Uncorrected Author Proof Restorative Neurology and Neuroscience xx (20xx) x–xx DOI 10.3233/RNN-140453 IOS Press 1 Cerebellar direct current stimulation modulates pain perception in humans 1 2 Tommaso Bocci a,b , Enrica Santarcangelo c , Beatrice Vannini a , Antonio Torzini b,d , Giancarlo Carli b , Roberta Ferrucci e , Alberto Priori e , Massimiliano Valeriani f ,g and Ferdinando Sartucci a,d,∗ 3 4 a Department of Clinical and Experimental Medicine, Unit of Neurology, Pisa University Medical School, Pisa, Italy 5 6 b Department of Medical and Surgical Sciences and Neuroscience, University of Siena, Siena, Italy 7 c Department of Translational Research and New Technologies in Medicine and Surgery, University of Pisa, Pisa, Italy 8 9 d Department of Clinical and Experimental Medicine, Cisanello Neurology Unit, Pisa University Medical School, Pisa, Italy 10 11 e Department of Neurological Sciences, University of Milan, Fondazione IRCCS Ospedale Maggiore Policlinico, Milan, Italy 12 13 f Division of Neurology, Ospedale Bambino Ges` u, IRCCS, Rome, Italy 14 g Center for Sensory-Motor Interaction, Aalborg University, Aalborg, Denmark 15 Abstract. 16 Purpose: The cerebellum is involved in a wide number of integrative functions, but its role in pain experience and in the nociceptive information processing is poorly understood. In healthy volunteers we evaluated the effects of transcranial cerebellar direct current stimulation (tcDCS) by studying the changes in the perceptive threshold, pain intensity at given stimulation intensities (VAS:0-10) and laser evoked potentials (LEPs) variables (N1 and N2/P2 amplitudes and latencies). 17 18 19 20 Methods: Fifteen normal subjects were studied before and after anodal, cathodal and sham tcDCS. LEPs were obtained using a neodymium:yttrium–aluminium–perovskite (Nd:YAP) laser and recorded from the dorsum of the left hand. VAS was evaluated by delivering laser pulses at two different intensities, respectively two and three times the perceptive threshold. 21 22 23 Results: Cathodal polarization dampened significantly the perceptive threshold and increased the VAS score, while the anodal one had opposite effects. Cathodal tcDCS increased significantly the N1 and N2/P2 amplitudes and decreased their latencies, whereas anodal tcDCS elicited opposite effects. Motor thresholds assessed through transcranial magnetic stimulation were not affected by cerebellar stimulation. 24 25 26 27 Conclusions: tcDCS modulates pain perception and its cortical correlates. Since it is effective on both N1 and N2/P2 components, we speculate that the cerebellum engagement in pain processing modulates the activity of both somatosensory and cingulate cortices. Present findings prompt investigation of the cerebellar direct current polarization as a possible novel and safe therapeutic tool in chronic pain patients. 28 29 30 31 Keywords: Pain cerebellum, cerebellar direct current stimulation, tDCS, laser evoked potentials, pain modulation 32 ∗ Corresponding author: Prof. Ferdinando Sartucci, M.D., Asso- ciate Professor of Neurology, Pisa University Medical School, Neuroscience Department, Neurology - Neurophysiology Units, Via Roma, n. 67; I 56126 Pisa, Italy. Tel.: +39 050 992176 (direct); Fax: +39 050 550563; 992405; E-mail: [email protected]. 1. Introduction 33 The cerebellum is involved in a wide number of 34 integrative functions, ranging from working memory 35 and associative learning to motor control (Schmah- 36 mann, 1991; Ito, 2006; Stoodley & Schmahmann, 37 0922-6028/15/$35.00 © 2015 – IOS Press and the authors. All rights reserved

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Unc
orre
cted
Aut
hor P
roof
Restorative Neurology and Neuroscience xx (20xx) xndashxxDOI 103233RNN-140453IOS Press
1
Cerebellar direct current stimulationmodulates pain perception in humans
1
2
Tommaso Bocciab Enrica Santarcangeloc Beatrice Vanninia Antonio Torzinibd Giancarlo CarlibRoberta Ferruccie Alberto Priorie Massimiliano Valerianifg and Ferdinando Sartucciadlowast
3
4
aDepartment of Clinical and Experimental Medicine Unit of Neurology Pisa University Medical School PisaItaly
5
6
bDepartment of Medical and Surgical Sciences and Neuroscience University of Siena Siena Italy7
cDepartment of Translational Research and New Technologies in Medicine and Surgery University of Pisa PisaItaly
8
9
dDepartment of Clinical and Experimental Medicine Cisanello Neurology Unit Pisa University Medical SchoolPisa Italy
10
11
eDepartment of Neurological Sciences University of Milan Fondazione IRCCS Ospedale Maggiore PoliclinicoMilan Italy
12
13
f Division of Neurology Ospedale Bambino Gesu IRCCS Rome Italy14
gCenter for Sensory-Motor Interaction Aalborg University Aalborg Denmark15
Abstract16
Purpose The cerebellum is involved in a wide number of integrative functions but its role in pain experience and in thenociceptive information processing is poorly understood In healthy volunteers we evaluated the effects of transcranial cerebellardirect current stimulation (tcDCS) by studying the changes in the perceptive threshold pain intensity at given stimulationintensities (VAS0-10) and laser evoked potentials (LEPs) variables (N1 and N2P2 amplitudes and latencies)
17
18
19
20
Methods Fifteen normal subjects were studied before and after anodal cathodal and sham tcDCS LEPs were obtained using aneodymiumyttriumndashaluminiumndashperovskite (NdYAP) laser and recorded from the dorsum of the left hand VAS was evaluatedby delivering laser pulses at two different intensities respectively two and three times the perceptive threshold
21
22
23
Results Cathodal polarization dampened significantly the perceptive threshold and increased the VAS score while the anodalone had opposite effects Cathodal tcDCS increased significantly the N1 and N2P2 amplitudes and decreased their latencieswhereas anodal tcDCS elicited opposite effects Motor thresholds assessed through transcranial magnetic stimulation were notaffected by cerebellar stimulation
24
25
26
27
Conclusions tcDCS modulates pain perception and its cortical correlates Since it is effective on both N1 and N2P2 componentswe speculate that the cerebellum engagement in pain processing modulates the activity of both somatosensory and cingulatecortices Present findings prompt investigation of the cerebellar direct current polarization as a possible novel and safe therapeutictool in chronic pain patients
28
29
30
31
Keywords Pain cerebellum cerebellar direct current stimulation tDCS laser evoked potentials pain modulation32
lowastCorresponding author Prof Ferdinando Sartucci MD Asso-ciate Professor of Neurology Pisa University Medical SchoolNeuroscience Department Neurology - Neurophysiology Units ViaRoma n 67 I 56126 Pisa Italy Tel +39 050 992176 (direct) Fax+39 050 550563 992405 E-mail fsartuccineuromedunipiit
1 Introduction 33
The cerebellum is involved in a wide number of 34
integrative functions ranging from working memory 35
and associative learning to motor control (Schmah- 36
mann 1991 Ito 2006 Stoodley amp Schmahmann 37
0922-602815$3500 copy 2015 ndash IOS Press and the authors All rights reserved
Unc
orre
cted
Aut
hor P
roof
2 T Bocci et al Cerebellum and pain A tcDCS study
2009 Strick et al 2009 Balsters et al 2013) It is38
also involved in the sensory cognitive (Borsook et39
al 2008) and affective dimensions of pain (Ploghaus40
et al 1999) In addition the cerebellum plays a role41
in the sensory-motor integration aimed at antinocicep-42
tive behaviour (Bingel et al 2002 Strigo et al 200343
Borsook et al 2008) as well as in salience-related44
affective and behavioral responses to nociceptive stim-45
ulation (Duerden amp Albanese 2013) In fact although46
it is not known how nociceptive information is encoded47
in the cerebellum it has been proposed that the cerebel-48
lum may integrate multiple effector systems including49
affective processing pain modulation and sensorimo-50
tor control51
Afferent inputs from nociceptors reach the cerebel-52
lum through two different and segregated pathways the53
spino-ponto-cerebellar and the spino-olivo-cerebellar54
route (Ekerot et al 1987a 1987b Ekerot et al55
1991a) and the cerebellar influence on pain process-56
ing closely resembles the inhibitory tone exerted by57
Purkinje cells over the primary motor cortex (M1) a58
phenomenon referred as cerebellum-brain inhibition59
(Kelly amp Strick 2003)60
Non-invasive brain stimulation (NIBS) techniques61
such as repetitive Transcranial Magnetic Stimulation62
(rTMS) and transcranial Direct Current Stimulation63
(tDCS) have recently emerged as interesting effective64
and promising tools for modulating pain experience65
(Antal amp Paulus 2010 Zaghi et al 2011) In fact66
a sufficient body of evidence shows analgesic effects67
of high-frequency rTMS of the primary motor cor-68
tex (M1) (Lefaucheur et al 2014) with effects likely69
arising from the restoration of defective intracortical70
inhibitory processes (Lefaucheur et al 2006) Among71
NIBS technique tDCS applied either over the motor72
(Fregni et al 2007 Mendonca et al 2011 Dasilva73
et al 2012 Reidler et al 2012) or the prefrontal cor-74
tex (Boggio et al 2008 2009 Mylius et al 2012) was75
also effective in pain modulation76
Only one study has assessed the effects of cerebellar77
rTMS suggesting that changes in pain perception were78
not specific for cerebellar stimulation (Zunhammer79
et al 2011) However no study has investigated to date80
the role of transcranial cerebellar direct current stim-81
ulation (tcDCS) a new and well-tolerated technique82
for modulating cerebellar excitability in modifying83
pain perception in humans (Ferrucci et al 2008 201284
Galea et al 2009 2011 Grimaldi et al 2014 Priori et85
al 2014) Notably despite some inter-individual dif-86
ferences recent modelling researches have revealed87
that during tcDCS the current spread to other struc- 88
tures outside the cerebellum is negligible and unlikely 89
to produce functional effects (Fig 1) (Parazzini et al 90
2013 2014a 2014b) 91
The aim of our study was to evaluate the effects 92
of tcDCS on pain perception and on its cortical cor- 93
relates We studied the changes in pain scores and in 94
laser evoked potentials (LEPs) variables (perceptive 95
threshold N1 and N2P2 amplitudes and latencies) 96
in participants undergoing direct current polarization 97
applied over the cerebellum 98
2 Materials and methods 99
21 Subjects 100
Fifteen healthy volunteers (mean age plusmn SD 101
258 plusmn 59 years 7 women) with no history of neuro- 102
logical disorders were enrolled in the study Women 103
were studied in the second week after their last menses 104
(Smith et al 1999) No subject had been under 105
medication in the month preceding the experimental 106
session which was scheduled at least 48 hours after 107
the last alcohol and caffeine consumption Written 108
informed consent was obtained from all participants 109
before enrollment in the study which was approved 110
by the local ethical Committee and followed the tenets 111
of the Declaration of Helsinki 112
22 Experimental design 113
As shown in Fig 2 at the beginning of each session 114
before cerebellar tDCS and immediately afterwards 115
the laser Perceptive Threshold (PT) corresponding to 116
the lowest intensity at which subjects perceived at least 117
50 of the stimuli (Cruccu et al 1999 Agostino et al 118
2000) was determined In order to minimize the num- 119
ber of nociceptive stimuli the nociceptive perception 120
threshold was not assessed A range of 10ndash40 stimuli 121
(mean SD 25 plusmn 5) was used to assess the perceptive 122
threshold before and after transcranial cerebellar stim- 123
ulation Less than 10 minutes were spent to determine 124
PT in line with previous reports (Truini et al 2011) 125
After the PT assessment participants were 126
instructed to pay attention to incoming laser noci- 127
ceptive stimuli in order to verbally rate the perceived 128
intensity about 2-3 seconds after each laser stimulation 129
which was performed before tcDCS (T0) immediately 130
after its termination (T1) and 60 min later (T2) 131
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 3
A
B
1
08
06
04
02
0
Fig 1 - Current density generated by cerebellar transcranial directcurrent stimulation (cerebellar tDCS) in humans A Top panel shows(viewed from the back) the electrode positions for cerebellar tDCSB Examples of segmented tissues in two human realistic VirtualFamily models (Ella and Duke) undergoing cerebellar tDCS Simu-lations were conducted using the simulation platform SEMCAD X(by SPEAG Schmid amp Partner Engineering AG Zurich Switzer-land) a lateral view of cerebellum pons midbrain medulla blateral view of the skull c back view of the cerebellum d and elateral and inferior views of normalized current density amplitudefield distributions over cortical subcortical and brainstem regionsf back view of normalized current density amplitude field distribu-tions over the cerebellum Values are normalized with respect to themaximum of the current density amplitude in the cerebellum Thespread of the current density (J) over the occipital cortex - quantifiedas the percentage of occipital volume where the amplitude of J-fieldis greater than 70 of the peak of J in the cerebellum - was only 4for ldquoDukerdquo and much less than 1 for ldquoEllardquo (modified from Prioriet al (2014) with permission)
Participants were blinded to the tcDCS polarity 132
anodal cathodal and sham tcDCS stimulations were 133
administered in three different sessions and separated 134
by at least 1 week to avoid possible carry-over effects 135
The order of interventions was randomized and bal- 136
anced across subjects Laser stimuli of intensity two 137
and three times the PT intensity (I1 I2) were delivered 138
by an experimenter (AT) whereas the evaluation of 139
electrophysiological parameters was done by FS both 140
blinded to the tcDCS polarity B V settled the tcDCS 141
polarity 142
221 Subjective experience 143
The perceived sensation was rated on the 0ndash10 144
Visual Analogue Scale (where 0 = no sensation and 145
10 = unbearable pain the intermediate levels being 146
1 = barely perceived 2 = lightly pricking not painful 147
3 = clearly pricking not painful 4 = barely painful 148
5 = painful prompting to rub the skin 6 = very painful 149
and distressing 7 and more strongly unpleasant pain) 150
VAS was studied in each subject after 10 nociceptive 151
laser I1 and I2 stimuli (VAS 1 VAS2) In each partic- 152
ipant individual VAS values were averaged for each 153
Time 154
Laser Evoked Potentials were obtained by stim- 155
uli corresponding to two times the Perceptive value 156
according with previous literature and guidelines 157
(Truini et al 2005 2010) 158
23 Procedures 159
231 Laser evoked potentials (LEPs) 160
The methods used for laser stimulation are 161
reported in detail elsewhere (Truini et al 2005 162
2010) A neodymiumyttriumndashaluminiumndashperovskite 163
(NdYAP) laser was used (wavelength 104 m pulse 164
duration 2ndash20 ms maximum energy 7 J) The laser 165
beam was transmitted from the generator to the stim- 166
ulating probe via a 10 m length optical fibre signals 167
were then amplified band pass filtered (01ndash200 Hz 168
time analysis 1000 ms) and fed to a computer for stor- 169
age and analysis (Cruccu et al 2008) The dorsum of 170
the left hand was stimulated by laser pulses (individ- 171
ual variability 389ndash1575 Jcm2) with short duration 172
(5 ms) and small diameter spots (5 mm Valeriani et al 173
2012) Ten stimuli whose intensity was established 174
on the basis of the Perceptive Threshold assessed for 175
each subject at T0 T1 and T2 were delivered and 176
the laser beam was shifted slightly between consec- 177
utive pulses to avoid skin lesions and reduce fatigue 178
Unc
orre
cted
Aut
hor P
roof
4 T Bocci et al Cerebellum and pain A tcDCS study
Fig 2 ndash Experimental protocol Psychophysical and electrophysiological variables evaluated at baseline (T0) and at two different time points(T1 T2) following anodal cathodal and sham tcDCS
of peripheral nociceptors (Truini et al 2005) The179
inter-stimulus interval was varied randomly (10ndash15 s)180
Participants were reclined on a couch and wore protec-181
tive goggles They were instructed to keep their eyes182
open and gaze slightly downwards since the N2P2183
amplitude is enhanced by attention (Lorenz amp Garcia-184
Larrea 2003 Truini et al 2005) they were requested185
to mentally count the number of stimuli The main186
Aδ-LEP vertex complex N2ndashP2 and the lateralised187
N1 component were recorded through standard disc188
non-polarizable AgAgCl surface electrodes (diameter189
10 mm Biomedreg Florence Italy) N2 and P2 compo-190
nents were recorded from the vertex (Cz) referenced191
to the earlobes the N1 component was recorded from192
the temporal leads (T4) referenced to Fz (Cruccu et al193
2008) Blinks and saccades were recorded with an EOG194
electrode placed on the supero-lateral right canthus195
connected to the system reference Ground was placed196
on the mid-forehead Skin impedance was kept below197
5 k198
232 Cerebellar transcutaneous direct current199
stimulation (tcDCS)200
tDCS was applied using a battery-driven constant201
current stimulator (HDCStim Newronika Italy) and202
a pair of electrodes in two saline-soaked synthetic203
sponges with a surface area of 25 cm2 For cathodal204
stimulation the cathode was centered on the median205
line 2 cm below the inion with its lateral borders about206
1 cm medially to the mastoid apophysis and the anode 207
over the right shoulder (Ferrucci et al 2008 2012 208
2013) For anodal stimulation the current flow was 209
reversed In the real tcDCS conditions direct current 210
was transcranially applied for 20 minutes with an inten- 211
sity of 20 mA and constant current flow was measured 212
by an ampere meter (current density asymp 008 mAcm2) 213
These values are similar to those previously reported 214
for cerebellar stimulation (Ferrucci et al 2008 2013) 215
are considered to be safe (Iyer et al 2005) and are 216
far below the threshold for tissue damage (Nitsche 217
et al 2003) Apart from occasional and short-lasting 218
tingling and burning sensations below the electrodes 219
direct current stimulation strength remained below the 220
sensory threshold throughout the experimental session 221
At the offset of tDCS the current was decreased in a 222
ramp-like manner a method shown to achieve a good 223
level of blinding among sessions (Gandiga et al 2006 224
Galea et al 2009) For a sham tDCS the current was 225
turned on only for 5 seconds at the beginning of the 226
sham session and then it was turned off in a ramp- 227
shaped fashion which induces initial skin sensations 228
indistinguishable from real tDCS 229
For all the electrophysiological recordings we chose 230
the left side to avoid interference from the return 231
electrode placed over the contralateral shoulder At 232
experimental debriefing subjects were not able to dis- 233
criminate between the applied anodal cathodal and 234
sham tDCS 235
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 5
Table 1
Row data (expressed as mean value plusmn 1 standard deviation a = anodal stimulation c = cathodal stimulation sh = sham condition)Both psychophysical and electrophysiological data for each subject are fully available as supplementary electronic material at
httpwwwenricasantarcangelocompublications
aT0 aT1 aT2 cT0 cT1 cT2 shT0 shT1 shT2
PT mean 462 607 609 485 376 368 472 466 489SD 080 095 092 086 062 067 098 062 081
VAS I1 mean 389 255 265 367 493 467 387 393 387SD 084 057 062 082 096 082 074 070 092
VAS I2 mean 540 402 403 524 673 665 533 549 530SD 063 082 071 048 047 049 078 069 064
N1 amplitude (V) mean 1292 848 801 1104 1496 1494 1101 1111 1121SD 318 298 258 265 258 333 250 267 283
N1 latency (ms) mean 12419 16146 15710 12704 10715 10405 12817 12867 13066SD 1090 1338 1368 1075 675 912 1320 1271 1209
N2P2 amplitude(V) mean 1114 738 757 1052 1453 1375 1114 1125 1147SD 262 237 233 265 296 329 272 269 216
N2P2 latency (ms) mean 15157 18932 18726 14878 12673 13230 15390 15108 15551SD 1312 1749 2139 2201 1849 1870 1433 1507 1675
233 Transcranial magnetic stimulation (TMS)236
Changes in Resting Motor Threshold (RMT) were237
evaluated at different intervals before and after the238
completion of tcDCS A Magstim Super Rapid Tran-239
scranial Magnetic Stimulator (Magstim Company240
Dyfed UK 22 T maximum field output) connected241
to a standard eight-shaped focal coil with wing diame-242
ters of 70 mm was used The handle of the eight-shaped243
focal coil was pointed backwards and rotated about 45244
deg to the mid-sagittal line to induce a tissue current245
perpendicular to the motor strip in the precentral sul-246
cus (Rossi et al 2009 Groppa et al 2012) RMT247
was defined as the minimum stimulator output that248
induces motor evoked potentials (MEPs) of more than249
50 V in at least five out of 10 trials when first dig-250
ital interosseus (FDI) muscle was completely relaxed251
(Ni et al 2007) The motor ldquohot spotrdquo for the targeted252
muscle was identified by single pulses of TMS deliv-253
ered at a slightly suprathreshold stimulus intensity and254
the magnetic stimuli induced monophasic pulses The255
coil was placed over the right motor cortex (centered on256
C4 according with the 10ndash20 EEG International Sys-257
tem) and electromyographic recordings were made by258
two standard non-polarizable AgAgCl surface elec-259
trodes (diameter 10 mm Biomedreg Florence Italy)260
one placed over the belly of the contralateral FDI261
muscle and the other on the skin overlying the first262
metacarpophalangeal joint of the first finger of the left263
hand RMT was evaluated to exclude possible cere-264
bellar stimulation spread out inducing motor cortex265
activation266
24 Variables and statistical analysis 267
We studied the subjective experience - percep- 268
tive threshold (PT) and pain intensity perceived 269
after laser I1 and I2 (VAS1 VAS2) - and elec- 270
trophysiological variables that is the peak-to-peak 271
amplitude of the N1 wave and N2P2 complex 272
the peak latency of N1 and N2 as reported in 273
previous papers using NdYAG laser (Lefaucheur 274
et al 2001 2002)275
Analyses were performed through SPSS15 sta- 276
tistical Package Psychophysical (PT VAS1 VAS2) 277
and electrophysiological variables (mean values of 278
ten traces N1amplitude and latency N2P2 ampli- 279
tude and latency) as well as Resting Motor Thresholds 280
(RMT) were analysed following a 3 Stimulation con- 281
ditions (anodal cathodal sham) times 3 Times (T0 T1 282
T2) design The Greenhouse-Geisser correction for 283
non sphericity was applied when necessary Con- 284
trast analysis between Times (F values) and paired 285
t tests between stimulations were alternatively used 286
for post-hoc comparisons when appropriate After 287
Bonferroni correction significance level was set at 288
p lt 0007289
The changes of all variable in T1 and T2 290
were expressed as ratio between post and pre 291
stimulation values (T1T0 T2T0) and compared 292
between each other according to a 2 Stimula- 293
tion (anodal cathodal) times 2 Times (T1To T2T0) 294
design 295
Unc
orre
cted
Aut
hor P
roof
6 T Bocci et al Cerebellum and pain A tcDCS study
Table 2
Contrast analyses all comparisons were highly significant(p lt 00001)
anodal cathodal shamPT time df F = 4430 F = 1867 ns
T0 vs Tl 228 F = 77669) F = 27523T0 vs T2 114 F = 78745 F = 27827Tl vs T2 114 ns ns
anodal vs sham cathodal vs shamT0 114 ns nsTl 114 t = 5069 t = 6991T2 114 t = 3709 t = 5849
VAS anodal cathodal sham
time 228 F = 41954 F = 31448 nsT0 vs Tl 114 F = 56968 F = 48596T0 vs T2 114 F = 52289 F = 525Tl vs T2 ns ns
anodal vs sham cathodal vs shamT0 114 ns nsTl 114 t = 644 t = 5916T2 114 t = 5294 t = 582
3 Results296
Row data (mean SD) are shown in Table 1 Base-297
line values were in line with those reported by earlier298
studies performed by using NdYAG laser (Lefaucheur299
et al 2001) Indeed only one study described a longer300
latency of the N2 wave (Cruccu et al 2008) The301
sham stimulation did not modulate any psychophys-302
ical and electrophysiological variable (Table 2) Since303
no pre-post difference was found for sham polarity this304
condition was not included in the comparison between305
Stimulations and Times306
31 Psychophysics307
PT exhibited a significant Stimulation effect308
(F(228) = 35055 p lt 00001 η2 = 0715) and a signifi-309
cant Stimulation times Time interaction (F(456) = 39464310
p lt 00001 η2 = 0738) Decomposition of the latter311
(Table 2) revealed that a) PT was higher for the anodal312
and lower for the cathodal stimulation conditions com-313
pared with the sham stimulation for both T1 and T2314
b) with respect to T0 PT increased in T1 and T2 in the315
anodal condition and decreased in the cathodal con-316
dition while no significant difference was observed317
between T1 and T2 (Fig 3A)318
Significantly different VAS1 and VAS2 were319
observed for the two stimulation intensities (F(114)320
= 54262 p lt 00001) and the three Stimulation con-321
ditions (F(218) = 88882 p lt 00001) Decomposition322
of the significant Stimulation times Time interaction323
(F(456) = 11596 p lt 00001) revealed that the reported 324
pain intensity for both stimulation intensities (VAS1 325
and VAS2) was higher for the cathodal and lower for 326
the anodal stimulation compared to the sham stimula- 327
tion (Table 2) It increased in T1 and decreased in T2 328
with respect to T0 whereas no significant difference 329
was found between T1 and T2 (Fig 3-B) 330
32 Laser evoked potentials 331
Figure 4-A shows the LEPs recorded in all experi- 332
mental conditions in a representative subject Both N1 333
and N2P2 amplitude (N1 F(456) = 10695 p lt 00001 334
η2 = 0884 N2P2 F(456) = 86864 p lt 00001 η2 = 335
0861) and latency (N1 F(456) = 110869 p lt 00001 336
η2 = 0888 N2P2 F(456) = 3660 p lt 00001 337
η2 = 0723) exhibited a significant Time times Stimulation 338
interaction Its decomposition (Table 3) showed 339
that both amplitudes increased and both latencies 340
decreased for cathodal stimulation in T1 and T2 with 341
respect to T0 the opposite occurred for the anodal 342
stimulation Both stimulations induced responses 343
significantly different from the sham condition 344
(Fig 4-B) The responses obtained after cathodal 345
stimulation were significantly improved (higher 346
amplitudes lower latencies) than those produced 347
by the anodal one (N1 amplitude F(114) = 41345 348
p lt 00001 N1 latency F(114) = 496228 p lt 00001 349
N2P2 amplitude (F(114) = 44537 p lt 00001 N2P2 350
latency F(114) = 119056 p lt 00001) 351
33 Resting motor thresholds 352
RMT values at baseline did not differ among exper- 353
imental conditions (mean plusmn SD sham 508 plusmn 83 354
anodal 491 plusmn 62 cathodal 503 plusmn 63) ANOVA 355
did not reveal any significant Stimulation (F(228) = 356
0882 p = 0425 η2 = 0059) Time (F(228) = 0212 357
p = 0810 η2 = 0015) and Stimulation times Time 358
effect (F(456) = 0339 p = 0851 η2 = 0024) for RMT 359
(Fig 5) 360
4 Discussion 361
Our study shows that cerebellar direct current 362
polarization modulates nociceptive perception and its 363
cortical correlates in healthy humans Specifically 364
cathodal tcDCS increases pain perception increases 365
amplitudes and decreases LEPs latencies likely though 366
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 7
A
B
Fig 3 - A Perceptive Threshold Changes (mean plusmn SD) at T1 and T2 with respect to baseline values (T1T0 T2T0) following sham (black)anodal (white) and cathodal (grey) tcDCS (lowastlowastp lt 0001 lowastlowastlowastp lt 00001) B Changes in visual analogue scale (VAS) scores over time VAS scoresat two different stimulus intensity respectively two (A left) and three (B right) times higher than the PT (lowastlowastp lt 0001 lowastlowastlowastp lt 00001)
A B
Fig 4 ndash A LEPs grand averaging traces were recorded at baseline (T0 black) and immediately after cerebellar polarization (T1 red) due tosham (left column) anodal (middle) and cathodal (right) tcDCS B Histograms showing LEPs variables and VAS scores changes (mean plusmn SD)after sham (black) anodal (white) or cathodal (grey) tcDCS with respect to baseline Top panels changes in N1 variables (amplitude and latency)over time bottom panels changes in N2P2 complex (lowastlowastp lt 0001 lowastlowastlowastp lt 00001)
Unc
orre
cted
Aut
hor P
roof
8 T Bocci et al Cerebellum and pain A tcDCS study
Table 3
LEPs post-hoc analyses p lt 00001 for all comparison except when explicitly indicated lowastlowastp lt 0002 lowast p lt 0005
N1 amplitude latency N2P2 amplitude latency
anodal cathodal shamdf
time 228 F = 67152 F = 96489 F = 134912 F = 34946 nsT0 vs T1 114 F = 109178 F = 18815 F = 165953 F = 64281T0 vs T2 114 F = 75143 F = 167697 F = 145125 F = 37818T1 vs T2 114 ns ns ns ns
anodal cathodaltime 228 F = 102281 F = 98717 F = 6577 F = 20918 nsT0 vs T1 114 F = 511186 F = 104027 F = 144112 F = 103864T0 vs T2 114 F = 96329 F = 116841 F = 105183 F = 14012lowastlowastT1 vs T2 114 ns ns ns ns ns
anodal vs sham anodal vs shamT0 114 ns ns ns nsT1 114 ns t = 925 t = 601 6262T2 114 ns t = 8128 t = 6731 5236
cathodal cathodal vs shamT0 114 ns ns t = 3281lowast nsT1 114 t = 16594 t = 8029 t = 8262 t = 520T2 114 t = 7309 t = 12669 t = 5048 t = 5301
Fig 5 - Resting Motor Thresholds Changes (mean plusmn SD) in Resting Motor threshold (RMT) expressed as percentage of the maximumstimulator output after sham (black) anodal (white) and cathodal (grey) tcDCS with respect to baseline marked as dotted line (lowastlowastp lt 0001lowastlowastlowastp lt 00001)
reduction of the inhibitory tone exerted by the cere-367
bellum on brain targets Anodal polarization elicits368
opposite effects producing analgesia Both findings369
support the role of the cerebellum in pain control370
it is noticeable that cathodal cerebellar stimulation371
induces hyperalgesia as occurs in patients with cere-372
bellar infarction (Ruscheweyh et al 2014)373
We would like to underline that in the present study374
LEPs were obtained at laser intensities depending on375
the perceptive threshold which varied as a function376
of anodal and cathodal stimulation This means that 377
the cerebellar stimulation has not a selective analgesic 378
effect as it influences both non nociceptive and noci- 379
ceptive perception A pre-eminent analgesic cannot be 380
assessed because the nociceptive threshold was not 381
evaluated 382
As tcDCS was effective on the modulation of 383
both N1 and N2P2 components and these responses 384
are generated by parallel and partially segregated 385
spinal pathways reaching different cortical targets 386
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 9
(Valeriani et al 2007) we may suggest that the cere-387
bellum is engaged in pain processing by modulating388
the activity of both somatosensory and cingulate cor-389
tices Indeed the cerebellum is involved in both the390
sensory-discriminative and emotional dimension of391
pain (Singer et al 2004 Moriguchi et al 2007) and392
non-invasive cerebellar current stimulation may modu-393
late pain experience and the associated cortical activity394
through many not alternative mechanisms In partic-395
ular changes in N1 reflects the modulation of the396
sensory component of pain while the vertex N2P2397
represents the neural correlate of affective aspects of398
pain experience (Garcia-Larrea et al 1997 Valeriani399
et al 2007) Notably tcDCS may act not only on spinal400
nociceptive neurons but also on wide-range cortical401
networks of the pain matrix (Singer et al 2004) thus402
influencing LEPs and pain experience through both403
top-down and bottom-up mechanisms404
The present study does not allow to hypothesize405
how and where tcDCS influences the cerebellar activ-406
ity A main role of Purkinje cells has been suggested407
as their activity modulation may affect the cerebellar408
inhibitory control of the cerebral cortex (Galea et al409
2009) This would be in line with the effects elicited410
by tDCS in the cerebral cortex which are observ-411
able after both short and long term delay likely also412
interfering with long-term potentiation (LTP)-like phe-413
nomena (Hamada et al 2012 Priori et al 2014)414
Moreover prolonged spiking activity in the cerebellar415
Golgi inhibitory neurons modulates the activity of the416
Purkinje cells and could partly account for the tcDCS417
after-effects (Hull et al 2013)418
The lack of changes in RMT indicates that the anal-419
gesic effects of anodal tcDCS are due to a specific420
modulation of the cerebellar activity and not to motor421
activation On the other hand tcDCS-induced cerebel-422
lar modulation (Purpura amp McMurtry 1965) could be423
not sufficient per se to activate the cerebello ndash thalamo424
- cortical motor pathway (Galea et al 2009) thus425
the reported analgesia and its cortical correlates can-426
not be sustained by the motor cortex activation This427
view is supported by the absence of any association428
between motor symptoms and pain perception in cere-429
bellar patients (Ruscheweyh et al 2014) In the same430
line in healthy subjects it has been recently shown431
that motor task-induced increased cortical excitability432
and analgesia are not associated (Volz et al 2012)433
Indeed RMT is a highly sensitive marker of motor434
tract excitability as it reflects activation of a small435
low-threshold and slow-conducting core of pyramidal436
neurons (Hess et al 1987 Rossini amp Rossi 2007) 437
although RMT may reflect changes in the activity of 438
different central nervous system structures it has been 439
satisfactorily used to assess motor cortex excitability 440
also in cerebellar patients (Battaglia et al 2006) 441
Another critical point is the possibility to modulate 442
with tcDCS both neural correlates underlying nocicep- 443
tive processing and pain perception Previous studies 444
using tDCS over motor cortex were inconsistent among 445
each other some works suggested that tDCS is able to 446
modify pain perception (Boggio et al 2008) while 447
others showed divergent effects on psychophysical 448
and neurophysiological outcome parameters (Luedtke 449
et al 2012 Ihle et al 2014) likely due to a possi- 450
ble overestimation of the role of motor areas on pain 451
processing (Antal et al 2008) 452
Our findings cannot be compared to the results 453
obtained by other Authors In fact the unique study 454
focused on the analgesic effects of non-invasive cere- 455
bellar stimulation reported till now (Zunhammer et al 456
2011) considered only subjective pain thresholds 457
In addition it described similar analgesic effects of 458
cerebellar and neck structures repetitive transcranial 459
magnetic stimulation (rTMS) thus denying any cere- 460
bellar specificity in the observed effects and suggesting 461
that the peripheral information passing through the 462
cerebellum may be responsible for analgesia The 463
main difference between the two studies possibly 464
accounting for different results consists of the neu- 465
romodulation techniques used 466
41 Limitations of the study 467
The present study has a few limitations First our 468
findings do not allow any hypothesis on the role of 469
the cerebellum in chronic pain The observations on 470
patients with cerebellar damage (Ruscheweyh et al 471
2014) suggest that their impaired inhibitory control 472
mechanisms may be not associated with the devel- 473
opment of chronic pain Second we cannot exclude 474
the possibility that tcDCS could modulate not only the 475
cerebellum but also surrounding areas such as the peri- 476
aqueductal gray However recent modelling researches 477
have revealed that during tcDCS the current spread 478
to other structures outside the cerebellum is negligi- 479
ble (Parazzini et al 2013 2014) Moreover several 480
studies have proved that in humans pain processing 481
is encoded within posterior areas of each cerebellar 482
hemisphere specifically in the hemispheric lobule VI 483
Crus I and VIIb (Moulton et al 2011) where the 484
Unc
orre
cted
Aut
hor P
roof
10 T Bocci et al Cerebellum and pain A tcDCS study
tcDCS-induced electrical field is strongly concentrated485
(Parazzini et al 2013) A further limitation is that we486
did not study the contribution of C-fibers the main487
component of spino-ponto-cerebellar and spino-olivo-488
cerebellar pathways In fact ultra-late LEPs related489
to C-fibers activations have not yet been standard-490
ized for clinical application and their occurrence could491
be markedly influenced by high order cognitive pro-492
cesses as they seem to be more affected by the level493
of consciousness and attention than A-delta responses494
(Qiu et al 2002 Opsommer et al 2003 Mouraux amp495
Plaghki 2006) Finally we wish to emphasize that in496
neuropathic patients the effects of the cerebellar stim-497
ulation could be quite different from those described498
here as both anatomical and functional connectivity499
are different from those observed in healthy partici-500
pants (Rocca et al 2010 Riedl et al 2011 Absinta et501
al 2012 Longo et al 2012 Ceko et al 2013)502
5 Conclusions503
Our findings indicate a cerebellar effect on pain504
experience and on its cortical correlates and prompt505
further investigation aimed at assessing whether the506
cerebellar direct current polarization could be used as a507
novel and safe therapeutic tool in chronic pain patients508
Acknowledgments509
We gratefully acknowledge the participation of all510
subjects as well as Mr D Barloscio and Mr C511
Orsini for their excellent technical assistance The512
paper was supported in part by the Italian operating513
and development MIUR PRIN grant year 2006 n514
2006062332 002515
RF and AP are founders and shareholders of516
Newronika srl Milan Italy517
References518
Absinta M Rocca MA Colombo B Falini A Comi G amp Fil-519
ippi M (2012) Selective decreased grey matter volume of the520
pain-matrix network in cluster headache Cephalalgia 32(2)521
109-115522
Agostino R Cruccu G Iannetti G Romaniello A Truini A523
amp Manfredi M (2000) Topographical distribution of pinprick524
and warmth thresholds to CO2 laser stimulation on the human525
skin Neurosci Lett 285(2) 115-118526
Antal A Brepohl N Poreisz C Boros K Csifcsak G amp 527
Paulus W (2008) Transcranial direct current stimulation over 528
somatosensory cortex decreases experimentally induced acute 529
pain perception Clin J Pain 24(1) 56-63 530
Antal A amp Paulus W (2010) Effects of transcranial theta-burst 531
stimulation on acute pain perception Restor Neurol Neurosci 532
28(4) 477-484 533
Apkarian AV Bushnell MC Treede RD amp Zubieta JK 534
(2005) Human brain mechanisms of pain perception and reg- 535
ulation in health and disease Eur J Pain 9(4) 463-484 536
Balsters JH Whelan CD Robertson IH amp Ramnani N (2013) 537
Cerebellum and cognition Evidence for the encoding of higher 538
order rules Cereb Cortex 23(6) 1433-1443 539
Battaglia F Quartarone A Ghilardi MF Dattola R Bagnato 540
S Rizzo V Morgante L amp Girlanda P (2006) Unilat- 541
eral cerebellar stroke disrupts movement preparation and motor 542
imagery Clin Neurophysiol 117(5) 1009-1016 543
Bingel U Quante M Knab R Bromm B Weiller C amp Buchel 544
C (2002) Subcortical structures involved in pain processing 545
Evidence from single-trial fMRI Pain 99(1-2) 313-321 546
Boggio PS Zaghi S amp Fregni F (2009) Modulation of emotions 547
associated with images of human pain using anodal transcranial 548
direct current stimulation (tDCS) Neuropsychologia 47(1) 549
212-217 550
Boggio PS Zaghi S Lopes M amp Fregni F (2008) Modulatory 551
effects of anodal transcranial direct current stimulation on per- 552
ception and pain thresholds in healthy volunteers Eur J Neurol 553
15(10) 1124-1130 554
Borsook D Moulton EA Tully S Schmahmann JD amp 555
Becerra L (2008) Human cerebellar responses to brush and 556
heat stimuli in healthy and neuropathic pain subjects Cerebel- 557
lum 7(3) 252-272 558
Ceko M Seminowicz DA Bushnell MC amp Olausson HW 559
(2013) Anatomical and functional enhancements of the insula 560
after loss of large primary somatosensory fibers Cereb Cortex 561
23(9) 2017-2024 562
Cruccu G Aminoff MJ Curio G Guerit JM Kakigi 563
R Mauguiere F Rossini PM Treede RD amp Garcia- 564
Larrea L (2008) Recommendations for the clinical use of 565
somatosensory-evoked potentials Clin Neurophysiol 119(8) 566
1705-1719 567
Cruccu G Romaniello A Amantini A Lombardi M Innocenti 568
P amp Manfredi M (1999) Assessment of trigeminal small- 569
fiber function Brain and reflex responses evoked by CO2-laser 570
stimulation Muscle Nerve 22(4) 508-516 571
Dasilva AF Mendonca ME Zaghi S Lopes M Dossantos 572
MF Spierings EL Bajwa Z Datta A Bikson M amp 573
Fregni F (2012) tDCS-induced analgesia and electrical fields 574
in pain-related neural networks in chronic migraine Headache 575
52(8) 1283-1295 576
Duerden EG amp Albanese MC (2013) Localization of pain- 577
related brain activation A meta-analysis of neuroimaging data 578
Hum Brain Mapp 34(1) 109-149 579
Ekerot CF Garwicz M amp Schouenborg J (1991a) The postsy- 580
naptic dorsal column pathway mediates cutaneous nociceptive 581
information to cerebellar climbing fibres in the cat J Physiol 582
441 275-284 583
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 11
Ekerot CF Garwicz M amp Schouenborg J (1991b) Topography584
and nociceptive receptive fields of climbing fibres projecting to585
the cerebellar anterior lobe in the cat J Physiol 441 257-274586
Ekerot CF Gustavsson P Oscarsson O amp Schouenborg J587
(1987a) Climbing fibres projecting to cat cerebellar anterior588
lobe activated by cutaneous A and C fibres J Physiol 386589
529-538590
Ekerot CF Oscarsson O amp Schouenborg J (1987b) Stimula-591
tion of cat cutaneous nociceptive C fibres causing tonic and592
synchronous activity in climbing fibres J Physiol 386 539-593
546594
Ferrucci R Brunoni AR Parazzini M Vergari M Rossi E595
Fumagalli M Mameli F Rosa M Giannicola G Zago S596
amp Priori A (2013) Modulating human procedural learning by597
cerebellar transcranial direct current stimulation Cerebellum598
12(4) 485-492599
Ferrucci R Giannicola G Rosa M Fumagalli M Boggio600
PS Hallett M Zago S amp Priori A (2012) Cerebellum601
and processing of negative facial emotions Cerebellar tran-602
scranial DC stimulation specifically enhances the emotional603
recognition of facial anger and sadness Cogn Emot 26(5)604
786-799605
Ferrucci R Marceglia S Vergari M Cogiamanian F Mrakic-606
Sposta S Mameli F Zago S Barbieri S amp Priori607
A (2008) Cerebellar transcranial direct current stimulation608
impairs the practice-dependent proficiency increase in working609
memory J Cogn Neurosci 20(9) 1687-1697610
Fregni F Freedman S amp Pascual-Leone A (2007) Recent611
advances in the treatment of chronic pain with non-invasive612
brain stimulation techniques Lancet Neurol 6(2) 188-191613
Galea JM Jayaram G Ajagbe L amp Celnik P (2009)614
Modulation of cerebellar excitability by polarity-specific615
noninvasive direct current stimulation J Neurosci 29(28)616
9115-9122617
Galea JM Vazquez A Pasricha N de Xivry JJ amp Celnik618
P (2011) Dissociating the roles of the cerebellum and motor619
cortex during adaptive learning The motor cortex retains what620
the cerebellum learns Cereb Cortex 21(8) 1761-1770621
Gandiga PC Hummel FC amp Cohen LG (2006) Tran-622
scranial DC stimulation (tDCS) A tool for double-blind623
sham-controlled clinical studies in brain stimulation Clin Neu-624
rophysiol 117(4) 845-850625
Garcia-Larrea L Peyron R Laurent B amp Mauguiere F (1997)626
Association and dissociation between laser-evoked potentials627
and pain perception Neuroreport 8(17) 3785-3789628
Grimaldi G Argyropoulos GP Boehringer A Celnik P629
Edwards MJ Ferrucci R Galea JM Groiss SJ Hiraoka630
K Kassavetis P Lesage E Manto M Miall RC Priori A631
Sadnicka A Ugawa Y amp Ziemann U (2014) Non-invasive632
cerebellar stimulationndasha consensus paper Cerebellum 13(1)633
121-138634
Groppa S Oliviero A Eisen A Quartarone A Cohen LG635
Mall V Kaelin-Lang A Mima T Rossi S Thickbroom636
GW Rossini PM Ziemann U Valls-Sole J amp Sieb-637
ner HR (2012) A practical guide to diagnostic transcranial638
magnetic stimulation Report of an IFCN committee Clin Neu-639
rophysiol 123(5) 858-882640
Hamada M Strigaro G Murase N Sadnicka A Galea JM 641
Edwards MJ amp Rothwell JC (2012) Cerebellar modulation 642
of human associative plasticity J Physiol 590(Pt 10) 2365- 643
2374 644
Hess CW Mills KR amp Murray NM (1987) Responses in small 645
hand muscles from magnetic stimulation of the human brain J 646
Physiol 388 397-419 647
Hull CA Chu Y Thanawala M amp Regehr WG (2013) Hyper- 648
polarization induces a long-term increase in the spontaneous 649
firing rate of cerebellar Golgi cells J Neurosci 33(14) 5895- 650
5902 651
Ihle K Rodriguez-Raecke R amp Luedtke K May A (2014) 652
tDCS modulates cortical nociceptive processing but has little 653
to no impact on pain perception Pain 155(10) 2080-2087 654
Ito M (2006) Cerebellar circuitry as a neuronal machine Prog 655
Neurobiol 78(3-5) 272-303 656
Iyer MB Mattu U Grafman J Lomarev M Sato S amp Wasser- 657
mann EM (2005) Safety and cognitive effect of frontal DC 658
brain polarization in healthy individuals Neurology 64(5) 872- 659
875 660
Kelly RM amp Strick PL (2003) Cerebellar loops with motor cor- 661
tex and prefrontal cortex of a nonhuman primate J Neurosci 662
23(23) 8432-8444 663
Lefaucheur JP Andre-Obadia N Antal A Ayache SS Baeken 664
C Benninger DH Cantello RM Cincotta M de Carvalho 665
M De Ridder D Devanne H Di Lazzaro V Filipovic SR 666
Hummel FC Jaaskelainen SK Kimiskidis VK Koch G 667
Langguth B Nyffeler T Oliviero A Padberg F Poulet E 668
Rossi S Rossini PM Rothwell JC Schonfeldt-Lecuona 669
C Siebner HR Slotema CW Stagg CJ Valls-Sole J 670
Ziemann U Paulus W amp Garcia-Larrea L (2014) Evidence- 671
based guidelines on the therapeutic use of repetitive transcranial 672
magnetic stimulation (rTMS) Clin Neurophysiol 125(11) 673
2150-2206 674
Lefaucheur JP Brusa A Creange A Drouot X amp Jarry G 675
(2002) Clinical application of laser evoked potentials using 676
the NdYAG laser Neurophysiol Clin 32(2) 91-98 677
Lefaucheur JP Debray S amp Jarry G (2001) Laser evoked poten- 678
tials using the NdYAG laser Muscle Nerve 24(4) 496-501 679
Lefaucheur JP Drouot X Menard-Lefaucheur I Keravel Y 680
amp Nguyen JP (2006) Motor cortex rTMS restores defective 681
intracortical inhibition in chronic neuropathic pain Neurology 682
67(9) 1568-1574 683
Longo MR Iannetti GD Mancini F Driver J amp Haggard P 684
(2012) Linking pain and the body Neural correlates of visually 685
induced analgesia J Neurosci 32(8) 2601-2607 686
Lorenz J amp Garcia-Larrea L (2003) Contribution of attentional 687
and cognitive factors to laser evoked brain potentials Neuro- 688
physiol Clin 33(6) 293-301 689
Lorenz J Minoshima S amp Casey KL (2003) Keeping pain out 690
of mind The role of the dorsolateral prefrontal cortex in pain 691
modulation Brain 126(Pt 5) 1079-1091 692
Luedtke K May A amp Jurgens TP (2012) No effect of a single 693
session of transcranial direct current stimulation on experimen- 694
tally induced pain in patients with chronic low back painndashan 695
exploratory study PLoS One 7(11) e48857 696
Unc
orre
cted
Aut
hor P
roof
12 T Bocci et al Cerebellum and pain A tcDCS study
Mendonca ME Santana MB Baptista AF Datta A Bikson697
M Fregni F amp Araujo CP (2011) Transcranial DC stimu-698
lation in fibromyalgia Optimized cortical target supported by699
high-resolution computational models J Pain 12(5) 610-617700
Moriguchi Y Decety J Ohnishi T Maeda M Mori T Nemoto701
K Matsuda H amp Komaki G (2007) Empathy and judg-702
ing otherrsquos pain An fMRI study of alexithymia Cereb Cortex703
17(9) 2223-2234704
Moulton EA Elman I Pendse G Schmahmann J Becerra L705
amp Borsook D (2011) Aversion-related circuitry in the cere-706
bellum Responses to noxious heat and unpleasant images J707
Neurosci 31(10) 3795-3804708
Moulton EA Schmahmann JD Becerra L amp Borsook D709
(2010) The cerebellum and pain Passive integrator or active710
participator Brain Res Rev 65(1) 14-27711
Mouraux A amp Plaghki L (2006) Are the processes reflected by712
late and ultra-late laser evoked potentials specific of nocicep-713
tion Suppl Clin Neurophysiol 59 197-204714
Mylius V Jung M Menzler K Haag A Khader PH Oer-715
tel WH Rosenow F amp Lefaucheur JP (2012) Effects of716
transcranial direct current stimulation on pain perception and717
working memory Eur J Pain 16(7) 974-982718
Ni Z Gunraj C amp Chen R (2007) Short interval intracortical719
inhibition and facilitation during the silent period in human J720
Physiol 583(Pt 3) 971-982721
Nitsche MA Liebetanz D Lang N Antal A Tergau F amp722
Paulus W (2003) Safety criteria for transcranial direct cur-723
rent stimulation (tDCS) in humans Clin Neurophysiol 114(11)724
2220-2222 author reply 2222-2223725
Opsommer E Guerit JM amp Plaghki L (2003) Exogenous726
and endogenous components of ultralate (C-fibre) evoked727
potentials following CO2 laser stimuli to tiny skin sur-728
face areas in healthy subjects Neurophysiol Clin 33(2)729
78-85730
Parazzini M Fiocchi S Liorni I Priori A amp Ravazzani P731
(2014) Computational modeling of transcranial direct current732
stimulation in the child brain Implications for the treatment of733
refractory childhood focal epilepsy Int J Neural Syst 24(2)734
1430006735
Parazzini M Rossi E Ferrucci R Fiocchi S Liorni I Priori736
A amp Ravazzani P (2013) Computational model of cerebellar737
transcranial direct current stimulation Conf Proc IEEE Eng738
Med Biol Soc 2013 237-240739
Parazzini M Rossi E Ferrucci R Liorni I Priori A amp Ravaz-740
zani P (2014) Modelling the electric field and the current741
density generated by cerebellar transcranial DC stimulation in742
humans Clin Neurophysiol 125(3) 577-584743
Perini I Bergstrand S amp Morrison I (2013) Where pain meets744
action in the human brain J Neurosci 33(40) 15930-15939745
Ploghaus A Tracey I Gati JS Clare S Menon RS Matthews746
PM amp Rawlins JN (1999) Dissociating pain from its antic-747
ipation in the human brain Science 284(5422) 1979-1981748
Priori A Ciocca M Parazzini M Vergari M amp Ferrucci R749
(2014) Transcranial cerebellar direct current stimulation and750
transcutaneous spinal cord direct current stimulation as innova-751
tive tools for neuroscientists J Physiol 592(Pt 16) 3345-3369752
Purpura DP amp McMurtry JC (1965) Intracellular activities and 753
evoked potential changes during polarization of motor cortex 754
J Neurophysiol 28 166-185 755
Qiu Y Inui K Wang X Tran TD amp Kakigi R (2002) Effects 756
of attention distraction and sleep on CO(2) laser evoked poten- 757
tials related to C-fibers in humans Clin Neurophysiol 113(10) 758
1579-1585 759
Reidler JS Mendonca ME Santana MB Wang X Lenkinski 760
R Motta AF Marchand S Latif L amp Fregni F (2012) 761
Effects of motor cortex modulation and descending inhibitory 762
systems on pain thresholds in healthy subjects J Pain 13(5) 763
450-458 764
Riedl V Valet M Woller A Sorg C Vogel D Sprenger 765
T Boecker H Wohlschlager AM amp Tolle TR (2011) 766
Repeated pain induces adaptations of intrinsic brain activity 767
to reflect past and predict future pain Neuroimage 57(1) 206- 768
213 769
Rocca MA Valsasina P Absinta M Colombo B Barcella V 770
Falini A Comi G amp Filippi M (2010) Central nervous 771
system dysregulation extends beyond the pain-matrix network 772
in cluster headache Cephalalgia 30(11) 1383-1391 773
Rossi S Hallett M Rossini PM amp Pascual-Leone A (2009) 774
Safety ethical considerations and application guidelines for 775
the use of transcranial magnetic stimulation in clinical practice 776
and research Clin Neurophysiol 120(12) 2008-2039 777
Rossini PM amp Rossi S (2007) Transcranial magnetic stimulation 778
Diagnostic therapeutic and research potential Neurology 779
68(7) 484-488 780
Ruscheweyh R Kuhnel M Filippopulos F Blum B Eggert 781
T amp Straube A (2014) Altered experimental pain perception 782
after cerebellar infarction Pain 155(7) 1303-1312 783
Schmahmann JD (1991) An emerging concept The cerebellar 784
contribution to higher function Arch Neurol 48(11) 1178- 785
1187 786
Singer T Seymour B OrsquoDoherty J Kaube H Dolan RJ amp 787
Frith CD (2004) Empathy for pain involves the affective but 788
not sensory components of pain Science 303(5661) 1157- 789
1162 790
Smith MJ Keel JC Greenberg BD Adams LF Schmidt PJ 791
Rubinow DA amp Wassermann EM (1999) Menstrual cycle 792
effects on cortical excitability Neurology 53(9) 2069-2072 793
Stoodley CJ amp Schmahmann JD (2009) Functional topography 794
in the human cerebellum A meta-analysis of neuroimaging 795
studies Neuroimage 44(2) 489-501 796
Strick PL Dum RP amp Fiez JA (2009) Cerebellum and non- 797
motor function Annu Rev Neurosci 32 413-434 798
Strigo IA Duncan GH Boivin M amp Bushnell MC (2003) 799
Differentiation of visceral and cutaneous pain in the human 800
brain J Neurophysiol 89(6) 3294-3303 801
Truini A Galeotti F Romaniello A Virtuoso M Iannetti GD 802
amp Cruccu G (2005) Laser-evoked potentials Normative val- 803
ues Clin Neurophysiol 116(4) 821-826 804
Truini A Panuccio G Galeotti F Maluccio MR Sartucci F 805
Avoli M amp Cruccu G (2010) Laser-evoked potentials as a 806
tool for assessing the efficacy of antinociceptive drugs Eur J 807
Pain 14(2) 222-225 808
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 13
Truini A Vergari M Biasiotta A La Cesa S Gabriele M Di809
Stefano G Cambieri C Cruccu G Inghilleri M amp Pri-810
ori A (2011) Transcutaneous spinal direct current stimulation811
inhibits nociceptive spinal pathway conduction and increases812
pain tolerance in humans Eur J Pain 15(10) 1023-1027813
Valeriani M Le Pera D Restuccia D de Armas L Mili-814
ucci R Betti V Vigevano F amp Tonali P (2007) Parallel815
spinal pathways generate the middle-latency N1 and the late P2816
components of the laser evoked potentials Clin Neurophysiol817
118(5) 1097-1104818
Valeriani M Pazzaglia C Cruccu G amp Truini A (2012) Clinical819
usefulness of laser evoked potentials Neurophysiol Clin 42(5)820
345-353821
Volz MS Mendonca M Pinheiro FS Cui H Santana M amp822
Fregni F (2012) Dissociation of motor task-induced cortical823
excitability and pain perception changes in healthy volunteers824
PLoS One 7(3) e34273825
Wager TD Rilling JK Smith EE Sokolik A Casey KL 826
Davidson RJ Kosslyn SM Rose RM amp Cohen JD 827
(2004) Placebo-induced changes in FMRI in the anticipation 828
and experience of pain Science 303(5661) 1162-1167 829
Zaghi S Thiele B Pimentel D Pimentel T amp Fregni 830
F (2011) Assessment and treatment of pain with non- 831
invasive cortical stimulation Restor Neurol Neurosci 29(6) 832
439-451 833
Zubieta JK Bueller JA Jackson LR Scott DJ Xu Y 834
Koeppe RA Nichols TE amp Stohler CS (2005) Placebo 835
effects mediated by endogenous opioid activity on mu-opioid 836
receptors J Neurosci 25(34) 7754-7762 837
Zunhammer M Busch V Griesbach F Landgrebe M Hajak G 838
amp Langguth B (2011) rTMS over the cerebellum modulates 839
temperature detection and pain thresholds through peripheral 840
mechanisms Brain Stimul 4(4) 210-7 e1 841

Unc
orre
cted
Aut
hor P
roof
2 T Bocci et al Cerebellum and pain A tcDCS study
2009 Strick et al 2009 Balsters et al 2013) It is38
also involved in the sensory cognitive (Borsook et39
al 2008) and affective dimensions of pain (Ploghaus40
et al 1999) In addition the cerebellum plays a role41
in the sensory-motor integration aimed at antinocicep-42
tive behaviour (Bingel et al 2002 Strigo et al 200343
Borsook et al 2008) as well as in salience-related44
affective and behavioral responses to nociceptive stim-45
ulation (Duerden amp Albanese 2013) In fact although46
it is not known how nociceptive information is encoded47
in the cerebellum it has been proposed that the cerebel-48
lum may integrate multiple effector systems including49
affective processing pain modulation and sensorimo-50
tor control51
Afferent inputs from nociceptors reach the cerebel-52
lum through two different and segregated pathways the53
spino-ponto-cerebellar and the spino-olivo-cerebellar54
route (Ekerot et al 1987a 1987b Ekerot et al55
1991a) and the cerebellar influence on pain process-56
ing closely resembles the inhibitory tone exerted by57
Purkinje cells over the primary motor cortex (M1) a58
phenomenon referred as cerebellum-brain inhibition59
(Kelly amp Strick 2003)60
Non-invasive brain stimulation (NIBS) techniques61
such as repetitive Transcranial Magnetic Stimulation62
(rTMS) and transcranial Direct Current Stimulation63
(tDCS) have recently emerged as interesting effective64
and promising tools for modulating pain experience65
(Antal amp Paulus 2010 Zaghi et al 2011) In fact66
a sufficient body of evidence shows analgesic effects67
of high-frequency rTMS of the primary motor cor-68
tex (M1) (Lefaucheur et al 2014) with effects likely69
arising from the restoration of defective intracortical70
inhibitory processes (Lefaucheur et al 2006) Among71
NIBS technique tDCS applied either over the motor72
(Fregni et al 2007 Mendonca et al 2011 Dasilva73
et al 2012 Reidler et al 2012) or the prefrontal cor-74
tex (Boggio et al 2008 2009 Mylius et al 2012) was75
also effective in pain modulation76
Only one study has assessed the effects of cerebellar77
rTMS suggesting that changes in pain perception were78
not specific for cerebellar stimulation (Zunhammer79
et al 2011) However no study has investigated to date80
the role of transcranial cerebellar direct current stim-81
ulation (tcDCS) a new and well-tolerated technique82
for modulating cerebellar excitability in modifying83
pain perception in humans (Ferrucci et al 2008 201284
Galea et al 2009 2011 Grimaldi et al 2014 Priori et85
al 2014) Notably despite some inter-individual dif-86
ferences recent modelling researches have revealed87
that during tcDCS the current spread to other struc- 88
tures outside the cerebellum is negligible and unlikely 89
to produce functional effects (Fig 1) (Parazzini et al 90
2013 2014a 2014b) 91
The aim of our study was to evaluate the effects 92
of tcDCS on pain perception and on its cortical cor- 93
relates We studied the changes in pain scores and in 94
laser evoked potentials (LEPs) variables (perceptive 95
threshold N1 and N2P2 amplitudes and latencies) 96
in participants undergoing direct current polarization 97
applied over the cerebellum 98
2 Materials and methods 99
21 Subjects 100
Fifteen healthy volunteers (mean age plusmn SD 101
258 plusmn 59 years 7 women) with no history of neuro- 102
logical disorders were enrolled in the study Women 103
were studied in the second week after their last menses 104
(Smith et al 1999) No subject had been under 105
medication in the month preceding the experimental 106
session which was scheduled at least 48 hours after 107
the last alcohol and caffeine consumption Written 108
informed consent was obtained from all participants 109
before enrollment in the study which was approved 110
by the local ethical Committee and followed the tenets 111
of the Declaration of Helsinki 112
22 Experimental design 113
As shown in Fig 2 at the beginning of each session 114
before cerebellar tDCS and immediately afterwards 115
the laser Perceptive Threshold (PT) corresponding to 116
the lowest intensity at which subjects perceived at least 117
50 of the stimuli (Cruccu et al 1999 Agostino et al 118
2000) was determined In order to minimize the num- 119
ber of nociceptive stimuli the nociceptive perception 120
threshold was not assessed A range of 10ndash40 stimuli 121
(mean SD 25 plusmn 5) was used to assess the perceptive 122
threshold before and after transcranial cerebellar stim- 123
ulation Less than 10 minutes were spent to determine 124
PT in line with previous reports (Truini et al 2011) 125
After the PT assessment participants were 126
instructed to pay attention to incoming laser noci- 127
ceptive stimuli in order to verbally rate the perceived 128
intensity about 2-3 seconds after each laser stimulation 129
which was performed before tcDCS (T0) immediately 130
after its termination (T1) and 60 min later (T2) 131
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 3
A
B
1
08
06
04
02
0
Fig 1 - Current density generated by cerebellar transcranial directcurrent stimulation (cerebellar tDCS) in humans A Top panel shows(viewed from the back) the electrode positions for cerebellar tDCSB Examples of segmented tissues in two human realistic VirtualFamily models (Ella and Duke) undergoing cerebellar tDCS Simu-lations were conducted using the simulation platform SEMCAD X(by SPEAG Schmid amp Partner Engineering AG Zurich Switzer-land) a lateral view of cerebellum pons midbrain medulla blateral view of the skull c back view of the cerebellum d and elateral and inferior views of normalized current density amplitudefield distributions over cortical subcortical and brainstem regionsf back view of normalized current density amplitude field distribu-tions over the cerebellum Values are normalized with respect to themaximum of the current density amplitude in the cerebellum Thespread of the current density (J) over the occipital cortex - quantifiedas the percentage of occipital volume where the amplitude of J-fieldis greater than 70 of the peak of J in the cerebellum - was only 4for ldquoDukerdquo and much less than 1 for ldquoEllardquo (modified from Prioriet al (2014) with permission)
Participants were blinded to the tcDCS polarity 132
anodal cathodal and sham tcDCS stimulations were 133
administered in three different sessions and separated 134
by at least 1 week to avoid possible carry-over effects 135
The order of interventions was randomized and bal- 136
anced across subjects Laser stimuli of intensity two 137
and three times the PT intensity (I1 I2) were delivered 138
by an experimenter (AT) whereas the evaluation of 139
electrophysiological parameters was done by FS both 140
blinded to the tcDCS polarity B V settled the tcDCS 141
polarity 142
221 Subjective experience 143
The perceived sensation was rated on the 0ndash10 144
Visual Analogue Scale (where 0 = no sensation and 145
10 = unbearable pain the intermediate levels being 146
1 = barely perceived 2 = lightly pricking not painful 147
3 = clearly pricking not painful 4 = barely painful 148
5 = painful prompting to rub the skin 6 = very painful 149
and distressing 7 and more strongly unpleasant pain) 150
VAS was studied in each subject after 10 nociceptive 151
laser I1 and I2 stimuli (VAS 1 VAS2) In each partic- 152
ipant individual VAS values were averaged for each 153
Time 154
Laser Evoked Potentials were obtained by stim- 155
uli corresponding to two times the Perceptive value 156
according with previous literature and guidelines 157
(Truini et al 2005 2010) 158
23 Procedures 159
231 Laser evoked potentials (LEPs) 160
The methods used for laser stimulation are 161
reported in detail elsewhere (Truini et al 2005 162
2010) A neodymiumyttriumndashaluminiumndashperovskite 163
(NdYAP) laser was used (wavelength 104 m pulse 164
duration 2ndash20 ms maximum energy 7 J) The laser 165
beam was transmitted from the generator to the stim- 166
ulating probe via a 10 m length optical fibre signals 167
were then amplified band pass filtered (01ndash200 Hz 168
time analysis 1000 ms) and fed to a computer for stor- 169
age and analysis (Cruccu et al 2008) The dorsum of 170
the left hand was stimulated by laser pulses (individ- 171
ual variability 389ndash1575 Jcm2) with short duration 172
(5 ms) and small diameter spots (5 mm Valeriani et al 173
2012) Ten stimuli whose intensity was established 174
on the basis of the Perceptive Threshold assessed for 175
each subject at T0 T1 and T2 were delivered and 176
the laser beam was shifted slightly between consec- 177
utive pulses to avoid skin lesions and reduce fatigue 178
Unc
orre
cted
Aut
hor P
roof
4 T Bocci et al Cerebellum and pain A tcDCS study
Fig 2 ndash Experimental protocol Psychophysical and electrophysiological variables evaluated at baseline (T0) and at two different time points(T1 T2) following anodal cathodal and sham tcDCS
of peripheral nociceptors (Truini et al 2005) The179
inter-stimulus interval was varied randomly (10ndash15 s)180
Participants were reclined on a couch and wore protec-181
tive goggles They were instructed to keep their eyes182
open and gaze slightly downwards since the N2P2183
amplitude is enhanced by attention (Lorenz amp Garcia-184
Larrea 2003 Truini et al 2005) they were requested185
to mentally count the number of stimuli The main186
Aδ-LEP vertex complex N2ndashP2 and the lateralised187
N1 component were recorded through standard disc188
non-polarizable AgAgCl surface electrodes (diameter189
10 mm Biomedreg Florence Italy) N2 and P2 compo-190
nents were recorded from the vertex (Cz) referenced191
to the earlobes the N1 component was recorded from192
the temporal leads (T4) referenced to Fz (Cruccu et al193
2008) Blinks and saccades were recorded with an EOG194
electrode placed on the supero-lateral right canthus195
connected to the system reference Ground was placed196
on the mid-forehead Skin impedance was kept below197
5 k198
232 Cerebellar transcutaneous direct current199
stimulation (tcDCS)200
tDCS was applied using a battery-driven constant201
current stimulator (HDCStim Newronika Italy) and202
a pair of electrodes in two saline-soaked synthetic203
sponges with a surface area of 25 cm2 For cathodal204
stimulation the cathode was centered on the median205
line 2 cm below the inion with its lateral borders about206
1 cm medially to the mastoid apophysis and the anode 207
over the right shoulder (Ferrucci et al 2008 2012 208
2013) For anodal stimulation the current flow was 209
reversed In the real tcDCS conditions direct current 210
was transcranially applied for 20 minutes with an inten- 211
sity of 20 mA and constant current flow was measured 212
by an ampere meter (current density asymp 008 mAcm2) 213
These values are similar to those previously reported 214
for cerebellar stimulation (Ferrucci et al 2008 2013) 215
are considered to be safe (Iyer et al 2005) and are 216
far below the threshold for tissue damage (Nitsche 217
et al 2003) Apart from occasional and short-lasting 218
tingling and burning sensations below the electrodes 219
direct current stimulation strength remained below the 220
sensory threshold throughout the experimental session 221
At the offset of tDCS the current was decreased in a 222
ramp-like manner a method shown to achieve a good 223
level of blinding among sessions (Gandiga et al 2006 224
Galea et al 2009) For a sham tDCS the current was 225
turned on only for 5 seconds at the beginning of the 226
sham session and then it was turned off in a ramp- 227
shaped fashion which induces initial skin sensations 228
indistinguishable from real tDCS 229
For all the electrophysiological recordings we chose 230
the left side to avoid interference from the return 231
electrode placed over the contralateral shoulder At 232
experimental debriefing subjects were not able to dis- 233
criminate between the applied anodal cathodal and 234
sham tDCS 235
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 5
Table 1
Row data (expressed as mean value plusmn 1 standard deviation a = anodal stimulation c = cathodal stimulation sh = sham condition)Both psychophysical and electrophysiological data for each subject are fully available as supplementary electronic material at
httpwwwenricasantarcangelocompublications
aT0 aT1 aT2 cT0 cT1 cT2 shT0 shT1 shT2
PT mean 462 607 609 485 376 368 472 466 489SD 080 095 092 086 062 067 098 062 081
VAS I1 mean 389 255 265 367 493 467 387 393 387SD 084 057 062 082 096 082 074 070 092
VAS I2 mean 540 402 403 524 673 665 533 549 530SD 063 082 071 048 047 049 078 069 064
N1 amplitude (V) mean 1292 848 801 1104 1496 1494 1101 1111 1121SD 318 298 258 265 258 333 250 267 283
N1 latency (ms) mean 12419 16146 15710 12704 10715 10405 12817 12867 13066SD 1090 1338 1368 1075 675 912 1320 1271 1209
N2P2 amplitude(V) mean 1114 738 757 1052 1453 1375 1114 1125 1147SD 262 237 233 265 296 329 272 269 216
N2P2 latency (ms) mean 15157 18932 18726 14878 12673 13230 15390 15108 15551SD 1312 1749 2139 2201 1849 1870 1433 1507 1675
233 Transcranial magnetic stimulation (TMS)236
Changes in Resting Motor Threshold (RMT) were237
evaluated at different intervals before and after the238
completion of tcDCS A Magstim Super Rapid Tran-239
scranial Magnetic Stimulator (Magstim Company240
Dyfed UK 22 T maximum field output) connected241
to a standard eight-shaped focal coil with wing diame-242
ters of 70 mm was used The handle of the eight-shaped243
focal coil was pointed backwards and rotated about 45244
deg to the mid-sagittal line to induce a tissue current245
perpendicular to the motor strip in the precentral sul-246
cus (Rossi et al 2009 Groppa et al 2012) RMT247
was defined as the minimum stimulator output that248
induces motor evoked potentials (MEPs) of more than249
50 V in at least five out of 10 trials when first dig-250
ital interosseus (FDI) muscle was completely relaxed251
(Ni et al 2007) The motor ldquohot spotrdquo for the targeted252
muscle was identified by single pulses of TMS deliv-253
ered at a slightly suprathreshold stimulus intensity and254
the magnetic stimuli induced monophasic pulses The255
coil was placed over the right motor cortex (centered on256
C4 according with the 10ndash20 EEG International Sys-257
tem) and electromyographic recordings were made by258
two standard non-polarizable AgAgCl surface elec-259
trodes (diameter 10 mm Biomedreg Florence Italy)260
one placed over the belly of the contralateral FDI261
muscle and the other on the skin overlying the first262
metacarpophalangeal joint of the first finger of the left263
hand RMT was evaluated to exclude possible cere-264
bellar stimulation spread out inducing motor cortex265
activation266
24 Variables and statistical analysis 267
We studied the subjective experience - percep- 268
tive threshold (PT) and pain intensity perceived 269
after laser I1 and I2 (VAS1 VAS2) - and elec- 270
trophysiological variables that is the peak-to-peak 271
amplitude of the N1 wave and N2P2 complex 272
the peak latency of N1 and N2 as reported in 273
previous papers using NdYAG laser (Lefaucheur 274
et al 2001 2002)275
Analyses were performed through SPSS15 sta- 276
tistical Package Psychophysical (PT VAS1 VAS2) 277
and electrophysiological variables (mean values of 278
ten traces N1amplitude and latency N2P2 ampli- 279
tude and latency) as well as Resting Motor Thresholds 280
(RMT) were analysed following a 3 Stimulation con- 281
ditions (anodal cathodal sham) times 3 Times (T0 T1 282
T2) design The Greenhouse-Geisser correction for 283
non sphericity was applied when necessary Con- 284
trast analysis between Times (F values) and paired 285
t tests between stimulations were alternatively used 286
for post-hoc comparisons when appropriate After 287
Bonferroni correction significance level was set at 288
p lt 0007289
The changes of all variable in T1 and T2 290
were expressed as ratio between post and pre 291
stimulation values (T1T0 T2T0) and compared 292
between each other according to a 2 Stimula- 293
tion (anodal cathodal) times 2 Times (T1To T2T0) 294
design 295
Unc
orre
cted
Aut
hor P
roof
6 T Bocci et al Cerebellum and pain A tcDCS study
Table 2
Contrast analyses all comparisons were highly significant(p lt 00001)
anodal cathodal shamPT time df F = 4430 F = 1867 ns
T0 vs Tl 228 F = 77669) F = 27523T0 vs T2 114 F = 78745 F = 27827Tl vs T2 114 ns ns
anodal vs sham cathodal vs shamT0 114 ns nsTl 114 t = 5069 t = 6991T2 114 t = 3709 t = 5849
VAS anodal cathodal sham
time 228 F = 41954 F = 31448 nsT0 vs Tl 114 F = 56968 F = 48596T0 vs T2 114 F = 52289 F = 525Tl vs T2 ns ns
anodal vs sham cathodal vs shamT0 114 ns nsTl 114 t = 644 t = 5916T2 114 t = 5294 t = 582
3 Results296
Row data (mean SD) are shown in Table 1 Base-297
line values were in line with those reported by earlier298
studies performed by using NdYAG laser (Lefaucheur299
et al 2001) Indeed only one study described a longer300
latency of the N2 wave (Cruccu et al 2008) The301
sham stimulation did not modulate any psychophys-302
ical and electrophysiological variable (Table 2) Since303
no pre-post difference was found for sham polarity this304
condition was not included in the comparison between305
Stimulations and Times306
31 Psychophysics307
PT exhibited a significant Stimulation effect308
(F(228) = 35055 p lt 00001 η2 = 0715) and a signifi-309
cant Stimulation times Time interaction (F(456) = 39464310
p lt 00001 η2 = 0738) Decomposition of the latter311
(Table 2) revealed that a) PT was higher for the anodal312
and lower for the cathodal stimulation conditions com-313
pared with the sham stimulation for both T1 and T2314
b) with respect to T0 PT increased in T1 and T2 in the315
anodal condition and decreased in the cathodal con-316
dition while no significant difference was observed317
between T1 and T2 (Fig 3A)318
Significantly different VAS1 and VAS2 were319
observed for the two stimulation intensities (F(114)320
= 54262 p lt 00001) and the three Stimulation con-321
ditions (F(218) = 88882 p lt 00001) Decomposition322
of the significant Stimulation times Time interaction323
(F(456) = 11596 p lt 00001) revealed that the reported 324
pain intensity for both stimulation intensities (VAS1 325
and VAS2) was higher for the cathodal and lower for 326
the anodal stimulation compared to the sham stimula- 327
tion (Table 2) It increased in T1 and decreased in T2 328
with respect to T0 whereas no significant difference 329
was found between T1 and T2 (Fig 3-B) 330
32 Laser evoked potentials 331
Figure 4-A shows the LEPs recorded in all experi- 332
mental conditions in a representative subject Both N1 333
and N2P2 amplitude (N1 F(456) = 10695 p lt 00001 334
η2 = 0884 N2P2 F(456) = 86864 p lt 00001 η2 = 335
0861) and latency (N1 F(456) = 110869 p lt 00001 336
η2 = 0888 N2P2 F(456) = 3660 p lt 00001 337
η2 = 0723) exhibited a significant Time times Stimulation 338
interaction Its decomposition (Table 3) showed 339
that both amplitudes increased and both latencies 340
decreased for cathodal stimulation in T1 and T2 with 341
respect to T0 the opposite occurred for the anodal 342
stimulation Both stimulations induced responses 343
significantly different from the sham condition 344
(Fig 4-B) The responses obtained after cathodal 345
stimulation were significantly improved (higher 346
amplitudes lower latencies) than those produced 347
by the anodal one (N1 amplitude F(114) = 41345 348
p lt 00001 N1 latency F(114) = 496228 p lt 00001 349
N2P2 amplitude (F(114) = 44537 p lt 00001 N2P2 350
latency F(114) = 119056 p lt 00001) 351
33 Resting motor thresholds 352
RMT values at baseline did not differ among exper- 353
imental conditions (mean plusmn SD sham 508 plusmn 83 354
anodal 491 plusmn 62 cathodal 503 plusmn 63) ANOVA 355
did not reveal any significant Stimulation (F(228) = 356
0882 p = 0425 η2 = 0059) Time (F(228) = 0212 357
p = 0810 η2 = 0015) and Stimulation times Time 358
effect (F(456) = 0339 p = 0851 η2 = 0024) for RMT 359
(Fig 5) 360
4 Discussion 361
Our study shows that cerebellar direct current 362
polarization modulates nociceptive perception and its 363
cortical correlates in healthy humans Specifically 364
cathodal tcDCS increases pain perception increases 365
amplitudes and decreases LEPs latencies likely though 366
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 7
A
B
Fig 3 - A Perceptive Threshold Changes (mean plusmn SD) at T1 and T2 with respect to baseline values (T1T0 T2T0) following sham (black)anodal (white) and cathodal (grey) tcDCS (lowastlowastp lt 0001 lowastlowastlowastp lt 00001) B Changes in visual analogue scale (VAS) scores over time VAS scoresat two different stimulus intensity respectively two (A left) and three (B right) times higher than the PT (lowastlowastp lt 0001 lowastlowastlowastp lt 00001)
A B
Fig 4 ndash A LEPs grand averaging traces were recorded at baseline (T0 black) and immediately after cerebellar polarization (T1 red) due tosham (left column) anodal (middle) and cathodal (right) tcDCS B Histograms showing LEPs variables and VAS scores changes (mean plusmn SD)after sham (black) anodal (white) or cathodal (grey) tcDCS with respect to baseline Top panels changes in N1 variables (amplitude and latency)over time bottom panels changes in N2P2 complex (lowastlowastp lt 0001 lowastlowastlowastp lt 00001)
Unc
orre
cted
Aut
hor P
roof
8 T Bocci et al Cerebellum and pain A tcDCS study
Table 3
LEPs post-hoc analyses p lt 00001 for all comparison except when explicitly indicated lowastlowastp lt 0002 lowast p lt 0005
N1 amplitude latency N2P2 amplitude latency
anodal cathodal shamdf
time 228 F = 67152 F = 96489 F = 134912 F = 34946 nsT0 vs T1 114 F = 109178 F = 18815 F = 165953 F = 64281T0 vs T2 114 F = 75143 F = 167697 F = 145125 F = 37818T1 vs T2 114 ns ns ns ns
anodal cathodaltime 228 F = 102281 F = 98717 F = 6577 F = 20918 nsT0 vs T1 114 F = 511186 F = 104027 F = 144112 F = 103864T0 vs T2 114 F = 96329 F = 116841 F = 105183 F = 14012lowastlowastT1 vs T2 114 ns ns ns ns ns
anodal vs sham anodal vs shamT0 114 ns ns ns nsT1 114 ns t = 925 t = 601 6262T2 114 ns t = 8128 t = 6731 5236
cathodal cathodal vs shamT0 114 ns ns t = 3281lowast nsT1 114 t = 16594 t = 8029 t = 8262 t = 520T2 114 t = 7309 t = 12669 t = 5048 t = 5301
Fig 5 - Resting Motor Thresholds Changes (mean plusmn SD) in Resting Motor threshold (RMT) expressed as percentage of the maximumstimulator output after sham (black) anodal (white) and cathodal (grey) tcDCS with respect to baseline marked as dotted line (lowastlowastp lt 0001lowastlowastlowastp lt 00001)
reduction of the inhibitory tone exerted by the cere-367
bellum on brain targets Anodal polarization elicits368
opposite effects producing analgesia Both findings369
support the role of the cerebellum in pain control370
it is noticeable that cathodal cerebellar stimulation371
induces hyperalgesia as occurs in patients with cere-372
bellar infarction (Ruscheweyh et al 2014)373
We would like to underline that in the present study374
LEPs were obtained at laser intensities depending on375
the perceptive threshold which varied as a function376
of anodal and cathodal stimulation This means that 377
the cerebellar stimulation has not a selective analgesic 378
effect as it influences both non nociceptive and noci- 379
ceptive perception A pre-eminent analgesic cannot be 380
assessed because the nociceptive threshold was not 381
evaluated 382
As tcDCS was effective on the modulation of 383
both N1 and N2P2 components and these responses 384
are generated by parallel and partially segregated 385
spinal pathways reaching different cortical targets 386
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 9
(Valeriani et al 2007) we may suggest that the cere-387
bellum is engaged in pain processing by modulating388
the activity of both somatosensory and cingulate cor-389
tices Indeed the cerebellum is involved in both the390
sensory-discriminative and emotional dimension of391
pain (Singer et al 2004 Moriguchi et al 2007) and392
non-invasive cerebellar current stimulation may modu-393
late pain experience and the associated cortical activity394
through many not alternative mechanisms In partic-395
ular changes in N1 reflects the modulation of the396
sensory component of pain while the vertex N2P2397
represents the neural correlate of affective aspects of398
pain experience (Garcia-Larrea et al 1997 Valeriani399
et al 2007) Notably tcDCS may act not only on spinal400
nociceptive neurons but also on wide-range cortical401
networks of the pain matrix (Singer et al 2004) thus402
influencing LEPs and pain experience through both403
top-down and bottom-up mechanisms404
The present study does not allow to hypothesize405
how and where tcDCS influences the cerebellar activ-406
ity A main role of Purkinje cells has been suggested407
as their activity modulation may affect the cerebellar408
inhibitory control of the cerebral cortex (Galea et al409
2009) This would be in line with the effects elicited410
by tDCS in the cerebral cortex which are observ-411
able after both short and long term delay likely also412
interfering with long-term potentiation (LTP)-like phe-413
nomena (Hamada et al 2012 Priori et al 2014)414
Moreover prolonged spiking activity in the cerebellar415
Golgi inhibitory neurons modulates the activity of the416
Purkinje cells and could partly account for the tcDCS417
after-effects (Hull et al 2013)418
The lack of changes in RMT indicates that the anal-419
gesic effects of anodal tcDCS are due to a specific420
modulation of the cerebellar activity and not to motor421
activation On the other hand tcDCS-induced cerebel-422
lar modulation (Purpura amp McMurtry 1965) could be423
not sufficient per se to activate the cerebello ndash thalamo424
- cortical motor pathway (Galea et al 2009) thus425
the reported analgesia and its cortical correlates can-426
not be sustained by the motor cortex activation This427
view is supported by the absence of any association428
between motor symptoms and pain perception in cere-429
bellar patients (Ruscheweyh et al 2014) In the same430
line in healthy subjects it has been recently shown431
that motor task-induced increased cortical excitability432
and analgesia are not associated (Volz et al 2012)433
Indeed RMT is a highly sensitive marker of motor434
tract excitability as it reflects activation of a small435
low-threshold and slow-conducting core of pyramidal436
neurons (Hess et al 1987 Rossini amp Rossi 2007) 437
although RMT may reflect changes in the activity of 438
different central nervous system structures it has been 439
satisfactorily used to assess motor cortex excitability 440
also in cerebellar patients (Battaglia et al 2006) 441
Another critical point is the possibility to modulate 442
with tcDCS both neural correlates underlying nocicep- 443
tive processing and pain perception Previous studies 444
using tDCS over motor cortex were inconsistent among 445
each other some works suggested that tDCS is able to 446
modify pain perception (Boggio et al 2008) while 447
others showed divergent effects on psychophysical 448
and neurophysiological outcome parameters (Luedtke 449
et al 2012 Ihle et al 2014) likely due to a possi- 450
ble overestimation of the role of motor areas on pain 451
processing (Antal et al 2008) 452
Our findings cannot be compared to the results 453
obtained by other Authors In fact the unique study 454
focused on the analgesic effects of non-invasive cere- 455
bellar stimulation reported till now (Zunhammer et al 456
2011) considered only subjective pain thresholds 457
In addition it described similar analgesic effects of 458
cerebellar and neck structures repetitive transcranial 459
magnetic stimulation (rTMS) thus denying any cere- 460
bellar specificity in the observed effects and suggesting 461
that the peripheral information passing through the 462
cerebellum may be responsible for analgesia The 463
main difference between the two studies possibly 464
accounting for different results consists of the neu- 465
romodulation techniques used 466
41 Limitations of the study 467
The present study has a few limitations First our 468
findings do not allow any hypothesis on the role of 469
the cerebellum in chronic pain The observations on 470
patients with cerebellar damage (Ruscheweyh et al 471
2014) suggest that their impaired inhibitory control 472
mechanisms may be not associated with the devel- 473
opment of chronic pain Second we cannot exclude 474
the possibility that tcDCS could modulate not only the 475
cerebellum but also surrounding areas such as the peri- 476
aqueductal gray However recent modelling researches 477
have revealed that during tcDCS the current spread 478
to other structures outside the cerebellum is negligi- 479
ble (Parazzini et al 2013 2014) Moreover several 480
studies have proved that in humans pain processing 481
is encoded within posterior areas of each cerebellar 482
hemisphere specifically in the hemispheric lobule VI 483
Crus I and VIIb (Moulton et al 2011) where the 484
Unc
orre
cted
Aut
hor P
roof
10 T Bocci et al Cerebellum and pain A tcDCS study
tcDCS-induced electrical field is strongly concentrated485
(Parazzini et al 2013) A further limitation is that we486
did not study the contribution of C-fibers the main487
component of spino-ponto-cerebellar and spino-olivo-488
cerebellar pathways In fact ultra-late LEPs related489
to C-fibers activations have not yet been standard-490
ized for clinical application and their occurrence could491
be markedly influenced by high order cognitive pro-492
cesses as they seem to be more affected by the level493
of consciousness and attention than A-delta responses494
(Qiu et al 2002 Opsommer et al 2003 Mouraux amp495
Plaghki 2006) Finally we wish to emphasize that in496
neuropathic patients the effects of the cerebellar stim-497
ulation could be quite different from those described498
here as both anatomical and functional connectivity499
are different from those observed in healthy partici-500
pants (Rocca et al 2010 Riedl et al 2011 Absinta et501
al 2012 Longo et al 2012 Ceko et al 2013)502
5 Conclusions503
Our findings indicate a cerebellar effect on pain504
experience and on its cortical correlates and prompt505
further investigation aimed at assessing whether the506
cerebellar direct current polarization could be used as a507
novel and safe therapeutic tool in chronic pain patients508
Acknowledgments509
We gratefully acknowledge the participation of all510
subjects as well as Mr D Barloscio and Mr C511
Orsini for their excellent technical assistance The512
paper was supported in part by the Italian operating513
and development MIUR PRIN grant year 2006 n514
2006062332 002515
RF and AP are founders and shareholders of516
Newronika srl Milan Italy517
References518
Absinta M Rocca MA Colombo B Falini A Comi G amp Fil-519
ippi M (2012) Selective decreased grey matter volume of the520
pain-matrix network in cluster headache Cephalalgia 32(2)521
109-115522
Agostino R Cruccu G Iannetti G Romaniello A Truini A523
amp Manfredi M (2000) Topographical distribution of pinprick524
and warmth thresholds to CO2 laser stimulation on the human525
skin Neurosci Lett 285(2) 115-118526
Antal A Brepohl N Poreisz C Boros K Csifcsak G amp 527
Paulus W (2008) Transcranial direct current stimulation over 528
somatosensory cortex decreases experimentally induced acute 529
pain perception Clin J Pain 24(1) 56-63 530
Antal A amp Paulus W (2010) Effects of transcranial theta-burst 531
stimulation on acute pain perception Restor Neurol Neurosci 532
28(4) 477-484 533
Apkarian AV Bushnell MC Treede RD amp Zubieta JK 534
(2005) Human brain mechanisms of pain perception and reg- 535
ulation in health and disease Eur J Pain 9(4) 463-484 536
Balsters JH Whelan CD Robertson IH amp Ramnani N (2013) 537
Cerebellum and cognition Evidence for the encoding of higher 538
order rules Cereb Cortex 23(6) 1433-1443 539
Battaglia F Quartarone A Ghilardi MF Dattola R Bagnato 540
S Rizzo V Morgante L amp Girlanda P (2006) Unilat- 541
eral cerebellar stroke disrupts movement preparation and motor 542
imagery Clin Neurophysiol 117(5) 1009-1016 543
Bingel U Quante M Knab R Bromm B Weiller C amp Buchel 544
C (2002) Subcortical structures involved in pain processing 545
Evidence from single-trial fMRI Pain 99(1-2) 313-321 546
Boggio PS Zaghi S amp Fregni F (2009) Modulation of emotions 547
associated with images of human pain using anodal transcranial 548
direct current stimulation (tDCS) Neuropsychologia 47(1) 549
212-217 550
Boggio PS Zaghi S Lopes M amp Fregni F (2008) Modulatory 551
effects of anodal transcranial direct current stimulation on per- 552
ception and pain thresholds in healthy volunteers Eur J Neurol 553
15(10) 1124-1130 554
Borsook D Moulton EA Tully S Schmahmann JD amp 555
Becerra L (2008) Human cerebellar responses to brush and 556
heat stimuli in healthy and neuropathic pain subjects Cerebel- 557
lum 7(3) 252-272 558
Ceko M Seminowicz DA Bushnell MC amp Olausson HW 559
(2013) Anatomical and functional enhancements of the insula 560
after loss of large primary somatosensory fibers Cereb Cortex 561
23(9) 2017-2024 562
Cruccu G Aminoff MJ Curio G Guerit JM Kakigi 563
R Mauguiere F Rossini PM Treede RD amp Garcia- 564
Larrea L (2008) Recommendations for the clinical use of 565
somatosensory-evoked potentials Clin Neurophysiol 119(8) 566
1705-1719 567
Cruccu G Romaniello A Amantini A Lombardi M Innocenti 568
P amp Manfredi M (1999) Assessment of trigeminal small- 569
fiber function Brain and reflex responses evoked by CO2-laser 570
stimulation Muscle Nerve 22(4) 508-516 571
Dasilva AF Mendonca ME Zaghi S Lopes M Dossantos 572
MF Spierings EL Bajwa Z Datta A Bikson M amp 573
Fregni F (2012) tDCS-induced analgesia and electrical fields 574
in pain-related neural networks in chronic migraine Headache 575
52(8) 1283-1295 576
Duerden EG amp Albanese MC (2013) Localization of pain- 577
related brain activation A meta-analysis of neuroimaging data 578
Hum Brain Mapp 34(1) 109-149 579
Ekerot CF Garwicz M amp Schouenborg J (1991a) The postsy- 580
naptic dorsal column pathway mediates cutaneous nociceptive 581
information to cerebellar climbing fibres in the cat J Physiol 582
441 275-284 583
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 11
Ekerot CF Garwicz M amp Schouenborg J (1991b) Topography584
and nociceptive receptive fields of climbing fibres projecting to585
the cerebellar anterior lobe in the cat J Physiol 441 257-274586
Ekerot CF Gustavsson P Oscarsson O amp Schouenborg J587
(1987a) Climbing fibres projecting to cat cerebellar anterior588
lobe activated by cutaneous A and C fibres J Physiol 386589
529-538590
Ekerot CF Oscarsson O amp Schouenborg J (1987b) Stimula-591
tion of cat cutaneous nociceptive C fibres causing tonic and592
synchronous activity in climbing fibres J Physiol 386 539-593
546594
Ferrucci R Brunoni AR Parazzini M Vergari M Rossi E595
Fumagalli M Mameli F Rosa M Giannicola G Zago S596
amp Priori A (2013) Modulating human procedural learning by597
cerebellar transcranial direct current stimulation Cerebellum598
12(4) 485-492599
Ferrucci R Giannicola G Rosa M Fumagalli M Boggio600
PS Hallett M Zago S amp Priori A (2012) Cerebellum601
and processing of negative facial emotions Cerebellar tran-602
scranial DC stimulation specifically enhances the emotional603
recognition of facial anger and sadness Cogn Emot 26(5)604
786-799605
Ferrucci R Marceglia S Vergari M Cogiamanian F Mrakic-606
Sposta S Mameli F Zago S Barbieri S amp Priori607
A (2008) Cerebellar transcranial direct current stimulation608
impairs the practice-dependent proficiency increase in working609
memory J Cogn Neurosci 20(9) 1687-1697610
Fregni F Freedman S amp Pascual-Leone A (2007) Recent611
advances in the treatment of chronic pain with non-invasive612
brain stimulation techniques Lancet Neurol 6(2) 188-191613
Galea JM Jayaram G Ajagbe L amp Celnik P (2009)614
Modulation of cerebellar excitability by polarity-specific615
noninvasive direct current stimulation J Neurosci 29(28)616
9115-9122617
Galea JM Vazquez A Pasricha N de Xivry JJ amp Celnik618
P (2011) Dissociating the roles of the cerebellum and motor619
cortex during adaptive learning The motor cortex retains what620
the cerebellum learns Cereb Cortex 21(8) 1761-1770621
Gandiga PC Hummel FC amp Cohen LG (2006) Tran-622
scranial DC stimulation (tDCS) A tool for double-blind623
sham-controlled clinical studies in brain stimulation Clin Neu-624
rophysiol 117(4) 845-850625
Garcia-Larrea L Peyron R Laurent B amp Mauguiere F (1997)626
Association and dissociation between laser-evoked potentials627
and pain perception Neuroreport 8(17) 3785-3789628
Grimaldi G Argyropoulos GP Boehringer A Celnik P629
Edwards MJ Ferrucci R Galea JM Groiss SJ Hiraoka630
K Kassavetis P Lesage E Manto M Miall RC Priori A631
Sadnicka A Ugawa Y amp Ziemann U (2014) Non-invasive632
cerebellar stimulationndasha consensus paper Cerebellum 13(1)633
121-138634
Groppa S Oliviero A Eisen A Quartarone A Cohen LG635
Mall V Kaelin-Lang A Mima T Rossi S Thickbroom636
GW Rossini PM Ziemann U Valls-Sole J amp Sieb-637
ner HR (2012) A practical guide to diagnostic transcranial638
magnetic stimulation Report of an IFCN committee Clin Neu-639
rophysiol 123(5) 858-882640
Hamada M Strigaro G Murase N Sadnicka A Galea JM 641
Edwards MJ amp Rothwell JC (2012) Cerebellar modulation 642
of human associative plasticity J Physiol 590(Pt 10) 2365- 643
2374 644
Hess CW Mills KR amp Murray NM (1987) Responses in small 645
hand muscles from magnetic stimulation of the human brain J 646
Physiol 388 397-419 647
Hull CA Chu Y Thanawala M amp Regehr WG (2013) Hyper- 648
polarization induces a long-term increase in the spontaneous 649
firing rate of cerebellar Golgi cells J Neurosci 33(14) 5895- 650
5902 651
Ihle K Rodriguez-Raecke R amp Luedtke K May A (2014) 652
tDCS modulates cortical nociceptive processing but has little 653
to no impact on pain perception Pain 155(10) 2080-2087 654
Ito M (2006) Cerebellar circuitry as a neuronal machine Prog 655
Neurobiol 78(3-5) 272-303 656
Iyer MB Mattu U Grafman J Lomarev M Sato S amp Wasser- 657
mann EM (2005) Safety and cognitive effect of frontal DC 658
brain polarization in healthy individuals Neurology 64(5) 872- 659
875 660
Kelly RM amp Strick PL (2003) Cerebellar loops with motor cor- 661
tex and prefrontal cortex of a nonhuman primate J Neurosci 662
23(23) 8432-8444 663
Lefaucheur JP Andre-Obadia N Antal A Ayache SS Baeken 664
C Benninger DH Cantello RM Cincotta M de Carvalho 665
M De Ridder D Devanne H Di Lazzaro V Filipovic SR 666
Hummel FC Jaaskelainen SK Kimiskidis VK Koch G 667
Langguth B Nyffeler T Oliviero A Padberg F Poulet E 668
Rossi S Rossini PM Rothwell JC Schonfeldt-Lecuona 669
C Siebner HR Slotema CW Stagg CJ Valls-Sole J 670
Ziemann U Paulus W amp Garcia-Larrea L (2014) Evidence- 671
based guidelines on the therapeutic use of repetitive transcranial 672
magnetic stimulation (rTMS) Clin Neurophysiol 125(11) 673
2150-2206 674
Lefaucheur JP Brusa A Creange A Drouot X amp Jarry G 675
(2002) Clinical application of laser evoked potentials using 676
the NdYAG laser Neurophysiol Clin 32(2) 91-98 677
Lefaucheur JP Debray S amp Jarry G (2001) Laser evoked poten- 678
tials using the NdYAG laser Muscle Nerve 24(4) 496-501 679
Lefaucheur JP Drouot X Menard-Lefaucheur I Keravel Y 680
amp Nguyen JP (2006) Motor cortex rTMS restores defective 681
intracortical inhibition in chronic neuropathic pain Neurology 682
67(9) 1568-1574 683
Longo MR Iannetti GD Mancini F Driver J amp Haggard P 684
(2012) Linking pain and the body Neural correlates of visually 685
induced analgesia J Neurosci 32(8) 2601-2607 686
Lorenz J amp Garcia-Larrea L (2003) Contribution of attentional 687
and cognitive factors to laser evoked brain potentials Neuro- 688
physiol Clin 33(6) 293-301 689
Lorenz J Minoshima S amp Casey KL (2003) Keeping pain out 690
of mind The role of the dorsolateral prefrontal cortex in pain 691
modulation Brain 126(Pt 5) 1079-1091 692
Luedtke K May A amp Jurgens TP (2012) No effect of a single 693
session of transcranial direct current stimulation on experimen- 694
tally induced pain in patients with chronic low back painndashan 695
exploratory study PLoS One 7(11) e48857 696
Unc
orre
cted
Aut
hor P
roof
12 T Bocci et al Cerebellum and pain A tcDCS study
Mendonca ME Santana MB Baptista AF Datta A Bikson697
M Fregni F amp Araujo CP (2011) Transcranial DC stimu-698
lation in fibromyalgia Optimized cortical target supported by699
high-resolution computational models J Pain 12(5) 610-617700
Moriguchi Y Decety J Ohnishi T Maeda M Mori T Nemoto701
K Matsuda H amp Komaki G (2007) Empathy and judg-702
ing otherrsquos pain An fMRI study of alexithymia Cereb Cortex703
17(9) 2223-2234704
Moulton EA Elman I Pendse G Schmahmann J Becerra L705
amp Borsook D (2011) Aversion-related circuitry in the cere-706
bellum Responses to noxious heat and unpleasant images J707
Neurosci 31(10) 3795-3804708
Moulton EA Schmahmann JD Becerra L amp Borsook D709
(2010) The cerebellum and pain Passive integrator or active710
participator Brain Res Rev 65(1) 14-27711
Mouraux A amp Plaghki L (2006) Are the processes reflected by712
late and ultra-late laser evoked potentials specific of nocicep-713
tion Suppl Clin Neurophysiol 59 197-204714
Mylius V Jung M Menzler K Haag A Khader PH Oer-715
tel WH Rosenow F amp Lefaucheur JP (2012) Effects of716
transcranial direct current stimulation on pain perception and717
working memory Eur J Pain 16(7) 974-982718
Ni Z Gunraj C amp Chen R (2007) Short interval intracortical719
inhibition and facilitation during the silent period in human J720
Physiol 583(Pt 3) 971-982721
Nitsche MA Liebetanz D Lang N Antal A Tergau F amp722
Paulus W (2003) Safety criteria for transcranial direct cur-723
rent stimulation (tDCS) in humans Clin Neurophysiol 114(11)724
2220-2222 author reply 2222-2223725
Opsommer E Guerit JM amp Plaghki L (2003) Exogenous726
and endogenous components of ultralate (C-fibre) evoked727
potentials following CO2 laser stimuli to tiny skin sur-728
face areas in healthy subjects Neurophysiol Clin 33(2)729
78-85730
Parazzini M Fiocchi S Liorni I Priori A amp Ravazzani P731
(2014) Computational modeling of transcranial direct current732
stimulation in the child brain Implications for the treatment of733
refractory childhood focal epilepsy Int J Neural Syst 24(2)734
1430006735
Parazzini M Rossi E Ferrucci R Fiocchi S Liorni I Priori736
A amp Ravazzani P (2013) Computational model of cerebellar737
transcranial direct current stimulation Conf Proc IEEE Eng738
Med Biol Soc 2013 237-240739
Parazzini M Rossi E Ferrucci R Liorni I Priori A amp Ravaz-740
zani P (2014) Modelling the electric field and the current741
density generated by cerebellar transcranial DC stimulation in742
humans Clin Neurophysiol 125(3) 577-584743
Perini I Bergstrand S amp Morrison I (2013) Where pain meets744
action in the human brain J Neurosci 33(40) 15930-15939745
Ploghaus A Tracey I Gati JS Clare S Menon RS Matthews746
PM amp Rawlins JN (1999) Dissociating pain from its antic-747
ipation in the human brain Science 284(5422) 1979-1981748
Priori A Ciocca M Parazzini M Vergari M amp Ferrucci R749
(2014) Transcranial cerebellar direct current stimulation and750
transcutaneous spinal cord direct current stimulation as innova-751
tive tools for neuroscientists J Physiol 592(Pt 16) 3345-3369752
Purpura DP amp McMurtry JC (1965) Intracellular activities and 753
evoked potential changes during polarization of motor cortex 754
J Neurophysiol 28 166-185 755
Qiu Y Inui K Wang X Tran TD amp Kakigi R (2002) Effects 756
of attention distraction and sleep on CO(2) laser evoked poten- 757
tials related to C-fibers in humans Clin Neurophysiol 113(10) 758
1579-1585 759
Reidler JS Mendonca ME Santana MB Wang X Lenkinski 760
R Motta AF Marchand S Latif L amp Fregni F (2012) 761
Effects of motor cortex modulation and descending inhibitory 762
systems on pain thresholds in healthy subjects J Pain 13(5) 763
450-458 764
Riedl V Valet M Woller A Sorg C Vogel D Sprenger 765
T Boecker H Wohlschlager AM amp Tolle TR (2011) 766
Repeated pain induces adaptations of intrinsic brain activity 767
to reflect past and predict future pain Neuroimage 57(1) 206- 768
213 769
Rocca MA Valsasina P Absinta M Colombo B Barcella V 770
Falini A Comi G amp Filippi M (2010) Central nervous 771
system dysregulation extends beyond the pain-matrix network 772
in cluster headache Cephalalgia 30(11) 1383-1391 773
Rossi S Hallett M Rossini PM amp Pascual-Leone A (2009) 774
Safety ethical considerations and application guidelines for 775
the use of transcranial magnetic stimulation in clinical practice 776
and research Clin Neurophysiol 120(12) 2008-2039 777
Rossini PM amp Rossi S (2007) Transcranial magnetic stimulation 778
Diagnostic therapeutic and research potential Neurology 779
68(7) 484-488 780
Ruscheweyh R Kuhnel M Filippopulos F Blum B Eggert 781
T amp Straube A (2014) Altered experimental pain perception 782
after cerebellar infarction Pain 155(7) 1303-1312 783
Schmahmann JD (1991) An emerging concept The cerebellar 784
contribution to higher function Arch Neurol 48(11) 1178- 785
1187 786
Singer T Seymour B OrsquoDoherty J Kaube H Dolan RJ amp 787
Frith CD (2004) Empathy for pain involves the affective but 788
not sensory components of pain Science 303(5661) 1157- 789
1162 790
Smith MJ Keel JC Greenberg BD Adams LF Schmidt PJ 791
Rubinow DA amp Wassermann EM (1999) Menstrual cycle 792
effects on cortical excitability Neurology 53(9) 2069-2072 793
Stoodley CJ amp Schmahmann JD (2009) Functional topography 794
in the human cerebellum A meta-analysis of neuroimaging 795
studies Neuroimage 44(2) 489-501 796
Strick PL Dum RP amp Fiez JA (2009) Cerebellum and non- 797
motor function Annu Rev Neurosci 32 413-434 798
Strigo IA Duncan GH Boivin M amp Bushnell MC (2003) 799
Differentiation of visceral and cutaneous pain in the human 800
brain J Neurophysiol 89(6) 3294-3303 801
Truini A Galeotti F Romaniello A Virtuoso M Iannetti GD 802
amp Cruccu G (2005) Laser-evoked potentials Normative val- 803
ues Clin Neurophysiol 116(4) 821-826 804
Truini A Panuccio G Galeotti F Maluccio MR Sartucci F 805
Avoli M amp Cruccu G (2010) Laser-evoked potentials as a 806
tool for assessing the efficacy of antinociceptive drugs Eur J 807
Pain 14(2) 222-225 808
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 13
Truini A Vergari M Biasiotta A La Cesa S Gabriele M Di809
Stefano G Cambieri C Cruccu G Inghilleri M amp Pri-810
ori A (2011) Transcutaneous spinal direct current stimulation811
inhibits nociceptive spinal pathway conduction and increases812
pain tolerance in humans Eur J Pain 15(10) 1023-1027813
Valeriani M Le Pera D Restuccia D de Armas L Mili-814
ucci R Betti V Vigevano F amp Tonali P (2007) Parallel815
spinal pathways generate the middle-latency N1 and the late P2816
components of the laser evoked potentials Clin Neurophysiol817
118(5) 1097-1104818
Valeriani M Pazzaglia C Cruccu G amp Truini A (2012) Clinical819
usefulness of laser evoked potentials Neurophysiol Clin 42(5)820
345-353821
Volz MS Mendonca M Pinheiro FS Cui H Santana M amp822
Fregni F (2012) Dissociation of motor task-induced cortical823
excitability and pain perception changes in healthy volunteers824
PLoS One 7(3) e34273825
Wager TD Rilling JK Smith EE Sokolik A Casey KL 826
Davidson RJ Kosslyn SM Rose RM amp Cohen JD 827
(2004) Placebo-induced changes in FMRI in the anticipation 828
and experience of pain Science 303(5661) 1162-1167 829
Zaghi S Thiele B Pimentel D Pimentel T amp Fregni 830
F (2011) Assessment and treatment of pain with non- 831
invasive cortical stimulation Restor Neurol Neurosci 29(6) 832
439-451 833
Zubieta JK Bueller JA Jackson LR Scott DJ Xu Y 834
Koeppe RA Nichols TE amp Stohler CS (2005) Placebo 835
effects mediated by endogenous opioid activity on mu-opioid 836
receptors J Neurosci 25(34) 7754-7762 837
Zunhammer M Busch V Griesbach F Landgrebe M Hajak G 838
amp Langguth B (2011) rTMS over the cerebellum modulates 839
temperature detection and pain thresholds through peripheral 840
mechanisms Brain Stimul 4(4) 210-7 e1 841

Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 3
A
B
1
08
06
04
02
0
Fig 1 - Current density generated by cerebellar transcranial directcurrent stimulation (cerebellar tDCS) in humans A Top panel shows(viewed from the back) the electrode positions for cerebellar tDCSB Examples of segmented tissues in two human realistic VirtualFamily models (Ella and Duke) undergoing cerebellar tDCS Simu-lations were conducted using the simulation platform SEMCAD X(by SPEAG Schmid amp Partner Engineering AG Zurich Switzer-land) a lateral view of cerebellum pons midbrain medulla blateral view of the skull c back view of the cerebellum d and elateral and inferior views of normalized current density amplitudefield distributions over cortical subcortical and brainstem regionsf back view of normalized current density amplitude field distribu-tions over the cerebellum Values are normalized with respect to themaximum of the current density amplitude in the cerebellum Thespread of the current density (J) over the occipital cortex - quantifiedas the percentage of occipital volume where the amplitude of J-fieldis greater than 70 of the peak of J in the cerebellum - was only 4for ldquoDukerdquo and much less than 1 for ldquoEllardquo (modified from Prioriet al (2014) with permission)
Participants were blinded to the tcDCS polarity 132
anodal cathodal and sham tcDCS stimulations were 133
administered in three different sessions and separated 134
by at least 1 week to avoid possible carry-over effects 135
The order of interventions was randomized and bal- 136
anced across subjects Laser stimuli of intensity two 137
and three times the PT intensity (I1 I2) were delivered 138
by an experimenter (AT) whereas the evaluation of 139
electrophysiological parameters was done by FS both 140
blinded to the tcDCS polarity B V settled the tcDCS 141
polarity 142
221 Subjective experience 143
The perceived sensation was rated on the 0ndash10 144
Visual Analogue Scale (where 0 = no sensation and 145
10 = unbearable pain the intermediate levels being 146
1 = barely perceived 2 = lightly pricking not painful 147
3 = clearly pricking not painful 4 = barely painful 148
5 = painful prompting to rub the skin 6 = very painful 149
and distressing 7 and more strongly unpleasant pain) 150
VAS was studied in each subject after 10 nociceptive 151
laser I1 and I2 stimuli (VAS 1 VAS2) In each partic- 152
ipant individual VAS values were averaged for each 153
Time 154
Laser Evoked Potentials were obtained by stim- 155
uli corresponding to two times the Perceptive value 156
according with previous literature and guidelines 157
(Truini et al 2005 2010) 158
23 Procedures 159
231 Laser evoked potentials (LEPs) 160
The methods used for laser stimulation are 161
reported in detail elsewhere (Truini et al 2005 162
2010) A neodymiumyttriumndashaluminiumndashperovskite 163
(NdYAP) laser was used (wavelength 104 m pulse 164
duration 2ndash20 ms maximum energy 7 J) The laser 165
beam was transmitted from the generator to the stim- 166
ulating probe via a 10 m length optical fibre signals 167
were then amplified band pass filtered (01ndash200 Hz 168
time analysis 1000 ms) and fed to a computer for stor- 169
age and analysis (Cruccu et al 2008) The dorsum of 170
the left hand was stimulated by laser pulses (individ- 171
ual variability 389ndash1575 Jcm2) with short duration 172
(5 ms) and small diameter spots (5 mm Valeriani et al 173
2012) Ten stimuli whose intensity was established 174
on the basis of the Perceptive Threshold assessed for 175
each subject at T0 T1 and T2 were delivered and 176
the laser beam was shifted slightly between consec- 177
utive pulses to avoid skin lesions and reduce fatigue 178
Unc
orre
cted
Aut
hor P
roof
4 T Bocci et al Cerebellum and pain A tcDCS study
Fig 2 ndash Experimental protocol Psychophysical and electrophysiological variables evaluated at baseline (T0) and at two different time points(T1 T2) following anodal cathodal and sham tcDCS
of peripheral nociceptors (Truini et al 2005) The179
inter-stimulus interval was varied randomly (10ndash15 s)180
Participants were reclined on a couch and wore protec-181
tive goggles They were instructed to keep their eyes182
open and gaze slightly downwards since the N2P2183
amplitude is enhanced by attention (Lorenz amp Garcia-184
Larrea 2003 Truini et al 2005) they were requested185
to mentally count the number of stimuli The main186
Aδ-LEP vertex complex N2ndashP2 and the lateralised187
N1 component were recorded through standard disc188
non-polarizable AgAgCl surface electrodes (diameter189
10 mm Biomedreg Florence Italy) N2 and P2 compo-190
nents were recorded from the vertex (Cz) referenced191
to the earlobes the N1 component was recorded from192
the temporal leads (T4) referenced to Fz (Cruccu et al193
2008) Blinks and saccades were recorded with an EOG194
electrode placed on the supero-lateral right canthus195
connected to the system reference Ground was placed196
on the mid-forehead Skin impedance was kept below197
5 k198
232 Cerebellar transcutaneous direct current199
stimulation (tcDCS)200
tDCS was applied using a battery-driven constant201
current stimulator (HDCStim Newronika Italy) and202
a pair of electrodes in two saline-soaked synthetic203
sponges with a surface area of 25 cm2 For cathodal204
stimulation the cathode was centered on the median205
line 2 cm below the inion with its lateral borders about206
1 cm medially to the mastoid apophysis and the anode 207
over the right shoulder (Ferrucci et al 2008 2012 208
2013) For anodal stimulation the current flow was 209
reversed In the real tcDCS conditions direct current 210
was transcranially applied for 20 minutes with an inten- 211
sity of 20 mA and constant current flow was measured 212
by an ampere meter (current density asymp 008 mAcm2) 213
These values are similar to those previously reported 214
for cerebellar stimulation (Ferrucci et al 2008 2013) 215
are considered to be safe (Iyer et al 2005) and are 216
far below the threshold for tissue damage (Nitsche 217
et al 2003) Apart from occasional and short-lasting 218
tingling and burning sensations below the electrodes 219
direct current stimulation strength remained below the 220
sensory threshold throughout the experimental session 221
At the offset of tDCS the current was decreased in a 222
ramp-like manner a method shown to achieve a good 223
level of blinding among sessions (Gandiga et al 2006 224
Galea et al 2009) For a sham tDCS the current was 225
turned on only for 5 seconds at the beginning of the 226
sham session and then it was turned off in a ramp- 227
shaped fashion which induces initial skin sensations 228
indistinguishable from real tDCS 229
For all the electrophysiological recordings we chose 230
the left side to avoid interference from the return 231
electrode placed over the contralateral shoulder At 232
experimental debriefing subjects were not able to dis- 233
criminate between the applied anodal cathodal and 234
sham tDCS 235
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 5
Table 1
Row data (expressed as mean value plusmn 1 standard deviation a = anodal stimulation c = cathodal stimulation sh = sham condition)Both psychophysical and electrophysiological data for each subject are fully available as supplementary electronic material at
httpwwwenricasantarcangelocompublications
aT0 aT1 aT2 cT0 cT1 cT2 shT0 shT1 shT2
PT mean 462 607 609 485 376 368 472 466 489SD 080 095 092 086 062 067 098 062 081
VAS I1 mean 389 255 265 367 493 467 387 393 387SD 084 057 062 082 096 082 074 070 092
VAS I2 mean 540 402 403 524 673 665 533 549 530SD 063 082 071 048 047 049 078 069 064
N1 amplitude (V) mean 1292 848 801 1104 1496 1494 1101 1111 1121SD 318 298 258 265 258 333 250 267 283
N1 latency (ms) mean 12419 16146 15710 12704 10715 10405 12817 12867 13066SD 1090 1338 1368 1075 675 912 1320 1271 1209
N2P2 amplitude(V) mean 1114 738 757 1052 1453 1375 1114 1125 1147SD 262 237 233 265 296 329 272 269 216
N2P2 latency (ms) mean 15157 18932 18726 14878 12673 13230 15390 15108 15551SD 1312 1749 2139 2201 1849 1870 1433 1507 1675
233 Transcranial magnetic stimulation (TMS)236
Changes in Resting Motor Threshold (RMT) were237
evaluated at different intervals before and after the238
completion of tcDCS A Magstim Super Rapid Tran-239
scranial Magnetic Stimulator (Magstim Company240
Dyfed UK 22 T maximum field output) connected241
to a standard eight-shaped focal coil with wing diame-242
ters of 70 mm was used The handle of the eight-shaped243
focal coil was pointed backwards and rotated about 45244
deg to the mid-sagittal line to induce a tissue current245
perpendicular to the motor strip in the precentral sul-246
cus (Rossi et al 2009 Groppa et al 2012) RMT247
was defined as the minimum stimulator output that248
induces motor evoked potentials (MEPs) of more than249
50 V in at least five out of 10 trials when first dig-250
ital interosseus (FDI) muscle was completely relaxed251
(Ni et al 2007) The motor ldquohot spotrdquo for the targeted252
muscle was identified by single pulses of TMS deliv-253
ered at a slightly suprathreshold stimulus intensity and254
the magnetic stimuli induced monophasic pulses The255
coil was placed over the right motor cortex (centered on256
C4 according with the 10ndash20 EEG International Sys-257
tem) and electromyographic recordings were made by258
two standard non-polarizable AgAgCl surface elec-259
trodes (diameter 10 mm Biomedreg Florence Italy)260
one placed over the belly of the contralateral FDI261
muscle and the other on the skin overlying the first262
metacarpophalangeal joint of the first finger of the left263
hand RMT was evaluated to exclude possible cere-264
bellar stimulation spread out inducing motor cortex265
activation266
24 Variables and statistical analysis 267
We studied the subjective experience - percep- 268
tive threshold (PT) and pain intensity perceived 269
after laser I1 and I2 (VAS1 VAS2) - and elec- 270
trophysiological variables that is the peak-to-peak 271
amplitude of the N1 wave and N2P2 complex 272
the peak latency of N1 and N2 as reported in 273
previous papers using NdYAG laser (Lefaucheur 274
et al 2001 2002)275
Analyses were performed through SPSS15 sta- 276
tistical Package Psychophysical (PT VAS1 VAS2) 277
and electrophysiological variables (mean values of 278
ten traces N1amplitude and latency N2P2 ampli- 279
tude and latency) as well as Resting Motor Thresholds 280
(RMT) were analysed following a 3 Stimulation con- 281
ditions (anodal cathodal sham) times 3 Times (T0 T1 282
T2) design The Greenhouse-Geisser correction for 283
non sphericity was applied when necessary Con- 284
trast analysis between Times (F values) and paired 285
t tests between stimulations were alternatively used 286
for post-hoc comparisons when appropriate After 287
Bonferroni correction significance level was set at 288
p lt 0007289
The changes of all variable in T1 and T2 290
were expressed as ratio between post and pre 291
stimulation values (T1T0 T2T0) and compared 292
between each other according to a 2 Stimula- 293
tion (anodal cathodal) times 2 Times (T1To T2T0) 294
design 295
Unc
orre
cted
Aut
hor P
roof
6 T Bocci et al Cerebellum and pain A tcDCS study
Table 2
Contrast analyses all comparisons were highly significant(p lt 00001)
anodal cathodal shamPT time df F = 4430 F = 1867 ns
T0 vs Tl 228 F = 77669) F = 27523T0 vs T2 114 F = 78745 F = 27827Tl vs T2 114 ns ns
anodal vs sham cathodal vs shamT0 114 ns nsTl 114 t = 5069 t = 6991T2 114 t = 3709 t = 5849
VAS anodal cathodal sham
time 228 F = 41954 F = 31448 nsT0 vs Tl 114 F = 56968 F = 48596T0 vs T2 114 F = 52289 F = 525Tl vs T2 ns ns
anodal vs sham cathodal vs shamT0 114 ns nsTl 114 t = 644 t = 5916T2 114 t = 5294 t = 582
3 Results296
Row data (mean SD) are shown in Table 1 Base-297
line values were in line with those reported by earlier298
studies performed by using NdYAG laser (Lefaucheur299
et al 2001) Indeed only one study described a longer300
latency of the N2 wave (Cruccu et al 2008) The301
sham stimulation did not modulate any psychophys-302
ical and electrophysiological variable (Table 2) Since303
no pre-post difference was found for sham polarity this304
condition was not included in the comparison between305
Stimulations and Times306
31 Psychophysics307
PT exhibited a significant Stimulation effect308
(F(228) = 35055 p lt 00001 η2 = 0715) and a signifi-309
cant Stimulation times Time interaction (F(456) = 39464310
p lt 00001 η2 = 0738) Decomposition of the latter311
(Table 2) revealed that a) PT was higher for the anodal312
and lower for the cathodal stimulation conditions com-313
pared with the sham stimulation for both T1 and T2314
b) with respect to T0 PT increased in T1 and T2 in the315
anodal condition and decreased in the cathodal con-316
dition while no significant difference was observed317
between T1 and T2 (Fig 3A)318
Significantly different VAS1 and VAS2 were319
observed for the two stimulation intensities (F(114)320
= 54262 p lt 00001) and the three Stimulation con-321
ditions (F(218) = 88882 p lt 00001) Decomposition322
of the significant Stimulation times Time interaction323
(F(456) = 11596 p lt 00001) revealed that the reported 324
pain intensity for both stimulation intensities (VAS1 325
and VAS2) was higher for the cathodal and lower for 326
the anodal stimulation compared to the sham stimula- 327
tion (Table 2) It increased in T1 and decreased in T2 328
with respect to T0 whereas no significant difference 329
was found between T1 and T2 (Fig 3-B) 330
32 Laser evoked potentials 331
Figure 4-A shows the LEPs recorded in all experi- 332
mental conditions in a representative subject Both N1 333
and N2P2 amplitude (N1 F(456) = 10695 p lt 00001 334
η2 = 0884 N2P2 F(456) = 86864 p lt 00001 η2 = 335
0861) and latency (N1 F(456) = 110869 p lt 00001 336
η2 = 0888 N2P2 F(456) = 3660 p lt 00001 337
η2 = 0723) exhibited a significant Time times Stimulation 338
interaction Its decomposition (Table 3) showed 339
that both amplitudes increased and both latencies 340
decreased for cathodal stimulation in T1 and T2 with 341
respect to T0 the opposite occurred for the anodal 342
stimulation Both stimulations induced responses 343
significantly different from the sham condition 344
(Fig 4-B) The responses obtained after cathodal 345
stimulation were significantly improved (higher 346
amplitudes lower latencies) than those produced 347
by the anodal one (N1 amplitude F(114) = 41345 348
p lt 00001 N1 latency F(114) = 496228 p lt 00001 349
N2P2 amplitude (F(114) = 44537 p lt 00001 N2P2 350
latency F(114) = 119056 p lt 00001) 351
33 Resting motor thresholds 352
RMT values at baseline did not differ among exper- 353
imental conditions (mean plusmn SD sham 508 plusmn 83 354
anodal 491 plusmn 62 cathodal 503 plusmn 63) ANOVA 355
did not reveal any significant Stimulation (F(228) = 356
0882 p = 0425 η2 = 0059) Time (F(228) = 0212 357
p = 0810 η2 = 0015) and Stimulation times Time 358
effect (F(456) = 0339 p = 0851 η2 = 0024) for RMT 359
(Fig 5) 360
4 Discussion 361
Our study shows that cerebellar direct current 362
polarization modulates nociceptive perception and its 363
cortical correlates in healthy humans Specifically 364
cathodal tcDCS increases pain perception increases 365
amplitudes and decreases LEPs latencies likely though 366
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 7
A
B
Fig 3 - A Perceptive Threshold Changes (mean plusmn SD) at T1 and T2 with respect to baseline values (T1T0 T2T0) following sham (black)anodal (white) and cathodal (grey) tcDCS (lowastlowastp lt 0001 lowastlowastlowastp lt 00001) B Changes in visual analogue scale (VAS) scores over time VAS scoresat two different stimulus intensity respectively two (A left) and three (B right) times higher than the PT (lowastlowastp lt 0001 lowastlowastlowastp lt 00001)
A B
Fig 4 ndash A LEPs grand averaging traces were recorded at baseline (T0 black) and immediately after cerebellar polarization (T1 red) due tosham (left column) anodal (middle) and cathodal (right) tcDCS B Histograms showing LEPs variables and VAS scores changes (mean plusmn SD)after sham (black) anodal (white) or cathodal (grey) tcDCS with respect to baseline Top panels changes in N1 variables (amplitude and latency)over time bottom panels changes in N2P2 complex (lowastlowastp lt 0001 lowastlowastlowastp lt 00001)
Unc
orre
cted
Aut
hor P
roof
8 T Bocci et al Cerebellum and pain A tcDCS study
Table 3
LEPs post-hoc analyses p lt 00001 for all comparison except when explicitly indicated lowastlowastp lt 0002 lowast p lt 0005
N1 amplitude latency N2P2 amplitude latency
anodal cathodal shamdf
time 228 F = 67152 F = 96489 F = 134912 F = 34946 nsT0 vs T1 114 F = 109178 F = 18815 F = 165953 F = 64281T0 vs T2 114 F = 75143 F = 167697 F = 145125 F = 37818T1 vs T2 114 ns ns ns ns
anodal cathodaltime 228 F = 102281 F = 98717 F = 6577 F = 20918 nsT0 vs T1 114 F = 511186 F = 104027 F = 144112 F = 103864T0 vs T2 114 F = 96329 F = 116841 F = 105183 F = 14012lowastlowastT1 vs T2 114 ns ns ns ns ns
anodal vs sham anodal vs shamT0 114 ns ns ns nsT1 114 ns t = 925 t = 601 6262T2 114 ns t = 8128 t = 6731 5236
cathodal cathodal vs shamT0 114 ns ns t = 3281lowast nsT1 114 t = 16594 t = 8029 t = 8262 t = 520T2 114 t = 7309 t = 12669 t = 5048 t = 5301
Fig 5 - Resting Motor Thresholds Changes (mean plusmn SD) in Resting Motor threshold (RMT) expressed as percentage of the maximumstimulator output after sham (black) anodal (white) and cathodal (grey) tcDCS with respect to baseline marked as dotted line (lowastlowastp lt 0001lowastlowastlowastp lt 00001)
reduction of the inhibitory tone exerted by the cere-367
bellum on brain targets Anodal polarization elicits368
opposite effects producing analgesia Both findings369
support the role of the cerebellum in pain control370
it is noticeable that cathodal cerebellar stimulation371
induces hyperalgesia as occurs in patients with cere-372
bellar infarction (Ruscheweyh et al 2014)373
We would like to underline that in the present study374
LEPs were obtained at laser intensities depending on375
the perceptive threshold which varied as a function376
of anodal and cathodal stimulation This means that 377
the cerebellar stimulation has not a selective analgesic 378
effect as it influences both non nociceptive and noci- 379
ceptive perception A pre-eminent analgesic cannot be 380
assessed because the nociceptive threshold was not 381
evaluated 382
As tcDCS was effective on the modulation of 383
both N1 and N2P2 components and these responses 384
are generated by parallel and partially segregated 385
spinal pathways reaching different cortical targets 386
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 9
(Valeriani et al 2007) we may suggest that the cere-387
bellum is engaged in pain processing by modulating388
the activity of both somatosensory and cingulate cor-389
tices Indeed the cerebellum is involved in both the390
sensory-discriminative and emotional dimension of391
pain (Singer et al 2004 Moriguchi et al 2007) and392
non-invasive cerebellar current stimulation may modu-393
late pain experience and the associated cortical activity394
through many not alternative mechanisms In partic-395
ular changes in N1 reflects the modulation of the396
sensory component of pain while the vertex N2P2397
represents the neural correlate of affective aspects of398
pain experience (Garcia-Larrea et al 1997 Valeriani399
et al 2007) Notably tcDCS may act not only on spinal400
nociceptive neurons but also on wide-range cortical401
networks of the pain matrix (Singer et al 2004) thus402
influencing LEPs and pain experience through both403
top-down and bottom-up mechanisms404
The present study does not allow to hypothesize405
how and where tcDCS influences the cerebellar activ-406
ity A main role of Purkinje cells has been suggested407
as their activity modulation may affect the cerebellar408
inhibitory control of the cerebral cortex (Galea et al409
2009) This would be in line with the effects elicited410
by tDCS in the cerebral cortex which are observ-411
able after both short and long term delay likely also412
interfering with long-term potentiation (LTP)-like phe-413
nomena (Hamada et al 2012 Priori et al 2014)414
Moreover prolonged spiking activity in the cerebellar415
Golgi inhibitory neurons modulates the activity of the416
Purkinje cells and could partly account for the tcDCS417
after-effects (Hull et al 2013)418
The lack of changes in RMT indicates that the anal-419
gesic effects of anodal tcDCS are due to a specific420
modulation of the cerebellar activity and not to motor421
activation On the other hand tcDCS-induced cerebel-422
lar modulation (Purpura amp McMurtry 1965) could be423
not sufficient per se to activate the cerebello ndash thalamo424
- cortical motor pathway (Galea et al 2009) thus425
the reported analgesia and its cortical correlates can-426
not be sustained by the motor cortex activation This427
view is supported by the absence of any association428
between motor symptoms and pain perception in cere-429
bellar patients (Ruscheweyh et al 2014) In the same430
line in healthy subjects it has been recently shown431
that motor task-induced increased cortical excitability432
and analgesia are not associated (Volz et al 2012)433
Indeed RMT is a highly sensitive marker of motor434
tract excitability as it reflects activation of a small435
low-threshold and slow-conducting core of pyramidal436
neurons (Hess et al 1987 Rossini amp Rossi 2007) 437
although RMT may reflect changes in the activity of 438
different central nervous system structures it has been 439
satisfactorily used to assess motor cortex excitability 440
also in cerebellar patients (Battaglia et al 2006) 441
Another critical point is the possibility to modulate 442
with tcDCS both neural correlates underlying nocicep- 443
tive processing and pain perception Previous studies 444
using tDCS over motor cortex were inconsistent among 445
each other some works suggested that tDCS is able to 446
modify pain perception (Boggio et al 2008) while 447
others showed divergent effects on psychophysical 448
and neurophysiological outcome parameters (Luedtke 449
et al 2012 Ihle et al 2014) likely due to a possi- 450
ble overestimation of the role of motor areas on pain 451
processing (Antal et al 2008) 452
Our findings cannot be compared to the results 453
obtained by other Authors In fact the unique study 454
focused on the analgesic effects of non-invasive cere- 455
bellar stimulation reported till now (Zunhammer et al 456
2011) considered only subjective pain thresholds 457
In addition it described similar analgesic effects of 458
cerebellar and neck structures repetitive transcranial 459
magnetic stimulation (rTMS) thus denying any cere- 460
bellar specificity in the observed effects and suggesting 461
that the peripheral information passing through the 462
cerebellum may be responsible for analgesia The 463
main difference between the two studies possibly 464
accounting for different results consists of the neu- 465
romodulation techniques used 466
41 Limitations of the study 467
The present study has a few limitations First our 468
findings do not allow any hypothesis on the role of 469
the cerebellum in chronic pain The observations on 470
patients with cerebellar damage (Ruscheweyh et al 471
2014) suggest that their impaired inhibitory control 472
mechanisms may be not associated with the devel- 473
opment of chronic pain Second we cannot exclude 474
the possibility that tcDCS could modulate not only the 475
cerebellum but also surrounding areas such as the peri- 476
aqueductal gray However recent modelling researches 477
have revealed that during tcDCS the current spread 478
to other structures outside the cerebellum is negligi- 479
ble (Parazzini et al 2013 2014) Moreover several 480
studies have proved that in humans pain processing 481
is encoded within posterior areas of each cerebellar 482
hemisphere specifically in the hemispheric lobule VI 483
Crus I and VIIb (Moulton et al 2011) where the 484
Unc
orre
cted
Aut
hor P
roof
10 T Bocci et al Cerebellum and pain A tcDCS study
tcDCS-induced electrical field is strongly concentrated485
(Parazzini et al 2013) A further limitation is that we486
did not study the contribution of C-fibers the main487
component of spino-ponto-cerebellar and spino-olivo-488
cerebellar pathways In fact ultra-late LEPs related489
to C-fibers activations have not yet been standard-490
ized for clinical application and their occurrence could491
be markedly influenced by high order cognitive pro-492
cesses as they seem to be more affected by the level493
of consciousness and attention than A-delta responses494
(Qiu et al 2002 Opsommer et al 2003 Mouraux amp495
Plaghki 2006) Finally we wish to emphasize that in496
neuropathic patients the effects of the cerebellar stim-497
ulation could be quite different from those described498
here as both anatomical and functional connectivity499
are different from those observed in healthy partici-500
pants (Rocca et al 2010 Riedl et al 2011 Absinta et501
al 2012 Longo et al 2012 Ceko et al 2013)502
5 Conclusions503
Our findings indicate a cerebellar effect on pain504
experience and on its cortical correlates and prompt505
further investigation aimed at assessing whether the506
cerebellar direct current polarization could be used as a507
novel and safe therapeutic tool in chronic pain patients508
Acknowledgments509
We gratefully acknowledge the participation of all510
subjects as well as Mr D Barloscio and Mr C511
Orsini for their excellent technical assistance The512
paper was supported in part by the Italian operating513
and development MIUR PRIN grant year 2006 n514
2006062332 002515
RF and AP are founders and shareholders of516
Newronika srl Milan Italy517
References518
Absinta M Rocca MA Colombo B Falini A Comi G amp Fil-519
ippi M (2012) Selective decreased grey matter volume of the520
pain-matrix network in cluster headache Cephalalgia 32(2)521
109-115522
Agostino R Cruccu G Iannetti G Romaniello A Truini A523
amp Manfredi M (2000) Topographical distribution of pinprick524
and warmth thresholds to CO2 laser stimulation on the human525
skin Neurosci Lett 285(2) 115-118526
Antal A Brepohl N Poreisz C Boros K Csifcsak G amp 527
Paulus W (2008) Transcranial direct current stimulation over 528
somatosensory cortex decreases experimentally induced acute 529
pain perception Clin J Pain 24(1) 56-63 530
Antal A amp Paulus W (2010) Effects of transcranial theta-burst 531
stimulation on acute pain perception Restor Neurol Neurosci 532
28(4) 477-484 533
Apkarian AV Bushnell MC Treede RD amp Zubieta JK 534
(2005) Human brain mechanisms of pain perception and reg- 535
ulation in health and disease Eur J Pain 9(4) 463-484 536
Balsters JH Whelan CD Robertson IH amp Ramnani N (2013) 537
Cerebellum and cognition Evidence for the encoding of higher 538
order rules Cereb Cortex 23(6) 1433-1443 539
Battaglia F Quartarone A Ghilardi MF Dattola R Bagnato 540
S Rizzo V Morgante L amp Girlanda P (2006) Unilat- 541
eral cerebellar stroke disrupts movement preparation and motor 542
imagery Clin Neurophysiol 117(5) 1009-1016 543
Bingel U Quante M Knab R Bromm B Weiller C amp Buchel 544
C (2002) Subcortical structures involved in pain processing 545
Evidence from single-trial fMRI Pain 99(1-2) 313-321 546
Boggio PS Zaghi S amp Fregni F (2009) Modulation of emotions 547
associated with images of human pain using anodal transcranial 548
direct current stimulation (tDCS) Neuropsychologia 47(1) 549
212-217 550
Boggio PS Zaghi S Lopes M amp Fregni F (2008) Modulatory 551
effects of anodal transcranial direct current stimulation on per- 552
ception and pain thresholds in healthy volunteers Eur J Neurol 553
15(10) 1124-1130 554
Borsook D Moulton EA Tully S Schmahmann JD amp 555
Becerra L (2008) Human cerebellar responses to brush and 556
heat stimuli in healthy and neuropathic pain subjects Cerebel- 557
lum 7(3) 252-272 558
Ceko M Seminowicz DA Bushnell MC amp Olausson HW 559
(2013) Anatomical and functional enhancements of the insula 560
after loss of large primary somatosensory fibers Cereb Cortex 561
23(9) 2017-2024 562
Cruccu G Aminoff MJ Curio G Guerit JM Kakigi 563
R Mauguiere F Rossini PM Treede RD amp Garcia- 564
Larrea L (2008) Recommendations for the clinical use of 565
somatosensory-evoked potentials Clin Neurophysiol 119(8) 566
1705-1719 567
Cruccu G Romaniello A Amantini A Lombardi M Innocenti 568
P amp Manfredi M (1999) Assessment of trigeminal small- 569
fiber function Brain and reflex responses evoked by CO2-laser 570
stimulation Muscle Nerve 22(4) 508-516 571
Dasilva AF Mendonca ME Zaghi S Lopes M Dossantos 572
MF Spierings EL Bajwa Z Datta A Bikson M amp 573
Fregni F (2012) tDCS-induced analgesia and electrical fields 574
in pain-related neural networks in chronic migraine Headache 575
52(8) 1283-1295 576
Duerden EG amp Albanese MC (2013) Localization of pain- 577
related brain activation A meta-analysis of neuroimaging data 578
Hum Brain Mapp 34(1) 109-149 579
Ekerot CF Garwicz M amp Schouenborg J (1991a) The postsy- 580
naptic dorsal column pathway mediates cutaneous nociceptive 581
information to cerebellar climbing fibres in the cat J Physiol 582
441 275-284 583
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 11
Ekerot CF Garwicz M amp Schouenborg J (1991b) Topography584
and nociceptive receptive fields of climbing fibres projecting to585
the cerebellar anterior lobe in the cat J Physiol 441 257-274586
Ekerot CF Gustavsson P Oscarsson O amp Schouenborg J587
(1987a) Climbing fibres projecting to cat cerebellar anterior588
lobe activated by cutaneous A and C fibres J Physiol 386589
529-538590
Ekerot CF Oscarsson O amp Schouenborg J (1987b) Stimula-591
tion of cat cutaneous nociceptive C fibres causing tonic and592
synchronous activity in climbing fibres J Physiol 386 539-593
546594
Ferrucci R Brunoni AR Parazzini M Vergari M Rossi E595
Fumagalli M Mameli F Rosa M Giannicola G Zago S596
amp Priori A (2013) Modulating human procedural learning by597
cerebellar transcranial direct current stimulation Cerebellum598
12(4) 485-492599
Ferrucci R Giannicola G Rosa M Fumagalli M Boggio600
PS Hallett M Zago S amp Priori A (2012) Cerebellum601
and processing of negative facial emotions Cerebellar tran-602
scranial DC stimulation specifically enhances the emotional603
recognition of facial anger and sadness Cogn Emot 26(5)604
786-799605
Ferrucci R Marceglia S Vergari M Cogiamanian F Mrakic-606
Sposta S Mameli F Zago S Barbieri S amp Priori607
A (2008) Cerebellar transcranial direct current stimulation608
impairs the practice-dependent proficiency increase in working609
memory J Cogn Neurosci 20(9) 1687-1697610
Fregni F Freedman S amp Pascual-Leone A (2007) Recent611
advances in the treatment of chronic pain with non-invasive612
brain stimulation techniques Lancet Neurol 6(2) 188-191613
Galea JM Jayaram G Ajagbe L amp Celnik P (2009)614
Modulation of cerebellar excitability by polarity-specific615
noninvasive direct current stimulation J Neurosci 29(28)616
9115-9122617
Galea JM Vazquez A Pasricha N de Xivry JJ amp Celnik618
P (2011) Dissociating the roles of the cerebellum and motor619
cortex during adaptive learning The motor cortex retains what620
the cerebellum learns Cereb Cortex 21(8) 1761-1770621
Gandiga PC Hummel FC amp Cohen LG (2006) Tran-622
scranial DC stimulation (tDCS) A tool for double-blind623
sham-controlled clinical studies in brain stimulation Clin Neu-624
rophysiol 117(4) 845-850625
Garcia-Larrea L Peyron R Laurent B amp Mauguiere F (1997)626
Association and dissociation between laser-evoked potentials627
and pain perception Neuroreport 8(17) 3785-3789628
Grimaldi G Argyropoulos GP Boehringer A Celnik P629
Edwards MJ Ferrucci R Galea JM Groiss SJ Hiraoka630
K Kassavetis P Lesage E Manto M Miall RC Priori A631
Sadnicka A Ugawa Y amp Ziemann U (2014) Non-invasive632
cerebellar stimulationndasha consensus paper Cerebellum 13(1)633
121-138634
Groppa S Oliviero A Eisen A Quartarone A Cohen LG635
Mall V Kaelin-Lang A Mima T Rossi S Thickbroom636
GW Rossini PM Ziemann U Valls-Sole J amp Sieb-637
ner HR (2012) A practical guide to diagnostic transcranial638
magnetic stimulation Report of an IFCN committee Clin Neu-639
rophysiol 123(5) 858-882640
Hamada M Strigaro G Murase N Sadnicka A Galea JM 641
Edwards MJ amp Rothwell JC (2012) Cerebellar modulation 642
of human associative plasticity J Physiol 590(Pt 10) 2365- 643
2374 644
Hess CW Mills KR amp Murray NM (1987) Responses in small 645
hand muscles from magnetic stimulation of the human brain J 646
Physiol 388 397-419 647
Hull CA Chu Y Thanawala M amp Regehr WG (2013) Hyper- 648
polarization induces a long-term increase in the spontaneous 649
firing rate of cerebellar Golgi cells J Neurosci 33(14) 5895- 650
5902 651
Ihle K Rodriguez-Raecke R amp Luedtke K May A (2014) 652
tDCS modulates cortical nociceptive processing but has little 653
to no impact on pain perception Pain 155(10) 2080-2087 654
Ito M (2006) Cerebellar circuitry as a neuronal machine Prog 655
Neurobiol 78(3-5) 272-303 656
Iyer MB Mattu U Grafman J Lomarev M Sato S amp Wasser- 657
mann EM (2005) Safety and cognitive effect of frontal DC 658
brain polarization in healthy individuals Neurology 64(5) 872- 659
875 660
Kelly RM amp Strick PL (2003) Cerebellar loops with motor cor- 661
tex and prefrontal cortex of a nonhuman primate J Neurosci 662
23(23) 8432-8444 663
Lefaucheur JP Andre-Obadia N Antal A Ayache SS Baeken 664
C Benninger DH Cantello RM Cincotta M de Carvalho 665
M De Ridder D Devanne H Di Lazzaro V Filipovic SR 666
Hummel FC Jaaskelainen SK Kimiskidis VK Koch G 667
Langguth B Nyffeler T Oliviero A Padberg F Poulet E 668
Rossi S Rossini PM Rothwell JC Schonfeldt-Lecuona 669
C Siebner HR Slotema CW Stagg CJ Valls-Sole J 670
Ziemann U Paulus W amp Garcia-Larrea L (2014) Evidence- 671
based guidelines on the therapeutic use of repetitive transcranial 672
magnetic stimulation (rTMS) Clin Neurophysiol 125(11) 673
2150-2206 674
Lefaucheur JP Brusa A Creange A Drouot X amp Jarry G 675
(2002) Clinical application of laser evoked potentials using 676
the NdYAG laser Neurophysiol Clin 32(2) 91-98 677
Lefaucheur JP Debray S amp Jarry G (2001) Laser evoked poten- 678
tials using the NdYAG laser Muscle Nerve 24(4) 496-501 679
Lefaucheur JP Drouot X Menard-Lefaucheur I Keravel Y 680
amp Nguyen JP (2006) Motor cortex rTMS restores defective 681
intracortical inhibition in chronic neuropathic pain Neurology 682
67(9) 1568-1574 683
Longo MR Iannetti GD Mancini F Driver J amp Haggard P 684
(2012) Linking pain and the body Neural correlates of visually 685
induced analgesia J Neurosci 32(8) 2601-2607 686
Lorenz J amp Garcia-Larrea L (2003) Contribution of attentional 687
and cognitive factors to laser evoked brain potentials Neuro- 688
physiol Clin 33(6) 293-301 689
Lorenz J Minoshima S amp Casey KL (2003) Keeping pain out 690
of mind The role of the dorsolateral prefrontal cortex in pain 691
modulation Brain 126(Pt 5) 1079-1091 692
Luedtke K May A amp Jurgens TP (2012) No effect of a single 693
session of transcranial direct current stimulation on experimen- 694
tally induced pain in patients with chronic low back painndashan 695
exploratory study PLoS One 7(11) e48857 696
Unc
orre
cted
Aut
hor P
roof
12 T Bocci et al Cerebellum and pain A tcDCS study
Mendonca ME Santana MB Baptista AF Datta A Bikson697
M Fregni F amp Araujo CP (2011) Transcranial DC stimu-698
lation in fibromyalgia Optimized cortical target supported by699
high-resolution computational models J Pain 12(5) 610-617700
Moriguchi Y Decety J Ohnishi T Maeda M Mori T Nemoto701
K Matsuda H amp Komaki G (2007) Empathy and judg-702
ing otherrsquos pain An fMRI study of alexithymia Cereb Cortex703
17(9) 2223-2234704
Moulton EA Elman I Pendse G Schmahmann J Becerra L705
amp Borsook D (2011) Aversion-related circuitry in the cere-706
bellum Responses to noxious heat and unpleasant images J707
Neurosci 31(10) 3795-3804708
Moulton EA Schmahmann JD Becerra L amp Borsook D709
(2010) The cerebellum and pain Passive integrator or active710
participator Brain Res Rev 65(1) 14-27711
Mouraux A amp Plaghki L (2006) Are the processes reflected by712
late and ultra-late laser evoked potentials specific of nocicep-713
tion Suppl Clin Neurophysiol 59 197-204714
Mylius V Jung M Menzler K Haag A Khader PH Oer-715
tel WH Rosenow F amp Lefaucheur JP (2012) Effects of716
transcranial direct current stimulation on pain perception and717
working memory Eur J Pain 16(7) 974-982718
Ni Z Gunraj C amp Chen R (2007) Short interval intracortical719
inhibition and facilitation during the silent period in human J720
Physiol 583(Pt 3) 971-982721
Nitsche MA Liebetanz D Lang N Antal A Tergau F amp722
Paulus W (2003) Safety criteria for transcranial direct cur-723
rent stimulation (tDCS) in humans Clin Neurophysiol 114(11)724
2220-2222 author reply 2222-2223725
Opsommer E Guerit JM amp Plaghki L (2003) Exogenous726
and endogenous components of ultralate (C-fibre) evoked727
potentials following CO2 laser stimuli to tiny skin sur-728
face areas in healthy subjects Neurophysiol Clin 33(2)729
78-85730
Parazzini M Fiocchi S Liorni I Priori A amp Ravazzani P731
(2014) Computational modeling of transcranial direct current732
stimulation in the child brain Implications for the treatment of733
refractory childhood focal epilepsy Int J Neural Syst 24(2)734
1430006735
Parazzini M Rossi E Ferrucci R Fiocchi S Liorni I Priori736
A amp Ravazzani P (2013) Computational model of cerebellar737
transcranial direct current stimulation Conf Proc IEEE Eng738
Med Biol Soc 2013 237-240739
Parazzini M Rossi E Ferrucci R Liorni I Priori A amp Ravaz-740
zani P (2014) Modelling the electric field and the current741
density generated by cerebellar transcranial DC stimulation in742
humans Clin Neurophysiol 125(3) 577-584743
Perini I Bergstrand S amp Morrison I (2013) Where pain meets744
action in the human brain J Neurosci 33(40) 15930-15939745
Ploghaus A Tracey I Gati JS Clare S Menon RS Matthews746
PM amp Rawlins JN (1999) Dissociating pain from its antic-747
ipation in the human brain Science 284(5422) 1979-1981748
Priori A Ciocca M Parazzini M Vergari M amp Ferrucci R749
(2014) Transcranial cerebellar direct current stimulation and750
transcutaneous spinal cord direct current stimulation as innova-751
tive tools for neuroscientists J Physiol 592(Pt 16) 3345-3369752
Purpura DP amp McMurtry JC (1965) Intracellular activities and 753
evoked potential changes during polarization of motor cortex 754
J Neurophysiol 28 166-185 755
Qiu Y Inui K Wang X Tran TD amp Kakigi R (2002) Effects 756
of attention distraction and sleep on CO(2) laser evoked poten- 757
tials related to C-fibers in humans Clin Neurophysiol 113(10) 758
1579-1585 759
Reidler JS Mendonca ME Santana MB Wang X Lenkinski 760
R Motta AF Marchand S Latif L amp Fregni F (2012) 761
Effects of motor cortex modulation and descending inhibitory 762
systems on pain thresholds in healthy subjects J Pain 13(5) 763
450-458 764
Riedl V Valet M Woller A Sorg C Vogel D Sprenger 765
T Boecker H Wohlschlager AM amp Tolle TR (2011) 766
Repeated pain induces adaptations of intrinsic brain activity 767
to reflect past and predict future pain Neuroimage 57(1) 206- 768
213 769
Rocca MA Valsasina P Absinta M Colombo B Barcella V 770
Falini A Comi G amp Filippi M (2010) Central nervous 771
system dysregulation extends beyond the pain-matrix network 772
in cluster headache Cephalalgia 30(11) 1383-1391 773
Rossi S Hallett M Rossini PM amp Pascual-Leone A (2009) 774
Safety ethical considerations and application guidelines for 775
the use of transcranial magnetic stimulation in clinical practice 776
and research Clin Neurophysiol 120(12) 2008-2039 777
Rossini PM amp Rossi S (2007) Transcranial magnetic stimulation 778
Diagnostic therapeutic and research potential Neurology 779
68(7) 484-488 780
Ruscheweyh R Kuhnel M Filippopulos F Blum B Eggert 781
T amp Straube A (2014) Altered experimental pain perception 782
after cerebellar infarction Pain 155(7) 1303-1312 783
Schmahmann JD (1991) An emerging concept The cerebellar 784
contribution to higher function Arch Neurol 48(11) 1178- 785
1187 786
Singer T Seymour B OrsquoDoherty J Kaube H Dolan RJ amp 787
Frith CD (2004) Empathy for pain involves the affective but 788
not sensory components of pain Science 303(5661) 1157- 789
1162 790
Smith MJ Keel JC Greenberg BD Adams LF Schmidt PJ 791
Rubinow DA amp Wassermann EM (1999) Menstrual cycle 792
effects on cortical excitability Neurology 53(9) 2069-2072 793
Stoodley CJ amp Schmahmann JD (2009) Functional topography 794
in the human cerebellum A meta-analysis of neuroimaging 795
studies Neuroimage 44(2) 489-501 796
Strick PL Dum RP amp Fiez JA (2009) Cerebellum and non- 797
motor function Annu Rev Neurosci 32 413-434 798
Strigo IA Duncan GH Boivin M amp Bushnell MC (2003) 799
Differentiation of visceral and cutaneous pain in the human 800
brain J Neurophysiol 89(6) 3294-3303 801
Truini A Galeotti F Romaniello A Virtuoso M Iannetti GD 802
amp Cruccu G (2005) Laser-evoked potentials Normative val- 803
ues Clin Neurophysiol 116(4) 821-826 804
Truini A Panuccio G Galeotti F Maluccio MR Sartucci F 805
Avoli M amp Cruccu G (2010) Laser-evoked potentials as a 806
tool for assessing the efficacy of antinociceptive drugs Eur J 807
Pain 14(2) 222-225 808
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 13
Truini A Vergari M Biasiotta A La Cesa S Gabriele M Di809
Stefano G Cambieri C Cruccu G Inghilleri M amp Pri-810
ori A (2011) Transcutaneous spinal direct current stimulation811
inhibits nociceptive spinal pathway conduction and increases812
pain tolerance in humans Eur J Pain 15(10) 1023-1027813
Valeriani M Le Pera D Restuccia D de Armas L Mili-814
ucci R Betti V Vigevano F amp Tonali P (2007) Parallel815
spinal pathways generate the middle-latency N1 and the late P2816
components of the laser evoked potentials Clin Neurophysiol817
118(5) 1097-1104818
Valeriani M Pazzaglia C Cruccu G amp Truini A (2012) Clinical819
usefulness of laser evoked potentials Neurophysiol Clin 42(5)820
345-353821
Volz MS Mendonca M Pinheiro FS Cui H Santana M amp822
Fregni F (2012) Dissociation of motor task-induced cortical823
excitability and pain perception changes in healthy volunteers824
PLoS One 7(3) e34273825
Wager TD Rilling JK Smith EE Sokolik A Casey KL 826
Davidson RJ Kosslyn SM Rose RM amp Cohen JD 827
(2004) Placebo-induced changes in FMRI in the anticipation 828
and experience of pain Science 303(5661) 1162-1167 829
Zaghi S Thiele B Pimentel D Pimentel T amp Fregni 830
F (2011) Assessment and treatment of pain with non- 831
invasive cortical stimulation Restor Neurol Neurosci 29(6) 832
439-451 833
Zubieta JK Bueller JA Jackson LR Scott DJ Xu Y 834
Koeppe RA Nichols TE amp Stohler CS (2005) Placebo 835
effects mediated by endogenous opioid activity on mu-opioid 836
receptors J Neurosci 25(34) 7754-7762 837
Zunhammer M Busch V Griesbach F Landgrebe M Hajak G 838
amp Langguth B (2011) rTMS over the cerebellum modulates 839
temperature detection and pain thresholds through peripheral 840
mechanisms Brain Stimul 4(4) 210-7 e1 841

Unc
orre
cted
Aut
hor P
roof
4 T Bocci et al Cerebellum and pain A tcDCS study
Fig 2 ndash Experimental protocol Psychophysical and electrophysiological variables evaluated at baseline (T0) and at two different time points(T1 T2) following anodal cathodal and sham tcDCS
of peripheral nociceptors (Truini et al 2005) The179
inter-stimulus interval was varied randomly (10ndash15 s)180
Participants were reclined on a couch and wore protec-181
tive goggles They were instructed to keep their eyes182
open and gaze slightly downwards since the N2P2183
amplitude is enhanced by attention (Lorenz amp Garcia-184
Larrea 2003 Truini et al 2005) they were requested185
to mentally count the number of stimuli The main186
Aδ-LEP vertex complex N2ndashP2 and the lateralised187
N1 component were recorded through standard disc188
non-polarizable AgAgCl surface electrodes (diameter189
10 mm Biomedreg Florence Italy) N2 and P2 compo-190
nents were recorded from the vertex (Cz) referenced191
to the earlobes the N1 component was recorded from192
the temporal leads (T4) referenced to Fz (Cruccu et al193
2008) Blinks and saccades were recorded with an EOG194
electrode placed on the supero-lateral right canthus195
connected to the system reference Ground was placed196
on the mid-forehead Skin impedance was kept below197
5 k198
232 Cerebellar transcutaneous direct current199
stimulation (tcDCS)200
tDCS was applied using a battery-driven constant201
current stimulator (HDCStim Newronika Italy) and202
a pair of electrodes in two saline-soaked synthetic203
sponges with a surface area of 25 cm2 For cathodal204
stimulation the cathode was centered on the median205
line 2 cm below the inion with its lateral borders about206
1 cm medially to the mastoid apophysis and the anode 207
over the right shoulder (Ferrucci et al 2008 2012 208
2013) For anodal stimulation the current flow was 209
reversed In the real tcDCS conditions direct current 210
was transcranially applied for 20 minutes with an inten- 211
sity of 20 mA and constant current flow was measured 212
by an ampere meter (current density asymp 008 mAcm2) 213
These values are similar to those previously reported 214
for cerebellar stimulation (Ferrucci et al 2008 2013) 215
are considered to be safe (Iyer et al 2005) and are 216
far below the threshold for tissue damage (Nitsche 217
et al 2003) Apart from occasional and short-lasting 218
tingling and burning sensations below the electrodes 219
direct current stimulation strength remained below the 220
sensory threshold throughout the experimental session 221
At the offset of tDCS the current was decreased in a 222
ramp-like manner a method shown to achieve a good 223
level of blinding among sessions (Gandiga et al 2006 224
Galea et al 2009) For a sham tDCS the current was 225
turned on only for 5 seconds at the beginning of the 226
sham session and then it was turned off in a ramp- 227
shaped fashion which induces initial skin sensations 228
indistinguishable from real tDCS 229
For all the electrophysiological recordings we chose 230
the left side to avoid interference from the return 231
electrode placed over the contralateral shoulder At 232
experimental debriefing subjects were not able to dis- 233
criminate between the applied anodal cathodal and 234
sham tDCS 235
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 5
Table 1
Row data (expressed as mean value plusmn 1 standard deviation a = anodal stimulation c = cathodal stimulation sh = sham condition)Both psychophysical and electrophysiological data for each subject are fully available as supplementary electronic material at
httpwwwenricasantarcangelocompublications
aT0 aT1 aT2 cT0 cT1 cT2 shT0 shT1 shT2
PT mean 462 607 609 485 376 368 472 466 489SD 080 095 092 086 062 067 098 062 081
VAS I1 mean 389 255 265 367 493 467 387 393 387SD 084 057 062 082 096 082 074 070 092
VAS I2 mean 540 402 403 524 673 665 533 549 530SD 063 082 071 048 047 049 078 069 064
N1 amplitude (V) mean 1292 848 801 1104 1496 1494 1101 1111 1121SD 318 298 258 265 258 333 250 267 283
N1 latency (ms) mean 12419 16146 15710 12704 10715 10405 12817 12867 13066SD 1090 1338 1368 1075 675 912 1320 1271 1209
N2P2 amplitude(V) mean 1114 738 757 1052 1453 1375 1114 1125 1147SD 262 237 233 265 296 329 272 269 216
N2P2 latency (ms) mean 15157 18932 18726 14878 12673 13230 15390 15108 15551SD 1312 1749 2139 2201 1849 1870 1433 1507 1675
233 Transcranial magnetic stimulation (TMS)236
Changes in Resting Motor Threshold (RMT) were237
evaluated at different intervals before and after the238
completion of tcDCS A Magstim Super Rapid Tran-239
scranial Magnetic Stimulator (Magstim Company240
Dyfed UK 22 T maximum field output) connected241
to a standard eight-shaped focal coil with wing diame-242
ters of 70 mm was used The handle of the eight-shaped243
focal coil was pointed backwards and rotated about 45244
deg to the mid-sagittal line to induce a tissue current245
perpendicular to the motor strip in the precentral sul-246
cus (Rossi et al 2009 Groppa et al 2012) RMT247
was defined as the minimum stimulator output that248
induces motor evoked potentials (MEPs) of more than249
50 V in at least five out of 10 trials when first dig-250
ital interosseus (FDI) muscle was completely relaxed251
(Ni et al 2007) The motor ldquohot spotrdquo for the targeted252
muscle was identified by single pulses of TMS deliv-253
ered at a slightly suprathreshold stimulus intensity and254
the magnetic stimuli induced monophasic pulses The255
coil was placed over the right motor cortex (centered on256
C4 according with the 10ndash20 EEG International Sys-257
tem) and electromyographic recordings were made by258
two standard non-polarizable AgAgCl surface elec-259
trodes (diameter 10 mm Biomedreg Florence Italy)260
one placed over the belly of the contralateral FDI261
muscle and the other on the skin overlying the first262
metacarpophalangeal joint of the first finger of the left263
hand RMT was evaluated to exclude possible cere-264
bellar stimulation spread out inducing motor cortex265
activation266
24 Variables and statistical analysis 267
We studied the subjective experience - percep- 268
tive threshold (PT) and pain intensity perceived 269
after laser I1 and I2 (VAS1 VAS2) - and elec- 270
trophysiological variables that is the peak-to-peak 271
amplitude of the N1 wave and N2P2 complex 272
the peak latency of N1 and N2 as reported in 273
previous papers using NdYAG laser (Lefaucheur 274
et al 2001 2002)275
Analyses were performed through SPSS15 sta- 276
tistical Package Psychophysical (PT VAS1 VAS2) 277
and electrophysiological variables (mean values of 278
ten traces N1amplitude and latency N2P2 ampli- 279
tude and latency) as well as Resting Motor Thresholds 280
(RMT) were analysed following a 3 Stimulation con- 281
ditions (anodal cathodal sham) times 3 Times (T0 T1 282
T2) design The Greenhouse-Geisser correction for 283
non sphericity was applied when necessary Con- 284
trast analysis between Times (F values) and paired 285
t tests between stimulations were alternatively used 286
for post-hoc comparisons when appropriate After 287
Bonferroni correction significance level was set at 288
p lt 0007289
The changes of all variable in T1 and T2 290
were expressed as ratio between post and pre 291
stimulation values (T1T0 T2T0) and compared 292
between each other according to a 2 Stimula- 293
tion (anodal cathodal) times 2 Times (T1To T2T0) 294
design 295
Unc
orre
cted
Aut
hor P
roof
6 T Bocci et al Cerebellum and pain A tcDCS study
Table 2
Contrast analyses all comparisons were highly significant(p lt 00001)
anodal cathodal shamPT time df F = 4430 F = 1867 ns
T0 vs Tl 228 F = 77669) F = 27523T0 vs T2 114 F = 78745 F = 27827Tl vs T2 114 ns ns
anodal vs sham cathodal vs shamT0 114 ns nsTl 114 t = 5069 t = 6991T2 114 t = 3709 t = 5849
VAS anodal cathodal sham
time 228 F = 41954 F = 31448 nsT0 vs Tl 114 F = 56968 F = 48596T0 vs T2 114 F = 52289 F = 525Tl vs T2 ns ns
anodal vs sham cathodal vs shamT0 114 ns nsTl 114 t = 644 t = 5916T2 114 t = 5294 t = 582
3 Results296
Row data (mean SD) are shown in Table 1 Base-297
line values were in line with those reported by earlier298
studies performed by using NdYAG laser (Lefaucheur299
et al 2001) Indeed only one study described a longer300
latency of the N2 wave (Cruccu et al 2008) The301
sham stimulation did not modulate any psychophys-302
ical and electrophysiological variable (Table 2) Since303
no pre-post difference was found for sham polarity this304
condition was not included in the comparison between305
Stimulations and Times306
31 Psychophysics307
PT exhibited a significant Stimulation effect308
(F(228) = 35055 p lt 00001 η2 = 0715) and a signifi-309
cant Stimulation times Time interaction (F(456) = 39464310
p lt 00001 η2 = 0738) Decomposition of the latter311
(Table 2) revealed that a) PT was higher for the anodal312
and lower for the cathodal stimulation conditions com-313
pared with the sham stimulation for both T1 and T2314
b) with respect to T0 PT increased in T1 and T2 in the315
anodal condition and decreased in the cathodal con-316
dition while no significant difference was observed317
between T1 and T2 (Fig 3A)318
Significantly different VAS1 and VAS2 were319
observed for the two stimulation intensities (F(114)320
= 54262 p lt 00001) and the three Stimulation con-321
ditions (F(218) = 88882 p lt 00001) Decomposition322
of the significant Stimulation times Time interaction323
(F(456) = 11596 p lt 00001) revealed that the reported 324
pain intensity for both stimulation intensities (VAS1 325
and VAS2) was higher for the cathodal and lower for 326
the anodal stimulation compared to the sham stimula- 327
tion (Table 2) It increased in T1 and decreased in T2 328
with respect to T0 whereas no significant difference 329
was found between T1 and T2 (Fig 3-B) 330
32 Laser evoked potentials 331
Figure 4-A shows the LEPs recorded in all experi- 332
mental conditions in a representative subject Both N1 333
and N2P2 amplitude (N1 F(456) = 10695 p lt 00001 334
η2 = 0884 N2P2 F(456) = 86864 p lt 00001 η2 = 335
0861) and latency (N1 F(456) = 110869 p lt 00001 336
η2 = 0888 N2P2 F(456) = 3660 p lt 00001 337
η2 = 0723) exhibited a significant Time times Stimulation 338
interaction Its decomposition (Table 3) showed 339
that both amplitudes increased and both latencies 340
decreased for cathodal stimulation in T1 and T2 with 341
respect to T0 the opposite occurred for the anodal 342
stimulation Both stimulations induced responses 343
significantly different from the sham condition 344
(Fig 4-B) The responses obtained after cathodal 345
stimulation were significantly improved (higher 346
amplitudes lower latencies) than those produced 347
by the anodal one (N1 amplitude F(114) = 41345 348
p lt 00001 N1 latency F(114) = 496228 p lt 00001 349
N2P2 amplitude (F(114) = 44537 p lt 00001 N2P2 350
latency F(114) = 119056 p lt 00001) 351
33 Resting motor thresholds 352
RMT values at baseline did not differ among exper- 353
imental conditions (mean plusmn SD sham 508 plusmn 83 354
anodal 491 plusmn 62 cathodal 503 plusmn 63) ANOVA 355
did not reveal any significant Stimulation (F(228) = 356
0882 p = 0425 η2 = 0059) Time (F(228) = 0212 357
p = 0810 η2 = 0015) and Stimulation times Time 358
effect (F(456) = 0339 p = 0851 η2 = 0024) for RMT 359
(Fig 5) 360
4 Discussion 361
Our study shows that cerebellar direct current 362
polarization modulates nociceptive perception and its 363
cortical correlates in healthy humans Specifically 364
cathodal tcDCS increases pain perception increases 365
amplitudes and decreases LEPs latencies likely though 366
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 7
A
B
Fig 3 - A Perceptive Threshold Changes (mean plusmn SD) at T1 and T2 with respect to baseline values (T1T0 T2T0) following sham (black)anodal (white) and cathodal (grey) tcDCS (lowastlowastp lt 0001 lowastlowastlowastp lt 00001) B Changes in visual analogue scale (VAS) scores over time VAS scoresat two different stimulus intensity respectively two (A left) and three (B right) times higher than the PT (lowastlowastp lt 0001 lowastlowastlowastp lt 00001)
A B
Fig 4 ndash A LEPs grand averaging traces were recorded at baseline (T0 black) and immediately after cerebellar polarization (T1 red) due tosham (left column) anodal (middle) and cathodal (right) tcDCS B Histograms showing LEPs variables and VAS scores changes (mean plusmn SD)after sham (black) anodal (white) or cathodal (grey) tcDCS with respect to baseline Top panels changes in N1 variables (amplitude and latency)over time bottom panels changes in N2P2 complex (lowastlowastp lt 0001 lowastlowastlowastp lt 00001)
Unc
orre
cted
Aut
hor P
roof
8 T Bocci et al Cerebellum and pain A tcDCS study
Table 3
LEPs post-hoc analyses p lt 00001 for all comparison except when explicitly indicated lowastlowastp lt 0002 lowast p lt 0005
N1 amplitude latency N2P2 amplitude latency
anodal cathodal shamdf
time 228 F = 67152 F = 96489 F = 134912 F = 34946 nsT0 vs T1 114 F = 109178 F = 18815 F = 165953 F = 64281T0 vs T2 114 F = 75143 F = 167697 F = 145125 F = 37818T1 vs T2 114 ns ns ns ns
anodal cathodaltime 228 F = 102281 F = 98717 F = 6577 F = 20918 nsT0 vs T1 114 F = 511186 F = 104027 F = 144112 F = 103864T0 vs T2 114 F = 96329 F = 116841 F = 105183 F = 14012lowastlowastT1 vs T2 114 ns ns ns ns ns
anodal vs sham anodal vs shamT0 114 ns ns ns nsT1 114 ns t = 925 t = 601 6262T2 114 ns t = 8128 t = 6731 5236
cathodal cathodal vs shamT0 114 ns ns t = 3281lowast nsT1 114 t = 16594 t = 8029 t = 8262 t = 520T2 114 t = 7309 t = 12669 t = 5048 t = 5301
Fig 5 - Resting Motor Thresholds Changes (mean plusmn SD) in Resting Motor threshold (RMT) expressed as percentage of the maximumstimulator output after sham (black) anodal (white) and cathodal (grey) tcDCS with respect to baseline marked as dotted line (lowastlowastp lt 0001lowastlowastlowastp lt 00001)
reduction of the inhibitory tone exerted by the cere-367
bellum on brain targets Anodal polarization elicits368
opposite effects producing analgesia Both findings369
support the role of the cerebellum in pain control370
it is noticeable that cathodal cerebellar stimulation371
induces hyperalgesia as occurs in patients with cere-372
bellar infarction (Ruscheweyh et al 2014)373
We would like to underline that in the present study374
LEPs were obtained at laser intensities depending on375
the perceptive threshold which varied as a function376
of anodal and cathodal stimulation This means that 377
the cerebellar stimulation has not a selective analgesic 378
effect as it influences both non nociceptive and noci- 379
ceptive perception A pre-eminent analgesic cannot be 380
assessed because the nociceptive threshold was not 381
evaluated 382
As tcDCS was effective on the modulation of 383
both N1 and N2P2 components and these responses 384
are generated by parallel and partially segregated 385
spinal pathways reaching different cortical targets 386
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 9
(Valeriani et al 2007) we may suggest that the cere-387
bellum is engaged in pain processing by modulating388
the activity of both somatosensory and cingulate cor-389
tices Indeed the cerebellum is involved in both the390
sensory-discriminative and emotional dimension of391
pain (Singer et al 2004 Moriguchi et al 2007) and392
non-invasive cerebellar current stimulation may modu-393
late pain experience and the associated cortical activity394
through many not alternative mechanisms In partic-395
ular changes in N1 reflects the modulation of the396
sensory component of pain while the vertex N2P2397
represents the neural correlate of affective aspects of398
pain experience (Garcia-Larrea et al 1997 Valeriani399
et al 2007) Notably tcDCS may act not only on spinal400
nociceptive neurons but also on wide-range cortical401
networks of the pain matrix (Singer et al 2004) thus402
influencing LEPs and pain experience through both403
top-down and bottom-up mechanisms404
The present study does not allow to hypothesize405
how and where tcDCS influences the cerebellar activ-406
ity A main role of Purkinje cells has been suggested407
as their activity modulation may affect the cerebellar408
inhibitory control of the cerebral cortex (Galea et al409
2009) This would be in line with the effects elicited410
by tDCS in the cerebral cortex which are observ-411
able after both short and long term delay likely also412
interfering with long-term potentiation (LTP)-like phe-413
nomena (Hamada et al 2012 Priori et al 2014)414
Moreover prolonged spiking activity in the cerebellar415
Golgi inhibitory neurons modulates the activity of the416
Purkinje cells and could partly account for the tcDCS417
after-effects (Hull et al 2013)418
The lack of changes in RMT indicates that the anal-419
gesic effects of anodal tcDCS are due to a specific420
modulation of the cerebellar activity and not to motor421
activation On the other hand tcDCS-induced cerebel-422
lar modulation (Purpura amp McMurtry 1965) could be423
not sufficient per se to activate the cerebello ndash thalamo424
- cortical motor pathway (Galea et al 2009) thus425
the reported analgesia and its cortical correlates can-426
not be sustained by the motor cortex activation This427
view is supported by the absence of any association428
between motor symptoms and pain perception in cere-429
bellar patients (Ruscheweyh et al 2014) In the same430
line in healthy subjects it has been recently shown431
that motor task-induced increased cortical excitability432
and analgesia are not associated (Volz et al 2012)433
Indeed RMT is a highly sensitive marker of motor434
tract excitability as it reflects activation of a small435
low-threshold and slow-conducting core of pyramidal436
neurons (Hess et al 1987 Rossini amp Rossi 2007) 437
although RMT may reflect changes in the activity of 438
different central nervous system structures it has been 439
satisfactorily used to assess motor cortex excitability 440
also in cerebellar patients (Battaglia et al 2006) 441
Another critical point is the possibility to modulate 442
with tcDCS both neural correlates underlying nocicep- 443
tive processing and pain perception Previous studies 444
using tDCS over motor cortex were inconsistent among 445
each other some works suggested that tDCS is able to 446
modify pain perception (Boggio et al 2008) while 447
others showed divergent effects on psychophysical 448
and neurophysiological outcome parameters (Luedtke 449
et al 2012 Ihle et al 2014) likely due to a possi- 450
ble overestimation of the role of motor areas on pain 451
processing (Antal et al 2008) 452
Our findings cannot be compared to the results 453
obtained by other Authors In fact the unique study 454
focused on the analgesic effects of non-invasive cere- 455
bellar stimulation reported till now (Zunhammer et al 456
2011) considered only subjective pain thresholds 457
In addition it described similar analgesic effects of 458
cerebellar and neck structures repetitive transcranial 459
magnetic stimulation (rTMS) thus denying any cere- 460
bellar specificity in the observed effects and suggesting 461
that the peripheral information passing through the 462
cerebellum may be responsible for analgesia The 463
main difference between the two studies possibly 464
accounting for different results consists of the neu- 465
romodulation techniques used 466
41 Limitations of the study 467
The present study has a few limitations First our 468
findings do not allow any hypothesis on the role of 469
the cerebellum in chronic pain The observations on 470
patients with cerebellar damage (Ruscheweyh et al 471
2014) suggest that their impaired inhibitory control 472
mechanisms may be not associated with the devel- 473
opment of chronic pain Second we cannot exclude 474
the possibility that tcDCS could modulate not only the 475
cerebellum but also surrounding areas such as the peri- 476
aqueductal gray However recent modelling researches 477
have revealed that during tcDCS the current spread 478
to other structures outside the cerebellum is negligi- 479
ble (Parazzini et al 2013 2014) Moreover several 480
studies have proved that in humans pain processing 481
is encoded within posterior areas of each cerebellar 482
hemisphere specifically in the hemispheric lobule VI 483
Crus I and VIIb (Moulton et al 2011) where the 484
Unc
orre
cted
Aut
hor P
roof
10 T Bocci et al Cerebellum and pain A tcDCS study
tcDCS-induced electrical field is strongly concentrated485
(Parazzini et al 2013) A further limitation is that we486
did not study the contribution of C-fibers the main487
component of spino-ponto-cerebellar and spino-olivo-488
cerebellar pathways In fact ultra-late LEPs related489
to C-fibers activations have not yet been standard-490
ized for clinical application and their occurrence could491
be markedly influenced by high order cognitive pro-492
cesses as they seem to be more affected by the level493
of consciousness and attention than A-delta responses494
(Qiu et al 2002 Opsommer et al 2003 Mouraux amp495
Plaghki 2006) Finally we wish to emphasize that in496
neuropathic patients the effects of the cerebellar stim-497
ulation could be quite different from those described498
here as both anatomical and functional connectivity499
are different from those observed in healthy partici-500
pants (Rocca et al 2010 Riedl et al 2011 Absinta et501
al 2012 Longo et al 2012 Ceko et al 2013)502
5 Conclusions503
Our findings indicate a cerebellar effect on pain504
experience and on its cortical correlates and prompt505
further investigation aimed at assessing whether the506
cerebellar direct current polarization could be used as a507
novel and safe therapeutic tool in chronic pain patients508
Acknowledgments509
We gratefully acknowledge the participation of all510
subjects as well as Mr D Barloscio and Mr C511
Orsini for their excellent technical assistance The512
paper was supported in part by the Italian operating513
and development MIUR PRIN grant year 2006 n514
2006062332 002515
RF and AP are founders and shareholders of516
Newronika srl Milan Italy517
References518
Absinta M Rocca MA Colombo B Falini A Comi G amp Fil-519
ippi M (2012) Selective decreased grey matter volume of the520
pain-matrix network in cluster headache Cephalalgia 32(2)521
109-115522
Agostino R Cruccu G Iannetti G Romaniello A Truini A523
amp Manfredi M (2000) Topographical distribution of pinprick524
and warmth thresholds to CO2 laser stimulation on the human525
skin Neurosci Lett 285(2) 115-118526
Antal A Brepohl N Poreisz C Boros K Csifcsak G amp 527
Paulus W (2008) Transcranial direct current stimulation over 528
somatosensory cortex decreases experimentally induced acute 529
pain perception Clin J Pain 24(1) 56-63 530
Antal A amp Paulus W (2010) Effects of transcranial theta-burst 531
stimulation on acute pain perception Restor Neurol Neurosci 532
28(4) 477-484 533
Apkarian AV Bushnell MC Treede RD amp Zubieta JK 534
(2005) Human brain mechanisms of pain perception and reg- 535
ulation in health and disease Eur J Pain 9(4) 463-484 536
Balsters JH Whelan CD Robertson IH amp Ramnani N (2013) 537
Cerebellum and cognition Evidence for the encoding of higher 538
order rules Cereb Cortex 23(6) 1433-1443 539
Battaglia F Quartarone A Ghilardi MF Dattola R Bagnato 540
S Rizzo V Morgante L amp Girlanda P (2006) Unilat- 541
eral cerebellar stroke disrupts movement preparation and motor 542
imagery Clin Neurophysiol 117(5) 1009-1016 543
Bingel U Quante M Knab R Bromm B Weiller C amp Buchel 544
C (2002) Subcortical structures involved in pain processing 545
Evidence from single-trial fMRI Pain 99(1-2) 313-321 546
Boggio PS Zaghi S amp Fregni F (2009) Modulation of emotions 547
associated with images of human pain using anodal transcranial 548
direct current stimulation (tDCS) Neuropsychologia 47(1) 549
212-217 550
Boggio PS Zaghi S Lopes M amp Fregni F (2008) Modulatory 551
effects of anodal transcranial direct current stimulation on per- 552
ception and pain thresholds in healthy volunteers Eur J Neurol 553
15(10) 1124-1130 554
Borsook D Moulton EA Tully S Schmahmann JD amp 555
Becerra L (2008) Human cerebellar responses to brush and 556
heat stimuli in healthy and neuropathic pain subjects Cerebel- 557
lum 7(3) 252-272 558
Ceko M Seminowicz DA Bushnell MC amp Olausson HW 559
(2013) Anatomical and functional enhancements of the insula 560
after loss of large primary somatosensory fibers Cereb Cortex 561
23(9) 2017-2024 562
Cruccu G Aminoff MJ Curio G Guerit JM Kakigi 563
R Mauguiere F Rossini PM Treede RD amp Garcia- 564
Larrea L (2008) Recommendations for the clinical use of 565
somatosensory-evoked potentials Clin Neurophysiol 119(8) 566
1705-1719 567
Cruccu G Romaniello A Amantini A Lombardi M Innocenti 568
P amp Manfredi M (1999) Assessment of trigeminal small- 569
fiber function Brain and reflex responses evoked by CO2-laser 570
stimulation Muscle Nerve 22(4) 508-516 571
Dasilva AF Mendonca ME Zaghi S Lopes M Dossantos 572
MF Spierings EL Bajwa Z Datta A Bikson M amp 573
Fregni F (2012) tDCS-induced analgesia and electrical fields 574
in pain-related neural networks in chronic migraine Headache 575
52(8) 1283-1295 576
Duerden EG amp Albanese MC (2013) Localization of pain- 577
related brain activation A meta-analysis of neuroimaging data 578
Hum Brain Mapp 34(1) 109-149 579
Ekerot CF Garwicz M amp Schouenborg J (1991a) The postsy- 580
naptic dorsal column pathway mediates cutaneous nociceptive 581
information to cerebellar climbing fibres in the cat J Physiol 582
441 275-284 583
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 11
Ekerot CF Garwicz M amp Schouenborg J (1991b) Topography584
and nociceptive receptive fields of climbing fibres projecting to585
the cerebellar anterior lobe in the cat J Physiol 441 257-274586
Ekerot CF Gustavsson P Oscarsson O amp Schouenborg J587
(1987a) Climbing fibres projecting to cat cerebellar anterior588
lobe activated by cutaneous A and C fibres J Physiol 386589
529-538590
Ekerot CF Oscarsson O amp Schouenborg J (1987b) Stimula-591
tion of cat cutaneous nociceptive C fibres causing tonic and592
synchronous activity in climbing fibres J Physiol 386 539-593
546594
Ferrucci R Brunoni AR Parazzini M Vergari M Rossi E595
Fumagalli M Mameli F Rosa M Giannicola G Zago S596
amp Priori A (2013) Modulating human procedural learning by597
cerebellar transcranial direct current stimulation Cerebellum598
12(4) 485-492599
Ferrucci R Giannicola G Rosa M Fumagalli M Boggio600
PS Hallett M Zago S amp Priori A (2012) Cerebellum601
and processing of negative facial emotions Cerebellar tran-602
scranial DC stimulation specifically enhances the emotional603
recognition of facial anger and sadness Cogn Emot 26(5)604
786-799605
Ferrucci R Marceglia S Vergari M Cogiamanian F Mrakic-606
Sposta S Mameli F Zago S Barbieri S amp Priori607
A (2008) Cerebellar transcranial direct current stimulation608
impairs the practice-dependent proficiency increase in working609
memory J Cogn Neurosci 20(9) 1687-1697610
Fregni F Freedman S amp Pascual-Leone A (2007) Recent611
advances in the treatment of chronic pain with non-invasive612
brain stimulation techniques Lancet Neurol 6(2) 188-191613
Galea JM Jayaram G Ajagbe L amp Celnik P (2009)614
Modulation of cerebellar excitability by polarity-specific615
noninvasive direct current stimulation J Neurosci 29(28)616
9115-9122617
Galea JM Vazquez A Pasricha N de Xivry JJ amp Celnik618
P (2011) Dissociating the roles of the cerebellum and motor619
cortex during adaptive learning The motor cortex retains what620
the cerebellum learns Cereb Cortex 21(8) 1761-1770621
Gandiga PC Hummel FC amp Cohen LG (2006) Tran-622
scranial DC stimulation (tDCS) A tool for double-blind623
sham-controlled clinical studies in brain stimulation Clin Neu-624
rophysiol 117(4) 845-850625
Garcia-Larrea L Peyron R Laurent B amp Mauguiere F (1997)626
Association and dissociation between laser-evoked potentials627
and pain perception Neuroreport 8(17) 3785-3789628
Grimaldi G Argyropoulos GP Boehringer A Celnik P629
Edwards MJ Ferrucci R Galea JM Groiss SJ Hiraoka630
K Kassavetis P Lesage E Manto M Miall RC Priori A631
Sadnicka A Ugawa Y amp Ziemann U (2014) Non-invasive632
cerebellar stimulationndasha consensus paper Cerebellum 13(1)633
121-138634
Groppa S Oliviero A Eisen A Quartarone A Cohen LG635
Mall V Kaelin-Lang A Mima T Rossi S Thickbroom636
GW Rossini PM Ziemann U Valls-Sole J amp Sieb-637
ner HR (2012) A practical guide to diagnostic transcranial638
magnetic stimulation Report of an IFCN committee Clin Neu-639
rophysiol 123(5) 858-882640
Hamada M Strigaro G Murase N Sadnicka A Galea JM 641
Edwards MJ amp Rothwell JC (2012) Cerebellar modulation 642
of human associative plasticity J Physiol 590(Pt 10) 2365- 643
2374 644
Hess CW Mills KR amp Murray NM (1987) Responses in small 645
hand muscles from magnetic stimulation of the human brain J 646
Physiol 388 397-419 647
Hull CA Chu Y Thanawala M amp Regehr WG (2013) Hyper- 648
polarization induces a long-term increase in the spontaneous 649
firing rate of cerebellar Golgi cells J Neurosci 33(14) 5895- 650
5902 651
Ihle K Rodriguez-Raecke R amp Luedtke K May A (2014) 652
tDCS modulates cortical nociceptive processing but has little 653
to no impact on pain perception Pain 155(10) 2080-2087 654
Ito M (2006) Cerebellar circuitry as a neuronal machine Prog 655
Neurobiol 78(3-5) 272-303 656
Iyer MB Mattu U Grafman J Lomarev M Sato S amp Wasser- 657
mann EM (2005) Safety and cognitive effect of frontal DC 658
brain polarization in healthy individuals Neurology 64(5) 872- 659
875 660
Kelly RM amp Strick PL (2003) Cerebellar loops with motor cor- 661
tex and prefrontal cortex of a nonhuman primate J Neurosci 662
23(23) 8432-8444 663
Lefaucheur JP Andre-Obadia N Antal A Ayache SS Baeken 664
C Benninger DH Cantello RM Cincotta M de Carvalho 665
M De Ridder D Devanne H Di Lazzaro V Filipovic SR 666
Hummel FC Jaaskelainen SK Kimiskidis VK Koch G 667
Langguth B Nyffeler T Oliviero A Padberg F Poulet E 668
Rossi S Rossini PM Rothwell JC Schonfeldt-Lecuona 669
C Siebner HR Slotema CW Stagg CJ Valls-Sole J 670
Ziemann U Paulus W amp Garcia-Larrea L (2014) Evidence- 671
based guidelines on the therapeutic use of repetitive transcranial 672
magnetic stimulation (rTMS) Clin Neurophysiol 125(11) 673
2150-2206 674
Lefaucheur JP Brusa A Creange A Drouot X amp Jarry G 675
(2002) Clinical application of laser evoked potentials using 676
the NdYAG laser Neurophysiol Clin 32(2) 91-98 677
Lefaucheur JP Debray S amp Jarry G (2001) Laser evoked poten- 678
tials using the NdYAG laser Muscle Nerve 24(4) 496-501 679
Lefaucheur JP Drouot X Menard-Lefaucheur I Keravel Y 680
amp Nguyen JP (2006) Motor cortex rTMS restores defective 681
intracortical inhibition in chronic neuropathic pain Neurology 682
67(9) 1568-1574 683
Longo MR Iannetti GD Mancini F Driver J amp Haggard P 684
(2012) Linking pain and the body Neural correlates of visually 685
induced analgesia J Neurosci 32(8) 2601-2607 686
Lorenz J amp Garcia-Larrea L (2003) Contribution of attentional 687
and cognitive factors to laser evoked brain potentials Neuro- 688
physiol Clin 33(6) 293-301 689
Lorenz J Minoshima S amp Casey KL (2003) Keeping pain out 690
of mind The role of the dorsolateral prefrontal cortex in pain 691
modulation Brain 126(Pt 5) 1079-1091 692
Luedtke K May A amp Jurgens TP (2012) No effect of a single 693
session of transcranial direct current stimulation on experimen- 694
tally induced pain in patients with chronic low back painndashan 695
exploratory study PLoS One 7(11) e48857 696
Unc
orre
cted
Aut
hor P
roof
12 T Bocci et al Cerebellum and pain A tcDCS study
Mendonca ME Santana MB Baptista AF Datta A Bikson697
M Fregni F amp Araujo CP (2011) Transcranial DC stimu-698
lation in fibromyalgia Optimized cortical target supported by699
high-resolution computational models J Pain 12(5) 610-617700
Moriguchi Y Decety J Ohnishi T Maeda M Mori T Nemoto701
K Matsuda H amp Komaki G (2007) Empathy and judg-702
ing otherrsquos pain An fMRI study of alexithymia Cereb Cortex703
17(9) 2223-2234704
Moulton EA Elman I Pendse G Schmahmann J Becerra L705
amp Borsook D (2011) Aversion-related circuitry in the cere-706
bellum Responses to noxious heat and unpleasant images J707
Neurosci 31(10) 3795-3804708
Moulton EA Schmahmann JD Becerra L amp Borsook D709
(2010) The cerebellum and pain Passive integrator or active710
participator Brain Res Rev 65(1) 14-27711
Mouraux A amp Plaghki L (2006) Are the processes reflected by712
late and ultra-late laser evoked potentials specific of nocicep-713
tion Suppl Clin Neurophysiol 59 197-204714
Mylius V Jung M Menzler K Haag A Khader PH Oer-715
tel WH Rosenow F amp Lefaucheur JP (2012) Effects of716
transcranial direct current stimulation on pain perception and717
working memory Eur J Pain 16(7) 974-982718
Ni Z Gunraj C amp Chen R (2007) Short interval intracortical719
inhibition and facilitation during the silent period in human J720
Physiol 583(Pt 3) 971-982721
Nitsche MA Liebetanz D Lang N Antal A Tergau F amp722
Paulus W (2003) Safety criteria for transcranial direct cur-723
rent stimulation (tDCS) in humans Clin Neurophysiol 114(11)724
2220-2222 author reply 2222-2223725
Opsommer E Guerit JM amp Plaghki L (2003) Exogenous726
and endogenous components of ultralate (C-fibre) evoked727
potentials following CO2 laser stimuli to tiny skin sur-728
face areas in healthy subjects Neurophysiol Clin 33(2)729
78-85730
Parazzini M Fiocchi S Liorni I Priori A amp Ravazzani P731
(2014) Computational modeling of transcranial direct current732
stimulation in the child brain Implications for the treatment of733
refractory childhood focal epilepsy Int J Neural Syst 24(2)734
1430006735
Parazzini M Rossi E Ferrucci R Fiocchi S Liorni I Priori736
A amp Ravazzani P (2013) Computational model of cerebellar737
transcranial direct current stimulation Conf Proc IEEE Eng738
Med Biol Soc 2013 237-240739
Parazzini M Rossi E Ferrucci R Liorni I Priori A amp Ravaz-740
zani P (2014) Modelling the electric field and the current741
density generated by cerebellar transcranial DC stimulation in742
humans Clin Neurophysiol 125(3) 577-584743
Perini I Bergstrand S amp Morrison I (2013) Where pain meets744
action in the human brain J Neurosci 33(40) 15930-15939745
Ploghaus A Tracey I Gati JS Clare S Menon RS Matthews746
PM amp Rawlins JN (1999) Dissociating pain from its antic-747
ipation in the human brain Science 284(5422) 1979-1981748
Priori A Ciocca M Parazzini M Vergari M amp Ferrucci R749
(2014) Transcranial cerebellar direct current stimulation and750
transcutaneous spinal cord direct current stimulation as innova-751
tive tools for neuroscientists J Physiol 592(Pt 16) 3345-3369752
Purpura DP amp McMurtry JC (1965) Intracellular activities and 753
evoked potential changes during polarization of motor cortex 754
J Neurophysiol 28 166-185 755
Qiu Y Inui K Wang X Tran TD amp Kakigi R (2002) Effects 756
of attention distraction and sleep on CO(2) laser evoked poten- 757
tials related to C-fibers in humans Clin Neurophysiol 113(10) 758
1579-1585 759
Reidler JS Mendonca ME Santana MB Wang X Lenkinski 760
R Motta AF Marchand S Latif L amp Fregni F (2012) 761
Effects of motor cortex modulation and descending inhibitory 762
systems on pain thresholds in healthy subjects J Pain 13(5) 763
450-458 764
Riedl V Valet M Woller A Sorg C Vogel D Sprenger 765
T Boecker H Wohlschlager AM amp Tolle TR (2011) 766
Repeated pain induces adaptations of intrinsic brain activity 767
to reflect past and predict future pain Neuroimage 57(1) 206- 768
213 769
Rocca MA Valsasina P Absinta M Colombo B Barcella V 770
Falini A Comi G amp Filippi M (2010) Central nervous 771
system dysregulation extends beyond the pain-matrix network 772
in cluster headache Cephalalgia 30(11) 1383-1391 773
Rossi S Hallett M Rossini PM amp Pascual-Leone A (2009) 774
Safety ethical considerations and application guidelines for 775
the use of transcranial magnetic stimulation in clinical practice 776
and research Clin Neurophysiol 120(12) 2008-2039 777
Rossini PM amp Rossi S (2007) Transcranial magnetic stimulation 778
Diagnostic therapeutic and research potential Neurology 779
68(7) 484-488 780
Ruscheweyh R Kuhnel M Filippopulos F Blum B Eggert 781
T amp Straube A (2014) Altered experimental pain perception 782
after cerebellar infarction Pain 155(7) 1303-1312 783
Schmahmann JD (1991) An emerging concept The cerebellar 784
contribution to higher function Arch Neurol 48(11) 1178- 785
1187 786
Singer T Seymour B OrsquoDoherty J Kaube H Dolan RJ amp 787
Frith CD (2004) Empathy for pain involves the affective but 788
not sensory components of pain Science 303(5661) 1157- 789
1162 790
Smith MJ Keel JC Greenberg BD Adams LF Schmidt PJ 791
Rubinow DA amp Wassermann EM (1999) Menstrual cycle 792
effects on cortical excitability Neurology 53(9) 2069-2072 793
Stoodley CJ amp Schmahmann JD (2009) Functional topography 794
in the human cerebellum A meta-analysis of neuroimaging 795
studies Neuroimage 44(2) 489-501 796
Strick PL Dum RP amp Fiez JA (2009) Cerebellum and non- 797
motor function Annu Rev Neurosci 32 413-434 798
Strigo IA Duncan GH Boivin M amp Bushnell MC (2003) 799
Differentiation of visceral and cutaneous pain in the human 800
brain J Neurophysiol 89(6) 3294-3303 801
Truini A Galeotti F Romaniello A Virtuoso M Iannetti GD 802
amp Cruccu G (2005) Laser-evoked potentials Normative val- 803
ues Clin Neurophysiol 116(4) 821-826 804
Truini A Panuccio G Galeotti F Maluccio MR Sartucci F 805
Avoli M amp Cruccu G (2010) Laser-evoked potentials as a 806
tool for assessing the efficacy of antinociceptive drugs Eur J 807
Pain 14(2) 222-225 808
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 13
Truini A Vergari M Biasiotta A La Cesa S Gabriele M Di809
Stefano G Cambieri C Cruccu G Inghilleri M amp Pri-810
ori A (2011) Transcutaneous spinal direct current stimulation811
inhibits nociceptive spinal pathway conduction and increases812
pain tolerance in humans Eur J Pain 15(10) 1023-1027813
Valeriani M Le Pera D Restuccia D de Armas L Mili-814
ucci R Betti V Vigevano F amp Tonali P (2007) Parallel815
spinal pathways generate the middle-latency N1 and the late P2816
components of the laser evoked potentials Clin Neurophysiol817
118(5) 1097-1104818
Valeriani M Pazzaglia C Cruccu G amp Truini A (2012) Clinical819
usefulness of laser evoked potentials Neurophysiol Clin 42(5)820
345-353821
Volz MS Mendonca M Pinheiro FS Cui H Santana M amp822
Fregni F (2012) Dissociation of motor task-induced cortical823
excitability and pain perception changes in healthy volunteers824
PLoS One 7(3) e34273825
Wager TD Rilling JK Smith EE Sokolik A Casey KL 826
Davidson RJ Kosslyn SM Rose RM amp Cohen JD 827
(2004) Placebo-induced changes in FMRI in the anticipation 828
and experience of pain Science 303(5661) 1162-1167 829
Zaghi S Thiele B Pimentel D Pimentel T amp Fregni 830
F (2011) Assessment and treatment of pain with non- 831
invasive cortical stimulation Restor Neurol Neurosci 29(6) 832
439-451 833
Zubieta JK Bueller JA Jackson LR Scott DJ Xu Y 834
Koeppe RA Nichols TE amp Stohler CS (2005) Placebo 835
effects mediated by endogenous opioid activity on mu-opioid 836
receptors J Neurosci 25(34) 7754-7762 837
Zunhammer M Busch V Griesbach F Landgrebe M Hajak G 838
amp Langguth B (2011) rTMS over the cerebellum modulates 839
temperature detection and pain thresholds through peripheral 840
mechanisms Brain Stimul 4(4) 210-7 e1 841

Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 5
Table 1
Row data (expressed as mean value plusmn 1 standard deviation a = anodal stimulation c = cathodal stimulation sh = sham condition)Both psychophysical and electrophysiological data for each subject are fully available as supplementary electronic material at
httpwwwenricasantarcangelocompublications
aT0 aT1 aT2 cT0 cT1 cT2 shT0 shT1 shT2
PT mean 462 607 609 485 376 368 472 466 489SD 080 095 092 086 062 067 098 062 081
VAS I1 mean 389 255 265 367 493 467 387 393 387SD 084 057 062 082 096 082 074 070 092
VAS I2 mean 540 402 403 524 673 665 533 549 530SD 063 082 071 048 047 049 078 069 064
N1 amplitude (V) mean 1292 848 801 1104 1496 1494 1101 1111 1121SD 318 298 258 265 258 333 250 267 283
N1 latency (ms) mean 12419 16146 15710 12704 10715 10405 12817 12867 13066SD 1090 1338 1368 1075 675 912 1320 1271 1209
N2P2 amplitude(V) mean 1114 738 757 1052 1453 1375 1114 1125 1147SD 262 237 233 265 296 329 272 269 216
N2P2 latency (ms) mean 15157 18932 18726 14878 12673 13230 15390 15108 15551SD 1312 1749 2139 2201 1849 1870 1433 1507 1675
233 Transcranial magnetic stimulation (TMS)236
Changes in Resting Motor Threshold (RMT) were237
evaluated at different intervals before and after the238
completion of tcDCS A Magstim Super Rapid Tran-239
scranial Magnetic Stimulator (Magstim Company240
Dyfed UK 22 T maximum field output) connected241
to a standard eight-shaped focal coil with wing diame-242
ters of 70 mm was used The handle of the eight-shaped243
focal coil was pointed backwards and rotated about 45244
deg to the mid-sagittal line to induce a tissue current245
perpendicular to the motor strip in the precentral sul-246
cus (Rossi et al 2009 Groppa et al 2012) RMT247
was defined as the minimum stimulator output that248
induces motor evoked potentials (MEPs) of more than249
50 V in at least five out of 10 trials when first dig-250
ital interosseus (FDI) muscle was completely relaxed251
(Ni et al 2007) The motor ldquohot spotrdquo for the targeted252
muscle was identified by single pulses of TMS deliv-253
ered at a slightly suprathreshold stimulus intensity and254
the magnetic stimuli induced monophasic pulses The255
coil was placed over the right motor cortex (centered on256
C4 according with the 10ndash20 EEG International Sys-257
tem) and electromyographic recordings were made by258
two standard non-polarizable AgAgCl surface elec-259
trodes (diameter 10 mm Biomedreg Florence Italy)260
one placed over the belly of the contralateral FDI261
muscle and the other on the skin overlying the first262
metacarpophalangeal joint of the first finger of the left263
hand RMT was evaluated to exclude possible cere-264
bellar stimulation spread out inducing motor cortex265
activation266
24 Variables and statistical analysis 267
We studied the subjective experience - percep- 268
tive threshold (PT) and pain intensity perceived 269
after laser I1 and I2 (VAS1 VAS2) - and elec- 270
trophysiological variables that is the peak-to-peak 271
amplitude of the N1 wave and N2P2 complex 272
the peak latency of N1 and N2 as reported in 273
previous papers using NdYAG laser (Lefaucheur 274
et al 2001 2002)275
Analyses were performed through SPSS15 sta- 276
tistical Package Psychophysical (PT VAS1 VAS2) 277
and electrophysiological variables (mean values of 278
ten traces N1amplitude and latency N2P2 ampli- 279
tude and latency) as well as Resting Motor Thresholds 280
(RMT) were analysed following a 3 Stimulation con- 281
ditions (anodal cathodal sham) times 3 Times (T0 T1 282
T2) design The Greenhouse-Geisser correction for 283
non sphericity was applied when necessary Con- 284
trast analysis between Times (F values) and paired 285
t tests between stimulations were alternatively used 286
for post-hoc comparisons when appropriate After 287
Bonferroni correction significance level was set at 288
p lt 0007289
The changes of all variable in T1 and T2 290
were expressed as ratio between post and pre 291
stimulation values (T1T0 T2T0) and compared 292
between each other according to a 2 Stimula- 293
tion (anodal cathodal) times 2 Times (T1To T2T0) 294
design 295
Unc
orre
cted
Aut
hor P
roof
6 T Bocci et al Cerebellum and pain A tcDCS study
Table 2
Contrast analyses all comparisons were highly significant(p lt 00001)
anodal cathodal shamPT time df F = 4430 F = 1867 ns
T0 vs Tl 228 F = 77669) F = 27523T0 vs T2 114 F = 78745 F = 27827Tl vs T2 114 ns ns
anodal vs sham cathodal vs shamT0 114 ns nsTl 114 t = 5069 t = 6991T2 114 t = 3709 t = 5849
VAS anodal cathodal sham
time 228 F = 41954 F = 31448 nsT0 vs Tl 114 F = 56968 F = 48596T0 vs T2 114 F = 52289 F = 525Tl vs T2 ns ns
anodal vs sham cathodal vs shamT0 114 ns nsTl 114 t = 644 t = 5916T2 114 t = 5294 t = 582
3 Results296
Row data (mean SD) are shown in Table 1 Base-297
line values were in line with those reported by earlier298
studies performed by using NdYAG laser (Lefaucheur299
et al 2001) Indeed only one study described a longer300
latency of the N2 wave (Cruccu et al 2008) The301
sham stimulation did not modulate any psychophys-302
ical and electrophysiological variable (Table 2) Since303
no pre-post difference was found for sham polarity this304
condition was not included in the comparison between305
Stimulations and Times306
31 Psychophysics307
PT exhibited a significant Stimulation effect308
(F(228) = 35055 p lt 00001 η2 = 0715) and a signifi-309
cant Stimulation times Time interaction (F(456) = 39464310
p lt 00001 η2 = 0738) Decomposition of the latter311
(Table 2) revealed that a) PT was higher for the anodal312
and lower for the cathodal stimulation conditions com-313
pared with the sham stimulation for both T1 and T2314
b) with respect to T0 PT increased in T1 and T2 in the315
anodal condition and decreased in the cathodal con-316
dition while no significant difference was observed317
between T1 and T2 (Fig 3A)318
Significantly different VAS1 and VAS2 were319
observed for the two stimulation intensities (F(114)320
= 54262 p lt 00001) and the three Stimulation con-321
ditions (F(218) = 88882 p lt 00001) Decomposition322
of the significant Stimulation times Time interaction323
(F(456) = 11596 p lt 00001) revealed that the reported 324
pain intensity for both stimulation intensities (VAS1 325
and VAS2) was higher for the cathodal and lower for 326
the anodal stimulation compared to the sham stimula- 327
tion (Table 2) It increased in T1 and decreased in T2 328
with respect to T0 whereas no significant difference 329
was found between T1 and T2 (Fig 3-B) 330
32 Laser evoked potentials 331
Figure 4-A shows the LEPs recorded in all experi- 332
mental conditions in a representative subject Both N1 333
and N2P2 amplitude (N1 F(456) = 10695 p lt 00001 334
η2 = 0884 N2P2 F(456) = 86864 p lt 00001 η2 = 335
0861) and latency (N1 F(456) = 110869 p lt 00001 336
η2 = 0888 N2P2 F(456) = 3660 p lt 00001 337
η2 = 0723) exhibited a significant Time times Stimulation 338
interaction Its decomposition (Table 3) showed 339
that both amplitudes increased and both latencies 340
decreased for cathodal stimulation in T1 and T2 with 341
respect to T0 the opposite occurred for the anodal 342
stimulation Both stimulations induced responses 343
significantly different from the sham condition 344
(Fig 4-B) The responses obtained after cathodal 345
stimulation were significantly improved (higher 346
amplitudes lower latencies) than those produced 347
by the anodal one (N1 amplitude F(114) = 41345 348
p lt 00001 N1 latency F(114) = 496228 p lt 00001 349
N2P2 amplitude (F(114) = 44537 p lt 00001 N2P2 350
latency F(114) = 119056 p lt 00001) 351
33 Resting motor thresholds 352
RMT values at baseline did not differ among exper- 353
imental conditions (mean plusmn SD sham 508 plusmn 83 354
anodal 491 plusmn 62 cathodal 503 plusmn 63) ANOVA 355
did not reveal any significant Stimulation (F(228) = 356
0882 p = 0425 η2 = 0059) Time (F(228) = 0212 357
p = 0810 η2 = 0015) and Stimulation times Time 358
effect (F(456) = 0339 p = 0851 η2 = 0024) for RMT 359
(Fig 5) 360
4 Discussion 361
Our study shows that cerebellar direct current 362
polarization modulates nociceptive perception and its 363
cortical correlates in healthy humans Specifically 364
cathodal tcDCS increases pain perception increases 365
amplitudes and decreases LEPs latencies likely though 366
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 7
A
B
Fig 3 - A Perceptive Threshold Changes (mean plusmn SD) at T1 and T2 with respect to baseline values (T1T0 T2T0) following sham (black)anodal (white) and cathodal (grey) tcDCS (lowastlowastp lt 0001 lowastlowastlowastp lt 00001) B Changes in visual analogue scale (VAS) scores over time VAS scoresat two different stimulus intensity respectively two (A left) and three (B right) times higher than the PT (lowastlowastp lt 0001 lowastlowastlowastp lt 00001)
A B
Fig 4 ndash A LEPs grand averaging traces were recorded at baseline (T0 black) and immediately after cerebellar polarization (T1 red) due tosham (left column) anodal (middle) and cathodal (right) tcDCS B Histograms showing LEPs variables and VAS scores changes (mean plusmn SD)after sham (black) anodal (white) or cathodal (grey) tcDCS with respect to baseline Top panels changes in N1 variables (amplitude and latency)over time bottom panels changes in N2P2 complex (lowastlowastp lt 0001 lowastlowastlowastp lt 00001)
Unc
orre
cted
Aut
hor P
roof
8 T Bocci et al Cerebellum and pain A tcDCS study
Table 3
LEPs post-hoc analyses p lt 00001 for all comparison except when explicitly indicated lowastlowastp lt 0002 lowast p lt 0005
N1 amplitude latency N2P2 amplitude latency
anodal cathodal shamdf
time 228 F = 67152 F = 96489 F = 134912 F = 34946 nsT0 vs T1 114 F = 109178 F = 18815 F = 165953 F = 64281T0 vs T2 114 F = 75143 F = 167697 F = 145125 F = 37818T1 vs T2 114 ns ns ns ns
anodal cathodaltime 228 F = 102281 F = 98717 F = 6577 F = 20918 nsT0 vs T1 114 F = 511186 F = 104027 F = 144112 F = 103864T0 vs T2 114 F = 96329 F = 116841 F = 105183 F = 14012lowastlowastT1 vs T2 114 ns ns ns ns ns
anodal vs sham anodal vs shamT0 114 ns ns ns nsT1 114 ns t = 925 t = 601 6262T2 114 ns t = 8128 t = 6731 5236
cathodal cathodal vs shamT0 114 ns ns t = 3281lowast nsT1 114 t = 16594 t = 8029 t = 8262 t = 520T2 114 t = 7309 t = 12669 t = 5048 t = 5301
Fig 5 - Resting Motor Thresholds Changes (mean plusmn SD) in Resting Motor threshold (RMT) expressed as percentage of the maximumstimulator output after sham (black) anodal (white) and cathodal (grey) tcDCS with respect to baseline marked as dotted line (lowastlowastp lt 0001lowastlowastlowastp lt 00001)
reduction of the inhibitory tone exerted by the cere-367
bellum on brain targets Anodal polarization elicits368
opposite effects producing analgesia Both findings369
support the role of the cerebellum in pain control370
it is noticeable that cathodal cerebellar stimulation371
induces hyperalgesia as occurs in patients with cere-372
bellar infarction (Ruscheweyh et al 2014)373
We would like to underline that in the present study374
LEPs were obtained at laser intensities depending on375
the perceptive threshold which varied as a function376
of anodal and cathodal stimulation This means that 377
the cerebellar stimulation has not a selective analgesic 378
effect as it influences both non nociceptive and noci- 379
ceptive perception A pre-eminent analgesic cannot be 380
assessed because the nociceptive threshold was not 381
evaluated 382
As tcDCS was effective on the modulation of 383
both N1 and N2P2 components and these responses 384
are generated by parallel and partially segregated 385
spinal pathways reaching different cortical targets 386
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 9
(Valeriani et al 2007) we may suggest that the cere-387
bellum is engaged in pain processing by modulating388
the activity of both somatosensory and cingulate cor-389
tices Indeed the cerebellum is involved in both the390
sensory-discriminative and emotional dimension of391
pain (Singer et al 2004 Moriguchi et al 2007) and392
non-invasive cerebellar current stimulation may modu-393
late pain experience and the associated cortical activity394
through many not alternative mechanisms In partic-395
ular changes in N1 reflects the modulation of the396
sensory component of pain while the vertex N2P2397
represents the neural correlate of affective aspects of398
pain experience (Garcia-Larrea et al 1997 Valeriani399
et al 2007) Notably tcDCS may act not only on spinal400
nociceptive neurons but also on wide-range cortical401
networks of the pain matrix (Singer et al 2004) thus402
influencing LEPs and pain experience through both403
top-down and bottom-up mechanisms404
The present study does not allow to hypothesize405
how and where tcDCS influences the cerebellar activ-406
ity A main role of Purkinje cells has been suggested407
as their activity modulation may affect the cerebellar408
inhibitory control of the cerebral cortex (Galea et al409
2009) This would be in line with the effects elicited410
by tDCS in the cerebral cortex which are observ-411
able after both short and long term delay likely also412
interfering with long-term potentiation (LTP)-like phe-413
nomena (Hamada et al 2012 Priori et al 2014)414
Moreover prolonged spiking activity in the cerebellar415
Golgi inhibitory neurons modulates the activity of the416
Purkinje cells and could partly account for the tcDCS417
after-effects (Hull et al 2013)418
The lack of changes in RMT indicates that the anal-419
gesic effects of anodal tcDCS are due to a specific420
modulation of the cerebellar activity and not to motor421
activation On the other hand tcDCS-induced cerebel-422
lar modulation (Purpura amp McMurtry 1965) could be423
not sufficient per se to activate the cerebello ndash thalamo424
- cortical motor pathway (Galea et al 2009) thus425
the reported analgesia and its cortical correlates can-426
not be sustained by the motor cortex activation This427
view is supported by the absence of any association428
between motor symptoms and pain perception in cere-429
bellar patients (Ruscheweyh et al 2014) In the same430
line in healthy subjects it has been recently shown431
that motor task-induced increased cortical excitability432
and analgesia are not associated (Volz et al 2012)433
Indeed RMT is a highly sensitive marker of motor434
tract excitability as it reflects activation of a small435
low-threshold and slow-conducting core of pyramidal436
neurons (Hess et al 1987 Rossini amp Rossi 2007) 437
although RMT may reflect changes in the activity of 438
different central nervous system structures it has been 439
satisfactorily used to assess motor cortex excitability 440
also in cerebellar patients (Battaglia et al 2006) 441
Another critical point is the possibility to modulate 442
with tcDCS both neural correlates underlying nocicep- 443
tive processing and pain perception Previous studies 444
using tDCS over motor cortex were inconsistent among 445
each other some works suggested that tDCS is able to 446
modify pain perception (Boggio et al 2008) while 447
others showed divergent effects on psychophysical 448
and neurophysiological outcome parameters (Luedtke 449
et al 2012 Ihle et al 2014) likely due to a possi- 450
ble overestimation of the role of motor areas on pain 451
processing (Antal et al 2008) 452
Our findings cannot be compared to the results 453
obtained by other Authors In fact the unique study 454
focused on the analgesic effects of non-invasive cere- 455
bellar stimulation reported till now (Zunhammer et al 456
2011) considered only subjective pain thresholds 457
In addition it described similar analgesic effects of 458
cerebellar and neck structures repetitive transcranial 459
magnetic stimulation (rTMS) thus denying any cere- 460
bellar specificity in the observed effects and suggesting 461
that the peripheral information passing through the 462
cerebellum may be responsible for analgesia The 463
main difference between the two studies possibly 464
accounting for different results consists of the neu- 465
romodulation techniques used 466
41 Limitations of the study 467
The present study has a few limitations First our 468
findings do not allow any hypothesis on the role of 469
the cerebellum in chronic pain The observations on 470
patients with cerebellar damage (Ruscheweyh et al 471
2014) suggest that their impaired inhibitory control 472
mechanisms may be not associated with the devel- 473
opment of chronic pain Second we cannot exclude 474
the possibility that tcDCS could modulate not only the 475
cerebellum but also surrounding areas such as the peri- 476
aqueductal gray However recent modelling researches 477
have revealed that during tcDCS the current spread 478
to other structures outside the cerebellum is negligi- 479
ble (Parazzini et al 2013 2014) Moreover several 480
studies have proved that in humans pain processing 481
is encoded within posterior areas of each cerebellar 482
hemisphere specifically in the hemispheric lobule VI 483
Crus I and VIIb (Moulton et al 2011) where the 484
Unc
orre
cted
Aut
hor P
roof
10 T Bocci et al Cerebellum and pain A tcDCS study
tcDCS-induced electrical field is strongly concentrated485
(Parazzini et al 2013) A further limitation is that we486
did not study the contribution of C-fibers the main487
component of spino-ponto-cerebellar and spino-olivo-488
cerebellar pathways In fact ultra-late LEPs related489
to C-fibers activations have not yet been standard-490
ized for clinical application and their occurrence could491
be markedly influenced by high order cognitive pro-492
cesses as they seem to be more affected by the level493
of consciousness and attention than A-delta responses494
(Qiu et al 2002 Opsommer et al 2003 Mouraux amp495
Plaghki 2006) Finally we wish to emphasize that in496
neuropathic patients the effects of the cerebellar stim-497
ulation could be quite different from those described498
here as both anatomical and functional connectivity499
are different from those observed in healthy partici-500
pants (Rocca et al 2010 Riedl et al 2011 Absinta et501
al 2012 Longo et al 2012 Ceko et al 2013)502
5 Conclusions503
Our findings indicate a cerebellar effect on pain504
experience and on its cortical correlates and prompt505
further investigation aimed at assessing whether the506
cerebellar direct current polarization could be used as a507
novel and safe therapeutic tool in chronic pain patients508
Acknowledgments509
We gratefully acknowledge the participation of all510
subjects as well as Mr D Barloscio and Mr C511
Orsini for their excellent technical assistance The512
paper was supported in part by the Italian operating513
and development MIUR PRIN grant year 2006 n514
2006062332 002515
RF and AP are founders and shareholders of516
Newronika srl Milan Italy517
References518
Absinta M Rocca MA Colombo B Falini A Comi G amp Fil-519
ippi M (2012) Selective decreased grey matter volume of the520
pain-matrix network in cluster headache Cephalalgia 32(2)521
109-115522
Agostino R Cruccu G Iannetti G Romaniello A Truini A523
amp Manfredi M (2000) Topographical distribution of pinprick524
and warmth thresholds to CO2 laser stimulation on the human525
skin Neurosci Lett 285(2) 115-118526
Antal A Brepohl N Poreisz C Boros K Csifcsak G amp 527
Paulus W (2008) Transcranial direct current stimulation over 528
somatosensory cortex decreases experimentally induced acute 529
pain perception Clin J Pain 24(1) 56-63 530
Antal A amp Paulus W (2010) Effects of transcranial theta-burst 531
stimulation on acute pain perception Restor Neurol Neurosci 532
28(4) 477-484 533
Apkarian AV Bushnell MC Treede RD amp Zubieta JK 534
(2005) Human brain mechanisms of pain perception and reg- 535
ulation in health and disease Eur J Pain 9(4) 463-484 536
Balsters JH Whelan CD Robertson IH amp Ramnani N (2013) 537
Cerebellum and cognition Evidence for the encoding of higher 538
order rules Cereb Cortex 23(6) 1433-1443 539
Battaglia F Quartarone A Ghilardi MF Dattola R Bagnato 540
S Rizzo V Morgante L amp Girlanda P (2006) Unilat- 541
eral cerebellar stroke disrupts movement preparation and motor 542
imagery Clin Neurophysiol 117(5) 1009-1016 543
Bingel U Quante M Knab R Bromm B Weiller C amp Buchel 544
C (2002) Subcortical structures involved in pain processing 545
Evidence from single-trial fMRI Pain 99(1-2) 313-321 546
Boggio PS Zaghi S amp Fregni F (2009) Modulation of emotions 547
associated with images of human pain using anodal transcranial 548
direct current stimulation (tDCS) Neuropsychologia 47(1) 549
212-217 550
Boggio PS Zaghi S Lopes M amp Fregni F (2008) Modulatory 551
effects of anodal transcranial direct current stimulation on per- 552
ception and pain thresholds in healthy volunteers Eur J Neurol 553
15(10) 1124-1130 554
Borsook D Moulton EA Tully S Schmahmann JD amp 555
Becerra L (2008) Human cerebellar responses to brush and 556
heat stimuli in healthy and neuropathic pain subjects Cerebel- 557
lum 7(3) 252-272 558
Ceko M Seminowicz DA Bushnell MC amp Olausson HW 559
(2013) Anatomical and functional enhancements of the insula 560
after loss of large primary somatosensory fibers Cereb Cortex 561
23(9) 2017-2024 562
Cruccu G Aminoff MJ Curio G Guerit JM Kakigi 563
R Mauguiere F Rossini PM Treede RD amp Garcia- 564
Larrea L (2008) Recommendations for the clinical use of 565
somatosensory-evoked potentials Clin Neurophysiol 119(8) 566
1705-1719 567
Cruccu G Romaniello A Amantini A Lombardi M Innocenti 568
P amp Manfredi M (1999) Assessment of trigeminal small- 569
fiber function Brain and reflex responses evoked by CO2-laser 570
stimulation Muscle Nerve 22(4) 508-516 571
Dasilva AF Mendonca ME Zaghi S Lopes M Dossantos 572
MF Spierings EL Bajwa Z Datta A Bikson M amp 573
Fregni F (2012) tDCS-induced analgesia and electrical fields 574
in pain-related neural networks in chronic migraine Headache 575
52(8) 1283-1295 576
Duerden EG amp Albanese MC (2013) Localization of pain- 577
related brain activation A meta-analysis of neuroimaging data 578
Hum Brain Mapp 34(1) 109-149 579
Ekerot CF Garwicz M amp Schouenborg J (1991a) The postsy- 580
naptic dorsal column pathway mediates cutaneous nociceptive 581
information to cerebellar climbing fibres in the cat J Physiol 582
441 275-284 583
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 11
Ekerot CF Garwicz M amp Schouenborg J (1991b) Topography584
and nociceptive receptive fields of climbing fibres projecting to585
the cerebellar anterior lobe in the cat J Physiol 441 257-274586
Ekerot CF Gustavsson P Oscarsson O amp Schouenborg J587
(1987a) Climbing fibres projecting to cat cerebellar anterior588
lobe activated by cutaneous A and C fibres J Physiol 386589
529-538590
Ekerot CF Oscarsson O amp Schouenborg J (1987b) Stimula-591
tion of cat cutaneous nociceptive C fibres causing tonic and592
synchronous activity in climbing fibres J Physiol 386 539-593
546594
Ferrucci R Brunoni AR Parazzini M Vergari M Rossi E595
Fumagalli M Mameli F Rosa M Giannicola G Zago S596
amp Priori A (2013) Modulating human procedural learning by597
cerebellar transcranial direct current stimulation Cerebellum598
12(4) 485-492599
Ferrucci R Giannicola G Rosa M Fumagalli M Boggio600
PS Hallett M Zago S amp Priori A (2012) Cerebellum601
and processing of negative facial emotions Cerebellar tran-602
scranial DC stimulation specifically enhances the emotional603
recognition of facial anger and sadness Cogn Emot 26(5)604
786-799605
Ferrucci R Marceglia S Vergari M Cogiamanian F Mrakic-606
Sposta S Mameli F Zago S Barbieri S amp Priori607
A (2008) Cerebellar transcranial direct current stimulation608
impairs the practice-dependent proficiency increase in working609
memory J Cogn Neurosci 20(9) 1687-1697610
Fregni F Freedman S amp Pascual-Leone A (2007) Recent611
advances in the treatment of chronic pain with non-invasive612
brain stimulation techniques Lancet Neurol 6(2) 188-191613
Galea JM Jayaram G Ajagbe L amp Celnik P (2009)614
Modulation of cerebellar excitability by polarity-specific615
noninvasive direct current stimulation J Neurosci 29(28)616
9115-9122617
Galea JM Vazquez A Pasricha N de Xivry JJ amp Celnik618
P (2011) Dissociating the roles of the cerebellum and motor619
cortex during adaptive learning The motor cortex retains what620
the cerebellum learns Cereb Cortex 21(8) 1761-1770621
Gandiga PC Hummel FC amp Cohen LG (2006) Tran-622
scranial DC stimulation (tDCS) A tool for double-blind623
sham-controlled clinical studies in brain stimulation Clin Neu-624
rophysiol 117(4) 845-850625
Garcia-Larrea L Peyron R Laurent B amp Mauguiere F (1997)626
Association and dissociation between laser-evoked potentials627
and pain perception Neuroreport 8(17) 3785-3789628
Grimaldi G Argyropoulos GP Boehringer A Celnik P629
Edwards MJ Ferrucci R Galea JM Groiss SJ Hiraoka630
K Kassavetis P Lesage E Manto M Miall RC Priori A631
Sadnicka A Ugawa Y amp Ziemann U (2014) Non-invasive632
cerebellar stimulationndasha consensus paper Cerebellum 13(1)633
121-138634
Groppa S Oliviero A Eisen A Quartarone A Cohen LG635
Mall V Kaelin-Lang A Mima T Rossi S Thickbroom636
GW Rossini PM Ziemann U Valls-Sole J amp Sieb-637
ner HR (2012) A practical guide to diagnostic transcranial638
magnetic stimulation Report of an IFCN committee Clin Neu-639
rophysiol 123(5) 858-882640
Hamada M Strigaro G Murase N Sadnicka A Galea JM 641
Edwards MJ amp Rothwell JC (2012) Cerebellar modulation 642
of human associative plasticity J Physiol 590(Pt 10) 2365- 643
2374 644
Hess CW Mills KR amp Murray NM (1987) Responses in small 645
hand muscles from magnetic stimulation of the human brain J 646
Physiol 388 397-419 647
Hull CA Chu Y Thanawala M amp Regehr WG (2013) Hyper- 648
polarization induces a long-term increase in the spontaneous 649
firing rate of cerebellar Golgi cells J Neurosci 33(14) 5895- 650
5902 651
Ihle K Rodriguez-Raecke R amp Luedtke K May A (2014) 652
tDCS modulates cortical nociceptive processing but has little 653
to no impact on pain perception Pain 155(10) 2080-2087 654
Ito M (2006) Cerebellar circuitry as a neuronal machine Prog 655
Neurobiol 78(3-5) 272-303 656
Iyer MB Mattu U Grafman J Lomarev M Sato S amp Wasser- 657
mann EM (2005) Safety and cognitive effect of frontal DC 658
brain polarization in healthy individuals Neurology 64(5) 872- 659
875 660
Kelly RM amp Strick PL (2003) Cerebellar loops with motor cor- 661
tex and prefrontal cortex of a nonhuman primate J Neurosci 662
23(23) 8432-8444 663
Lefaucheur JP Andre-Obadia N Antal A Ayache SS Baeken 664
C Benninger DH Cantello RM Cincotta M de Carvalho 665
M De Ridder D Devanne H Di Lazzaro V Filipovic SR 666
Hummel FC Jaaskelainen SK Kimiskidis VK Koch G 667
Langguth B Nyffeler T Oliviero A Padberg F Poulet E 668
Rossi S Rossini PM Rothwell JC Schonfeldt-Lecuona 669
C Siebner HR Slotema CW Stagg CJ Valls-Sole J 670
Ziemann U Paulus W amp Garcia-Larrea L (2014) Evidence- 671
based guidelines on the therapeutic use of repetitive transcranial 672
magnetic stimulation (rTMS) Clin Neurophysiol 125(11) 673
2150-2206 674
Lefaucheur JP Brusa A Creange A Drouot X amp Jarry G 675
(2002) Clinical application of laser evoked potentials using 676
the NdYAG laser Neurophysiol Clin 32(2) 91-98 677
Lefaucheur JP Debray S amp Jarry G (2001) Laser evoked poten- 678
tials using the NdYAG laser Muscle Nerve 24(4) 496-501 679
Lefaucheur JP Drouot X Menard-Lefaucheur I Keravel Y 680
amp Nguyen JP (2006) Motor cortex rTMS restores defective 681
intracortical inhibition in chronic neuropathic pain Neurology 682
67(9) 1568-1574 683
Longo MR Iannetti GD Mancini F Driver J amp Haggard P 684
(2012) Linking pain and the body Neural correlates of visually 685
induced analgesia J Neurosci 32(8) 2601-2607 686
Lorenz J amp Garcia-Larrea L (2003) Contribution of attentional 687
and cognitive factors to laser evoked brain potentials Neuro- 688
physiol Clin 33(6) 293-301 689
Lorenz J Minoshima S amp Casey KL (2003) Keeping pain out 690
of mind The role of the dorsolateral prefrontal cortex in pain 691
modulation Brain 126(Pt 5) 1079-1091 692
Luedtke K May A amp Jurgens TP (2012) No effect of a single 693
session of transcranial direct current stimulation on experimen- 694
tally induced pain in patients with chronic low back painndashan 695
exploratory study PLoS One 7(11) e48857 696
Unc
orre
cted
Aut
hor P
roof
12 T Bocci et al Cerebellum and pain A tcDCS study
Mendonca ME Santana MB Baptista AF Datta A Bikson697
M Fregni F amp Araujo CP (2011) Transcranial DC stimu-698
lation in fibromyalgia Optimized cortical target supported by699
high-resolution computational models J Pain 12(5) 610-617700
Moriguchi Y Decety J Ohnishi T Maeda M Mori T Nemoto701
K Matsuda H amp Komaki G (2007) Empathy and judg-702
ing otherrsquos pain An fMRI study of alexithymia Cereb Cortex703
17(9) 2223-2234704
Moulton EA Elman I Pendse G Schmahmann J Becerra L705
amp Borsook D (2011) Aversion-related circuitry in the cere-706
bellum Responses to noxious heat and unpleasant images J707
Neurosci 31(10) 3795-3804708
Moulton EA Schmahmann JD Becerra L amp Borsook D709
(2010) The cerebellum and pain Passive integrator or active710
participator Brain Res Rev 65(1) 14-27711
Mouraux A amp Plaghki L (2006) Are the processes reflected by712
late and ultra-late laser evoked potentials specific of nocicep-713
tion Suppl Clin Neurophysiol 59 197-204714
Mylius V Jung M Menzler K Haag A Khader PH Oer-715
tel WH Rosenow F amp Lefaucheur JP (2012) Effects of716
transcranial direct current stimulation on pain perception and717
working memory Eur J Pain 16(7) 974-982718
Ni Z Gunraj C amp Chen R (2007) Short interval intracortical719
inhibition and facilitation during the silent period in human J720
Physiol 583(Pt 3) 971-982721
Nitsche MA Liebetanz D Lang N Antal A Tergau F amp722
Paulus W (2003) Safety criteria for transcranial direct cur-723
rent stimulation (tDCS) in humans Clin Neurophysiol 114(11)724
2220-2222 author reply 2222-2223725
Opsommer E Guerit JM amp Plaghki L (2003) Exogenous726
and endogenous components of ultralate (C-fibre) evoked727
potentials following CO2 laser stimuli to tiny skin sur-728
face areas in healthy subjects Neurophysiol Clin 33(2)729
78-85730
Parazzini M Fiocchi S Liorni I Priori A amp Ravazzani P731
(2014) Computational modeling of transcranial direct current732
stimulation in the child brain Implications for the treatment of733
refractory childhood focal epilepsy Int J Neural Syst 24(2)734
1430006735
Parazzini M Rossi E Ferrucci R Fiocchi S Liorni I Priori736
A amp Ravazzani P (2013) Computational model of cerebellar737
transcranial direct current stimulation Conf Proc IEEE Eng738
Med Biol Soc 2013 237-240739
Parazzini M Rossi E Ferrucci R Liorni I Priori A amp Ravaz-740
zani P (2014) Modelling the electric field and the current741
density generated by cerebellar transcranial DC stimulation in742
humans Clin Neurophysiol 125(3) 577-584743
Perini I Bergstrand S amp Morrison I (2013) Where pain meets744
action in the human brain J Neurosci 33(40) 15930-15939745
Ploghaus A Tracey I Gati JS Clare S Menon RS Matthews746
PM amp Rawlins JN (1999) Dissociating pain from its antic-747
ipation in the human brain Science 284(5422) 1979-1981748
Priori A Ciocca M Parazzini M Vergari M amp Ferrucci R749
(2014) Transcranial cerebellar direct current stimulation and750
transcutaneous spinal cord direct current stimulation as innova-751
tive tools for neuroscientists J Physiol 592(Pt 16) 3345-3369752
Purpura DP amp McMurtry JC (1965) Intracellular activities and 753
evoked potential changes during polarization of motor cortex 754
J Neurophysiol 28 166-185 755
Qiu Y Inui K Wang X Tran TD amp Kakigi R (2002) Effects 756
of attention distraction and sleep on CO(2) laser evoked poten- 757
tials related to C-fibers in humans Clin Neurophysiol 113(10) 758
1579-1585 759
Reidler JS Mendonca ME Santana MB Wang X Lenkinski 760
R Motta AF Marchand S Latif L amp Fregni F (2012) 761
Effects of motor cortex modulation and descending inhibitory 762
systems on pain thresholds in healthy subjects J Pain 13(5) 763
450-458 764
Riedl V Valet M Woller A Sorg C Vogel D Sprenger 765
T Boecker H Wohlschlager AM amp Tolle TR (2011) 766
Repeated pain induces adaptations of intrinsic brain activity 767
to reflect past and predict future pain Neuroimage 57(1) 206- 768
213 769
Rocca MA Valsasina P Absinta M Colombo B Barcella V 770
Falini A Comi G amp Filippi M (2010) Central nervous 771
system dysregulation extends beyond the pain-matrix network 772
in cluster headache Cephalalgia 30(11) 1383-1391 773
Rossi S Hallett M Rossini PM amp Pascual-Leone A (2009) 774
Safety ethical considerations and application guidelines for 775
the use of transcranial magnetic stimulation in clinical practice 776
and research Clin Neurophysiol 120(12) 2008-2039 777
Rossini PM amp Rossi S (2007) Transcranial magnetic stimulation 778
Diagnostic therapeutic and research potential Neurology 779
68(7) 484-488 780
Ruscheweyh R Kuhnel M Filippopulos F Blum B Eggert 781
T amp Straube A (2014) Altered experimental pain perception 782
after cerebellar infarction Pain 155(7) 1303-1312 783
Schmahmann JD (1991) An emerging concept The cerebellar 784
contribution to higher function Arch Neurol 48(11) 1178- 785
1187 786
Singer T Seymour B OrsquoDoherty J Kaube H Dolan RJ amp 787
Frith CD (2004) Empathy for pain involves the affective but 788
not sensory components of pain Science 303(5661) 1157- 789
1162 790
Smith MJ Keel JC Greenberg BD Adams LF Schmidt PJ 791
Rubinow DA amp Wassermann EM (1999) Menstrual cycle 792
effects on cortical excitability Neurology 53(9) 2069-2072 793
Stoodley CJ amp Schmahmann JD (2009) Functional topography 794
in the human cerebellum A meta-analysis of neuroimaging 795
studies Neuroimage 44(2) 489-501 796
Strick PL Dum RP amp Fiez JA (2009) Cerebellum and non- 797
motor function Annu Rev Neurosci 32 413-434 798
Strigo IA Duncan GH Boivin M amp Bushnell MC (2003) 799
Differentiation of visceral and cutaneous pain in the human 800
brain J Neurophysiol 89(6) 3294-3303 801
Truini A Galeotti F Romaniello A Virtuoso M Iannetti GD 802
amp Cruccu G (2005) Laser-evoked potentials Normative val- 803
ues Clin Neurophysiol 116(4) 821-826 804
Truini A Panuccio G Galeotti F Maluccio MR Sartucci F 805
Avoli M amp Cruccu G (2010) Laser-evoked potentials as a 806
tool for assessing the efficacy of antinociceptive drugs Eur J 807
Pain 14(2) 222-225 808
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 13
Truini A Vergari M Biasiotta A La Cesa S Gabriele M Di809
Stefano G Cambieri C Cruccu G Inghilleri M amp Pri-810
ori A (2011) Transcutaneous spinal direct current stimulation811
inhibits nociceptive spinal pathway conduction and increases812
pain tolerance in humans Eur J Pain 15(10) 1023-1027813
Valeriani M Le Pera D Restuccia D de Armas L Mili-814
ucci R Betti V Vigevano F amp Tonali P (2007) Parallel815
spinal pathways generate the middle-latency N1 and the late P2816
components of the laser evoked potentials Clin Neurophysiol817
118(5) 1097-1104818
Valeriani M Pazzaglia C Cruccu G amp Truini A (2012) Clinical819
usefulness of laser evoked potentials Neurophysiol Clin 42(5)820
345-353821
Volz MS Mendonca M Pinheiro FS Cui H Santana M amp822
Fregni F (2012) Dissociation of motor task-induced cortical823
excitability and pain perception changes in healthy volunteers824
PLoS One 7(3) e34273825
Wager TD Rilling JK Smith EE Sokolik A Casey KL 826
Davidson RJ Kosslyn SM Rose RM amp Cohen JD 827
(2004) Placebo-induced changes in FMRI in the anticipation 828
and experience of pain Science 303(5661) 1162-1167 829
Zaghi S Thiele B Pimentel D Pimentel T amp Fregni 830
F (2011) Assessment and treatment of pain with non- 831
invasive cortical stimulation Restor Neurol Neurosci 29(6) 832
439-451 833
Zubieta JK Bueller JA Jackson LR Scott DJ Xu Y 834
Koeppe RA Nichols TE amp Stohler CS (2005) Placebo 835
effects mediated by endogenous opioid activity on mu-opioid 836
receptors J Neurosci 25(34) 7754-7762 837
Zunhammer M Busch V Griesbach F Landgrebe M Hajak G 838
amp Langguth B (2011) rTMS over the cerebellum modulates 839
temperature detection and pain thresholds through peripheral 840
mechanisms Brain Stimul 4(4) 210-7 e1 841

Unc
orre
cted
Aut
hor P
roof
6 T Bocci et al Cerebellum and pain A tcDCS study
Table 2
Contrast analyses all comparisons were highly significant(p lt 00001)
anodal cathodal shamPT time df F = 4430 F = 1867 ns
T0 vs Tl 228 F = 77669) F = 27523T0 vs T2 114 F = 78745 F = 27827Tl vs T2 114 ns ns
anodal vs sham cathodal vs shamT0 114 ns nsTl 114 t = 5069 t = 6991T2 114 t = 3709 t = 5849
VAS anodal cathodal sham
time 228 F = 41954 F = 31448 nsT0 vs Tl 114 F = 56968 F = 48596T0 vs T2 114 F = 52289 F = 525Tl vs T2 ns ns
anodal vs sham cathodal vs shamT0 114 ns nsTl 114 t = 644 t = 5916T2 114 t = 5294 t = 582
3 Results296
Row data (mean SD) are shown in Table 1 Base-297
line values were in line with those reported by earlier298
studies performed by using NdYAG laser (Lefaucheur299
et al 2001) Indeed only one study described a longer300
latency of the N2 wave (Cruccu et al 2008) The301
sham stimulation did not modulate any psychophys-302
ical and electrophysiological variable (Table 2) Since303
no pre-post difference was found for sham polarity this304
condition was not included in the comparison between305
Stimulations and Times306
31 Psychophysics307
PT exhibited a significant Stimulation effect308
(F(228) = 35055 p lt 00001 η2 = 0715) and a signifi-309
cant Stimulation times Time interaction (F(456) = 39464310
p lt 00001 η2 = 0738) Decomposition of the latter311
(Table 2) revealed that a) PT was higher for the anodal312
and lower for the cathodal stimulation conditions com-313
pared with the sham stimulation for both T1 and T2314
b) with respect to T0 PT increased in T1 and T2 in the315
anodal condition and decreased in the cathodal con-316
dition while no significant difference was observed317
between T1 and T2 (Fig 3A)318
Significantly different VAS1 and VAS2 were319
observed for the two stimulation intensities (F(114)320
= 54262 p lt 00001) and the three Stimulation con-321
ditions (F(218) = 88882 p lt 00001) Decomposition322
of the significant Stimulation times Time interaction323
(F(456) = 11596 p lt 00001) revealed that the reported 324
pain intensity for both stimulation intensities (VAS1 325
and VAS2) was higher for the cathodal and lower for 326
the anodal stimulation compared to the sham stimula- 327
tion (Table 2) It increased in T1 and decreased in T2 328
with respect to T0 whereas no significant difference 329
was found between T1 and T2 (Fig 3-B) 330
32 Laser evoked potentials 331
Figure 4-A shows the LEPs recorded in all experi- 332
mental conditions in a representative subject Both N1 333
and N2P2 amplitude (N1 F(456) = 10695 p lt 00001 334
η2 = 0884 N2P2 F(456) = 86864 p lt 00001 η2 = 335
0861) and latency (N1 F(456) = 110869 p lt 00001 336
η2 = 0888 N2P2 F(456) = 3660 p lt 00001 337
η2 = 0723) exhibited a significant Time times Stimulation 338
interaction Its decomposition (Table 3) showed 339
that both amplitudes increased and both latencies 340
decreased for cathodal stimulation in T1 and T2 with 341
respect to T0 the opposite occurred for the anodal 342
stimulation Both stimulations induced responses 343
significantly different from the sham condition 344
(Fig 4-B) The responses obtained after cathodal 345
stimulation were significantly improved (higher 346
amplitudes lower latencies) than those produced 347
by the anodal one (N1 amplitude F(114) = 41345 348
p lt 00001 N1 latency F(114) = 496228 p lt 00001 349
N2P2 amplitude (F(114) = 44537 p lt 00001 N2P2 350
latency F(114) = 119056 p lt 00001) 351
33 Resting motor thresholds 352
RMT values at baseline did not differ among exper- 353
imental conditions (mean plusmn SD sham 508 plusmn 83 354
anodal 491 plusmn 62 cathodal 503 plusmn 63) ANOVA 355
did not reveal any significant Stimulation (F(228) = 356
0882 p = 0425 η2 = 0059) Time (F(228) = 0212 357
p = 0810 η2 = 0015) and Stimulation times Time 358
effect (F(456) = 0339 p = 0851 η2 = 0024) for RMT 359
(Fig 5) 360
4 Discussion 361
Our study shows that cerebellar direct current 362
polarization modulates nociceptive perception and its 363
cortical correlates in healthy humans Specifically 364
cathodal tcDCS increases pain perception increases 365
amplitudes and decreases LEPs latencies likely though 366
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 7
A
B
Fig 3 - A Perceptive Threshold Changes (mean plusmn SD) at T1 and T2 with respect to baseline values (T1T0 T2T0) following sham (black)anodal (white) and cathodal (grey) tcDCS (lowastlowastp lt 0001 lowastlowastlowastp lt 00001) B Changes in visual analogue scale (VAS) scores over time VAS scoresat two different stimulus intensity respectively two (A left) and three (B right) times higher than the PT (lowastlowastp lt 0001 lowastlowastlowastp lt 00001)
A B
Fig 4 ndash A LEPs grand averaging traces were recorded at baseline (T0 black) and immediately after cerebellar polarization (T1 red) due tosham (left column) anodal (middle) and cathodal (right) tcDCS B Histograms showing LEPs variables and VAS scores changes (mean plusmn SD)after sham (black) anodal (white) or cathodal (grey) tcDCS with respect to baseline Top panels changes in N1 variables (amplitude and latency)over time bottom panels changes in N2P2 complex (lowastlowastp lt 0001 lowastlowastlowastp lt 00001)
Unc
orre
cted
Aut
hor P
roof
8 T Bocci et al Cerebellum and pain A tcDCS study
Table 3
LEPs post-hoc analyses p lt 00001 for all comparison except when explicitly indicated lowastlowastp lt 0002 lowast p lt 0005
N1 amplitude latency N2P2 amplitude latency
anodal cathodal shamdf
time 228 F = 67152 F = 96489 F = 134912 F = 34946 nsT0 vs T1 114 F = 109178 F = 18815 F = 165953 F = 64281T0 vs T2 114 F = 75143 F = 167697 F = 145125 F = 37818T1 vs T2 114 ns ns ns ns
anodal cathodaltime 228 F = 102281 F = 98717 F = 6577 F = 20918 nsT0 vs T1 114 F = 511186 F = 104027 F = 144112 F = 103864T0 vs T2 114 F = 96329 F = 116841 F = 105183 F = 14012lowastlowastT1 vs T2 114 ns ns ns ns ns
anodal vs sham anodal vs shamT0 114 ns ns ns nsT1 114 ns t = 925 t = 601 6262T2 114 ns t = 8128 t = 6731 5236
cathodal cathodal vs shamT0 114 ns ns t = 3281lowast nsT1 114 t = 16594 t = 8029 t = 8262 t = 520T2 114 t = 7309 t = 12669 t = 5048 t = 5301
Fig 5 - Resting Motor Thresholds Changes (mean plusmn SD) in Resting Motor threshold (RMT) expressed as percentage of the maximumstimulator output after sham (black) anodal (white) and cathodal (grey) tcDCS with respect to baseline marked as dotted line (lowastlowastp lt 0001lowastlowastlowastp lt 00001)
reduction of the inhibitory tone exerted by the cere-367
bellum on brain targets Anodal polarization elicits368
opposite effects producing analgesia Both findings369
support the role of the cerebellum in pain control370
it is noticeable that cathodal cerebellar stimulation371
induces hyperalgesia as occurs in patients with cere-372
bellar infarction (Ruscheweyh et al 2014)373
We would like to underline that in the present study374
LEPs were obtained at laser intensities depending on375
the perceptive threshold which varied as a function376
of anodal and cathodal stimulation This means that 377
the cerebellar stimulation has not a selective analgesic 378
effect as it influences both non nociceptive and noci- 379
ceptive perception A pre-eminent analgesic cannot be 380
assessed because the nociceptive threshold was not 381
evaluated 382
As tcDCS was effective on the modulation of 383
both N1 and N2P2 components and these responses 384
are generated by parallel and partially segregated 385
spinal pathways reaching different cortical targets 386
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 9
(Valeriani et al 2007) we may suggest that the cere-387
bellum is engaged in pain processing by modulating388
the activity of both somatosensory and cingulate cor-389
tices Indeed the cerebellum is involved in both the390
sensory-discriminative and emotional dimension of391
pain (Singer et al 2004 Moriguchi et al 2007) and392
non-invasive cerebellar current stimulation may modu-393
late pain experience and the associated cortical activity394
through many not alternative mechanisms In partic-395
ular changes in N1 reflects the modulation of the396
sensory component of pain while the vertex N2P2397
represents the neural correlate of affective aspects of398
pain experience (Garcia-Larrea et al 1997 Valeriani399
et al 2007) Notably tcDCS may act not only on spinal400
nociceptive neurons but also on wide-range cortical401
networks of the pain matrix (Singer et al 2004) thus402
influencing LEPs and pain experience through both403
top-down and bottom-up mechanisms404
The present study does not allow to hypothesize405
how and where tcDCS influences the cerebellar activ-406
ity A main role of Purkinje cells has been suggested407
as their activity modulation may affect the cerebellar408
inhibitory control of the cerebral cortex (Galea et al409
2009) This would be in line with the effects elicited410
by tDCS in the cerebral cortex which are observ-411
able after both short and long term delay likely also412
interfering with long-term potentiation (LTP)-like phe-413
nomena (Hamada et al 2012 Priori et al 2014)414
Moreover prolonged spiking activity in the cerebellar415
Golgi inhibitory neurons modulates the activity of the416
Purkinje cells and could partly account for the tcDCS417
after-effects (Hull et al 2013)418
The lack of changes in RMT indicates that the anal-419
gesic effects of anodal tcDCS are due to a specific420
modulation of the cerebellar activity and not to motor421
activation On the other hand tcDCS-induced cerebel-422
lar modulation (Purpura amp McMurtry 1965) could be423
not sufficient per se to activate the cerebello ndash thalamo424
- cortical motor pathway (Galea et al 2009) thus425
the reported analgesia and its cortical correlates can-426
not be sustained by the motor cortex activation This427
view is supported by the absence of any association428
between motor symptoms and pain perception in cere-429
bellar patients (Ruscheweyh et al 2014) In the same430
line in healthy subjects it has been recently shown431
that motor task-induced increased cortical excitability432
and analgesia are not associated (Volz et al 2012)433
Indeed RMT is a highly sensitive marker of motor434
tract excitability as it reflects activation of a small435
low-threshold and slow-conducting core of pyramidal436
neurons (Hess et al 1987 Rossini amp Rossi 2007) 437
although RMT may reflect changes in the activity of 438
different central nervous system structures it has been 439
satisfactorily used to assess motor cortex excitability 440
also in cerebellar patients (Battaglia et al 2006) 441
Another critical point is the possibility to modulate 442
with tcDCS both neural correlates underlying nocicep- 443
tive processing and pain perception Previous studies 444
using tDCS over motor cortex were inconsistent among 445
each other some works suggested that tDCS is able to 446
modify pain perception (Boggio et al 2008) while 447
others showed divergent effects on psychophysical 448
and neurophysiological outcome parameters (Luedtke 449
et al 2012 Ihle et al 2014) likely due to a possi- 450
ble overestimation of the role of motor areas on pain 451
processing (Antal et al 2008) 452
Our findings cannot be compared to the results 453
obtained by other Authors In fact the unique study 454
focused on the analgesic effects of non-invasive cere- 455
bellar stimulation reported till now (Zunhammer et al 456
2011) considered only subjective pain thresholds 457
In addition it described similar analgesic effects of 458
cerebellar and neck structures repetitive transcranial 459
magnetic stimulation (rTMS) thus denying any cere- 460
bellar specificity in the observed effects and suggesting 461
that the peripheral information passing through the 462
cerebellum may be responsible for analgesia The 463
main difference between the two studies possibly 464
accounting for different results consists of the neu- 465
romodulation techniques used 466
41 Limitations of the study 467
The present study has a few limitations First our 468
findings do not allow any hypothesis on the role of 469
the cerebellum in chronic pain The observations on 470
patients with cerebellar damage (Ruscheweyh et al 471
2014) suggest that their impaired inhibitory control 472
mechanisms may be not associated with the devel- 473
opment of chronic pain Second we cannot exclude 474
the possibility that tcDCS could modulate not only the 475
cerebellum but also surrounding areas such as the peri- 476
aqueductal gray However recent modelling researches 477
have revealed that during tcDCS the current spread 478
to other structures outside the cerebellum is negligi- 479
ble (Parazzini et al 2013 2014) Moreover several 480
studies have proved that in humans pain processing 481
is encoded within posterior areas of each cerebellar 482
hemisphere specifically in the hemispheric lobule VI 483
Crus I and VIIb (Moulton et al 2011) where the 484
Unc
orre
cted
Aut
hor P
roof
10 T Bocci et al Cerebellum and pain A tcDCS study
tcDCS-induced electrical field is strongly concentrated485
(Parazzini et al 2013) A further limitation is that we486
did not study the contribution of C-fibers the main487
component of spino-ponto-cerebellar and spino-olivo-488
cerebellar pathways In fact ultra-late LEPs related489
to C-fibers activations have not yet been standard-490
ized for clinical application and their occurrence could491
be markedly influenced by high order cognitive pro-492
cesses as they seem to be more affected by the level493
of consciousness and attention than A-delta responses494
(Qiu et al 2002 Opsommer et al 2003 Mouraux amp495
Plaghki 2006) Finally we wish to emphasize that in496
neuropathic patients the effects of the cerebellar stim-497
ulation could be quite different from those described498
here as both anatomical and functional connectivity499
are different from those observed in healthy partici-500
pants (Rocca et al 2010 Riedl et al 2011 Absinta et501
al 2012 Longo et al 2012 Ceko et al 2013)502
5 Conclusions503
Our findings indicate a cerebellar effect on pain504
experience and on its cortical correlates and prompt505
further investigation aimed at assessing whether the506
cerebellar direct current polarization could be used as a507
novel and safe therapeutic tool in chronic pain patients508
Acknowledgments509
We gratefully acknowledge the participation of all510
subjects as well as Mr D Barloscio and Mr C511
Orsini for their excellent technical assistance The512
paper was supported in part by the Italian operating513
and development MIUR PRIN grant year 2006 n514
2006062332 002515
RF and AP are founders and shareholders of516
Newronika srl Milan Italy517
References518
Absinta M Rocca MA Colombo B Falini A Comi G amp Fil-519
ippi M (2012) Selective decreased grey matter volume of the520
pain-matrix network in cluster headache Cephalalgia 32(2)521
109-115522
Agostino R Cruccu G Iannetti G Romaniello A Truini A523
amp Manfredi M (2000) Topographical distribution of pinprick524
and warmth thresholds to CO2 laser stimulation on the human525
skin Neurosci Lett 285(2) 115-118526
Antal A Brepohl N Poreisz C Boros K Csifcsak G amp 527
Paulus W (2008) Transcranial direct current stimulation over 528
somatosensory cortex decreases experimentally induced acute 529
pain perception Clin J Pain 24(1) 56-63 530
Antal A amp Paulus W (2010) Effects of transcranial theta-burst 531
stimulation on acute pain perception Restor Neurol Neurosci 532
28(4) 477-484 533
Apkarian AV Bushnell MC Treede RD amp Zubieta JK 534
(2005) Human brain mechanisms of pain perception and reg- 535
ulation in health and disease Eur J Pain 9(4) 463-484 536
Balsters JH Whelan CD Robertson IH amp Ramnani N (2013) 537
Cerebellum and cognition Evidence for the encoding of higher 538
order rules Cereb Cortex 23(6) 1433-1443 539
Battaglia F Quartarone A Ghilardi MF Dattola R Bagnato 540
S Rizzo V Morgante L amp Girlanda P (2006) Unilat- 541
eral cerebellar stroke disrupts movement preparation and motor 542
imagery Clin Neurophysiol 117(5) 1009-1016 543
Bingel U Quante M Knab R Bromm B Weiller C amp Buchel 544
C (2002) Subcortical structures involved in pain processing 545
Evidence from single-trial fMRI Pain 99(1-2) 313-321 546
Boggio PS Zaghi S amp Fregni F (2009) Modulation of emotions 547
associated with images of human pain using anodal transcranial 548
direct current stimulation (tDCS) Neuropsychologia 47(1) 549
212-217 550
Boggio PS Zaghi S Lopes M amp Fregni F (2008) Modulatory 551
effects of anodal transcranial direct current stimulation on per- 552
ception and pain thresholds in healthy volunteers Eur J Neurol 553
15(10) 1124-1130 554
Borsook D Moulton EA Tully S Schmahmann JD amp 555
Becerra L (2008) Human cerebellar responses to brush and 556
heat stimuli in healthy and neuropathic pain subjects Cerebel- 557
lum 7(3) 252-272 558
Ceko M Seminowicz DA Bushnell MC amp Olausson HW 559
(2013) Anatomical and functional enhancements of the insula 560
after loss of large primary somatosensory fibers Cereb Cortex 561
23(9) 2017-2024 562
Cruccu G Aminoff MJ Curio G Guerit JM Kakigi 563
R Mauguiere F Rossini PM Treede RD amp Garcia- 564
Larrea L (2008) Recommendations for the clinical use of 565
somatosensory-evoked potentials Clin Neurophysiol 119(8) 566
1705-1719 567
Cruccu G Romaniello A Amantini A Lombardi M Innocenti 568
P amp Manfredi M (1999) Assessment of trigeminal small- 569
fiber function Brain and reflex responses evoked by CO2-laser 570
stimulation Muscle Nerve 22(4) 508-516 571
Dasilva AF Mendonca ME Zaghi S Lopes M Dossantos 572
MF Spierings EL Bajwa Z Datta A Bikson M amp 573
Fregni F (2012) tDCS-induced analgesia and electrical fields 574
in pain-related neural networks in chronic migraine Headache 575
52(8) 1283-1295 576
Duerden EG amp Albanese MC (2013) Localization of pain- 577
related brain activation A meta-analysis of neuroimaging data 578
Hum Brain Mapp 34(1) 109-149 579
Ekerot CF Garwicz M amp Schouenborg J (1991a) The postsy- 580
naptic dorsal column pathway mediates cutaneous nociceptive 581
information to cerebellar climbing fibres in the cat J Physiol 582
441 275-284 583
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 11
Ekerot CF Garwicz M amp Schouenborg J (1991b) Topography584
and nociceptive receptive fields of climbing fibres projecting to585
the cerebellar anterior lobe in the cat J Physiol 441 257-274586
Ekerot CF Gustavsson P Oscarsson O amp Schouenborg J587
(1987a) Climbing fibres projecting to cat cerebellar anterior588
lobe activated by cutaneous A and C fibres J Physiol 386589
529-538590
Ekerot CF Oscarsson O amp Schouenborg J (1987b) Stimula-591
tion of cat cutaneous nociceptive C fibres causing tonic and592
synchronous activity in climbing fibres J Physiol 386 539-593
546594
Ferrucci R Brunoni AR Parazzini M Vergari M Rossi E595
Fumagalli M Mameli F Rosa M Giannicola G Zago S596
amp Priori A (2013) Modulating human procedural learning by597
cerebellar transcranial direct current stimulation Cerebellum598
12(4) 485-492599
Ferrucci R Giannicola G Rosa M Fumagalli M Boggio600
PS Hallett M Zago S amp Priori A (2012) Cerebellum601
and processing of negative facial emotions Cerebellar tran-602
scranial DC stimulation specifically enhances the emotional603
recognition of facial anger and sadness Cogn Emot 26(5)604
786-799605
Ferrucci R Marceglia S Vergari M Cogiamanian F Mrakic-606
Sposta S Mameli F Zago S Barbieri S amp Priori607
A (2008) Cerebellar transcranial direct current stimulation608
impairs the practice-dependent proficiency increase in working609
memory J Cogn Neurosci 20(9) 1687-1697610
Fregni F Freedman S amp Pascual-Leone A (2007) Recent611
advances in the treatment of chronic pain with non-invasive612
brain stimulation techniques Lancet Neurol 6(2) 188-191613
Galea JM Jayaram G Ajagbe L amp Celnik P (2009)614
Modulation of cerebellar excitability by polarity-specific615
noninvasive direct current stimulation J Neurosci 29(28)616
9115-9122617
Galea JM Vazquez A Pasricha N de Xivry JJ amp Celnik618
P (2011) Dissociating the roles of the cerebellum and motor619
cortex during adaptive learning The motor cortex retains what620
the cerebellum learns Cereb Cortex 21(8) 1761-1770621
Gandiga PC Hummel FC amp Cohen LG (2006) Tran-622
scranial DC stimulation (tDCS) A tool for double-blind623
sham-controlled clinical studies in brain stimulation Clin Neu-624
rophysiol 117(4) 845-850625
Garcia-Larrea L Peyron R Laurent B amp Mauguiere F (1997)626
Association and dissociation between laser-evoked potentials627
and pain perception Neuroreport 8(17) 3785-3789628
Grimaldi G Argyropoulos GP Boehringer A Celnik P629
Edwards MJ Ferrucci R Galea JM Groiss SJ Hiraoka630
K Kassavetis P Lesage E Manto M Miall RC Priori A631
Sadnicka A Ugawa Y amp Ziemann U (2014) Non-invasive632
cerebellar stimulationndasha consensus paper Cerebellum 13(1)633
121-138634
Groppa S Oliviero A Eisen A Quartarone A Cohen LG635
Mall V Kaelin-Lang A Mima T Rossi S Thickbroom636
GW Rossini PM Ziemann U Valls-Sole J amp Sieb-637
ner HR (2012) A practical guide to diagnostic transcranial638
magnetic stimulation Report of an IFCN committee Clin Neu-639
rophysiol 123(5) 858-882640
Hamada M Strigaro G Murase N Sadnicka A Galea JM 641
Edwards MJ amp Rothwell JC (2012) Cerebellar modulation 642
of human associative plasticity J Physiol 590(Pt 10) 2365- 643
2374 644
Hess CW Mills KR amp Murray NM (1987) Responses in small 645
hand muscles from magnetic stimulation of the human brain J 646
Physiol 388 397-419 647
Hull CA Chu Y Thanawala M amp Regehr WG (2013) Hyper- 648
polarization induces a long-term increase in the spontaneous 649
firing rate of cerebellar Golgi cells J Neurosci 33(14) 5895- 650
5902 651
Ihle K Rodriguez-Raecke R amp Luedtke K May A (2014) 652
tDCS modulates cortical nociceptive processing but has little 653
to no impact on pain perception Pain 155(10) 2080-2087 654
Ito M (2006) Cerebellar circuitry as a neuronal machine Prog 655
Neurobiol 78(3-5) 272-303 656
Iyer MB Mattu U Grafman J Lomarev M Sato S amp Wasser- 657
mann EM (2005) Safety and cognitive effect of frontal DC 658
brain polarization in healthy individuals Neurology 64(5) 872- 659
875 660
Kelly RM amp Strick PL (2003) Cerebellar loops with motor cor- 661
tex and prefrontal cortex of a nonhuman primate J Neurosci 662
23(23) 8432-8444 663
Lefaucheur JP Andre-Obadia N Antal A Ayache SS Baeken 664
C Benninger DH Cantello RM Cincotta M de Carvalho 665
M De Ridder D Devanne H Di Lazzaro V Filipovic SR 666
Hummel FC Jaaskelainen SK Kimiskidis VK Koch G 667
Langguth B Nyffeler T Oliviero A Padberg F Poulet E 668
Rossi S Rossini PM Rothwell JC Schonfeldt-Lecuona 669
C Siebner HR Slotema CW Stagg CJ Valls-Sole J 670
Ziemann U Paulus W amp Garcia-Larrea L (2014) Evidence- 671
based guidelines on the therapeutic use of repetitive transcranial 672
magnetic stimulation (rTMS) Clin Neurophysiol 125(11) 673
2150-2206 674
Lefaucheur JP Brusa A Creange A Drouot X amp Jarry G 675
(2002) Clinical application of laser evoked potentials using 676
the NdYAG laser Neurophysiol Clin 32(2) 91-98 677
Lefaucheur JP Debray S amp Jarry G (2001) Laser evoked poten- 678
tials using the NdYAG laser Muscle Nerve 24(4) 496-501 679
Lefaucheur JP Drouot X Menard-Lefaucheur I Keravel Y 680
amp Nguyen JP (2006) Motor cortex rTMS restores defective 681
intracortical inhibition in chronic neuropathic pain Neurology 682
67(9) 1568-1574 683
Longo MR Iannetti GD Mancini F Driver J amp Haggard P 684
(2012) Linking pain and the body Neural correlates of visually 685
induced analgesia J Neurosci 32(8) 2601-2607 686
Lorenz J amp Garcia-Larrea L (2003) Contribution of attentional 687
and cognitive factors to laser evoked brain potentials Neuro- 688
physiol Clin 33(6) 293-301 689
Lorenz J Minoshima S amp Casey KL (2003) Keeping pain out 690
of mind The role of the dorsolateral prefrontal cortex in pain 691
modulation Brain 126(Pt 5) 1079-1091 692
Luedtke K May A amp Jurgens TP (2012) No effect of a single 693
session of transcranial direct current stimulation on experimen- 694
tally induced pain in patients with chronic low back painndashan 695
exploratory study PLoS One 7(11) e48857 696
Unc
orre
cted
Aut
hor P
roof
12 T Bocci et al Cerebellum and pain A tcDCS study
Mendonca ME Santana MB Baptista AF Datta A Bikson697
M Fregni F amp Araujo CP (2011) Transcranial DC stimu-698
lation in fibromyalgia Optimized cortical target supported by699
high-resolution computational models J Pain 12(5) 610-617700
Moriguchi Y Decety J Ohnishi T Maeda M Mori T Nemoto701
K Matsuda H amp Komaki G (2007) Empathy and judg-702
ing otherrsquos pain An fMRI study of alexithymia Cereb Cortex703
17(9) 2223-2234704
Moulton EA Elman I Pendse G Schmahmann J Becerra L705
amp Borsook D (2011) Aversion-related circuitry in the cere-706
bellum Responses to noxious heat and unpleasant images J707
Neurosci 31(10) 3795-3804708
Moulton EA Schmahmann JD Becerra L amp Borsook D709
(2010) The cerebellum and pain Passive integrator or active710
participator Brain Res Rev 65(1) 14-27711
Mouraux A amp Plaghki L (2006) Are the processes reflected by712
late and ultra-late laser evoked potentials specific of nocicep-713
tion Suppl Clin Neurophysiol 59 197-204714
Mylius V Jung M Menzler K Haag A Khader PH Oer-715
tel WH Rosenow F amp Lefaucheur JP (2012) Effects of716
transcranial direct current stimulation on pain perception and717
working memory Eur J Pain 16(7) 974-982718
Ni Z Gunraj C amp Chen R (2007) Short interval intracortical719
inhibition and facilitation during the silent period in human J720
Physiol 583(Pt 3) 971-982721
Nitsche MA Liebetanz D Lang N Antal A Tergau F amp722
Paulus W (2003) Safety criteria for transcranial direct cur-723
rent stimulation (tDCS) in humans Clin Neurophysiol 114(11)724
2220-2222 author reply 2222-2223725
Opsommer E Guerit JM amp Plaghki L (2003) Exogenous726
and endogenous components of ultralate (C-fibre) evoked727
potentials following CO2 laser stimuli to tiny skin sur-728
face areas in healthy subjects Neurophysiol Clin 33(2)729
78-85730
Parazzini M Fiocchi S Liorni I Priori A amp Ravazzani P731
(2014) Computational modeling of transcranial direct current732
stimulation in the child brain Implications for the treatment of733
refractory childhood focal epilepsy Int J Neural Syst 24(2)734
1430006735
Parazzini M Rossi E Ferrucci R Fiocchi S Liorni I Priori736
A amp Ravazzani P (2013) Computational model of cerebellar737
transcranial direct current stimulation Conf Proc IEEE Eng738
Med Biol Soc 2013 237-240739
Parazzini M Rossi E Ferrucci R Liorni I Priori A amp Ravaz-740
zani P (2014) Modelling the electric field and the current741
density generated by cerebellar transcranial DC stimulation in742
humans Clin Neurophysiol 125(3) 577-584743
Perini I Bergstrand S amp Morrison I (2013) Where pain meets744
action in the human brain J Neurosci 33(40) 15930-15939745
Ploghaus A Tracey I Gati JS Clare S Menon RS Matthews746
PM amp Rawlins JN (1999) Dissociating pain from its antic-747
ipation in the human brain Science 284(5422) 1979-1981748
Priori A Ciocca M Parazzini M Vergari M amp Ferrucci R749
(2014) Transcranial cerebellar direct current stimulation and750
transcutaneous spinal cord direct current stimulation as innova-751
tive tools for neuroscientists J Physiol 592(Pt 16) 3345-3369752
Purpura DP amp McMurtry JC (1965) Intracellular activities and 753
evoked potential changes during polarization of motor cortex 754
J Neurophysiol 28 166-185 755
Qiu Y Inui K Wang X Tran TD amp Kakigi R (2002) Effects 756
of attention distraction and sleep on CO(2) laser evoked poten- 757
tials related to C-fibers in humans Clin Neurophysiol 113(10) 758
1579-1585 759
Reidler JS Mendonca ME Santana MB Wang X Lenkinski 760
R Motta AF Marchand S Latif L amp Fregni F (2012) 761
Effects of motor cortex modulation and descending inhibitory 762
systems on pain thresholds in healthy subjects J Pain 13(5) 763
450-458 764
Riedl V Valet M Woller A Sorg C Vogel D Sprenger 765
T Boecker H Wohlschlager AM amp Tolle TR (2011) 766
Repeated pain induces adaptations of intrinsic brain activity 767
to reflect past and predict future pain Neuroimage 57(1) 206- 768
213 769
Rocca MA Valsasina P Absinta M Colombo B Barcella V 770
Falini A Comi G amp Filippi M (2010) Central nervous 771
system dysregulation extends beyond the pain-matrix network 772
in cluster headache Cephalalgia 30(11) 1383-1391 773
Rossi S Hallett M Rossini PM amp Pascual-Leone A (2009) 774
Safety ethical considerations and application guidelines for 775
the use of transcranial magnetic stimulation in clinical practice 776
and research Clin Neurophysiol 120(12) 2008-2039 777
Rossini PM amp Rossi S (2007) Transcranial magnetic stimulation 778
Diagnostic therapeutic and research potential Neurology 779
68(7) 484-488 780
Ruscheweyh R Kuhnel M Filippopulos F Blum B Eggert 781
T amp Straube A (2014) Altered experimental pain perception 782
after cerebellar infarction Pain 155(7) 1303-1312 783
Schmahmann JD (1991) An emerging concept The cerebellar 784
contribution to higher function Arch Neurol 48(11) 1178- 785
1187 786
Singer T Seymour B OrsquoDoherty J Kaube H Dolan RJ amp 787
Frith CD (2004) Empathy for pain involves the affective but 788
not sensory components of pain Science 303(5661) 1157- 789
1162 790
Smith MJ Keel JC Greenberg BD Adams LF Schmidt PJ 791
Rubinow DA amp Wassermann EM (1999) Menstrual cycle 792
effects on cortical excitability Neurology 53(9) 2069-2072 793
Stoodley CJ amp Schmahmann JD (2009) Functional topography 794
in the human cerebellum A meta-analysis of neuroimaging 795
studies Neuroimage 44(2) 489-501 796
Strick PL Dum RP amp Fiez JA (2009) Cerebellum and non- 797
motor function Annu Rev Neurosci 32 413-434 798
Strigo IA Duncan GH Boivin M amp Bushnell MC (2003) 799
Differentiation of visceral and cutaneous pain in the human 800
brain J Neurophysiol 89(6) 3294-3303 801
Truini A Galeotti F Romaniello A Virtuoso M Iannetti GD 802
amp Cruccu G (2005) Laser-evoked potentials Normative val- 803
ues Clin Neurophysiol 116(4) 821-826 804
Truini A Panuccio G Galeotti F Maluccio MR Sartucci F 805
Avoli M amp Cruccu G (2010) Laser-evoked potentials as a 806
tool for assessing the efficacy of antinociceptive drugs Eur J 807
Pain 14(2) 222-225 808
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 13
Truini A Vergari M Biasiotta A La Cesa S Gabriele M Di809
Stefano G Cambieri C Cruccu G Inghilleri M amp Pri-810
ori A (2011) Transcutaneous spinal direct current stimulation811
inhibits nociceptive spinal pathway conduction and increases812
pain tolerance in humans Eur J Pain 15(10) 1023-1027813
Valeriani M Le Pera D Restuccia D de Armas L Mili-814
ucci R Betti V Vigevano F amp Tonali P (2007) Parallel815
spinal pathways generate the middle-latency N1 and the late P2816
components of the laser evoked potentials Clin Neurophysiol817
118(5) 1097-1104818
Valeriani M Pazzaglia C Cruccu G amp Truini A (2012) Clinical819
usefulness of laser evoked potentials Neurophysiol Clin 42(5)820
345-353821
Volz MS Mendonca M Pinheiro FS Cui H Santana M amp822
Fregni F (2012) Dissociation of motor task-induced cortical823
excitability and pain perception changes in healthy volunteers824
PLoS One 7(3) e34273825
Wager TD Rilling JK Smith EE Sokolik A Casey KL 826
Davidson RJ Kosslyn SM Rose RM amp Cohen JD 827
(2004) Placebo-induced changes in FMRI in the anticipation 828
and experience of pain Science 303(5661) 1162-1167 829
Zaghi S Thiele B Pimentel D Pimentel T amp Fregni 830
F (2011) Assessment and treatment of pain with non- 831
invasive cortical stimulation Restor Neurol Neurosci 29(6) 832
439-451 833
Zubieta JK Bueller JA Jackson LR Scott DJ Xu Y 834
Koeppe RA Nichols TE amp Stohler CS (2005) Placebo 835
effects mediated by endogenous opioid activity on mu-opioid 836
receptors J Neurosci 25(34) 7754-7762 837
Zunhammer M Busch V Griesbach F Landgrebe M Hajak G 838
amp Langguth B (2011) rTMS over the cerebellum modulates 839
temperature detection and pain thresholds through peripheral 840
mechanisms Brain Stimul 4(4) 210-7 e1 841

Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 7
A
B
Fig 3 - A Perceptive Threshold Changes (mean plusmn SD) at T1 and T2 with respect to baseline values (T1T0 T2T0) following sham (black)anodal (white) and cathodal (grey) tcDCS (lowastlowastp lt 0001 lowastlowastlowastp lt 00001) B Changes in visual analogue scale (VAS) scores over time VAS scoresat two different stimulus intensity respectively two (A left) and three (B right) times higher than the PT (lowastlowastp lt 0001 lowastlowastlowastp lt 00001)
A B
Fig 4 ndash A LEPs grand averaging traces were recorded at baseline (T0 black) and immediately after cerebellar polarization (T1 red) due tosham (left column) anodal (middle) and cathodal (right) tcDCS B Histograms showing LEPs variables and VAS scores changes (mean plusmn SD)after sham (black) anodal (white) or cathodal (grey) tcDCS with respect to baseline Top panels changes in N1 variables (amplitude and latency)over time bottom panels changes in N2P2 complex (lowastlowastp lt 0001 lowastlowastlowastp lt 00001)
Unc
orre
cted
Aut
hor P
roof
8 T Bocci et al Cerebellum and pain A tcDCS study
Table 3
LEPs post-hoc analyses p lt 00001 for all comparison except when explicitly indicated lowastlowastp lt 0002 lowast p lt 0005
N1 amplitude latency N2P2 amplitude latency
anodal cathodal shamdf
time 228 F = 67152 F = 96489 F = 134912 F = 34946 nsT0 vs T1 114 F = 109178 F = 18815 F = 165953 F = 64281T0 vs T2 114 F = 75143 F = 167697 F = 145125 F = 37818T1 vs T2 114 ns ns ns ns
anodal cathodaltime 228 F = 102281 F = 98717 F = 6577 F = 20918 nsT0 vs T1 114 F = 511186 F = 104027 F = 144112 F = 103864T0 vs T2 114 F = 96329 F = 116841 F = 105183 F = 14012lowastlowastT1 vs T2 114 ns ns ns ns ns
anodal vs sham anodal vs shamT0 114 ns ns ns nsT1 114 ns t = 925 t = 601 6262T2 114 ns t = 8128 t = 6731 5236
cathodal cathodal vs shamT0 114 ns ns t = 3281lowast nsT1 114 t = 16594 t = 8029 t = 8262 t = 520T2 114 t = 7309 t = 12669 t = 5048 t = 5301
Fig 5 - Resting Motor Thresholds Changes (mean plusmn SD) in Resting Motor threshold (RMT) expressed as percentage of the maximumstimulator output after sham (black) anodal (white) and cathodal (grey) tcDCS with respect to baseline marked as dotted line (lowastlowastp lt 0001lowastlowastlowastp lt 00001)
reduction of the inhibitory tone exerted by the cere-367
bellum on brain targets Anodal polarization elicits368
opposite effects producing analgesia Both findings369
support the role of the cerebellum in pain control370
it is noticeable that cathodal cerebellar stimulation371
induces hyperalgesia as occurs in patients with cere-372
bellar infarction (Ruscheweyh et al 2014)373
We would like to underline that in the present study374
LEPs were obtained at laser intensities depending on375
the perceptive threshold which varied as a function376
of anodal and cathodal stimulation This means that 377
the cerebellar stimulation has not a selective analgesic 378
effect as it influences both non nociceptive and noci- 379
ceptive perception A pre-eminent analgesic cannot be 380
assessed because the nociceptive threshold was not 381
evaluated 382
As tcDCS was effective on the modulation of 383
both N1 and N2P2 components and these responses 384
are generated by parallel and partially segregated 385
spinal pathways reaching different cortical targets 386
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 9
(Valeriani et al 2007) we may suggest that the cere-387
bellum is engaged in pain processing by modulating388
the activity of both somatosensory and cingulate cor-389
tices Indeed the cerebellum is involved in both the390
sensory-discriminative and emotional dimension of391
pain (Singer et al 2004 Moriguchi et al 2007) and392
non-invasive cerebellar current stimulation may modu-393
late pain experience and the associated cortical activity394
through many not alternative mechanisms In partic-395
ular changes in N1 reflects the modulation of the396
sensory component of pain while the vertex N2P2397
represents the neural correlate of affective aspects of398
pain experience (Garcia-Larrea et al 1997 Valeriani399
et al 2007) Notably tcDCS may act not only on spinal400
nociceptive neurons but also on wide-range cortical401
networks of the pain matrix (Singer et al 2004) thus402
influencing LEPs and pain experience through both403
top-down and bottom-up mechanisms404
The present study does not allow to hypothesize405
how and where tcDCS influences the cerebellar activ-406
ity A main role of Purkinje cells has been suggested407
as their activity modulation may affect the cerebellar408
inhibitory control of the cerebral cortex (Galea et al409
2009) This would be in line with the effects elicited410
by tDCS in the cerebral cortex which are observ-411
able after both short and long term delay likely also412
interfering with long-term potentiation (LTP)-like phe-413
nomena (Hamada et al 2012 Priori et al 2014)414
Moreover prolonged spiking activity in the cerebellar415
Golgi inhibitory neurons modulates the activity of the416
Purkinje cells and could partly account for the tcDCS417
after-effects (Hull et al 2013)418
The lack of changes in RMT indicates that the anal-419
gesic effects of anodal tcDCS are due to a specific420
modulation of the cerebellar activity and not to motor421
activation On the other hand tcDCS-induced cerebel-422
lar modulation (Purpura amp McMurtry 1965) could be423
not sufficient per se to activate the cerebello ndash thalamo424
- cortical motor pathway (Galea et al 2009) thus425
the reported analgesia and its cortical correlates can-426
not be sustained by the motor cortex activation This427
view is supported by the absence of any association428
between motor symptoms and pain perception in cere-429
bellar patients (Ruscheweyh et al 2014) In the same430
line in healthy subjects it has been recently shown431
that motor task-induced increased cortical excitability432
and analgesia are not associated (Volz et al 2012)433
Indeed RMT is a highly sensitive marker of motor434
tract excitability as it reflects activation of a small435
low-threshold and slow-conducting core of pyramidal436
neurons (Hess et al 1987 Rossini amp Rossi 2007) 437
although RMT may reflect changes in the activity of 438
different central nervous system structures it has been 439
satisfactorily used to assess motor cortex excitability 440
also in cerebellar patients (Battaglia et al 2006) 441
Another critical point is the possibility to modulate 442
with tcDCS both neural correlates underlying nocicep- 443
tive processing and pain perception Previous studies 444
using tDCS over motor cortex were inconsistent among 445
each other some works suggested that tDCS is able to 446
modify pain perception (Boggio et al 2008) while 447
others showed divergent effects on psychophysical 448
and neurophysiological outcome parameters (Luedtke 449
et al 2012 Ihle et al 2014) likely due to a possi- 450
ble overestimation of the role of motor areas on pain 451
processing (Antal et al 2008) 452
Our findings cannot be compared to the results 453
obtained by other Authors In fact the unique study 454
focused on the analgesic effects of non-invasive cere- 455
bellar stimulation reported till now (Zunhammer et al 456
2011) considered only subjective pain thresholds 457
In addition it described similar analgesic effects of 458
cerebellar and neck structures repetitive transcranial 459
magnetic stimulation (rTMS) thus denying any cere- 460
bellar specificity in the observed effects and suggesting 461
that the peripheral information passing through the 462
cerebellum may be responsible for analgesia The 463
main difference between the two studies possibly 464
accounting for different results consists of the neu- 465
romodulation techniques used 466
41 Limitations of the study 467
The present study has a few limitations First our 468
findings do not allow any hypothesis on the role of 469
the cerebellum in chronic pain The observations on 470
patients with cerebellar damage (Ruscheweyh et al 471
2014) suggest that their impaired inhibitory control 472
mechanisms may be not associated with the devel- 473
opment of chronic pain Second we cannot exclude 474
the possibility that tcDCS could modulate not only the 475
cerebellum but also surrounding areas such as the peri- 476
aqueductal gray However recent modelling researches 477
have revealed that during tcDCS the current spread 478
to other structures outside the cerebellum is negligi- 479
ble (Parazzini et al 2013 2014) Moreover several 480
studies have proved that in humans pain processing 481
is encoded within posterior areas of each cerebellar 482
hemisphere specifically in the hemispheric lobule VI 483
Crus I and VIIb (Moulton et al 2011) where the 484
Unc
orre
cted
Aut
hor P
roof
10 T Bocci et al Cerebellum and pain A tcDCS study
tcDCS-induced electrical field is strongly concentrated485
(Parazzini et al 2013) A further limitation is that we486
did not study the contribution of C-fibers the main487
component of spino-ponto-cerebellar and spino-olivo-488
cerebellar pathways In fact ultra-late LEPs related489
to C-fibers activations have not yet been standard-490
ized for clinical application and their occurrence could491
be markedly influenced by high order cognitive pro-492
cesses as they seem to be more affected by the level493
of consciousness and attention than A-delta responses494
(Qiu et al 2002 Opsommer et al 2003 Mouraux amp495
Plaghki 2006) Finally we wish to emphasize that in496
neuropathic patients the effects of the cerebellar stim-497
ulation could be quite different from those described498
here as both anatomical and functional connectivity499
are different from those observed in healthy partici-500
pants (Rocca et al 2010 Riedl et al 2011 Absinta et501
al 2012 Longo et al 2012 Ceko et al 2013)502
5 Conclusions503
Our findings indicate a cerebellar effect on pain504
experience and on its cortical correlates and prompt505
further investigation aimed at assessing whether the506
cerebellar direct current polarization could be used as a507
novel and safe therapeutic tool in chronic pain patients508
Acknowledgments509
We gratefully acknowledge the participation of all510
subjects as well as Mr D Barloscio and Mr C511
Orsini for their excellent technical assistance The512
paper was supported in part by the Italian operating513
and development MIUR PRIN grant year 2006 n514
2006062332 002515
RF and AP are founders and shareholders of516
Newronika srl Milan Italy517
References518
Absinta M Rocca MA Colombo B Falini A Comi G amp Fil-519
ippi M (2012) Selective decreased grey matter volume of the520
pain-matrix network in cluster headache Cephalalgia 32(2)521
109-115522
Agostino R Cruccu G Iannetti G Romaniello A Truini A523
amp Manfredi M (2000) Topographical distribution of pinprick524
and warmth thresholds to CO2 laser stimulation on the human525
skin Neurosci Lett 285(2) 115-118526
Antal A Brepohl N Poreisz C Boros K Csifcsak G amp 527
Paulus W (2008) Transcranial direct current stimulation over 528
somatosensory cortex decreases experimentally induced acute 529
pain perception Clin J Pain 24(1) 56-63 530
Antal A amp Paulus W (2010) Effects of transcranial theta-burst 531
stimulation on acute pain perception Restor Neurol Neurosci 532
28(4) 477-484 533
Apkarian AV Bushnell MC Treede RD amp Zubieta JK 534
(2005) Human brain mechanisms of pain perception and reg- 535
ulation in health and disease Eur J Pain 9(4) 463-484 536
Balsters JH Whelan CD Robertson IH amp Ramnani N (2013) 537
Cerebellum and cognition Evidence for the encoding of higher 538
order rules Cereb Cortex 23(6) 1433-1443 539
Battaglia F Quartarone A Ghilardi MF Dattola R Bagnato 540
S Rizzo V Morgante L amp Girlanda P (2006) Unilat- 541
eral cerebellar stroke disrupts movement preparation and motor 542
imagery Clin Neurophysiol 117(5) 1009-1016 543
Bingel U Quante M Knab R Bromm B Weiller C amp Buchel 544
C (2002) Subcortical structures involved in pain processing 545
Evidence from single-trial fMRI Pain 99(1-2) 313-321 546
Boggio PS Zaghi S amp Fregni F (2009) Modulation of emotions 547
associated with images of human pain using anodal transcranial 548
direct current stimulation (tDCS) Neuropsychologia 47(1) 549
212-217 550
Boggio PS Zaghi S Lopes M amp Fregni F (2008) Modulatory 551
effects of anodal transcranial direct current stimulation on per- 552
ception and pain thresholds in healthy volunteers Eur J Neurol 553
15(10) 1124-1130 554
Borsook D Moulton EA Tully S Schmahmann JD amp 555
Becerra L (2008) Human cerebellar responses to brush and 556
heat stimuli in healthy and neuropathic pain subjects Cerebel- 557
lum 7(3) 252-272 558
Ceko M Seminowicz DA Bushnell MC amp Olausson HW 559
(2013) Anatomical and functional enhancements of the insula 560
after loss of large primary somatosensory fibers Cereb Cortex 561
23(9) 2017-2024 562
Cruccu G Aminoff MJ Curio G Guerit JM Kakigi 563
R Mauguiere F Rossini PM Treede RD amp Garcia- 564
Larrea L (2008) Recommendations for the clinical use of 565
somatosensory-evoked potentials Clin Neurophysiol 119(8) 566
1705-1719 567
Cruccu G Romaniello A Amantini A Lombardi M Innocenti 568
P amp Manfredi M (1999) Assessment of trigeminal small- 569
fiber function Brain and reflex responses evoked by CO2-laser 570
stimulation Muscle Nerve 22(4) 508-516 571
Dasilva AF Mendonca ME Zaghi S Lopes M Dossantos 572
MF Spierings EL Bajwa Z Datta A Bikson M amp 573
Fregni F (2012) tDCS-induced analgesia and electrical fields 574
in pain-related neural networks in chronic migraine Headache 575
52(8) 1283-1295 576
Duerden EG amp Albanese MC (2013) Localization of pain- 577
related brain activation A meta-analysis of neuroimaging data 578
Hum Brain Mapp 34(1) 109-149 579
Ekerot CF Garwicz M amp Schouenborg J (1991a) The postsy- 580
naptic dorsal column pathway mediates cutaneous nociceptive 581
information to cerebellar climbing fibres in the cat J Physiol 582
441 275-284 583
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 11
Ekerot CF Garwicz M amp Schouenborg J (1991b) Topography584
and nociceptive receptive fields of climbing fibres projecting to585
the cerebellar anterior lobe in the cat J Physiol 441 257-274586
Ekerot CF Gustavsson P Oscarsson O amp Schouenborg J587
(1987a) Climbing fibres projecting to cat cerebellar anterior588
lobe activated by cutaneous A and C fibres J Physiol 386589
529-538590
Ekerot CF Oscarsson O amp Schouenborg J (1987b) Stimula-591
tion of cat cutaneous nociceptive C fibres causing tonic and592
synchronous activity in climbing fibres J Physiol 386 539-593
546594
Ferrucci R Brunoni AR Parazzini M Vergari M Rossi E595
Fumagalli M Mameli F Rosa M Giannicola G Zago S596
amp Priori A (2013) Modulating human procedural learning by597
cerebellar transcranial direct current stimulation Cerebellum598
12(4) 485-492599
Ferrucci R Giannicola G Rosa M Fumagalli M Boggio600
PS Hallett M Zago S amp Priori A (2012) Cerebellum601
and processing of negative facial emotions Cerebellar tran-602
scranial DC stimulation specifically enhances the emotional603
recognition of facial anger and sadness Cogn Emot 26(5)604
786-799605
Ferrucci R Marceglia S Vergari M Cogiamanian F Mrakic-606
Sposta S Mameli F Zago S Barbieri S amp Priori607
A (2008) Cerebellar transcranial direct current stimulation608
impairs the practice-dependent proficiency increase in working609
memory J Cogn Neurosci 20(9) 1687-1697610
Fregni F Freedman S amp Pascual-Leone A (2007) Recent611
advances in the treatment of chronic pain with non-invasive612
brain stimulation techniques Lancet Neurol 6(2) 188-191613
Galea JM Jayaram G Ajagbe L amp Celnik P (2009)614
Modulation of cerebellar excitability by polarity-specific615
noninvasive direct current stimulation J Neurosci 29(28)616
9115-9122617
Galea JM Vazquez A Pasricha N de Xivry JJ amp Celnik618
P (2011) Dissociating the roles of the cerebellum and motor619
cortex during adaptive learning The motor cortex retains what620
the cerebellum learns Cereb Cortex 21(8) 1761-1770621
Gandiga PC Hummel FC amp Cohen LG (2006) Tran-622
scranial DC stimulation (tDCS) A tool for double-blind623
sham-controlled clinical studies in brain stimulation Clin Neu-624
rophysiol 117(4) 845-850625
Garcia-Larrea L Peyron R Laurent B amp Mauguiere F (1997)626
Association and dissociation between laser-evoked potentials627
and pain perception Neuroreport 8(17) 3785-3789628
Grimaldi G Argyropoulos GP Boehringer A Celnik P629
Edwards MJ Ferrucci R Galea JM Groiss SJ Hiraoka630
K Kassavetis P Lesage E Manto M Miall RC Priori A631
Sadnicka A Ugawa Y amp Ziemann U (2014) Non-invasive632
cerebellar stimulationndasha consensus paper Cerebellum 13(1)633
121-138634
Groppa S Oliviero A Eisen A Quartarone A Cohen LG635
Mall V Kaelin-Lang A Mima T Rossi S Thickbroom636
GW Rossini PM Ziemann U Valls-Sole J amp Sieb-637
ner HR (2012) A practical guide to diagnostic transcranial638
magnetic stimulation Report of an IFCN committee Clin Neu-639
rophysiol 123(5) 858-882640
Hamada M Strigaro G Murase N Sadnicka A Galea JM 641
Edwards MJ amp Rothwell JC (2012) Cerebellar modulation 642
of human associative plasticity J Physiol 590(Pt 10) 2365- 643
2374 644
Hess CW Mills KR amp Murray NM (1987) Responses in small 645
hand muscles from magnetic stimulation of the human brain J 646
Physiol 388 397-419 647
Hull CA Chu Y Thanawala M amp Regehr WG (2013) Hyper- 648
polarization induces a long-term increase in the spontaneous 649
firing rate of cerebellar Golgi cells J Neurosci 33(14) 5895- 650
5902 651
Ihle K Rodriguez-Raecke R amp Luedtke K May A (2014) 652
tDCS modulates cortical nociceptive processing but has little 653
to no impact on pain perception Pain 155(10) 2080-2087 654
Ito M (2006) Cerebellar circuitry as a neuronal machine Prog 655
Neurobiol 78(3-5) 272-303 656
Iyer MB Mattu U Grafman J Lomarev M Sato S amp Wasser- 657
mann EM (2005) Safety and cognitive effect of frontal DC 658
brain polarization in healthy individuals Neurology 64(5) 872- 659
875 660
Kelly RM amp Strick PL (2003) Cerebellar loops with motor cor- 661
tex and prefrontal cortex of a nonhuman primate J Neurosci 662
23(23) 8432-8444 663
Lefaucheur JP Andre-Obadia N Antal A Ayache SS Baeken 664
C Benninger DH Cantello RM Cincotta M de Carvalho 665
M De Ridder D Devanne H Di Lazzaro V Filipovic SR 666
Hummel FC Jaaskelainen SK Kimiskidis VK Koch G 667
Langguth B Nyffeler T Oliviero A Padberg F Poulet E 668
Rossi S Rossini PM Rothwell JC Schonfeldt-Lecuona 669
C Siebner HR Slotema CW Stagg CJ Valls-Sole J 670
Ziemann U Paulus W amp Garcia-Larrea L (2014) Evidence- 671
based guidelines on the therapeutic use of repetitive transcranial 672
magnetic stimulation (rTMS) Clin Neurophysiol 125(11) 673
2150-2206 674
Lefaucheur JP Brusa A Creange A Drouot X amp Jarry G 675
(2002) Clinical application of laser evoked potentials using 676
the NdYAG laser Neurophysiol Clin 32(2) 91-98 677
Lefaucheur JP Debray S amp Jarry G (2001) Laser evoked poten- 678
tials using the NdYAG laser Muscle Nerve 24(4) 496-501 679
Lefaucheur JP Drouot X Menard-Lefaucheur I Keravel Y 680
amp Nguyen JP (2006) Motor cortex rTMS restores defective 681
intracortical inhibition in chronic neuropathic pain Neurology 682
67(9) 1568-1574 683
Longo MR Iannetti GD Mancini F Driver J amp Haggard P 684
(2012) Linking pain and the body Neural correlates of visually 685
induced analgesia J Neurosci 32(8) 2601-2607 686
Lorenz J amp Garcia-Larrea L (2003) Contribution of attentional 687
and cognitive factors to laser evoked brain potentials Neuro- 688
physiol Clin 33(6) 293-301 689
Lorenz J Minoshima S amp Casey KL (2003) Keeping pain out 690
of mind The role of the dorsolateral prefrontal cortex in pain 691
modulation Brain 126(Pt 5) 1079-1091 692
Luedtke K May A amp Jurgens TP (2012) No effect of a single 693
session of transcranial direct current stimulation on experimen- 694
tally induced pain in patients with chronic low back painndashan 695
exploratory study PLoS One 7(11) e48857 696
Unc
orre
cted
Aut
hor P
roof
12 T Bocci et al Cerebellum and pain A tcDCS study
Mendonca ME Santana MB Baptista AF Datta A Bikson697
M Fregni F amp Araujo CP (2011) Transcranial DC stimu-698
lation in fibromyalgia Optimized cortical target supported by699
high-resolution computational models J Pain 12(5) 610-617700
Moriguchi Y Decety J Ohnishi T Maeda M Mori T Nemoto701
K Matsuda H amp Komaki G (2007) Empathy and judg-702
ing otherrsquos pain An fMRI study of alexithymia Cereb Cortex703
17(9) 2223-2234704
Moulton EA Elman I Pendse G Schmahmann J Becerra L705
amp Borsook D (2011) Aversion-related circuitry in the cere-706
bellum Responses to noxious heat and unpleasant images J707
Neurosci 31(10) 3795-3804708
Moulton EA Schmahmann JD Becerra L amp Borsook D709
(2010) The cerebellum and pain Passive integrator or active710
participator Brain Res Rev 65(1) 14-27711
Mouraux A amp Plaghki L (2006) Are the processes reflected by712
late and ultra-late laser evoked potentials specific of nocicep-713
tion Suppl Clin Neurophysiol 59 197-204714
Mylius V Jung M Menzler K Haag A Khader PH Oer-715
tel WH Rosenow F amp Lefaucheur JP (2012) Effects of716
transcranial direct current stimulation on pain perception and717
working memory Eur J Pain 16(7) 974-982718
Ni Z Gunraj C amp Chen R (2007) Short interval intracortical719
inhibition and facilitation during the silent period in human J720
Physiol 583(Pt 3) 971-982721
Nitsche MA Liebetanz D Lang N Antal A Tergau F amp722
Paulus W (2003) Safety criteria for transcranial direct cur-723
rent stimulation (tDCS) in humans Clin Neurophysiol 114(11)724
2220-2222 author reply 2222-2223725
Opsommer E Guerit JM amp Plaghki L (2003) Exogenous726
and endogenous components of ultralate (C-fibre) evoked727
potentials following CO2 laser stimuli to tiny skin sur-728
face areas in healthy subjects Neurophysiol Clin 33(2)729
78-85730
Parazzini M Fiocchi S Liorni I Priori A amp Ravazzani P731
(2014) Computational modeling of transcranial direct current732
stimulation in the child brain Implications for the treatment of733
refractory childhood focal epilepsy Int J Neural Syst 24(2)734
1430006735
Parazzini M Rossi E Ferrucci R Fiocchi S Liorni I Priori736
A amp Ravazzani P (2013) Computational model of cerebellar737
transcranial direct current stimulation Conf Proc IEEE Eng738
Med Biol Soc 2013 237-240739
Parazzini M Rossi E Ferrucci R Liorni I Priori A amp Ravaz-740
zani P (2014) Modelling the electric field and the current741
density generated by cerebellar transcranial DC stimulation in742
humans Clin Neurophysiol 125(3) 577-584743
Perini I Bergstrand S amp Morrison I (2013) Where pain meets744
action in the human brain J Neurosci 33(40) 15930-15939745
Ploghaus A Tracey I Gati JS Clare S Menon RS Matthews746
PM amp Rawlins JN (1999) Dissociating pain from its antic-747
ipation in the human brain Science 284(5422) 1979-1981748
Priori A Ciocca M Parazzini M Vergari M amp Ferrucci R749
(2014) Transcranial cerebellar direct current stimulation and750
transcutaneous spinal cord direct current stimulation as innova-751
tive tools for neuroscientists J Physiol 592(Pt 16) 3345-3369752
Purpura DP amp McMurtry JC (1965) Intracellular activities and 753
evoked potential changes during polarization of motor cortex 754
J Neurophysiol 28 166-185 755
Qiu Y Inui K Wang X Tran TD amp Kakigi R (2002) Effects 756
of attention distraction and sleep on CO(2) laser evoked poten- 757
tials related to C-fibers in humans Clin Neurophysiol 113(10) 758
1579-1585 759
Reidler JS Mendonca ME Santana MB Wang X Lenkinski 760
R Motta AF Marchand S Latif L amp Fregni F (2012) 761
Effects of motor cortex modulation and descending inhibitory 762
systems on pain thresholds in healthy subjects J Pain 13(5) 763
450-458 764
Riedl V Valet M Woller A Sorg C Vogel D Sprenger 765
T Boecker H Wohlschlager AM amp Tolle TR (2011) 766
Repeated pain induces adaptations of intrinsic brain activity 767
to reflect past and predict future pain Neuroimage 57(1) 206- 768
213 769
Rocca MA Valsasina P Absinta M Colombo B Barcella V 770
Falini A Comi G amp Filippi M (2010) Central nervous 771
system dysregulation extends beyond the pain-matrix network 772
in cluster headache Cephalalgia 30(11) 1383-1391 773
Rossi S Hallett M Rossini PM amp Pascual-Leone A (2009) 774
Safety ethical considerations and application guidelines for 775
the use of transcranial magnetic stimulation in clinical practice 776
and research Clin Neurophysiol 120(12) 2008-2039 777
Rossini PM amp Rossi S (2007) Transcranial magnetic stimulation 778
Diagnostic therapeutic and research potential Neurology 779
68(7) 484-488 780
Ruscheweyh R Kuhnel M Filippopulos F Blum B Eggert 781
T amp Straube A (2014) Altered experimental pain perception 782
after cerebellar infarction Pain 155(7) 1303-1312 783
Schmahmann JD (1991) An emerging concept The cerebellar 784
contribution to higher function Arch Neurol 48(11) 1178- 785
1187 786
Singer T Seymour B OrsquoDoherty J Kaube H Dolan RJ amp 787
Frith CD (2004) Empathy for pain involves the affective but 788
not sensory components of pain Science 303(5661) 1157- 789
1162 790
Smith MJ Keel JC Greenberg BD Adams LF Schmidt PJ 791
Rubinow DA amp Wassermann EM (1999) Menstrual cycle 792
effects on cortical excitability Neurology 53(9) 2069-2072 793
Stoodley CJ amp Schmahmann JD (2009) Functional topography 794
in the human cerebellum A meta-analysis of neuroimaging 795
studies Neuroimage 44(2) 489-501 796
Strick PL Dum RP amp Fiez JA (2009) Cerebellum and non- 797
motor function Annu Rev Neurosci 32 413-434 798
Strigo IA Duncan GH Boivin M amp Bushnell MC (2003) 799
Differentiation of visceral and cutaneous pain in the human 800
brain J Neurophysiol 89(6) 3294-3303 801
Truini A Galeotti F Romaniello A Virtuoso M Iannetti GD 802
amp Cruccu G (2005) Laser-evoked potentials Normative val- 803
ues Clin Neurophysiol 116(4) 821-826 804
Truini A Panuccio G Galeotti F Maluccio MR Sartucci F 805
Avoli M amp Cruccu G (2010) Laser-evoked potentials as a 806
tool for assessing the efficacy of antinociceptive drugs Eur J 807
Pain 14(2) 222-225 808
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 13
Truini A Vergari M Biasiotta A La Cesa S Gabriele M Di809
Stefano G Cambieri C Cruccu G Inghilleri M amp Pri-810
ori A (2011) Transcutaneous spinal direct current stimulation811
inhibits nociceptive spinal pathway conduction and increases812
pain tolerance in humans Eur J Pain 15(10) 1023-1027813
Valeriani M Le Pera D Restuccia D de Armas L Mili-814
ucci R Betti V Vigevano F amp Tonali P (2007) Parallel815
spinal pathways generate the middle-latency N1 and the late P2816
components of the laser evoked potentials Clin Neurophysiol817
118(5) 1097-1104818
Valeriani M Pazzaglia C Cruccu G amp Truini A (2012) Clinical819
usefulness of laser evoked potentials Neurophysiol Clin 42(5)820
345-353821
Volz MS Mendonca M Pinheiro FS Cui H Santana M amp822
Fregni F (2012) Dissociation of motor task-induced cortical823
excitability and pain perception changes in healthy volunteers824
PLoS One 7(3) e34273825
Wager TD Rilling JK Smith EE Sokolik A Casey KL 826
Davidson RJ Kosslyn SM Rose RM amp Cohen JD 827
(2004) Placebo-induced changes in FMRI in the anticipation 828
and experience of pain Science 303(5661) 1162-1167 829
Zaghi S Thiele B Pimentel D Pimentel T amp Fregni 830
F (2011) Assessment and treatment of pain with non- 831
invasive cortical stimulation Restor Neurol Neurosci 29(6) 832
439-451 833
Zubieta JK Bueller JA Jackson LR Scott DJ Xu Y 834
Koeppe RA Nichols TE amp Stohler CS (2005) Placebo 835
effects mediated by endogenous opioid activity on mu-opioid 836
receptors J Neurosci 25(34) 7754-7762 837
Zunhammer M Busch V Griesbach F Landgrebe M Hajak G 838
amp Langguth B (2011) rTMS over the cerebellum modulates 839
temperature detection and pain thresholds through peripheral 840
mechanisms Brain Stimul 4(4) 210-7 e1 841
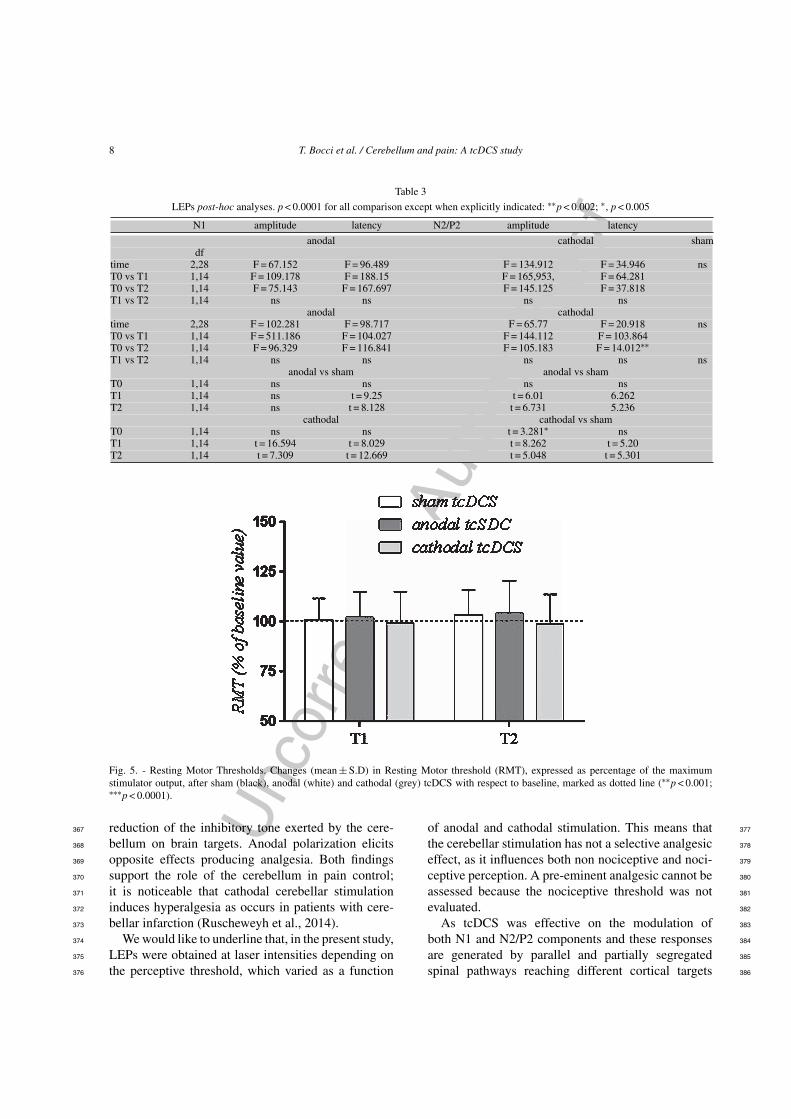
Unc
orre
cted
Aut
hor P
roof
8 T Bocci et al Cerebellum and pain A tcDCS study
Table 3
LEPs post-hoc analyses p lt 00001 for all comparison except when explicitly indicated lowastlowastp lt 0002 lowast p lt 0005
N1 amplitude latency N2P2 amplitude latency
anodal cathodal shamdf
time 228 F = 67152 F = 96489 F = 134912 F = 34946 nsT0 vs T1 114 F = 109178 F = 18815 F = 165953 F = 64281T0 vs T2 114 F = 75143 F = 167697 F = 145125 F = 37818T1 vs T2 114 ns ns ns ns
anodal cathodaltime 228 F = 102281 F = 98717 F = 6577 F = 20918 nsT0 vs T1 114 F = 511186 F = 104027 F = 144112 F = 103864T0 vs T2 114 F = 96329 F = 116841 F = 105183 F = 14012lowastlowastT1 vs T2 114 ns ns ns ns ns
anodal vs sham anodal vs shamT0 114 ns ns ns nsT1 114 ns t = 925 t = 601 6262T2 114 ns t = 8128 t = 6731 5236
cathodal cathodal vs shamT0 114 ns ns t = 3281lowast nsT1 114 t = 16594 t = 8029 t = 8262 t = 520T2 114 t = 7309 t = 12669 t = 5048 t = 5301
Fig 5 - Resting Motor Thresholds Changes (mean plusmn SD) in Resting Motor threshold (RMT) expressed as percentage of the maximumstimulator output after sham (black) anodal (white) and cathodal (grey) tcDCS with respect to baseline marked as dotted line (lowastlowastp lt 0001lowastlowastlowastp lt 00001)
reduction of the inhibitory tone exerted by the cere-367
bellum on brain targets Anodal polarization elicits368
opposite effects producing analgesia Both findings369
support the role of the cerebellum in pain control370
it is noticeable that cathodal cerebellar stimulation371
induces hyperalgesia as occurs in patients with cere-372
bellar infarction (Ruscheweyh et al 2014)373
We would like to underline that in the present study374
LEPs were obtained at laser intensities depending on375
the perceptive threshold which varied as a function376
of anodal and cathodal stimulation This means that 377
the cerebellar stimulation has not a selective analgesic 378
effect as it influences both non nociceptive and noci- 379
ceptive perception A pre-eminent analgesic cannot be 380
assessed because the nociceptive threshold was not 381
evaluated 382
As tcDCS was effective on the modulation of 383
both N1 and N2P2 components and these responses 384
are generated by parallel and partially segregated 385
spinal pathways reaching different cortical targets 386
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 9
(Valeriani et al 2007) we may suggest that the cere-387
bellum is engaged in pain processing by modulating388
the activity of both somatosensory and cingulate cor-389
tices Indeed the cerebellum is involved in both the390
sensory-discriminative and emotional dimension of391
pain (Singer et al 2004 Moriguchi et al 2007) and392
non-invasive cerebellar current stimulation may modu-393
late pain experience and the associated cortical activity394
through many not alternative mechanisms In partic-395
ular changes in N1 reflects the modulation of the396
sensory component of pain while the vertex N2P2397
represents the neural correlate of affective aspects of398
pain experience (Garcia-Larrea et al 1997 Valeriani399
et al 2007) Notably tcDCS may act not only on spinal400
nociceptive neurons but also on wide-range cortical401
networks of the pain matrix (Singer et al 2004) thus402
influencing LEPs and pain experience through both403
top-down and bottom-up mechanisms404
The present study does not allow to hypothesize405
how and where tcDCS influences the cerebellar activ-406
ity A main role of Purkinje cells has been suggested407
as their activity modulation may affect the cerebellar408
inhibitory control of the cerebral cortex (Galea et al409
2009) This would be in line with the effects elicited410
by tDCS in the cerebral cortex which are observ-411
able after both short and long term delay likely also412
interfering with long-term potentiation (LTP)-like phe-413
nomena (Hamada et al 2012 Priori et al 2014)414
Moreover prolonged spiking activity in the cerebellar415
Golgi inhibitory neurons modulates the activity of the416
Purkinje cells and could partly account for the tcDCS417
after-effects (Hull et al 2013)418
The lack of changes in RMT indicates that the anal-419
gesic effects of anodal tcDCS are due to a specific420
modulation of the cerebellar activity and not to motor421
activation On the other hand tcDCS-induced cerebel-422
lar modulation (Purpura amp McMurtry 1965) could be423
not sufficient per se to activate the cerebello ndash thalamo424
- cortical motor pathway (Galea et al 2009) thus425
the reported analgesia and its cortical correlates can-426
not be sustained by the motor cortex activation This427
view is supported by the absence of any association428
between motor symptoms and pain perception in cere-429
bellar patients (Ruscheweyh et al 2014) In the same430
line in healthy subjects it has been recently shown431
that motor task-induced increased cortical excitability432
and analgesia are not associated (Volz et al 2012)433
Indeed RMT is a highly sensitive marker of motor434
tract excitability as it reflects activation of a small435
low-threshold and slow-conducting core of pyramidal436
neurons (Hess et al 1987 Rossini amp Rossi 2007) 437
although RMT may reflect changes in the activity of 438
different central nervous system structures it has been 439
satisfactorily used to assess motor cortex excitability 440
also in cerebellar patients (Battaglia et al 2006) 441
Another critical point is the possibility to modulate 442
with tcDCS both neural correlates underlying nocicep- 443
tive processing and pain perception Previous studies 444
using tDCS over motor cortex were inconsistent among 445
each other some works suggested that tDCS is able to 446
modify pain perception (Boggio et al 2008) while 447
others showed divergent effects on psychophysical 448
and neurophysiological outcome parameters (Luedtke 449
et al 2012 Ihle et al 2014) likely due to a possi- 450
ble overestimation of the role of motor areas on pain 451
processing (Antal et al 2008) 452
Our findings cannot be compared to the results 453
obtained by other Authors In fact the unique study 454
focused on the analgesic effects of non-invasive cere- 455
bellar stimulation reported till now (Zunhammer et al 456
2011) considered only subjective pain thresholds 457
In addition it described similar analgesic effects of 458
cerebellar and neck structures repetitive transcranial 459
magnetic stimulation (rTMS) thus denying any cere- 460
bellar specificity in the observed effects and suggesting 461
that the peripheral information passing through the 462
cerebellum may be responsible for analgesia The 463
main difference between the two studies possibly 464
accounting for different results consists of the neu- 465
romodulation techniques used 466
41 Limitations of the study 467
The present study has a few limitations First our 468
findings do not allow any hypothesis on the role of 469
the cerebellum in chronic pain The observations on 470
patients with cerebellar damage (Ruscheweyh et al 471
2014) suggest that their impaired inhibitory control 472
mechanisms may be not associated with the devel- 473
opment of chronic pain Second we cannot exclude 474
the possibility that tcDCS could modulate not only the 475
cerebellum but also surrounding areas such as the peri- 476
aqueductal gray However recent modelling researches 477
have revealed that during tcDCS the current spread 478
to other structures outside the cerebellum is negligi- 479
ble (Parazzini et al 2013 2014) Moreover several 480
studies have proved that in humans pain processing 481
is encoded within posterior areas of each cerebellar 482
hemisphere specifically in the hemispheric lobule VI 483
Crus I and VIIb (Moulton et al 2011) where the 484
Unc
orre
cted
Aut
hor P
roof
10 T Bocci et al Cerebellum and pain A tcDCS study
tcDCS-induced electrical field is strongly concentrated485
(Parazzini et al 2013) A further limitation is that we486
did not study the contribution of C-fibers the main487
component of spino-ponto-cerebellar and spino-olivo-488
cerebellar pathways In fact ultra-late LEPs related489
to C-fibers activations have not yet been standard-490
ized for clinical application and their occurrence could491
be markedly influenced by high order cognitive pro-492
cesses as they seem to be more affected by the level493
of consciousness and attention than A-delta responses494
(Qiu et al 2002 Opsommer et al 2003 Mouraux amp495
Plaghki 2006) Finally we wish to emphasize that in496
neuropathic patients the effects of the cerebellar stim-497
ulation could be quite different from those described498
here as both anatomical and functional connectivity499
are different from those observed in healthy partici-500
pants (Rocca et al 2010 Riedl et al 2011 Absinta et501
al 2012 Longo et al 2012 Ceko et al 2013)502
5 Conclusions503
Our findings indicate a cerebellar effect on pain504
experience and on its cortical correlates and prompt505
further investigation aimed at assessing whether the506
cerebellar direct current polarization could be used as a507
novel and safe therapeutic tool in chronic pain patients508
Acknowledgments509
We gratefully acknowledge the participation of all510
subjects as well as Mr D Barloscio and Mr C511
Orsini for their excellent technical assistance The512
paper was supported in part by the Italian operating513
and development MIUR PRIN grant year 2006 n514
2006062332 002515
RF and AP are founders and shareholders of516
Newronika srl Milan Italy517
References518
Absinta M Rocca MA Colombo B Falini A Comi G amp Fil-519
ippi M (2012) Selective decreased grey matter volume of the520
pain-matrix network in cluster headache Cephalalgia 32(2)521
109-115522
Agostino R Cruccu G Iannetti G Romaniello A Truini A523
amp Manfredi M (2000) Topographical distribution of pinprick524
and warmth thresholds to CO2 laser stimulation on the human525
skin Neurosci Lett 285(2) 115-118526
Antal A Brepohl N Poreisz C Boros K Csifcsak G amp 527
Paulus W (2008) Transcranial direct current stimulation over 528
somatosensory cortex decreases experimentally induced acute 529
pain perception Clin J Pain 24(1) 56-63 530
Antal A amp Paulus W (2010) Effects of transcranial theta-burst 531
stimulation on acute pain perception Restor Neurol Neurosci 532
28(4) 477-484 533
Apkarian AV Bushnell MC Treede RD amp Zubieta JK 534
(2005) Human brain mechanisms of pain perception and reg- 535
ulation in health and disease Eur J Pain 9(4) 463-484 536
Balsters JH Whelan CD Robertson IH amp Ramnani N (2013) 537
Cerebellum and cognition Evidence for the encoding of higher 538
order rules Cereb Cortex 23(6) 1433-1443 539
Battaglia F Quartarone A Ghilardi MF Dattola R Bagnato 540
S Rizzo V Morgante L amp Girlanda P (2006) Unilat- 541
eral cerebellar stroke disrupts movement preparation and motor 542
imagery Clin Neurophysiol 117(5) 1009-1016 543
Bingel U Quante M Knab R Bromm B Weiller C amp Buchel 544
C (2002) Subcortical structures involved in pain processing 545
Evidence from single-trial fMRI Pain 99(1-2) 313-321 546
Boggio PS Zaghi S amp Fregni F (2009) Modulation of emotions 547
associated with images of human pain using anodal transcranial 548
direct current stimulation (tDCS) Neuropsychologia 47(1) 549
212-217 550
Boggio PS Zaghi S Lopes M amp Fregni F (2008) Modulatory 551
effects of anodal transcranial direct current stimulation on per- 552
ception and pain thresholds in healthy volunteers Eur J Neurol 553
15(10) 1124-1130 554
Borsook D Moulton EA Tully S Schmahmann JD amp 555
Becerra L (2008) Human cerebellar responses to brush and 556
heat stimuli in healthy and neuropathic pain subjects Cerebel- 557
lum 7(3) 252-272 558
Ceko M Seminowicz DA Bushnell MC amp Olausson HW 559
(2013) Anatomical and functional enhancements of the insula 560
after loss of large primary somatosensory fibers Cereb Cortex 561
23(9) 2017-2024 562
Cruccu G Aminoff MJ Curio G Guerit JM Kakigi 563
R Mauguiere F Rossini PM Treede RD amp Garcia- 564
Larrea L (2008) Recommendations for the clinical use of 565
somatosensory-evoked potentials Clin Neurophysiol 119(8) 566
1705-1719 567
Cruccu G Romaniello A Amantini A Lombardi M Innocenti 568
P amp Manfredi M (1999) Assessment of trigeminal small- 569
fiber function Brain and reflex responses evoked by CO2-laser 570
stimulation Muscle Nerve 22(4) 508-516 571
Dasilva AF Mendonca ME Zaghi S Lopes M Dossantos 572
MF Spierings EL Bajwa Z Datta A Bikson M amp 573
Fregni F (2012) tDCS-induced analgesia and electrical fields 574
in pain-related neural networks in chronic migraine Headache 575
52(8) 1283-1295 576
Duerden EG amp Albanese MC (2013) Localization of pain- 577
related brain activation A meta-analysis of neuroimaging data 578
Hum Brain Mapp 34(1) 109-149 579
Ekerot CF Garwicz M amp Schouenborg J (1991a) The postsy- 580
naptic dorsal column pathway mediates cutaneous nociceptive 581
information to cerebellar climbing fibres in the cat J Physiol 582
441 275-284 583
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 11
Ekerot CF Garwicz M amp Schouenborg J (1991b) Topography584
and nociceptive receptive fields of climbing fibres projecting to585
the cerebellar anterior lobe in the cat J Physiol 441 257-274586
Ekerot CF Gustavsson P Oscarsson O amp Schouenborg J587
(1987a) Climbing fibres projecting to cat cerebellar anterior588
lobe activated by cutaneous A and C fibres J Physiol 386589
529-538590
Ekerot CF Oscarsson O amp Schouenborg J (1987b) Stimula-591
tion of cat cutaneous nociceptive C fibres causing tonic and592
synchronous activity in climbing fibres J Physiol 386 539-593
546594
Ferrucci R Brunoni AR Parazzini M Vergari M Rossi E595
Fumagalli M Mameli F Rosa M Giannicola G Zago S596
amp Priori A (2013) Modulating human procedural learning by597
cerebellar transcranial direct current stimulation Cerebellum598
12(4) 485-492599
Ferrucci R Giannicola G Rosa M Fumagalli M Boggio600
PS Hallett M Zago S amp Priori A (2012) Cerebellum601
and processing of negative facial emotions Cerebellar tran-602
scranial DC stimulation specifically enhances the emotional603
recognition of facial anger and sadness Cogn Emot 26(5)604
786-799605
Ferrucci R Marceglia S Vergari M Cogiamanian F Mrakic-606
Sposta S Mameli F Zago S Barbieri S amp Priori607
A (2008) Cerebellar transcranial direct current stimulation608
impairs the practice-dependent proficiency increase in working609
memory J Cogn Neurosci 20(9) 1687-1697610
Fregni F Freedman S amp Pascual-Leone A (2007) Recent611
advances in the treatment of chronic pain with non-invasive612
brain stimulation techniques Lancet Neurol 6(2) 188-191613
Galea JM Jayaram G Ajagbe L amp Celnik P (2009)614
Modulation of cerebellar excitability by polarity-specific615
noninvasive direct current stimulation J Neurosci 29(28)616
9115-9122617
Galea JM Vazquez A Pasricha N de Xivry JJ amp Celnik618
P (2011) Dissociating the roles of the cerebellum and motor619
cortex during adaptive learning The motor cortex retains what620
the cerebellum learns Cereb Cortex 21(8) 1761-1770621
Gandiga PC Hummel FC amp Cohen LG (2006) Tran-622
scranial DC stimulation (tDCS) A tool for double-blind623
sham-controlled clinical studies in brain stimulation Clin Neu-624
rophysiol 117(4) 845-850625
Garcia-Larrea L Peyron R Laurent B amp Mauguiere F (1997)626
Association and dissociation between laser-evoked potentials627
and pain perception Neuroreport 8(17) 3785-3789628
Grimaldi G Argyropoulos GP Boehringer A Celnik P629
Edwards MJ Ferrucci R Galea JM Groiss SJ Hiraoka630
K Kassavetis P Lesage E Manto M Miall RC Priori A631
Sadnicka A Ugawa Y amp Ziemann U (2014) Non-invasive632
cerebellar stimulationndasha consensus paper Cerebellum 13(1)633
121-138634
Groppa S Oliviero A Eisen A Quartarone A Cohen LG635
Mall V Kaelin-Lang A Mima T Rossi S Thickbroom636
GW Rossini PM Ziemann U Valls-Sole J amp Sieb-637
ner HR (2012) A practical guide to diagnostic transcranial638
magnetic stimulation Report of an IFCN committee Clin Neu-639
rophysiol 123(5) 858-882640
Hamada M Strigaro G Murase N Sadnicka A Galea JM 641
Edwards MJ amp Rothwell JC (2012) Cerebellar modulation 642
of human associative plasticity J Physiol 590(Pt 10) 2365- 643
2374 644
Hess CW Mills KR amp Murray NM (1987) Responses in small 645
hand muscles from magnetic stimulation of the human brain J 646
Physiol 388 397-419 647
Hull CA Chu Y Thanawala M amp Regehr WG (2013) Hyper- 648
polarization induces a long-term increase in the spontaneous 649
firing rate of cerebellar Golgi cells J Neurosci 33(14) 5895- 650
5902 651
Ihle K Rodriguez-Raecke R amp Luedtke K May A (2014) 652
tDCS modulates cortical nociceptive processing but has little 653
to no impact on pain perception Pain 155(10) 2080-2087 654
Ito M (2006) Cerebellar circuitry as a neuronal machine Prog 655
Neurobiol 78(3-5) 272-303 656
Iyer MB Mattu U Grafman J Lomarev M Sato S amp Wasser- 657
mann EM (2005) Safety and cognitive effect of frontal DC 658
brain polarization in healthy individuals Neurology 64(5) 872- 659
875 660
Kelly RM amp Strick PL (2003) Cerebellar loops with motor cor- 661
tex and prefrontal cortex of a nonhuman primate J Neurosci 662
23(23) 8432-8444 663
Lefaucheur JP Andre-Obadia N Antal A Ayache SS Baeken 664
C Benninger DH Cantello RM Cincotta M de Carvalho 665
M De Ridder D Devanne H Di Lazzaro V Filipovic SR 666
Hummel FC Jaaskelainen SK Kimiskidis VK Koch G 667
Langguth B Nyffeler T Oliviero A Padberg F Poulet E 668
Rossi S Rossini PM Rothwell JC Schonfeldt-Lecuona 669
C Siebner HR Slotema CW Stagg CJ Valls-Sole J 670
Ziemann U Paulus W amp Garcia-Larrea L (2014) Evidence- 671
based guidelines on the therapeutic use of repetitive transcranial 672
magnetic stimulation (rTMS) Clin Neurophysiol 125(11) 673
2150-2206 674
Lefaucheur JP Brusa A Creange A Drouot X amp Jarry G 675
(2002) Clinical application of laser evoked potentials using 676
the NdYAG laser Neurophysiol Clin 32(2) 91-98 677
Lefaucheur JP Debray S amp Jarry G (2001) Laser evoked poten- 678
tials using the NdYAG laser Muscle Nerve 24(4) 496-501 679
Lefaucheur JP Drouot X Menard-Lefaucheur I Keravel Y 680
amp Nguyen JP (2006) Motor cortex rTMS restores defective 681
intracortical inhibition in chronic neuropathic pain Neurology 682
67(9) 1568-1574 683
Longo MR Iannetti GD Mancini F Driver J amp Haggard P 684
(2012) Linking pain and the body Neural correlates of visually 685
induced analgesia J Neurosci 32(8) 2601-2607 686
Lorenz J amp Garcia-Larrea L (2003) Contribution of attentional 687
and cognitive factors to laser evoked brain potentials Neuro- 688
physiol Clin 33(6) 293-301 689
Lorenz J Minoshima S amp Casey KL (2003) Keeping pain out 690
of mind The role of the dorsolateral prefrontal cortex in pain 691
modulation Brain 126(Pt 5) 1079-1091 692
Luedtke K May A amp Jurgens TP (2012) No effect of a single 693
session of transcranial direct current stimulation on experimen- 694
tally induced pain in patients with chronic low back painndashan 695
exploratory study PLoS One 7(11) e48857 696
Unc
orre
cted
Aut
hor P
roof
12 T Bocci et al Cerebellum and pain A tcDCS study
Mendonca ME Santana MB Baptista AF Datta A Bikson697
M Fregni F amp Araujo CP (2011) Transcranial DC stimu-698
lation in fibromyalgia Optimized cortical target supported by699
high-resolution computational models J Pain 12(5) 610-617700
Moriguchi Y Decety J Ohnishi T Maeda M Mori T Nemoto701
K Matsuda H amp Komaki G (2007) Empathy and judg-702
ing otherrsquos pain An fMRI study of alexithymia Cereb Cortex703
17(9) 2223-2234704
Moulton EA Elman I Pendse G Schmahmann J Becerra L705
amp Borsook D (2011) Aversion-related circuitry in the cere-706
bellum Responses to noxious heat and unpleasant images J707
Neurosci 31(10) 3795-3804708
Moulton EA Schmahmann JD Becerra L amp Borsook D709
(2010) The cerebellum and pain Passive integrator or active710
participator Brain Res Rev 65(1) 14-27711
Mouraux A amp Plaghki L (2006) Are the processes reflected by712
late and ultra-late laser evoked potentials specific of nocicep-713
tion Suppl Clin Neurophysiol 59 197-204714
Mylius V Jung M Menzler K Haag A Khader PH Oer-715
tel WH Rosenow F amp Lefaucheur JP (2012) Effects of716
transcranial direct current stimulation on pain perception and717
working memory Eur J Pain 16(7) 974-982718
Ni Z Gunraj C amp Chen R (2007) Short interval intracortical719
inhibition and facilitation during the silent period in human J720
Physiol 583(Pt 3) 971-982721
Nitsche MA Liebetanz D Lang N Antal A Tergau F amp722
Paulus W (2003) Safety criteria for transcranial direct cur-723
rent stimulation (tDCS) in humans Clin Neurophysiol 114(11)724
2220-2222 author reply 2222-2223725
Opsommer E Guerit JM amp Plaghki L (2003) Exogenous726
and endogenous components of ultralate (C-fibre) evoked727
potentials following CO2 laser stimuli to tiny skin sur-728
face areas in healthy subjects Neurophysiol Clin 33(2)729
78-85730
Parazzini M Fiocchi S Liorni I Priori A amp Ravazzani P731
(2014) Computational modeling of transcranial direct current732
stimulation in the child brain Implications for the treatment of733
refractory childhood focal epilepsy Int J Neural Syst 24(2)734
1430006735
Parazzini M Rossi E Ferrucci R Fiocchi S Liorni I Priori736
A amp Ravazzani P (2013) Computational model of cerebellar737
transcranial direct current stimulation Conf Proc IEEE Eng738
Med Biol Soc 2013 237-240739
Parazzini M Rossi E Ferrucci R Liorni I Priori A amp Ravaz-740
zani P (2014) Modelling the electric field and the current741
density generated by cerebellar transcranial DC stimulation in742
humans Clin Neurophysiol 125(3) 577-584743
Perini I Bergstrand S amp Morrison I (2013) Where pain meets744
action in the human brain J Neurosci 33(40) 15930-15939745
Ploghaus A Tracey I Gati JS Clare S Menon RS Matthews746
PM amp Rawlins JN (1999) Dissociating pain from its antic-747
ipation in the human brain Science 284(5422) 1979-1981748
Priori A Ciocca M Parazzini M Vergari M amp Ferrucci R749
(2014) Transcranial cerebellar direct current stimulation and750
transcutaneous spinal cord direct current stimulation as innova-751
tive tools for neuroscientists J Physiol 592(Pt 16) 3345-3369752
Purpura DP amp McMurtry JC (1965) Intracellular activities and 753
evoked potential changes during polarization of motor cortex 754
J Neurophysiol 28 166-185 755
Qiu Y Inui K Wang X Tran TD amp Kakigi R (2002) Effects 756
of attention distraction and sleep on CO(2) laser evoked poten- 757
tials related to C-fibers in humans Clin Neurophysiol 113(10) 758
1579-1585 759
Reidler JS Mendonca ME Santana MB Wang X Lenkinski 760
R Motta AF Marchand S Latif L amp Fregni F (2012) 761
Effects of motor cortex modulation and descending inhibitory 762
systems on pain thresholds in healthy subjects J Pain 13(5) 763
450-458 764
Riedl V Valet M Woller A Sorg C Vogel D Sprenger 765
T Boecker H Wohlschlager AM amp Tolle TR (2011) 766
Repeated pain induces adaptations of intrinsic brain activity 767
to reflect past and predict future pain Neuroimage 57(1) 206- 768
213 769
Rocca MA Valsasina P Absinta M Colombo B Barcella V 770
Falini A Comi G amp Filippi M (2010) Central nervous 771
system dysregulation extends beyond the pain-matrix network 772
in cluster headache Cephalalgia 30(11) 1383-1391 773
Rossi S Hallett M Rossini PM amp Pascual-Leone A (2009) 774
Safety ethical considerations and application guidelines for 775
the use of transcranial magnetic stimulation in clinical practice 776
and research Clin Neurophysiol 120(12) 2008-2039 777
Rossini PM amp Rossi S (2007) Transcranial magnetic stimulation 778
Diagnostic therapeutic and research potential Neurology 779
68(7) 484-488 780
Ruscheweyh R Kuhnel M Filippopulos F Blum B Eggert 781
T amp Straube A (2014) Altered experimental pain perception 782
after cerebellar infarction Pain 155(7) 1303-1312 783
Schmahmann JD (1991) An emerging concept The cerebellar 784
contribution to higher function Arch Neurol 48(11) 1178- 785
1187 786
Singer T Seymour B OrsquoDoherty J Kaube H Dolan RJ amp 787
Frith CD (2004) Empathy for pain involves the affective but 788
not sensory components of pain Science 303(5661) 1157- 789
1162 790
Smith MJ Keel JC Greenberg BD Adams LF Schmidt PJ 791
Rubinow DA amp Wassermann EM (1999) Menstrual cycle 792
effects on cortical excitability Neurology 53(9) 2069-2072 793
Stoodley CJ amp Schmahmann JD (2009) Functional topography 794
in the human cerebellum A meta-analysis of neuroimaging 795
studies Neuroimage 44(2) 489-501 796
Strick PL Dum RP amp Fiez JA (2009) Cerebellum and non- 797
motor function Annu Rev Neurosci 32 413-434 798
Strigo IA Duncan GH Boivin M amp Bushnell MC (2003) 799
Differentiation of visceral and cutaneous pain in the human 800
brain J Neurophysiol 89(6) 3294-3303 801
Truini A Galeotti F Romaniello A Virtuoso M Iannetti GD 802
amp Cruccu G (2005) Laser-evoked potentials Normative val- 803
ues Clin Neurophysiol 116(4) 821-826 804
Truini A Panuccio G Galeotti F Maluccio MR Sartucci F 805
Avoli M amp Cruccu G (2010) Laser-evoked potentials as a 806
tool for assessing the efficacy of antinociceptive drugs Eur J 807
Pain 14(2) 222-225 808
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 13
Truini A Vergari M Biasiotta A La Cesa S Gabriele M Di809
Stefano G Cambieri C Cruccu G Inghilleri M amp Pri-810
ori A (2011) Transcutaneous spinal direct current stimulation811
inhibits nociceptive spinal pathway conduction and increases812
pain tolerance in humans Eur J Pain 15(10) 1023-1027813
Valeriani M Le Pera D Restuccia D de Armas L Mili-814
ucci R Betti V Vigevano F amp Tonali P (2007) Parallel815
spinal pathways generate the middle-latency N1 and the late P2816
components of the laser evoked potentials Clin Neurophysiol817
118(5) 1097-1104818
Valeriani M Pazzaglia C Cruccu G amp Truini A (2012) Clinical819
usefulness of laser evoked potentials Neurophysiol Clin 42(5)820
345-353821
Volz MS Mendonca M Pinheiro FS Cui H Santana M amp822
Fregni F (2012) Dissociation of motor task-induced cortical823
excitability and pain perception changes in healthy volunteers824
PLoS One 7(3) e34273825
Wager TD Rilling JK Smith EE Sokolik A Casey KL 826
Davidson RJ Kosslyn SM Rose RM amp Cohen JD 827
(2004) Placebo-induced changes in FMRI in the anticipation 828
and experience of pain Science 303(5661) 1162-1167 829
Zaghi S Thiele B Pimentel D Pimentel T amp Fregni 830
F (2011) Assessment and treatment of pain with non- 831
invasive cortical stimulation Restor Neurol Neurosci 29(6) 832
439-451 833
Zubieta JK Bueller JA Jackson LR Scott DJ Xu Y 834
Koeppe RA Nichols TE amp Stohler CS (2005) Placebo 835
effects mediated by endogenous opioid activity on mu-opioid 836
receptors J Neurosci 25(34) 7754-7762 837
Zunhammer M Busch V Griesbach F Landgrebe M Hajak G 838
amp Langguth B (2011) rTMS over the cerebellum modulates 839
temperature detection and pain thresholds through peripheral 840
mechanisms Brain Stimul 4(4) 210-7 e1 841

Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 9
(Valeriani et al 2007) we may suggest that the cere-387
bellum is engaged in pain processing by modulating388
the activity of both somatosensory and cingulate cor-389
tices Indeed the cerebellum is involved in both the390
sensory-discriminative and emotional dimension of391
pain (Singer et al 2004 Moriguchi et al 2007) and392
non-invasive cerebellar current stimulation may modu-393
late pain experience and the associated cortical activity394
through many not alternative mechanisms In partic-395
ular changes in N1 reflects the modulation of the396
sensory component of pain while the vertex N2P2397
represents the neural correlate of affective aspects of398
pain experience (Garcia-Larrea et al 1997 Valeriani399
et al 2007) Notably tcDCS may act not only on spinal400
nociceptive neurons but also on wide-range cortical401
networks of the pain matrix (Singer et al 2004) thus402
influencing LEPs and pain experience through both403
top-down and bottom-up mechanisms404
The present study does not allow to hypothesize405
how and where tcDCS influences the cerebellar activ-406
ity A main role of Purkinje cells has been suggested407
as their activity modulation may affect the cerebellar408
inhibitory control of the cerebral cortex (Galea et al409
2009) This would be in line with the effects elicited410
by tDCS in the cerebral cortex which are observ-411
able after both short and long term delay likely also412
interfering with long-term potentiation (LTP)-like phe-413
nomena (Hamada et al 2012 Priori et al 2014)414
Moreover prolonged spiking activity in the cerebellar415
Golgi inhibitory neurons modulates the activity of the416
Purkinje cells and could partly account for the tcDCS417
after-effects (Hull et al 2013)418
The lack of changes in RMT indicates that the anal-419
gesic effects of anodal tcDCS are due to a specific420
modulation of the cerebellar activity and not to motor421
activation On the other hand tcDCS-induced cerebel-422
lar modulation (Purpura amp McMurtry 1965) could be423
not sufficient per se to activate the cerebello ndash thalamo424
- cortical motor pathway (Galea et al 2009) thus425
the reported analgesia and its cortical correlates can-426
not be sustained by the motor cortex activation This427
view is supported by the absence of any association428
between motor symptoms and pain perception in cere-429
bellar patients (Ruscheweyh et al 2014) In the same430
line in healthy subjects it has been recently shown431
that motor task-induced increased cortical excitability432
and analgesia are not associated (Volz et al 2012)433
Indeed RMT is a highly sensitive marker of motor434
tract excitability as it reflects activation of a small435
low-threshold and slow-conducting core of pyramidal436
neurons (Hess et al 1987 Rossini amp Rossi 2007) 437
although RMT may reflect changes in the activity of 438
different central nervous system structures it has been 439
satisfactorily used to assess motor cortex excitability 440
also in cerebellar patients (Battaglia et al 2006) 441
Another critical point is the possibility to modulate 442
with tcDCS both neural correlates underlying nocicep- 443
tive processing and pain perception Previous studies 444
using tDCS over motor cortex were inconsistent among 445
each other some works suggested that tDCS is able to 446
modify pain perception (Boggio et al 2008) while 447
others showed divergent effects on psychophysical 448
and neurophysiological outcome parameters (Luedtke 449
et al 2012 Ihle et al 2014) likely due to a possi- 450
ble overestimation of the role of motor areas on pain 451
processing (Antal et al 2008) 452
Our findings cannot be compared to the results 453
obtained by other Authors In fact the unique study 454
focused on the analgesic effects of non-invasive cere- 455
bellar stimulation reported till now (Zunhammer et al 456
2011) considered only subjective pain thresholds 457
In addition it described similar analgesic effects of 458
cerebellar and neck structures repetitive transcranial 459
magnetic stimulation (rTMS) thus denying any cere- 460
bellar specificity in the observed effects and suggesting 461
that the peripheral information passing through the 462
cerebellum may be responsible for analgesia The 463
main difference between the two studies possibly 464
accounting for different results consists of the neu- 465
romodulation techniques used 466
41 Limitations of the study 467
The present study has a few limitations First our 468
findings do not allow any hypothesis on the role of 469
the cerebellum in chronic pain The observations on 470
patients with cerebellar damage (Ruscheweyh et al 471
2014) suggest that their impaired inhibitory control 472
mechanisms may be not associated with the devel- 473
opment of chronic pain Second we cannot exclude 474
the possibility that tcDCS could modulate not only the 475
cerebellum but also surrounding areas such as the peri- 476
aqueductal gray However recent modelling researches 477
have revealed that during tcDCS the current spread 478
to other structures outside the cerebellum is negligi- 479
ble (Parazzini et al 2013 2014) Moreover several 480
studies have proved that in humans pain processing 481
is encoded within posterior areas of each cerebellar 482
hemisphere specifically in the hemispheric lobule VI 483
Crus I and VIIb (Moulton et al 2011) where the 484
Unc
orre
cted
Aut
hor P
roof
10 T Bocci et al Cerebellum and pain A tcDCS study
tcDCS-induced electrical field is strongly concentrated485
(Parazzini et al 2013) A further limitation is that we486
did not study the contribution of C-fibers the main487
component of spino-ponto-cerebellar and spino-olivo-488
cerebellar pathways In fact ultra-late LEPs related489
to C-fibers activations have not yet been standard-490
ized for clinical application and their occurrence could491
be markedly influenced by high order cognitive pro-492
cesses as they seem to be more affected by the level493
of consciousness and attention than A-delta responses494
(Qiu et al 2002 Opsommer et al 2003 Mouraux amp495
Plaghki 2006) Finally we wish to emphasize that in496
neuropathic patients the effects of the cerebellar stim-497
ulation could be quite different from those described498
here as both anatomical and functional connectivity499
are different from those observed in healthy partici-500
pants (Rocca et al 2010 Riedl et al 2011 Absinta et501
al 2012 Longo et al 2012 Ceko et al 2013)502
5 Conclusions503
Our findings indicate a cerebellar effect on pain504
experience and on its cortical correlates and prompt505
further investigation aimed at assessing whether the506
cerebellar direct current polarization could be used as a507
novel and safe therapeutic tool in chronic pain patients508
Acknowledgments509
We gratefully acknowledge the participation of all510
subjects as well as Mr D Barloscio and Mr C511
Orsini for their excellent technical assistance The512
paper was supported in part by the Italian operating513
and development MIUR PRIN grant year 2006 n514
2006062332 002515
RF and AP are founders and shareholders of516
Newronika srl Milan Italy517
References518
Absinta M Rocca MA Colombo B Falini A Comi G amp Fil-519
ippi M (2012) Selective decreased grey matter volume of the520
pain-matrix network in cluster headache Cephalalgia 32(2)521
109-115522
Agostino R Cruccu G Iannetti G Romaniello A Truini A523
amp Manfredi M (2000) Topographical distribution of pinprick524
and warmth thresholds to CO2 laser stimulation on the human525
skin Neurosci Lett 285(2) 115-118526
Antal A Brepohl N Poreisz C Boros K Csifcsak G amp 527
Paulus W (2008) Transcranial direct current stimulation over 528
somatosensory cortex decreases experimentally induced acute 529
pain perception Clin J Pain 24(1) 56-63 530
Antal A amp Paulus W (2010) Effects of transcranial theta-burst 531
stimulation on acute pain perception Restor Neurol Neurosci 532
28(4) 477-484 533
Apkarian AV Bushnell MC Treede RD amp Zubieta JK 534
(2005) Human brain mechanisms of pain perception and reg- 535
ulation in health and disease Eur J Pain 9(4) 463-484 536
Balsters JH Whelan CD Robertson IH amp Ramnani N (2013) 537
Cerebellum and cognition Evidence for the encoding of higher 538
order rules Cereb Cortex 23(6) 1433-1443 539
Battaglia F Quartarone A Ghilardi MF Dattola R Bagnato 540
S Rizzo V Morgante L amp Girlanda P (2006) Unilat- 541
eral cerebellar stroke disrupts movement preparation and motor 542
imagery Clin Neurophysiol 117(5) 1009-1016 543
Bingel U Quante M Knab R Bromm B Weiller C amp Buchel 544
C (2002) Subcortical structures involved in pain processing 545
Evidence from single-trial fMRI Pain 99(1-2) 313-321 546
Boggio PS Zaghi S amp Fregni F (2009) Modulation of emotions 547
associated with images of human pain using anodal transcranial 548
direct current stimulation (tDCS) Neuropsychologia 47(1) 549
212-217 550
Boggio PS Zaghi S Lopes M amp Fregni F (2008) Modulatory 551
effects of anodal transcranial direct current stimulation on per- 552
ception and pain thresholds in healthy volunteers Eur J Neurol 553
15(10) 1124-1130 554
Borsook D Moulton EA Tully S Schmahmann JD amp 555
Becerra L (2008) Human cerebellar responses to brush and 556
heat stimuli in healthy and neuropathic pain subjects Cerebel- 557
lum 7(3) 252-272 558
Ceko M Seminowicz DA Bushnell MC amp Olausson HW 559
(2013) Anatomical and functional enhancements of the insula 560
after loss of large primary somatosensory fibers Cereb Cortex 561
23(9) 2017-2024 562
Cruccu G Aminoff MJ Curio G Guerit JM Kakigi 563
R Mauguiere F Rossini PM Treede RD amp Garcia- 564
Larrea L (2008) Recommendations for the clinical use of 565
somatosensory-evoked potentials Clin Neurophysiol 119(8) 566
1705-1719 567
Cruccu G Romaniello A Amantini A Lombardi M Innocenti 568
P amp Manfredi M (1999) Assessment of trigeminal small- 569
fiber function Brain and reflex responses evoked by CO2-laser 570
stimulation Muscle Nerve 22(4) 508-516 571
Dasilva AF Mendonca ME Zaghi S Lopes M Dossantos 572
MF Spierings EL Bajwa Z Datta A Bikson M amp 573
Fregni F (2012) tDCS-induced analgesia and electrical fields 574
in pain-related neural networks in chronic migraine Headache 575
52(8) 1283-1295 576
Duerden EG amp Albanese MC (2013) Localization of pain- 577
related brain activation A meta-analysis of neuroimaging data 578
Hum Brain Mapp 34(1) 109-149 579
Ekerot CF Garwicz M amp Schouenborg J (1991a) The postsy- 580
naptic dorsal column pathway mediates cutaneous nociceptive 581
information to cerebellar climbing fibres in the cat J Physiol 582
441 275-284 583
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 11
Ekerot CF Garwicz M amp Schouenborg J (1991b) Topography584
and nociceptive receptive fields of climbing fibres projecting to585
the cerebellar anterior lobe in the cat J Physiol 441 257-274586
Ekerot CF Gustavsson P Oscarsson O amp Schouenborg J587
(1987a) Climbing fibres projecting to cat cerebellar anterior588
lobe activated by cutaneous A and C fibres J Physiol 386589
529-538590
Ekerot CF Oscarsson O amp Schouenborg J (1987b) Stimula-591
tion of cat cutaneous nociceptive C fibres causing tonic and592
synchronous activity in climbing fibres J Physiol 386 539-593
546594
Ferrucci R Brunoni AR Parazzini M Vergari M Rossi E595
Fumagalli M Mameli F Rosa M Giannicola G Zago S596
amp Priori A (2013) Modulating human procedural learning by597
cerebellar transcranial direct current stimulation Cerebellum598
12(4) 485-492599
Ferrucci R Giannicola G Rosa M Fumagalli M Boggio600
PS Hallett M Zago S amp Priori A (2012) Cerebellum601
and processing of negative facial emotions Cerebellar tran-602
scranial DC stimulation specifically enhances the emotional603
recognition of facial anger and sadness Cogn Emot 26(5)604
786-799605
Ferrucci R Marceglia S Vergari M Cogiamanian F Mrakic-606
Sposta S Mameli F Zago S Barbieri S amp Priori607
A (2008) Cerebellar transcranial direct current stimulation608
impairs the practice-dependent proficiency increase in working609
memory J Cogn Neurosci 20(9) 1687-1697610
Fregni F Freedman S amp Pascual-Leone A (2007) Recent611
advances in the treatment of chronic pain with non-invasive612
brain stimulation techniques Lancet Neurol 6(2) 188-191613
Galea JM Jayaram G Ajagbe L amp Celnik P (2009)614
Modulation of cerebellar excitability by polarity-specific615
noninvasive direct current stimulation J Neurosci 29(28)616
9115-9122617
Galea JM Vazquez A Pasricha N de Xivry JJ amp Celnik618
P (2011) Dissociating the roles of the cerebellum and motor619
cortex during adaptive learning The motor cortex retains what620
the cerebellum learns Cereb Cortex 21(8) 1761-1770621
Gandiga PC Hummel FC amp Cohen LG (2006) Tran-622
scranial DC stimulation (tDCS) A tool for double-blind623
sham-controlled clinical studies in brain stimulation Clin Neu-624
rophysiol 117(4) 845-850625
Garcia-Larrea L Peyron R Laurent B amp Mauguiere F (1997)626
Association and dissociation between laser-evoked potentials627
and pain perception Neuroreport 8(17) 3785-3789628
Grimaldi G Argyropoulos GP Boehringer A Celnik P629
Edwards MJ Ferrucci R Galea JM Groiss SJ Hiraoka630
K Kassavetis P Lesage E Manto M Miall RC Priori A631
Sadnicka A Ugawa Y amp Ziemann U (2014) Non-invasive632
cerebellar stimulationndasha consensus paper Cerebellum 13(1)633
121-138634
Groppa S Oliviero A Eisen A Quartarone A Cohen LG635
Mall V Kaelin-Lang A Mima T Rossi S Thickbroom636
GW Rossini PM Ziemann U Valls-Sole J amp Sieb-637
ner HR (2012) A practical guide to diagnostic transcranial638
magnetic stimulation Report of an IFCN committee Clin Neu-639
rophysiol 123(5) 858-882640
Hamada M Strigaro G Murase N Sadnicka A Galea JM 641
Edwards MJ amp Rothwell JC (2012) Cerebellar modulation 642
of human associative plasticity J Physiol 590(Pt 10) 2365- 643
2374 644
Hess CW Mills KR amp Murray NM (1987) Responses in small 645
hand muscles from magnetic stimulation of the human brain J 646
Physiol 388 397-419 647
Hull CA Chu Y Thanawala M amp Regehr WG (2013) Hyper- 648
polarization induces a long-term increase in the spontaneous 649
firing rate of cerebellar Golgi cells J Neurosci 33(14) 5895- 650
5902 651
Ihle K Rodriguez-Raecke R amp Luedtke K May A (2014) 652
tDCS modulates cortical nociceptive processing but has little 653
to no impact on pain perception Pain 155(10) 2080-2087 654
Ito M (2006) Cerebellar circuitry as a neuronal machine Prog 655
Neurobiol 78(3-5) 272-303 656
Iyer MB Mattu U Grafman J Lomarev M Sato S amp Wasser- 657
mann EM (2005) Safety and cognitive effect of frontal DC 658
brain polarization in healthy individuals Neurology 64(5) 872- 659
875 660
Kelly RM amp Strick PL (2003) Cerebellar loops with motor cor- 661
tex and prefrontal cortex of a nonhuman primate J Neurosci 662
23(23) 8432-8444 663
Lefaucheur JP Andre-Obadia N Antal A Ayache SS Baeken 664
C Benninger DH Cantello RM Cincotta M de Carvalho 665
M De Ridder D Devanne H Di Lazzaro V Filipovic SR 666
Hummel FC Jaaskelainen SK Kimiskidis VK Koch G 667
Langguth B Nyffeler T Oliviero A Padberg F Poulet E 668
Rossi S Rossini PM Rothwell JC Schonfeldt-Lecuona 669
C Siebner HR Slotema CW Stagg CJ Valls-Sole J 670
Ziemann U Paulus W amp Garcia-Larrea L (2014) Evidence- 671
based guidelines on the therapeutic use of repetitive transcranial 672
magnetic stimulation (rTMS) Clin Neurophysiol 125(11) 673
2150-2206 674
Lefaucheur JP Brusa A Creange A Drouot X amp Jarry G 675
(2002) Clinical application of laser evoked potentials using 676
the NdYAG laser Neurophysiol Clin 32(2) 91-98 677
Lefaucheur JP Debray S amp Jarry G (2001) Laser evoked poten- 678
tials using the NdYAG laser Muscle Nerve 24(4) 496-501 679
Lefaucheur JP Drouot X Menard-Lefaucheur I Keravel Y 680
amp Nguyen JP (2006) Motor cortex rTMS restores defective 681
intracortical inhibition in chronic neuropathic pain Neurology 682
67(9) 1568-1574 683
Longo MR Iannetti GD Mancini F Driver J amp Haggard P 684
(2012) Linking pain and the body Neural correlates of visually 685
induced analgesia J Neurosci 32(8) 2601-2607 686
Lorenz J amp Garcia-Larrea L (2003) Contribution of attentional 687
and cognitive factors to laser evoked brain potentials Neuro- 688
physiol Clin 33(6) 293-301 689
Lorenz J Minoshima S amp Casey KL (2003) Keeping pain out 690
of mind The role of the dorsolateral prefrontal cortex in pain 691
modulation Brain 126(Pt 5) 1079-1091 692
Luedtke K May A amp Jurgens TP (2012) No effect of a single 693
session of transcranial direct current stimulation on experimen- 694
tally induced pain in patients with chronic low back painndashan 695
exploratory study PLoS One 7(11) e48857 696
Unc
orre
cted
Aut
hor P
roof
12 T Bocci et al Cerebellum and pain A tcDCS study
Mendonca ME Santana MB Baptista AF Datta A Bikson697
M Fregni F amp Araujo CP (2011) Transcranial DC stimu-698
lation in fibromyalgia Optimized cortical target supported by699
high-resolution computational models J Pain 12(5) 610-617700
Moriguchi Y Decety J Ohnishi T Maeda M Mori T Nemoto701
K Matsuda H amp Komaki G (2007) Empathy and judg-702
ing otherrsquos pain An fMRI study of alexithymia Cereb Cortex703
17(9) 2223-2234704
Moulton EA Elman I Pendse G Schmahmann J Becerra L705
amp Borsook D (2011) Aversion-related circuitry in the cere-706
bellum Responses to noxious heat and unpleasant images J707
Neurosci 31(10) 3795-3804708
Moulton EA Schmahmann JD Becerra L amp Borsook D709
(2010) The cerebellum and pain Passive integrator or active710
participator Brain Res Rev 65(1) 14-27711
Mouraux A amp Plaghki L (2006) Are the processes reflected by712
late and ultra-late laser evoked potentials specific of nocicep-713
tion Suppl Clin Neurophysiol 59 197-204714
Mylius V Jung M Menzler K Haag A Khader PH Oer-715
tel WH Rosenow F amp Lefaucheur JP (2012) Effects of716
transcranial direct current stimulation on pain perception and717
working memory Eur J Pain 16(7) 974-982718
Ni Z Gunraj C amp Chen R (2007) Short interval intracortical719
inhibition and facilitation during the silent period in human J720
Physiol 583(Pt 3) 971-982721
Nitsche MA Liebetanz D Lang N Antal A Tergau F amp722
Paulus W (2003) Safety criteria for transcranial direct cur-723
rent stimulation (tDCS) in humans Clin Neurophysiol 114(11)724
2220-2222 author reply 2222-2223725
Opsommer E Guerit JM amp Plaghki L (2003) Exogenous726
and endogenous components of ultralate (C-fibre) evoked727
potentials following CO2 laser stimuli to tiny skin sur-728
face areas in healthy subjects Neurophysiol Clin 33(2)729
78-85730
Parazzini M Fiocchi S Liorni I Priori A amp Ravazzani P731
(2014) Computational modeling of transcranial direct current732
stimulation in the child brain Implications for the treatment of733
refractory childhood focal epilepsy Int J Neural Syst 24(2)734
1430006735
Parazzini M Rossi E Ferrucci R Fiocchi S Liorni I Priori736
A amp Ravazzani P (2013) Computational model of cerebellar737
transcranial direct current stimulation Conf Proc IEEE Eng738
Med Biol Soc 2013 237-240739
Parazzini M Rossi E Ferrucci R Liorni I Priori A amp Ravaz-740
zani P (2014) Modelling the electric field and the current741
density generated by cerebellar transcranial DC stimulation in742
humans Clin Neurophysiol 125(3) 577-584743
Perini I Bergstrand S amp Morrison I (2013) Where pain meets744
action in the human brain J Neurosci 33(40) 15930-15939745
Ploghaus A Tracey I Gati JS Clare S Menon RS Matthews746
PM amp Rawlins JN (1999) Dissociating pain from its antic-747
ipation in the human brain Science 284(5422) 1979-1981748
Priori A Ciocca M Parazzini M Vergari M amp Ferrucci R749
(2014) Transcranial cerebellar direct current stimulation and750
transcutaneous spinal cord direct current stimulation as innova-751
tive tools for neuroscientists J Physiol 592(Pt 16) 3345-3369752
Purpura DP amp McMurtry JC (1965) Intracellular activities and 753
evoked potential changes during polarization of motor cortex 754
J Neurophysiol 28 166-185 755
Qiu Y Inui K Wang X Tran TD amp Kakigi R (2002) Effects 756
of attention distraction and sleep on CO(2) laser evoked poten- 757
tials related to C-fibers in humans Clin Neurophysiol 113(10) 758
1579-1585 759
Reidler JS Mendonca ME Santana MB Wang X Lenkinski 760
R Motta AF Marchand S Latif L amp Fregni F (2012) 761
Effects of motor cortex modulation and descending inhibitory 762
systems on pain thresholds in healthy subjects J Pain 13(5) 763
450-458 764
Riedl V Valet M Woller A Sorg C Vogel D Sprenger 765
T Boecker H Wohlschlager AM amp Tolle TR (2011) 766
Repeated pain induces adaptations of intrinsic brain activity 767
to reflect past and predict future pain Neuroimage 57(1) 206- 768
213 769
Rocca MA Valsasina P Absinta M Colombo B Barcella V 770
Falini A Comi G amp Filippi M (2010) Central nervous 771
system dysregulation extends beyond the pain-matrix network 772
in cluster headache Cephalalgia 30(11) 1383-1391 773
Rossi S Hallett M Rossini PM amp Pascual-Leone A (2009) 774
Safety ethical considerations and application guidelines for 775
the use of transcranial magnetic stimulation in clinical practice 776
and research Clin Neurophysiol 120(12) 2008-2039 777
Rossini PM amp Rossi S (2007) Transcranial magnetic stimulation 778
Diagnostic therapeutic and research potential Neurology 779
68(7) 484-488 780
Ruscheweyh R Kuhnel M Filippopulos F Blum B Eggert 781
T amp Straube A (2014) Altered experimental pain perception 782
after cerebellar infarction Pain 155(7) 1303-1312 783
Schmahmann JD (1991) An emerging concept The cerebellar 784
contribution to higher function Arch Neurol 48(11) 1178- 785
1187 786
Singer T Seymour B OrsquoDoherty J Kaube H Dolan RJ amp 787
Frith CD (2004) Empathy for pain involves the affective but 788
not sensory components of pain Science 303(5661) 1157- 789
1162 790
Smith MJ Keel JC Greenberg BD Adams LF Schmidt PJ 791
Rubinow DA amp Wassermann EM (1999) Menstrual cycle 792
effects on cortical excitability Neurology 53(9) 2069-2072 793
Stoodley CJ amp Schmahmann JD (2009) Functional topography 794
in the human cerebellum A meta-analysis of neuroimaging 795
studies Neuroimage 44(2) 489-501 796
Strick PL Dum RP amp Fiez JA (2009) Cerebellum and non- 797
motor function Annu Rev Neurosci 32 413-434 798
Strigo IA Duncan GH Boivin M amp Bushnell MC (2003) 799
Differentiation of visceral and cutaneous pain in the human 800
brain J Neurophysiol 89(6) 3294-3303 801
Truini A Galeotti F Romaniello A Virtuoso M Iannetti GD 802
amp Cruccu G (2005) Laser-evoked potentials Normative val- 803
ues Clin Neurophysiol 116(4) 821-826 804
Truini A Panuccio G Galeotti F Maluccio MR Sartucci F 805
Avoli M amp Cruccu G (2010) Laser-evoked potentials as a 806
tool for assessing the efficacy of antinociceptive drugs Eur J 807
Pain 14(2) 222-225 808
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 13
Truini A Vergari M Biasiotta A La Cesa S Gabriele M Di809
Stefano G Cambieri C Cruccu G Inghilleri M amp Pri-810
ori A (2011) Transcutaneous spinal direct current stimulation811
inhibits nociceptive spinal pathway conduction and increases812
pain tolerance in humans Eur J Pain 15(10) 1023-1027813
Valeriani M Le Pera D Restuccia D de Armas L Mili-814
ucci R Betti V Vigevano F amp Tonali P (2007) Parallel815
spinal pathways generate the middle-latency N1 and the late P2816
components of the laser evoked potentials Clin Neurophysiol817
118(5) 1097-1104818
Valeriani M Pazzaglia C Cruccu G amp Truini A (2012) Clinical819
usefulness of laser evoked potentials Neurophysiol Clin 42(5)820
345-353821
Volz MS Mendonca M Pinheiro FS Cui H Santana M amp822
Fregni F (2012) Dissociation of motor task-induced cortical823
excitability and pain perception changes in healthy volunteers824
PLoS One 7(3) e34273825
Wager TD Rilling JK Smith EE Sokolik A Casey KL 826
Davidson RJ Kosslyn SM Rose RM amp Cohen JD 827
(2004) Placebo-induced changes in FMRI in the anticipation 828
and experience of pain Science 303(5661) 1162-1167 829
Zaghi S Thiele B Pimentel D Pimentel T amp Fregni 830
F (2011) Assessment and treatment of pain with non- 831
invasive cortical stimulation Restor Neurol Neurosci 29(6) 832
439-451 833
Zubieta JK Bueller JA Jackson LR Scott DJ Xu Y 834
Koeppe RA Nichols TE amp Stohler CS (2005) Placebo 835
effects mediated by endogenous opioid activity on mu-opioid 836
receptors J Neurosci 25(34) 7754-7762 837
Zunhammer M Busch V Griesbach F Landgrebe M Hajak G 838
amp Langguth B (2011) rTMS over the cerebellum modulates 839
temperature detection and pain thresholds through peripheral 840
mechanisms Brain Stimul 4(4) 210-7 e1 841

Unc
orre
cted
Aut
hor P
roof
10 T Bocci et al Cerebellum and pain A tcDCS study
tcDCS-induced electrical field is strongly concentrated485
(Parazzini et al 2013) A further limitation is that we486
did not study the contribution of C-fibers the main487
component of spino-ponto-cerebellar and spino-olivo-488
cerebellar pathways In fact ultra-late LEPs related489
to C-fibers activations have not yet been standard-490
ized for clinical application and their occurrence could491
be markedly influenced by high order cognitive pro-492
cesses as they seem to be more affected by the level493
of consciousness and attention than A-delta responses494
(Qiu et al 2002 Opsommer et al 2003 Mouraux amp495
Plaghki 2006) Finally we wish to emphasize that in496
neuropathic patients the effects of the cerebellar stim-497
ulation could be quite different from those described498
here as both anatomical and functional connectivity499
are different from those observed in healthy partici-500
pants (Rocca et al 2010 Riedl et al 2011 Absinta et501
al 2012 Longo et al 2012 Ceko et al 2013)502
5 Conclusions503
Our findings indicate a cerebellar effect on pain504
experience and on its cortical correlates and prompt505
further investigation aimed at assessing whether the506
cerebellar direct current polarization could be used as a507
novel and safe therapeutic tool in chronic pain patients508
Acknowledgments509
We gratefully acknowledge the participation of all510
subjects as well as Mr D Barloscio and Mr C511
Orsini for their excellent technical assistance The512
paper was supported in part by the Italian operating513
and development MIUR PRIN grant year 2006 n514
2006062332 002515
RF and AP are founders and shareholders of516
Newronika srl Milan Italy517
References518
Absinta M Rocca MA Colombo B Falini A Comi G amp Fil-519
ippi M (2012) Selective decreased grey matter volume of the520
pain-matrix network in cluster headache Cephalalgia 32(2)521
109-115522
Agostino R Cruccu G Iannetti G Romaniello A Truini A523
amp Manfredi M (2000) Topographical distribution of pinprick524
and warmth thresholds to CO2 laser stimulation on the human525
skin Neurosci Lett 285(2) 115-118526
Antal A Brepohl N Poreisz C Boros K Csifcsak G amp 527
Paulus W (2008) Transcranial direct current stimulation over 528
somatosensory cortex decreases experimentally induced acute 529
pain perception Clin J Pain 24(1) 56-63 530
Antal A amp Paulus W (2010) Effects of transcranial theta-burst 531
stimulation on acute pain perception Restor Neurol Neurosci 532
28(4) 477-484 533
Apkarian AV Bushnell MC Treede RD amp Zubieta JK 534
(2005) Human brain mechanisms of pain perception and reg- 535
ulation in health and disease Eur J Pain 9(4) 463-484 536
Balsters JH Whelan CD Robertson IH amp Ramnani N (2013) 537
Cerebellum and cognition Evidence for the encoding of higher 538
order rules Cereb Cortex 23(6) 1433-1443 539
Battaglia F Quartarone A Ghilardi MF Dattola R Bagnato 540
S Rizzo V Morgante L amp Girlanda P (2006) Unilat- 541
eral cerebellar stroke disrupts movement preparation and motor 542
imagery Clin Neurophysiol 117(5) 1009-1016 543
Bingel U Quante M Knab R Bromm B Weiller C amp Buchel 544
C (2002) Subcortical structures involved in pain processing 545
Evidence from single-trial fMRI Pain 99(1-2) 313-321 546
Boggio PS Zaghi S amp Fregni F (2009) Modulation of emotions 547
associated with images of human pain using anodal transcranial 548
direct current stimulation (tDCS) Neuropsychologia 47(1) 549
212-217 550
Boggio PS Zaghi S Lopes M amp Fregni F (2008) Modulatory 551
effects of anodal transcranial direct current stimulation on per- 552
ception and pain thresholds in healthy volunteers Eur J Neurol 553
15(10) 1124-1130 554
Borsook D Moulton EA Tully S Schmahmann JD amp 555
Becerra L (2008) Human cerebellar responses to brush and 556
heat stimuli in healthy and neuropathic pain subjects Cerebel- 557
lum 7(3) 252-272 558
Ceko M Seminowicz DA Bushnell MC amp Olausson HW 559
(2013) Anatomical and functional enhancements of the insula 560
after loss of large primary somatosensory fibers Cereb Cortex 561
23(9) 2017-2024 562
Cruccu G Aminoff MJ Curio G Guerit JM Kakigi 563
R Mauguiere F Rossini PM Treede RD amp Garcia- 564
Larrea L (2008) Recommendations for the clinical use of 565
somatosensory-evoked potentials Clin Neurophysiol 119(8) 566
1705-1719 567
Cruccu G Romaniello A Amantini A Lombardi M Innocenti 568
P amp Manfredi M (1999) Assessment of trigeminal small- 569
fiber function Brain and reflex responses evoked by CO2-laser 570
stimulation Muscle Nerve 22(4) 508-516 571
Dasilva AF Mendonca ME Zaghi S Lopes M Dossantos 572
MF Spierings EL Bajwa Z Datta A Bikson M amp 573
Fregni F (2012) tDCS-induced analgesia and electrical fields 574
in pain-related neural networks in chronic migraine Headache 575
52(8) 1283-1295 576
Duerden EG amp Albanese MC (2013) Localization of pain- 577
related brain activation A meta-analysis of neuroimaging data 578
Hum Brain Mapp 34(1) 109-149 579
Ekerot CF Garwicz M amp Schouenborg J (1991a) The postsy- 580
naptic dorsal column pathway mediates cutaneous nociceptive 581
information to cerebellar climbing fibres in the cat J Physiol 582
441 275-284 583
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 11
Ekerot CF Garwicz M amp Schouenborg J (1991b) Topography584
and nociceptive receptive fields of climbing fibres projecting to585
the cerebellar anterior lobe in the cat J Physiol 441 257-274586
Ekerot CF Gustavsson P Oscarsson O amp Schouenborg J587
(1987a) Climbing fibres projecting to cat cerebellar anterior588
lobe activated by cutaneous A and C fibres J Physiol 386589
529-538590
Ekerot CF Oscarsson O amp Schouenborg J (1987b) Stimula-591
tion of cat cutaneous nociceptive C fibres causing tonic and592
synchronous activity in climbing fibres J Physiol 386 539-593
546594
Ferrucci R Brunoni AR Parazzini M Vergari M Rossi E595
Fumagalli M Mameli F Rosa M Giannicola G Zago S596
amp Priori A (2013) Modulating human procedural learning by597
cerebellar transcranial direct current stimulation Cerebellum598
12(4) 485-492599
Ferrucci R Giannicola G Rosa M Fumagalli M Boggio600
PS Hallett M Zago S amp Priori A (2012) Cerebellum601
and processing of negative facial emotions Cerebellar tran-602
scranial DC stimulation specifically enhances the emotional603
recognition of facial anger and sadness Cogn Emot 26(5)604
786-799605
Ferrucci R Marceglia S Vergari M Cogiamanian F Mrakic-606
Sposta S Mameli F Zago S Barbieri S amp Priori607
A (2008) Cerebellar transcranial direct current stimulation608
impairs the practice-dependent proficiency increase in working609
memory J Cogn Neurosci 20(9) 1687-1697610
Fregni F Freedman S amp Pascual-Leone A (2007) Recent611
advances in the treatment of chronic pain with non-invasive612
brain stimulation techniques Lancet Neurol 6(2) 188-191613
Galea JM Jayaram G Ajagbe L amp Celnik P (2009)614
Modulation of cerebellar excitability by polarity-specific615
noninvasive direct current stimulation J Neurosci 29(28)616
9115-9122617
Galea JM Vazquez A Pasricha N de Xivry JJ amp Celnik618
P (2011) Dissociating the roles of the cerebellum and motor619
cortex during adaptive learning The motor cortex retains what620
the cerebellum learns Cereb Cortex 21(8) 1761-1770621
Gandiga PC Hummel FC amp Cohen LG (2006) Tran-622
scranial DC stimulation (tDCS) A tool for double-blind623
sham-controlled clinical studies in brain stimulation Clin Neu-624
rophysiol 117(4) 845-850625
Garcia-Larrea L Peyron R Laurent B amp Mauguiere F (1997)626
Association and dissociation between laser-evoked potentials627
and pain perception Neuroreport 8(17) 3785-3789628
Grimaldi G Argyropoulos GP Boehringer A Celnik P629
Edwards MJ Ferrucci R Galea JM Groiss SJ Hiraoka630
K Kassavetis P Lesage E Manto M Miall RC Priori A631
Sadnicka A Ugawa Y amp Ziemann U (2014) Non-invasive632
cerebellar stimulationndasha consensus paper Cerebellum 13(1)633
121-138634
Groppa S Oliviero A Eisen A Quartarone A Cohen LG635
Mall V Kaelin-Lang A Mima T Rossi S Thickbroom636
GW Rossini PM Ziemann U Valls-Sole J amp Sieb-637
ner HR (2012) A practical guide to diagnostic transcranial638
magnetic stimulation Report of an IFCN committee Clin Neu-639
rophysiol 123(5) 858-882640
Hamada M Strigaro G Murase N Sadnicka A Galea JM 641
Edwards MJ amp Rothwell JC (2012) Cerebellar modulation 642
of human associative plasticity J Physiol 590(Pt 10) 2365- 643
2374 644
Hess CW Mills KR amp Murray NM (1987) Responses in small 645
hand muscles from magnetic stimulation of the human brain J 646
Physiol 388 397-419 647
Hull CA Chu Y Thanawala M amp Regehr WG (2013) Hyper- 648
polarization induces a long-term increase in the spontaneous 649
firing rate of cerebellar Golgi cells J Neurosci 33(14) 5895- 650
5902 651
Ihle K Rodriguez-Raecke R amp Luedtke K May A (2014) 652
tDCS modulates cortical nociceptive processing but has little 653
to no impact on pain perception Pain 155(10) 2080-2087 654
Ito M (2006) Cerebellar circuitry as a neuronal machine Prog 655
Neurobiol 78(3-5) 272-303 656
Iyer MB Mattu U Grafman J Lomarev M Sato S amp Wasser- 657
mann EM (2005) Safety and cognitive effect of frontal DC 658
brain polarization in healthy individuals Neurology 64(5) 872- 659
875 660
Kelly RM amp Strick PL (2003) Cerebellar loops with motor cor- 661
tex and prefrontal cortex of a nonhuman primate J Neurosci 662
23(23) 8432-8444 663
Lefaucheur JP Andre-Obadia N Antal A Ayache SS Baeken 664
C Benninger DH Cantello RM Cincotta M de Carvalho 665
M De Ridder D Devanne H Di Lazzaro V Filipovic SR 666
Hummel FC Jaaskelainen SK Kimiskidis VK Koch G 667
Langguth B Nyffeler T Oliviero A Padberg F Poulet E 668
Rossi S Rossini PM Rothwell JC Schonfeldt-Lecuona 669
C Siebner HR Slotema CW Stagg CJ Valls-Sole J 670
Ziemann U Paulus W amp Garcia-Larrea L (2014) Evidence- 671
based guidelines on the therapeutic use of repetitive transcranial 672
magnetic stimulation (rTMS) Clin Neurophysiol 125(11) 673
2150-2206 674
Lefaucheur JP Brusa A Creange A Drouot X amp Jarry G 675
(2002) Clinical application of laser evoked potentials using 676
the NdYAG laser Neurophysiol Clin 32(2) 91-98 677
Lefaucheur JP Debray S amp Jarry G (2001) Laser evoked poten- 678
tials using the NdYAG laser Muscle Nerve 24(4) 496-501 679
Lefaucheur JP Drouot X Menard-Lefaucheur I Keravel Y 680
amp Nguyen JP (2006) Motor cortex rTMS restores defective 681
intracortical inhibition in chronic neuropathic pain Neurology 682
67(9) 1568-1574 683
Longo MR Iannetti GD Mancini F Driver J amp Haggard P 684
(2012) Linking pain and the body Neural correlates of visually 685
induced analgesia J Neurosci 32(8) 2601-2607 686
Lorenz J amp Garcia-Larrea L (2003) Contribution of attentional 687
and cognitive factors to laser evoked brain potentials Neuro- 688
physiol Clin 33(6) 293-301 689
Lorenz J Minoshima S amp Casey KL (2003) Keeping pain out 690
of mind The role of the dorsolateral prefrontal cortex in pain 691
modulation Brain 126(Pt 5) 1079-1091 692
Luedtke K May A amp Jurgens TP (2012) No effect of a single 693
session of transcranial direct current stimulation on experimen- 694
tally induced pain in patients with chronic low back painndashan 695
exploratory study PLoS One 7(11) e48857 696
Unc
orre
cted
Aut
hor P
roof
12 T Bocci et al Cerebellum and pain A tcDCS study
Mendonca ME Santana MB Baptista AF Datta A Bikson697
M Fregni F amp Araujo CP (2011) Transcranial DC stimu-698
lation in fibromyalgia Optimized cortical target supported by699
high-resolution computational models J Pain 12(5) 610-617700
Moriguchi Y Decety J Ohnishi T Maeda M Mori T Nemoto701
K Matsuda H amp Komaki G (2007) Empathy and judg-702
ing otherrsquos pain An fMRI study of alexithymia Cereb Cortex703
17(9) 2223-2234704
Moulton EA Elman I Pendse G Schmahmann J Becerra L705
amp Borsook D (2011) Aversion-related circuitry in the cere-706
bellum Responses to noxious heat and unpleasant images J707
Neurosci 31(10) 3795-3804708
Moulton EA Schmahmann JD Becerra L amp Borsook D709
(2010) The cerebellum and pain Passive integrator or active710
participator Brain Res Rev 65(1) 14-27711
Mouraux A amp Plaghki L (2006) Are the processes reflected by712
late and ultra-late laser evoked potentials specific of nocicep-713
tion Suppl Clin Neurophysiol 59 197-204714
Mylius V Jung M Menzler K Haag A Khader PH Oer-715
tel WH Rosenow F amp Lefaucheur JP (2012) Effects of716
transcranial direct current stimulation on pain perception and717
working memory Eur J Pain 16(7) 974-982718
Ni Z Gunraj C amp Chen R (2007) Short interval intracortical719
inhibition and facilitation during the silent period in human J720
Physiol 583(Pt 3) 971-982721
Nitsche MA Liebetanz D Lang N Antal A Tergau F amp722
Paulus W (2003) Safety criteria for transcranial direct cur-723
rent stimulation (tDCS) in humans Clin Neurophysiol 114(11)724
2220-2222 author reply 2222-2223725
Opsommer E Guerit JM amp Plaghki L (2003) Exogenous726
and endogenous components of ultralate (C-fibre) evoked727
potentials following CO2 laser stimuli to tiny skin sur-728
face areas in healthy subjects Neurophysiol Clin 33(2)729
78-85730
Parazzini M Fiocchi S Liorni I Priori A amp Ravazzani P731
(2014) Computational modeling of transcranial direct current732
stimulation in the child brain Implications for the treatment of733
refractory childhood focal epilepsy Int J Neural Syst 24(2)734
1430006735
Parazzini M Rossi E Ferrucci R Fiocchi S Liorni I Priori736
A amp Ravazzani P (2013) Computational model of cerebellar737
transcranial direct current stimulation Conf Proc IEEE Eng738
Med Biol Soc 2013 237-240739
Parazzini M Rossi E Ferrucci R Liorni I Priori A amp Ravaz-740
zani P (2014) Modelling the electric field and the current741
density generated by cerebellar transcranial DC stimulation in742
humans Clin Neurophysiol 125(3) 577-584743
Perini I Bergstrand S amp Morrison I (2013) Where pain meets744
action in the human brain J Neurosci 33(40) 15930-15939745
Ploghaus A Tracey I Gati JS Clare S Menon RS Matthews746
PM amp Rawlins JN (1999) Dissociating pain from its antic-747
ipation in the human brain Science 284(5422) 1979-1981748
Priori A Ciocca M Parazzini M Vergari M amp Ferrucci R749
(2014) Transcranial cerebellar direct current stimulation and750
transcutaneous spinal cord direct current stimulation as innova-751
tive tools for neuroscientists J Physiol 592(Pt 16) 3345-3369752
Purpura DP amp McMurtry JC (1965) Intracellular activities and 753
evoked potential changes during polarization of motor cortex 754
J Neurophysiol 28 166-185 755
Qiu Y Inui K Wang X Tran TD amp Kakigi R (2002) Effects 756
of attention distraction and sleep on CO(2) laser evoked poten- 757
tials related to C-fibers in humans Clin Neurophysiol 113(10) 758
1579-1585 759
Reidler JS Mendonca ME Santana MB Wang X Lenkinski 760
R Motta AF Marchand S Latif L amp Fregni F (2012) 761
Effects of motor cortex modulation and descending inhibitory 762
systems on pain thresholds in healthy subjects J Pain 13(5) 763
450-458 764
Riedl V Valet M Woller A Sorg C Vogel D Sprenger 765
T Boecker H Wohlschlager AM amp Tolle TR (2011) 766
Repeated pain induces adaptations of intrinsic brain activity 767
to reflect past and predict future pain Neuroimage 57(1) 206- 768
213 769
Rocca MA Valsasina P Absinta M Colombo B Barcella V 770
Falini A Comi G amp Filippi M (2010) Central nervous 771
system dysregulation extends beyond the pain-matrix network 772
in cluster headache Cephalalgia 30(11) 1383-1391 773
Rossi S Hallett M Rossini PM amp Pascual-Leone A (2009) 774
Safety ethical considerations and application guidelines for 775
the use of transcranial magnetic stimulation in clinical practice 776
and research Clin Neurophysiol 120(12) 2008-2039 777
Rossini PM amp Rossi S (2007) Transcranial magnetic stimulation 778
Diagnostic therapeutic and research potential Neurology 779
68(7) 484-488 780
Ruscheweyh R Kuhnel M Filippopulos F Blum B Eggert 781
T amp Straube A (2014) Altered experimental pain perception 782
after cerebellar infarction Pain 155(7) 1303-1312 783
Schmahmann JD (1991) An emerging concept The cerebellar 784
contribution to higher function Arch Neurol 48(11) 1178- 785
1187 786
Singer T Seymour B OrsquoDoherty J Kaube H Dolan RJ amp 787
Frith CD (2004) Empathy for pain involves the affective but 788
not sensory components of pain Science 303(5661) 1157- 789
1162 790
Smith MJ Keel JC Greenberg BD Adams LF Schmidt PJ 791
Rubinow DA amp Wassermann EM (1999) Menstrual cycle 792
effects on cortical excitability Neurology 53(9) 2069-2072 793
Stoodley CJ amp Schmahmann JD (2009) Functional topography 794
in the human cerebellum A meta-analysis of neuroimaging 795
studies Neuroimage 44(2) 489-501 796
Strick PL Dum RP amp Fiez JA (2009) Cerebellum and non- 797
motor function Annu Rev Neurosci 32 413-434 798
Strigo IA Duncan GH Boivin M amp Bushnell MC (2003) 799
Differentiation of visceral and cutaneous pain in the human 800
brain J Neurophysiol 89(6) 3294-3303 801
Truini A Galeotti F Romaniello A Virtuoso M Iannetti GD 802
amp Cruccu G (2005) Laser-evoked potentials Normative val- 803
ues Clin Neurophysiol 116(4) 821-826 804
Truini A Panuccio G Galeotti F Maluccio MR Sartucci F 805
Avoli M amp Cruccu G (2010) Laser-evoked potentials as a 806
tool for assessing the efficacy of antinociceptive drugs Eur J 807
Pain 14(2) 222-225 808
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 13
Truini A Vergari M Biasiotta A La Cesa S Gabriele M Di809
Stefano G Cambieri C Cruccu G Inghilleri M amp Pri-810
ori A (2011) Transcutaneous spinal direct current stimulation811
inhibits nociceptive spinal pathway conduction and increases812
pain tolerance in humans Eur J Pain 15(10) 1023-1027813
Valeriani M Le Pera D Restuccia D de Armas L Mili-814
ucci R Betti V Vigevano F amp Tonali P (2007) Parallel815
spinal pathways generate the middle-latency N1 and the late P2816
components of the laser evoked potentials Clin Neurophysiol817
118(5) 1097-1104818
Valeriani M Pazzaglia C Cruccu G amp Truini A (2012) Clinical819
usefulness of laser evoked potentials Neurophysiol Clin 42(5)820
345-353821
Volz MS Mendonca M Pinheiro FS Cui H Santana M amp822
Fregni F (2012) Dissociation of motor task-induced cortical823
excitability and pain perception changes in healthy volunteers824
PLoS One 7(3) e34273825
Wager TD Rilling JK Smith EE Sokolik A Casey KL 826
Davidson RJ Kosslyn SM Rose RM amp Cohen JD 827
(2004) Placebo-induced changes in FMRI in the anticipation 828
and experience of pain Science 303(5661) 1162-1167 829
Zaghi S Thiele B Pimentel D Pimentel T amp Fregni 830
F (2011) Assessment and treatment of pain with non- 831
invasive cortical stimulation Restor Neurol Neurosci 29(6) 832
439-451 833
Zubieta JK Bueller JA Jackson LR Scott DJ Xu Y 834
Koeppe RA Nichols TE amp Stohler CS (2005) Placebo 835
effects mediated by endogenous opioid activity on mu-opioid 836
receptors J Neurosci 25(34) 7754-7762 837
Zunhammer M Busch V Griesbach F Landgrebe M Hajak G 838
amp Langguth B (2011) rTMS over the cerebellum modulates 839
temperature detection and pain thresholds through peripheral 840
mechanisms Brain Stimul 4(4) 210-7 e1 841

Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 11
Ekerot CF Garwicz M amp Schouenborg J (1991b) Topography584
and nociceptive receptive fields of climbing fibres projecting to585
the cerebellar anterior lobe in the cat J Physiol 441 257-274586
Ekerot CF Gustavsson P Oscarsson O amp Schouenborg J587
(1987a) Climbing fibres projecting to cat cerebellar anterior588
lobe activated by cutaneous A and C fibres J Physiol 386589
529-538590
Ekerot CF Oscarsson O amp Schouenborg J (1987b) Stimula-591
tion of cat cutaneous nociceptive C fibres causing tonic and592
synchronous activity in climbing fibres J Physiol 386 539-593
546594
Ferrucci R Brunoni AR Parazzini M Vergari M Rossi E595
Fumagalli M Mameli F Rosa M Giannicola G Zago S596
amp Priori A (2013) Modulating human procedural learning by597
cerebellar transcranial direct current stimulation Cerebellum598
12(4) 485-492599
Ferrucci R Giannicola G Rosa M Fumagalli M Boggio600
PS Hallett M Zago S amp Priori A (2012) Cerebellum601
and processing of negative facial emotions Cerebellar tran-602
scranial DC stimulation specifically enhances the emotional603
recognition of facial anger and sadness Cogn Emot 26(5)604
786-799605
Ferrucci R Marceglia S Vergari M Cogiamanian F Mrakic-606
Sposta S Mameli F Zago S Barbieri S amp Priori607
A (2008) Cerebellar transcranial direct current stimulation608
impairs the practice-dependent proficiency increase in working609
memory J Cogn Neurosci 20(9) 1687-1697610
Fregni F Freedman S amp Pascual-Leone A (2007) Recent611
advances in the treatment of chronic pain with non-invasive612
brain stimulation techniques Lancet Neurol 6(2) 188-191613
Galea JM Jayaram G Ajagbe L amp Celnik P (2009)614
Modulation of cerebellar excitability by polarity-specific615
noninvasive direct current stimulation J Neurosci 29(28)616
9115-9122617
Galea JM Vazquez A Pasricha N de Xivry JJ amp Celnik618
P (2011) Dissociating the roles of the cerebellum and motor619
cortex during adaptive learning The motor cortex retains what620
the cerebellum learns Cereb Cortex 21(8) 1761-1770621
Gandiga PC Hummel FC amp Cohen LG (2006) Tran-622
scranial DC stimulation (tDCS) A tool for double-blind623
sham-controlled clinical studies in brain stimulation Clin Neu-624
rophysiol 117(4) 845-850625
Garcia-Larrea L Peyron R Laurent B amp Mauguiere F (1997)626
Association and dissociation between laser-evoked potentials627
and pain perception Neuroreport 8(17) 3785-3789628
Grimaldi G Argyropoulos GP Boehringer A Celnik P629
Edwards MJ Ferrucci R Galea JM Groiss SJ Hiraoka630
K Kassavetis P Lesage E Manto M Miall RC Priori A631
Sadnicka A Ugawa Y amp Ziemann U (2014) Non-invasive632
cerebellar stimulationndasha consensus paper Cerebellum 13(1)633
121-138634
Groppa S Oliviero A Eisen A Quartarone A Cohen LG635
Mall V Kaelin-Lang A Mima T Rossi S Thickbroom636
GW Rossini PM Ziemann U Valls-Sole J amp Sieb-637
ner HR (2012) A practical guide to diagnostic transcranial638
magnetic stimulation Report of an IFCN committee Clin Neu-639
rophysiol 123(5) 858-882640
Hamada M Strigaro G Murase N Sadnicka A Galea JM 641
Edwards MJ amp Rothwell JC (2012) Cerebellar modulation 642
of human associative plasticity J Physiol 590(Pt 10) 2365- 643
2374 644
Hess CW Mills KR amp Murray NM (1987) Responses in small 645
hand muscles from magnetic stimulation of the human brain J 646
Physiol 388 397-419 647
Hull CA Chu Y Thanawala M amp Regehr WG (2013) Hyper- 648
polarization induces a long-term increase in the spontaneous 649
firing rate of cerebellar Golgi cells J Neurosci 33(14) 5895- 650
5902 651
Ihle K Rodriguez-Raecke R amp Luedtke K May A (2014) 652
tDCS modulates cortical nociceptive processing but has little 653
to no impact on pain perception Pain 155(10) 2080-2087 654
Ito M (2006) Cerebellar circuitry as a neuronal machine Prog 655
Neurobiol 78(3-5) 272-303 656
Iyer MB Mattu U Grafman J Lomarev M Sato S amp Wasser- 657
mann EM (2005) Safety and cognitive effect of frontal DC 658
brain polarization in healthy individuals Neurology 64(5) 872- 659
875 660
Kelly RM amp Strick PL (2003) Cerebellar loops with motor cor- 661
tex and prefrontal cortex of a nonhuman primate J Neurosci 662
23(23) 8432-8444 663
Lefaucheur JP Andre-Obadia N Antal A Ayache SS Baeken 664
C Benninger DH Cantello RM Cincotta M de Carvalho 665
M De Ridder D Devanne H Di Lazzaro V Filipovic SR 666
Hummel FC Jaaskelainen SK Kimiskidis VK Koch G 667
Langguth B Nyffeler T Oliviero A Padberg F Poulet E 668
Rossi S Rossini PM Rothwell JC Schonfeldt-Lecuona 669
C Siebner HR Slotema CW Stagg CJ Valls-Sole J 670
Ziemann U Paulus W amp Garcia-Larrea L (2014) Evidence- 671
based guidelines on the therapeutic use of repetitive transcranial 672
magnetic stimulation (rTMS) Clin Neurophysiol 125(11) 673
2150-2206 674
Lefaucheur JP Brusa A Creange A Drouot X amp Jarry G 675
(2002) Clinical application of laser evoked potentials using 676
the NdYAG laser Neurophysiol Clin 32(2) 91-98 677
Lefaucheur JP Debray S amp Jarry G (2001) Laser evoked poten- 678
tials using the NdYAG laser Muscle Nerve 24(4) 496-501 679
Lefaucheur JP Drouot X Menard-Lefaucheur I Keravel Y 680
amp Nguyen JP (2006) Motor cortex rTMS restores defective 681
intracortical inhibition in chronic neuropathic pain Neurology 682
67(9) 1568-1574 683
Longo MR Iannetti GD Mancini F Driver J amp Haggard P 684
(2012) Linking pain and the body Neural correlates of visually 685
induced analgesia J Neurosci 32(8) 2601-2607 686
Lorenz J amp Garcia-Larrea L (2003) Contribution of attentional 687
and cognitive factors to laser evoked brain potentials Neuro- 688
physiol Clin 33(6) 293-301 689
Lorenz J Minoshima S amp Casey KL (2003) Keeping pain out 690
of mind The role of the dorsolateral prefrontal cortex in pain 691
modulation Brain 126(Pt 5) 1079-1091 692
Luedtke K May A amp Jurgens TP (2012) No effect of a single 693
session of transcranial direct current stimulation on experimen- 694
tally induced pain in patients with chronic low back painndashan 695
exploratory study PLoS One 7(11) e48857 696
Unc
orre
cted
Aut
hor P
roof
12 T Bocci et al Cerebellum and pain A tcDCS study
Mendonca ME Santana MB Baptista AF Datta A Bikson697
M Fregni F amp Araujo CP (2011) Transcranial DC stimu-698
lation in fibromyalgia Optimized cortical target supported by699
high-resolution computational models J Pain 12(5) 610-617700
Moriguchi Y Decety J Ohnishi T Maeda M Mori T Nemoto701
K Matsuda H amp Komaki G (2007) Empathy and judg-702
ing otherrsquos pain An fMRI study of alexithymia Cereb Cortex703
17(9) 2223-2234704
Moulton EA Elman I Pendse G Schmahmann J Becerra L705
amp Borsook D (2011) Aversion-related circuitry in the cere-706
bellum Responses to noxious heat and unpleasant images J707
Neurosci 31(10) 3795-3804708
Moulton EA Schmahmann JD Becerra L amp Borsook D709
(2010) The cerebellum and pain Passive integrator or active710
participator Brain Res Rev 65(1) 14-27711
Mouraux A amp Plaghki L (2006) Are the processes reflected by712
late and ultra-late laser evoked potentials specific of nocicep-713
tion Suppl Clin Neurophysiol 59 197-204714
Mylius V Jung M Menzler K Haag A Khader PH Oer-715
tel WH Rosenow F amp Lefaucheur JP (2012) Effects of716
transcranial direct current stimulation on pain perception and717
working memory Eur J Pain 16(7) 974-982718
Ni Z Gunraj C amp Chen R (2007) Short interval intracortical719
inhibition and facilitation during the silent period in human J720
Physiol 583(Pt 3) 971-982721
Nitsche MA Liebetanz D Lang N Antal A Tergau F amp722
Paulus W (2003) Safety criteria for transcranial direct cur-723
rent stimulation (tDCS) in humans Clin Neurophysiol 114(11)724
2220-2222 author reply 2222-2223725
Opsommer E Guerit JM amp Plaghki L (2003) Exogenous726
and endogenous components of ultralate (C-fibre) evoked727
potentials following CO2 laser stimuli to tiny skin sur-728
face areas in healthy subjects Neurophysiol Clin 33(2)729
78-85730
Parazzini M Fiocchi S Liorni I Priori A amp Ravazzani P731
(2014) Computational modeling of transcranial direct current732
stimulation in the child brain Implications for the treatment of733
refractory childhood focal epilepsy Int J Neural Syst 24(2)734
1430006735
Parazzini M Rossi E Ferrucci R Fiocchi S Liorni I Priori736
A amp Ravazzani P (2013) Computational model of cerebellar737
transcranial direct current stimulation Conf Proc IEEE Eng738
Med Biol Soc 2013 237-240739
Parazzini M Rossi E Ferrucci R Liorni I Priori A amp Ravaz-740
zani P (2014) Modelling the electric field and the current741
density generated by cerebellar transcranial DC stimulation in742
humans Clin Neurophysiol 125(3) 577-584743
Perini I Bergstrand S amp Morrison I (2013) Where pain meets744
action in the human brain J Neurosci 33(40) 15930-15939745
Ploghaus A Tracey I Gati JS Clare S Menon RS Matthews746
PM amp Rawlins JN (1999) Dissociating pain from its antic-747
ipation in the human brain Science 284(5422) 1979-1981748
Priori A Ciocca M Parazzini M Vergari M amp Ferrucci R749
(2014) Transcranial cerebellar direct current stimulation and750
transcutaneous spinal cord direct current stimulation as innova-751
tive tools for neuroscientists J Physiol 592(Pt 16) 3345-3369752
Purpura DP amp McMurtry JC (1965) Intracellular activities and 753
evoked potential changes during polarization of motor cortex 754
J Neurophysiol 28 166-185 755
Qiu Y Inui K Wang X Tran TD amp Kakigi R (2002) Effects 756
of attention distraction and sleep on CO(2) laser evoked poten- 757
tials related to C-fibers in humans Clin Neurophysiol 113(10) 758
1579-1585 759
Reidler JS Mendonca ME Santana MB Wang X Lenkinski 760
R Motta AF Marchand S Latif L amp Fregni F (2012) 761
Effects of motor cortex modulation and descending inhibitory 762
systems on pain thresholds in healthy subjects J Pain 13(5) 763
450-458 764
Riedl V Valet M Woller A Sorg C Vogel D Sprenger 765
T Boecker H Wohlschlager AM amp Tolle TR (2011) 766
Repeated pain induces adaptations of intrinsic brain activity 767
to reflect past and predict future pain Neuroimage 57(1) 206- 768
213 769
Rocca MA Valsasina P Absinta M Colombo B Barcella V 770
Falini A Comi G amp Filippi M (2010) Central nervous 771
system dysregulation extends beyond the pain-matrix network 772
in cluster headache Cephalalgia 30(11) 1383-1391 773
Rossi S Hallett M Rossini PM amp Pascual-Leone A (2009) 774
Safety ethical considerations and application guidelines for 775
the use of transcranial magnetic stimulation in clinical practice 776
and research Clin Neurophysiol 120(12) 2008-2039 777
Rossini PM amp Rossi S (2007) Transcranial magnetic stimulation 778
Diagnostic therapeutic and research potential Neurology 779
68(7) 484-488 780
Ruscheweyh R Kuhnel M Filippopulos F Blum B Eggert 781
T amp Straube A (2014) Altered experimental pain perception 782
after cerebellar infarction Pain 155(7) 1303-1312 783
Schmahmann JD (1991) An emerging concept The cerebellar 784
contribution to higher function Arch Neurol 48(11) 1178- 785
1187 786
Singer T Seymour B OrsquoDoherty J Kaube H Dolan RJ amp 787
Frith CD (2004) Empathy for pain involves the affective but 788
not sensory components of pain Science 303(5661) 1157- 789
1162 790
Smith MJ Keel JC Greenberg BD Adams LF Schmidt PJ 791
Rubinow DA amp Wassermann EM (1999) Menstrual cycle 792
effects on cortical excitability Neurology 53(9) 2069-2072 793
Stoodley CJ amp Schmahmann JD (2009) Functional topography 794
in the human cerebellum A meta-analysis of neuroimaging 795
studies Neuroimage 44(2) 489-501 796
Strick PL Dum RP amp Fiez JA (2009) Cerebellum and non- 797
motor function Annu Rev Neurosci 32 413-434 798
Strigo IA Duncan GH Boivin M amp Bushnell MC (2003) 799
Differentiation of visceral and cutaneous pain in the human 800
brain J Neurophysiol 89(6) 3294-3303 801
Truini A Galeotti F Romaniello A Virtuoso M Iannetti GD 802
amp Cruccu G (2005) Laser-evoked potentials Normative val- 803
ues Clin Neurophysiol 116(4) 821-826 804
Truini A Panuccio G Galeotti F Maluccio MR Sartucci F 805
Avoli M amp Cruccu G (2010) Laser-evoked potentials as a 806
tool for assessing the efficacy of antinociceptive drugs Eur J 807
Pain 14(2) 222-225 808
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 13
Truini A Vergari M Biasiotta A La Cesa S Gabriele M Di809
Stefano G Cambieri C Cruccu G Inghilleri M amp Pri-810
ori A (2011) Transcutaneous spinal direct current stimulation811
inhibits nociceptive spinal pathway conduction and increases812
pain tolerance in humans Eur J Pain 15(10) 1023-1027813
Valeriani M Le Pera D Restuccia D de Armas L Mili-814
ucci R Betti V Vigevano F amp Tonali P (2007) Parallel815
spinal pathways generate the middle-latency N1 and the late P2816
components of the laser evoked potentials Clin Neurophysiol817
118(5) 1097-1104818
Valeriani M Pazzaglia C Cruccu G amp Truini A (2012) Clinical819
usefulness of laser evoked potentials Neurophysiol Clin 42(5)820
345-353821
Volz MS Mendonca M Pinheiro FS Cui H Santana M amp822
Fregni F (2012) Dissociation of motor task-induced cortical823
excitability and pain perception changes in healthy volunteers824
PLoS One 7(3) e34273825
Wager TD Rilling JK Smith EE Sokolik A Casey KL 826
Davidson RJ Kosslyn SM Rose RM amp Cohen JD 827
(2004) Placebo-induced changes in FMRI in the anticipation 828
and experience of pain Science 303(5661) 1162-1167 829
Zaghi S Thiele B Pimentel D Pimentel T amp Fregni 830
F (2011) Assessment and treatment of pain with non- 831
invasive cortical stimulation Restor Neurol Neurosci 29(6) 832
439-451 833
Zubieta JK Bueller JA Jackson LR Scott DJ Xu Y 834
Koeppe RA Nichols TE amp Stohler CS (2005) Placebo 835
effects mediated by endogenous opioid activity on mu-opioid 836
receptors J Neurosci 25(34) 7754-7762 837
Zunhammer M Busch V Griesbach F Landgrebe M Hajak G 838
amp Langguth B (2011) rTMS over the cerebellum modulates 839
temperature detection and pain thresholds through peripheral 840
mechanisms Brain Stimul 4(4) 210-7 e1 841

Unc
orre
cted
Aut
hor P
roof
12 T Bocci et al Cerebellum and pain A tcDCS study
Mendonca ME Santana MB Baptista AF Datta A Bikson697
M Fregni F amp Araujo CP (2011) Transcranial DC stimu-698
lation in fibromyalgia Optimized cortical target supported by699
high-resolution computational models J Pain 12(5) 610-617700
Moriguchi Y Decety J Ohnishi T Maeda M Mori T Nemoto701
K Matsuda H amp Komaki G (2007) Empathy and judg-702
ing otherrsquos pain An fMRI study of alexithymia Cereb Cortex703
17(9) 2223-2234704
Moulton EA Elman I Pendse G Schmahmann J Becerra L705
amp Borsook D (2011) Aversion-related circuitry in the cere-706
bellum Responses to noxious heat and unpleasant images J707
Neurosci 31(10) 3795-3804708
Moulton EA Schmahmann JD Becerra L amp Borsook D709
(2010) The cerebellum and pain Passive integrator or active710
participator Brain Res Rev 65(1) 14-27711
Mouraux A amp Plaghki L (2006) Are the processes reflected by712
late and ultra-late laser evoked potentials specific of nocicep-713
tion Suppl Clin Neurophysiol 59 197-204714
Mylius V Jung M Menzler K Haag A Khader PH Oer-715
tel WH Rosenow F amp Lefaucheur JP (2012) Effects of716
transcranial direct current stimulation on pain perception and717
working memory Eur J Pain 16(7) 974-982718
Ni Z Gunraj C amp Chen R (2007) Short interval intracortical719
inhibition and facilitation during the silent period in human J720
Physiol 583(Pt 3) 971-982721
Nitsche MA Liebetanz D Lang N Antal A Tergau F amp722
Paulus W (2003) Safety criteria for transcranial direct cur-723
rent stimulation (tDCS) in humans Clin Neurophysiol 114(11)724
2220-2222 author reply 2222-2223725
Opsommer E Guerit JM amp Plaghki L (2003) Exogenous726
and endogenous components of ultralate (C-fibre) evoked727
potentials following CO2 laser stimuli to tiny skin sur-728
face areas in healthy subjects Neurophysiol Clin 33(2)729
78-85730
Parazzini M Fiocchi S Liorni I Priori A amp Ravazzani P731
(2014) Computational modeling of transcranial direct current732
stimulation in the child brain Implications for the treatment of733
refractory childhood focal epilepsy Int J Neural Syst 24(2)734
1430006735
Parazzini M Rossi E Ferrucci R Fiocchi S Liorni I Priori736
A amp Ravazzani P (2013) Computational model of cerebellar737
transcranial direct current stimulation Conf Proc IEEE Eng738
Med Biol Soc 2013 237-240739
Parazzini M Rossi E Ferrucci R Liorni I Priori A amp Ravaz-740
zani P (2014) Modelling the electric field and the current741
density generated by cerebellar transcranial DC stimulation in742
humans Clin Neurophysiol 125(3) 577-584743
Perini I Bergstrand S amp Morrison I (2013) Where pain meets744
action in the human brain J Neurosci 33(40) 15930-15939745
Ploghaus A Tracey I Gati JS Clare S Menon RS Matthews746
PM amp Rawlins JN (1999) Dissociating pain from its antic-747
ipation in the human brain Science 284(5422) 1979-1981748
Priori A Ciocca M Parazzini M Vergari M amp Ferrucci R749
(2014) Transcranial cerebellar direct current stimulation and750
transcutaneous spinal cord direct current stimulation as innova-751
tive tools for neuroscientists J Physiol 592(Pt 16) 3345-3369752
Purpura DP amp McMurtry JC (1965) Intracellular activities and 753
evoked potential changes during polarization of motor cortex 754
J Neurophysiol 28 166-185 755
Qiu Y Inui K Wang X Tran TD amp Kakigi R (2002) Effects 756
of attention distraction and sleep on CO(2) laser evoked poten- 757
tials related to C-fibers in humans Clin Neurophysiol 113(10) 758
1579-1585 759
Reidler JS Mendonca ME Santana MB Wang X Lenkinski 760
R Motta AF Marchand S Latif L amp Fregni F (2012) 761
Effects of motor cortex modulation and descending inhibitory 762
systems on pain thresholds in healthy subjects J Pain 13(5) 763
450-458 764
Riedl V Valet M Woller A Sorg C Vogel D Sprenger 765
T Boecker H Wohlschlager AM amp Tolle TR (2011) 766
Repeated pain induces adaptations of intrinsic brain activity 767
to reflect past and predict future pain Neuroimage 57(1) 206- 768
213 769
Rocca MA Valsasina P Absinta M Colombo B Barcella V 770
Falini A Comi G amp Filippi M (2010) Central nervous 771
system dysregulation extends beyond the pain-matrix network 772
in cluster headache Cephalalgia 30(11) 1383-1391 773
Rossi S Hallett M Rossini PM amp Pascual-Leone A (2009) 774
Safety ethical considerations and application guidelines for 775
the use of transcranial magnetic stimulation in clinical practice 776
and research Clin Neurophysiol 120(12) 2008-2039 777
Rossini PM amp Rossi S (2007) Transcranial magnetic stimulation 778
Diagnostic therapeutic and research potential Neurology 779
68(7) 484-488 780
Ruscheweyh R Kuhnel M Filippopulos F Blum B Eggert 781
T amp Straube A (2014) Altered experimental pain perception 782
after cerebellar infarction Pain 155(7) 1303-1312 783
Schmahmann JD (1991) An emerging concept The cerebellar 784
contribution to higher function Arch Neurol 48(11) 1178- 785
1187 786
Singer T Seymour B OrsquoDoherty J Kaube H Dolan RJ amp 787
Frith CD (2004) Empathy for pain involves the affective but 788
not sensory components of pain Science 303(5661) 1157- 789
1162 790
Smith MJ Keel JC Greenberg BD Adams LF Schmidt PJ 791
Rubinow DA amp Wassermann EM (1999) Menstrual cycle 792
effects on cortical excitability Neurology 53(9) 2069-2072 793
Stoodley CJ amp Schmahmann JD (2009) Functional topography 794
in the human cerebellum A meta-analysis of neuroimaging 795
studies Neuroimage 44(2) 489-501 796
Strick PL Dum RP amp Fiez JA (2009) Cerebellum and non- 797
motor function Annu Rev Neurosci 32 413-434 798
Strigo IA Duncan GH Boivin M amp Bushnell MC (2003) 799
Differentiation of visceral and cutaneous pain in the human 800
brain J Neurophysiol 89(6) 3294-3303 801
Truini A Galeotti F Romaniello A Virtuoso M Iannetti GD 802
amp Cruccu G (2005) Laser-evoked potentials Normative val- 803
ues Clin Neurophysiol 116(4) 821-826 804
Truini A Panuccio G Galeotti F Maluccio MR Sartucci F 805
Avoli M amp Cruccu G (2010) Laser-evoked potentials as a 806
tool for assessing the efficacy of antinociceptive drugs Eur J 807
Pain 14(2) 222-225 808
Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 13
Truini A Vergari M Biasiotta A La Cesa S Gabriele M Di809
Stefano G Cambieri C Cruccu G Inghilleri M amp Pri-810
ori A (2011) Transcutaneous spinal direct current stimulation811
inhibits nociceptive spinal pathway conduction and increases812
pain tolerance in humans Eur J Pain 15(10) 1023-1027813
Valeriani M Le Pera D Restuccia D de Armas L Mili-814
ucci R Betti V Vigevano F amp Tonali P (2007) Parallel815
spinal pathways generate the middle-latency N1 and the late P2816
components of the laser evoked potentials Clin Neurophysiol817
118(5) 1097-1104818
Valeriani M Pazzaglia C Cruccu G amp Truini A (2012) Clinical819
usefulness of laser evoked potentials Neurophysiol Clin 42(5)820
345-353821
Volz MS Mendonca M Pinheiro FS Cui H Santana M amp822
Fregni F (2012) Dissociation of motor task-induced cortical823
excitability and pain perception changes in healthy volunteers824
PLoS One 7(3) e34273825
Wager TD Rilling JK Smith EE Sokolik A Casey KL 826
Davidson RJ Kosslyn SM Rose RM amp Cohen JD 827
(2004) Placebo-induced changes in FMRI in the anticipation 828
and experience of pain Science 303(5661) 1162-1167 829
Zaghi S Thiele B Pimentel D Pimentel T amp Fregni 830
F (2011) Assessment and treatment of pain with non- 831
invasive cortical stimulation Restor Neurol Neurosci 29(6) 832
439-451 833
Zubieta JK Bueller JA Jackson LR Scott DJ Xu Y 834
Koeppe RA Nichols TE amp Stohler CS (2005) Placebo 835
effects mediated by endogenous opioid activity on mu-opioid 836
receptors J Neurosci 25(34) 7754-7762 837
Zunhammer M Busch V Griesbach F Landgrebe M Hajak G 838
amp Langguth B (2011) rTMS over the cerebellum modulates 839
temperature detection and pain thresholds through peripheral 840
mechanisms Brain Stimul 4(4) 210-7 e1 841

Unc
orre
cted
Aut
hor P
roof
T Bocci et al Cerebellum and pain A tcDCS study 13
Truini A Vergari M Biasiotta A La Cesa S Gabriele M Di809
Stefano G Cambieri C Cruccu G Inghilleri M amp Pri-810
ori A (2011) Transcutaneous spinal direct current stimulation811
inhibits nociceptive spinal pathway conduction and increases812
pain tolerance in humans Eur J Pain 15(10) 1023-1027813
Valeriani M Le Pera D Restuccia D de Armas L Mili-814
ucci R Betti V Vigevano F amp Tonali P (2007) Parallel815
spinal pathways generate the middle-latency N1 and the late P2816
components of the laser evoked potentials Clin Neurophysiol817
118(5) 1097-1104818
Valeriani M Pazzaglia C Cruccu G amp Truini A (2012) Clinical819
usefulness of laser evoked potentials Neurophysiol Clin 42(5)820
345-353821
Volz MS Mendonca M Pinheiro FS Cui H Santana M amp822
Fregni F (2012) Dissociation of motor task-induced cortical823
excitability and pain perception changes in healthy volunteers824
PLoS One 7(3) e34273825
Wager TD Rilling JK Smith EE Sokolik A Casey KL 826
Davidson RJ Kosslyn SM Rose RM amp Cohen JD 827
(2004) Placebo-induced changes in FMRI in the anticipation 828
and experience of pain Science 303(5661) 1162-1167 829
Zaghi S Thiele B Pimentel D Pimentel T amp Fregni 830
F (2011) Assessment and treatment of pain with non- 831
invasive cortical stimulation Restor Neurol Neurosci 29(6) 832
439-451 833
Zubieta JK Bueller JA Jackson LR Scott DJ Xu Y 834
Koeppe RA Nichols TE amp Stohler CS (2005) Placebo 835
effects mediated by endogenous opioid activity on mu-opioid 836
receptors J Neurosci 25(34) 7754-7762 837
Zunhammer M Busch V Griesbach F Landgrebe M Hajak G 838
amp Langguth B (2011) rTMS over the cerebellum modulates 839
temperature detection and pain thresholds through peripheral 840
mechanisms Brain Stimul 4(4) 210-7 e1 841
Related Documents











