ISSN: 1524-4539 Copyright © 2008 American Heart Association. All rights reserved. Print ISSN: 0009-7322. Online 72514 Circulation is published by the American Heart Association. 7272 Greenville Avenue, Dallas, TX DOI: 10.1161/CIRCULATIONAHA.107.745539 2008;117;1405-1413; originally published online Mar 3, 2008; Circulation Magnuson, Colin G. Nichols, William A. Coetzee and David J. Lefer John W. Elrod, Maddison Harrell, Thomas P. Flagg, Susheel Gundewar, Mark A. Channels in Myocardial Ischemia/Reperfusion Injury Role of Sulfonylurea Receptor Type 1 Subunits of ATP-Sensitive Potassium http://circ.ahajournals.org/cgi/content/full/CIRCULATIONAHA.107.745539/DC1 Data Supplement (unedited) at: http://circ.ahajournals.org/cgi/content/full/circulationaha;117/18/e332 An erratum has been published regarding this article. Please see the attached page or: http://circ.ahajournals.org/cgi/content/full/117/11/1405 located on the World Wide Web at: The online version of this article, along with updated information and services, is http://www.lww.com/reprints Reprints: Information about reprints can be found online at [email protected] 410-528-8550. E-mail: Fax: Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters http://circ.ahajournals.org/subscriptions/ Subscriptions: Information about subscribing to Circulation is online at at UCLA BIOMED LIBRARY SERIALS on February 23, 2010 circ.ahajournals.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ISSN: 1524-4539 Copyright © 2008 American Heart Association. All rights reserved. Print ISSN: 0009-7322. Online
72514Circulation is published by the American Heart Association. 7272 Greenville Avenue, Dallas, TX
DOI: 10.1161/CIRCULATIONAHA.107.745539 2008;117;1405-1413; originally published online Mar 3, 2008; Circulation
Magnuson, Colin G. Nichols, William A. Coetzee and David J. Lefer John W. Elrod, Maddison Harrell, Thomas P. Flagg, Susheel Gundewar, Mark A.
Channels in Myocardial Ischemia/Reperfusion InjuryRole of Sulfonylurea Receptor Type 1 Subunits of ATP-Sensitive Potassium
http://circ.ahajournals.org/cgi/content/full/CIRCULATIONAHA.107.745539/DC1Data Supplement (unedited) at:
http://circ.ahajournals.org/cgi/content/full/circulationaha;117/18/e332
An erratum has been published regarding this article. Please see the attached page or:
http://circ.ahajournals.org/cgi/content/full/117/11/1405located on the World Wide Web at:
The online version of this article, along with updated information and services, is
http://www.lww.com/reprintsReprints: Information about reprints can be found online at
[email protected]. E-mail:
Fax:Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters
http://circ.ahajournals.org/subscriptions/Subscriptions: Information about subscribing to Circulation is online at
at UCLA BIOMED LIBRARY SERIALS on February 23, 2010 circ.ahajournals.orgDownloaded from

Role of Sulfonylurea Receptor Type 1 Subunits ofATP-Sensitive Potassium Channels in Myocardial
Ischemia/Reperfusion InjuryJohn W. Elrod, PhD; Maddison Harrell, BS; Thomas P. Flagg, PhD; Susheel Gundewar, MD;
Mark A. Magnuson, MD; Colin G. Nichols, PhD; William A. Coetzee, PhD; David J. Lefer, PhD
Background—Opening of cardiac ATP-sensitive potassium channels (KATP channels) is a well-characterized protectivemechanism against ischemia and reperfusion injury. Evidence exists for an involvement of both sarcolemmal andmitochondrial KATP channels in such protection. Classically, cardiac sarcolemmal KATP channels are thought to becomposed of Kir6.2 (inward-rectifier potassium channel 6.2) and SUR2A (sulfonylurea receptor type 2A) subunits;however, the evidence is strong that SUR1 (sulfonylurea receptor type 1) subunits are also expressed in the heart andthat they may have a functional role. The aim of this study, therefore, was to examine the role of SUR1 in myocardialinfarction.
Methods and Results—We subjected mice lacking SUR1 subunits to in vivo myocardial ischemia/reperfusion injury.Interestingly, the SUR1-null mice were markedly protected against the ischemic insult, displaying a reduced infarct sizeand preservation of left ventricular function, which suggests a role for this KATP channel subunit in cardiovascularfunction during conditions of stress.
Conclusions—SUR1 subunits have a high sensitivity toward many sulfonylureas and certain KATP channel–opening drugs.Their potential role during ischemic events should therefore be considered both in the interpretation of experimental datawith pharmacological agents and in the clinical arena when the cardiovascular outcome of patients treated withantidiabetic sulfonylureas is being considered. (Circulation. 2008;117:1405-1413.)
Key Words: KATP channels � ischemia � ion channels � infarction � myocardial infarction
Even in the initial description of the ATP-sensitive potas-sium (KATP) channel in the heart,1 it was postulated that
the opening of these channels during hypoxic or ischemicconditions might be cardioprotective. In general, this hasproved to be the case, and a multitude of in vivo and in vitrostudies have demonstrated that KATP channel opening gener-ally protects against ischemic insults.2 KATP channel openinghas also been demonstrated to play a key role in the eventsthat lead to the protection afforded by ischemic precondition-ing. A role for KATP channels present both at the sarcolemmaand in mitochondria has been reported in the phenomenon ofischemic preconditioning.3
Clinical Perspective p 1413
Many of the arguments about the roles of KATP channels inischemia are derived from experiments using putativelyspecific pharmacological compounds, the action of which
may depend on the metabolic state of the cell.4 The nonspeci-ficity of these agents is a cause for concern in the interpre-tation of experimental results. The molecular characterizationof subunits of KATP channels (inward-rectifier potassiumchannels 6.1 and 6.2 [Kir6.1 and Kir6.2] and sulfonylureareceptor [SUR] type 1 [SUR1] and type 2 [SUR2]) allowedgenetic approaches to be used to assess the roles of KATP
channels in a variety of physiological and pathophysiologicalprocesses.5 Mouse models have been created that are genet-ically deficient for each of these respective subunits.5 Thecardiac KATP channel has been proposed to be composed ofKir6.2 subunits in combination with sulfonylurea receptortype 2A (SUR2A) subunits5,6; indeed, mouse hearts deficientin Kir6.2 subunits lack functional cardiac KATP channelactivity.7 These mice also lack the protective effect ofischemic preconditioning,8 which is consistent with a protec-tive role for cardiac KATP channels during ischemia. Mice
Received October 11, 2007; accepted January 22, 2008.From the Department of Medicine and Pathology (J.W.E., S.G., D.J.L.), Albert Einstein College of Medicine, New York, NY; Department of Pediatrics
(M.H., W.A.C.), NYU School of Medicine, New York, NY; Department of Cell Biology and Physiology (T.P.F., C.G.N.), Washington University Schoolof Medicine, St Louis, Mo; and Department of Molecular Physiology and Biophysics (M.A.M.), Vanderbilt University School of Medicine, Nashville,Tenn.
The online-only Data Supplement, consisting of a table and a figure, is available with this article at http://circ.ahajournals.org/cgi/content/full/CIRCULATIONAHA.107.745539/DC1.
Correspondence to Dr William A. Coetzee, Pediatric Cardiology, NYU School of Medicine, 560 First Ave, TCH-521, New York, NY 10016. [email protected]
© 2008 American Heart Association, Inc.
Circulation is available at http://circ.ahajournals.org DOI: 10.1161/CIRCULATIONAHA.107.745539
1405
Molecular Cardiology
at UCLA BIOMED LIBRARY SERIALS on February 23, 2010 circ.ahajournals.orgDownloaded from

deficient in SUR2 subunits have an elevated blood pressureand elevated coronary perfusion pressure and exhibit sponta-neous coronary vasospasm,9 which is consistent with expres-sion of SUR type 2B (SUR2B) subunits in the vasculature5
and the known functional role of KATP channels in themaintenance of vascular tone and blood flow.10 Although thevascular phenotype may complicate interpretations, it hasrecently been reported that mouse hearts deficient in SUR2subunits are protected against global ischemia.11 This surpris-ing finding is not predicted from the simple view of SUR2-containing KATP channels being involved in ischemic protec-tion, because loss of the KATP channel–mediated protectionwould be predicted to exacerbate ischemia.
Several reports discuss the expression of (and a possiblerole for) SUR1 subunits in the heart.12–15 To assess thepossible contribution of SUR1 subunits in protection duringmyocardial ischemia, we used a genetic mouse model thatlacks SUR1 subunits16 and subjected the mice to an in vivoischemia/reperfusion protocol. The data indicate a novel andunexpected role for SUR1 subunits in the heart.
MethodsGeneration of Genetic Mouse ModelsMice deficient in the ABCC8 gene (SUR1-null mice) have beendescribed previously.16 All animals received humane care in com-pliance with the “Principles of Laboratory Animal Care” formulatedby the National Society of Medical Research and the “Guide for theCare and Use of Laboratory Animals” published by the NationalInstitutes of Health (NIH publication No. 85-23, revised 1996). Theexperimental protocol was reviewed and approved by the institu-tional animal care and use committees of Albert Einstein College ofMedicine and NYU School of Medicine.
Myocardial Ischemia and Reperfusion ProtocolAll surgical procedures were performed with aseptic techniques asdescribed in detail previously.17 Briefly, mice were anesthetized viaan intraperitoneal injection of sodium pentobarbital (50 mg/kg) andketamine (60 mg/kg). Mice were intubated and ventilated with 100%oxygen by a rodent ventilator (Minivent, model 845, Hugo SachsElektronik, March-Hugstetten, Germany) at a rate of 110 strokes/minand 230-�L tidal volume. A thoracotomy was performed, and the leftcoronary artery was visually identified and ligated with a 7-0 silksuture to render the left ventricle (LV) ischemic.
Assessment of Myocardial Infarct SizeAfter 24 hours of reperfusion, infarct size was determined asdescribed previously.17 Briefly, Evan’s blue dye was injected into thecarotid artery catheter to delineate the ischemic zone from thenonischemic zone. The heart was rapidly excised and cross-sectionedinto 1-mm-thick slices, which were then incubated in 1.0% 2,3,5-triphenyl-tetrazolium chloride solution (TTC) to demarcate theviable and nonviable myocardium within the risk zone. Images ofeach side of each section were acquired and weighed. The areas ofinfarction, risk, and nonischemic LV were assessed in a blindedfashion with computer-assisted planimetry (NIH Image 1.57, Na-tional Institutes of Health, Bethesda, Md).
Echocardiographic Assessment of LV Structureand FunctionMice were lightly anesthetized with isoflurane in 100% O2 withcontinuous monitoring of ECG, core body temperature, and respira-tion rate. Transthoracic high-resolution echocardiography of the LVwith a 30-MHz RMV (real-time microvisualization) scanhead(probe) interfaced with a Vevo 770 (VisualSonics, Toronto, Canada)was performed 1 week before myocardial infarction. Parasternal
long- and short-axis B-mode and M-mode images were acquired.Offline measurements and calculations were made with VisualSonicssoftware (version 2.2.3) for assessment of LV structure and function.Seven days after myocardial infarction, mice again underwentechocardiography just before the evaluation of infarct size.
Histological Analysis of Myocardial Infarct SizeHearts were excised rapidly and fixed in conventional fixing solu-tions (10% buffered formalin) after 30 minutes of left coronary arteryischemia and 7 days reperfusion. Hearts were cross-sectioned into1-mm-thick slices with a tissue chopper. Hearts were embedded in astandard fashion and stained with hematoxylin and eosin. Digitalimages of the slides were then captured and analyzed in a blindedfashion with computer-assisted planimetry with Image J software(version 1.37, National Institutes of Health) to measure the area ofinfarct or scar relative to the LV. For each heart, we analyzed 4sections taken from each 1-mm–thick slice and then averaged thosenumbers to obtain the size of infarct or scar per LV for each animal.
Isolation of Ventricular Cardiac MyocytesLV myocytes were isolated from adult mice (aged 4 to 6 months;weight 25 g to 35 g) by a standard enzymatic technique describedpreviously18 with some minor modifications. Briefly, mice wereanesthetized by intraperitoneal injection (0.01 mL/g) that containedketamine (1.5 mg/mL) and xylazine (15 mg/mL) diluted in PBS. Theheart was excised, cannulated within 2 minutes, and perfusedretrogradely at 3 mL/min with PBS containing (in mmol/L) NaCl113, KCl 4.7, KH2PO4 0.6, Na2HPO4 0.6, MgSO4.7H20 1.2, phenolred 0.032, NaHCO3 12, KHCO3 10, HEPES 10, taurine 30, 2,3-butanedione monoxine (BDM) 10, and glucose 10 (pH 7.2 at 37°C).After a 5-minute wash, saline containing 20 mg of collagenase (356U/mg; type II, Worthington Biochemicals, Lakewood, NJ), 3 mg ofprotease (5.2 U/mg; type XIV, Sigma-Aldrich, St Louis, Mo), andtrypsin (0.14 mg/mL; 10X solution; GIBCO/Invitrogen, Carlsbad,Calif) was perfused. After 13 to 15 minutes of perfusion, the heartwas removed from the cannula and gently minced in enzyme-free,Ca2�-free perfusion buffer containing BSA (1 mg/mL) and BDM(0.2 mg/mL). The cell suspension was filtered though 250-�m nylonmesh (Small Parts, Inc, Miami, Fla), and myocytes were separatedinto pellets by gravity and subsequently washed with perfusionbuffer without BDM and BSA. The myocytes were plated in 50-mmnonstick Valmark Petri dishes (Midwest Scientific, St Louis, Mo)and subjected to stepwise Ca2� reintroduction. Myocytes were storedat room temperature (20°C to 22°C) in perfusion buffer containing1 mmol/L Ca2� and used within 8 hours after isolation.
Patch-Clamp RecordingsRecordings of single-KATP channel activity were made by standardpatch-clamp techniques.19 Patch electrodes were made from thick-walled glass capillaries (outside diameter 1.5 mm, inside diameter1.12 mm; World Precision Instruments Inc, Sarasota, Fla) with ahorizontal puller (Zeitz Instrumente Universal puller, Augsburg,Germany) and heat-polished. The pipette solution (extracellularmedium) contained (in mmol/L) gluconate 110, KCl 30, EGTA 1,MgCl2 1.2, and HEPES 10 (pH 7.2 with KOH). The bath solution(intracellular solution) contained (in mmol/L) gluconate 110, KCl30, CaCl2 2, MgCl2 1, and HEPES 10 (pH 7.4 with KOH). Whenfilled, electrode resistances ranged between 4 and 9 M�. Recordingswere made with a patch-clamp amplifier (Axopatch 200, AxonInstruments, Foster City, Calif). Solution changes were made withinwith a rapid solution changer (�100 ms; RSC-160, Biological,Cliax, France). The current signals were amplified (20 to 200mV/pA), low-pass filtered at 1 kHz, and digitized at 5 kHz withpClamp software (Axon Instruments). Unless otherwise stated, thepipette potential was kept at 100 mV. Data were analyzed withpClamp (Axon Instruments) or Origin for Windows (MicroCalSoftware, Northampton, Mass) software. The unitary current ampli-tude was determined from an amplitude histogram of 15 to 20seconds of recorded data. The histogram was fitted to a sum ofgaussian distributions. The difference between 2 adjacent gaussian
1406 Circulation March 18, 2008
at UCLA BIOMED LIBRARY SERIALS on February 23, 2010 circ.ahajournals.orgDownloaded from
peaks was taken as a measure of the unitary current amplitude.Because most recordings contained many more than a single KATP
channel, no attempts were made to study the distribution of channeldwell times. Where possible, results are expressed as mean�SEM.Results were repeated in a minimum of 3 patches to ensurereproducibility.
Reverse-Transcription Polymerase Chain ReactionExpression AssaysReal-time reverse-transcription polymerase chain reaction (RT-PCR)was performed essentially as described previously.20 Total RNA wasextracted from ventricular tissue by the acid phenol guanidiniummethod (TriReagent, Sigma). RNA quality was determined byagarose electrophoresis (intact 18S and 28S RNA bands), and theconcentration was determined spectrophotometrically. Total RNAwas reverse transcribed with random hexamer primers according tothe manufacturer’s guidelines (Superscript III; Invitrogen). PCRreactions were performed with an ABI Prism 7900HT sequencedetection system (PerkinElmer Applied Biosystems, Foster City,Calif) with a SYBR green master mix (PerkinElmer Applied Bio-systems). The thermal cycling conditions comprised an initial dena-turing step at 95°C for 10 minutes and 40 cycles at 95°C for 5seconds, 60°C for 15 seconds, and 72°C for 15 seconds, followed bya melting curve analysis.20 The primers used in the PCR reactions areshown in the Table. Because of the nature of the primer design usedfor real-time RT-PCR (amplicons to be within 80 to 120 bp), it wasnot possible to specifically amplify SUR2B. Conventional RT-PCRexperiments were therefore performed to determine the relativeexpression of SUR2A and SUR2B (30 cycles with an annealingtemperature of 55°C) with primers that produce different ampliconsizes for SUR2A and SUR2B21 (Table).
Statistical AnalysisAll data in the present study are expressed as mean�SEM. Differ-ences in data between groups were compared with Prism 4 software(GraphPad Software, Inc, San Diego, Calif) with Student t test or1-way ANOVA where appropriate. For the ANOVA, if a significantvariance was found, the Tukey test was used as the post hoc analysis.P�0.05 was considered significant.
The authors had full access to and take full responsibility for theintegrity of the data. All authors have read and agree to themanuscript as written.
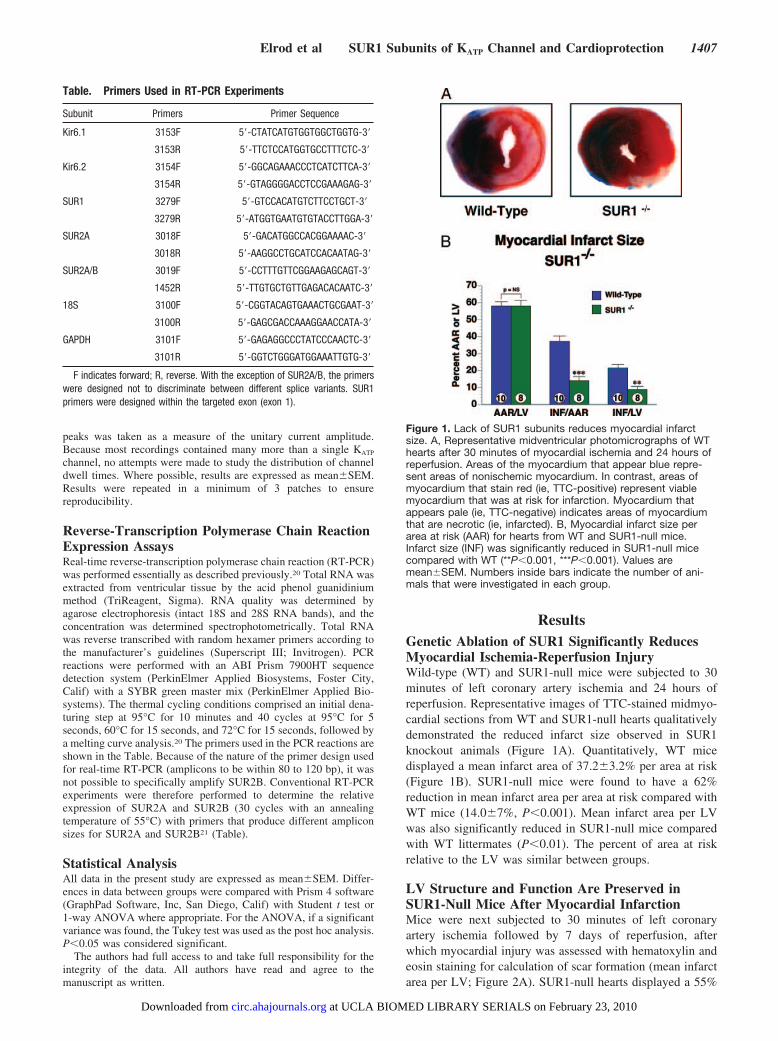
ResultsGenetic Ablation of SUR1 Significantly ReducesMyocardial Ischemia-Reperfusion InjuryWild-type (WT) and SUR1-null mice were subjected to 30minutes of left coronary artery ischemia and 24 hours ofreperfusion. Representative images of TTC-stained midmyo-cardial sections from WT and SUR1-null hearts qualitativelydemonstrated the reduced infarct size observed in SUR1knockout animals (Figure 1A). Quantitatively, WT micedisplayed a mean infarct area of 37.2�3.2% per area at risk(Figure 1B). SUR1-null mice were found to have a 62%reduction in mean infarct area per area at risk compared withWT mice (14.0�7%, P�0.001). Mean infarct area per LVwas also significantly reduced in SUR1-null mice comparedwith WT littermates (P�0.01). The percent of area at riskrelative to the LV was similar between groups.
LV Structure and Function Are Preserved inSUR1-Null Mice After Myocardial InfarctionMice were next subjected to 30 minutes of left coronaryartery ischemia followed by 7 days of reperfusion, afterwhich myocardial injury was assessed with hematoxylin andeosin staining for calculation of scar formation (mean infarctarea per LV; Figure 2A). SUR1-null hearts displayed a 55%
Table. Primers Used in RT-PCR Experiments
Subunit Primers Primer Sequence
Kir6.1 3153F 5�-CTATCATGTGGTGGCTGGTG-3�
3153R 5�-TTCTCCATGGTGCCTTTCTC-3�
Kir6.2 3154F 5�-GGCAGAAACCCTCATCTTCA-3�
3154R 5�-GTAGGGGACCTCCGAAAGAG-3�
SUR1 3279F 5�-GTCCACATGTCTTCCTGCT-3�
3279R 5�-ATGGTGAATGTGTACCTTGGA-3�
SUR2A 3018F 5�-GACATGGCCACGGAAAAC-3�
3018R 5�-AAGGCCTGCATCCACAATAG-3�
SUR2A/B 3019F 5�-CCTTTGTTCGGAAGAGCAGT-3�
1452R 5�-TTGTGCTGTTGAGACACAATC-3�
18S 3100F 5�-CGGTACAGTGAAACTGCGAAT-3�
3100R 5�-GAGCGACCAAAGGAACCATA-3�
GAPDH 3101F 5�-GAGAGGCCCTATCCCAACTC-3�
3101R 5�-GGTCTGGGATGGAAATTGTG-3�
F indicates forward; R, reverse. With the exception of SUR2A/B, the primerswere designed not to discriminate between different splice variants. SUR1primers were designed within the targeted exon (exon 1).
Figure 1. Lack of SUR1 subunits reduces myocardial infarctsize. A, Representative midventricular photomicrographs of WThearts after 30 minutes of myocardial ischemia and 24 hours ofreperfusion. Areas of the myocardium that appear blue repre-sent areas of nonischemic myocardium. In contrast, areas ofmyocardium that stain red (ie, TTC-positive) represent viablemyocardium that was at risk for infarction. Myocardium thatappears pale (ie, TTC-negative) indicates areas of myocardiumthat are necrotic (ie, infarcted). B, Myocardial infarct size perarea at risk (AAR) for hearts from WT and SUR1-null mice.Infarct size (INF) was significantly reduced in SUR1-null micecompared with WT (**P�0.001, ***P�0.001). Values aremean�SEM. Numbers inside bars indicate the number of ani-mals that were investigated in each group.
Elrod et al SUR1 Subunits of KATP Channel and Cardioprotection 1407
at UCLA BIOMED LIBRARY SERIALS on February 23, 2010 circ.ahajournals.orgDownloaded from
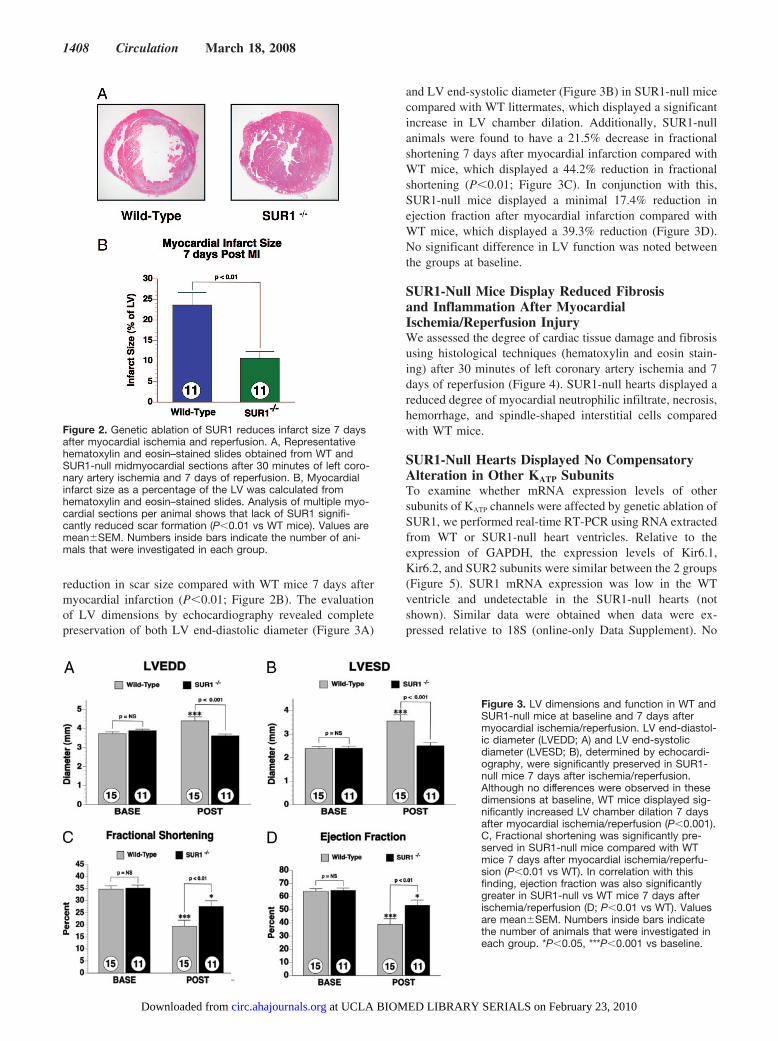
reduction in scar size compared with WT mice 7 days aftermyocardial infarction (P�0.01; Figure 2B). The evaluationof LV dimensions by echocardiography revealed completepreservation of both LV end-diastolic diameter (Figure 3A)
and LV end-systolic diameter (Figure 3B) in SUR1-null micecompared with WT littermates, which displayed a significantincrease in LV chamber dilation. Additionally, SUR1-nullanimals were found to have a 21.5% decrease in fractionalshortening 7 days after myocardial infarction compared withWT mice, which displayed a 44.2% reduction in fractionalshortening (P�0.01; Figure 3C). In conjunction with this,SUR1-null mice displayed a minimal 17.4% reduction inejection fraction after myocardial infarction compared withWT mice, which displayed a 39.3% reduction (Figure 3D).No significant difference in LV function was noted betweenthe groups at baseline.
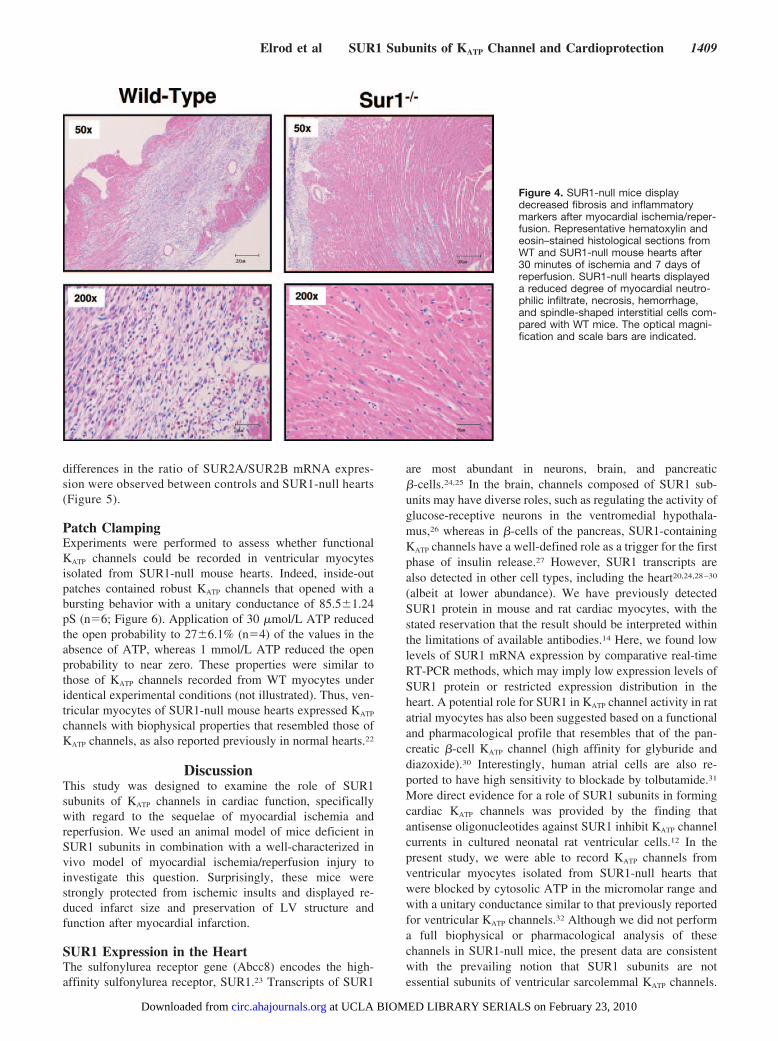
SUR1-Null Mice Display Reduced Fibrosisand Inflammation After MyocardialIschemia/Reperfusion InjuryWe assessed the degree of cardiac tissue damage and fibrosisusing histological techniques (hematoxylin and eosin stain-ing) after 30 minutes of left coronary artery ischemia and 7days of reperfusion (Figure 4). SUR1-null hearts displayed areduced degree of myocardial neutrophilic infiltrate, necrosis,hemorrhage, and spindle-shaped interstitial cells comparedwith WT mice.
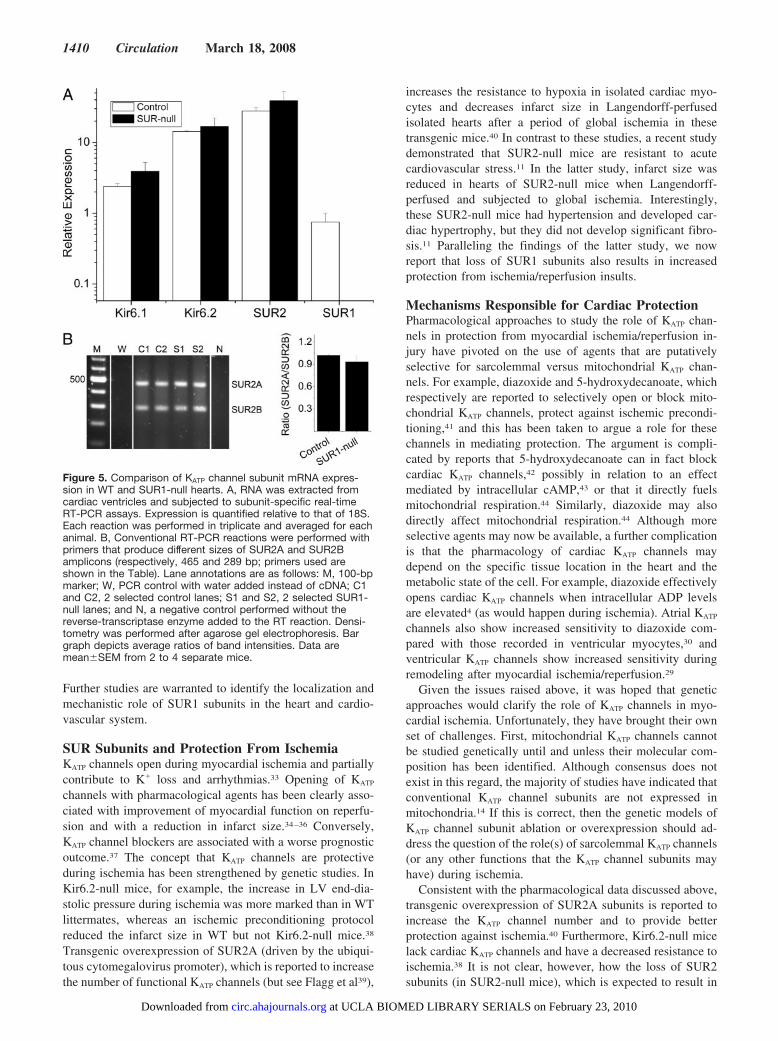
SUR1-Null Hearts Displayed No CompensatoryAlteration in Other KATP SubunitsTo examine whether mRNA expression levels of othersubunits of KATP channels were affected by genetic ablation ofSUR1, we performed real-time RT-PCR using RNA extractedfrom WT or SUR1-null heart ventricles. Relative to theexpression of GAPDH, the expression levels of Kir6.1,Kir6.2, and SUR2 subunits were similar between the 2 groups(Figure 5). SUR1 mRNA expression was low in the WTventricle and undetectable in the SUR1-null hearts (notshown). Similar data were obtained when data were ex-pressed relative to 18S (online-only Data Supplement). No
Figure 2. Genetic ablation of SUR1 reduces infarct size 7 daysafter myocardial ischemia and reperfusion. A, Representativehematoxylin and eosin–stained slides obtained from WT andSUR1-null midmyocardial sections after 30 minutes of left coro-nary artery ischemia and 7 days of reperfusion. B, Myocardialinfarct size as a percentage of the LV was calculated fromhematoxylin and eosin–stained slides. Analysis of multiple myo-cardial sections per animal shows that lack of SUR1 signifi-cantly reduced scar formation (P�0.01 vs WT mice). Values aremean�SEM. Numbers inside bars indicate the number of ani-mals that were investigated in each group.
Figure 3. LV dimensions and function in WT andSUR1-null mice at baseline and 7 days aftermyocardial ischemia/reperfusion. LV end-diastol-ic diameter (LVEDD; A) and LV end-systolicdiameter (LVESD; B), determined by echocardi-ography, were significantly preserved in SUR1-null mice 7 days after ischemia/reperfusion.Although no differences were observed in thesedimensions at baseline, WT mice displayed sig-nificantly increased LV chamber dilation 7 daysafter myocardial ischemia/reperfusion (P�0.001).C, Fractional shortening was significantly pre-served in SUR1-null mice compared with WTmice 7 days after myocardial ischemia/reperfu-sion (P�0.01 vs WT). In correlation with thisfinding, ejection fraction was also significantlygreater in SUR1-null vs WT mice 7 days afterischemia/reperfusion (D; P�0.01 vs WT). Valuesare mean�SEM. Numbers inside bars indicatethe number of animals that were investigated ineach group. *P�0.05, ***P�0.001 vs baseline.
1408 Circulation March 18, 2008
at UCLA BIOMED LIBRARY SERIALS on February 23, 2010 circ.ahajournals.orgDownloaded from
differences in the ratio of SUR2A/SUR2B mRNA expres-sion were observed between controls and SUR1-null hearts(Figure 5).
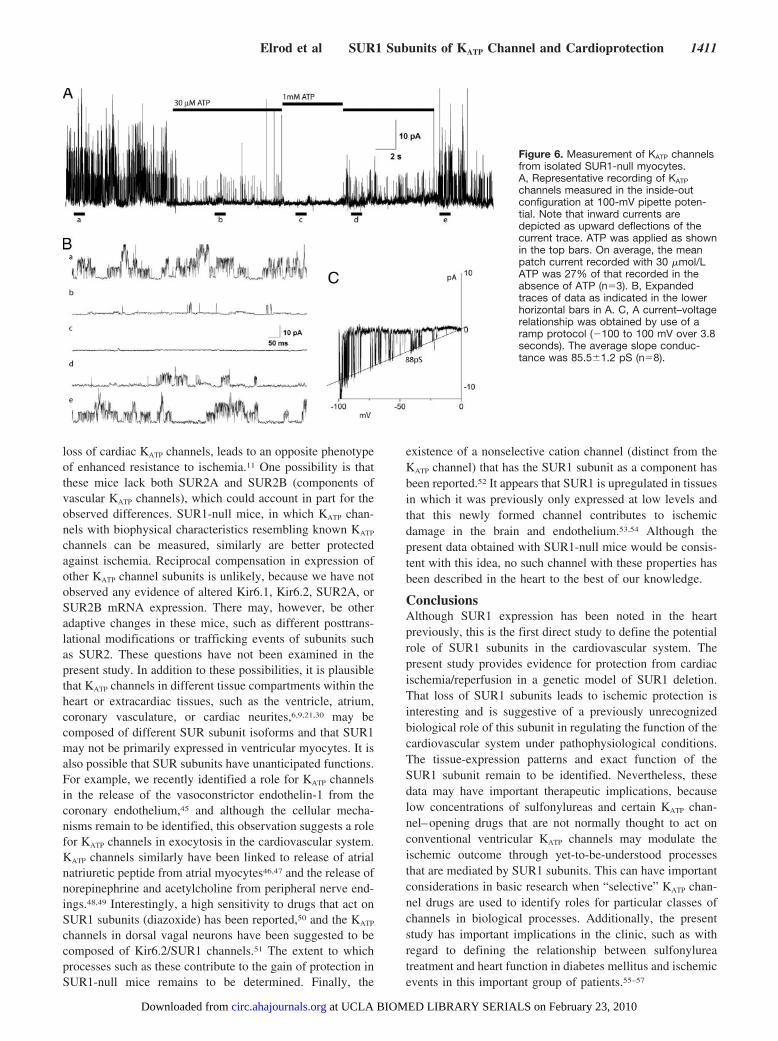
Patch ClampingExperiments were performed to assess whether functionalKATP channels could be recorded in ventricular myocytesisolated from SUR1-null mouse hearts. Indeed, inside-outpatches contained robust KATP channels that opened with abursting behavior with a unitary conductance of 85.5�1.24pS (n�6; Figure 6). Application of 30 �mol/L ATP reducedthe open probability to 27�6.1% (n�4) of the values in theabsence of ATP, whereas 1 mmol/L ATP reduced the openprobability to near zero. These properties were similar tothose of KATP channels recorded from WT myocytes underidentical experimental conditions (not illustrated). Thus, ven-tricular myocytes of SUR1-null mouse hearts expressed KATP
channels with biophysical properties that resembled those ofKATP channels, as also reported previously in normal hearts.22
DiscussionThis study was designed to examine the role of SUR1subunits of KATP channels in cardiac function, specificallywith regard to the sequelae of myocardial ischemia andreperfusion. We used an animal model of mice deficient inSUR1 subunits in combination with a well-characterized invivo model of myocardial ischemia/reperfusion injury toinvestigate this question. Surprisingly, these mice werestrongly protected from ischemic insults and displayed re-duced infarct size and preservation of LV structure andfunction after myocardial infarction.
SUR1 Expression in the HeartThe sulfonylurea receptor gene (Abcc8) encodes the high-affinity sulfonylurea receptor, SUR1.23 Transcripts of SUR1
are most abundant in neurons, brain, and pancreatic�-cells.24,25 In the brain, channels composed of SUR1 sub-units may have diverse roles, such as regulating the activity ofglucose-receptive neurons in the ventromedial hypothala-mus,26 whereas in �-cells of the pancreas, SUR1-containingKATP channels have a well-defined role as a trigger for the firstphase of insulin release.27 However, SUR1 transcripts arealso detected in other cell types, including the heart20,24,28–30
(albeit at lower abundance). We have previously detectedSUR1 protein in mouse and rat cardiac myocytes, with thestated reservation that the result should be interpreted withinthe limitations of available antibodies.14 Here, we found lowlevels of SUR1 mRNA expression by comparative real-timeRT-PCR methods, which may imply low expression levels ofSUR1 protein or restricted expression distribution in theheart. A potential role for SUR1 in KATP channel activity in ratatrial myocytes has also been suggested based on a functionaland pharmacological profile that resembles that of the pan-creatic �-cell KATP channel (high affinity for glyburide anddiazoxide).30 Interestingly, human atrial cells are also re-ported to have high sensitivity to blockade by tolbutamide.31
More direct evidence for a role of SUR1 subunits in formingcardiac KATP channels was provided by the finding thatantisense oligonucleotides against SUR1 inhibit KATP channelcurrents in cultured neonatal rat ventricular cells.12 In thepresent study, we were able to record KATP channels fromventricular myocytes isolated from SUR1-null hearts thatwere blocked by cytosolic ATP in the micromolar range andwith a unitary conductance similar to that previously reportedfor ventricular KATP channels.32 Although we did not performa full biophysical or pharmacological analysis of thesechannels in SUR1-null mice, the present data are consistentwith the prevailing notion that SUR1 subunits are notessential subunits of ventricular sarcolemmal KATP channels.
Figure 4. SUR1-null mice displaydecreased fibrosis and inflammatorymarkers after myocardial ischemia/reper-fusion. Representative hematoxylin andeosin–stained histological sections fromWT and SUR1-null mouse hearts after30 minutes of ischemia and 7 days ofreperfusion. SUR1-null hearts displayeda reduced degree of myocardial neutro-philic infiltrate, necrosis, hemorrhage,and spindle-shaped interstitial cells com-pared with WT mice. The optical magni-fication and scale bars are indicated.
Elrod et al SUR1 Subunits of KATP Channel and Cardioprotection 1409
at UCLA BIOMED LIBRARY SERIALS on February 23, 2010 circ.ahajournals.orgDownloaded from
Further studies are warranted to identify the localization andmechanistic role of SUR1 subunits in the heart and cardio-vascular system.
SUR Subunits and Protection From IschemiaKATP channels open during myocardial ischemia and partiallycontribute to K� loss and arrhythmias.33 Opening of KATP
channels with pharmacological agents has been clearly asso-ciated with improvement of myocardial function on reperfu-sion and with a reduction in infarct size.34–36 Conversely,KATP channel blockers are associated with a worse prognosticoutcome.37 The concept that KATP channels are protectiveduring ischemia has been strengthened by genetic studies. InKir6.2-null mice, for example, the increase in LV end-dia-stolic pressure during ischemia was more marked than in WTlittermates, whereas an ischemic preconditioning protocolreduced the infarct size in WT but not Kir6.2-null mice.38
Transgenic overexpression of SUR2A (driven by the ubiqui-tous cytomegalovirus promoter), which is reported to increasethe number of functional KATP channels (but see Flagg et al39),
increases the resistance to hypoxia in isolated cardiac myo-cytes and decreases infarct size in Langendorff-perfusedisolated hearts after a period of global ischemia in thesetransgenic mice.40 In contrast to these studies, a recent studydemonstrated that SUR2-null mice are resistant to acutecardiovascular stress.11 In the latter study, infarct size wasreduced in hearts of SUR2-null mice when Langendorff-perfused and subjected to global ischemia. Interestingly,these SUR2-null mice had hypertension and developed car-diac hypertrophy, but they did not develop significant fibro-sis.11 Paralleling the findings of the latter study, we nowreport that loss of SUR1 subunits also results in increasedprotection from ischemia/reperfusion insults.
Mechanisms Responsible for Cardiac ProtectionPharmacological approaches to study the role of KATP chan-nels in protection from myocardial ischemia/reperfusion in-jury have pivoted on the use of agents that are putativelyselective for sarcolemmal versus mitochondrial KATP chan-nels. For example, diazoxide and 5-hydroxydecanoate, whichrespectively are reported to selectively open or block mito-chondrial KATP channels, protect against ischemic precondi-tioning,41 and this has been taken to argue a role for thesechannels in mediating protection. The argument is compli-cated by reports that 5-hydroxydecanoate can in fact blockcardiac KATP channels,42 possibly in relation to an effectmediated by intracellular cAMP,43 or that it directly fuelsmitochondrial respiration.44 Similarly, diazoxide may alsodirectly affect mitochondrial respiration.44 Although moreselective agents may now be available, a further complicationis that the pharmacology of cardiac KATP channels maydepend on the specific tissue location in the heart and themetabolic state of the cell. For example, diazoxide effectivelyopens cardiac KATP channels when intracellular ADP levelsare elevated4 (as would happen during ischemia). Atrial KATP
channels also show increased sensitivity to diazoxide com-pared with those recorded in ventricular myocytes,30 andventricular KATP channels show increased sensitivity duringremodeling after myocardial ischemia/reperfusion.29
Given the issues raised above, it was hoped that geneticapproaches would clarify the role of KATP channels in myo-cardial ischemia. Unfortunately, they have brought their ownset of challenges. First, mitochondrial KATP channels cannotbe studied genetically until and unless their molecular com-position has been identified. Although consensus does notexist in this regard, the majority of studies have indicated thatconventional KATP channel subunits are not expressed inmitochondria.14 If this is correct, then the genetic models ofKATP channel subunit ablation or overexpression should ad-dress the question of the role(s) of sarcolemmal KATP channels(or any other functions that the KATP channel subunits mayhave) during ischemia.
Consistent with the pharmacological data discussed above,transgenic overexpression of SUR2A subunits is reported toincrease the KATP channel number and to provide betterprotection against ischemia.40 Furthermore, Kir6.2-null micelack cardiac KATP channels and have a decreased resistance toischemia.38 It is not clear, however, how the loss of SUR2subunits (in SUR2-null mice), which is expected to result in
Figure 5. Comparison of KATP channel subunit mRNA expres-sion in WT and SUR1-null hearts. A, RNA was extracted fromcardiac ventricles and subjected to subunit-specific real-timeRT-PCR assays. Expression is quantified relative to that of 18S.Each reaction was performed in triplicate and averaged for eachanimal. B, Conventional RT-PCR reactions were performed withprimers that produce different sizes of SUR2A and SUR2Bamplicons (respectively, 465 and 289 bp; primers used areshown in the Table). Lane annotations are as follows: M, 100-bpmarker; W, PCR control with water added instead of cDNA; C1and C2, 2 selected control lanes; S1 and S2, 2 selected SUR1-null lanes; and N, a negative control performed without thereverse-transcriptase enzyme added to the RT reaction. Densi-tometry was performed after agarose gel electrophoresis. Bargraph depicts average ratios of band intensities. Data aremean�SEM from 2 to 4 separate mice.
1410 Circulation March 18, 2008
at UCLA BIOMED LIBRARY SERIALS on February 23, 2010 circ.ahajournals.orgDownloaded from
loss of cardiac KATP channels, leads to an opposite phenotypeof enhanced resistance to ischemia.11 One possibility is thatthese mice lack both SUR2A and SUR2B (components ofvascular KATP channels), which could account in part for theobserved differences. SUR1-null mice, in which KATP chan-nels with biophysical characteristics resembling known KATP
channels can be measured, similarly are better protectedagainst ischemia. Reciprocal compensation in expression ofother KATP channel subunits is unlikely, because we have notobserved any evidence of altered Kir6.1, Kir6.2, SUR2A, orSUR2B mRNA expression. There may, however, be otheradaptive changes in these mice, such as different posttrans-lational modifications or trafficking events of subunits suchas SUR2. These questions have not been examined in thepresent study. In addition to these possibilities, it is plausiblethat KATP channels in different tissue compartments within theheart or extracardiac tissues, such as the ventricle, atrium,coronary vasculature, or cardiac neurites,6,9,21,30 may becomposed of different SUR subunit isoforms and that SUR1may not be primarily expressed in ventricular myocytes. It isalso possible that SUR subunits have unanticipated functions.For example, we recently identified a role for KATP channelsin the release of the vasoconstrictor endothelin-1 from thecoronary endothelium,45 and although the cellular mecha-nisms remain to be identified, this observation suggests a rolefor KATP channels in exocytosis in the cardiovascular system.KATP channels similarly have been linked to release of atrialnatriuretic peptide from atrial myocytes46,47 and the release ofnorepinephrine and acetylcholine from peripheral nerve end-ings.48,49 Interestingly, a high sensitivity to drugs that act onSUR1 subunits (diazoxide) has been reported,50 and the KATP
channels in dorsal vagal neurons have been suggested to becomposed of Kir6.2/SUR1 channels.51 The extent to whichprocesses such as these contribute to the gain of protection inSUR1-null mice remains to be determined. Finally, the
existence of a nonselective cation channel (distinct from theKATP channel) that has the SUR1 subunit as a component hasbeen reported.52 It appears that SUR1 is upregulated in tissuesin which it was previously only expressed at low levels andthat this newly formed channel contributes to ischemicdamage in the brain and endothelium.53,54 Although thepresent data obtained with SUR1-null mice would be consis-tent with this idea, no such channel with these properties hasbeen described in the heart to the best of our knowledge.
ConclusionsAlthough SUR1 expression has been noted in the heartpreviously, this is the first direct study to define the potentialrole of SUR1 subunits in the cardiovascular system. Thepresent study provides evidence for protection from cardiacischemia/reperfusion in a genetic model of SUR1 deletion.That loss of SUR1 subunits leads to ischemic protection isinteresting and is suggestive of a previously unrecognizedbiological role of this subunit in regulating the function of thecardiovascular system under pathophysiological conditions.The tissue-expression patterns and exact function of theSUR1 subunit remain to be identified. Nevertheless, thesedata may have important therapeutic implications, becauselow concentrations of sulfonylureas and certain KATP chan-nel–opening drugs that are not normally thought to act onconventional ventricular KATP channels may modulate theischemic outcome through yet-to-be-understood processesthat are mediated by SUR1 subunits. This can have importantconsiderations in basic research when “selective” KATP chan-nel drugs are used to identify roles for particular classes ofchannels in biological processes. Additionally, the presentstudy has important implications in the clinic, such as withregard to defining the relationship between sulfonylureatreatment and heart function in diabetes mellitus and ischemicevents in this important group of patients.55–57
Figure 6. Measurement of KATP channelsfrom isolated SUR1-null myocytes.A, Representative recording of KATP
channels measured in the inside-outconfiguration at 100-mV pipette poten-tial. Note that inward currents aredepicted as upward deflections of thecurrent trace. ATP was applied as shownin the top bars. On average, the meanpatch current recorded with 30 �mol/LATP was 27% of that recorded in theabsence of ATP (n�3). B, Expandedtraces of data as indicated in the lowerhorizontal bars in A. C, A current–voltagerelationship was obtained by use of aramp protocol (�100 to 100 mV over 3.8seconds). The average slope conduc-tance was 85.5�1.2 pS (n�8).
Elrod et al SUR1 Subunits of KATP Channel and Cardioprotection 1411
at UCLA BIOMED LIBRARY SERIALS on February 23, 2010 circ.ahajournals.orgDownloaded from

AcknowledgmentsThe authors are grateful to Laura L. Carihill for assistance with someof the RT-PCR experiments.
Sources of FundingThis study was supported by grants from the National Institutes ofHealth (HL060849 to Dr Lefer and HL45742 to Dr Nichols), theAmerican Diabetes Association (7-04-RA-59 to Dr Lefer), and theMasonic Seventh Manhattan District Association (to Dr Coetzee).
DisclosuresNone.
References1. Noma A. ATP-regulated K� channels in cardiac muscle. Nature. 1983;
305:147–148.2. Gross GJ, Auchampach JA. Role of ATP dependent potassium channels
in myocardial ischaemia. Cardiovasc Res. 1992;26:1011–1016.3. Gross GJ, Peart JN. KATP channels and myocardial preconditioning: an
update. Am J Physiol Heart Circ Physiol. 2003;285:H921–H930.4. D’hahan N, Moreau C, Prost AL, Jacquet H, Alekseev AE, Terzic A,
Vivaudou M. Pharmacological plasticity of cardiac ATP-sensitivepotassium channels toward diazoxide revealed by ADP. Proc Natl AcadSci U S A. 1999;96:12162–12167.
5. Seino S, Miki T. Physiological and pathophysiological roles of ATP-sensitive K� channels. Prog Biophys Mol Biol. 2003;81:133–176.
6. Babenko AP, Gonzalez G, Aguilar-Bryan L, Bryan J. Reconstitutedhuman cardiac KATP channels: functional identity with the nativechannels from the sarcolemma of human ventricular cells. Circ Res.1998;83:1132–1143.
7. Suzuki M, Li RA, Miki T, Uemura H, Sakamoto N, Ohmoto-Sekine Y,Tamagawa M, Ogura T, Seino S, Marban E, Nakaya H. Functional rolesof cardiac and vascular ATP-sensitive potassium channels clarified byKir6.2-knockout mice. Circ Res. 2001;88:570–577.
8. Gumina RJ, Pucar D, Bast P, Hodgson DM, Kurtz CE, Dzeja PP, Miki T,Seino S, Terzic A. Knockout of Kir6.2 negates ischemic preconditioning-induced protection of myocardial energetics. Am J Physiol Heart CircPhysiol. 2003;284:H2106–H2113.
9. Chutkow WA, Pu J, Wheeler MT, Wada T, Makielski JC, Burant CF,McNally EM. Episodic coronary artery vasospasm and hypertensiondevelop in the absence of Sur2 K(ATP) channels. J Clin Invest. 2002;110:203–208.
10. Brayden JE. Functional roles of KATP channels in vascular smoothmuscle. Clin Exp Pharmacol Physiol. 2002;29:312–316.
11. Stoller D, Kakkar R, Smelley M, Chalupsky K, Earley JU, Shi NQ,Makielski JC, McNally EM. Mice lacking sulfonylurea receptor 2(SUR2) ATP-sensitive potassium channels are resistant to acute cardio-vascular stress. J Mol Cell Cardiol. 2007;43:445–454.
12. Yokoshiki H, Sunagawa M, Seki T, Sperelakis N. Antisense oligode-oxynucleotides of sulfonylurea receptors inhibit ATP-sensitive K�
channels in cultured neonatal rat ventricular cells. Pflugers Arch. 1999;437:400–408.
13. Hambrock A, Preisig-Muller R, Russ U, Piehl A, Hanley PJ, Ray J, DautJ, Quast U, Derst C. Four novel splice variants of sulfonylurea receptor 1.Am J Physiol Cell Physiol. 2002;283:C587–C598.
14. Morrissey A, Rosner E, Lanning J, Parachuru L, Dhar Chowdhury P, HanS, Lopez G, Tong X, Yoshida H, Nakamura TY, Artman M, Giblin JP,Tinker A, Coetzee WA. Immunolocalization of K(ATP) channel subunitsin mouse and rat cardiac myocytes and the coronary vasculature. BMCPhysiol. 2005;5:1.
15. Shi NQ, Ye B, Makielski JC. Function and distribution of the SURisoforms and splice variants. J Mol Cell Cardiol. 2005;39:51–60.
16. Shiota C, Larsson O, Shelton KD, Shiota M, Efanov AM, Hoy M,Lindner J, Kooptiwut S, Juntti-Berggren L, Gromada J, Berggren PO,Magnuson MA. Sulfonylurea receptor type 1 knock-out mice have intactfeeding-stimulated insulin secretion despite marked impairment in theirresponse to glucose. J Biol Chem. 2002;277:37176–37183.
17. Elrod JW, Greer JJ, Bryan NS, Langston W, Szot JF, Gebregzlabher H,Janssens S, Feelisch M, Lefer DJ. Cardiomyocyte-specific overexpressionof NO synthase-3 protects against myocardial ischemia-reperfusioninjury. Arterioscler Thromb Vasc Biol. 2006;26:1517–1523.
18. Tong X, Porter LM, Liu G, Dhar Chowdhury P, Srivastava S, PountneyD, Yoshida H, Artman M, Fishman GI, Yu C, Iyer R, Morley GE,
Gutstein DE, Coetzee WA. Consequences of cardiac myocyte-specificablation of KATP channels in transgenic mice expressing dominantnegative Kir6 subunits. Am J Physiol Heart Circ Physiol. 2006;291:H543–H551.
19. Hamill OP, Marty A, Neher E, Sakmann B, Sigworth FJ. Improvedpatch-clamp techniques for high-resolution current recording from cellsand cell-free membrane patches. Pflugers Arch. 1981;391:85–100.
20. Harrell MD, Harbi S, Hoffman JF, Zavadil J, Coetzee WA. Large-scaleanalysis of ion channel gene expression in the mouse heart duringperinatal development. Physiol Genomics. 2007;28:273–283.
21. Yoshida H, Feig J, Ghiu IA, Artman M, Coetzee WA. K(ATP) channelsof primary human coronary artery endothelial cells consist of a hetero-multimeric complex of Kir6.1, Kir6.2, and SUR2B subunits. J Mol CellCardiol. 2004;37:857–869.
22. Lederer WJ, Nichols CG. Nucleotide modulation of the activity of ratheart ATP-sensitive K� channels in isolated membrane patches.J Physiol. 1989;419:193–211.
23. Aguilar-Bryan L, Nichols CG, Wechsler SW, Clement JP, Boyd AE III,Gonzalez G, Herrera-Sosa H, Nguy K, Bryan J, Nelson DA. Cloning ofthe beta cell high-affinity sulfonylurea receptor: a regulator of insulinsecretion. Science. 1995;268:423–426.
24. Inagaki N, Gonoi T, Clement JP, Namba N, Inazawa J, Gonzalez G,Aguilar-Bryan L, Seino S, Bryan J. Reconstitution of IKATP: an inwardrectifier subunit plus the sulfonylurea receptor. Science. 1995;270:1166–1170.
25. Karschin C, Ecke C, Ashcroft FM, Karschin A. Overlapping distributionof KATP channel-forming Kir6.2 subunit and the sulfonylurea receptorSUR1 in rodent brain. FEBS Lett. 1997;401:59–64.
26. Lee K, Dixon AK, Richardson JA, Pinnock RD. Glucose-receptiveneurones in the rat ventromedial hypothalamus express KATP channelscomposed of Kir6.1 and SUR1 subunits. J Physiol. 1999;515(pt2):439–452.
27. Nichols CG. KATP channels as molecular sensors of cellular metabolism.Nature. 2006;440:470–476.
28. Hernandez-Sanchez C, Ito Y, Ferrer J, Reitman M, LeRoith D. Charac-terization of the mouse sulfonylurea receptor 1 promoter and its regu-lation. J Biol Chem. 1999;274:18261–18270.
29. Isidoro TN, Philip-Couderc P, Papageorgiou I, Baertschi AJ, Lerch R,Montessuit C. Expression and function of ATP-dependent potassiumchannels in late post-infarction remodeling. J Mol Cell Cardiol. 2007;42:1016–1025.
30. Baron A, van Bever L, Monnier D, Roatti A, Baertschi AJ. A novelK(ATP) current in cultured neonatal rat atrial appendage cardiomyocytes.Circ Res. 1999;85:707–715.
31. Zunkler BJ, Henning B, Ott T, Hildebrandt AG, Fleck E. Effects oftolbutamide on ATP-sensitive K� channels from human right atrialcardiac myocytes. Pharmacol Toxicol. 1997;80:69–75.
32. Nichols CG, Lederer WJ, Cannell MB. ATP dependence of KATP channelkinetics in isolated membrane patches from rat ventricle. Biophys J.1991;60:1164–1177.
33. Kantor PF, Coetzee WA, Carmeliet E, Dennis SC, Opie LH. Reduction ofischemic K� loss and arrhythmias in rat hearts: effect of glibenclamide, asulfonylurea. Circ Res. 1990;66:478–485.
34. Gross GJ, Pieper GM, Warltier DC. Comparative effects of nicorandil,nitroglycerin, nicotinic acid, and SG-86 on the metabolic status andfunctional recovery of the ischemic-reperfused myocardium. J Car-diovasc Pharmacol. 1987;10(suppl 8):S76–S84.
35. Grover GJ, Sleph PG, Parham CS. Nicorandil improves postischemiccontractile function independently of direct myocardial effects. J Car-diovasc Pharmacol. 1990;15:698–705.
36. Hearse DJ. Activation of ATP-sensitive potassium channels: a novelpharmacological approach to myocardial protection. Cardiovasc Res.1995;30:1–17.
37. Thornton JD, Thornton CS, Sterling DL, Downey JM. Blockade ofATP-sensitive potassium channels increases infarct size but does notprevent preconditioning in rabbit hearts. Circ Res. 1993;72:44–49.
38. Suzuki M, Sasaki N, Miki T, Sakamoto N, Ohmoto-Sekine Y, TamagawaM, Seino S, Marban E, Nakaya H. Role of sarcolemmal K(ATP) channelsin cardioprotection against ischemia/reperfusion injury in mice. J ClinInvest. 2002;109:509–516.
39. Flagg TP, Remedi MS, Masia R, Gomes J, McLerie M, Lopatin AN,Nichols CG. Transgenic overexpression of SUR1 in the heart suppressessarcolemmal K(ATP). J Mol Cell Cardiol. 2005;39:647–656.
40. Du Q, Jovanovic S, Clelland A, Sukhodub A, Budas G, Phelan K,Murray-Tait V, Malone L, Jovanovic A. Overexpression of SUR2A
1412 Circulation March 18, 2008
at UCLA BIOMED LIBRARY SERIALS on February 23, 2010 circ.ahajournals.orgDownloaded from

generates a cardiac phenotype resistant to ischemia. FASEB J. 2006;20:1131–1141.
41. Grover GJ, Garlid KD. ATP-sensitive potassium channels: a review oftheir cardioprotective pharmacology. J Mol Cell Cardiol. 2000;32:677–695.
42. Notsu T, Ohhashi K, Tanaka I, Ishikawa H, Niho T, Fukutake K, MizotaM. 5-Hydroxydecanoate inhibits ATP-sensitive K� channel currents inguinea-pig single ventricular myocytes. Eur J Pharmacol. 1992;220:35–41.
43. Notsu T, Tanaka I, Mizota M, Yanagibashi K, Fukutake K. A cAMP-dependent protein kinase inhibitor modulates the blocking action of ATPand 5-hydroxydecanoate on the ATP-sensitive K� channel. Life Sci.1992;51:1851–1856.
44. Hanley PJ, Mickel M, Loffler M, Brandt U, Daut J. K(ATP) channel-independent targets of diazoxide and 5-hydroxydecanoate in the heart.J Physiol. 2002;542:735–741.
45. Malester B, Tong X, Ghiu IA, Kontogeorgis A, Gutstein DE, Xu J,Hendricks-Munoz K, Coetzee WA. Transgenic expression of a dominantnegative KATP channel subunit in the mouse endothelium: effects oncoronary flow and endothelin-1 secretion. FASEB J. 2007;21:2162–2172.
46. Kim SH, Cho KW, Chang SH, Kim SZ, Chae SW. Glibenclamide sup-presses stretch-activated ANP secretion: involvements of K�ATPchannels and L-type Ca2� channel modulation. Pflugers Arch. 1997;434:362–372.
47. Saegusa N, Sato T, Saito T, Tamagawa M, Komuro I, Nakaya H. Kir6.2-deficient mice are susceptible to stimulated ANP secretion: K(ATP)channel acts as a negative feedback mechanism? Cardiovasc Res. 2005;67:60–68.
48. Oe K, Sperlagh B, Santha E, Matko I, Nagashima H, Foldes FF, Vizi ES.Modulation of norepinephrine release by ATP-dependent K(�)-channelactivators and inhibitors in guinea-pig and human isolated right atrium.Cardiovasc Res. 1999;43:125–134.
49. Kilbinger H, Krause A, Mang CF, Englert H, Wirth K. Effects of K(ATP)channel modulators on acetylcholine release from guinea-pig isolatedatria and small intestine. Naunyn Schmiedebergs Arch Pharmacol. 2002;365:371–377.
50. Mohan RM, Paterson DJ. Activation of sulphonylurea-sensitive channelsand the NO-cGMP pathway decreases the heart rate response to sympa-thetic nerve stimulation. Cardiovasc Res. 2000;47:81–89.
51. Karschin A, Brockhaus J, Ballanyi K. KATP channel formation by thesulphonylurea receptors SUR1 with Kir6.2 subunits in rat dorsal vagalneurons in situ. J Physiol. 1998;509:339–346.
52. Chen M, Dong Y, Simard JM. Functional coupling between sulfonylureareceptor type 1 and a nonselective cation channel in reactive astrocytesfrom adult rat brain. J Neurosci. 2003;23:8568–8577.
53. Simard JM, Chen M, Tarasov KV, Bhatta S, Ivanova S, Melnitchenko L,Tsymbalyuk N, West GA, Gerzanich V. Newly expressed SUR1-regulated NC(Ca-ATP) channel mediates cerebral edema after ischemicstroke. Nat Med. 2006;12:433–440.
54. Simard JM, Tsymbalyuk O, Ivanov A, Ivanova S, Bhatta S, Geng Z, WooSK, Gerzanich V. Endothelial sulfonylurea receptor 1-regulated NCCa-ATP channels mediate progressive hemorrhagic necrosis followingspinal cord injury. J Clin Invest. 2007;117:2105–2113.
55. Pogatsa G, Koltai MZ, Jermendy G, Simon J, Aranyi Z, Ballagi-PordanyG. The effect of sulphonylurea therapy on the outcome of coronary heartdiseases in diabetic patients. Acta Med Hung. 1992;49:39–51.
56. Klepzig H, Kober G, Matter C, Luus H, Schneider H, Boedeker KH,Kiowski W, Amann FW, Gruber D, Harris S, Burger W. Sulfonylureasand ischaemic preconditioning: a double-blind, placebo-controlled eval-uation of glimepiride and glibenclamide. Eur Heart J. 1999;20:439–446.
57. Johnsen SP, Monster TB, Olsen ML, Thisted H, McLaughlin JK,Sorensen HT, Lervang HH, Rungby J. Risk and short-term prognosis ofmyocardial infarction among users of antidiabetic drugs. Am J Ther.2006;13:134–140.
CLINICAL PERSPECTIVEEarly studies (eg, the University Group Diabetes Program) have suggested an excess of cardiovascular mortality intolbutamide-treated subjects. Second-generation sulfonylureas are thought to pose a lesser cardiovascular risk. The effectsof sulfonylureas on cardiovascular infarct size and ischemic arrhythmias remain controversial. Molecular studies areneeded to resolve this complex question. In cardiac myocytes, the relevant KATP channel subunits are generally thought tobe Kir6.2 and SUR2A; however, expression of other KATP channel subunit family members (Kir6.1 and SUR1) occurs inthe heart, where their role remains undefined. The present study investigated the cardiovascular role of SUR1 subunits andfound that mice lacking these subunits are markedly protected from ischemia/reperfusion injury. Sulfonylureas bind toSUR1 subunits of the KATP channel and inhibit K� efflux. These data may have important ramifications for thecardiovascular risk of diabetic patients treated with sulfonylureas. Specifically, the prediction would be that blockade ofSUR1-containing channels with low sulfonylurea concentrations in the cardiovascular system would be cardioprotectivevia a reduction in infarct size after ischemia/reperfusion. A caveat is that blockade of a potassium channel would also beproexcitatory and potentially proarrhythmic. Effects on arrhythmias have not been investigated in the present study, andfurther studies are needed to evaluate the cardiovascular effects of SUR1-containing channels in the cardiovascular system.Nevertheless, this study suggests a previously unrecognized diversity of cardiovascular KATP channels that may affectcardiovascular outcome with sulfonylurea treatment.
Elrod et al SUR1 Subunits of KATP Channel and Cardioprotection 1413
at UCLA BIOMED LIBRARY SERIALS on February 23, 2010 circ.ahajournals.orgDownloaded from

In the article by Elrod et al, “Role of Sulfonylurea Receptor Type 1 Subunits of ATP-SensitivePotassium Channels in Myocardial Ischemia/Reperfusion Injury,” which appeared in the March18, 2008, issue of the journal (Circulation. 2008:117:1405–1413), the following correction shouldbe made:
On page 1406, in the first sentence of Methods, “Mice deficient in the ABCC9 gene” shouldbe changed to “Mice deficient in the ABCC8 gene.”
This correction has been made in the current online version of the article. The authors regret thiserror.
DOI: 10.1161/CIRCULATIONAHA.107.189620
(Circulation. 2008;117:e332.)© 2008 American Heart Association, Inc.
Circulation is available at http://circ.ahajournals.org
e332
Correction
at UCLA BIOMED LIBRARY SERIALS on February 23, 2010 circ.ahajournals.orgDownloaded from
Related Documents











