Changes in the midgut diverticula in the harvestmen Amilenus aurantiacus (Phalangiidae, Opiliones) during winter diapause Sa ska Lipov sek a, b, c , Tone Novak b , Franc Jan zekovi c b , Gerd Leitinger d, * a Medical Faculty, University of Maribor, Taborska ulica 8, SI-2000 Maribor, Slovenia b Department of Biology, Faculty of Natural Sciences and Mathematics, University of Maribor, Koro ska cesta 160, SI-2000 Maribor, Slovenia c Faculty of Chemistry and Chemical Engineering, Smetanova ulica 17, University of Maribor, SI-2000 Maribor, Slovenia d Institute of Cell Biology, Histology and Embryology, Research Unit Electron Microscopic Techniques, Medical University of Graz, Harrachgasse 21, Graz A-8010, Austria article info Article history: Received 21 August 2014 Accepted 15 December 2014 Available online 26 December 2014 Keywords: Arachnids Autophagy Diapause Glycogen Lipids Ultrastructure abstract The harvestmen Amilenus aurantiacus overwinter in diapause in hypogean habitats. The midgut diver- ticula have been studied microscopically (light microscopy, TEM) and biochemically (energy-storing compounds: lipids and glycogen) to analyze changes during this programmed starvation period. Throughout the investigated period, the epithelium of the midgut diverticula is composed of secretory cells, digestive cells and adipocytes. Additionally, after the middle of overwintering, the excretory cells appear, and two assemblages of secretory cells are present: the SC1 secretory cells are characterized by electron-dense cytoplasm with numerous protein granules, and the SC2 cells by an electron-lucent cytoplasm with fewer protein granules. The autophagic activity is observed from the middle of over- wintering, indicating its vital role in providing nutrients during this non-feeding period. Lipids and glycogen are present in the midgut diverticula cells, except in the excretory cells. Measurements of the lipid droplet diameters and the lipid quantities yielded quite comparable information on their con- sumption. Lipids are gradually spent in both sexes, more rapidly in females, owing to ripening of the ovaries. Glycogen rates decrease towards the middle, and increase just before the end of overwintering, indicating that individuals are preparing for the epigean active ecophase. © 2014 Elsevier Ltd. All rights reserved. 1. Introduction The midgut is the largest organ in harvestmen and performs many vital functions, including digestion, nutrient absorption, nutrient and waste storage and water uptake (Becker and Peters, 1985; Shultz and Pinto-da-Rocha, 2007). It is divided into an anterior ventriculus and a posterior postventriculus by a dorsal transverse fold. Numerous prosomal and opisthosomal tubular lateral branches or diverticules arise from the lateral walls of the ventriculus, sometimes referred to collectively as the midgut gland (Shultz and Pinto-da-Rocha, 2007). The luminal side of the midgut diverticula is composed of a single-layer epithelium of columnar cells. Their apical plasma membrane is differentiated into microvilli projecting into the midgut lumen (Shultz and Pinto-da-Rocha, 2007). Amilenus aurantiacus hatch in late summer (Martens, 1978). After the first autumn frost in November or December, they enter hypogean habitats and overwinter there (Martens, 1969, 1978). During overwintering in caves, they moult twice and undergo a diapause (Bourne, 1978b). In March or April, adults or subadults (own unpublished data) return to the epigean habitats. They mature in late spring and die in the late summer or early autumn (Martens, 1969, 1978; Bourne, 1978a). During winter, in some caves individuals occur in mass; they do not feed in that period of their life cycle (Jequier, 1964; Martens, 1969; Bourne, 1978a; Novak et al., 1984, 2012, 2013). In central Europe, Amilenus aurantiacus is the most abundant non-troglobiotic terrestrial macroinvertebrate species overwintering in caves (Novak et al., 2013). Bourne (1978b) provided the first information on the digestion of A. aurantiacus during its winter diapause. The liquid part of the consumed food passes into the midgut diverticula, while solid particles with a little liquid are retained in the midgut. In the * Corresponding author. Tel.: þ43 316 380 4237; fax: þ43 316 385 9625. E-mail addresses: [email protected] (S. Lipov sek), gerd.leitinger@ medunigraz.at (G. Leitinger). Contents lists available at ScienceDirect Arthropod Structure & Development journal homepage: www.elsevier.com/locate/asd http://dx.doi.org/10.1016/j.asd.2014.12.002 1467-8039/© 2014 Elsevier Ltd. All rights reserved. Arthropod Structure & Development 44 (2015) 131e141

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

lable at ScienceDirect
Arthropod Structure & Development 44 (2015) 131e141
Contents lists avai
Arthropod Structure & Development
journal homepage: www.elsevier .com/locate/asd
Changes in the midgut diverticula in the harvestmen Amilenusaurantiacus (Phalangiidae, Opiliones) during winter diapause
Sa�ska Lipov�sek a, b, c, Tone Novak b, Franc Jan�zekovi�c b, Gerd Leitinger d, *
a Medical Faculty, University of Maribor, Taborska ulica 8, SI-2000 Maribor, Sloveniab Department of Biology, Faculty of Natural Sciences and Mathematics, University of Maribor, Koro�ska cesta 160, SI-2000 Maribor, Sloveniac Faculty of Chemistry and Chemical Engineering, Smetanova ulica 17, University of Maribor, SI-2000 Maribor, Sloveniad Institute of Cell Biology, Histology and Embryology, Research Unit Electron Microscopic Techniques, Medical University of Graz, Harrachgasse 21,Graz A-8010, Austria
a r t i c l e i n f o
Article history:Received 21 August 2014Accepted 15 December 2014Available online 26 December 2014
Keywords:ArachnidsAutophagyDiapauseGlycogenLipidsUltrastructure
* Corresponding author. Tel.: þ43 316 380 4237; faE-mail addresses: [email protected] (S.
medunigraz.at (G. Leitinger).
http://dx.doi.org/10.1016/j.asd.2014.12.0021467-8039/© 2014 Elsevier Ltd. All rights reserved.
a b s t r a c t
The harvestmen Amilenus aurantiacus overwinter in diapause in hypogean habitats. The midgut diver-ticula have been studied microscopically (light microscopy, TEM) and biochemically (energy-storingcompounds: lipids and glycogen) to analyze changes during this programmed starvation period.Throughout the investigated period, the epithelium of the midgut diverticula is composed of secretorycells, digestive cells and adipocytes. Additionally, after the middle of overwintering, the excretory cellsappear, and two assemblages of secretory cells are present: the SC1 secretory cells are characterized byelectron-dense cytoplasm with numerous protein granules, and the SC2 cells by an electron-lucentcytoplasm with fewer protein granules. The autophagic activity is observed from the middle of over-wintering, indicating its vital role in providing nutrients during this non-feeding period. Lipids andglycogen are present in the midgut diverticula cells, except in the excretory cells. Measurements of thelipid droplet diameters and the lipid quantities yielded quite comparable information on their con-sumption. Lipids are gradually spent in both sexes, more rapidly in females, owing to ripening of theovaries. Glycogen rates decrease towards the middle, and increase just before the end of overwintering,indicating that individuals are preparing for the epigean active ecophase.
© 2014 Elsevier Ltd. All rights reserved.
1. Introduction
The midgut is the largest organ in harvestmen and performsmany vital functions, including digestion, nutrient absorption,nutrient and waste storage and water uptake (Becker and Peters,1985; Shultz and Pinto-da-Rocha, 2007). It is divided into ananterior ventriculus and a posterior postventriculus by a dorsaltransverse fold. Numerous prosomal and opisthosomal tubularlateral branches or diverticules arise from the lateral walls of theventriculus, sometimes referred to collectively as the midgut gland(Shultz and Pinto-da-Rocha, 2007). The luminal side of the midgutdiverticula is composed of a single-layer epithelium of columnarcells. Their apical plasmamembrane is differentiated intomicrovilli
x: þ43 316 385 9625.Lipov�sek), gerd.leitinger@
projecting into the midgut lumen (Shultz and Pinto-da-Rocha,2007).
Amilenus aurantiacus hatch in late summer (Martens, 1978).After the first autumn frost in November or December, they enterhypogean habitats and overwinter there (Martens, 1969, 1978).During overwintering in caves, they moult twice and undergo adiapause (Bourne, 1978b). In March or April, adults or subadults(own unpublished data) return to the epigean habitats. Theymature in late spring and die in the late summer or early autumn(Martens, 1969, 1978; Bourne, 1978a). During winter, in some cavesindividuals occur in mass; they do not feed in that period of theirlife cycle (Jequier, 1964; Martens, 1969; Bourne, 1978a; Novak et al.,1984, 2012, 2013). In central Europe, Amilenus aurantiacus is themost abundant non-troglobiotic terrestrial macroinvertebratespecies overwintering in caves (Novak et al., 2013).
Bourne (1978b) provided the first information on the digestionof A. aurantiacus during its winter diapause. The liquid part of theconsumed food passes into the midgut diverticula, while solidparticles with a little liquid are retained in the midgut. In the
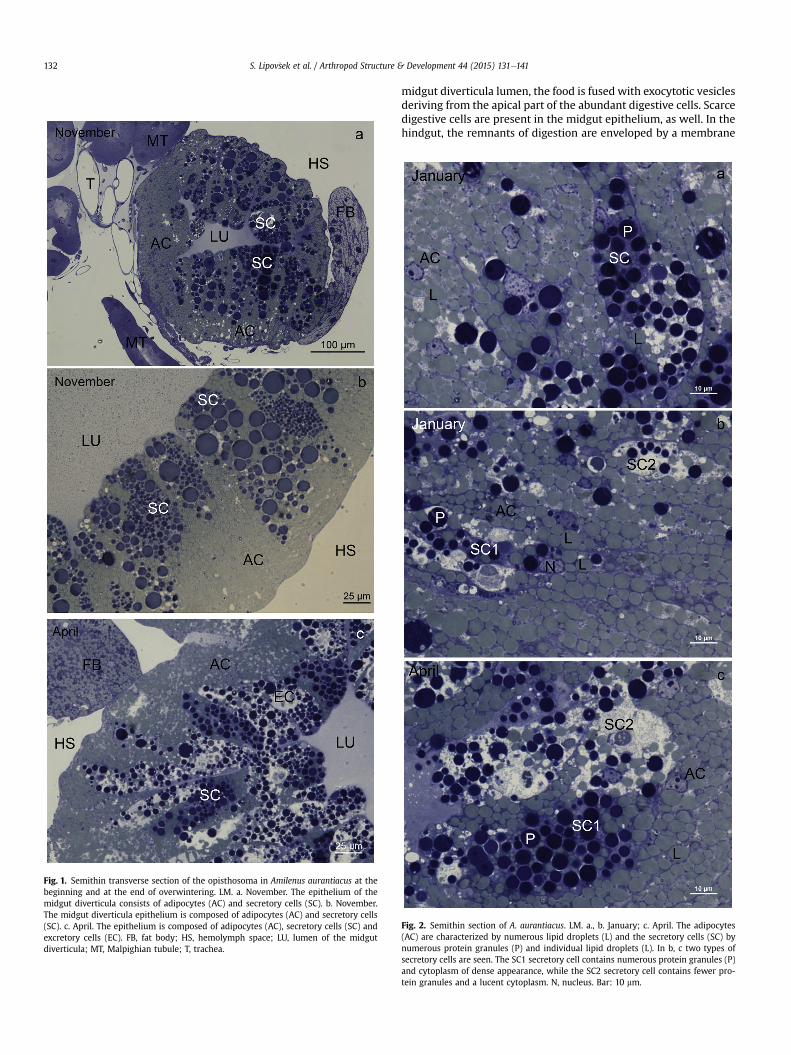
Fig. 1. Semithin transverse section of the opisthosoma in Amilenus aurantiacus at thebeginning and at the end of overwintering. LM. a. November. The epithelium of themidgut diverticula consists of adipocytes (AC) and secretory cells (SC). b. November.The midgut diverticula epithelium is composed of adipocytes (AC) and secretory cells(SC). c. April. The epithelium is composed of adipocytes (AC), secretory cells (SC) andexcretory cells (EC). FB, fat body; HS, hemolymph space; LU, lumen of the midgutdiverticula; MT, Malpighian tubule; T, trachea.
S. Lipov�sek et al. / Arthropod Structure & Development 44 (2015) 131e141132
midgut diverticula lumen, the food is fused with exocytotic vesiclesderiving from the apical part of the abundant digestive cells. Scarcedigestive cells are present in the midgut epithelium, as well. In thehindgut, the remnants of digestion are enveloped by a membrane
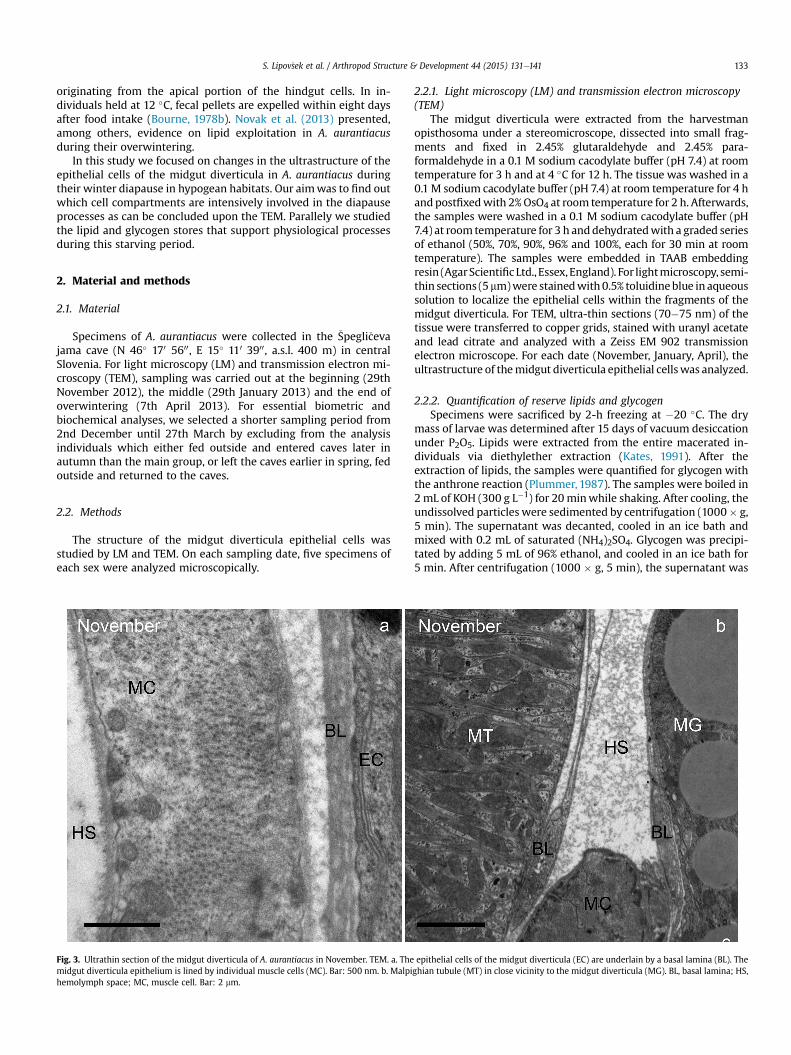
Fig. 2. Semithin section of A. aurantiacus. LM. a., b. January; c. April. The adipocytes(AC) are characterized by numerous lipid droplets (L) and the secretory cells (SC) bynumerous protein granules (P) and individual lipid droplets (L). In b, c two types ofsecretory cells are seen. The SC1 secretory cell contains numerous protein granules (P)and cytoplasm of dense appearance, while the SC2 secretory cell contains fewer pro-tein granules and a lucent cytoplasm. N, nucleus. Bar: 10 mm.
S. Lipov�sek et al. / Arthropod Structure & Development 44 (2015) 131e141 133
originating from the apical portion of the hindgut cells. In in-dividuals held at 12 �C, fecal pellets are expelled within eight daysafter food intake (Bourne, 1978b). Novak et al. (2013) presented,among others, evidence on lipid exploitation in A. aurantiacusduring their overwintering.
In this study we focused on changes in the ultrastructure of theepithelial cells of the midgut diverticula in A. aurantiacus duringtheir winter diapause in hypogean habitats. Our aimwas to find outwhich cell compartments are intensively involved in the diapauseprocesses as can be concluded upon the TEM. Parallely we studiedthe lipid and glycogen stores that support physiological processesduring this starving period.
2. Material and methods
2.1. Material
Specimens of A. aurantiacus were collected in the �Spegli�cevajama cave (N 46� 170 5600, E 15� 110 3900, a.s.l. 400 m) in centralSlovenia. For light microscopy (LM) and transmission electron mi-croscopy (TEM), sampling was carried out at the beginning (29thNovember 2012), the middle (29th January 2013) and the end ofoverwintering (7th April 2013). For essential biometric andbiochemical analyses, we selected a shorter sampling period from2nd December until 27th March by excluding from the analysisindividuals which either fed outside and entered caves later inautumn than the main group, or left the caves earlier in spring, fedoutside and returned to the caves.
2.2. Methods
The structure of the midgut diverticula epithelial cells wasstudied by LM and TEM. On each sampling date, five specimens ofeach sex were analyzed microscopically.
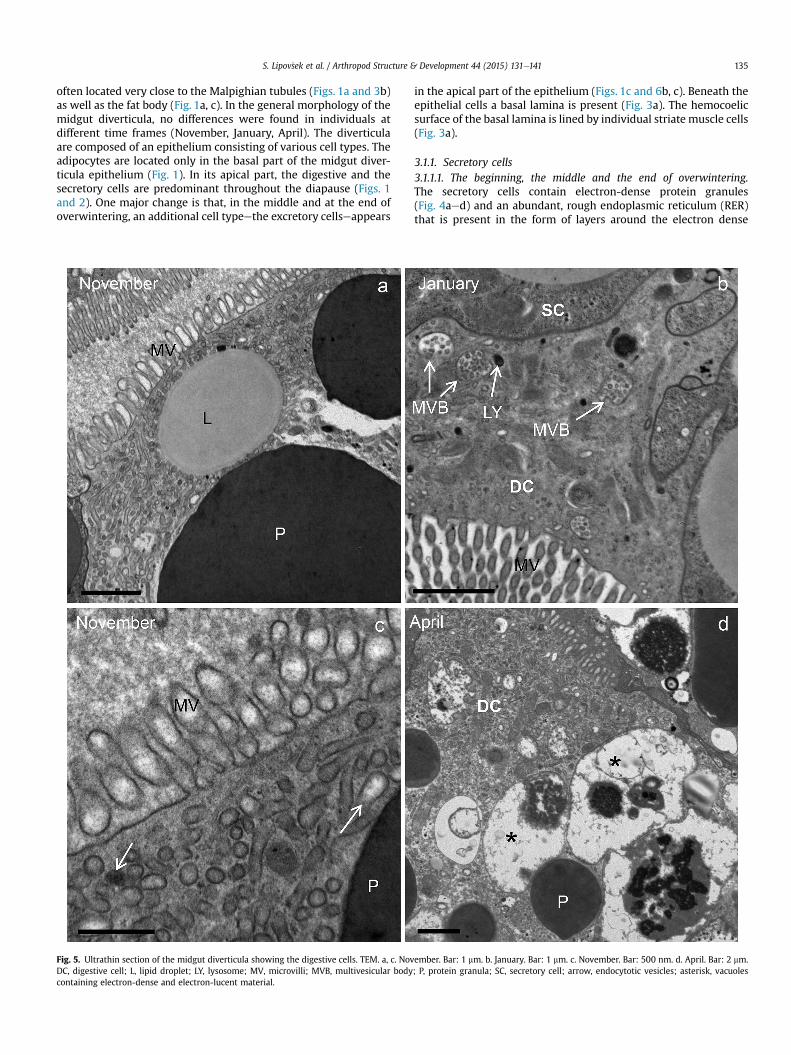
Fig. 3. Ultrathin section of the midgut diverticula of A. aurantiacus in November. TEM. a. Themidgut diverticula epithelium is lined by individual muscle cells (MC). Bar: 500 nm. b. Malpihemolymph space; MC, muscle cell. Bar: 2 mm.
2.2.1. Light microscopy (LM) and transmission electron microscopy(TEM)
The midgut diverticula were extracted from the harvestmanopisthosoma under a stereomicroscope, dissected into small frag-ments and fixed in 2.45% glutaraldehyde and 2.45% para-formaldehyde in a 0.1 M sodium cacodylate buffer (pH 7.4) at roomtemperature for 3 h and at 4 �C for 12 h. The tissue was washed in a0.1 M sodium cacodylate buffer (pH 7.4) at room temperature for 4 handpostfixedwith 2%OsO4 at room temperature for 2 h. Afterwards,the samples were washed in a 0.1 M sodium cacodylate buffer (pH7.4) at room temperature for 3 h anddehydratedwith a graded seriesof ethanol (50%, 70%, 90%, 96% and 100%, each for 30 min at roomtemperature). The samples were embedded in TAAB embeddingresin (AgarScientific Ltd., Essex,England). For lightmicroscopy, semi-thin sections (5mm)were stainedwith0.5% toluidineblue in aqueoussolution to localize the epithelial cells within the fragments of themidgut diverticula. For TEM, ultra-thin sections (70�75 nm) of thetissue were transferred to copper grids, stained with uranyl acetateand lead citrate and analyzed with a Zeiss EM 902 transmissionelectron microscope. For each date (November, January, April), theultrastructure of themidgut diverticula epithelial cellswas analyzed.
2.2.2. Quantification of reserve lipids and glycogenSpecimens were sacrificed by 2-h freezing at �20 �C. The dry
mass of larvae was determined after 15 days of vacuum desiccationunder P2O5. Lipids were extracted from the entire macerated in-dividuals via diethylether extraction (Kates, 1991). After theextraction of lipids, the samples were quantified for glycogen withthe anthrone reaction (Plummer, 1987). The samples were boiled in2mL of KOH (300 g L�1) for 20minwhile shaking. After cooling, theundissolved particles were sedimented by centrifugation (1000� g,5 min). The supernatant was decanted, cooled in an ice bath andmixed with 0.2 mL of saturated (NH4)2SO4. Glycogen was precipi-tated by adding 5 mL of 96% ethanol, and cooled in an ice bath for5 min. After centrifugation (1000 � g, 5 min), the supernatant was
epithelial cells of the midgut diverticula (EC) are underlain by a basal lamina (BL). Theghian tubule (MT) in close vicinity to the midgut diverticula (MG). BL, basal lamina; HS,

S. Lipov�sek et al. / Arthropod Structure & Development 44 (2015) 131e141134
discharged, and the sedimented glycogen dried at 105 �C, weighed,and then dissolved in 4 mL water and warmed to 100 �C. TwomL ofthe anthrone reagent (2 g l�1 in conc. H2SO4) was added to 0.5mL ofthe glycogen solution and warmed in a boiling water bath for10 min. After cooling, the absorption was recorded at 620 nmagainst a reagent blank. A glucose solution (0.1 g L�1) was used asthe reference, and a correction factor of 0.9 was used in calculatingthe glycogen amounts.
2.3. Statistical analysis
The t-test was used in testing differences between means of themeasured parameters in males and females. One-way ANOVA was
Fig. 4. Ultrathin section of the midgut diverticula showing the secretory cells and the digestBar: 1 mm. AL, autolysosome; DC, digestive cell; L, lipid droplet; MV, microvilli; P, protein g
used in testing differences between the three time frames. The dataanalysis was carried out with the statistical software SPSS 21.0.
3. Results
3.1. Structure investigations
In all the diapausing A. aurantiacus, between the beginning andthe end of overwintering the digestive tract is empty. Thedescription of the midgut diverticula of A. aurantiacus relates toboth male and female juveniles (in the following text: males andfemales), since no differences are observed between the sexesregarding the organization of this organ. The midgut diverticula are
ive cells. TEM. a, c. November. Bar: 2 mm. b. January. Bar: 2 mm. c. Bar: 500 nm. d. April.ranulum; RER, rough endoplasmic reticulum; S, spherite; SC, secretory cell.
S. Lipov�sek et al. / Arthropod Structure & Development 44 (2015) 131e141 135
often located very close to the Malpighian tubules (Figs. 1a and 3b)as well as the fat body (Fig. 1a, c). In the general morphology of themidgut diverticula, no differences were found in individuals atdifferent time frames (November, January, April). The diverticulaare composed of an epithelium consisting of various cell types. Theadipocytes are located only in the basal part of the midgut diver-ticula epithelium (Fig. 1). In its apical part, the digestive and thesecretory cells are predominant throughout the diapause (Figs. 1and 2). One major change is that, in the middle and at the end ofoverwintering, an additional cell typeethe excretory cellseappears
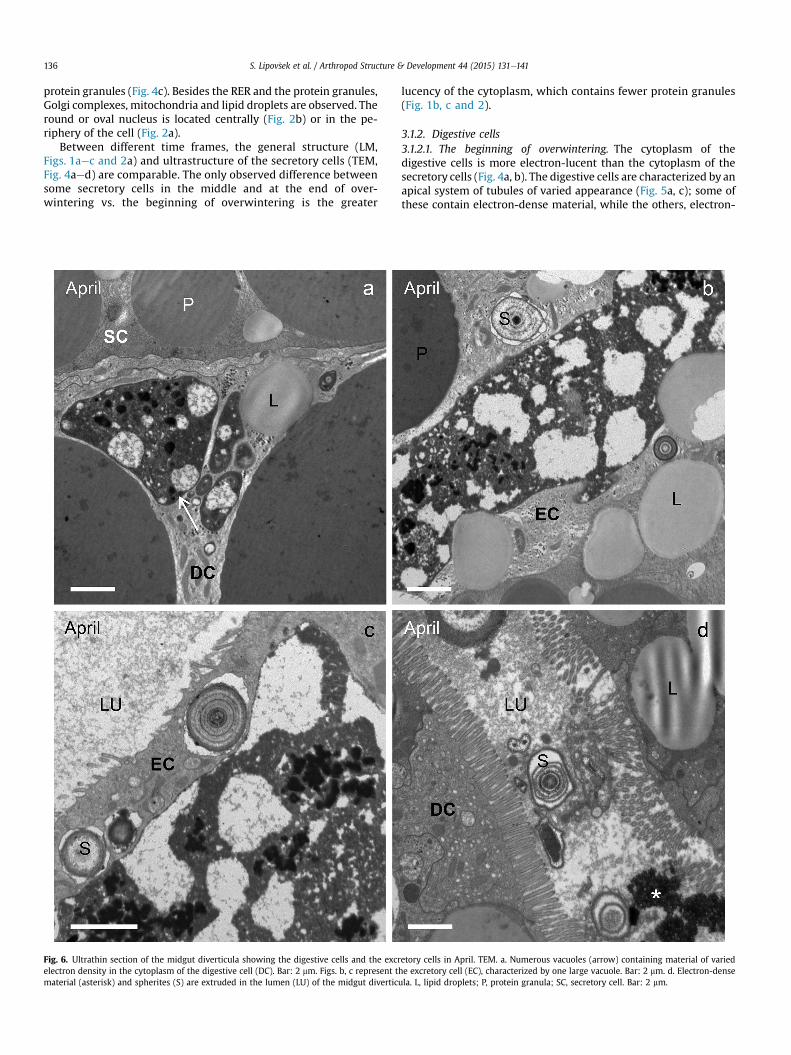
Fig. 5. Ultrathin section of the midgut diverticula showing the digestive cells. TEM. a, c. NovDC, digestive cell; L, lipid droplet; LY, lysosome; MV, microvilli; MVB, multivesicular bodycontaining electron-dense and electron-lucent material.
in the apical part of the epithelium (Figs. 1c and 6b, c). Beneath theepithelial cells a basal lamina is present (Fig. 3a). The hemocoelicsurface of the basal lamina is lined by individual striate muscle cells(Fig. 3a).
3.1.1. Secretory cells3.1.1.1. The beginning, the middle and the end of overwintering.The secretory cells contain electron-dense protein granules(Fig. 4aed) and an abundant, rough endoplasmic reticulum (RER)that is present in the form of layers around the electron dense
ember. Bar: 1 mm. b. January. Bar: 1 mm. c. November. Bar: 500 nm. d. April. Bar: 2 mm.; P, protein granula; SC, secretory cell; arrow, endocytotic vesicles; asterisk, vacuoles
S. Lipov�sek et al. / Arthropod Structure & Development 44 (2015) 131e141136
protein granules (Fig. 4c). Besides the RER and the protein granules,Golgi complexes, mitochondria and lipid droplets are observed. Theround or oval nucleus is located centrally (Fig. 2b) or in the pe-riphery of the cell (Fig. 2a).
Between different time frames, the general structure (LM,Figs. 1aec and 2a) and ultrastructure of the secretory cells (TEM,Fig. 4aed) are comparable. The only observed difference betweensome secretory cells in the middle and at the end of over-wintering vs. the beginning of overwintering is the greater
Fig. 6. Ultrathin section of the midgut diverticula showing the digestive cells and the excrelectron density in the cytoplasm of the digestive cell (DC). Bar: 2 mm. Figs. b, c represent tmaterial (asterisk) and spherites (S) are extruded in the lumen (LU) of the midgut divertic
lucency of the cytoplasm, which contains fewer protein granules(Fig. 1b, c and 2).
3.1.2. Digestive cells3.1.2.1. The beginning of overwintering. The cytoplasm of thedigestive cells is more electron-lucent than the cytoplasm of thesecretory cells (Fig. 4a, b). The digestive cells are characterized by anapical system of tubules of varied appearance (Fig. 5a, c); some ofthese contain electron-dense material, while the others, electron-
etory cells in April. TEM. a. Numerous vacuoles (arrow) containing material of variedhe excretory cell (EC), characterized by one large vacuole. Bar: 2 mm. d. Electron-denseula. L, lipid droplets; P, protein granula; SC, secretory cell. Bar: 2 mm.
S. Lipov�sek et al. / Arthropod Structure & Development 44 (2015) 131e141 137
lucent material (Fig. 5c). The cytoplasm contains electron-densevacuoles, lipid droplets, glycogen granules and spherites. The RERis less abundant than in the secretory cell. The round to oval nu-cleus is located in the center or in the basal part of the cell.
3.1.2.2. The middle and the end of overwintering. In the middle ofoverwintering, in some digestive cells multivesicular bodies andlysosomes are observed (Fig. 5b). At the end of overwintering,autolysosomes, characterized by vesicles containing eitherelectron-dense material or the material of various electron den-sities (Fig. 5d), are present in the cytoplasm. Autolysosomes areabsent at the beginning of overwintering, scarce in the middle and
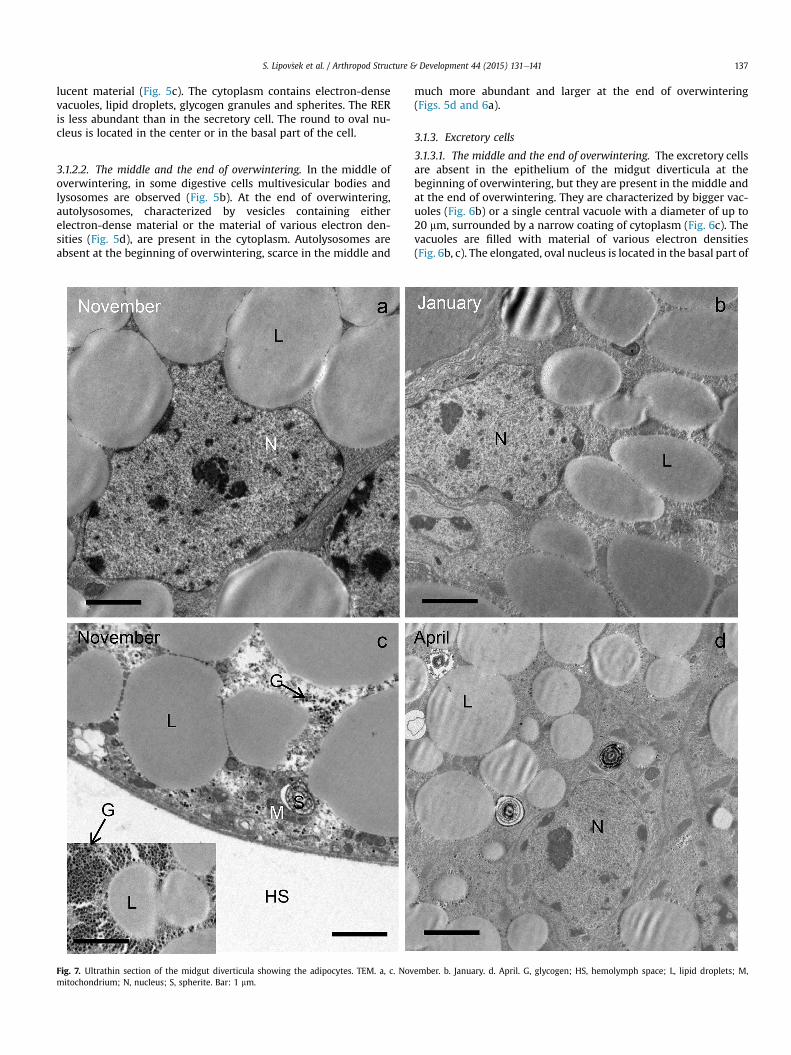
Fig. 7. Ultrathin section of the midgut diverticula showing the adipocytes. TEM. a, c. Novmitochondrium; N, nucleus; S, spherite. Bar: 1 mm.
much more abundant and larger at the end of overwintering(Figs. 5d and 6a).
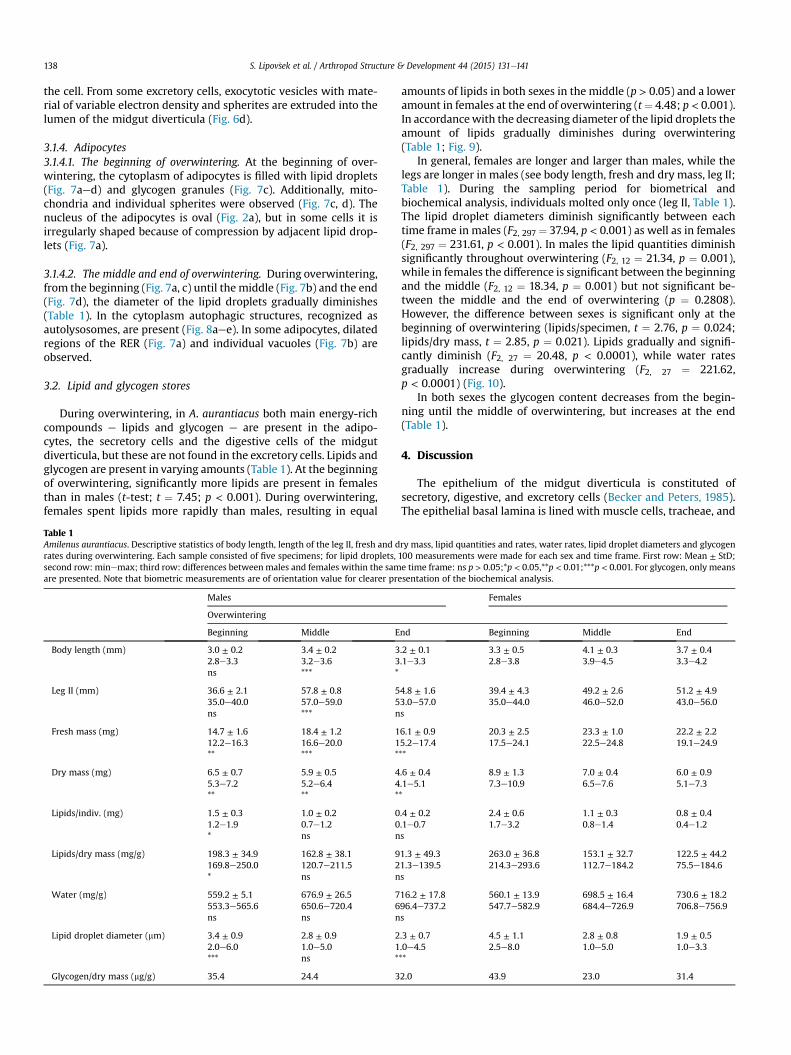
3.1.3. Excretory cells
3.1.3.1. The middle and the end of overwintering. The excretory cellsare absent in the epithelium of the midgut diverticula at thebeginning of overwintering, but they are present in the middle andat the end of overwintering. They are characterized by bigger vac-uoles (Fig. 6b) or a single central vacuole with a diameter of up to20 mm, surrounded by a narrow coating of cytoplasm (Fig. 6c). Thevacuoles are filled with material of various electron densities(Fig. 6b, c). The elongated, oval nucleus is located in the basal part of
ember. b. January. d. April. G, glycogen; HS, hemolymph space; L, lipid droplets; M,
S. Lipov�sek et al. / Arthropod Structure & Development 44 (2015) 131e141138
the cell. From some excretory cells, exocytotic vesicles with mate-rial of variable electron density and spherites are extruded into thelumen of the midgut diverticula (Fig. 6d).
3.1.4. Adipocytes3.1.4.1. The beginning of overwintering. At the beginning of over-wintering, the cytoplasm of adipocytes is filled with lipid droplets(Fig. 7aed) and glycogen granules (Fig. 7c). Additionally, mito-chondria and individual spherites were observed (Fig. 7c, d). Thenucleus of the adipocytes is oval (Fig. 2a), but in some cells it isirregularly shaped because of compression by adjacent lipid drop-lets (Fig. 7a).
3.1.4.2. The middle and end of overwintering. During overwintering,from the beginning (Fig. 7a, c) until the middle (Fig. 7b) and the end(Fig. 7d), the diameter of the lipid droplets gradually diminishes(Table 1). In the cytoplasm autophagic structures, recognized asautolysosomes, are present (Fig. 8aee). In some adipocytes, dilatedregions of the RER (Fig. 7a) and individual vacuoles (Fig. 7b) areobserved.
3.2. Lipid and glycogen stores
During overwintering, in A. aurantiacus both main energy-richcompounds e lipids and glycogen e are present in the adipo-cytes, the secretory cells and the digestive cells of the midgutdiverticula, but these are not found in the excretory cells. Lipids andglycogen are present in varying amounts (Table 1). At the beginningof overwintering, significantly more lipids are present in femalesthan in males (t-test; t ¼ 7.45; p < 0.001). During overwintering,females spent lipids more rapidly than males, resulting in equal
Table 1Amilenus aurantiacus. Descriptive statistics of body length, length of the leg II, fresh and drates during overwintering. Each sample consisted of five specimens; for lipid droplets,second row: minemax; third row: differences betweenmales and females within the samare presented. Note that biometric measurements are of orientation value for clearer pre
Males
Overwintering
Beginning Middle E
Body length (mm) 3.0 ± 0.2 3.4 ± 0.2 32.8e3.3 3.2e3.6 3ns *** *
Leg II (mm) 36.6 ± 2.1 57.8 ± 0.8 535.0e40.0 57.0e59.0 5ns *** n
Fresh mass (mg) 14.7 ± 1.6 18.4 ± 1.2 112.2e16.3 16.6e20.0 1** *** *
Dry mass (mg) 6.5 ± 0.7 5.9 ± 0.5 45.3e7.2 5.2e6.4 4** ** *
Lipids/indiv. (mg) 1.5 ± 0.3 1.0 ± 0.2 01.2e1.9 0.7e1.2 0* ns n
Lipids/dry mass (mg/g) 198.3 ± 34.9 162.8 ± 38.1 9169.8e250.0 120.7e211.5 2* ns n
Water (mg/g) 559.2 ± 5.1 676.9 ± 26.5 7553.3e565.6 650.6e720.4 6ns ns n
Lipid droplet diameter (mm) 3.4 ± 0.9 2.8 ± 0.9 22.0e6.0 1.0e5.0 1*** ns *
Glycogen/dry mass (mg/g) 35.4 24.4 3
amounts of lipids in both sexes in the middle (p > 0.05) and a loweramount in females at the end of overwintering (t¼ 4.48; p < 0.001).In accordancewith the decreasing diameter of the lipid droplets theamount of lipids gradually diminishes during overwintering(Table 1; Fig. 9).
In general, females are longer and larger than males, while thelegs are longer in males (see body length, fresh and dry mass, leg II;Table 1). During the sampling period for biometrical andbiochemical analysis, individuals molted only once (leg II, Table 1).The lipid droplet diameters diminish significantly between eachtime frame in males (F2, 297 ¼ 37.94, p < 0.001) as well as in females(F2, 297 ¼ 231.61, p < 0.001). In males the lipid quantities diminishsignificantly throughout overwintering (F2, 12 ¼ 21.34, p ¼ 0.001),while in females the difference is significant between the beginningand the middle (F2, 12 ¼ 18.34, p ¼ 0.001) but not significant be-tween the middle and the end of overwintering (p ¼ 0.2808).However, the difference between sexes is significant only at thebeginning of overwintering (lipids/specimen, t ¼ 2.76, p ¼ 0.024;lipids/dry mass, t ¼ 2.85, p ¼ 0.021). Lipids gradually and signifi-cantly diminish (F2, 27 ¼ 20.48, p < 0.0001), while water ratesgradually increase during overwintering (F2, 27 ¼ 221.62,p < 0.0001) (Fig. 10).
In both sexes the glycogen content decreases from the begin-ning until the middle of overwintering, but increases at the end(Table 1).
4. Discussion
The epithelium of the midgut diverticula is constituted ofsecretory, digestive, and excretory cells (Becker and Peters, 1985).The epithelial basal lamina is lined with muscle cells, tracheae, and
ry mass, lipid quantities and rates, water rates, lipid droplet diameters and glycogen100 measurements were made for each sex and time frame. First row: Mean ± StD;e time frame: ns p > 0.05;*p < 0.05,**p < 0.01;***p < 0.001. For glycogen, only meanssentation of the biochemical analysis.
Females
nd Beginning Middle End
.2 ± 0.1 3.3 ± 0.5 4.1 ± 0.3 3.7 ± 0.4
.1e3.3 2.8e3.8 3.9e4.5 3.3e4.2
4.8 ± 1.6 39.4 ± 4.3 49.2 ± 2.6 51.2 ± 4.93.0e57.0 35.0e44.0 46.0e52.0 43.0e56.0s
6.1 ± 0.9 20.3 ± 2.5 23.3 ± 1.0 22.2 ± 2.25.2e17.4 17.5e24.1 22.5e24.8 19.1e24.9**
.6 ± 0.4 8.9 ± 1.3 7.0 ± 0.4 6.0 ± 0.9
.1e5.1 7.3e10.9 6.5e7.6 5.1e7.3*
.4 ± 0.2 2.4 ± 0.6 1.1 ± 0.3 0.8 ± 0.4
.1e0.7 1.7e3.2 0.8e1.4 0.4e1.2s
1.3 ± 49.3 263.0 ± 36.8 153.1 ± 32.7 122.5 ± 44.21.3e139.5 214.3e293.6 112.7e184.2 75.5e184.6s
16.2 ± 17.8 560.1 ± 13.9 698.5 ± 16.4 730.6 ± 18.296.4e737.2 547.7e582.9 684.4e726.9 706.8e756.9s
.3 ± 0.7 4.5 ± 1.1 2.8 ± 0.8 1.9 ± 0.5
.0e4.5 2.5e8.0 1.0e5.0 1.0e3.3**
2.0 43.9 23.0 31.4
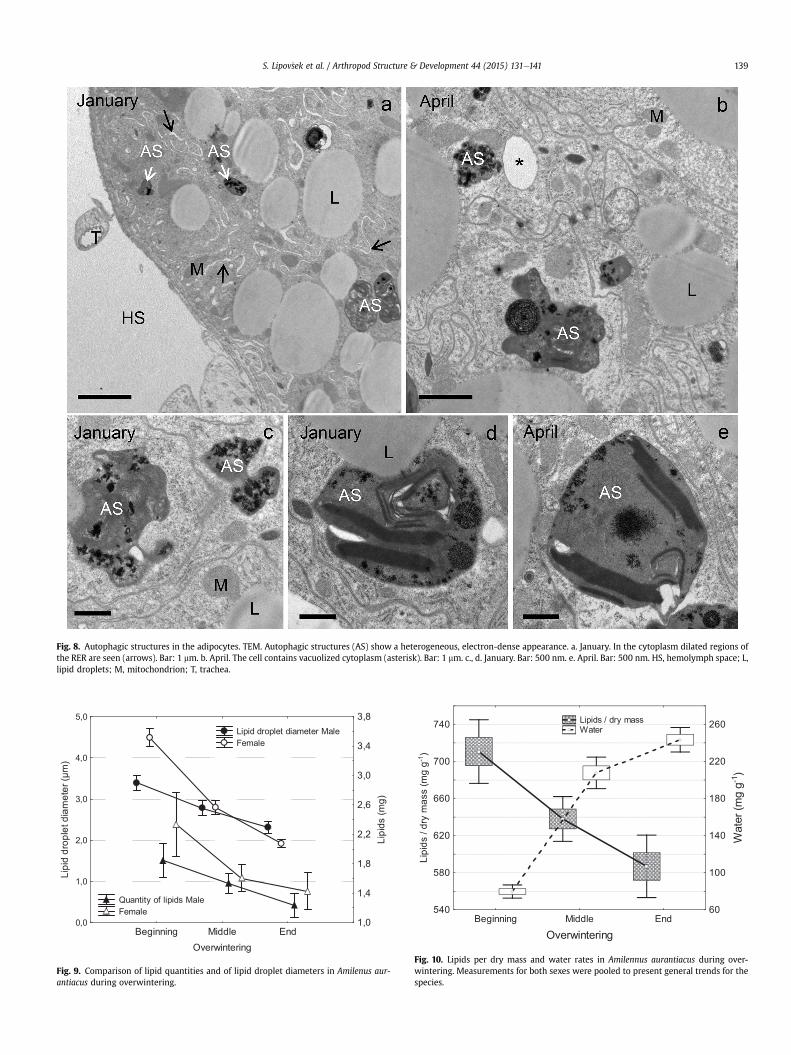
Fig. 8. Autophagic structures in the adipocytes. TEM. Autophagic structures (AS) show a heterogeneous, electron-dense appearance. a. January. In the cytoplasm dilated regions ofthe RER are seen (arrows). Bar: 1 mm. b. April. The cell contains vacuolized cytoplasm (asterisk). Bar: 1 mm. c., d. January. Bar: 500 nm. e. April. Bar: 500 nm. HS, hemolymph space; L,lipid droplets; M, mitochondrion; T, trachea.
Beginning Middle EndOverwintering
0,0
1,0
2,0
3,0
4,0
5,0
Lipi
d dr
ople
t dia
met
er (μ
m)
1,0
1,4
1,8
2,2
2,6
3,0
3,4
3,8
Lipi
ds (m
g)
Fig. 9. Comparison of lipid quantities and of lipid droplet diameters in Amilenus aur-antiacus during overwintering.
Beginning Middle End
Overwintering
540
580
620
660
700
740
Lipi
ds /
dry
mas
s (m
g g-1
)
60
100
140
180
220
260W
ater
(mg
g-1)
Lipids / dry mass Water
Fig. 10. Lipids per dry mass and water rates in Amilennus aurantiacus during over-wintering. Measurements for both sexes were pooled to present general trends for thespecies.
S. Lipov�sek et al. / Arthropod Structure & Development 44 (2015) 131e141 139

S. Lipov�sek et al. / Arthropod Structure & Development 44 (2015) 131e141140
strands of intermediate tissue (Shultz and Pinto-da-Rocha, 2007).Harvestmen are considered as primarily predaceous to omnivorousand scavenging arachnids, with a preference for carnivory, and theyfeed on solids as well as on liquids (K€astner, 1935; Juberthie, 1964;Martens, 1978; Gnaspini, 1996; Acosta and Machado, 2007;Wijnhoven, 2011). After the food enters the anterior part of theventriculus, it is transported into the spacious midgut diverticula.In harvestmen, the midgut diverticula were previously described inPhalangium opilio (Alberti and Storch, 1983; Becker and Peters,1985; Ludwig and Alberti, 1990), Gyas annulatus and Gyas titanus(Lipov�sek et al., 2004).
The midgut diverticula in the opisthosoma of A. aurantiacus aresimilar to those in other harvestmen, e.g., Phalangium opilio (Albertiand Storch, 1983; Becker and Peters, 1985; Ludwig and Alberti,1990), Gyas annulatus and Gyas titanus (Lipov�sek et al., 2004). InPh. opilio, Becker and Peters (1985) described three types of midgutdiverticula epithelial cells: digestive, secretory and excretory cells.In overwintering A. aurantiacus, all three cell types are present.While, on the one hand, the digestive cells gradually transform intoexcretory cells, on the other hand, the secretory cells form, ac-cording to their appearance, two distinctive cell assemblages fromthe middle of overwintering on. The first cell group shows astructure comparable to the one in the cells at the beginning ofoverwintering, while the second assemblage has a more lucentcytoplasm containing fewer protein granules. These differencescould be the consequence of gradual, successive, asynchronousexploitation of protein granules within separate cells. The functionof the secretory cells is to secrete enzymes and by pinocytosis totake up the predigested nutrients which undergo further digestionin the digestive cells (Becker and Peters, 1985). At the moment wehave no evidence for such a successive activation of the secretorycells in A. aurantiacus; it should be investigated in the future.
Ultrastructural differences in the midgut diverticula epithelialcells between the sexes were absent in this study, probably for twomain reasons: first, although external-morphologically distin-guishable, juveniles of both sexes, while starting a diapause, areanatomically very similar suggesting their physiological similarity.Second, during starvation, there are no conspicuous changes in thedigestive systems and the intracellular processes are not sex-dependent. Thus, such a result had been expected.
Autophagy is known to be a pro-survival process in response tostarvation (Scott et al., 2004; Chera et al., 2009; Kourtis andTavernarakis, 2009; Suzuki et al., 2011) or other stress factors, e.g.a microsporidian infection of the midgut (Rost-Roszkowska et al.,2011). The presence of autophagic structures in A. aurantiacus inall cell types after the middle of overwintering indicates that thesecells use their own compounds for maintaining their basal meta-bolism. According to their structural characteristics (Xie andKlionsky, 2007; Malagoli et al., 2010; Yang and Klionsky, 2010),the autophagic structures in A. aurantiacus have been recognized asautolysosomes. As autolysosomeswere absent at the beginning andtheir abundance gradually increased towards the end of over-wintering, this indicates their direct involvement in the processessupporting the basal metabolism.
Multivesicular bodies were present in the cytoplasm of somedigestive cells after the middle of overwintering. Besides, in someadipocytes, dilated regions of RER and individual small vacuoleswere observed in the cytoplasm. Similar vacuoles are reported fromsome columnar cells in starved cockroaches Periplaneta americana(Park et al., 2009) and in starved antlion larvae Euroleon nostras(Lipov�sek et al., 2012). In P. americana the columnar cells lost RERand exhibited many small vacuoles, probably degenerated RER(Park et al., 2009). Materials originating from the exocytotic vesiclesand discharged into the lumen of the midgut diverticula constitutepart of an overwintering fecal pellet which in A. aurantiacus is also
probably expelled after the diapause termination. Since these pel-lets differ from the postprandial ones, their chemical analysis couldprovide important further insight into the digestive and autophagicprocesses during overwintering.
In overwintering A. aurantiacus, lipids and glycogen are presentin the adipocytes, the secretory cells and the digestive cells of themidgut diverticula. Gradual diminution of the lipid droplets in theadipocytes in both sexes indicates the constant exploitation oflipids during this starvation period. The more intense use of lipidsin females reflects their more intense metabolism, which is relatedto the ripening of the ovaries during the diapause.
Comparison of the results of measurement of the lipid dropletdiameters, on the one hand, and the lipid quantities, on the other,clearly denotes a tight correlation between these two methods.Both analyses are thus of comparable value; either one is relevantto evaluation of the lipid metabolism in A. aurantiacus duringoverwintering. This is probably true in most arthropod species thatbase their overwintering metabolism on lipid storage. On the otherhand, conversion of glycogen to glycerol is reversible and temper-ature dependent, so that as insects warm in the spring, much of theglycerol is recovered and stored again as glycogen (Hahn andDenlinger, 2011). The increasing glycogen levels at the end ofoverwintering thus show that this happens in A. aurantiacus beforetheir diapause termination and leaving the caves.
Acknowledgments
We would like to thank Elisabeth Bock und Rudi Schmied(Medical University Graz) for their excellent technical assistance.Michelle Gadpaille valuably improved the English of the manu-script. We are indebted to the Editor and to the Reviewers for theirinsightful suggestions and comments. This study was partly sup-ported by the Slovenian Research Agency within the BiodiversityResearch Programme (Grant No. P1-0078) and by grant “Infra-strukturni program” University of Maribor (IP-0552).
References
Acosta, L.E., Machado, G., 2007. Diet and foraging. In: Pinto-da-Rocha, R.,Machado, G., Giribet, G. (Eds.), Harvestmen. The Biology of Opiliones,pp. 309e338.
Alberti, G., Storch, V., 1983. Zur Ultrastruktur der Mitteldarmdrüsen von Spinnen-tieren (Scorpiones, Araneae, Acari) unter verschiedenen Ern€ahrungsbedinungen.Zool. Anz. 11, 145e160.
Becker, A., Peters, W., 1985. Fine structure of the midgut gland of Phalangium opilio(Chelicerata, Phalangida). Zoomorphology 105, 317e325.
Bourne, J.D., 1978a. Donn�ees pr�eliminaires sur l’�ecologie et la biologie de l'opiliontroglox�ene Amilenus aurantiacus (Simon). In: Actes 6e Congr�es Suisse deSp�el�eologie. Porrentruy, pp. 17e24.
Bourne, J.D., 1978b. Observations pr�eliminaires sur l’intestin de l’opilion troglox�eneAmilenus aurantiacus (Simon) pendant son sejour souterrain. M�emoires Bio-sp�eol. 8, 39e42.
Chera, S., Buzgariu, W., Ghila, L., Galliot, B., 2009. Autophagy in Hydra: a response tostarvation and stress in early animal evolution. Biochim. Biophys. Acta 1793,1432e1443.
Gnaspini, P., 1996. Population ecology of Goniosoma spelaeum, a caverniculousharvestman from southeastern Brazil (Arachnida: Opiliones, Gonyleptidae).J. Zool. 239, 417e435.
Hahn, D.A., Denlinger, D.L., 2011. Energetics of insect diapause. Annu. Rev. Entomol.56, 103e121.
Jequier, J.-P., 1964. �Etude �ecologique et statistique de la faune terrestre d'une cav-erne du Jura Suisse au cours d'une ann�ee d'observations. Rev. Suisse Zool. 71,13e370.
Juberthie, C., 1964. Recherches sur la biologie des Opilions. Ann. Splol. 19, 1e238.K€astner, A., 1935. Die Funktion der sogenannten sympathischen Ganglien und die
Exkretion bei den Phalangiiden. Zool. Anz. 109, 273e288.Kates, M., 1991. Techniques in Lipidology. Elsevier, Amsterdam.Kourtis, N., Tavernarakis, N., 2009. Autophagy and cell death in model organisms.
Cell Death Differ. 16, 21e30.Lipov�sek, S., Novak, T., Jan�zekovi�c, F., Sen�ci�c, L., Pabst, M.A., 2004. A contribution to
the functional morphology of the midgut gland in phalangiid harvestmen Gyasannulatus and Gyas titanus during their life cycle. Tissue Cell 36, 275e282.

S. Lipov�sek et al. / Arthropod Structure & Development 44 (2015) 131e141 141
Lipov�sek, S., Letofsky-Papst, I., Hofer, F., Leitinger, G., Devetak, D., 2012. The evi-dence on the degradation processes in the midgut epithelial cells of the larvalantlion Euroleon nostras (Geoffroy in Fourcroy, 1785) (Myrmeleontidae, Neu-roptera). Micron 43, 651e665.
Ludwig, M., Alberti, G., 1990. Peculiarities of arachnid midgut glands. Acta Zool.Fenica 190, 255e259.
Malagoli, D., Abdalla, F.C., Cao, Y., Feng, Q., Fujisaki, K., Gregorc, A., Matsuo, T.,Nezis, I.P., Papassideri, I.S., Sass, M., Silva-Zacarin, E.C.M., Tettamanti, G., Ume-miya-Shirafuji, R., 2010. Autophagy and its physiological relevance in arthro-pods: current knowledge and perspectives (Review). Autophagy 6, 575e588.
Martens, J., 1969. Systematische Stellung von Amilenus aurantiacus (Simon) (Opi-liones, Phalangiidae). Senckenberg. Biol. 50, 219e224.
Martens, J., 1978. Weberknechte, Opiliones. Die Tierwelt Deutschlands 64 (FischerVerlag).
Novak, T., Gruber, J., Slana, L., 1984. Remarks on Opiliones from cavities in Slovenia(Yugoslavia). M�emoires Biospeleol. 11, 185e197.
Novak, T., Perc, M., Lipov�sek, S., Jan�zekovi�c, F., 2012. Duality of terrestrial subter-ranean fauna. Int. J. Speleol. 41, 181e188. http://dx.doi.org/10.5038/1827-806X.41.2.5.
Novak, T., Jan�zekovi�c, F., Lipov�sek, S., 2013. Contribution of non-troglobioticterrestrial invertebrates to carbon input in hypogean habitats. Acta Carsol. 42,301e309.
Park, M.S., Park, P., Takeda, M., 2009. Starvation induces apoptosis in the midgutnidi of Periplaneta americana: a histochemical and ultrastructural study. CellTissue Res. 335, 631e638.
Plummer, D.T., 1987. An Introduction to Practical Biochemistry. McGraw-Hill, London.Rost-Roszkowska, M.M., Poprawa, I., Kaczmarek, L., 2011. Autophagy as the cell
survival in response to a microsporidian infection of the midgut epithelium ofIsohypsibius granulifer granulifer (Eutardigrada: Hypsibiidae). Acta Zool. http://dx.doi.org/10.1111/j.1463-6395.2011.00552.x.
Scott, R.C., Schuldiner, O., Neufeld, T.P., 2004. Role and regulation of starvation-induced autophagy in the Drosophila fat body. Dev. Cell 7, 167e178.
Shultz, J.W., Pinto-da-Rocha, R., 2007. Morphology and functional anatomy. In:Pinto-da-Rocha, R., Machado, G., Giribet, G. (Eds.), Harvestmen. The Biology ofOpiliones, pp. 14e61.
Suzuki, S.W., Onodera, J., Ohsumi, Y., 2011. Starvation induced cell death inautophagy-defective yeast mutants is caused by mitochondria dysfunction.PLoS ONE 6, e17412.
Wijnhoven, H., 2011. Notes on the biology of the unidentified invasive harvestmanLeiobunum sp. (Arachnida: Opiliones). Arachnol. Mittl. 41, 17e30.
Xie, Z., Klionsky, D.J., 2007. Autophagosome formation: core machinery and adap-tations. Nat. Cell Biol. 9, 1102e1109.
Yang, Z., Klionsky, D.J., 2010. Eaten alive: a history of macroautophagy. Nat. Cell Biol.12, 814e822.
Related Documents











