Copyright © 2013 Pearson Canada Inc. 26 - 1 Chapter 26 DNA Restructuring: Repair, Recombination, Rearrangement, Amplification

Chapter 26
Jan 14, 2016
Chapter 26. DNA Restructuring: Repair, Recombination, Rearrangement, Amplification. Chapter 26 Outline :. DNA Repair Recombination Gene Rearrangements Gene Amplification. A summary of the major processes in information restructuring:. Endogenous DNA-damaging reactions:. - PowerPoint PPT Presentation
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Copyright © 2013 Pearson Canada Inc.26 - 1
Chapter 26
DNA Restructuring:
Repair, Recombination, Rearrangement, Amplification

Biochemistry, 4th Edition
Copyright © 2013 Pearson Canada Inc. 26 - 2
Chapter 26 Outline:
• DNA Repair• Recombination• Gene Rearrangements• Gene Amplification

Biochemistry, 4th Edition
A summary of the major processes in information restructuring:
Copyright © 2013 Pearson Canada Inc. 26 - 3

Biochemistry, 4th Edition
Endogenous DNA-damaging reactions:
Copyright © 2013 Pearson Canada Inc. 26 - 4
The approximate frequency of each reaction, in number of lesions per mammalian cell per day, is indicated.
Biochemistry, 4th Edition
Environmental DNA-damaging agents
Copyright © 2013 Pearson Canada Inc. 26 - 5
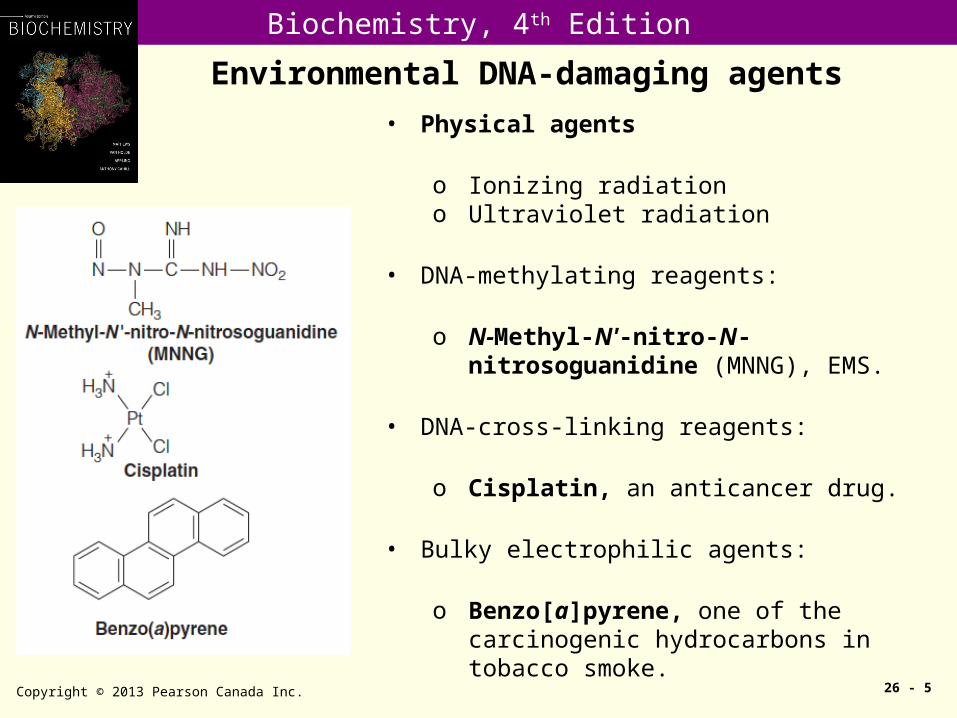
• Physical agents
o Ionizing radiationo Ultraviolet radiation
• DNA-methylating reagents:
o N-Methyl-N'-nitro-N-nitrosoguanidine (MNNG), EMS.
• DNA-cross-linking reagents:
o Cisplatin, an anticancer drug.
• Bulky electrophilic agents:
o Benzo[a]pyrene, one of the carcinogenic hydrocarbons in tobacco smoke.
Biochemistry, 4th Edition
Structures of UV-induced pyrimidine dimers
Copyright © 2013 Pearson Canada Inc. 26 - 6
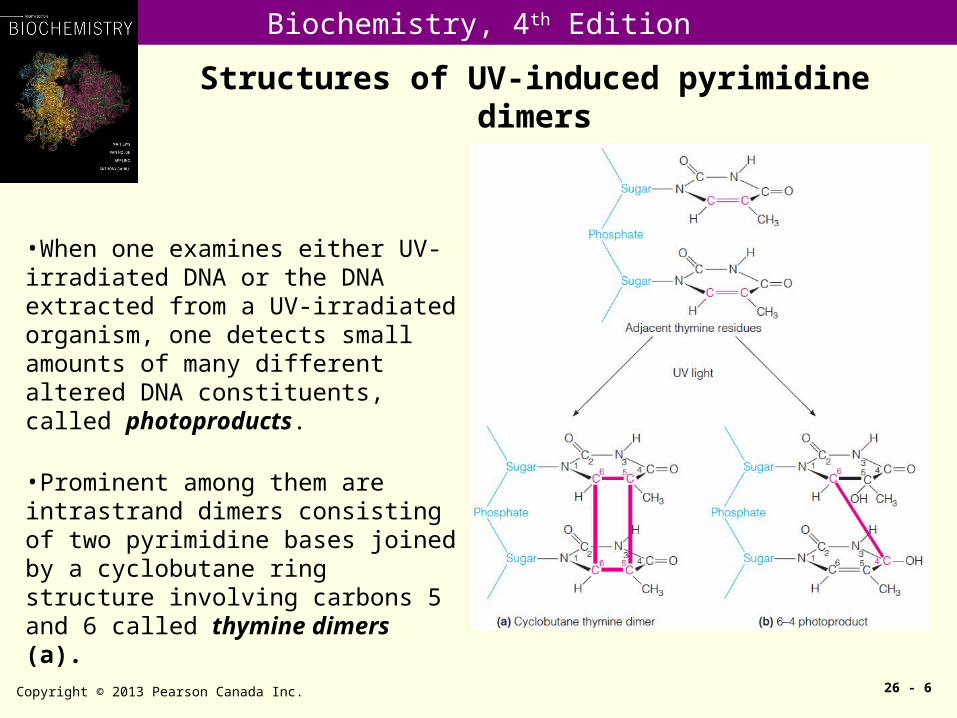
•When one examines either UV-irradiated DNA or the DNA extracted from a UV-irradiated organism, one detects small amounts of many different altered DNA constituents, called photoproducts.
•Prominent among them are intrastrand dimers consisting of two pyrimidine bases joined by a cyclobutane ring structure involving carbons 5 and 6 called thymine dimers (a).

Biochemistry, 4th Edition
Types and consequences of DNA damage
Copyright © 2013 Pearson Canada Inc. 26 - 7
(1) DNA damage may be classified as: (i) strand breaks, (ii) base loss (AP site), (iii) base damages, (iv) adducts, (v) cross-links, (vi) sugar damages, (vii) DNA-protein cross links.
(2) DNA can be repaired directly, by changing a damaged base to a normal one (direct repair), or indirectly, by replacing a DNA segment containing the damaged nucleotide (excision repair).
(3) DNA damage, if not repaired, may affect replication and transcription, leading to mutation or cell death. Coding lesions won’t interfere with replication but may produce mutation. Non-coding lesions will interfere with replication and may lead to formation of daughter-strand gaps (DSG) or double-strand breaks (DSB), which may be repaired by recombination or other mechanism.

Biochemistry, 4th Edition
Biochemistry, 4th Edition
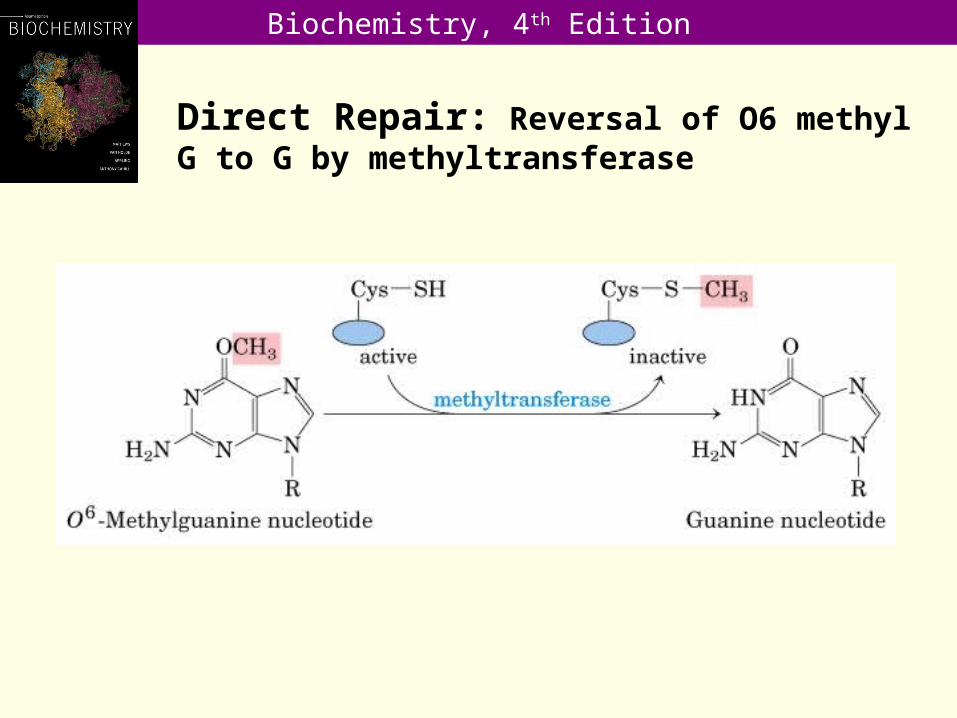
Direct Repair: Reversal of O6 methyl G to G by methyltransferase

Biochemistry, 4th Edition
Mispairing of O6-methylguanine with thymine in a DNA duplex:
Copyright © 2013 Pearson Canada Inc. 26 - 10
•O6-alkylguanine, is mutagenic because the modified base has a very high probability of pairing with thymine when the modified strand replicates.

Biochemistry, 4th Edition
Copyright © 2013 Pearson Canada Inc. 26 - 11
• Thus, alkylation of a DNA-guanine stimulates a GC AT transition.
• Repair of this type of damage involves an unusual enzyme, O6-alkylguanine alkyltransferase, which transfers a methyl or ethyl group from an O6–methylguanine or O6-ethylguanine residue to a cysteine residue in the active site of the protein.
• Having become alkylated, it cannot remove the alkyl group, and the protein molecule turns over.

Biochemistry, 4th Edition
Direct Repair: Photoreactivation by photolyase
Copyright © 2013 Pearson Canada Inc. 26 - 12
The photolyase:
•Specific for cyclobutane-type pyrimidine dimers.•Requires chromophores, such as FAD and 5,10-methylenetetrahydrofolate, as cofactors.•Chromophore absorbs light and transferring an electron to the dimer to break the pyrimidine-pyrimidine bond by a free radical mechanism.

Biochemistry, 4th Edition
Nucleotide Excision Repair
Copyright © 2013 Pearson Canada Inc. 26 - 13
Excision repair of thymine dimers by the UvrABC excinuclease of E. coli:
•A complex of A and B proteins tracks along DNA until it reaches a thymine dimer or other damaged site, where it halts and forces the DNA to bend.
•UvrA (a “molecular matchmaker”) then dissociates, allowing UvrC to bind to B.
•The BC complex cuts on both sides of the dimer.
•Helicase, polymerase, and ligase remove the damaged undecamer and replace it with new DNA.
•This system may use DNA polymerase II as well as pol I.
Biochemistry, 4th Edition
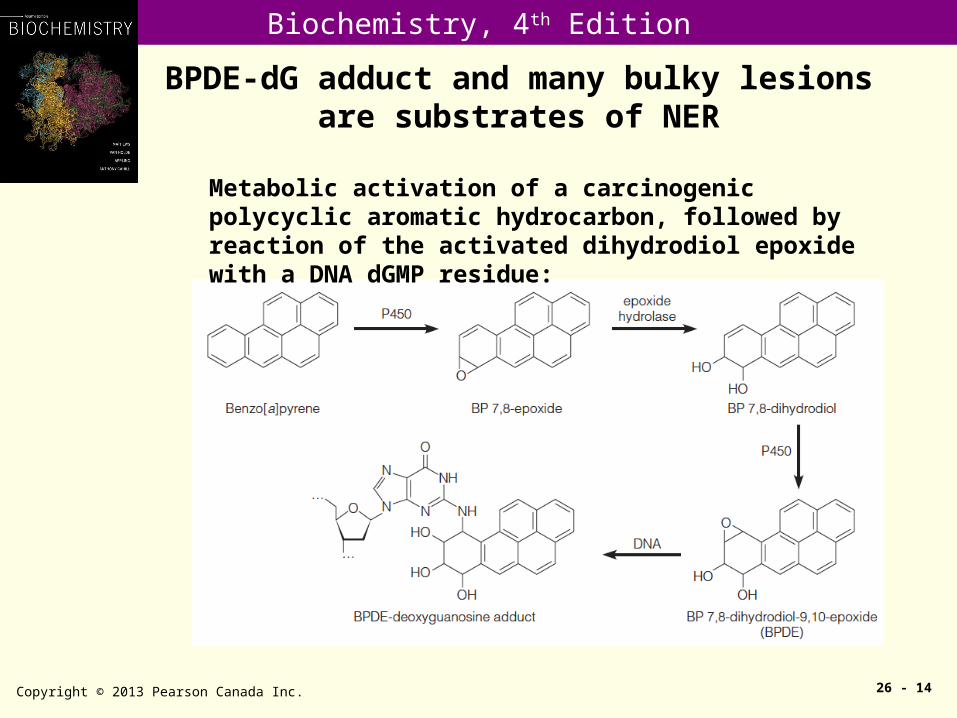
BPDE-dG adduct and many bulky lesions are substrates of NER
Copyright © 2013 Pearson Canada Inc. 26 - 14
Metabolic activation of a carcinogenic polycyclic aromatic hydrocarbon, followed by reaction of the activated dihydrodiol epoxide with a DNA dGMP residue:

Biochemistry, 4th Edition
Base excision repair (BER)
Copyright © 2013 Pearson Canada Inc. 26 - 15
Replacement of uracil in DNA by BER:
•Uracil-DNA N-glycosylase (also called Ung) removes uracil, leaving an apyrimidinic site.
•A specific endonuclease recognizes this 5’ site and cleaves on the side.
•DNA polymerase I replaces the missing nucleotide, leaving deoxyribose-5-phosphate on the 5’ side of the nick.
•This is removed hydrolytically, and DNA ligase seals the nick.
Biochemistry, 4th Edition
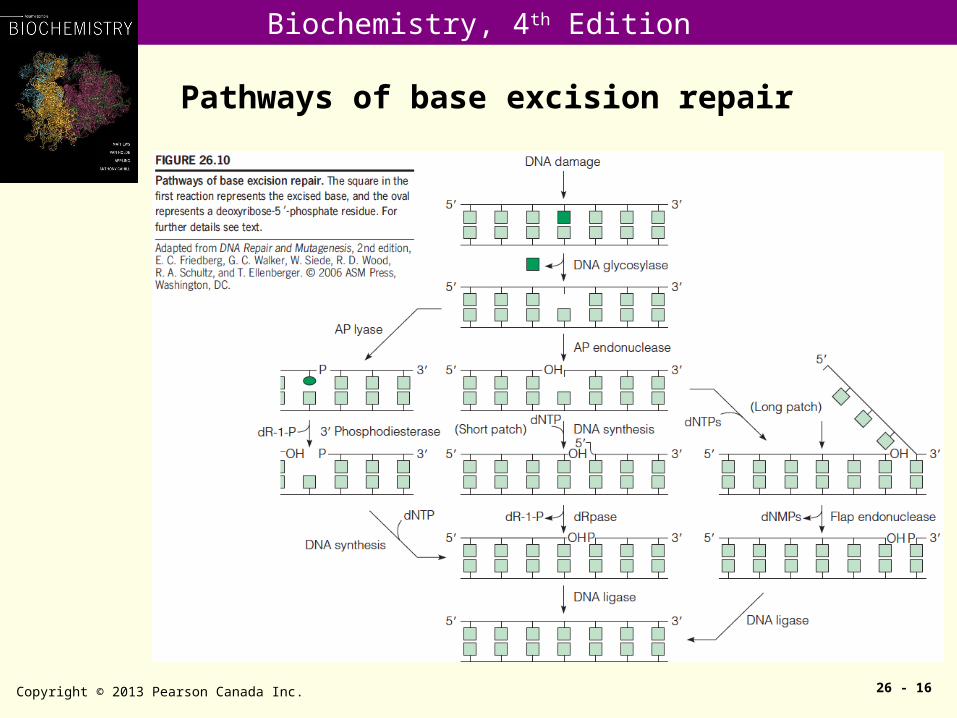
Pathways of base excision repair
Copyright © 2013 Pearson Canada Inc. 26 - 16
Biochemistry, 4th Edition
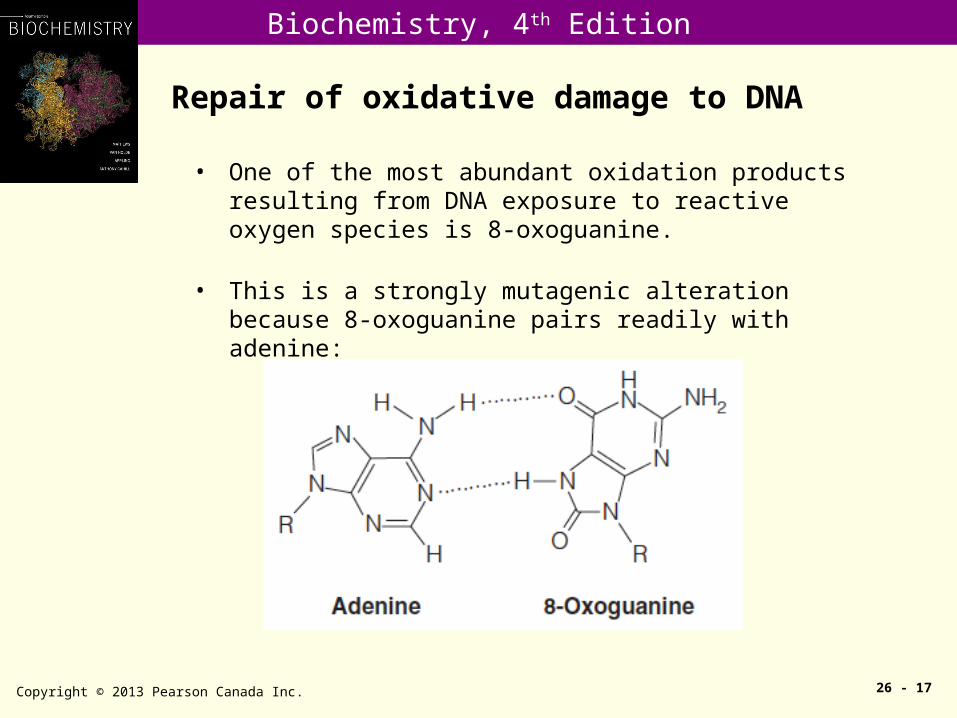
Repair of oxidative damage to DNA
Copyright © 2013 Pearson Canada Inc. 26 - 17
• One of the most abundant oxidation products resulting from DNA exposure to reactive oxygen species is 8-oxoguanine.
• This is a strongly mutagenic alteration because 8-oxoguanine pairs readily with adenine:
Biochemistry, 4th Edition
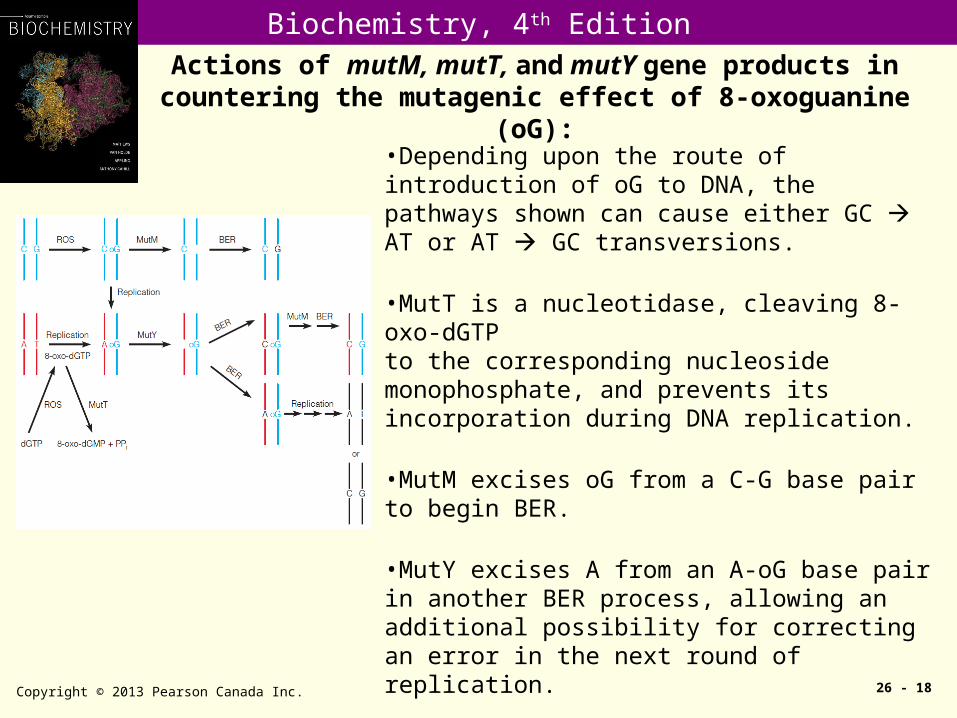
Actions of mutM, mutT, and mutY gene products in countering the mutagenic effect of 8-oxoguanine (oG):
Copyright © 2013 Pearson Canada Inc. 26 - 18
•Depending upon the route of introduction of oG to DNA, the pathways shown can cause either GC AT or AT GC transversions.
•MutT is a nucleotidase, cleaving 8-oxo-dGTPto the corresponding nucleoside monophosphate, and prevents its incorporation during DNA replication.
•MutM excises oG from a C-G base pair to begin BER.
•MutY excises A from an A-oG base pair in another BER process, allowing an additional possibility for correcting an error in the next round of replication.
Biochemistry, 4th Edition
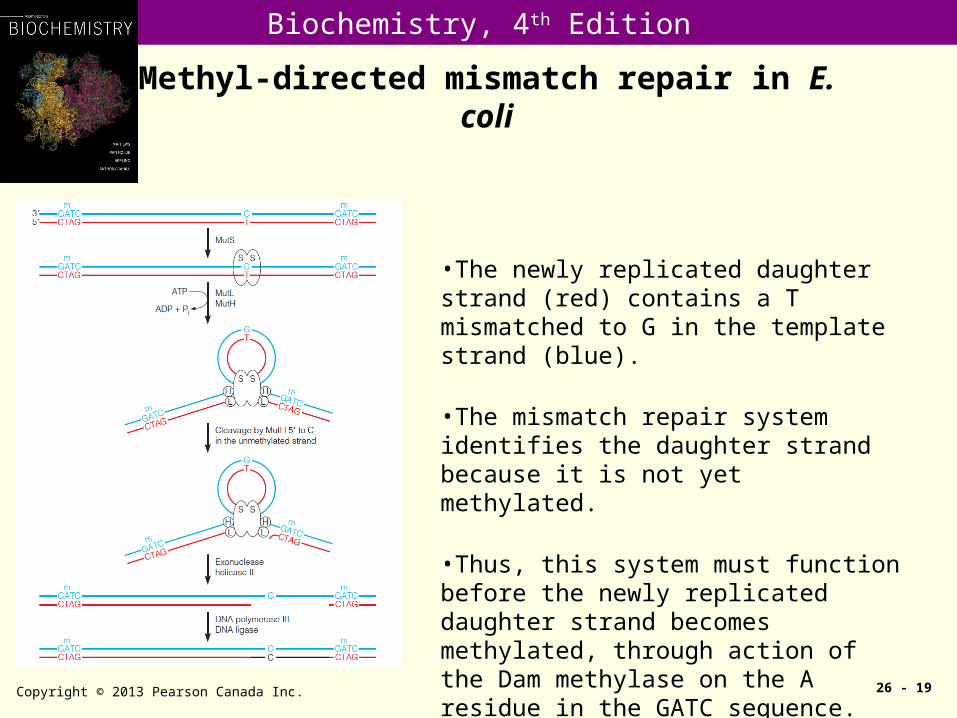
Methyl-directed mismatch repair in E. coli
Copyright © 2013 Pearson Canada Inc. 26 - 19
•The newly replicated daughter strand (red) contains a T mismatched to G in the template strand (blue).
•The mismatch repair system identifies the daughter strand because it is not yet methylated.
•Thus, this system must function before the newly replicated daughter strand becomes methylated, through action of the Dam methylase on the A residue in the GATC sequence.

Biochemistry, 4th Edition
Mismatch repair deficiency in HNPCC patients
Copyright © 2013 Pearson Canada Inc. 26 - 20
• Mutations in mismatch repair proteins were found in tumor cells from individuals with an inherited cancer predisposition called HNPCC (heritable nonpolyposis colon cancer).
• Germ-line mutations in the genes for five different mismatch repair proteins have been found to be associated with HNPCC.
• Tumor cells from those affected with HNPCC exhibit phenomenon called microsatellite instability—a large number of mutations in regions of the genome containing repeats of single-, double-, and triple-nucleotide sequences, usually with large increases in the numbers of repeating units in such sequences.
• These data suggest that the product and template strands can normally slip at such sites so that DNA polymerase copies a short repeating sequence more than once, or else skips a segment.
• This creates a heteroduplex with a short loop.
Biochemistry, 4th Edition
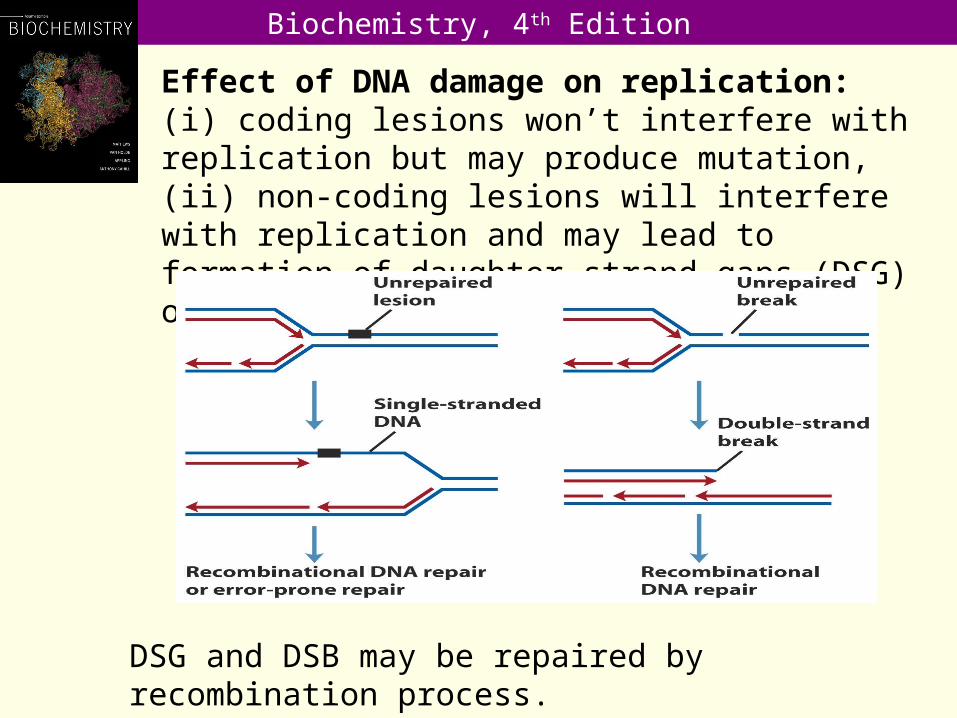
Effect of DNA damage on replication: (i) coding lesions won’t interfere with replication but may produce mutation, (ii) non-coding lesions will interfere with replication and may lead to formation of daughter-strand gaps (DSG) or double-strand breaks (DSB).
DSG and DSB may be repaired by recombination process.

Biochemistry, 4th Edition
Daughter-strand Gap Repair by recombination or translesion synthesis (SOS response)
Copyright © 2013 Pearson Canada Inc. 26 - 22
•The undamaged parental DNA strand is transferred to the gap in the daughter strand, formed by the inability of DNA polymerase to replicate past it.
•The remaining steps occur by mechanisms similar or identical to those in homologous recombination.
•SOS response may induce new DNA polymerase(s) which are capable of translesion synthesis.
Biochemistry, 4th Edition
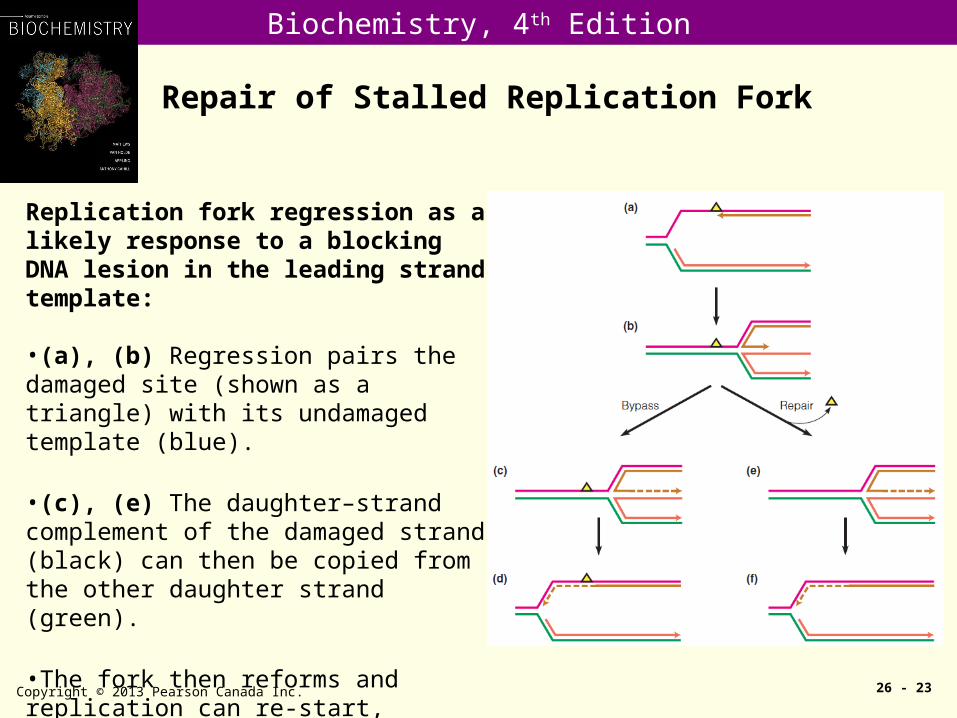
Repair of Stalled Replication Fork
Copyright © 2013 Pearson Canada Inc. 26 - 23
Replication fork regression as a likely response to a blocking DNA lesion in the leading strand template:
•(a), (b) Regression pairs the damaged site (shown as a triangle) with its undamaged template (blue).
•(c), (e) The daughter–strand complement of the damaged strand (black) can then be copied from the other daughter strand (green).
•The fork then reforms and replication can re-start, whether the damage has been repaired (f) or not (d).

Biochemistry, 4th Edition
Repair of DNA double-strand breaks by NHEJ
Copyright © 2013 Pearson Canada Inc. 26 - 24
• Double-strand DNA breaks can be repaired either by homologous recombination or by nonhomologous end joining (NHEJ), a process that does not require DNA sequence homology at the ends being joined.
A pathway for nonhomologous end joining:
• The figure shows involvement of some proteins in addition to those identified in the text, including DNA-dependent protein kinase (DNA-PK), the processing protein Artemis, and the X4-L4 complex, which includes DNA ligase IV.
Biochemistry, 4th Edition
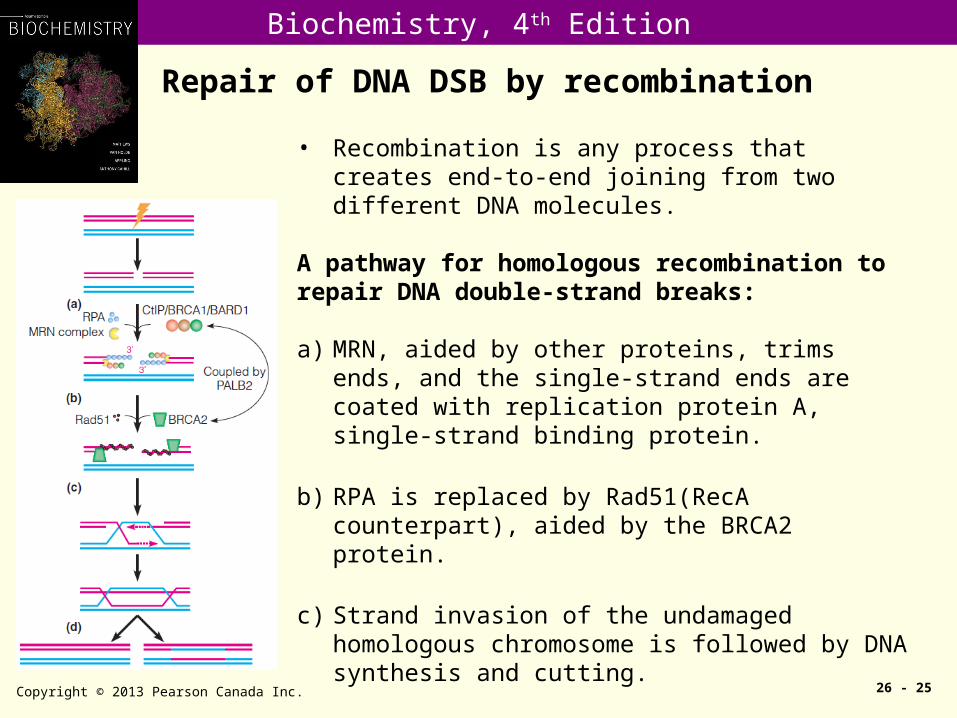
Repair of DNA DSB by recombination
Copyright © 2013 Pearson Canada Inc. 26 - 25
• Recombination is any process that creates end-to-end joining from two different DNA molecules.
A pathway for homologous recombination torepair DNA double-strand breaks:
a) MRN, aided by other proteins, trims ends, and the single-strand ends are coated with replication protein A, single-strand binding protein.
b) RPA is replaced by Rad51(RecA counterpart), aided by the BRCA2 protein.
c) Strand invasion of the undamaged homologous chromosome is followed by DNA synthesis and cutting.
d) Which can yield either recombinant (right) or nonrecombinant (left) chromosomes.

Biochemistry, 4th Edition
DNA repair and cancer
• Defects in the genes encoding the proteins involved in nucleotide-excision repair, mismatch repair, and recombination repair have all been linked to human cancer.
• Examples are: (i) xeroderma pigmentosum (or XP) patients with defects in nucleotide-excision repair, (ii) HNPCC (hereditary nonpoplyposis colon cancer) patients with defects in hMLH1 and hMSH2, and (3) breast cancer patients with inherited defects in BRCA1 and Brca2, which are known to interact with Rad 51 (the eukaryotic homolog of RecA) and therefore may have defective recombination repair.

Biochemistry, 4th Edition
DNA Recombination
• Segments of DNA can rearrange their location – within a chromosome
– from one chromosome to another
• Such recombination is involved in many biological processes– Repair of DNA
– Segregation of chromosomes during meiosis
– Enhancement of generic diversity
• In sexually reproducing organism, recombination and mutations are two driving forces of evolution
• Recombination of co-infecting viral genomes may enhance virulence and provide resistance to antivirals
Biochemistry, 4th Edition
DNA Recombination
• Homologous recombination or generalized recombination.
• Site-specific recombinataion.• Transposition.• Illegitimate recombination

Biochemistry, 4th Edition
Recombination
Copyright © 2013 Pearson Canada Inc. 26 - 29
Biochemistry, 4th Edition
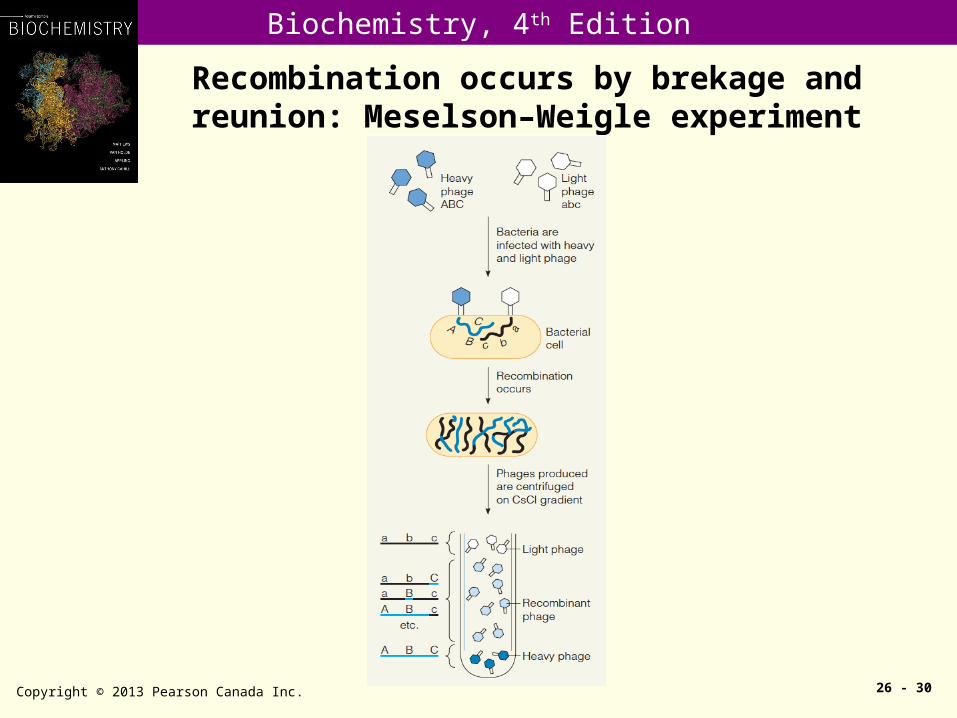
Recombination occurs by brekage and reunion: Meselson–Weigle experiment
Copyright © 2013 Pearson Canada Inc. 26 - 30

Biochemistry, 4th Edition
The Holliday model for homologous recombination
26 - 31
•A, a, Z, and z are genetic markers.•Heteroduplexes (one strand comes from one parent and the other from the second parent) are formed at sites of recombination.
Copyright © 2013 Pearson Canada Inc.

Biochemistry, 4th Edition
The Meselson–Radding model for homologous recombination
Copyright © 2013 Pearson Canada Inc. 26 - 32

Biochemistry, 4th Edition
Homologous recombination is catalyzed by enzymes
• The most well characterized recombination enzymes are derived from studies with E. coli cells.
• Presynapsis: helicase and/or nuclease to generate single-strand DNA with 3’-OH end (RecBCD).
• Synapsis: joint molecule formation to generate Holliday juncture (RecA).
• Postsynapsis: branch migration and resolution of Holliday juncture (RuvABC).

Biochemistry, 4th Edition
RecA forms nucleoprotein filament on single-strand DNA, promotes joint molecule formation
and strand exchange
Copyright © 2013 Pearson Canada Inc. 26 - 34
A model for RecA-mediated strand exchange:
•At the left, a RecA–ssDNA filament, with ssDNA shown in red.
•In the middle, a joint molecule, with triplestranded DNA; the original ssDNA is wrapped in the minor groove of the duplex DNA.
•At the right, strand exchange is occurring.
•The red ssDNA is complementary in sequence to the yellow strand of the duplex, and RecA action is displacing the green strand, coincident with formation of a new red–yellow dsDNA.

Biochemistry, 4th Edition
A model for RecA-mediated strand exchange
Copyright © 2013 Pearson Canada Inc. 26 - 35
•The black strand associated with RecA at the outset is complementary to the red strand in the duplex.
•Synapsis occurs to (rightward in the center diagram), with each RecA molecule displaced after its associated DNA has base-paired with its complement.
•In this model reaction the displaced blue strand is wrapped around the newly formed duplex.

Biochemistry, 4th Edition
Helicase and nuclease activities of the RecBCD
Copyright © 2013 Pearson Canada Inc. 26 - 36
A model for the action of RecBCD, () sites, and RecA in initiating homologous recombination:

Biochemistry, 4th Edition
RuvABC
• RuvA (22 kDa) binds a Holliday junction with high affinity, and together with RuvB (37 kDa) promotes ATP-dependent branch migration of the junctions leading to the formation of heteroduplex DNA.
• RuvC (19 kDa) resolves Holliday juncture into recombinant products.

Biochemistry, 4th Edition
A model for the RuvA-RuvB-Holliday junction structure
Copyright © 2013 Pearson Canada Inc. 26 - 38
•This is based upon crystal structures of RuvA and RuvB.
•Branch migration is believed to involve spooling of DNA to left and right through the RuvB twin pumps, with the upper and lower arms being drawn into the center and eventually out through the pumps.

Biochemistry, 4th Edition
Site-specific Recombination
Copyright © 2013 Pearson Canada Inc. 26 - 39
Integration of bacteriophage : •The phage chromosome circularizes between genes A and R, and recombination takes place between the attP site and a corresponding region, attB, on the E. coli chromosome between the gal and bio markers. •The enzyme integrase carries out the site-specific recombinational event with the help of a bacterial protein.
Biochemistry, 4th Edition
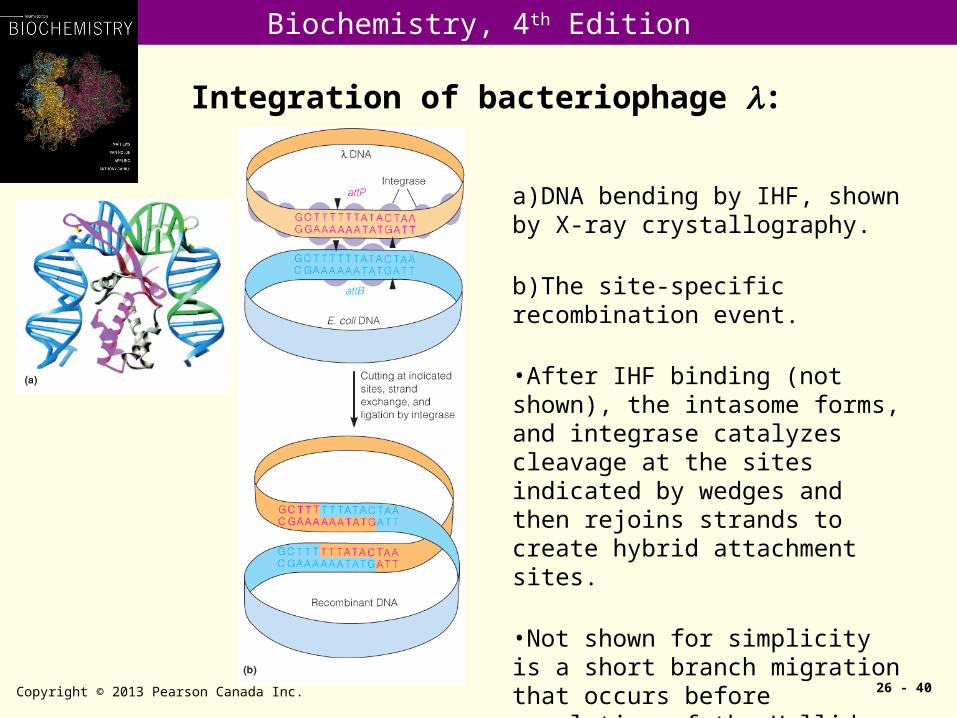
Integration of bacteriophage :
Copyright © 2013 Pearson Canada Inc. 26 - 40
a)DNA bending by IHF, shown by X-ray crystallography.
b)The site-specific recombination event.
•After IHF binding (not shown), the intasome forms, and integrase catalyzes cleavage at the sites indicated by wedges and then rejoins strands to create hybrid attachment sites.
•Not shown for simplicity is a short branch migration that occurs before resolution of the Holliday junction.

Biochemistry, 4th Edition
VDJ recombination of immunoglobin gene
Copyright © 2013 Pearson Canada Inc. 26 - 41
Gene rearrangements in antibody gene maturation:
•The rearrangements of C, V, and J sequences produce one mature light chain gene, and the transcription, processing, and translation of this gene produce an antibody light chain.
Biochemistry, 4th Edition
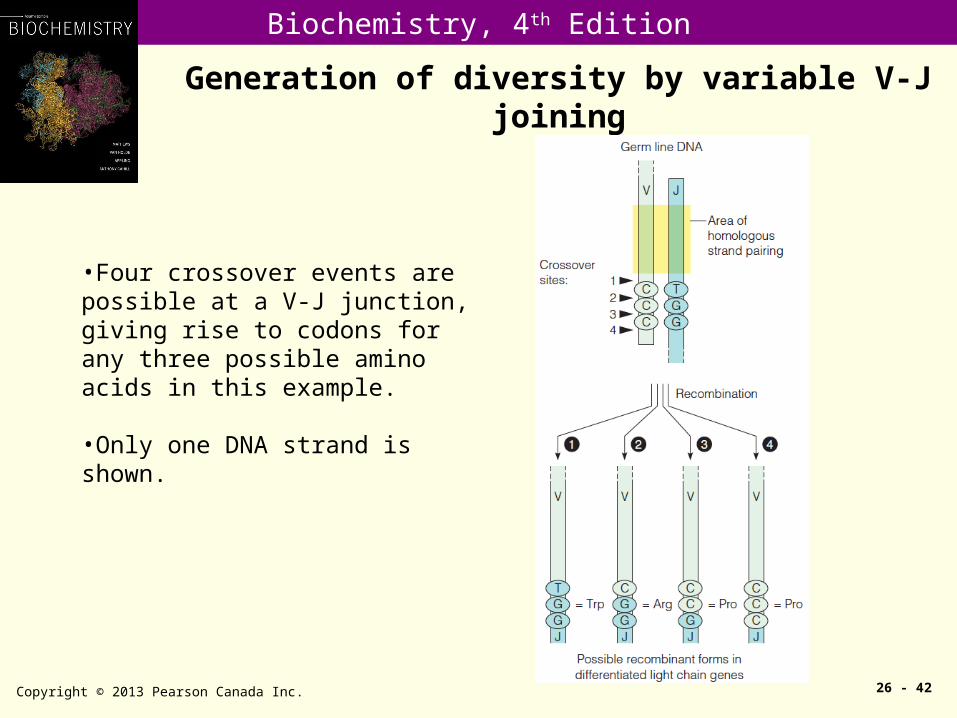
Generation of diversity by variable V-J joining
Copyright © 2013 Pearson Canada Inc. 26 - 42
•Four crossover events are possible at a V-J junction, giving rise to codons for any three possible amino acids in this example.
•Only one DNA strand is shown.
Biochemistry, 4th Edition
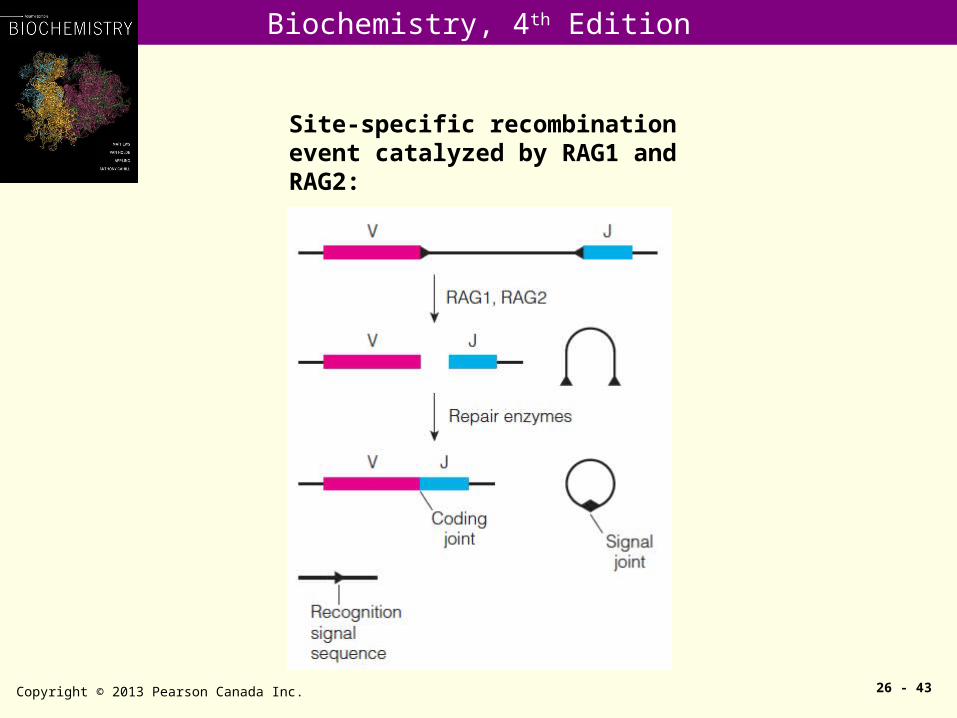
Copyright © 2013 Pearson Canada Inc. 26 - 43
Site-specific recombination event catalyzed by RAG1 and RAG2:

Biochemistry, 4th Edition
Recognition signal sequences
Copyright © 2013 Pearson Canada Inc. 26 - 44
• Related DNA sequences are found to the side of each V sequence and to the side of each J sequence, and they represent recognition sites for the enzymes involved in the joining reaction.
• Those sequences, which are called recognition signal sequences, are as follows:
• Note that the homologous regions of these sequences are inverted repeats.

Biochemistry, 4th Edition
Effects of recombination on DNA structure
Copyright © 2013 Pearson Canada Inc. 26 - 45
•Depending on the orientation of the two copies, either
a) inversion or
b) deletion can result.

Biochemistry, 4th Edition
Transposons
Copyright © 2013 Pearson Canada Inc. 26 - 46
Structures of class I, class II, and class III mobile genetic elements: •Inverted repeats are shown in purple.•The “drug resistance element” may be one of a variety of protein-coding genes.

Biochemistry, 4th Edition
Copyright © 2013 Pearson Canada Inc. 26 - 47

Biochemistry, 4th Edition
Transposition
Copyright © 2013 Pearson Canada Inc. 26 - 48
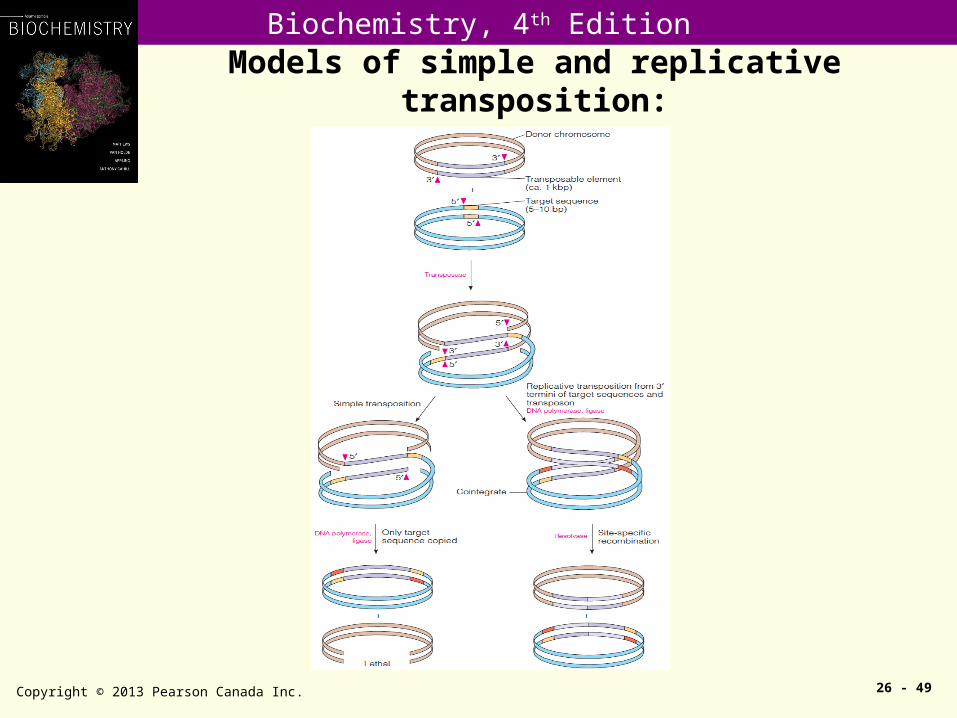
Transposition is characterized by duplication of direct repeats (5-9 bps in most cases) at target site.
Biochemistry, 4th Edition
Models of simple and replicative transposition:
Copyright © 2013 Pearson Canada Inc. 26 - 49

Biochemistry, 4th Edition
Retrotransposon: Transposition mediated through a RNA intermediate.
Copyright © 2013 Pearson Canada Inc. 26 - 50
Structure of retroviral genomes in the integrated state:
a)A nononcogenic virus. b)An oncogenic virus such as Rous sarcoma virus, showing the viral oncogene downstream (rightward) from the viral replication genes. c)A defective oncogenic virus, such as Moloney murine sarcoma virus, with the viral oncogene replacing part or all of a gene (env) essential to viral replication. •In each case the LTRs are direct repeats, flanked by short inverted repeats.
Biochemistry, 4th Edition
Copyright © 2013 Pearson Canada Inc. 26 - 51
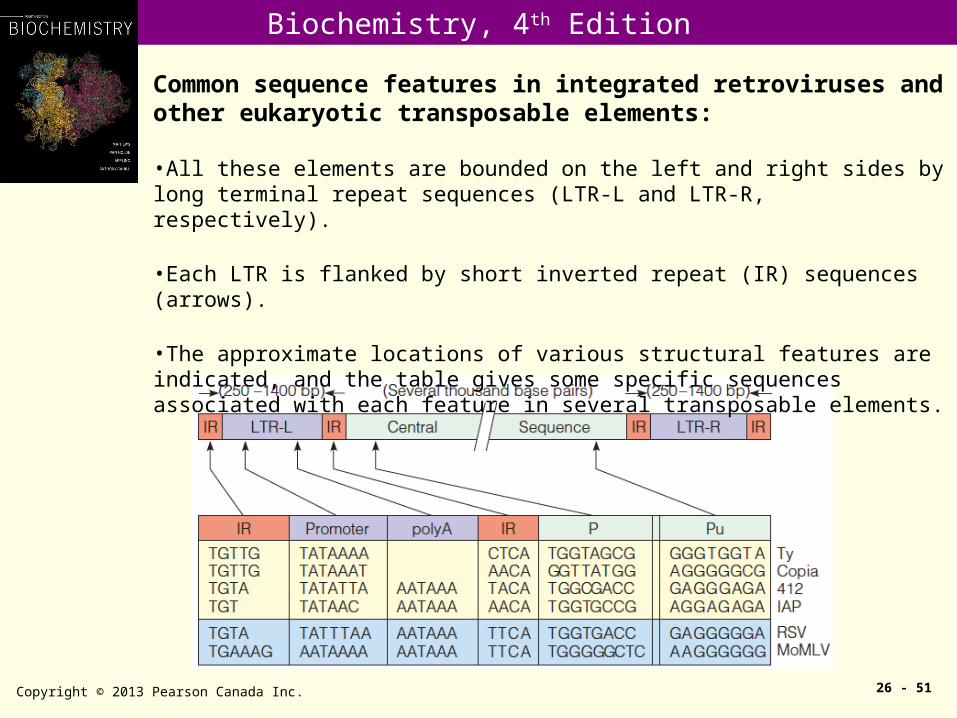
Common sequence features in integrated retroviruses and other eukaryotic transposable elements:
•All these elements are bounded on the left and right sides by long terminal repeat sequences (LTR-L and LTR-R, respectively).
•Each LTR is flanked by short inverted repeat (IR) sequences (arrows).
•The approximate locations of various structural features are indicated, and the table gives some specific sequences associated with each feature in several transposable elements.
Biochemistry, 4th Edition
Gene Amplification
Copyright © 2013 Pearson Canada Inc. 26 - 52
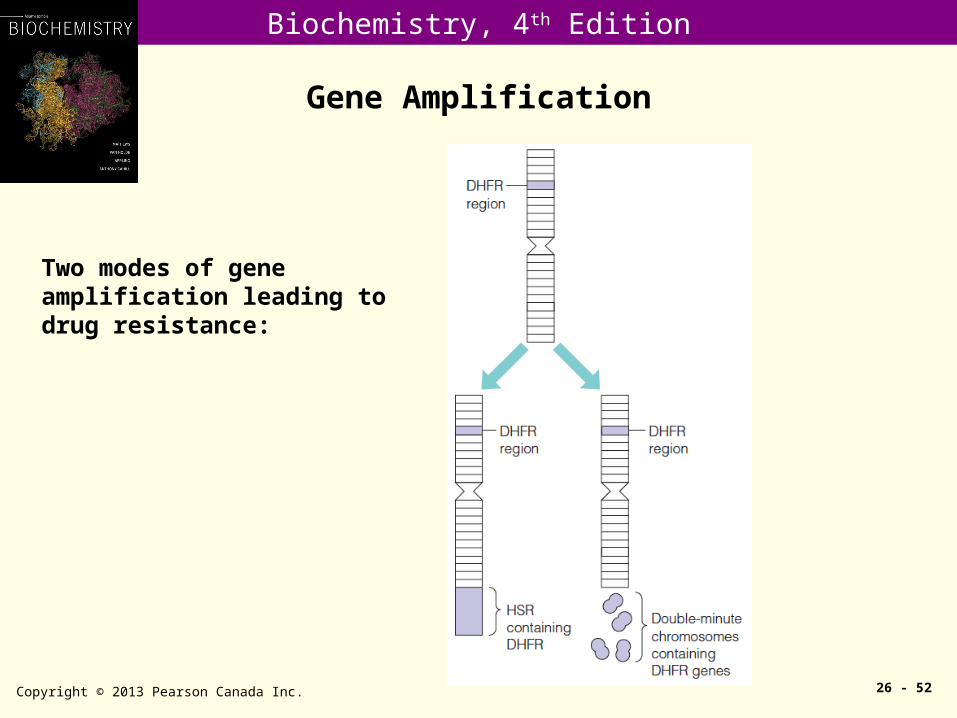
Two modes of gene amplification leading to drug resistance:

Biochemistry, 4th EditionChromosome structural changes that accompany
dihydrofolate reductase gene amplification
Copyright © 2013 Pearson Canada Inc. 26 - 53
•The micrograph shows metaphase chromosomes from Chinese hamster ovary cells that are highly resistant to methotrexate.
•Chromosomal DNA was subjected to hybridization in situ with a fluorescence-labeled DHFR gene probe.
•White arrows point to single-copy genes.
•The amplified chromosomal sequences are on a giant form of the chromosome that also contains one of the original single-copy sequences.
Related Documents




