loop res-idue of a putative zinc finger. EMBO J 9:1355–1364
Marzluf GA (1997) Genetic regulation of nitrogen metabolism in thefungi. Microbiol Mol Biol Rev 61:17–32
Montenegro E, Fierro F, Fernández FJ, Gutiérrez S, Martín JF (1992)Resolution of chromosomes III and VI of Aspergillus nidulansby pulsed-field gel electrophoresis shows that the penicillin bio-synthetic pathway genes pcbAB, pcbC, and penDE are clusteredon chromosome VI (3.0 megabases). J Bacteriol 174:7063–7067
Moye WS, Amuro N, Rao JKM, Zalkin H (1985) Nucleotide se-quence of yeast GDH1 encoding nicotinamide adenine dinucle-otide phosphate-dependent glutamate dehydrogenase. J BiolChem 260:8502–8508
Piddington CS, Houston CS, Paloheimo M, Cantrell M, Miettinen-Oinonen A, Nevalainen H, Rambosek J (1993) The cloning andsequencing of the genes encoding phytase (phy) and pH 2.5-op-timum acid phosphatase (aph) from Aspergillus niger var. awa-mori. Gene 133:55–62
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular cloning: a la-boratory manual. Cold Spring Harbor Laboratory, Cold SpringHarbor, New York
Sanger F, Nicklen S, Coulson AR (1977) DNA sequencing withchain-terminating inhibitors. Proc Natl Acad Sci USA 74:5463–5467
Shen Y-Q, Heim J, Solomon NA, Wolfe S, Demain AL (1984) Re-pression of β-lactam production in Cephalosporium acremoniumby nitrogen sources. J Antibiot 37:503–511
Specht CA, DiRusso CC, Novotny CP, Ullrich RC (1982) A meth-od for extracting high-molecular-weight deoxyribonucleic acidfrom fungi. Anal Biochem 119:158–163
Thompson SA (1990) Molecular cloning and deletion of the gene en-coding aspergillopepsin A from Aspergillus awamori. Gene96:313
Verdoes JC, Punt PJ, van den Hondel CAMJJ (1995) Molecular ge-netic strain improvement for the over-production of fungal pro-teins by filamentous fungi. Appl Microbiol Biotechnol 43:195–205
Yelton MM, Hamer JE, Timberlake WE (1984) Transformation ofAspergillus nidulans by using a trpC plasmid. Proc Natl AcadSci USA 81:1470–1474
59
Related Documents











