CHAPTER 5 ION CHANNEL

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

CHAPTER 5ION CHANNEL

Molecules are diffused into the cell
1. Lipid soluble molecules
2.Through channels

Plasma membrane
FIGURE 3-15 DIFFUSION ACROSS THE PLASMA MEMBRANE
EXTRACELLULAR FLUID
CYTOPLASM
Lipid-soluble moleculesdiffuse through theplasma membrane
Channelprotein
Small water-solublemolecules and ionsdiffuse throughmembrane channels
Large molecules that cannotdiffuse through lipids cannotcross the plasma membraneunless they are transportedby a carrier mechanism

Molecules are actively transported into the cell
What means ‘actively’?
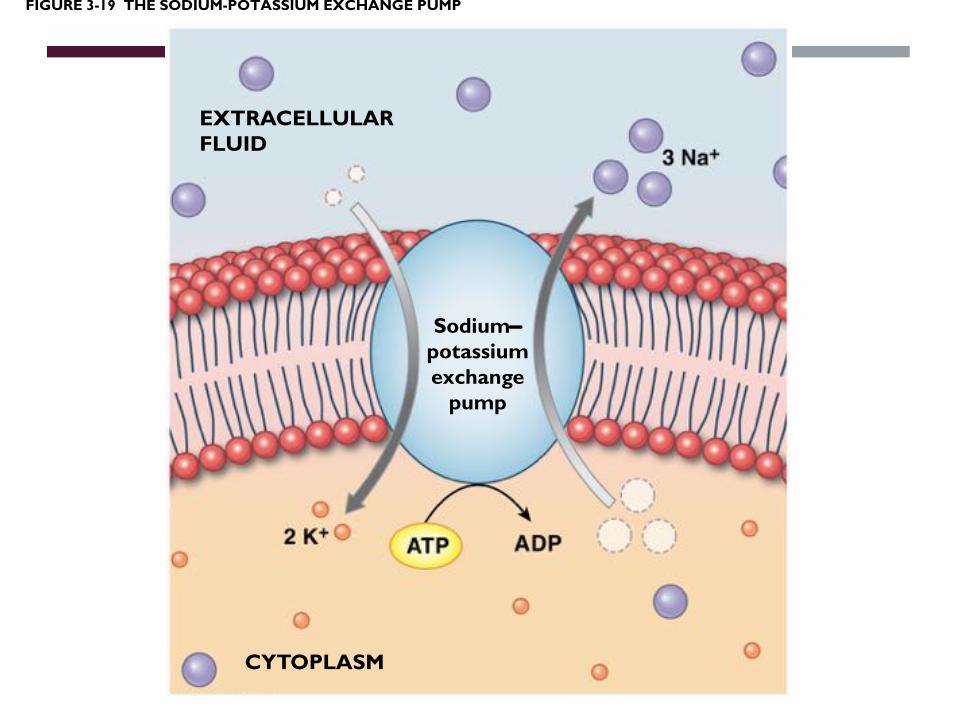
FIGURE 3-19 THE SODIUM-POTASSIUM EXCHANGE PUMP
EXTRACELLULARFLUID
Sodium-potassiumexchange
pump
CYTOPLASM

Endocytosis: getting molecules inside
Exocytosis: spitting molecules outside
vs.


THE PERMEABILITY OF THE CELL MEMBRANE
Cell membrane
Permeable?
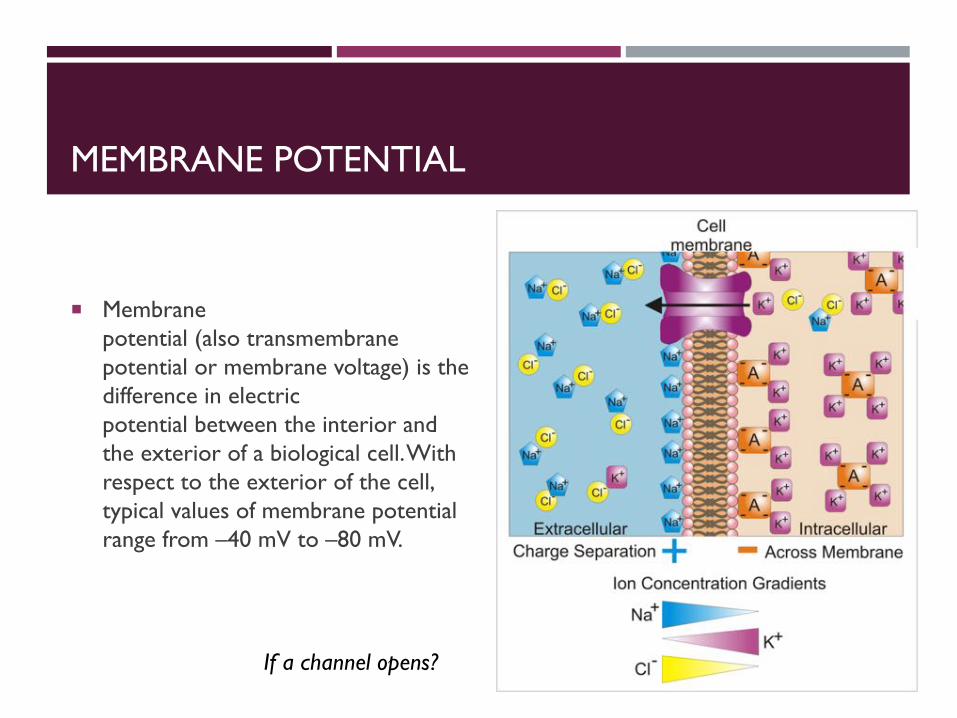
MEMBRANE POTENTIAL
¡ Membrane potential (also transmembrane potential or membrane voltage) is the difference in electric potential between the interior and the exterior of a biological cell. With respect to the exterior of the cell, typical values of membrane potential range from –40 mV to –80 mV.
If a channel opens?

HOW TO DETECT THE CURRENT?

CHARACTERISTICS OF THE CURRENT
open
close
openclose
openclose
openclose

CURRENT-VOLTAGE
Ohmic channel Rectifying channel
Characters of channels
Linear Non-linear

CHANNEL STRUCTURE & FUNCTION

THE NOBEL PRIZE IN CHEMISTRY 2003
Roderick MacKinnon Peter Agre
The Nobel Prize in Chemistry 2003 was awarded "for discoveries concerning channels in cell membranes" jointly with one half to Peter Agre "for the discovery of water channels" and with one half to Roderick MacKinnon "for structural and mechanistic studies of ion channels".
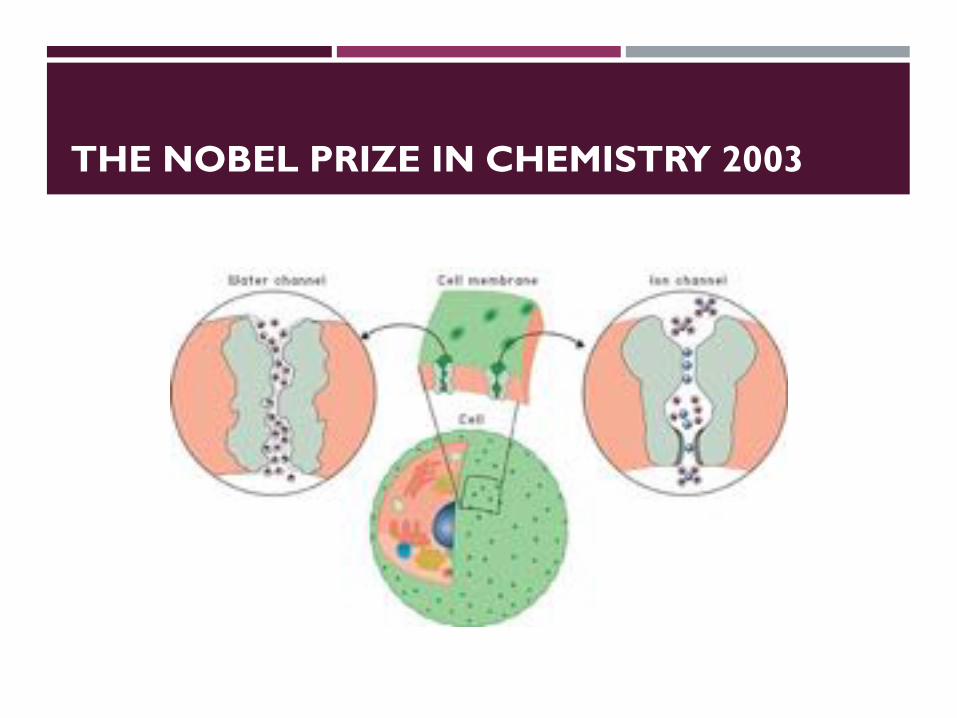
THE NOBEL PRIZE IN CHEMISTRY 2003

MACKINNON PAPERS
POTASSIUM CHANNELS AND THE ATOMIC BASISOF SELECTIVE ION CONDUCTION
Nobel Lecture, December 8, 2003
by
Roderick MacKinnon
Howard Hughes Medical Institute, Laboratory of Molecular Neurobiologyand Biophysics, Rockefeller University, 1230 York Avenue, New York, NY 10021, USA.
INTRODUCTION
All living cells are surrounded by a thin, approximately 40 Å thick lipid bilay-er called the cell membrane. The cell membrane holds the contents of a cellin one place so that the chemistry of life can occur, but it is a barrier to themovement of certain essential ingredients including the ions Na+, K+, Ca2+
and Cl-. The barrier to ion flow across the membrane – known as the dielec-tric barrier – can be understood at an intuitive level: the cell membrane inte-rior is an oily substance and ions are more stable in water than in oil. The en-ergetic preference of an ion for water arises from the electric field around theion and its interaction with neighboring molecules. Water is an electricallypolarizable substance, which means that its molecules rearrange in an ion’selectric field, pointing negative oxygen atoms in the direction of cations andpositive hydrogen atoms toward anions. These electrically stabilizing interac-tions are much weaker in a less polarizable substance such as oil. Thus, an ionwill tend to stay in the water on either side of a cell membrane rather than en-ter and cross the membrane. And yet numerous cellular processes, rangingfrom electrolyte transport across epithelia to electrical signal production inneurons, depend on the flow of ions across the membrane. To mediate theflow, specific protein catalysts known as ion channels exist in the cell mem-brane. Ion channels exhibit the following three essential properties: (1) theyconduct ions rapidly, (2) many ion channels are highly selective, meaning onlycertain ion species flow while others are excluded, (3) their function is regu-lated by processes known as gating, that is, ion conduction is turned on andoff in response to specific environmental stimuli. Figure 1 summarizes theseproperties (figure 1).
The modern history of ion channels began in 1952 when Hodgkin andHuxley published their seminal papers on the theory of the action potentialin the squid giant axon (Hodgkin and Huxley, 1952a; Hodgkin and Huxley,1952b; Hodgkin and Huxley, 1952c; Hodgkin and Huxley, 1952d). A funda-mental element of their theory was that the axon membrane undergoeschanges in its permeability to Na+ and K+ ions. The Hodgkin-Huxley theory
214
The Structure of the PotassiumChannel: Molecular Basis of K!
Conduction and SelectivityDeclan A. Doyle, Joao Morais Cabral, Richard A. Pfuetzner,
Anling Kuo, Jacqueline M. Gulbis, Steven L. Cohen,Brian T. Chait, Roderick MacKinnon*
The potassium channel from Streptomyces lividans is an integral membrane protein withsequence similarity to all known K! channels, particularly in the pore region. X-rayanalysis with data to 3.2 angstroms reveals that four identical subunits create an invertedteepee, or cone, cradling the selectivity filter of the pore in its outer end. The narrowselectivity filter is only 12 angstroms long, whereas the remainder of the pore is widerand lined with hydrophobic amino acids. A large water-filled cavity and helix dipoles arepositioned so as to overcome electrostatic destabilization of an ion in the pore at thecenter of the bilayer. Main chain carbonyl oxygen atoms from the K! channel signaturesequence line the selectivity filter, which is held open by structural constraints to co-ordinate K! ions but not smaller Na! ions. The selectivity filter contains two K! ions about7.5 angstroms apart. This configuration promotes ion conduction by exploiting electro-static repulsive forces to overcome attractive forces between K! ions and the selectivityfilter. The architecture of the pore establishes the physical principles underlying selectiveK! conduction.
Potassium ions diffuse rapidly across cellmembranes through proteins called K!
channels. This movement underlies manyfundamental biological processes, includ-ing electrical signaling in the nervous sys-tem. Potassium channels use diversemechanisms of gating (the processes bywhich the pore opens and closes), but theyall exhibit very similar ion permeabilitycharacteristics (1). All K! channels showa selectivity sequence of K! " Rb! #Cs!, whereas permeability for the smallestalkali metal ions Na! and Li! is immea-surably low. Potassium is at least 10,000times more permeant than Na!, a featurethat is essential to the function of K!
channels. Potassium channels also share aconstellation of permeability characteris-tics that is indicative of a multi-ionconduction mechanism: The flux of ionsin one direction shows high-order cou-pling to flux in the opposite direction, andionic mixtures result in anomalous con-duction behavior (2). Because of these
properties, K! channels are classifiedas “long pore channels,” invoking thenotion that multiple ions queue inside along, narrow pore in single file. Inaddition, the pores of all K! channelscan be blocked by tetraethylammonium(TEA) ions (3).
Molecular cloning and mutagenesis ex-periments have reinforced the conclusionthat all K! channels have essentially thesame pore constitution. Without exception,all contain a critical amino acid sequence,the K! channel signature sequence. Muta-tion of these amino acids disrupts the chan-nel’s ability to discriminate between K!
and Na! ions (4).Two aspects of ion conduction by K!
channels have tantalized biophysicists forthe past quarter century. First, what is thechemical basis of the impressive fidelitywith which the channel distinguishes be-tween K! and Na! ions, which are feature-less spheres of Pauling radius 1.33 Å and0.95 Å, respectively? Second, how can K!
channels be so highly selective and at thesame time, apparently paradoxically, exhib-it a throughput rate approaching the diffu-sion limit? The 104 margin by which K! isselected over Na! implies strong energeticinteractions between K! ions and the pore.And yet strong energetic interactions seemincongruent with throughput rates up to108 ions per second. How can these twoessential features of the K! channel pore bereconciled?
Potassium Channel Architecture
The amino acid sequence of the K! chan-nel from Streptomyces lividans (KcsA K!
channel) (5) is similar to that of other K!
channels, including vertebrate and inverte-brate voltage-dependent K! channels, ver-tebrate inward rectifier and Ca2!-activatedK! channels, K! channels from plants andbacteria, and cyclic nucleotide-gated cationchannels (Fig. 1). On the basis of hydro-phobicity analysis, there are two closelyrelated varieties of K! channels, those con-taining two membrane-spanning segmentsper subunit and those containing six. In allcases, the functional K! channel protein isa tetramer (6), typically of four identicalsubunits (7). Subunits of the two mem-brane-spanning variety appear to be short-ened versions of their larger counterparts, asif they simply lack the first four membrane-spanning segments. Although the KcsA K!
channel is a two membrane-spanning K!
channel, its amino acid sequence is actuallycloser to those of eukaryotic six membrane-spanning K! channels. In particular, itssequence in the pore region, located be-tween the membrane-spanning stretchesand containing the K! channel signaturesequence, is nearly identical to that foundin the Drosophila (Shaker) and vertebratevoltage-gated K! channels (Fig. 1). In anaccompanying paper, through a study of theKcsA K! channel interaction with eukary-otic K! channel toxins, we confirm thatthe KcsA pore structure is indeed very sim-ilar to that of eukaryotic K! channels andthat its structure is maintained when it isremoved from the membrane with deter-gent (8).
We have determined the KcsA K!
channel structure from residue position 23to 119 by x-ray crystallography (Table 1).The cytoplasmic carboxyl terminus (resi-dues 126 to 158) was removed in the prep-aration and the remaining residues weredisordered. The KcsA K! channel crystalsare radiation-sensitive and the diffractionpattern is anisotropic, with reflections ob-served along the best and worst directionsat 2.5 Å and 3.5 Å Bragg spacings, respec-tively. By data selection, anisotropy correc-tion, introduction of heavy atom sites bysite-directed mutagenesis, averaging, andsolvent flattening, an interpretable electrondensity map was calculated (Fig. 2, Athrough C). This map was without mainchain breaks and showed strong side chaindensity (Fig. 2C). The model was refinedwith data to 3.2 Å (the data set was 93 %complete to 3.2 Å with 67% completenessbetween 3.3 Å and 3.2 Å), maintaininghighly restrained stereochemistry and keep-ing tight noncrystallographic symmetry re-straints. The refinement procedure was
D. A. Doyle, R. A. Pfuetzner, A. Kuo, and R. MacKinnonare in the Laboratory of Molecular Neurobiology and Bio-physics and the Howard Hughes Medical Institute, Rock-efeller University, 1230 York Avenue, New York, NY10021, USA. J. M. Cabral and J. M. Gulbis are in theLaboratory of Molecular Neurobiology and Biophysics,Rockefeller University, 1230 York Avenue, New York, NY10021, USA. S. L. Cohen and B. T. Chait are in theLaboratory of Mass Spectrometry and Gaseous IonChemistry, Rockefeller University, 1230 York Avenue,New York, NY 10021, USA.
*To whom correspondence should be addressed. E-mail:[email protected]
RESEARCH ARTICLES
www.sciencemag.org ! SCIENCE ! VOL. 280 ! 3 APRIL 1998 69
on
Augu
st 8
, 201
1w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

THE NOBEL PRIZE IN CHEMISTRY 2003
Passage of water molecules through the aquaporin AQP1. Because of the positive charge at the center of the channel, positively charged ions such as H3O+, are deflected. This prevents proton leakage through the channel.

THE NOBEL PRIZE IN CHEMISTRY 2003
The ion channel permits passage of potassium ions but not sodium ions. The oxygen atoms of the ion filter form an environment very similar to the water environment outside the filter. The cell may also control opening and closing of the channel.

HOW IT WORKS? - OPENING

THREE PHYSICAL MODELS OF CHANNEL
Overall structural change
Part of structural change
Blocking particle

HOW IT WORKS? – REGULATION OF ACTIVITY
?

SEVERAL TYPES OF STIMULI CONTROL THE OPENING AND CLOSING
Ligand
Phosphorylation
Voltage-gated
Stress-gated

HOW IT WORKS? - INACTIVATION
Inactivation

VOLTAGE-GATED CHANNELS: TWO MECHANISMS
Ion binding (ex) Calcium binding
Refraction

EXOGENOUS LIGANDS CAN BIAS AN ION CHANNEL TOWARD AN OPEN OR CLOSED STATE.
Exogenous ligand…
Reversibledrug Irreversible
drugEndogenousligand…Endogenous
drug…
ReversibleReaction
IrreversibleReaction

STRUCTURE OF CHANNEL
Transmembrane domain
Property of membrane?

THE SECONDARY STRUCTURE OF MEMBRANE-SPANNING PROTEINS
Type 1 & II ?
Acetylcholine receptor
Hyd
roph
obic
ity
Amino acids

THREE SUPERFAMILIES
Ligand-opening
Gap channel
Voltage gated

P REGION
¡ The more external (i.e., more extracellular) portion of the pore is formed by the "P-loops" (the region between S5 and S6) of the four domains. This region is the most narrow part of the pore and is responsible for its ion selectivity.

FOUR RELATED FAMILIES OF ION CHANNELS WITH P-REGIONS
Voltage gated K+ channel
Inward rectifyingK+ channel
Glutamate openingchannel
K+ channelWith two p regions

FOUR RELATED FAMILIES OF ION CHANNELS WITH P-REGIONS
(Sandoz & Levitz, 2013)

FOUR RELATED FAMILIES OF ION CHANNELS WITH P-REGIONS
Honor, 2007
GENE EXPRESSION IN DEVELOPMENT
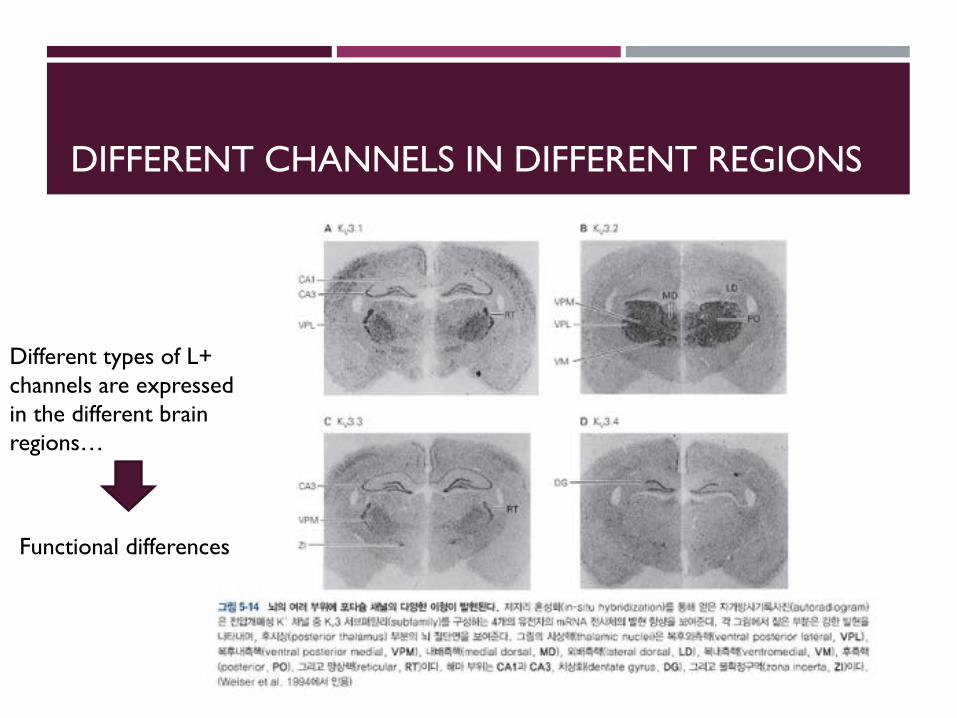
DIFFERENT CHANNELS IN DIFFERENT REGIONS
Different types of L+ channels are expressed in the different brain regions…
Functional differences

X-RAY CRYSTAL STRUCTURE

GATING OF BACTERIAL POTASSIUM CHANNELS
Close state Open state
Glycine

CIC FAMILY OF CHLORIDE CHANNELS AND TRANSPORTERS
Can you see some difference?
Link between function and structure…


COMPARISON OF GENERAL ARCHITECTURE
Parallel Anti-parallel
K+ channel Cl- channel

SPECIFICITY

HOW TO GET THE SPECIFICITY?
https://www.youtube.com/watch?v=4zms9bXM2FA

MACKINNON PAPERS
POTASSIUM CHANNELS AND THE ATOMIC BASISOF SELECTIVE ION CONDUCTION
Nobel Lecture, December 8, 2003
by
Roderick MacKinnon
Howard Hughes Medical Institute, Laboratory of Molecular Neurobiologyand Biophysics, Rockefeller University, 1230 York Avenue, New York, NY 10021, USA.
INTRODUCTION
All living cells are surrounded by a thin, approximately 40 Å thick lipid bilay-er called the cell membrane. The cell membrane holds the contents of a cellin one place so that the chemistry of life can occur, but it is a barrier to themovement of certain essential ingredients including the ions Na+, K+, Ca2+
and Cl-. The barrier to ion flow across the membrane – known as the dielec-tric barrier – can be understood at an intuitive level: the cell membrane inte-rior is an oily substance and ions are more stable in water than in oil. The en-ergetic preference of an ion for water arises from the electric field around theion and its interaction with neighboring molecules. Water is an electricallypolarizable substance, which means that its molecules rearrange in an ion’selectric field, pointing negative oxygen atoms in the direction of cations andpositive hydrogen atoms toward anions. These electrically stabilizing interac-tions are much weaker in a less polarizable substance such as oil. Thus, an ionwill tend to stay in the water on either side of a cell membrane rather than en-ter and cross the membrane. And yet numerous cellular processes, rangingfrom electrolyte transport across epithelia to electrical signal production inneurons, depend on the flow of ions across the membrane. To mediate theflow, specific protein catalysts known as ion channels exist in the cell mem-brane. Ion channels exhibit the following three essential properties: (1) theyconduct ions rapidly, (2) many ion channels are highly selective, meaning onlycertain ion species flow while others are excluded, (3) their function is regu-lated by processes known as gating, that is, ion conduction is turned on andoff in response to specific environmental stimuli. Figure 1 summarizes theseproperties (figure 1).
The modern history of ion channels began in 1952 when Hodgkin andHuxley published their seminal papers on the theory of the action potentialin the squid giant axon (Hodgkin and Huxley, 1952a; Hodgkin and Huxley,1952b; Hodgkin and Huxley, 1952c; Hodgkin and Huxley, 1952d). A funda-mental element of their theory was that the axon membrane undergoeschanges in its permeability to Na+ and K+ ions. The Hodgkin-Huxley theory
214
The Structure of the PotassiumChannel: Molecular Basis of K!
Conduction and SelectivityDeclan A. Doyle, Joao Morais Cabral, Richard A. Pfuetzner,
Anling Kuo, Jacqueline M. Gulbis, Steven L. Cohen,Brian T. Chait, Roderick MacKinnon*
The potassium channel from Streptomyces lividans is an integral membrane protein withsequence similarity to all known K! channels, particularly in the pore region. X-rayanalysis with data to 3.2 angstroms reveals that four identical subunits create an invertedteepee, or cone, cradling the selectivity filter of the pore in its outer end. The narrowselectivity filter is only 12 angstroms long, whereas the remainder of the pore is widerand lined with hydrophobic amino acids. A large water-filled cavity and helix dipoles arepositioned so as to overcome electrostatic destabilization of an ion in the pore at thecenter of the bilayer. Main chain carbonyl oxygen atoms from the K! channel signaturesequence line the selectivity filter, which is held open by structural constraints to co-ordinate K! ions but not smaller Na! ions. The selectivity filter contains two K! ions about7.5 angstroms apart. This configuration promotes ion conduction by exploiting electro-static repulsive forces to overcome attractive forces between K! ions and the selectivityfilter. The architecture of the pore establishes the physical principles underlying selectiveK! conduction.
Potassium ions diffuse rapidly across cellmembranes through proteins called K!
channels. This movement underlies manyfundamental biological processes, includ-ing electrical signaling in the nervous sys-tem. Potassium channels use diversemechanisms of gating (the processes bywhich the pore opens and closes), but theyall exhibit very similar ion permeabilitycharacteristics (1). All K! channels showa selectivity sequence of K! " Rb! #Cs!, whereas permeability for the smallestalkali metal ions Na! and Li! is immea-surably low. Potassium is at least 10,000times more permeant than Na!, a featurethat is essential to the function of K!
channels. Potassium channels also share aconstellation of permeability characteris-tics that is indicative of a multi-ionconduction mechanism: The flux of ionsin one direction shows high-order cou-pling to flux in the opposite direction, andionic mixtures result in anomalous con-duction behavior (2). Because of these
properties, K! channels are classifiedas “long pore channels,” invoking thenotion that multiple ions queue inside along, narrow pore in single file. Inaddition, the pores of all K! channelscan be blocked by tetraethylammonium(TEA) ions (3).
Molecular cloning and mutagenesis ex-periments have reinforced the conclusionthat all K! channels have essentially thesame pore constitution. Without exception,all contain a critical amino acid sequence,the K! channel signature sequence. Muta-tion of these amino acids disrupts the chan-nel’s ability to discriminate between K!
and Na! ions (4).Two aspects of ion conduction by K!
channels have tantalized biophysicists forthe past quarter century. First, what is thechemical basis of the impressive fidelitywith which the channel distinguishes be-tween K! and Na! ions, which are feature-less spheres of Pauling radius 1.33 Å and0.95 Å, respectively? Second, how can K!
channels be so highly selective and at thesame time, apparently paradoxically, exhib-it a throughput rate approaching the diffu-sion limit? The 104 margin by which K! isselected over Na! implies strong energeticinteractions between K! ions and the pore.And yet strong energetic interactions seemincongruent with throughput rates up to108 ions per second. How can these twoessential features of the K! channel pore bereconciled?
Potassium Channel Architecture
The amino acid sequence of the K! chan-nel from Streptomyces lividans (KcsA K!
channel) (5) is similar to that of other K!
channels, including vertebrate and inverte-brate voltage-dependent K! channels, ver-tebrate inward rectifier and Ca2!-activatedK! channels, K! channels from plants andbacteria, and cyclic nucleotide-gated cationchannels (Fig. 1). On the basis of hydro-phobicity analysis, there are two closelyrelated varieties of K! channels, those con-taining two membrane-spanning segmentsper subunit and those containing six. In allcases, the functional K! channel protein isa tetramer (6), typically of four identicalsubunits (7). Subunits of the two mem-brane-spanning variety appear to be short-ened versions of their larger counterparts, asif they simply lack the first four membrane-spanning segments. Although the KcsA K!
channel is a two membrane-spanning K!
channel, its amino acid sequence is actuallycloser to those of eukaryotic six membrane-spanning K! channels. In particular, itssequence in the pore region, located be-tween the membrane-spanning stretchesand containing the K! channel signaturesequence, is nearly identical to that foundin the Drosophila (Shaker) and vertebratevoltage-gated K! channels (Fig. 1). In anaccompanying paper, through a study of theKcsA K! channel interaction with eukary-otic K! channel toxins, we confirm thatthe KcsA pore structure is indeed very sim-ilar to that of eukaryotic K! channels andthat its structure is maintained when it isremoved from the membrane with deter-gent (8).
We have determined the KcsA K!
channel structure from residue position 23to 119 by x-ray crystallography (Table 1).The cytoplasmic carboxyl terminus (resi-dues 126 to 158) was removed in the prep-aration and the remaining residues weredisordered. The KcsA K! channel crystalsare radiation-sensitive and the diffractionpattern is anisotropic, with reflections ob-served along the best and worst directionsat 2.5 Å and 3.5 Å Bragg spacings, respec-tively. By data selection, anisotropy correc-tion, introduction of heavy atom sites bysite-directed mutagenesis, averaging, andsolvent flattening, an interpretable electrondensity map was calculated (Fig. 2, Athrough C). This map was without mainchain breaks and showed strong side chaindensity (Fig. 2C). The model was refinedwith data to 3.2 Å (the data set was 93 %complete to 3.2 Å with 67% completenessbetween 3.3 Å and 3.2 Å), maintaininghighly restrained stereochemistry and keep-ing tight noncrystallographic symmetry re-straints. The refinement procedure was
D. A. Doyle, R. A. Pfuetzner, A. Kuo, and R. MacKinnonare in the Laboratory of Molecular Neurobiology and Bio-physics and the Howard Hughes Medical Institute, Rock-efeller University, 1230 York Avenue, New York, NY10021, USA. J. M. Cabral and J. M. Gulbis are in theLaboratory of Molecular Neurobiology and Biophysics,Rockefeller University, 1230 York Avenue, New York, NY10021, USA. S. L. Cohen and B. T. Chait are in theLaboratory of Mass Spectrometry and Gaseous IonChemistry, Rockefeller University, 1230 York Avenue,New York, NY 10021, USA.
*To whom correspondence should be addressed. E-mail:[email protected]
RESEARCH ARTICLES
www.sciencemag.org ! SCIENCE ! VOL. 280 ! 3 APRIL 1998 69
on
Augu
st 8
, 201
1w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

NEXT…
Signal conduction
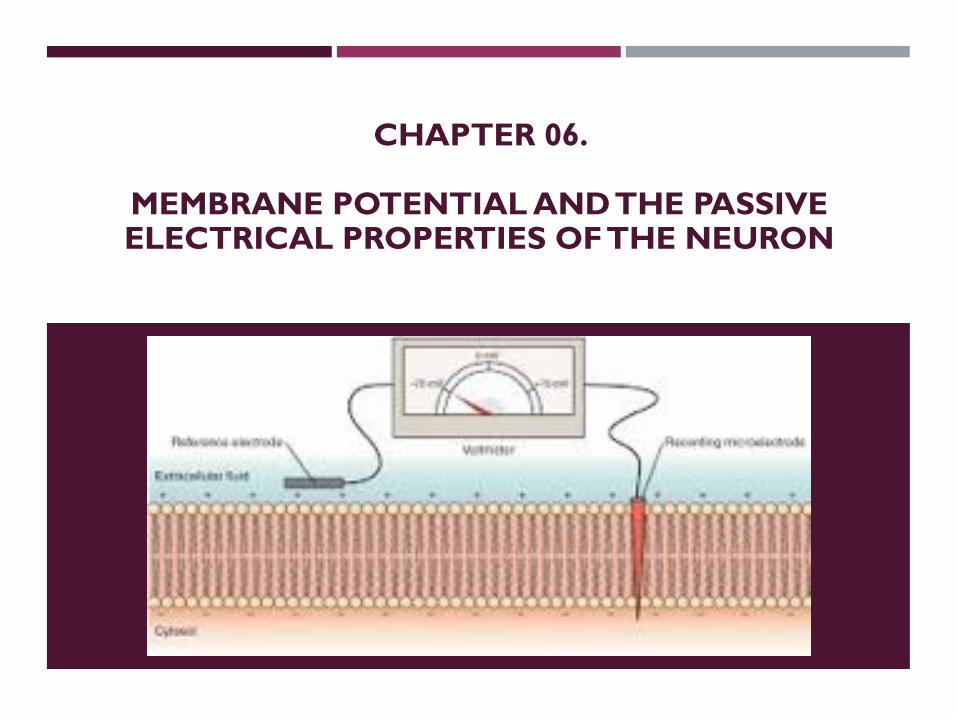
CHAPTER 06.
MEMBRANE POTENTIAL AND THE PASSIVE ELECTRICAL PROPERTIES OF THE NEURON

NeuronalconditionMembrane
Potential, Vm
Vm = Vin -Vout
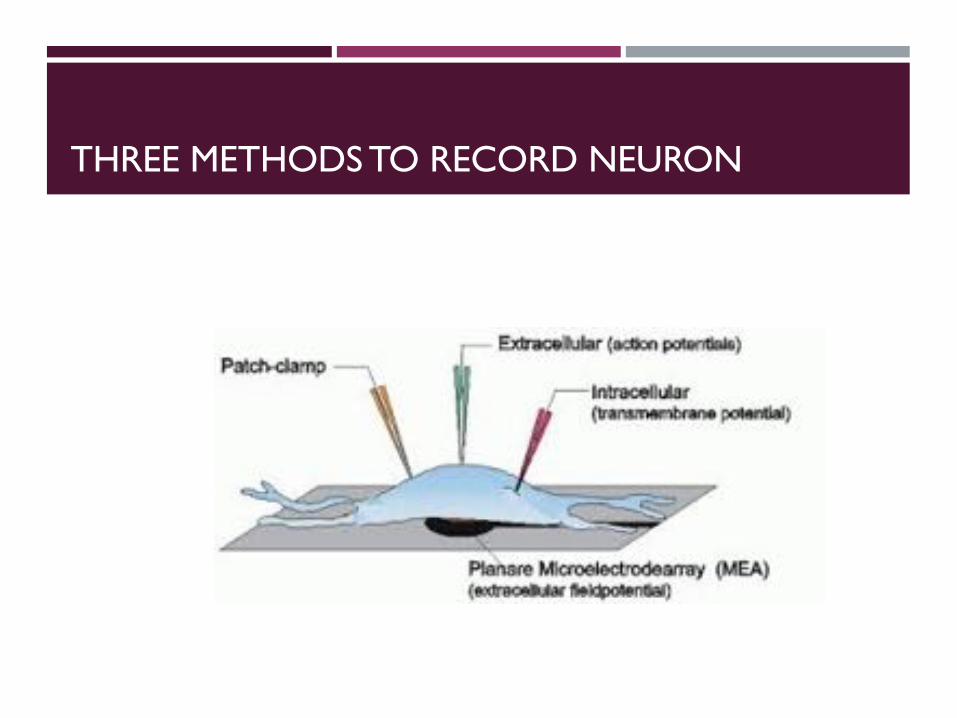
THREE METHODS TO RECORD NEURON

PATCH CLAMP RECORDING


EXTRACELLULAR RECORDING
Good for in vivo recording…

INTRA VS. EXTRACELLULAR RECORDING

Let’s talk about the intracellular recording!
Concept of Depolarization & Hyperpolarization.

Resting potential: -60 ~ - 70 mV
Imagine…


Depolarization

Hyperpolarization

How do they send information through the long neurites?

How do they send information through the long neurites? Input here?!
Ion influx occurs and then??

How do they send information through the long neurites?

Action potential(Cole & Curtis, 1939)

(Cole & Curtis, 1939)
Action potential

THE ACTION POTENTIAL IS A RAPID CHANGE IN MEMBRANE POTENTIAL
1. Depolarizationphase
2. Repolarizationphase
3. Hyperpolarization phase
Resting potential
Threshold potential

VOLTAGE-GATED CHANNELS
How voltage-gated channels work
At the resting potential, voltage-gated Na+ channels are closed.
Conformational changes openvoltage-gated channels whenthe membrane is depolarized.
Two important types:1.) Na+ voltage gated channels2.) K+ voltage gated channels

Resting Potential - Both voltage gated Na+ and K+ channels are closed.
Simplified model of AP

Initial Depolarization - Some Na+ channels open. If enough Na+ channels open, then the threshold is surpassed and an action potential is initiated.

Na+ channels open quickly. K+ channels are still closed.
PNa+ > PK+

Na+ channels self-inactivate, K+ channels are open.
PK+ >> PNa+

Emembrane ≈ E K+
PK+ > PK+ at resting state

Resting Potential - Both Na+ and K+ channels are closed.

https://www.youtube.com/watch?v=OZG8M_ldA1M
Action potential
https://www.youtube.com/watch?v=ifD1YG07fB8

NEXT…
How to travel the long, long way through axon?

READING PAPER
Nature © Macmillan Publishers Ltd 1997
The capsaicin receptor: aheat-activated ion channelin the pain pathwayMichael J. Caterina*,MarkA.Schumacher†k,MakotoTominaga*k, TobiasA.Rosen*, JonD. Levine‡ &David Julius*
Departments of * Cellular and Molecular Pharmacology, † Anesthesia, and ‡ Medicine, University of California, San Francisco, California 94143-0450, USA
k These authors contributed equally to this study.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Capsaicin, the main pungent ingredient in ‘hot’ chilli peppers, elicits a sensation of burning pain by selectivelyactivating sensoryneuronsthat convey information about noxiousstimuli to thecentral nervoussystem.Wehaveusedanexpressioncloningstrategy basedoncalcium influx to isolate a functional cDNAencodingacapsaicin receptor fromsensory neurons. This receptor is a non-selective cation channel that is structurally related to members of the TRPfamily of ion channels. The cloned capsaicin receptor is also activated by increases in temperature in the noxiousrange, suggesting that it functions as a transducer of painful thermal stimuli in vivo.
Pain is initiated when the peripheral terminals of a subgroup ofsensory neurons are activated by noxious chemical, mechanical orthermal stimuli. These neurons, called nociceptors, transmit infor-mation regarding tissue damage to pain-processing centres in thespinal cord and brain1. Nociceptors are characterized, in part, bytheir sensitivity to capsaicin, a natural product of capsicum peppersthat is the active ingredient of many ‘hot’ and spicy foods. Inmammals, exposure of nociceptor terminals to capsaicin leadsinitially to excitation of the neuron and the consequent perceptionof pain and local release of inflammatory mediators. With pro-longed exposure, nociceptor terminals become insensitive to cap-saicin, as well as to other noxious stimuli2. This latter phenomenonof nociceptor desensitization underlies the seemingly paradoxicaluse of capsaicin as an analgesic agent in the treatment of painfuldisorders ranging from viral and diabetic neuropathies to rheuma-toid arthritis3,4. Some of this decreased sensitivity to noxious stimulimay result from reversible changes in the nociceptor, but the long-term loss of responsiveness can be explained by death of thenociceptor or destruction of its peripheral terminals followingexposure to capsaicin2,5.
The cellular specificity of capsaicin action and its ability to evokethe sensation of burning pain have led to speculation that the targetof capsaicin action plays an important physiological role in thedetection of painful stimuli. Indeed, capsaicin may elicit theperception of pain by mimicking the actions of a physiologicalstimulus or an endogenous ligand produced during tissue injury6.Although the excitatory and neurotoxic properties of capsaicinhave been used extensively to define and study nociceptiveneurons, its precise mechanism of action has remained elusive.Electrophysiological7,8 and biochemical9 studies have shown thatcapsaicin excites nociceptors by increasing the permeability of theplasma membrane to cations, but the molecular mechanism under-lying this phenomenon is unclear. Proposed models range from thedirect perturbation of membrane lipids by hydrophobic capsaicinmolecules10 to the activation of a specific receptor on or withinsensory neurons6. Because capsaicin derivatives show structure–function relationships and evoke responses in a dose-dependentmanner11,12, the existence of a receptor site represents the most likelymechanism. This model has been strengthened by the developmentof capsazepine, a competitive capsaicin antagonist13, and by thediscovery of resiniferatoxin, an extremely potent capsaicin analoguefrom Euphorbia plants that mimics the cellular actions ofcapsaicin14,15. The potency of resiniferatoxin at nanomolar quantities
has led to its use as a high-affinity radioligand to visualize saturable,capsaicin- and capsazepine-sensitive binding sites on nociceptors16.
A more detailed understanding of the molecular nature ofcapsaicin action and its relationship to endogenous pain signallingmechanisms might be obtained through the cloning of a geneencoding a capsaicin receptor. To achieve this we used a functionalscreening assay to isolate a cDNA clone that reconstitutes capsaicinresponsiveness in non-neuronal cells. The deduced amino-acidsequence of this clone demonstrates that the capsaicin receptor isan integral membrane protein with homology to a family of putativestore-operated calcium channels. The cloned receptor seems to be
articles
816 NATURE | VOL 389 | 23 OCTOBER 1997
Figure 1 Expression cloning of a capsaicin receptor using calcium imaging.
HEK293 cells transiently transfected with pools of clones from a rodent dorsal root
ganglion (DRG) cDNA library were subjected to microscopic fluorescent calcium
imaging before (left) and during (right) treatment with 3 mM capsaicin. Cells
transfected with vector alone (pCDNA3; top) exhibited no response to capsaicin.
Between 1% and 5% of cells transfected with pool 11 exhibited marked increases
in cytoplasmiccalcium (middle, arrowheads). This poolwas iteratively subdivided
and reassayed until a single positive clone (VR1) was isolated (bottom). Elevated
relative calcium concentrations are indicated by an increased ratio of Fura-2
emission at 340 versus 380nm wavelength excitation (see colour bar).

ACTION POTENTIALS AND CONDUCTION

Neuron
F8-2
• Axons carry information from the cell body to the axon terminals.
• Axon terminals communicate with their target cells at synapses.
How to travel?

T3-5
• Difference in ion concentration between compartments gives rise to the resting membrane potential (RMP). Membrane permeability to these ions also influences the RMP.
• Transient changes from the RMP produce electrical signals which transmit information in nerve cells.
Changes in the Membrane Potential Produce Electric Signals in Nerve Cells

Terminology Associated with Changes in Membrane Potential
F8-7, F8-8
• Depolarization- a decrease in the potential difference between the inside and outside of the cell.
•Hyperpolarization- an increase in the potential difference between the inside and outside of the cell.
• Repolarization- returning to the RMP from either direction.
•Overshoot- when the inside of the cell becomes +ve due to the reversal of the membrane potential polarity.

• In the nervous system, different channel types are responsible for transmitting electrical signals over long and short distances:
•A) Graded potentials travel over short distances and are activated by the opening of mechanically or chemically gated channels.
•B) Action potentials travel over long distances and they are generated by the opening of voltage-gated channels.
Gated Channels Are Involved in Neuronal Signalling

Graded Potentials
F8-9
•Graded potentials are depolarizations or hyperpolarizations whose strength is proportional to the strength of the triggering event.
•Graded potentials lose their strength as they move through the cell due to the leakage of charge across the membrane (eg. leaky water hose).
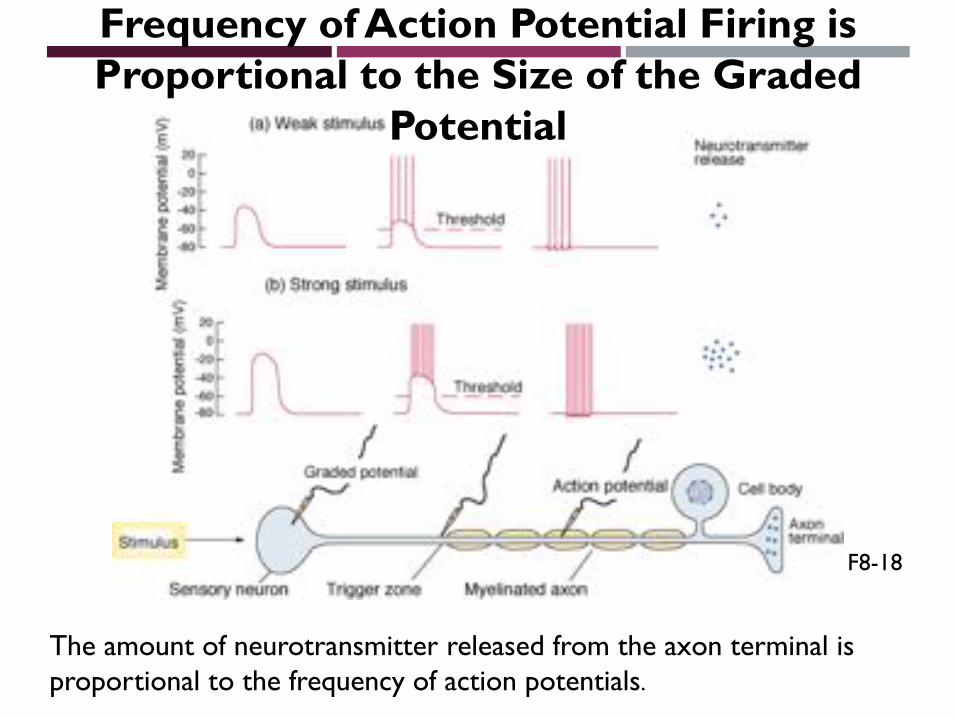
Frequency of Action Potential Firing is Proportional to the Size of the Graded
Potential
The amount of neurotransmitter released from the axon terminal is proportional to the frequency of action potentials.
F8-18

•A graded potential depolarization is called excitatory postsynaptic potential (EPSP). A graded potential hyperpolarization is called an inhibitory postsynaptic potentials(IPSP).
•They occur in the cell body and dendrites of the neuron.
•The wave of depolarization or hyperpolarization which moves through the cell with a graded potential is known as local current flow.
Question: EPSP or IPSP?

Question: See through the AP!!!

•Graded potentials travel through the neuron until they reach the trigger zone. If they depolarize the membrane above threshold voltage (about -55 mV in mammals), an action potential is triggered and it travels down the axon.
F8-10
Graded Potentials Above Threshold Voltage Trigger Action Potentials
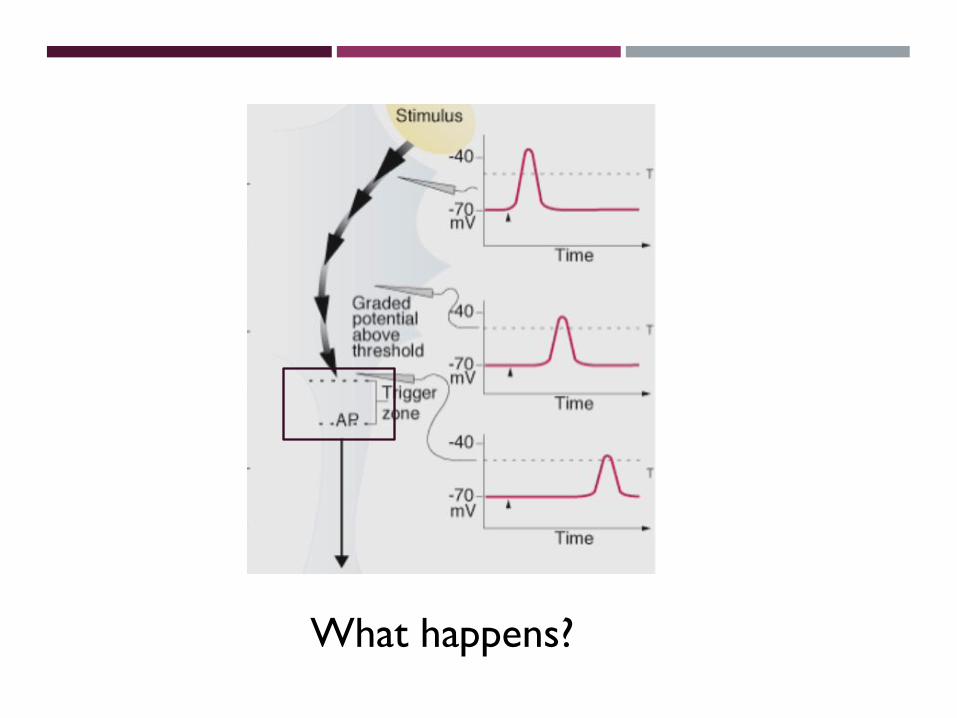
What happens?

Graded potential summation= Information summation

Spatial Summation
• A neuron may receive greater than 10, 000 inputs from presynaptic neurons.
• The initiation of an action potential from several simultaneous subthreshold graded potentials, originating from different locations, is known as spatial summation.
F8-12

Temporal Summation
• When summation occurs from graded potentials overlapping in time, it is called temporal summation.
• Summation of graded potentials demonstrates a key property of neurons: postsynaptic integration.
F8-13

Then Action potential occurs!
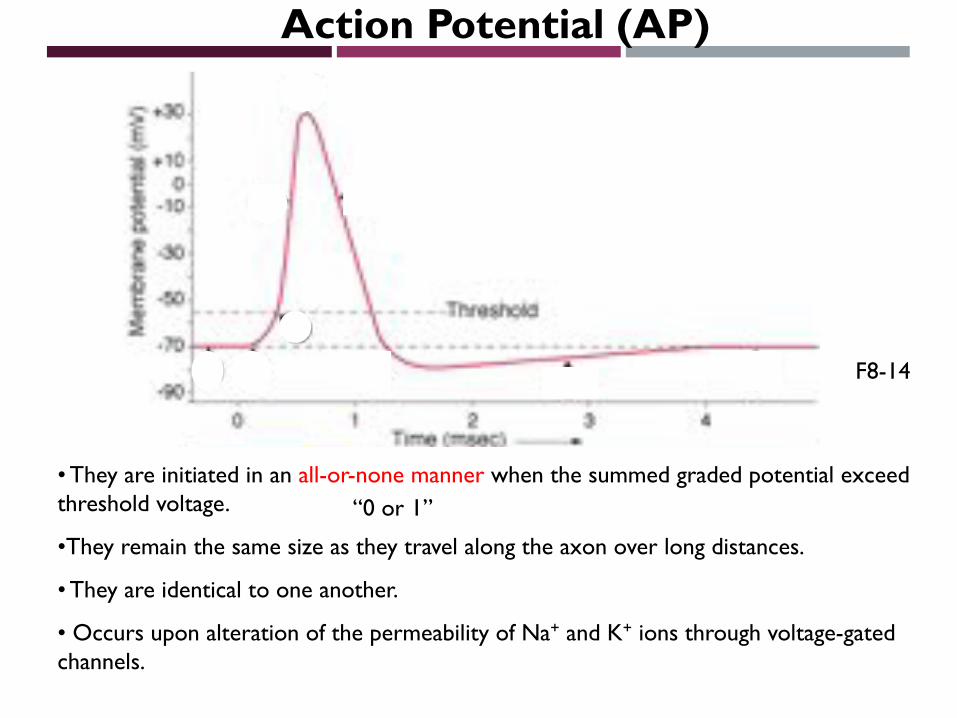
Action Potential (AP)
• They are initiated in an all-or-none manner when the summed graded potential exceed threshold voltage.
•They remain the same size as they travel along the axon over long distances.
• They are identical to one another.
• Occurs upon alteration of the permeability of Na+ and K+ ions through voltage-gated channels.
F8-14
“0 or 1”

Graded Potential vs Action Potential

Timecourse of the Action Potential

Na+ Channels Have Two Gates
F8-15

• The movement of the inactivation gate is coupled to the movement of the activation gate, but its response time is slower.
• When the activation gate is open, the signal passes along the channel protein to the inactivation gate.
•Na+/K+-ATPase Has No Direct Role to Play in the Action Potential.



Figure 1. Schematic representation of voltage-gated sodium channel α-subunits and Tetrodotoxin (TTX) binding site. Voltage-gated sodium channel α-subunits are formed by four homologous domains (DI-IV), each consisting of 6 α-helical transmembrane segments (1–6). Segment 4 (dark red) corresponds to the voltage sensors. Sites of phosphorylation by protein kinase A (PKA) and protein kinase C (PKC) are represented by yellow circles and brown squares, respectively. The fast inactivation gate (IFM motif) is located in the intracellular loop between domains 3 and 4 and is represented by h (in pink oval); pink circles show the sites involved in forming the inactivation gate receptor. P-loops are located between helices 5 and 6 (in blue), which are the pore-lining segments (as shown in the lower figure). Outer (EEDD motif) and inner (DEKA motif) rings, represented by a green and purple band, respectively (in both the upper and lower figures), are formed by the amino acids indicated by circles of the same color. The TTX molecule interacts with the amino acid residues of these two rings in the pore of the channel, as detailed in the lower figure.
Tetrodotoxin (TTX) as a Therapeutic Agent for Pain

P REGION
¡ The more external (i.e., more extracellular) portion of the pore is formed by the "P-loops" (the region between S5 and S6) of the four domains. This region is the most narrow part of the pore and is responsible for its ion selectivity.
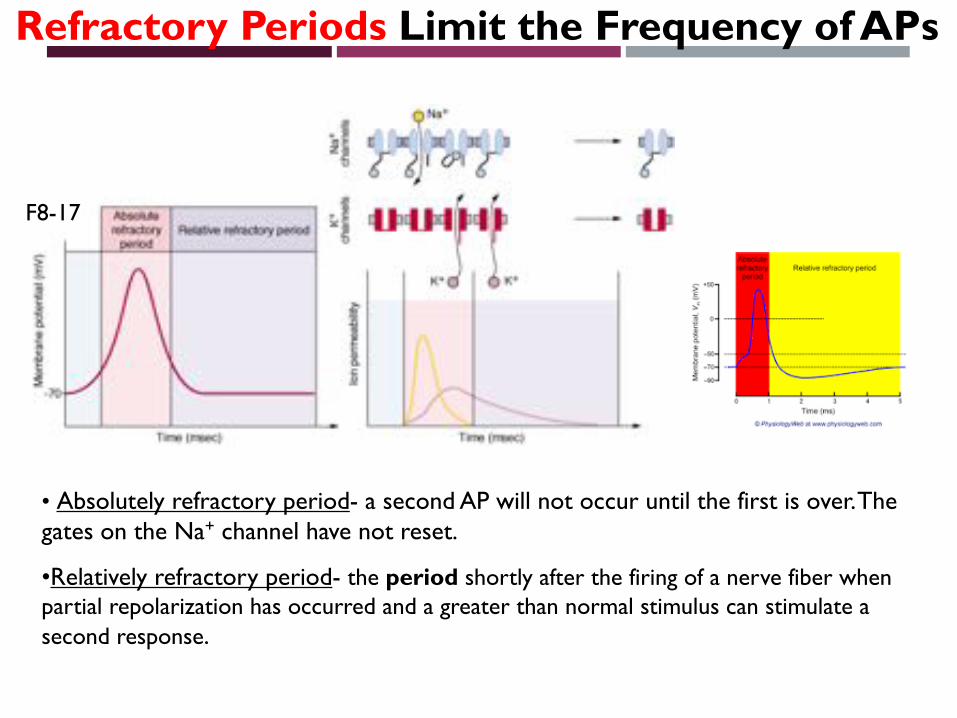
• Absolutely refractory period- a second AP will not occur until the first is over. The gates on the Na+ channel have not reset.
•Relatively refractory period- the period shortly after the firing of a nerve fiber when partial repolarization has occurred and a greater than normal stimulus can stimulate a second response.
Refractory Periods Limit the Frequency of APs
F8-17

Action Potential Conduction
• Movement of the AP along the axon at high speed is called conduction.
• A wave of action potentials travel down the axon. GP =>AP=>GP=>…
• Each section of the axon is experiencing a different phase of the AP (see figure).
F8-19

If you stimulated here…
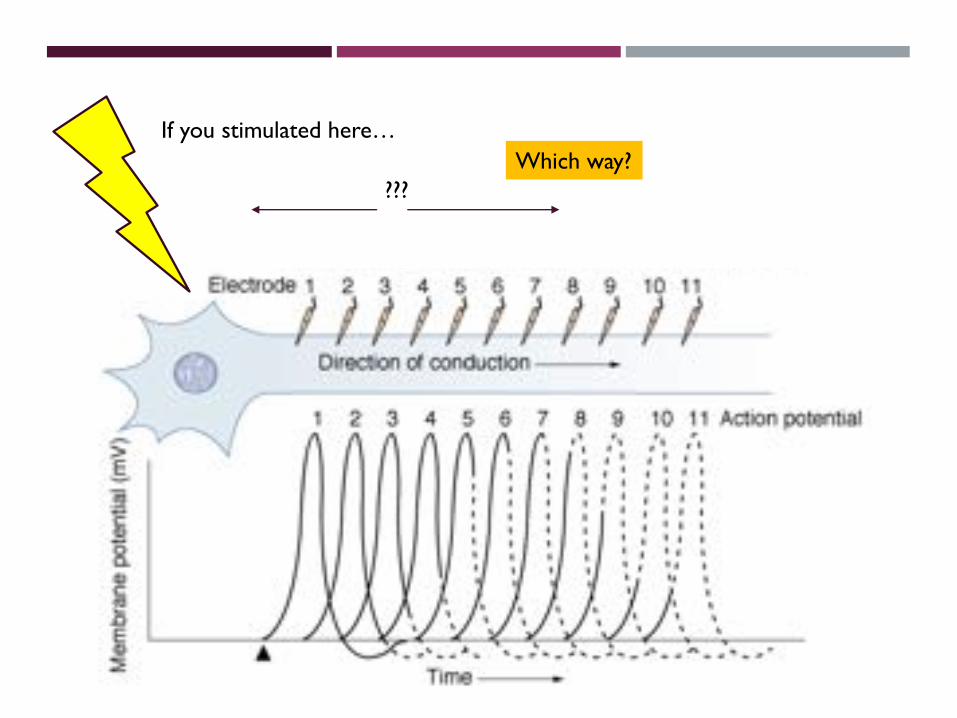
If you stimulated here…
???Which way?
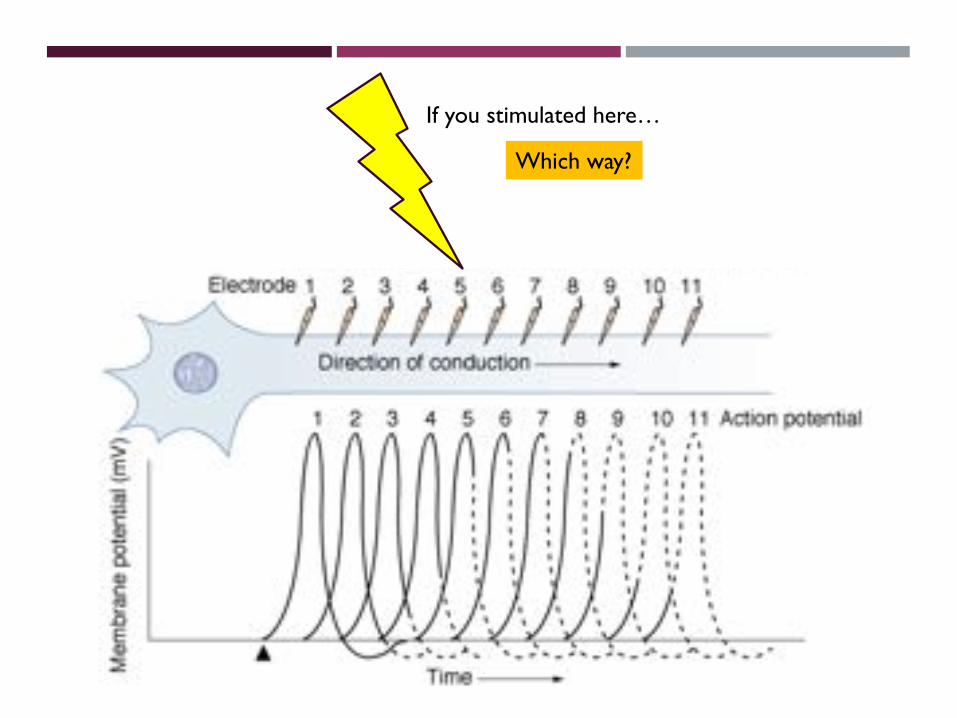
If you stimulated here…
Which way?

•Absolute refractory periods prevent back propagation of APs into the cell body.
•Refractory periods limit the rate at which signals can be transmitted down a neuron. Limit is around 100 impulses/s.

Let’s see the steps of action potential again!

• Graded potential triggers AP. Opens voltage-gated Na+ channels.
F8-20a

•The Na+ spreads in all directions attracted by the -ve ions in adjacent regions (3,4). Opens Na+ channels and initiates AP in the adjacent region along the axon (4), but not in the cell body where there are no voltage-gated Na+ channels (3).
F8-20b

•K+ channels have opened in the initial segment (5) and the Na+ (6) ions cannot trigger an AP in that region since its absolutely refractory. Na+
ions initiate action potentials in segment (7).
F8-20c

•K+ channels have opened in the initial segment (5) and the Na+ (6) ions cannot trigger an AP in that region since its absolutely refractory. Na+
ions initiate action potentials in segment (7).
F8-20c
Conduction speed?
How to make a better condition
for ion movement…
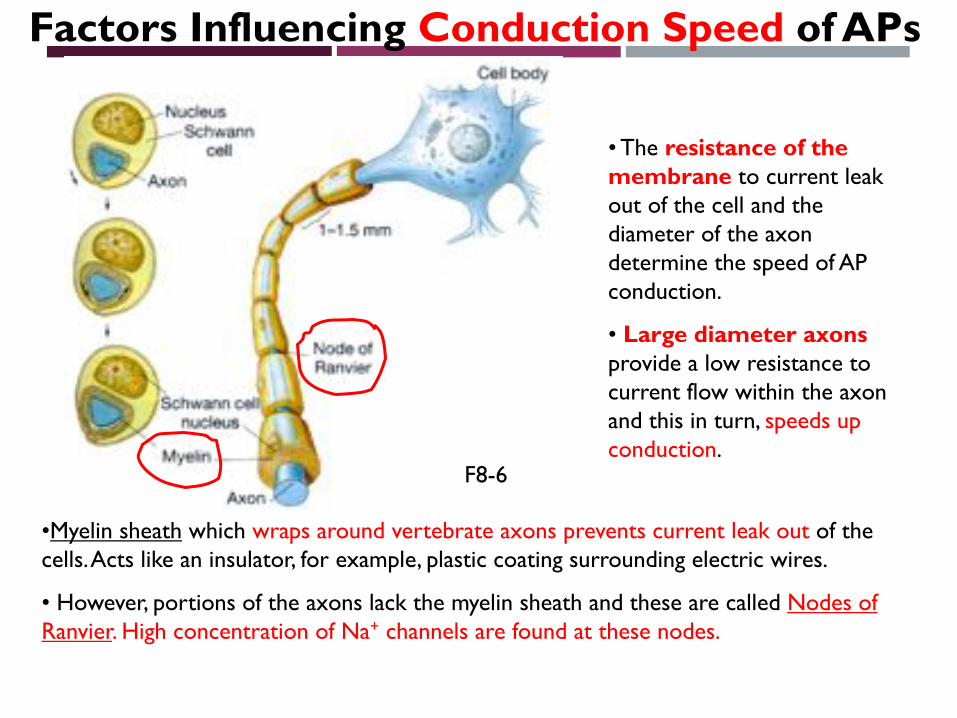
Factors Influencing Conduction Speed of APs
• The resistance of the membrane to current leak out of the cell and the diameter of the axon determine the speed of AP conduction.
• Large diameter axons provide a low resistance to current flow within the axon and this in turn, speeds up conduction.
•Myelin sheath which wraps around vertebrate axons prevents current leak out of the cells. Acts like an insulator, for example, plastic coating surrounding electric wires.
• However, portions of the axons lack the myelin sheath and these are called Nodes of Ranvier. High concentration of Na+ channels are found at these nodes.
F8-6

Saltatory Conduction
• When depolarization reaches a node, Na+ enters the axon through open channels.
• At the nodes, Na+ entry reinforces the depolarization to keep the amplitude of the AP constant, but slows the current flow due to a loss of charge to the extracellular fluid.
• However, it speeds up again when the depolarization encounters the next node.
•The apparent leapfrogging of APs from node to node along the axon is called saltatoryconduction.
F8-22
from the Latin saltare, to hop or leap

Which one is faster?

Multiple Sclerosis

Jacqueline du Pré died of Multiple Sclerosis
• In demylinating diseases, such as multiple sclerosis, the loss of myelin in the nervous system slows down the conduction of APs. Multiple sclerosis patients complain of muscle weakness, fatigue, difficulty with walking and a loss of vision.

NEXT…
Membrane potential and its property…

READING PAPER
Nature © Macmillan Publishers Ltd 1997
The capsaicin receptor: aheat-activated ion channelin the pain pathwayMichael J. Caterina*,MarkA.Schumacher†k,MakotoTominaga*k, TobiasA.Rosen*, JonD. Levine‡ &David Julius*
Departments of * Cellular and Molecular Pharmacology, † Anesthesia, and ‡ Medicine, University of California, San Francisco, California 94143-0450, USA
k These authors contributed equally to this study.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Capsaicin, the main pungent ingredient in ‘hot’ chilli peppers, elicits a sensation of burning pain by selectivelyactivating sensoryneuronsthat convey information about noxiousstimuli to thecentral nervoussystem.Wehaveusedanexpressioncloningstrategy basedoncalcium influx to isolate a functional cDNAencodingacapsaicin receptor fromsensory neurons. This receptor is a non-selective cation channel that is structurally related to members of the TRPfamily of ion channels. The cloned capsaicin receptor is also activated by increases in temperature in the noxiousrange, suggesting that it functions as a transducer of painful thermal stimuli in vivo.
Pain is initiated when the peripheral terminals of a subgroup ofsensory neurons are activated by noxious chemical, mechanical orthermal stimuli. These neurons, called nociceptors, transmit infor-mation regarding tissue damage to pain-processing centres in thespinal cord and brain1. Nociceptors are characterized, in part, bytheir sensitivity to capsaicin, a natural product of capsicum peppersthat is the active ingredient of many ‘hot’ and spicy foods. Inmammals, exposure of nociceptor terminals to capsaicin leadsinitially to excitation of the neuron and the consequent perceptionof pain and local release of inflammatory mediators. With pro-longed exposure, nociceptor terminals become insensitive to cap-saicin, as well as to other noxious stimuli2. This latter phenomenonof nociceptor desensitization underlies the seemingly paradoxicaluse of capsaicin as an analgesic agent in the treatment of painfuldisorders ranging from viral and diabetic neuropathies to rheuma-toid arthritis3,4. Some of this decreased sensitivity to noxious stimulimay result from reversible changes in the nociceptor, but the long-term loss of responsiveness can be explained by death of thenociceptor or destruction of its peripheral terminals followingexposure to capsaicin2,5.
The cellular specificity of capsaicin action and its ability to evokethe sensation of burning pain have led to speculation that the targetof capsaicin action plays an important physiological role in thedetection of painful stimuli. Indeed, capsaicin may elicit theperception of pain by mimicking the actions of a physiologicalstimulus or an endogenous ligand produced during tissue injury6.Although the excitatory and neurotoxic properties of capsaicinhave been used extensively to define and study nociceptiveneurons, its precise mechanism of action has remained elusive.Electrophysiological7,8 and biochemical9 studies have shown thatcapsaicin excites nociceptors by increasing the permeability of theplasma membrane to cations, but the molecular mechanism under-lying this phenomenon is unclear. Proposed models range from thedirect perturbation of membrane lipids by hydrophobic capsaicinmolecules10 to the activation of a specific receptor on or withinsensory neurons6. Because capsaicin derivatives show structure–function relationships and evoke responses in a dose-dependentmanner11,12, the existence of a receptor site represents the most likelymechanism. This model has been strengthened by the developmentof capsazepine, a competitive capsaicin antagonist13, and by thediscovery of resiniferatoxin, an extremely potent capsaicin analoguefrom Euphorbia plants that mimics the cellular actions ofcapsaicin14,15. The potency of resiniferatoxin at nanomolar quantities
has led to its use as a high-affinity radioligand to visualize saturable,capsaicin- and capsazepine-sensitive binding sites on nociceptors16.
A more detailed understanding of the molecular nature ofcapsaicin action and its relationship to endogenous pain signallingmechanisms might be obtained through the cloning of a geneencoding a capsaicin receptor. To achieve this we used a functionalscreening assay to isolate a cDNA clone that reconstitutes capsaicinresponsiveness in non-neuronal cells. The deduced amino-acidsequence of this clone demonstrates that the capsaicin receptor isan integral membrane protein with homology to a family of putativestore-operated calcium channels. The cloned receptor seems to be
articles
816 NATURE | VOL 389 | 23 OCTOBER 1997
Figure 1 Expression cloning of a capsaicin receptor using calcium imaging.
HEK293 cells transiently transfected with pools of clones from a rodent dorsal root
ganglion (DRG) cDNA library were subjected to microscopic fluorescent calcium
imaging before (left) and during (right) treatment with 3 mM capsaicin. Cells
transfected with vector alone (pCDNA3; top) exhibited no response to capsaicin.
Between 1% and 5% of cells transfected with pool 11 exhibited marked increases
in cytoplasmiccalcium (middle, arrowheads). This poolwas iteratively subdivided
and reassayed until a single positive clone (VR1) was isolated (bottom). Elevated
relative calcium concentrations are indicated by an increased ratio of Fura-2
emission at 340 versus 380nm wavelength excitation (see colour bar).
Cellular/Molecular
Action Potential Broadening in Capsaicin-Sensitive DRGNeurons from Frequency-Dependent Reduction of Kv3Current
Pin W. Liu, Nathaniel T. Blair, and Bruce P. BeanDepartment of Neurobiology, Harvard Medical School, Boston, Massachusetts 02115
Action potential (AP) shape is a key determinant of cellular electrophysiological behavior. We found that in small-diameter, capsaicin-sensitive dorsal root ganglia neurons corresponding to nociceptors (from rats of either sex), stimulation at frequencies as low as 1 Hzproduced progressive broadening of the APs. Stimulation at 10 Hz for 3 s resulted in an increase in AP width by an average of 76 ! 7% at22°C and by 38 ! 3% at 35°C. AP clamp experiments showed that spike broadening results from frequency-dependent reduction ofpotassium current during spike repolarization. The major current responsible for frequency-dependent reduction of overall spike-repolarizing potassium current was identified as Kv3 current by its sensitivity to low concentrations of 4-aminopyridine (IC50 "100 !M)and block by the peptide inhibitor blood depressing substance I (BDS-I). There was a small component of Kv1-mediated current duringAP repolarization, but this current did not show frequency-dependent reduction. In a small fraction of cells, there was a component ofcalcium-dependent potassium current that showed frequency-dependent reduction, but the contribution to overall potassium currentreduction was almost always much smaller than that of Kv3-mediated current. These results show that Kv3 channels make a majorcontribution to spike repolarization in small-diameter DRG neurons and undergo frequency-dependent reduction, leading to spikebroadening at moderate firing frequencies. Spike broadening from frequency-dependent reduction in Kv3 current could mitigate thefrequency-dependent decreases in conduction velocity typical of C-fiber axons.
Key words: action potential; Kv3 channels; nociceptor; spike broadening
IntroductionThe sensation of pain originates in primary afferent sensory neu-rons and some pathophysiological pain likely involves alteredexcitability of these neurons (Liu et al., 2000; Chung and Chung,2002; Katz and Gold, 2006). The excitability of dorsal root ganglia
(DRG) neurons corresponding to pain-sensing C-fibers is con-trolled by the many voltage-dependent ion channels that theyexpress, including multiple types of sodium channels, calciumchannels, and potassium channels (Gold et al., 1996; Rasband etal., 2001; Rush et al., 2007; Dib-Hajj et al., 2009; Zamponi et al.,2009). Potassium currents in DRG neurons comprise an espe-cially complex mixture of components, including multiple inac-tivating and sustained voltage-gated potassium currents (Gold etal., 1996; Safronov et al., 1996; Everill et al., 1998; Rola et al.,2003), calcium-activated potassium currents (Gold et al., 1996;Scholz et al., 1998), and sodium-activated potassium current(Nuwer et al., 2010; Martinez-Espinosa et al., 2015) Reducedpotassium currents likely contribute to aberrant nociceptor ac-tivity produced by inflammation (Nicol et al., 1997; Vaughn and
Received June 19, 2017; revised Aug. 29, 2017; accepted Aug. 30, 2017.Author contributions: P.W.L., N.T.B., and B.P.B. designed research; P.W.L. and N.T.B. performed research; P.W.L.,
N.T.B., and B.P.B. analyzed data; P.W.L., N.T.B., and B.P.B. wrote the paper.This work was supported by the National Institutes of Health (Grant NS036855).The authors declare no competing financial interests.N.T. Blair’s present address: Hydra Biosciences, 45 Moulton St., Cambridge, MA 02138.Correspondence should be addressed to Bruce P. Bean, Department of Neurobiology, Harvard Medical School,
220 Longwood Avenue, Boston, MA 02115. E-mail: [email protected]:10.1523/JNEUROSCI.1703-17.2017
Copyright © 2017 the authors 0270-6474/17/379705-10$15.00/0
Significance Statement
Small-diameter dorsal root ganglia (DRG) neurons mediating nociception and other sensory modalities express many types ofpotassium channels, but how they combine to control firing patterns and conduction is not well understood. We found that actionpotentials of small-diameter rat DRG neurons showed spike broadening at frequencies as low as 1 Hz and that spike broadeningresulted predominantly from frequency-dependent inactivation of Kv3 channels. Spike width helps to control transmitter release,conduction velocity, and firing patterns and understanding the role of particular potassium channels can help to guide newpharmacological strategies for targeting pain-sensing neurons selectively.
The Journal of Neuroscience, October 4, 2017 • 37(40):9705–9714 • 9705
In the years since the Hodgkin–Huxley analysis of the squid axon action potential1, it has become clear that most neurons contain far more than the two voltage-dependent conductances found in the squid axon2,3. Action potentials serve a very different function in neu-ronal cell bodies, where they encode information in their frequency and pattern, than in axons, where they serve primarily to rapidly propagate signals over distance. The membrane of the squid axon is a poor encoder, as it fires only over a narrow range of frequencies when stimulated by the injection of widely-varying current levels4. By contrast, most neuronal cell bodies (in both vertebrate and invertebrate animals) can fire over a far wider range of frequencies and can respond to small changes in input currents with significant changes in firing frequency5–10. Clearly, this richer firing behaviour depends on the expression of more types of voltage-dependent ion chan-nels. Interestingly, although the squid axon is strikingly deficient as an encoder, some other invertebrate axons can fire over a wide frequency range11 and have a richer repertoire of ion channel types12, as do at least some mammalian axons13.
The presence of multiple channel types in most neurons has been appreciated since at least the 1970s. However, few were prepared for the staggering number of distinct kinds of ion channels revealed over the last two decades by the convergent techniques of patch-clamp recording, heterologous expression of cloned chan-nels and genomic analysis — including, for example, more than 100 principal subunits of potassium chan-nels14. Even more surprising, perhaps, was the gradual realization of just how many distinct voltage-depend-ent conductances are expressed by individual neurons in the mammalian brain — commonly including 2 or 3 components of sodium current, 4 or 5 different
components of voltage-dependent calcium currents, at least 4 or 5 different components of voltage-activated potassium current, at least 2 to 3 types of calcium-acti-vated potassium currents, the hyperpolarization-acti-vated current Ih, and others. Because of this complexity, our understanding of how different conductances inter-act to form the action potentials of even the best-studied central neurons is still incomplete, even though Hodgkin and Huxley devised the basic experimental approach still being used — voltage-clamp analysis of individual time- and voltage-dependent conductances and reconstruction of the whole by numerical modelling — more than half a century ago1. In this review I discuss differences in the shape, rate and pattern of firing of action potentials between various types of neurons, focusing on mam-malian central neurons, and review recent advances in understanding the role of specific types of ion channels in generating these differences.
All spikes are not alikeThe shape of action potentials (BOX 1) differs consid-erably among various types of neurons in the mam-malian brain (FIG. 1). For example, in the cortex and hippocampus, GABA (γ-aminobutyric acid)-releasing interneurons generally have narrower spikes than gluta-matergic pyramidal neurons. This is seen most clearly in intracellular recordings, in which spike shape can be determined precisely8,15,16 (FIG. 1), but the difference in spike width is also evident from extracellular record-ings in vivo17. Cells with narrow spikes also commonly (but not always8) display ‘fast-spiking’ behaviour: being capable of firing at high frequencies with little decrease in frequency during prolonged stimulation5,6,8,9,15,18–20. Recently, the fast-spiking phenotype has been related to expression of the Kv3 family of voltage-gated potassium
Harvard Medical School, Department of Neurobiology, 220 Longwood Avenue, Boston, Massachusetts 02115, USA.e-mail: [email protected] doi:10.1038/nrn2148
Heterologous expressionExpression of protein molecules by the injection of complementary RNA into the cytoplasm (or complementary DNA into the nucleus) of host cells that do not normally express the proteins, such as Xenopus oocytes or mammalian cell lines.
SpikeAnother term for an action potential (especially the portion with the most rapidly changing voltage).
The action potential in mammalian central neuronsBruce P. Bean
Abstract | The action potential of the squid giant axon is formed by just two voltage-dependent conductances in the cell membrane, yet mammalian central neurons typically express more than a dozen different types of voltage-dependent ion channels. This rich repertoire of channels allows neurons to encode information by generating action potentials with a wide range of shapes, frequencies and patterns. Recent work offers an increasingly detailed understanding of how the expression of particular channel types underlies the remarkably diverse firing behaviour of various types of neurons.
R E V I E W S
NATURE REVIEWS | NEUROSCIENCE VOLUME 8 | JUNE 2007 | 451
In the years since the Hodgkin–Huxley analysis of the squid axon action potential1, it has become clear that most neurons contain far more than the two voltage-dependent conductances found in the squid axon2,3. Action potentials serve a very different function in neu-ronal cell bodies, where they encode information in their frequency and pattern, than in axons, where they serve primarily to rapidly propagate signals over distance. The membrane of the squid axon is a poor encoder, as it fires only over a narrow range of frequencies when stimulated by the injection of widely-varying current levels4. By contrast, most neuronal cell bodies (in both vertebrate and invertebrate animals) can fire over a far wider range of frequencies and can respond to small changes in input currents with significant changes in firing frequency5–10. Clearly, this richer firing behaviour depends on the expression of more types of voltage-dependent ion chan-nels. Interestingly, although the squid axon is strikingly deficient as an encoder, some other invertebrate axons can fire over a wide frequency range11 and have a richer repertoire of ion channel types12, as do at least some mammalian axons13.
The presence of multiple channel types in most neurons has been appreciated since at least the 1970s. However, few were prepared for the staggering number of distinct kinds of ion channels revealed over the last two decades by the convergent techniques of patch-clamp recording, heterologous expression of cloned chan-nels and genomic analysis — including, for example, more than 100 principal subunits of potassium chan-nels14. Even more surprising, perhaps, was the gradual realization of just how many distinct voltage-depend-ent conductances are expressed by individual neurons in the mammalian brain — commonly including 2 or 3 components of sodium current, 4 or 5 different
components of voltage-dependent calcium currents, at least 4 or 5 different components of voltage-activated potassium current, at least 2 to 3 types of calcium-acti-vated potassium currents, the hyperpolarization-acti-vated current Ih, and others. Because of this complexity, our understanding of how different conductances inter-act to form the action potentials of even the best-studied central neurons is still incomplete, even though Hodgkin and Huxley devised the basic experimental approach still being used — voltage-clamp analysis of individual time- and voltage-dependent conductances and reconstruction of the whole by numerical modelling — more than half a century ago1. In this review I discuss differences in the shape, rate and pattern of firing of action potentials between various types of neurons, focusing on mam-malian central neurons, and review recent advances in understanding the role of specific types of ion channels in generating these differences.
All spikes are not alikeThe shape of action potentials (BOX 1) differs consid-erably among various types of neurons in the mam-malian brain (FIG. 1). For example, in the cortex and hippocampus, GABA (γ-aminobutyric acid)-releasing interneurons generally have narrower spikes than gluta-matergic pyramidal neurons. This is seen most clearly in intracellular recordings, in which spike shape can be determined precisely8,15,16 (FIG. 1), but the difference in spike width is also evident from extracellular record-ings in vivo17. Cells with narrow spikes also commonly (but not always8) display ‘fast-spiking’ behaviour: being capable of firing at high frequencies with little decrease in frequency during prolonged stimulation5,6,8,9,15,18–20. Recently, the fast-spiking phenotype has been related to expression of the Kv3 family of voltage-gated potassium
Harvard Medical School, Department of Neurobiology, 220 Longwood Avenue, Boston, Massachusetts 02115, USA.e-mail: [email protected] doi:10.1038/nrn2148
Heterologous expressionExpression of protein molecules by the injection of complementary RNA into the cytoplasm (or complementary DNA into the nucleus) of host cells that do not normally express the proteins, such as Xenopus oocytes or mammalian cell lines.
SpikeAnother term for an action potential (especially the portion with the most rapidly changing voltage).
The action potential in mammalian central neuronsBruce P. Bean
Abstract | The action potential of the squid giant axon is formed by just two voltage-dependent conductances in the cell membrane, yet mammalian central neurons typically express more than a dozen different types of voltage-dependent ion channels. This rich repertoire of channels allows neurons to encode information by generating action potentials with a wide range of shapes, frequencies and patterns. Recent work offers an increasingly detailed understanding of how the expression of particular channel types underlies the remarkably diverse firing behaviour of various types of neurons.
R E V I E W S
NATURE REVIEWS | NEUROSCIENCE VOLUME 8 | JUNE 2007 | 451
© 2007 Nature Publishing Group
Related Documents

![Evidence of Ca2+-Dependent Carbohydrate Association ... · Ca2+I2+ and [2Lex + Ca2+]2+. The CID experiments of the [2Lex-LacCer + Ca2+I2+ dimers resulted in a neutral loss covalently](https://static.cupdf.com/doc/110x72/5f8af1f17b5f935beb015692/evidence-of-ca2-dependent-carbohydrate-association-ca2i2-and-2lex-ca22.jpg)









