EQUINE EXERCISE PHYSIOLOGY 6 Equine vet. J., Suppl. 34 (2002) 199-204 199 Summary This study examined changes in skeletal muscle GLUT4 content glucose transport in isolated muscle membranes (GT) from horses before and 2 min after standardised submaximal exercise tests (SET) prior to and after completion of 6 weeks of training. Seven horses, age 3–9 years, body mass mean ± s.e. 530 ± 19 kg, and sedentary for at least 4 months, completed 6 weeks of training on a treadmill. An initial SET (UT) was performed on a 4° incline at a speed equivalent to 55% of pretraining VO 2max and was repeated post-training at the same absolute workload (ABS). A third SET (REL) was performed at 55% of post-training VO 2max . There was no significant pre- to postexercise change in GLUT4 content before or after training. Following training, total GLUT4 content was increased 2- or 3-fold in pre-exercise biopsies (pre UT: 0.30 ± 0.05; pre ABS: 1.05 ± 0.32; pre REL: 1.34 ± 0.28 arbitrary units) (P<0.05) with similar increases in postexercise GLUT4 content (P<0.05) (post UT: 0.33 ± 0.06; post ABS: 1.19 ± 0.44; post REL: 1.43 ± 0.31). GT increased 2.5- to 6-fold in postexercise muscle membrane vesicles in UT over a range of glucose concentrations. After training (ABS and REL), there was a 25–50% attenuation (P<0.05) in membrane GT in response to exercise in ABS and REL. These finding indicate that moderate intensity exercise training increased middle gluteal muscle GLUT4 content, but this change was not reflected in an increase in muscle membrane glucose transport activity in postexercise muscle samples. Introduction During exercise, skeletal muscle glucose utilisation reflects the combined effect of glucose supply, glucose transport capacity and glucose metabolism. Regardless of training state, maintenance of glucose supply to working skeletal muscle (from muscle glycogen and plasma glucose) is critical to sustain work output during moderately intense exercise (>60–65% VO 2max , in human subjects) and, when hepatic glucose output fails to match the demands of working muscle, the decline in plasma glucose supply contributes to the onset of fatigue. Exercise training influences the pattern of substrate utilisation during exercise with one of the main adaptations being a decrease in glucose utilisation. In human athletes, this decrease has been attributed to a reduction in glucose transport activity (Kristiansen et al. 1998). A crucial and often rate-limiting step in glucose metabolism in skeletal muscle is the ability to transport glucose across the plasma membrane into the cell (DeFronzo et al. 1981; Richter et al. 2001). Such transport is facilitated by a family of glucose transporter proteins, of which GLUT4 and GLUT1 have been identified as the isoforms of importance in skeletal muscle. GLUT1 is responsible for basal glucose transport independent of insulin and is primarily associated with the plasma membrane. By comparison, the GLUT4 isoform is insulin-responsive and is translocated from intracellular endosomes either by a complex signalling pathway triggered by insulin or by an undefined pathway triggered by exercise (Marette et al. 1992). In rats and man, GLUT4 content is increased with physical conditioning and there is a concomitant increase in insulin-stimulated glucose transport activity (Hirshman et al. 1993) suggesting a mechanism for improved glucose clearance in the trained state. However, evidence of a post-training reduction in glucose utilisation during submaximal exercise indicates that more information is needed to understand the link between the content of GLUT4 in the muscle and its intrinsic activity. In horses, there are few data pertaining to the effects of exercise training on regulation of glucose transport into muscle during submaximal exercise. It is not known whether training results in a reduction in the utilisation of, or increased reliance on, plasma glucose for maintenance of carbohydrate supply to working muscle. An increase in glucose transporter protein content and/or glucose uptake could improve the availability of glucose to the muscle cell. Therefore, the objective of this study was to determine the effects of moderate intensity exercise training on mechanisms that affect glucose transport in skeletal muscle during exercise. Specifically, our objective was to determine the effects of a 6 week period of exercise training in horses on skeletal muscle GLUT4 content and muscle membrane glucose transport capacity in samples of middle gluteal muscle obtained before and after exercise in the untrained and trained state. Our hypothesis was that a 6 week period of exercise training would result in increases in skeletal muscle GLUT4 protein content and muscle membrane glucose transport capacity. As training results in an increase in maximum aerobic capacity, the pattern of fuel utilisation and, as a result, the expectations for glucose uptake could be altered in the trained state. Therefore, in this study, post-training testing was performed at the same relative and same absolute workload. Changes in skeletal muscle GLUT4 content and muscle membrane glucose transport following 6 weeks of exercise training L. J. MCCUTCHEON*, R. J. GEOR † and K. W. HINCHCLIFF † Department of Pathobiology, Ontario Veterinary College, University of Guelph, Guelph, Ontario N1G 2W1, Canada and † Department of Veterinary Clinical Sciences, College of Veterinary Medicine, The Ohio State University, Columbus, Ohio, USA. Keywords: horse; glucoregulation; insulin; glycogen *Author to whom correspondence should be addressed.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

EQUINE EXERCISE PHYSIOLOGY 6Equine vet. J., Suppl. 34 (2002) 199-204
199
Summary
This study examined changes in skeletal muscle GLUT4content glucose transport in isolated muscle membranes (GT)from horses before and 2 min after standardised submaximalexercise tests (SET) prior to and after completion of 6 weeksof training. Seven horses, age 3–9 years, body mass mean ±s.e. 530 ± 19 kg, and sedentary for at least 4 months,completed 6 weeks of training on a treadmill. An initial SET(UT) was performed on a 4° incline at a speed equivalent to55% of pretraining VO2max and was repeated post-training atthe same absolute workload (ABS). A third SET (REL) wasperformed at 55% of post-training VO2max. There was nosignificant pre- to postexercise change in GLUT4 contentbefore or after training. Following training, total GLUT4content was increased 2- or 3-fold in pre-exercise biopsies(pre UT: 0.30 ± 0.05; pre ABS: 1.05 ± 0.32; pre REL: 1.34 ±0.28 arbitrary units) (P<0.05) with similar increases inpostexercise GLUT4 content (P<0.05) (post UT: 0.33 ± 0.06;post ABS: 1.19 ± 0.44; post REL: 1.43 ± 0.31). GT increased2.5- to 6-fold in postexercise muscle membrane vesicles in UTover a range of glucose concentrations. After training (ABSand REL), there was a 25–50% attenuation (P<0.05) inmembrane GT in response to exercise in ABS and REL. Thesefinding indicate that moderate intensity exercise trainingincreased middle gluteal muscle GLUT4 content, but thischange was not reflected in an increase in muscle membraneglucose transport activity in postexercise muscle samples.
Introduction
During exercise, skeletal muscle glucose utilisation reflects thecombined effect of glucose supply, glucose transport capacityand glucose metabolism. Regardless of training state,maintenance of glucose supply to working skeletal muscle (frommuscle glycogen and plasma glucose) is critical to sustain workoutput during moderately intense exercise (>60–65% VO2max, inhuman subjects) and, when hepatic glucose output fails to matchthe demands of working muscle, the decline in plasma glucosesupply contributes to the onset of fatigue. Exercise traininginfluences the pattern of substrate utilisation during exercisewith one of the main adaptations being a decrease in glucose
utilisation. In human athletes, this decrease has been attributed toa reduction in glucose transport activity (Kristiansen et al. 1998).
A crucial and often rate-limiting step in glucose metabolismin skeletal muscle is the ability to transport glucose across theplasma membrane into the cell (DeFronzo et al. 1981; Richter etal. 2001). Such transport is facilitated by a family of glucosetransporter proteins, of which GLUT4 and GLUT1 have beenidentified as the isoforms of importance in skeletal muscle.GLUT1 is responsible for basal glucose transport independent ofinsulin and is primarily associated with the plasma membrane.By comparison, the GLUT4 isoform is insulin-responsive and istranslocated from intracellular endosomes either by a complexsignalling pathway triggered by insulin or by an undefinedpathway triggered by exercise (Marette et al. 1992). In rats andman, GLUT4 content is increased with physical conditioningand there is a concomitant increase in insulin-stimulated glucosetransport activity (Hirshman et al. 1993) suggesting amechanism for improved glucose clearance in the trained state.However, evidence of a post-training reduction in glucoseutilisation during submaximal exercise indicates that moreinformation is needed to understand the link between the contentof GLUT4 in the muscle and its intrinsic activity.
In horses, there are few data pertaining to the effects of exercisetraining on regulation of glucose transport into muscle duringsubmaximal exercise. It is not known whether training results in areduction in the utilisation of, or increased reliance on, plasmaglucose for maintenance of carbohydrate supply to workingmuscle. An increase in glucose transporter protein content and/orglucose uptake could improve the availability of glucose to themuscle cell. Therefore, the objective of this study was to determinethe effects of moderate intensity exercise training on mechanismsthat affect glucose transport in skeletal muscle during exercise.Specifically, our objective was to determine the effects of a 6 weekperiod of exercise training in horses on skeletal muscle GLUT4content and muscle membrane glucose transport capacity insamples of middle gluteal muscle obtained before and afterexercise in the untrained and trained state. Our hypothesis was thata 6 week period of exercise training would result in increases inskeletal muscle GLUT4 protein content and muscle membraneglucose transport capacity. As training results in an increase inmaximum aerobic capacity, the pattern of fuel utilisation and, as aresult, the expectations for glucose uptake could be altered in thetrained state. Therefore, in this study, post-training testing wasperformed at the same relative and same absolute workload.
Changes in skeletal muscle GLUT4 content and musclemembrane glucose transport following 6 weeks ofexercise trainingL. J. MCCUTCHEON*, R. J. GEOR† and K. W. HINCHCLIFF†
Department of Pathobiology, Ontario Veterinary College, University of Guelph, Guelph, Ontario N1G 2W1,Canada and †Department of Veterinary Clinical Sciences, College of Veterinary Medicine, The Ohio StateUniversity, Columbus, Ohio, USA.
Keywords: horse; glucoregulation; insulin; glycogen
*Author to whom correspondence should be addressed.

200 Changes in muscle GLUT-4 content following training
Materials and methods
All animal experiments were conducted after approval by theInstitutional Laboratory Animal Care and Use Committee of TheOhio State University and were performed in compliance withtheir guidelines and recommendations.
Horses
The subjects were 7 horses (2 Standardbred and 5Thoroughbred; all geldings), age 3–9 years and mean ± s.e. bodymass 530 ± 19.1 kg (range 441–589 kg). Prior to this study,horses had been paddock-rested for a period of at least 6 months.During the study, the horses were housed indoors and fed a dietof timothy grass/alfalfa hay and a pelleted concentrate ration(Bridle Bits 1200)1 (4 kg/day), had access to a salt/mineral blockand received treadmill exercise only.
Experimental design
The investigation was designed as a longitudinal study duringwhich all horses undertook a standardised exercise test (SET)prior to and following 6 weeks of training (see Training). Beforethe experiment, there was an initial 5 day period of acclimatisationto the treadmill consisting of short (3–5 min) walking and trottingsessions. Horses were also acclimated to a loose-fitting facemaskfor measurement of respiratory gas exchange. Each horsecompleted an incremental exercise test (IET) for determination ofmaximum aerobic capacity (VO2max), 5 days before thecommencement of training. Three days prior to training, horsesperformed an SET at 55% of VO2max (UT). Upon completion oftraining, the horses completed a second IET. Followingcompletion of training, and in random order, the horses performedSETs at 55% of pretraining VO2max (ABS) and at 55% of post-training VO2max (REL), corresponding to the same absolute andrelative workload before and after training, respectively. The post-training SETs were separated by 3 days. All SETs and trainingwere undertaken on a treadmill set at a 4° incline.
Incremental exercise tests
For each horse, the maximal oxygen consumption (VO2max) andthe relationship between VO2 and speed were determined duringan incremental exercise test (IET) 5 days before the firstexperiment and 2 days following the 6 week training period.Each IET consisted of the horse running on a high speedtreadmill2 inclined at 4° for 90 s at 4 m/s, after which thetreadmill speed was increased by 1 m/s every 90 s until the horsewas no longer able to maintain its position on the treadmill. VO2was measured every 10 s during the exercise test. VO2max wasdefined as the value at which VO2 reached a plateau, despitefurther increases in speed. A plateau was defined as a change inVO2 of <4 ml/kg/min with an increase in speed. From linearregression analysis (speeds below VO2max), the running speedthat elicited 55% of VO2max was calculated for each horse.
Training regimen
Training consisted of 25 min of treadmill exercise 5 days/weekfor 6 weeks, with weekly increases in the intensity of exercise. InWeek 1, horses completed 1 min at a walk (1.8 m/s), 2 min at aslow trot (3 m/s) followed by 20 min at 55% of pretrainingVO2max (treadmill belt speed 4.2–5.8 m/s) and a final 2 min at awalk. During Weeks 2 to 6 of training, horses also ran for 3 min
at 75% of pretraining VO2max (6.0–8.0 m/s) with a correspondingdecrease in the duration of exercise at 55% of pretraining VO2max.
Standardised submaximal exercise test
The horses were fasted for 12 h before each experiment and hadbeen confined to their stalls during this period. Thereafter, thehorses were positioned on the treadmill (4° incline) and a loose-fitting facemask for measurement of respiratory gas exchangewas applied. The horses completed a 5 min warm-up (3 m/streadmill belt speed), followed by 60 min running at a speedcalculated to elicit 55% VO2max for 60 min or until developmentof fatigue, as determined by an inability to keep pace with thetreadmill despite verbal encouragement. During the exercise test,fans mounted 0.5 m in front and to the sides of the treadmill wereused to maintain an air velocity of 3.5–4.0 m/s over the horse.Ambient conditions were similar for all trials; mean ± s.e. valuesfor room temperature and relative humidity during theexperiments were 18.3 ± 0.2°C and 44 ± 2%, respectively.
Respiratory gas exchange measurements
Rates of oxygen consumption (VO2) and carbon dioxide production(VCO2), were measured with an open-circuit calorimeter (Oxymax-XL)3, as described previously (Hinchcliff et al. 1993). Flowthrough the system was ~1500 l/min STP with the horse stationaryand 9000 l/min during running. The gas analysis system wascalibrated before the start of each exercise test using gas mixtureswith O2 and CO2 concentrations that spanned the measurementrange. The overall accuracy of the system was verified repeatedlyby the nitrogen dilution method (Fedak et al. 1981).
Sampling procedures
A 5 cm square area of skin over each middle gluteal muscle wasclipped and shaved. After aseptic preparation and localanaesthesia of the overlying skin, a sample of middle glutealmuscle was obtained by percutaneous biopsy at a depth of 4 cmusing the needle biopsy technique 5 min before commencementof exercise and within 2 min of cessation of exercise. Musclesamples (1–1.5 g) were placed immediately in liquid nitrogenand stored at -80°C until analysis.
Analytical techniques
Muscle GLUT4 content: Measurement of glucose tranporterprotein to assess GLUT4 content in skeletal muscle wasdetermined as described by Ploug et al. (1993) withmodifications as described by Johannsson et al. (1996). Briefly,frozen muscle (50–80 mg) was homogenised in buffer [210mmol/l sucrose, 2 mmol/l ethylene glycol-bis(β-aminoethylether)-N,N,N’,N’-tetraacetic acid, 40 mmol/l NaCl,30 mmol/l N-2-hydrozyethylpiperazine-N’-2-ethanesulfonicacid, pH 7.4, and 2 mmol/l phenylmethylsulphonyl fluoride]and was mixed with 3 ml KCl (1.167 mol/l) and pyrophosphate(58.3 mmol/l), centrifuged (50,000 g for 75 min at 4°C) and lefton ice for 15 min. Supernatant was removed and the pelletresuspended and homogenised (10 min) in 600 µl buffer, afterwhich 200 µl 16% sodium dodecyl sulphate (SDS) was added,samples were centrifuged (3000 g) 15 min and the supernatantrecovered for protein determination and electrophoresis.GLUT4 was determined using Western blotting procedures asdescribed previously (Megeny et al. 1993, 1994; Johannsson etal. 1996). Briefly, protein (50 µg) was mixed with 62.5 µl
L. J. McCutcheon et al. 201
Laemmli sample buffer containing 2.5% dithiothreitol andbrought to 125 ml with buffer containing 25 mmol/l tris(hydrozymethyl) aminomethane (Tris, pH 8.3), 0.19 mmol/lglycine and 1% sodium dodecyl sulphate (SDS). Samples wereseparated by SDS-polyacrylamide electrophoresis on a 12%resolving gel and transferred to an Immobilon membrane4 byelectromembrane transfer for 90 min. GLUT4 protein wasdetected by incubation with a polyclonal immuno-A purifiedantibody5 followed by incubation with a horseradishperoxidase-labelled second anti-rabbit immunoglobulin G6. Theantibody recognises the 12 amino acid sequence in the carboxyterminus of GLUT4, an area conserved across several speciesincluding the rat, rabbit and man. After the measurement ofGLUT4, the membrane was stripped with buffer (100 mmol/l 2-mercaptoethanol, 12% SDS, 62.5 mmol/l Tris.HCl) for 30 minat 50°C. GLUT4 protein was visualised using an enhancedchemiluminescence detection system5 according to theinstructions of the manufacturer. Autoradiographs of thewestern blots were quantified using scanning densitometry andappropriate software (Model GS300 with GS370 software)7.The GLUT4 protein/µg protein was expressed in arbitrary unitsrelative to a rat heart standard.
Muscle membrane glucose uptake: Glucose uptake was measuredin muscle membrane vesicles from skeletal muscle by thetechnique described by Burton et al. (1988), modified by Ploug etal. (1993) and further modified by Richter et al. (1998). Briefly,frozen muscle (50–80 mg) was homogenised in buffer asdescribed above and total membranes were recovered bycentrifugation (256,000 g). Glucose transport was measured atroom temperature. In a 1.5 µl Eppendorf tube, 30 µl of membranevesicle solution was mixed with 10 µl of a solution containing 0.5 µCi d-[3H]glucose, 0.1 µCi L-[14C]glucose and severalconcentrations of cold D-glucose with the mannitol concentrationbeing varied reciprocally such that the total concentration ofsugars was maintained at 40 mmol/l. Glucose uptakemeasurements were performed at glucose concentrations of 0, 5,10, 25 and 50 mmol/l. After mixing, entry of glucose was stoppedat one of a series of designated time points between 0 and 120 sby addition of 1 ml chilled stop solution (200 µmol/l phloretin inKCl-HEPES buffer). The stopped mixture was spun for 1 minusing a high speed centrifuge, after which the supernatant wasaspirated and the remaining pellet was mixed with 2.5 mlscintillation fluid. For each measurement, D-glucose uptake wascalculated by subtracting L-glucose from total D-glucose uptake(Ploug et al. 1992). Ethanol alone (0.5% final concentration) wasadded in control experiments and cytochalasin B (in 50% ethanolsolution; final concentration 20 µmol/l cytochalasin B and 0.5%ethanol) was used to inhibit specific D-glucose transport. Alldeterminations were performed in triplicate.
Statistical analyses
Data are presented as means ± s.e. The data were analysed by useof a 2-way repeated measures analysis of variance, withtreatment (UT, ABS, REL) and time as independent factors. Apaired Student’s t test was used to compare pre- and post-training VO2max. The null hypothesis was rejected at α = 0.05 forthe main effects (time and treatment). Significant differencesidentified by ANOVA were isolated using the Student-Newman-Keul’s post hoc test. The Sigmastat 2.0 software package8 wasused for statistical computations.
Results
One horse did not complete the REL trial due to lameness and,therefore, data for REL are expressed as means ± s.e. for 6 horses.Mean bodyweight decreased (P<0.05) after 6 weeks of training(pretraining 530 ± 19 kg; post-training 512 ± 16 kg). There was a14.9% increase (P<0.001) in mass-specific VO2max (pretraining118.4 ± 7.4 ml/kg/min; post-training 136.1 ± 7.8 ml/kg/min); inabsolute terms this change represented a 10.9% increase in VO2max(62.3 ± 3.2 l/min vs. 69.1 ± 3.1 l/min). In UT and ABS, meanrunning speed was 5.1 ± 2 m/s (range 4.3–6.0 m/s), representing57.0 ± 1.1% of pretraining VO2max. With an increase in aerobiccapacity, the relative workload during ABS represented only 49.5 ± 0.8% of post-training VO2max. Mean running speed in RELwas 5.9 ± 0.2 m/s (range 5.4–6.8 m/s), corresponding to a relativeworkload of 57.1 ± 1.2% VO2max. Mean exercise duration waslonger (P<0.05) in ABS (57 ± 1.9 min) than in UT (46 ± 3.9 min)and REL (49 ± 4.6 min). One, 3 and 5 horses completed 60 min ofexercise in UT, REL and ABS, respectively.
GLUT4 content
Pre- and postexercise skeletal muscle GLUT4 protein contentwas increased 2–3-fold following training (UT: pre 0.30 ± 0.05and post 0.33 ± 0.06; ABS: pre 1.05 ± 0.32 and post 1.19 ± 0.44;
Fig 1: Concentration of GLUT4 expressed in arbitrary units/mg proteinhomogenate from gluteus medius biopsies obtained from 7 horses before(Pre) and after (Post) approximately 60 min of exercise prior to (UT)and following 6 weeks of exercise training (ABS and REL). Exercise wasconducted at 55% of pretraining VO2max in UT and ABS and at 55% ofpost-training VO2max in REL. Values are mean ± s.e. *Significantlydifferent from UT (P<0.05).
GLU
T-4
cont
ent
(arb
itrar
y un
its)
2.0
1.5
1.0
0.5
0.0UT ABS REL
RestExercise
*
* **
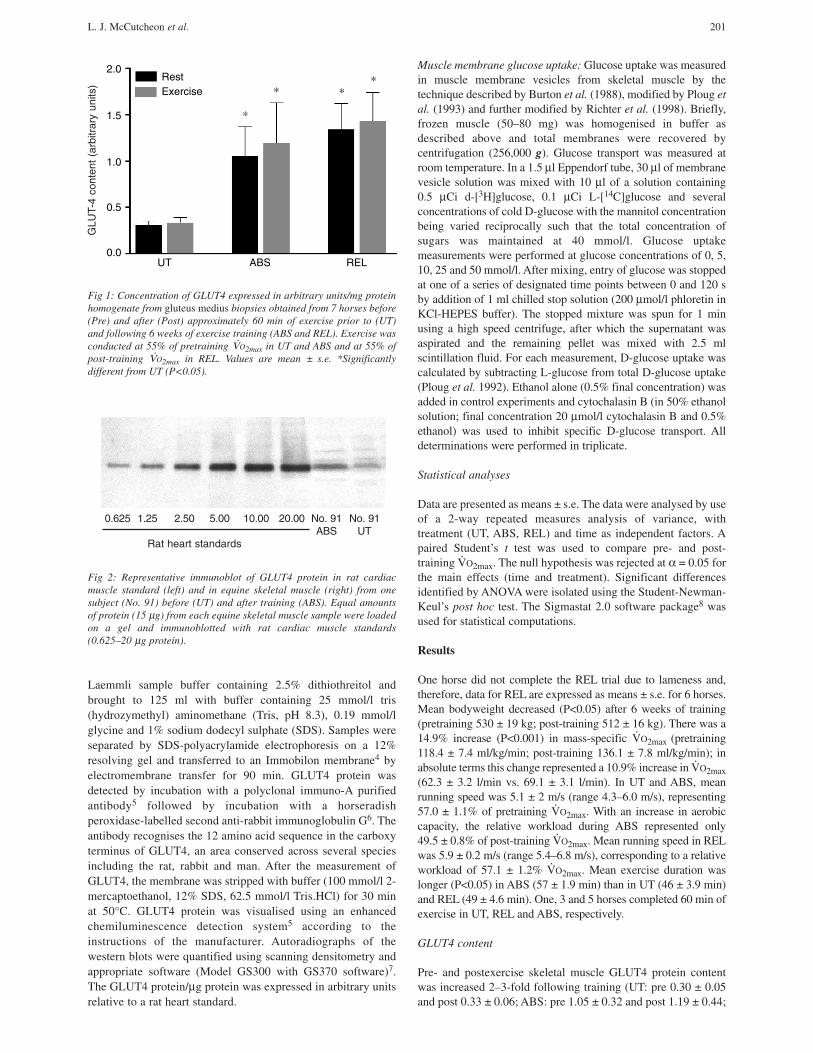
Fig 2: Representative immunoblot of GLUT4 protein in rat cardiacmuscle standard (left) and in equine skeletal muscle (right) from onesubject (No. 91) before (UT) and after training (ABS). Equal amountsof protein (15 µg) from each equine skeletal muscle sample were loadedon a gel and immunoblotted with rat cardiac muscle standards(0.625–20 µg protein).
0.625 1.25 2.50 5.00 10.00 20.00 No. 91 No. 91ABS UT
Rat heart standards
202 Changes in muscle GLUT-4 content following training
REL: pre 1.34 ± 0.28 and post 1.43 ± 0.31 arbitrary units)(P<0.05; Fig 1) with no significant difference in GLUT4 proteincontent evident (P>0.05) between ABS and REL post-training.Before or after training, there was no change (P>0.05) in GLUT4protein content with exercise.
Glucose transport
Glucose transport capacity, as determined by glucose uptake inmuscle membrane vesicles, was measured over a range ofglucose concentrations from 0 to 50 mmol/l. Values for glucosetransport at 0 mmol/l were not different from controlexperiments. In all samples, uptake was linear over the first 25 s
after which it became curvilinear. Initial rate of glucose uptakeby vesicles (measured as the 25 s uptake) at 5 and 10 mmol/lglucose concentration, was increased 3- to 5-fold in UT whencomparing pre- and postexercise muscle membrane samples(P<0.05; Figs 2, 3) with similar increases at 25 and 50 mmol/lconcentrations. After training (ABS and REL), the pre- topostexercise increase in glucose uptake was reduced 25–50%over the same range of glucose concentrations (P<0.05) whencompared to UT.
Discussion
The present study examined changes in equine skeletal muscleGLUT4 protein content and glucose transport in musclemembrane vesicles associated with 6 weeks of exercise trainingand a 60 min standardised submaximal treadmill exercise test.The main findings of the study were: 1) exercise training resultedin a 2–3-fold increase in whole muscle GLUT4 protein content inpre-exercise middle gluteal muscle samples; 2) regardless oftraining state, postexercise GLUT4 content did not differ fromvalues measured in pre-exercise samples; 3) postexercise rate ofglucose transport into muscle membrane vesicles was increasedover a range of glucose concentrations and 4) the extent of thepostexercise increase in glucose transport capacity, as measuredby rate of glucose uptake into muscle membrane vesicles, wasblunted following exercise training.
The 10.9% increase in maximal aerobic capacity for thehorses in this study demonstrates a training effect consistent withprevious studies in which horses have undertaken daily exerciseat 40–75% of their VO2max for a period of 2–8 weeks (Knight etal. 1991; Geor et al. 1999). The improved aerobic capacity of thehorses in the trained state is supported by the increase in meanexercise duration (22%) in ABS, with a higher number ofsubjects able to complete 60 min exercise.
In this study, the GLUT4 content measured in musclehomogenate from samples obtained after 6 weeks of training wasincreased when compared to samples acquired prior to training.The findings are similar to those of Kristiansen et al. (2000) whonoted a 66% increase in GLUT4 in human subjects after 3 weekstraining. Glucose transport capacity assayed in muscle membranevesicles derived from these samples increased in samples obtainedwithin 2 min after exercise, compared to samples collected priorto exercise. However, contrary to our hypothesis, the extent of thepostexercise glucose transport capacity as assayed in these musclemembrane vesicles was reduced in samples obtained from horsesin the trained compared to the untrained state. Therefore, despitethe increase in GLUT4 protein content of whole muscle withtraining, the increase in glucose transport capacity of the isolatedmuscle cell membranes from rest to 2 min postexercise was not asgreat in the trained vs. the untrained state during moderate-intensity exercise. It has been suggested that the energy status ofthe muscle, as well as the size of the GLUT4 pool, determines theextent of glucose uptake following exercise (Ihlemann et al. 1999;Kristiansen et al. 2000).
Studies in rats have indicated that 4 days of swim-trainingwere sufficient to induce up to a 100% increase in skeletalmuscle GLUT4 content and in vitro insulin-stimulated glucosetransport (Ren et al. 1994; Reynolds et al. 1997). More recentevidence suggests that the increase in GLUT4 content closelyreflects the intensity of exercise performed, with as little as 14 x20 s bouts of exercise resulting in an increase comparable to thatattained after 360 min/day for 8 days (Terada et al. 2001).Conversely, Host et al. (1998) have demonstrated that increasesin GLUT4 protein and in insulin-stimulated glucose transportwere completely reversed within 40 h after the last exercise bout,
Fig 3: Initial rate of D-glucose-specific uptake into muscle membranevesicles measured at 5 mmol/l D-glucose. Muscle membrane vesicleswere prepared from gluteus medius biopsies obtained from horses before(Pre) and after (Post) approximately 60 min of exercise prior to (UT)and following 6 weeks of exercise training (ABS and REL). Exercise wasconducted at 55% of pretraining VO2max in UT and ABS and at 55% ofpost-training VO2max in REL. Values are mean ± s.e. *Significantlydifferent from Pre; #significantly different from Pre and from Post (ABSand REL) (P<0.05).
Glu
cose
tra
nspo
rt (
pmol
/µg/
min
)
100
75
50
25
0Pre Post
UT
ABS
REL
#*
*
*
Fig 4: Effect of training on postexercise D-glucose-specific uptake intomuscle membrane vesicles measured at 0, 5, 10, 25 and 50 mmol/l of D-glucose concentration. Vesicles were prepared from gluteus mediusbiopsies obtained from horses after approximately 60 min exercise priorto (UT) and following 6 weeks exercise training (ABS and REL).Exercise was conducted at 55% of pretraining VO2max in UT and ABSand at 55% of post-training VO2max in REL. Values are mean ± s.e.*Significantly different from UT (P<0.05)
300
250
200
150
100
50
0
Glu
cose
upt
ake
(pm
ol/µ
g/m
in)
0 10 20 30 40 50Glucose concentration (mmol/l)
UT
ABS
REL
*
*
*
*

L. J. McCutcheon et al. 203
suggesting a short half-life of the GLUT4 protein. In the presentstudy, the increases in muscle GLUT4 content after 6 weeks oftraining for 25 min/day were substantial. Whether greaterchanges in GLUT4 content could have been achieved byemploying a different exercise protocol is not known. However,based on work in other species, it is possible that a similarelevation in GLUT4 might have been achieved with higherintensity training over a shorter training period.
One of the outcomes of endurance training is a reduction inplasma glucose utilisation during exercise at a givensubmaximal workload. However, while decreases in fuelutilisation have been shown when work is performed in theuntrained and trained state at the same absolute workload,changes in fuel utilisation are minimal or nonexistent whenwork is performed at the same relative intensity. Recently, wehave demonstrated that moderate-intensity training in horsesdecreases glucose flux during submaximal exercise at the sameabsolute, but not relative, workload (Geor et al. 2002). Thisobservation might seem contradictory in light of thedemonstrated increase in GLUT4 protein content of skeletalmuscle following training in the present study and in previousinvestigations in rats and man (Houmard et al. 1995; Terada etal. 2001). By analogy, the post-training increases in GLUT4content might be expected to result in greater, not decreased,utilisation of glucose during exercise. The fact that the increasein glucose transporter protein post-training was not reflected insarcolemmal glucose transport activity leads us to speculate thatthere was a reduction in the extent of translocation of GLUT4to the sarcolemma in trained muscle.
There are varying findings as to the effects of training onglucose transport capacity. In rats and man, a training-inducedincrease in GLUT4 content and a concomitant increase ininsulin-stimulated glucose transport activity has been reported(Mikines et al. 1989; Dela et al. 1993). In contrast, Richter et al.(1998a) recently reported that training blunted the exercise-associated translocation of GLUT4 protein to the sarcolemma,in turn leading to diminished exercise-induced sarcolemmalglucose transport capacity (Richter et al. 1998b; Wojtaszewskiand Richter 1998). The investigators interpreted the decreasedglucose transport to be due to decreased GLUT4 translocationto the sarcolemma. This reduction in GLUT4 translocation inhuman subjects was demonstrated in experiments thatemployed the sarcolemmal vesicle technique that was also usedin the present study. Importantly, whereas our findings indicatethat glucose transport activity is reduced over a range of glucoseconcentrations (Figs 2, 3), this decrease in GLUT4 translocationis evident at glucose concentrations of 5–10 mmol/l, the rangeover which glucose concentration varies during submaximalexercise in horses (Geor et al. 2002). Based on recentinvestigations by Kristiansen et al. (2000), a higher rate ofglucose uptake can be achieved after training when working atthe same relative workload in a glycogen-depleted state.However, these investigators indicate that this post-trainingincrease in glucose uptake was only possible when theworkload was extremely high (80–100% VO2max). Thesefindings support the recent observations of Richter et al. (2001)that contraction-induced muscle glucose uptake does notrepresent the product of a single signalling pathway but is morelikely to be the result of the interaction of several signallingpathways activated to different extents according to the specificmetabolic needs of any given situation.
The 2 main pathways identified for stimulation of glucosetransport in skeletal muscle are activation by insulin andactivation by exercise (or muscular contractions) (Holloszy andHansen 1996; Hayashi et al. 1997). Maximally insulin-
stimulated glucose transport appears to increase in proportionwith the increase in skeletal muscle GLUT4 post-training, withthe maximal effects of insulin and exercise being additive (Renet al. 1994; Hayashi et al. 1997). A third possible influence forglucose transport is the glycogen content of the muscle. Thereare conflicting reports as to the possible effects of muscleglycogen content on glucose transport activity. Overall, itappears that 1) reduced muscle glycogen is associated withincreased glucose transport and insulin sensitivity of thetransport process and 2) high muscle glycogen reverses theincrease in insulin sensitivity normally associated with exercise(Youn and Buchanan 1993; Jensen et al. 1997; Host et al. 1998).We have recently reported a 15% increase in resting gluteusmedius muscle glycogen content and higher muscle glycogenfollowing exercise after moderate-intensity training in horses(Geor et al. 2002). It is, therefore, possible that a reduction inglucose transport could reflect an attenuation of stimulus fortransport associated with an elevation in resting muscle glycogenafter training. However, at this time mechanisms underlying therelationship between muscle glycogen concentration andglucose transport are still largely speculative. Furthermore, themethodology used in the present study did not allow us toexplore the possible influence of glycogen content on theobserved reduction in muscle membrane glucose transport.
As previously indicated, both GLUT1 and GLUT4 isoformsof the glucose transporter protein are present in skeletal muscle.However, it has been estimated that the majority (>60%) of theGLUT1 isoform is located with free T-tubule membranes andintramuscular nerve branches (Ploug et al. 1989; Hanberg et al.1992). Work by Ploug et al. (1993) demonstrated that the use oflarge sarcolemmal vesicles resulted in a preparation thatcontained little or no GLUT1, thereby allowing the study of thekinetics of GLUT4 transporters alone. However, while GLUT4is preferentially concentrated in these membrane preparations, itcannot be assumed that GLUT1 is entirely excluded in vesicleformation. In addition, the work of Ploug et al. (1993)demonstrated that the recruitment of GLUT4 transporters isprobably from a single intracellular site, as there was no additiveeffect of insulin on glucose transport in giant vesicles followingmuscle contractions. In the present study, we did not attempt toassess the muscle membrane preparations for the presence ofGLUT1 transporters and it is, therefore, possible that a fractionof the total transport activity reported could represent the activityof the GLUT1 isoform. However, our results demonstrate ablunting of glucose transport following moderate-intensityexercise training rather than an enhancement of transport activityas might be expected if additional transport protein was present.
It is concluded that moderate-intensity exercise training inhorses results in an increase in skeletal muscle GLUT4.However, following 60 min of submaximal exercise, theincrease in exercise-induced glucose transporter proteindetermined in post-training muscle homogenates was notreflected in muscle membrane glucose transport activity,suggesting a reduction in the extent of translocation of GLUT4to the sarcolemma in trained muscle.
Acknowledgements
The authors gratefully acknowledge the technical assistance ofHua Shen and Vivian Nicholson at the University of Guelph andLeia Hill, Candace Sack and Steve Clinard for their assistance atthe Ohio State University. This study was supported by grantsfrom the Equine Research Funds at the Ohio State Universityand the Equine Research Program of the Ontario Ministry ofAgriculture, Food and Rural Affairs.

204 Changes in muscle GLUT-4 content following training
Manufacturers’ addresses
1Countrymark Cooperative, Indianapolis, Indiana, USA.2Sato, Uppsala, Sweden.3Columbus Instruments, Columbus, Ohio, USA.4Millipore, USA.5Biogenesis Inc., Sandown, New Hampshire, USA.6Amersham, Arlington Heights, Illinois, USA.7Hoefer, San Francisco, California, USA.8Jandel Scientific, San Rafael, California, USA.
References
Burton, F., Dorstelmann, U. and Hutter, O.F. (1988) Single-channel activity insarcolemmal vesicles from human and other mammalian muscles. MuscleNerve 11, 1029-1033.
DeFronzo, R.A., Jacot, E., Jequier, E., Maeder, E., Wahren, J. and Felber, J.P.(1981) The effect of insulin on the disposal of intravenous glucose. Diabetes30, 1000-1007.
Dela, F.K., Handberg, A., Mikines, K.J., Vinten, J. and Galbo, H. (1993) GLUT4and insulin receptor binding and kinase activity in trained human muscle. J.Physiol. (London) 469, 615-624.
Fedak, M.A., Rome, L. and Seeherman, H.J. (1981) One-step N2-dilutiontechnique for calibrating open-circuit VO2 measuring systems. J. appl. Physiol.51, 772-776.
Geor, R.J., McCutcheon, L.J. and Shen, H. (1999) Muscular and metabolicresponses to moderate intensity short-term training. Equine vet. J., Suppl. 30,311-317.
Geor, R.J., McCutcheon, L.J. Hinchcliff, K.W. and Sams, R.A. (2002) Training-induced alterations in glucose metabolism during moderate-intensity exercise.Equine vet. J., Suppl. 34, 22-28.
Handberg, A., Kayser, L. Hoyer, P.E. and Vinten, J. (1992) A substantial part ofGLUT-1 in crude membranes from muscle originates from perineural sheets.Am. J. Physiol. 262 (Endocrinol. Metab. 21), E721-E727.
Hayashi, T. Woitaszewski, J.F.P. and Goodyear, L.J. (1997) Exercise regulation ofglucose transport in skeletal muscle. Am. J. Physiol. 273 (Endocrinol. Metab.),E1039-E1051.
Hinchcliff, K.W., McKeever, K.H., Muir, W.W. and Sams, R.A. (1993) Effect offurosemide and weight carriage on energetic responses of horses toincremental exertion. Am. J. vet. Res. 54, 1500-1504.
Hirshman, M.F., Wardzala, L.J., Goodyear, L.J., Fuller, S.P., Horton, E.D. andHorton, E.S. (1993) Exercise training increases GLUT-4 protein in rat adiposecells. Am. J. Physiol. 264 (Endrocrinol. Metab. 27), E882-E889.
Holloszy, J.O. and Hansen, P.A. (1996) Regulation of glucose transport intoskeletal muscle. Rev. Physiol. Biochem. Pharmacol. 128, 99-193.
Host, H.H., Hansen, P.A. Nolte, L.A., Chen, M.A. and Holloszy, J.O. (1998)Glycogen supercompensation masks the effect of a training-induced increasein GLUT-4 on muscle glucose transport. J. appl. Physiol. 85, 133-138.
Houmard, J.A., Hickey, M.S., Tyndall, G.L., Gavigan, K.E. and Dohm, G.L. (1995)Seven days of exercise increase GLUT-4 protein content in human skeletalmuscle. J. appl. Physiol. 79, 1936-1938.
Ihlemann, J., Ploug, T., Hellsten, Y. and Galbo, H. (1999) Effect of tension oncontraction-induced glucose transport in rat skeletal muscle. Am. J. Physiol.277 (Endocrinol. Metab. 38), E208-E214.
Jensen, J., Aslesen, R., Ivy, J.L., and Brors, O. (1997) Role of glycogenconcentration and eipnephrine on glucose uptake in rat epitrochlearis muscle.Am. J. Physiol. 272 (Endocrinol. Metab. 35), E649-E655.
Johannsson, E., McCullagh, K.J.A., Han, X., Fernando, P.K., Jensen, J., Dahl, H.A. and Bonen, A. (1996) Effect of overexpressing GLUT-1 and GLUT-4 oninsulin and contraction-stimulated glucose transport in muscle. Am. J. Physiol.271 (Endocrinol. Metab. 34), E547-E555.
Knight, P.K., Sinha, A.K. and Rose, R.J. (1991) Effects of training intensity onmaximum oxygen uptake. In: Equine Exercise Physiology 3, Eds: S.G.B.Persson, A. Lindholm and L.B. Jeffcott, ICEEP Publications, Davis, California.pp 77-82.
Kristiansen, S., Gade, J., Wojtaszewski, J.F.P, Kiens, B. and Richter, E.A. (2000)Glucose uptake is increased in trained vs. untrained muscle during heavyexercise. J. appl. Physiol. 89, 1151-1158.
Marette, A., Richardson, J.M., Ramlal, T., Balon, T.W., Vranic, M., Pessin, J.E. andKlip, A. (1992) Abundance, locatization, and insulin-induced translocation ofglucose transporters in red and white muscles. Am. J. Physiol. 263 (CellPhysiol. 32), C443-C452.
Megeny, L.A., Prasad, M., Tan, M.H. and Bonen, A. (1994) Expression of theinsulin-regulatable transport GLUT-4 is influenced by neurogenic factors. Am.J. Physiol. 266 (Endocrinol. Metab. 29), E813-E816.
Megeny, L.A., Neufer, P.D., Dohm, G.L., Tan, M.H., Blewett, C.A., Elder, G.C.B.and Bonen, A. (1993) Effects of muscle activity and fiber composition onglucose transport and GLUT-4. Am. J. Physiol. 264 (Endocrinol. Metab. 27),E583-E593.
Mikines, K.J., Sonne, B., Tronier, B. and Galbo, H. (1989) Effects of acute exerciseand detraining on insulin action in trained men. J. appl. Physiol. 66, 704-711.
Ploug, T., Galbo, H., Ohkuwa, T., Tranum-Jensen, J. and Vinten, J. (1992) Kineticsof glucose transport in rat skeletal muscle membrane vesicles: effects of insulinand contractions. Am. J. Physiol. 262 (Endocrinol. Metab. 25), E700-E711.
Ploug, T., Wojtaszewski, J., Kristiansen, S., Hespel, P., Galbo, H. and Richter, E.(1993) Glucose transport and transporters in muscle giant vesicles: differentialeffects of insulin and contractions. Am. J. Physiol. 264 (Endocrinol. Metab.27), E270-E278.
Ren, J.-M., Semenkovich, C.F., Gulve, E.A., Gao, J. and Holloszy, J.O. (1994)Exercise induces rapid increases in GLUT4 expression, glucose transportcapacity, and insulin-stimulated glycogen storage in muscle. J. Biol. Chem.269, 14369-14401.
Reynolds, T.H., Brozinick, J.T., Rogers, M.A. and Cushman, S.W. (1997) Effectsof exercise training on insulin- and hypoxia-stimulated glucose transport inisolated rat epitrochlearis muscle: assessment of cell-surface GLUT-4 usingATB-BMPA photolabeling. Am. J. Physiol. 272 (Endocrinol. Metab. 35),E320-E325.
Richter, E.A., Jensen, P., Kiens, B. and Kristiansen, S. (1998a) Sarcolemmalglucose transport and GLUT-4 translocation during exercise are diminished byendurance training. Am. J. Physiol. 274 (Endocrinol. Metab.), E89-E95.
Richter, E.A., Kristiansen, S. Wojtaszewski, J. Daugaard, J.R., Asp, S., Hespel, P.and Kiens, B. (1998b) Training effects on muscle glucose transport duringexercise. Adv. Exp. Med. Biol. 441, 107-116.
Richter, E.A., Derave, W. and Wojtaszewski, J.F.P. (2001) Glucose, exercise, andinsulin: emerging concepts. J. Physiol. 535.2, 313-322.
Terada, S., Yokoziki, T., Kawanaka, K., Ogawa, K., Higuchi, M. Ezaki, O. andTabata, I. (2001) Effects of high-intensity swimming training on GLUT-4 andglucose transport activity in rat skeletal muscle. J. appl. Physiol. 90, 2019-2024.
Wojtaszewski, J.F.P. and Richter, E.A. (1998) Glucose utilization during exercise:influence of endurance training. Acta Physiol. Scand. 162, 351-358.
Youn, J.H. and Buchanan, T.A. (1993) Fasting does not impair insulin-stimulatedglucose uptake but alters intracellular glucose metabolism in conscious rats.Diabetes 42, 757-763.
Related Documents