VIROLOGY 226, 374–383 (1996) ARTICLE NO. 0665 Cellular Proteins Bind to Multiple Sites of the Leader Region of Cauliflower Mosaic Virus 35S RNA DIANA I. DOMINGUEZ, THOMAS HOHN, 1 and WALTRAUD SCHMIDT-PUCHTA Friedrich Miescher-Institut, P.O. Box 2543, CH-4002 Basel, Switzerland Received August 15, 1996; accepted September 24, 1996 UV crosslinking assays were performed to characterize interactions between the leader of the cauliflower mosaic virus (CaMV) 35S RNA and cellular, cytoplasmic proteins. From turnip, a host for CaMV, three different proteins, p35, p49, and p100, interacted with multiple binding sites within the leader. p35 binds to RNA nonspecifically and p49 binds with low specificity, whereas p100 interacts specifically with viral sequences. The expression of the proteins is not induced upon virus infection, as there is no difference in the protein pattern between healthy and infected cell extracts. In a cellular fractionation, p35 and p100 remain in the high-speed supernatant whereas p49 cosediments with polysomes. A possible involvement of these proteins in the translation of the CaMV 35S RNA is discussed. q 1996 Academic Press, Inc. INTRODUCTION 5–10% of mRNAs carry upstream small open reading frames (sORFs) that per se hinder translation (Kozak, Cauliflower mosaic virus (CaMV) is the type member 1987). Compared to cellular mRNAs, the CaMV 35S RNA of caulimoviruses, a group of plant pararetroviruses [for leader is exceptional in several respects: it is unusually a review see Rothnie et al. (1994)]. In CaMV-infected long and folds into a complex stem–loop structure (Fu ¨t- cells, mRNAs are transcribed from two different promot- terer et al., 1988). Furthermore, it contains, depending on ers (Fig. 1): the polycistronic 35S RNA serves as the the strain, 6 to 7 sORFs, 1 to 35 codons in length (Fu ¨tterer template for reverse transcription [for a review see et al., 1988; Hohn et al., 1989). Upstream sORFs and Bonneville and Hohn (1993)], as mRNA for viral proteins long structured leader sequences are features of mRNAs (Dixon and Hohn, 1984), and also as substrate for various whose expression is highly regulated, such as mRNAs splicing events (Kiss-La ´ szlo ´ et al., 1995); and the subgen- of transcription factors [e.g., Lc (Damiani and Wessler, omic, monocistronic 19S RNA encodes the viral inclusion 1993)], proteins involved in signal transduction [e.g., body protein (Covey and Hull, 1981), which acts also opaque-2 (Lohmer et al., 1993); GCN4 (Mu ¨ller and Hin- as translational transactivator (Bonneville et al., 1989; nebusch, 1986); ornithine decarboxylase (Manzella and Fu ¨tterer and Hohn, 1992; Sha et al., 1995; Zijlstra and Blackshear, 1991)] and viral proteins [e.g., picornavirus Hohn, 1992) and probably has additional functions (Him- proteins (Palmenberg, 1987)]. melbach et al., 1996). The negative effect of the leader sORFs on CaMV The terminally redundant 35S RNA contains a 600- translation is partially compensated for by cis-acting sig- nt-long leader (Fig. 2A) that includes important signals nals present in the leader. It was proposed that these controlling different functions of the viral life cycle: it com- act through a shunt mechanism allowing the scanning prises all the signals required for polyadenylation (Roth- complex to bypass the central leader region (Fu ¨tterer nie et al., 1994; Sanfac ¸on et al., 1991), the primer binding et al., 1990, 1993). Ribosome shunting has also been site for reverse transcription (Pfeiffer and Hohn, 1983), a observed in a badnavirus, in rice tungro bacilliform virus splice donor (Kiss-La ´ szlo ´ et al., 1995), and translation (RTBV) (Fu ¨tterer et al., 1996), and in adenovirus late control signals (Fu ¨tterer et al., 1990; Fu ¨tterer et al., 1993). mRNAs, directed by a complex group of secondary struc- It has also been proposed to contain the packaging sig- tures in the tripartite leader (Yueh and Schneider, 1996). nal(s) (Fu ¨tterer et al., 1988; Richins et al., 1987). The molecular mechanism of the shunt is still unclear. Translation of most eukaryotic mRNAs is preceded by However, since shunting can occur in the absence of scanning of the 43S preinitiation complex along the RNA any viral gene product, it is possible that (one or more) and initiation at the first AUG in a favorable context [scan- cellular factors could participate in the process. ning model of translation (Kozak, 1989)]. However, about Protein – RNA interactions between leader or trailer re- gions and cellular proteins were studied in a number of 1 To whom correspondence and reprint requests should be ad- dressed. Fax: (4161) 6973976. animal viruses [e.g., Leopardi et al., 1993; Pogue et al., 374 0042-6822/96 $18.00 Copyright q 1996 by Academic Press, Inc. All rights of reproduction in any form reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

VIROLOGY 226, 374–383 (1996)ARTICLE NO. 0665
Cellular Proteins Bind to Multiple Sites of the Leader Region ofCauliflower Mosaic Virus 35S RNA
DIANA I. DOMINGUEZ, THOMAS HOHN,1 and WALTRAUD SCHMIDT-PUCHTA
Friedrich Miescher-Institut, P.O. Box 2543, CH-4002 Basel, Switzerland
Received August 15, 1996; accepted September 24, 1996
UV crosslinking assays were performed to characterize interactions between the leader of the cauliflower mosaic virus(CaMV) 35S RNA and cellular, cytoplasmic proteins. From turnip, a host for CaMV, three different proteins, p35, p49, andp100, interacted with multiple binding sites within the leader. p35 binds to RNA nonspecifically and p49 binds with lowspecificity, whereas p100 interacts specifically with viral sequences. The expression of the proteins is not induced uponvirus infection, as there is no difference in the protein pattern between healthy and infected cell extracts. In a cellularfractionation, p35 and p100 remain in the high-speed supernatant whereas p49 cosediments with polysomes. A possibleinvolvement of these proteins in the translation of the CaMV 35S RNA is discussed. q 1996 Academic Press, Inc.
INTRODUCTION 5–10% of mRNAs carry upstream small open readingframes (sORFs) that per se hinder translation (Kozak,
Cauliflower mosaic virus (CaMV) is the type member1987). Compared to cellular mRNAs, the CaMV 35S RNA
of caulimoviruses, a group of plant pararetroviruses [forleader is exceptional in several respects: it is unusually
a review see Rothnie et al. (1994)]. In CaMV-infectedlong and folds into a complex stem–loop structure (Fut-cells, mRNAs are transcribed from two different promot-terer et al., 1988). Furthermore, it contains, depending oners (Fig. 1): the polycistronic 35S RNA serves as thethe strain, 6 to 7 sORFs, 1 to 35 codons in length (Futterertemplate for reverse transcription [for a review seeet al., 1988; Hohn et al., 1989). Upstream sORFs andBonneville and Hohn (1993)], as mRNA for viral proteinslong structured leader sequences are features of mRNAs(Dixon and Hohn, 1984), and also as substrate for variouswhose expression is highly regulated, such as mRNAssplicing events (Kiss-Laszlo et al., 1995); and the subgen-of transcription factors [e.g., Lc (Damiani and Wessler,omic, monocistronic 19S RNA encodes the viral inclusion1993)], proteins involved in signal transduction [e.g.,body protein (Covey and Hull, 1981), which acts alsoopaque-2 (Lohmer et al., 1993); GCN4 (Muller and Hin-as translational transactivator (Bonneville et al., 1989;nebusch, 1986); ornithine decarboxylase (Manzella andFutterer and Hohn, 1992; Sha et al., 1995; Zijlstra andBlackshear, 1991)] and viral proteins [e.g., picornavirusHohn, 1992) and probably has additional functions (Him-proteins (Palmenberg, 1987)].melbach et al., 1996).
The negative effect of the leader sORFs on CaMVThe terminally redundant 35S RNA contains a 600-translation is partially compensated for by cis-acting sig-nt-long leader (Fig. 2A) that includes important signalsnals present in the leader. It was proposed that thesecontrolling different functions of the viral life cycle: it com-act through a shunt mechanism allowing the scanningprises all the signals required for polyadenylation (Roth-complex to bypass the central leader region (Futterernie et al., 1994; Sanfacon et al., 1991), the primer bindinget al., 1990, 1993). Ribosome shunting has also beensite for reverse transcription (Pfeiffer and Hohn, 1983), aobserved in a badnavirus, in rice tungro bacilliform virussplice donor (Kiss-Laszlo et al., 1995), and translation(RTBV) (Futterer et al., 1996), and in adenovirus latecontrol signals (Futterer et al., 1990; Futterer et al., 1993).mRNAs, directed by a complex group of secondary struc-It has also been proposed to contain the packaging sig-tures in the tripartite leader (Yueh and Schneider, 1996).nal(s) (Futterer et al., 1988; Richins et al., 1987).The molecular mechanism of the shunt is still unclear.Translation of most eukaryotic mRNAs is preceded byHowever, since shunting can occur in the absence ofscanning of the 43S preinitiation complex along the RNAany viral gene product, it is possible that (one or more)and initiation at the first AUG in a favorable context [scan-cellular factors could participate in the process.ning model of translation (Kozak, 1989)]. However, about
Protein–RNA interactions between leader or trailer re-gions and cellular proteins were studied in a number of1 To whom correspondence and reprint requests should be ad-
dressed. Fax: (4161) 6973976. animal viruses [e.g., Leopardi et al., 1993; Pogue et al.,
3740042-6822/96 $18.00Copyright q 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.
AID VY 8250 / 6a25$$$681 11-13-96 14:39:11 viras AP: Virology

375BINDING OF CELLULAR PROTEINS TO CaMV 35S RNA LEADER
MATERIALS AND METHODS
Preparation of cell extracts
Protoplasts were prepared from noninfected or CaMV-infected turnip leaves (Brassica rapa, ‘‘Just right’’) as de-scribed in Futterer et al. (1989). Cytoplasmic S12 extractswere prepared by freezing and thawing the protoplasts in1 vol of homogenization buffer containing 50 mM Tris/HCl,pH 8.0, 5% glycerol, 100 mM KCl, 1 mM PMSF, 5 mM e-aminocapronic acid, 10 mM EGTA, 5 mM DTT, 1 mg/mlantipain (Gegenheimer, 1990). The mixture was spun at12,000 g at 47 for 20 min. The supernatant, S12, was usedeither directly for UV crosslinking experiments or centri-fuged at 120,000 g for 2 hr at 47 to prepare a polysomalfraction (P) and a high-speed supernatant (S120), respec-
FIG. 1. Circular map of the CaMV genome. The ORFs are indicated tively (Dasgupta et al., 1980). Protein concentration in eachby shaded arrows in the outer circle. ORF VII (no known function), I
sample was determined by the method of Bradford (1976).(M, cell-to-cell movement), II (ITF, insect transmission factor), III (noAliquots of all fractions were stored at 0807.known function), IV (GAG, structural proteins), V (enzymatic functions;
PRO, protease; POL, polymerase and RNaseH), VI (TAV, transactivator).The two major viral RNAs are shown as arrows along the respective Plasmid constructionscoding region.
Construction of plasmid pSLC20 has been previouslydescribed (Futterer et al., 1989). Briefly, this plasmid con-tains an SP6 promoter followed by the entire 614 nt of the1993] and elaborated extensively for picornaviruses (DelCaMV leader (strain S) plus the chloramphenicol acetylAngel et al., 1989; Jang and Wimmer, 1990; Luz and Beck,transferase (CAT) gene fused to ORF VII. Fragments Ia1991; Meerovitch et al., 1989). A cytoplasmic 57-kDa pro-and IVa (Fig. 2B) and the plasmids used for the mutationtein was described to bind to several regions of the inter-and deletion analysis of fragment IVa were constructednal ribosomal entry site (IRES) and this binding was cor-by introducing synthetic oligonucleotides into the vectorrelated with efficient translatability in vitro (Hellen et al.,pT3T7 (Boehringer Mannheim). The remaining fragments1993; Jang and Wimmer, 1990; Pestova et al., 1991). Evi-were obtained by subcloning appropriate regions of thedence had been obtained that p57 is identical to theleader into the vectors pSP65 and pGEM1 (Promega).pyrimidine tract binding protein, PTB (Hellen et al., 1993),The numbering of the CaMV leader is taken from Futterera protein that has been assigned to function in pre-mRNAet al. (1990).splicing (Patton et al., 1991). p52, a second protein in-
volved in poliovirus internal initiation, turned out to beIn vitro transcription
the La-autoantigen (Meerovitch et al., 1993), a nuclearprotein involved in transcription initiation and termination Uniformly labeled RNAs were prepared by in vitro tran-
scription of the linearized plasmids, using SP6 (Boeh-of RNA polymerase III transcripts (Maraia, 1996).ringer Mannheim), T3 (Boehringer Mannheim), or T7 (Bio-Little is known about RNA–protein interactions involv-finex) RNA polymerase and [a-32P]UTP (800 Ci/mmol;ing plant viruses and their host factors. In tobacco mo-NEN) according to the manufacturer’s instructions. Tran-saic virus (TMV), the V sequence at the 5*-end and thescripts were purified on 6% denaturing polyacrylamidepseudoknot region at the 3*-end play an important rolegels. In competition experiments transcripts were pre-in increasing translation efficiency and they specificallypared in the presence of tracer amounts of [a-32P]UTPbind a cellular protein (Leathers et al., 1993; Tanguayin order to allow precise quantification of the amount ofand Gallie, 1996).RNA added.In this report we describe the interaction of turnip cyto-
plasmic proteins with the 5*-leader of the CaMV 35S RNA.UV crosslinking of RNA–protein complexesThree proteins bind to multiple sites in this region. p100
interacts strongly with two defined regions within the Protein extracts (5–10 mg protein) were preincubatedleader; p49, which usually appears as a doublet, binds with at room temperature for 10 min with 100 mg/ml tRNAlower specificity to different subfragments of the leader, but (Boehringer Mannheim) as a nonspecific competitor in ait interacts with neither the first 60 nucleotides of the leader buffer containing 10 mM HEPES (pH 7.6), 8 mM MgCl2 ,nor the vector-derived RNA sequences; p35 binds nonspe- 1 mM DTT, 4 mM spermidine, 1 mg/ml heparin, 5% glyc-cifically to RNA. One of the p100 binding sites was further erol, 20 U RNase inhibitor (Boehringer Mannheim) in a
final volume of 10 ml. In competition experiments, com-characterized by mutational analysis.
AID VY 8250 / 6a25$$$681 11-13-96 14:39:11 viras AP: Virology
376 DOMINGUEZ, HOHN, AND SCHMIDT-PUCHTA
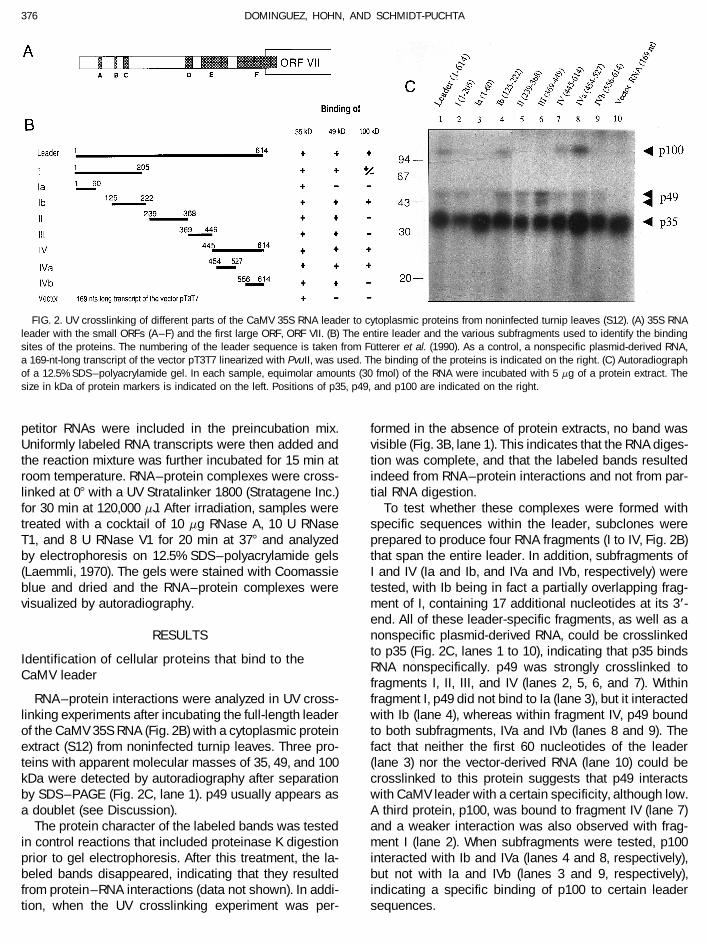
FIG. 2. UV crosslinking of different parts of the CaMV 35S RNA leader to cytoplasmic proteins from noninfected turnip leaves (S12). (A) 35S RNAleader with the small ORFs (A – F) and the first large ORF, ORF VII. (B) The entire leader and the various subfragments used to identify the bindingsites of the proteins. The numbering of the leader sequence is taken from Futterer et al. (1990). As a control, a nonspecific plasmid-derived RNA,a 169-nt-long transcript of the vector pT3T7 linearized with PvuII, was used. The binding of the proteins is indicated on the right. (C) Autoradiographof a 12.5% SDS–polyacrylamide gel. In each sample, equimolar amounts (30 fmol) of the RNA were incubated with 5 mg of a protein extract. Thesize in kDa of protein markers is indicated on the left. Positions of p35, p49, and p100 are indicated on the right.
petitor RNAs were included in the preincubation mix. formed in the absence of protein extracts, no band wasvisible (Fig. 3B, lane 1). This indicates that the RNA diges-Uniformly labeled RNA transcripts were then added and
the reaction mixture was further incubated for 15 min at tion was complete, and that the labeled bands resultedindeed from RNA–protein interactions and not from par-room temperature. RNA–protein complexes were cross-
linked at 07 with a UV Stratalinker 1800 (Stratagene Inc.) tial RNA digestion.To test whether these complexes were formed withfor 30 min at 120,000 mJ. After irradiation, samples were
treated with a cocktail of 10 mg RNase A, 10 U RNase specific sequences within the leader, subclones wereprepared to produce four RNA fragments (I to IV, Fig. 2B)T1, and 8 U RNase V1 for 20 min at 377 and analyzed
by electrophoresis on 12.5% SDS–polyacrylamide gels that span the entire leader. In addition, subfragments ofI and IV (Ia and Ib, and IVa and IVb, respectively) were(Laemmli, 1970). The gels were stained with Coomassie
blue and dried and the RNA–protein complexes were tested, with Ib being in fact a partially overlapping frag-ment of I, containing 17 additional nucleotides at its 3*-visualized by autoradiography.end. All of these leader-specific fragments, as well as anonspecific plasmid-derived RNA, could be crosslinkedRESULTSto p35 (Fig. 2C, lanes 1 to 10), indicating that p35 binds
Identification of cellular proteins that bind to theRNA nonspecifically. p49 was strongly crosslinked to
CaMV leaderfragments I, II, III, and IV (lanes 2, 5, 6, and 7). Withinfragment I, p49 did not bind to Ia (lane 3), but it interactedRNA–protein interactions were analyzed in UV cross-
linking experiments after incubating the full-length leader with Ib (lane 4), whereas within fragment IV, p49 boundto both subfragments, IVa and IVb (lanes 8 and 9). Theof the CaMV 35S RNA (Fig. 2B) with a cytoplasmic protein
extract (S12) from noninfected turnip leaves. Three pro- fact that neither the first 60 nucleotides of the leader(lane 3) nor the vector-derived RNA (lane 10) could beteins with apparent molecular masses of 35, 49, and 100
kDa were detected by autoradiography after separation crosslinked to this protein suggests that p49 interactswith CaMV leader with a certain specificity, although low.by SDS–PAGE (Fig. 2C, lane 1). p49 usually appears as
a doublet (see Discussion). A third protein, p100, was bound to fragment IV (lane 7)and a weaker interaction was also observed with frag-The protein character of the labeled bands was tested
in control reactions that included proteinase K digestion ment I (lane 2). When subfragments were tested, p100interacted with Ib and IVa (lanes 4 and 8, respectively),prior to gel electrophoresis. After this treatment, the la-
beled bands disappeared, indicating that they resulted but not with Ia and IVb (lanes 3 and 9, respectively),indicating a specific binding of p100 to certain leaderfrom protein–RNA interactions (data not shown). In addi-
tion, when the UV crosslinking experiment was per- sequences.
AID VY 8250 / 6a25$$$681 11-13-96 14:39:11 viras AP: Virology
377BINDING OF CELLULAR PROTEINS TO CaMV 35S RNA LEADER
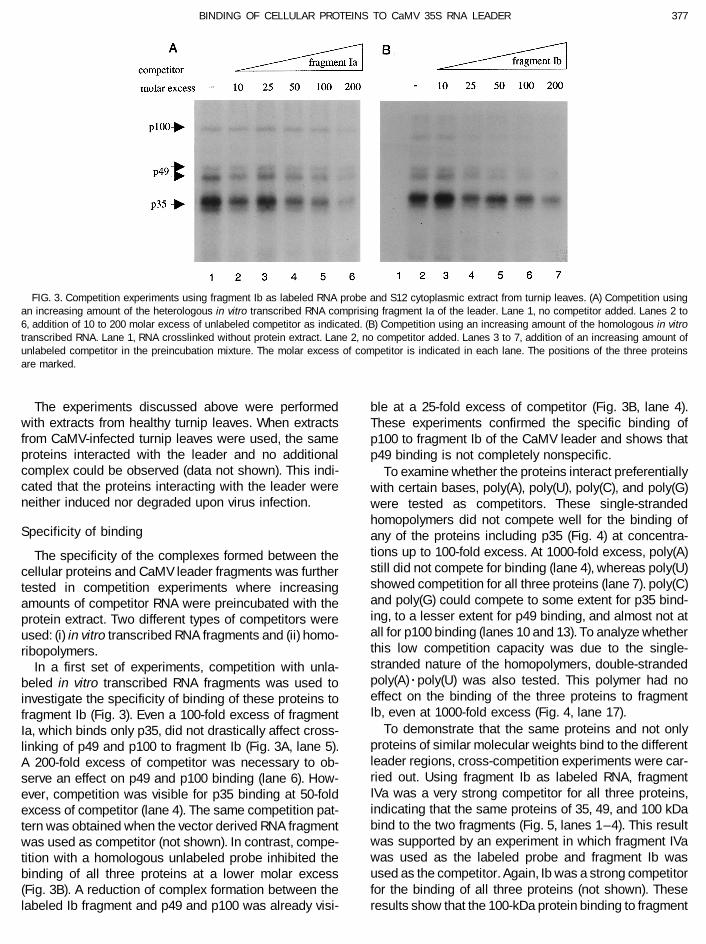
FIG. 3. Competition experiments using fragment Ib as labeled RNA probe and S12 cytoplasmic extract from turnip leaves. (A) Competition usingan increasing amount of the heterologous in vitro transcribed RNA comprising fragment Ia of the leader. Lane 1, no competitor added. Lanes 2 to6, addition of 10 to 200 molar excess of unlabeled competitor as indicated. (B) Competition using an increasing amount of the homologous in vitrotranscribed RNA. Lane 1, RNA crosslinked without protein extract. Lane 2, no competitor added. Lanes 3 to 7, addition of an increasing amount ofunlabeled competitor in the preincubation mixture. The molar excess of competitor is indicated in each lane. The positions of the three proteinsare marked.
The experiments discussed above were performed ble at a 25-fold excess of competitor (Fig. 3B, lane 4).with extracts from healthy turnip leaves. When extracts These experiments confirmed the specific binding offrom CaMV-infected turnip leaves were used, the same p100 to fragment Ib of the CaMV leader and shows thatproteins interacted with the leader and no additional p49 binding is not completely nonspecific.complex could be observed (data not shown). This indi- To examine whether the proteins interact preferentiallycated that the proteins interacting with the leader were with certain bases, poly(A), poly(U), poly(C), and poly(G)neither induced nor degraded upon virus infection. were tested as competitors. These single-stranded
homopolymers did not compete well for the binding ofSpecificity of binding any of the proteins including p35 (Fig. 4) at concentra-
tions up to 100-fold excess. At 1000-fold excess, poly(A)The specificity of the complexes formed between thestill did not compete for binding (lane 4), whereas poly(U)cellular proteins and CaMV leader fragments was furthershowed competition for all three proteins (lane 7). poly(C)tested in competition experiments where increasingand poly(G) could compete to some extent for p35 bind-amounts of competitor RNA were preincubated with theing, to a lesser extent for p49 binding, and almost not atprotein extract. Two different types of competitors wereall for p100 binding (lanes 10 and 13). To analyze whetherused: (i) in vitro transcribed RNA fragments and (ii) homo-this low competition capacity was due to the single-ribopolymers.stranded nature of the homopolymers, double-strandedIn a first set of experiments, competition with unla-poly(A)rpoly(U) was also tested. This polymer had nobeled in vitro transcribed RNA fragments was used toeffect on the binding of the three proteins to fragmentinvestigate the specificity of binding of these proteins toIb, even at 1000-fold excess (Fig. 4, lane 17).fragment Ib (Fig. 3). Even a 100-fold excess of fragment
To demonstrate that the same proteins and not onlyIa, which binds only p35, did not drastically affect cross-proteins of similar molecular weights bind to the differentlinking of p49 and p100 to fragment Ib (Fig. 3A, lane 5).leader regions, cross-competition experiments were car-A 200-fold excess of competitor was necessary to ob-ried out. Using fragment Ib as labeled RNA, fragmentserve an effect on p49 and p100 binding (lane 6). How-IVa was a very strong competitor for all three proteins,ever, competition was visible for p35 binding at 50-foldindicating that the same proteins of 35, 49, and 100 kDaexcess of competitor (lane 4). The same competition pat-bind to the two fragments (Fig. 5, lanes 1–4). This resulttern was obtained when the vector derived RNA fragmentwas supported by an experiment in which fragment IVawas used as competitor (not shown). In contrast, compe-was used as the labeled probe and fragment Ib wastition with a homologous unlabeled probe inhibited theused as the competitor. Again, Ib was a strong competitorbinding of all three proteins at a lower molar excessfor the binding of all three proteins (not shown). These(Fig. 3B). A reduction of complex formation between the
labeled Ib fragment and p49 and p100 was already visi- results show that the 100-kDa protein binding to fragment
AID VY 8250 / 6a25$$$681 11-13-96 14:39:11 viras AP: Virology
378 DOMINGUEZ, HOHN, AND SCHMIDT-PUCHTA
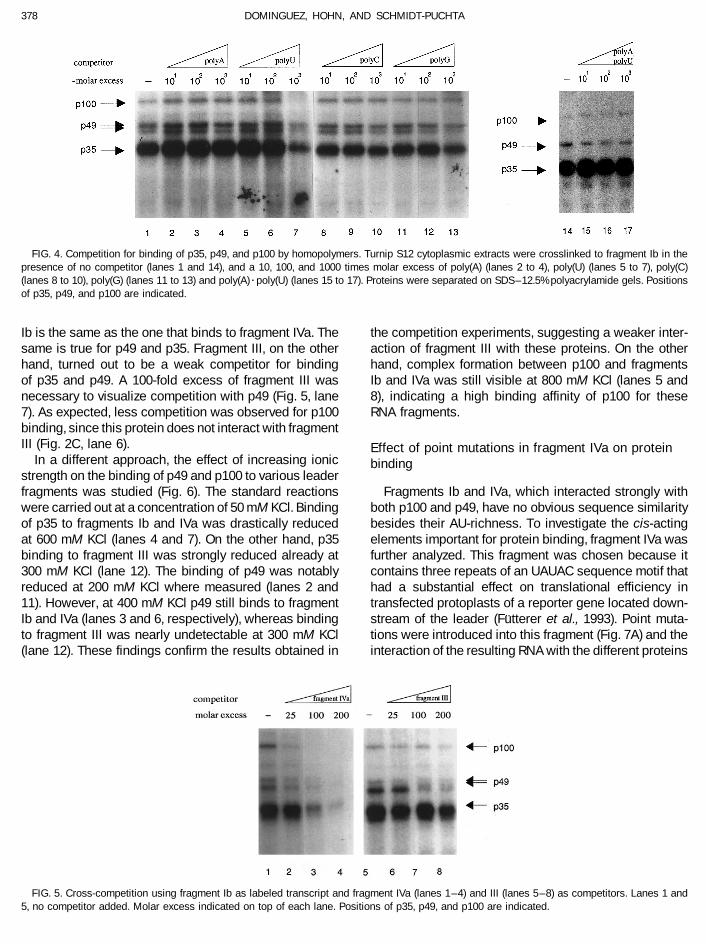
FIG. 4. Competition for binding of p35, p49, and p100 by homopolymers. Turnip S12 cytoplasmic extracts were crosslinked to fragment Ib in thepresence of no competitor (lanes 1 and 14), and a 10, 100, and 1000 times molar excess of poly(A) (lanes 2 to 4), poly(U) (lanes 5 to 7), poly(C)(lanes 8 to 10), poly(G) (lanes 11 to 13) and poly(A)rpoly(U) (lanes 15 to 17). Proteins were separated on SDS–12.5% polyacrylamide gels. Positionsof p35, p49, and p100 are indicated.
Ib is the same as the one that binds to fragment IVa. The the competition experiments, suggesting a weaker inter-action of fragment III with these proteins. On the othersame is true for p49 and p35. Fragment III, on the other
hand, turned out to be a weak competitor for binding hand, complex formation between p100 and fragmentsIb and IVa was still visible at 800 mM KCl (lanes 5 andof p35 and p49. A 100-fold excess of fragment III was
necessary to visualize competition with p49 (Fig. 5, lane 8), indicating a high binding affinity of p100 for theseRNA fragments.7). As expected, less competition was observed for p100
binding, since this protein does not interact with fragmentIII (Fig. 2C, lane 6). Effect of point mutations in fragment IVa on protein
In a different approach, the effect of increasing ionic bindingstrength on the binding of p49 and p100 to various leaderfragments was studied (Fig. 6). The standard reactions Fragments Ib and IVa, which interacted strongly with
both p100 and p49, have no obvious sequence similaritywere carried out at a concentration of 50 mM KCl. Bindingof p35 to fragments Ib and IVa was drastically reduced besides their AU-richness. To investigate the cis-acting
elements important for protein binding, fragment IVa wasat 600 mM KCl (lanes 4 and 7). On the other hand, p35binding to fragment III was strongly reduced already at further analyzed. This fragment was chosen because it
contains three repeats of an UAUAC sequence motif that300 mM KCl (lane 12). The binding of p49 was notablyreduced at 200 mM KCl where measured (lanes 2 and had a substantial effect on translational efficiency in
transfected protoplasts of a reporter gene located down-11). However, at 400 mM KCl p49 still binds to fragmentIb and IVa (lanes 3 and 6, respectively), whereas binding stream of the leader (Futterer et al., 1993). Point muta-
tions were introduced into this fragment (Fig. 7A) and theto fragment III was nearly undetectable at 300 mM KCl(lane 12). These findings confirm the results obtained in interaction of the resulting RNA with the different proteins
FIG. 5. Cross-competition using fragment Ib as labeled transcript and fragment IVa (lanes 1–4) and III (lanes 5–8) as competitors. Lanes 1 and5, no competitor added. Molar excess indicated on top of each lane. Positions of p35, p49, and p100 are indicated.
AID VY 8250 / 6a25$$$681 11-13-96 14:39:11 viras AP: Virology
379BINDING OF CELLULAR PROTEINS TO CaMV 35S RNA LEADER
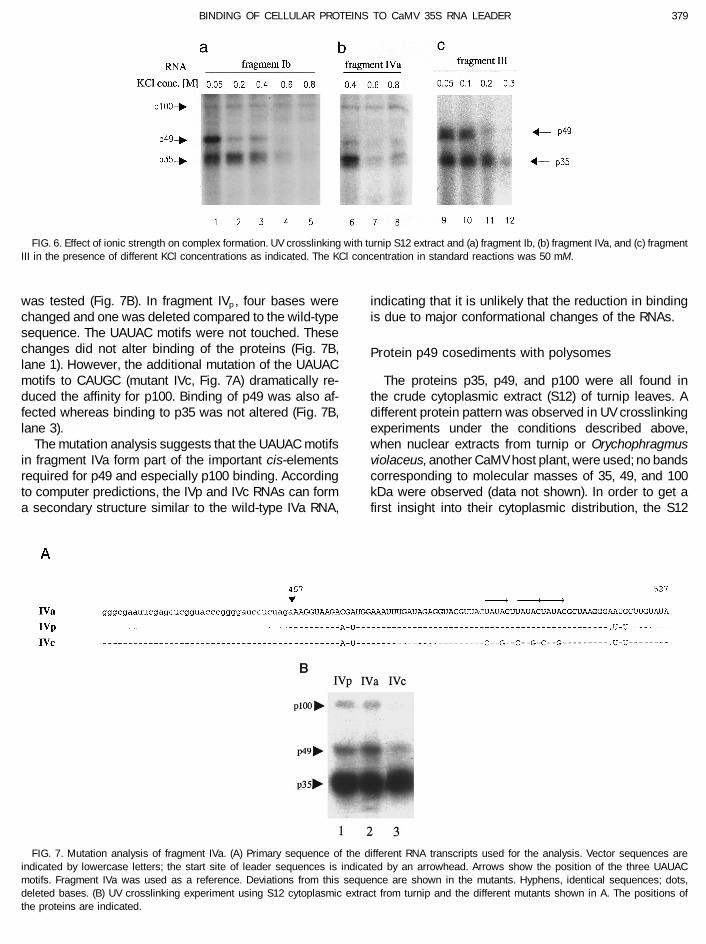
FIG. 6. Effect of ionic strength on complex formation. UV crosslinking with turnip S12 extract and (a) fragment Ib, (b) fragment IVa, and (c) fragmentIII in the presence of different KCl concentrations as indicated. The KCl concentration in standard reactions was 50 mM.
was tested (Fig. 7B). In fragment IVp , four bases were indicating that it is unlikely that the reduction in bindingis due to major conformational changes of the RNAs.changed and one was deleted compared to the wild-type
sequence. The UAUAC motifs were not touched. Thesechanges did not alter binding of the proteins (Fig. 7B, Protein p49 cosediments with polysomeslane 1). However, the additional mutation of the UAUACmotifs to CAUGC (mutant IVc, Fig. 7A) dramatically re- The proteins p35, p49, and p100 were all found in
the crude cytoplasmic extract (S12) of turnip leaves. Aduced the affinity for p100. Binding of p49 was also af-fected whereas binding to p35 was not altered (Fig. 7B, different protein pattern was observed in UV crosslinking
experiments under the conditions described above,lane 3).The mutation analysis suggests that the UAUAC motifs when nuclear extracts from turnip or Orychophragmus
violaceus, another CaMV host plant, were used; no bandsin fragment IVa form part of the important cis-elementsrequired for p49 and especially p100 binding. According corresponding to molecular masses of 35, 49, and 100
kDa were observed (data not shown). In order to get ato computer predictions, the IVp and IVc RNAs can forma secondary structure similar to the wild-type IVa RNA, first insight into their cytoplasmic distribution, the S12
FIG. 7. Mutation analysis of fragment IVa. (A) Primary sequence of the different RNA transcripts used for the analysis. Vector sequences areindicated by lowercase letters; the start site of leader sequences is indicated by an arrowhead. Arrows show the position of the three UAUACmotifs. Fragment IVa was used as a reference. Deviations from this sequence are shown in the mutants. Hyphens, identical sequences; dots,deleted bases. (B) UV crosslinking experiment using S12 cytoplasmic extract from turnip and the different mutants shown in A. The positions ofthe proteins are indicated.
AID VY 8250 / 6a25$$$681 11-13-96 14:39:11 viras AP: Virology

380 DOMINGUEZ, HOHN, AND SCHMIDT-PUCHTA
V leader and to the pseudoknot domain in the 3*-UTR ofTMV RNA and has been proposed to mediate the positiveeffect that these regions have on translation (Tanguayand Gallie, 1996).
The leader sequences of CaMV and RTBV pregenomicRNAs and adenovirus late RNAs have been proposed todirect translation by a mechanism known as ribosomeshunting or jumping. (Futterer et al., 1993, 1996; Yuehand Schneider, 1996). The fact that shunting can occurin the absence of viral gene products prompted us tosearch for cellular proteins binding to CaMV leader aspossible candidates to participate in the process. The UVcrosslinking experiments reported here revealed threecytoplasmic proteins that interact with the 5* leader ofCaMV 35S RNA (Fig. 2). The relative amount of each ofthe three proteins that was crosslinked to RNA was vari-able between experiments. The reason for this is un-known, but it could reflect variations in the amount ofeach of the proteins in the different batches of cellularextracts. p35 was shown to bind rather nonspecificallyto all RNAs tested. p49 did not bind to a nonviral plasmid-
FIG. 8. Localization of the proteins. Different protein fractions (see derived RNA; however, it interacted with most leaderMaterials and Methods) were crosslinked to fragment IVa. S12, crude fragments (Fig. 2C), with the exception of the first 60cytoplasmic extract (lane 1); P, polysomal pellet (lane 2) and S120, high-
nucleotides of the leader (fragment Ia). In many experi-speed supernatant (lane 3). The positions of the proteins are indicated.ments p49 appeared as a double band. Whether thissecond band corresponds to a posttranslational modifi-
preparation was further centrifuged at 120,000 g to obtain cation of p49, a degradation product, or, alternatively, toa high-speed supernatant (S120) and a polysomal pellet an additional protein is not yet known. However, since(P). Figure 8 shows that p35 and p100 both remained in this second band shows the same specificity as p49 (seethe high-speed supernatant (lane 3), whereas p49 cosed- competition experiments, Figs. 3 and 4) and the sameimented with the polysomes (lane 2). cellular localization (Fig. 8), it is most probably related
to p49 and not an additional protein. A more specificDISCUSSION interaction with only two defined regions of the leader,
subfragments Ib and IVa, could be demonstrated forIn eukaryotic cells posttranscriptional mechanismsp100. This interaction seems to be strong, since it re-play a considerable role in regulating gene expressionquires more than 800 mM KCl to dissociate the complexand there is a multitude of proteins interacting with(Fig. 6). This is comparable to the binding affinity of themRNA leader or trailer regions involved in mediating thisTMV p30 protein to RNA (Citovsky et al., 1990). Besidesregulation (e.g., Burd and Dreyfuss, 1994; Curtis et al.,these interactions, p100 was also bound to the larger1995; Mc Carthy and Kollmus, 1995; Standart and Jack-fragments I and IV (Fig. 2C). The fact that p100 interactsson, 1994). Leader-binding regulatory proteins may in-more weakly with fragments I and IV than with the sub-hibit or enhance translation by stabilizing secondaryfragments Ib and IVa could have different interpretations.structures. For instance, translation of ferritin is re-One possibility is that the structure of the RNA is im-pressed by the IRP protein that binds to the iron respon-portant for binding; a more favorable structure might besive element (IRE) present in the 5*-untranslated regionformed by fragment Ib and IVa, whereas fragments I and(UTR) of the ferritin mRNA (Klausner et al., 1993). PositiveIV might adopt a conformation for which p100 wouldtranslational regulation has mainly been observed in con-have less affinity. For fragment I, another possibility exits:nection with viral RNAs; for instance, cellular proteins,subfragment Ib contains 17 additional nucleotides (Fig.like p57, identified as the polypyrimidine tract binding2B) which could be important for p100 recognition.protein (PTB) and p52, identified as the La-autoantigen,
By cross-competition experiments it was shown thatare proposed to be involved in the internal translationalthe same p35, p49, and p100 protein species bind toinitiation of picornaviral RNA (Hellen et al., 1993; Kamin-the different leader fragments and not different proteinsski et al., 1995; Meerovitch et al., 1993; Svitkin et al.,having similar mobility in SDS–PAGE. These experi-1994). A plant protein of 102 kDa molecular mass, iso-
lated from wheat germ, has been shown to bind to the ments also indicate that within each protein the same
AID VY 8250 / 6a25$$$681 11-13-96 14:39:11 viras AP: Virology

381BINDING OF CELLULAR PROTEINS TO CaMV 35S RNA LEADER
binding site is used for interaction with the different ever this is not a prerequisite for proteins involved intranslation, since some initiation and elongation factorsRNAs.
The entire CaMV 35S RNA leader is AU-rich. A compar- are isolated from the supernatants of cytoplasmic ex-tracts after a high-speed centrifugation to pellet the poly-ison of the different binding sites for p100, however, did
not reveal any apparent sequence similarity. The specific somal fraction (Lax et al., 1986). In addition, both bindingsites of p100 overlap with regions of the CaMV leaderbinding of a protein to different RNAs without any appar-
ent sequence similarity was also described for the bind- that have been defined as important for the shunt mecha-nism of translation in transient expression experimentsing of p57 to different parts of the internal ribosome entry
site of poliovirus RNA (Hellen et al., 1994) and foot and (Futterer et al., 1990, 1993). In particular, mutations intro-duced into the UAUAC motifs of fragment IVa reducedmouth disease virus RNA (Luz and Beck, 1991) and for
the binding of the wheat germ protein p102 to the 5*- translation efficiently in protoplasts and drastically re-duced binding of p100 and also p49.and 3*-UTRs of TMV RNA (Tanguay and Gallie, 1996).
To determine more precisely cis-acting elements re- In picornaviruses, the RNA binding proteins arethought to be part of a larger complex (Jang and Wimmer,quired for binding, fragment IVa was further investigated.
In transient expression experiments this region was 1990). This could also be true for the CaMV leader bind-ing proteins. The two binding sites of p100 are about 200shown to be important for translation of a reporter gene
downstream of the leader. It contains three copies of a nt apart in the primary sequence but are very close toeach other in the proposed secondary structure (FuttererUAUAC motif, and mutations in these motifs affect trans-
lation efficiency (Futterer et al., 1993). Point mutations et al., 1988). An interaction between different proteinsand different parts of the leader might stabilize a specificintroduced into these motifs considerably reduced bind-
ing of p49 and p100 (Fig. 7B). The effect of the mutation tertiary conformation. Such a complex might allow theshunt mechanism to take place.is more drastic for p100 binding. From these results we
conclude that the motifs are important for the binding of The 35S RNA is terminally redundant and the first 180nt of the leader, including most of fragment Ib, are dupli-both proteins to fragment IVa. On the other hand, the
exact primary sequence of the motifs does not seem to cated at the 3*-end of the RNA. Therefore an even morecomplex interaction between the 5*- and 3*-nontrans-be absolutely required for binding, since they are not
present in fragment Ib to which the proteins also bind. lated ends of the CaMV 35S RNA could be considered.Studies are underway to purify the leader binding pro-In addition, the requirements of both proteins must be
different, since p49 interacts with RNA molecules to teins and to investigate further their function in the virallife cycle, and as they are also present in noninfectedwhich p100 does not bind. One possibility is that, in
addition to the primary sequence, secondary and tertiary cells, their function in the host.structural elements are involved in the recognition; thefact that p100 interacts weakly with fragments I and IV ACKNOWLEDGMENTSbut strongly with the subfragments Ib and IVa also points
We thank Karl Gordon for providing the fragment Ib clone, his com-in this direction. Several RNA binding proteins are knownmunication concerning the pseudoknot structure, and helpful discus-
to recognize such elements, mostly short stems with spe- sions. We very much appreciate the technical assistance of Matthiascific bulges or loops [e.g., Rop (Predki et al., 1995), IRE Muller. We thank Helen Rothnie, Johannes Futterer, Witold Filipowicz,
and Lyubov Ryabova for critical reading of the manuscript. W.S.-P. wasbinding protein (Jaffrey et al., 1993), Rev (Cullen, 1992),supported by an EMBO fellowship.Tat (Mark et al., 1993)]. The entire leader forms an exten-
sive secondary structure. The two regions to which p100binds are predicted to be able to form pseudoknots (K. REFERENCESGordon, personal communication). One of the proposed
Bonneville, J. M., and Hohn, T. (1993). ‘‘Reverse Transcriptase’’ (N.pseudoknots would be included in fragment Ib and the Skalka and S. Goff, Eds.), pp. 357–390, Cold Spring Harbor Labora-other in fragment IVa. The latter covers exactly the tory Press, Cold Spring Harbor, NY.
Bonneville, J. M., Sanfacon, H., Futterer, J., and Hohn, T. (1989). Posttran-UAUAC motifs. In nondenaturing electrophoresis gels,scriptional trans-activation in cauliflower mosaic virus. Cell 59, 1135–fragment IVa RNA appears as two bands of different1143.electrophoretic mobility, indicating that in fact two iso-
Bradford, M. (1976). A rapid and sensitive method for the quantificationforms exist (D.D., unpublished results). Pseudoknots of microgram quantities of protein utilizing the principle of proteinpresent in the 3*-UTR of TMV are the targets for specific dye binding. Anal. Biochem. 76, 248–254.
Burd, C. G., and Dreyfuss, G. (1994). Conserved structure and diversityprotein binding and play a role in the translational effi-of functions of RNA-binding proteins. Science 265, 615–621.ciency of TMV RNA (Leathers et al., 1993; Tanguay and
Citovsky, V., Knorr, D., Schuster, G., and Zambryski, P. (1990). TheGallie, 1996).P30 movement protein of tobacco mosaic virus is a single-stranded
Several hints suggest that p49 and p100 could be in- nucleic acid binding protein. Cell 60, 637–647.volved in translation of the 35S RNA. First of all, p49 Covey, S. N., and Hull, R. (1981). Transcription of cauliflower mosaic
virus DNA. Detection of transcripts, properties, and location of thecosediments with the polysomal fraction (Fig. 8). How-
AID VY 8250 / 6a25$$$681 11-13-96 14:39:11 viras AP: Virology

382 DOMINGUEZ, HOHN, AND SCHMIDT-PUCHTA
gene encoding the virus inclusion body protein. Virology 111, 463– for internal initiation of translation of encephalomyocarditis virusRNA. RNA 1, 924–938.474.
Cullen, B. R. (1992). Mechanism of action of regulatory proteins en- Kiss-Laszlo, Z., Blanc, S., and Hohn, T. (1995). Splicing of cauliflowermosaic virus 35S RNA is essential for viral infectivity. EMBO J. 14,coded by complex retroviruses. Microbiol. Rev. 56, 2498–2501.
Curtis, D., Lehmann, R., and Zamore, P. D. (1995). Translational regula- 3552–3562.Klausner, R. D., Rouault, T. A., and Harford, J. B. (1993). Regulating thetion in development. Cell 81, 171–178.
Damiani, R. D., and Wessler, S. (1993). An upstream open reading frame fate of mRNA: The control of cellular iron metabolism. Cell 72, 19–28.represses expression of Lc, a member of the R/B family of maize
transcriptional activators. Proc. Natl. Acad. Sci. USA 90, 8244–8248. Kozak, M. (1987). An analysis of 5*-noncoding sequences from 699vertebrate messenger RNAs. Nucleic Acids Res. 15, 8125–8148.Dasgupta, A., Zabel, P., and Baltimore, D. (1980). Dependence of the
activity of the poliovirus replicase on a host cell protein. Cell 19, Kozak, M. (1989). The scanning model for translation—An update. J.Cell Biol. 108, 229–241.423–429.
Del Angel, R. M., Papavassiliou, A. G., Fernandez-Tomas, C., Silverstein, Laemmli, U. K. (1970). Cleavage of structural proteins during the assem-bly of the head of bacteriophage T4. Nature 227, 680–685.S. J., and Racaniello, V. R. (1989). Cell proteins bind to multiple sites
within the 5*-untranslated region of poliovirus RNA. Proc. Natl. Acad. Lax, S. R., Lauer, S. J., Browning, K. S., and Ravel, J. M. (1986). Purifica-tion and properties of protein synthesis initiation and elongationSci. USA 86, 8299–8303.
Dixon, L. K., and Hohn, T. (1984). Initiation of translation of the cauli- factors from wheat germ. Methods Enzymol. 118, 109–128.Leathers, V., Tanguay, R., Kobayashi, M., and Gallie, D. R. (1993). Aflower mosaic virus genome from a polycistronic mRNA: Evidence
from deletion mutagenesis. EMBO J. 3, 2731–2736. phylogenetically conserved sequence within viral 3* untranslatedRNA pseudoknots regulates translation. Mol. Cell. Biol. 13, 5331–Futterer, J., Gordon, K., Bonneville, J. M., Sanfacon, H., Pisan, B., Pens-
wick, J., and Hohn, T. (1988). The leading sequence of caulimovirus 5347.Leopardi, R., Hukkanen, V., Vainionaa, R., and Salmi, A. A. (1993). Celllarge RNA can be folded into a large stem-loop structure. Nucleic
Acids Res. 16, 8377–8390. proteins bind to sites within the 3* noncoding region and the positive-strand leader sequence of measles virus RNA. J. Virol. 67, 785–790.Futterer, J., Gordon, K., Pfeiffer, P., Sanfacon, H., Pisan, B., Bonneville,
J., and Hohn, T. (1989). Differential inhibition of downstream gene Lohmer, S., Maddaloni, M., Motto, M., Salamini, F., and Thompson, R. D.(1993). Translation of the mRNA of the maize transcriptional activatorexpression by the cauliflower mosaic virus 35S RNA leader. Virus
Genes 3, 45–55. opaque-2 is inhibited by upstream open reading frames present inthe leader sequence. Plant Cell 5, 65–73.Futterer, J., Gordon, K., Sanfacon, H., Bonneville, J., and Hohn, T. (1990).
Positive and negative control of translation by the leader of cauli- Luz, N., and Beck, E. (1991). Interaction of a cellular 57-kilodalton pro-tein with the internal translation initiation site of foot-and-mouth dis-flower mosaic virus pregenomic 35S RNA. EMBO J. 9, 1697–1707.
Futterer, J., and Hohn, T. (1992). Role of an upstream open reading ease virus. J. Virol. 65, 6486–6494.Manzella, J. M., and Blackshear, P. J. (1991). Regulation of rat ornithineframe in the translation of polycistronic mRNA in plant cells. Nucleic
Acids Res. 20, 3851–3857. decarboxylase mRNA translation by its 5*-untranslated region. J.Virol. 65, 6486–6494.Futterer, J., Kiss-Laszlo, Z., and Hohn, T. (1993). Nonlinear ribosome
migration on cauliflower mosaic virus 35S RNA. Cell 73, 789–802. Maraia, R. J. (1996). Transcription termination factor La is also an initia-tion factor for RNA polymerase III. Proc. Natl. Acad. Sci. USA 93,Futterer, J., Potrykus, I., Bao, Y., Li, L., Burns, T. M., Hull, R., and Hohn,
T. (1996). Position-dependent ATT initiation during plant pararetrovi- 3383–3387.Mark, J. C., Lamont, C., Hamy, F., Dingwall, C., Green, S. M., Lowe, A. D.,rus rice tungro bacilliform virus translation. J. Virol. 70, 2999–3010.
Gegenheimer, P. (1990). in ‘‘Methods in Enzymology’’ (M. Deutscher, Butler, P. J. G., Gait, M. J., and Karn, J. (1993). High affinity binding ofTAR RNA by the human immunodeficiency virus type-1 tat proteinEd.), pp. 174–193, Academic Press, San Diego.
Hellen, C. U. T., Pestova, T. V., Litterst, M., and Wimmer, E. (1994). The requires base-pairs in the RNA stem and animo acid residues flank-ing the basic region. J. Mol. Biol. 230, 90–110.cellular polypeptide p57 (pyrimidine tract-binding protein) binds to
multiple sites in the poliovirus 5* nontranslated region. J. Virol. 68, Mc Carthy, J. E. G., and Kollmus, H. (1995). Cytoplasmic mRNA–proteininteractions in eukaryotic gene expression. Trends Biochem. Sci. 20,941–950.
Hellen, C. U. T., Witherell, G. W., Schmid, M., Shin, S. H., Pestova, T. V., 191–197.Meerovitch, K., Pelletier, J., and Sonenberg, N. (1989). A cellular proteinGil, A., and Wimmer, E. (1993). A cytoplasmic 57-kD protein that is
required for translation of picornavirus RNA by internal ribosomal that binds to the 5*-noncoding region of poliovirus RNA: Implicationsfor internal translation initiation. Genes Dev. 3, 1026–1034.entry is identical to the nuclear pyrimidine tract-binding protein. Proc.
Natl. Acad. Sci. USA 90, 7642 –7646. Meerovitch, K., Svitkin, Y. V., Lee, H. S., Lejbkowicz, F., Kenan, D. J.,Chan, E. K. L., Agol, V., Keene, J. D., and Sonenberg, N. (1993). LaHimmelbach, A., Chapdelaine, Y., and Hohn, T. (1996). Interaction be-
tween Cauliflower Mosaic Virus inclusion body protein and capsid autoantigen enhances and corrects aberrant translation of poliovirusRNA in reticulocyte lysate. J. Virol. 67, 3798–3807.protein: Implications for viral assembly. Virology 217, 147–157.
Hohn, T., Bonneville, J. M., Futterer, J., Gordon, K., Pisan, B., Sanfacon, Muller, P. P., and Hinnebusch, A. G. (1986). Multiple upstream AUGcodons mediate translational control of GCN4. Cell 45, 201–207.H., Schultze, M., and Jiricny, J. (1989). in ‘‘UCLA Symposium on the
Molecular Biology of Plant Pathogen Interactions’’ (B. Staskawics Palmenberg, A. C. (1987). in ‘‘Positive Strand RNA Viruses’’ (M. A. Brintonand R. R. Rueckert, Eds.), UCLA Symposia Cellular Biology, pp. 25–and P. Ahlquist, Eds.), pp. 153 –165, A. R. Liss, New York.
Jaffrey, S. R., Haile, D. J., Klausner, R. D., and Harford, J. B. (1993). The 34, A. R. Liss, New York.Patton, J. G., Meyer, S., Tempst, P., and Nadal-Ginard, B. (1991). Charac-interaction between iron-responsive element binding protein and its
cognate RNA is highly dependent upon both RNA sequence and terization and molecular cloning of polypyrimidine tract-binding pro-tein: A component of a complex necessary for pre-mRNA splicing.structure. Nucleic Acids Res. 21, 4627–4631.
Jang, S. K., and Wimmer, E. (1990). Cap-independent translation of en- Genes Dev. 5, 1237–1251.Pestova, T. V., Hellen, C. U. T., and Wimmer, E. (1991). Translation ofcephalomyocarditis virus RNA: Structural elements of the internal
ribosome entry site and essential binding of a cellular 57 kDa protein. poliovirus RNA: Role of an essential cis-acting oligopyrimidine ele-ment within the 5* nontranslated region and involvement of a cellularGenes Dev. 4, 1560–1572.
Kaminski, A., Hunt, S. L., Patton, J. G., and Jackson, R. J. (1995). Direct 57-kilodalton protein. J. Virol. 65, 6194–6204.Pfeiffer, P., and Hohn, T. (1983). Involvement of reverse transcription inevidence that polypyrimidine tract binding protein (PTB) is essential
AID VY 8250 / 6a25$$$681 11-13-96 14:39:11 viras AP: Virology

383BINDING OF CELLULAR PROTEINS TO CaMV 35S RNA LEADER
the replication of cauliflower mosaic virus: A detailed model and test Sha, Y., Broglio, E. P., Cannopn, J. F., and Schoelz, J. E. (1995). Expres-of some aspects. Cell 33, 781 –789. sion of a plant viral polycistronic mRNA in yeast, Saccharomyces
Pogue, G. P., Cao, X.-Q., Singh, N. K., and Nakhasi, H. L. (1993). 5* cerevisiae, mediated by a plant virus translational transactivator.sequences of rubella virus RNA stimulate translation of chimeric Proc. Natl. Acad. Sci. USA 92, 8911–8915.RNAs and specificallly interact with two host-encoded proteins. J. Standart, N., and Jackson, R. J. (1994). Regulation of translation by spe-Virol. 67, 7106–7117. cific protein–mRNA interactions. Biochemie 76, 867–879.
Predki, P. F., Nayak, L. M., Gottlieb, M. C. B., and Regan, L. (1995). Dis- Svitkin, Y. V., Meerovitch, K., Lee, H. S., Dholakia, J. N., Kenan, D. J.,secting RNA–protein interactions: RNA–RNA recognition by Rop. Agol, V., and Sonenberg, N. (1994). Internal translation initiation onCell 80, 41–50. poliovirus RNA: Further characterization of La funtion in poliovirus
Richins, R. D., Scholthof, H. B., and Shepherd, R. J. (1987). Sequence of translated in vitro. J. Virol. 68, 1544–1550.figwort mosaic virus DNA (caulimovirus group). Nucleic Acids Res.
Tanguay, R. L., and Gallie, D. R. (1996). Isolation and characterization15, 8451–8466.
of the 102-kilodalton RNA-binding protein that binds to the 5* and 3*Rothnie, H. M., Chapdelaine, Y., and Hohn, T. (1994). Pararetrovirusestranslational enhancers of tobacco mosaic virus. J. Biol. Chem. 271,and retroviruses: A comparative review of viral structure and gene14316–14322.expression strategies. Adv. Virus Res. 44, 1–67.
Yueh, A., and Schneider, R. J. (1996). Selective translation initiation byRothnie, H. M., Reid, J., and Hohn, T. (1994). The contribution of AAUAAAribosome jumping in adenovirus-infected and heat shocked cells.and the upstream element UUUGUA to the efficiency of mRNA 3*-Genes Dev. 10, 1557–1567.end formation in plants. EMBO J. 13, 2200–2210.
Zijlstra, C., and Hohn, T. (1992). Cauliflower mosaic virus gene VI con-Sanfacon, H., Brodman, P., and Hohn, T. (1991). A dissection of thetrols translation from dicistronic expression units in transgenic Arabi-cauliflower mosaic virus RNA polyadenylation signal. Genes Dev. 5,
141–149. dopsis plants. Plant Cell 4, 1471–1484.
AID VY 8250 / 6a25$$$681 11-13-96 14:39:11 viras AP: Virology
Related Documents











