of February 6, 2015. This information is current as Cells Basophilic Leukemia Mast Cells and Jurkat T Tyrosine Phosphatase PTP-MEG2 in Rat Enlargement of Secretory Vesicles by Protein Tomas Mustelin Edvard Monosov, Mats Stridsberg, Minoru Fukuda and Xiaodong Wang, Huong Huynh, Anette Gjörloff-Wingren, http://www.jimmunol.org/content/168/9/4612 doi: 10.4049/jimmunol.168.9.4612 2002; 168:4612-4619; ; J Immunol References http://www.jimmunol.org/content/168/9/4612.full#ref-list-1 , 18 of which you can access for free at: cites 35 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2002 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on February 6, 2015 http://www.jimmunol.org/ Downloaded from by guest on February 6, 2015 http://www.jimmunol.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

of February 6, 2015.This information is current as
CellsBasophilic Leukemia Mast Cells and Jurkat TTyrosine Phosphatase PTP-MEG2 in Rat Enlargement of Secretory Vesicles by Protein
Tomas MustelinEdvard Monosov, Mats Stridsberg, Minoru Fukuda and Xiaodong Wang, Huong Huynh, Anette Gjörloff-Wingren,
http://www.jimmunol.org/content/168/9/4612doi: 10.4049/jimmunol.168.9.4612
2002; 168:4612-4619; ;J Immunol
Referenceshttp://www.jimmunol.org/content/168/9/4612.full#ref-list-1
, 18 of which you can access for free at: cites 35 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2002 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on February 6, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on February 6, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

Enlargement of Secretory Vesicles by Protein TyrosinePhosphatase PTP-MEG2 in Rat Basophilic Leukemia MastCells and Jurkat T Cells1
Xiaodong Wang,2* Huong Huynh,* Anette Gjorloff-Wingren,3* Edvard Monosov,†
Mats Stridsberg,§ Minoru Fukuda,‡ and Tomas Mustelin4*
Stimulus-induced secretion of bioactive polypeptides is a fundamental aspect of the immune system. Secretory proteins aresynthesized in the endoplasmic reticulum and are transported through the Golgi apparatus to the trans-Golgi network, where theyare sorted into transport vesicles that bud off and fuse into condensing vacuoles, which subsequently undergo an editing andconcentration process to become mature secretory vesicles. In this study, we report that the PTP-MEG2 protein tyrosine phos-phatase is located on these vesicles in mast cells. Expression of PTP-MEG2 caused a striking enlargement of these vesicles in bothrat basophilic leukemia mast cells and Jurkat T leukemia cells into giant vesicles with diameters of up to several micrometers. Thefused vesicles did not acquire markers for other compartments and were adjacent to the trans-Golgi network, contained car-boxypeptidase E, chromogranin C, and IL-2, and had an electron-dense core typical of secretory vesicles. Expression of PTP-MEG2 also caused a reduction in the secretion of IL-2 from stimulated Jurkat cells. The effects of PTP-MEG2 on secretory vesiclesrequired the catalytic activity of PTP-MEG2 and was rapidly reversed by pervanadate. We propose that PTP-MEG2 representsa novel connection between tyrosine dephosphorylation and the regulation of secretory vesicles in hematopoietic cells. TheJournal of Immunology, 2002, 168: 4612–4619.
T he reversible modification of cellular proteins by phos-phorylation of the hydroxyl group of specific tyrosine res-idues is a widely used mechanism for regulation of cel-
lular proliferation, differentiation, and signal transduction (1–3).Protein tyrosine phosphorylation is catalyzed by protein tyrosinekinases, many of which are crucial for the development and dif-ferentiation of hematopoietic cell lineages and organs (3–5), and,conversely, aberrantly expressed or regulated in many types ofretroviral or spontaneous neoplastic diseases (3, 6–8). Tyrosinephosphorylation is counteracted by cellular protein tyrosine phos-phatases (PTPases),5 which are generally considered to be antion-cogenic and to promote cell differentiation (2). PTPases are a di-verse family of enzymes with a conserved 280-aa catalytic domain,but widely differing N- and C-termini, which often fulfill bothtargeting and regulatory functions for the holoenzyme (9, 10).
These noncatalytic regions often contain protein-protein interac-tion domains, such as Src homology 2 domains, PSD-95, Dlg, Z01homology domains or modules found in cytoskeletal proteins.There are also PTPases with nuclear localization or endoplasmicreticulum retention signals (9, 10).
The 68-kDa PTP-MEG2 (11) is the sole mammalian represen-tative of a group of PTPases that contain a unique 250-aa N-ter-minal region with homology to cellular retinaldehyde-binding pro-tein and Sec14p, a yeast protein with phosphatidylinositol transferactivity. Two PTPases related to PTP-MEG2, termed PTPX1 andPTPX10, have been cloned from Xenopus laevis (12). In addition,Sec14p homology domains are also found in �-tocopherol-bindingprotein (13), a 45-kDa secretory protein from olfactory epithelium(14), human Sec14p homologs (15), and a large number of otherproteins from yeast and plants to humans (16). A common themeamong these proteins is binding of hydrophobic molecules andinvolvement in secretion.
Very little is known about the biology of PTP-MEG2. The en-zyme is expressed in many cell types (11), including at low levelsin Jurkat T cells (17), and its expression is induced during mono-cyte/macrophage differentiation of U937 cells (18). We have re-cently found that PTP-MEG2 differed from 13 other tested intra-cellular PTPases in that its expression resulted in its accumulationin large intracellular vesicles of unknown nature (17). In this study,we demonstrate that these vesicles represent secretory vesicles,which fuse and swell in a manner that depends on the catalyticactivity of PTP-MEG2. Based on our findings, we discuss a pos-sible physiological function of PTP-MEG2 in the biogenesis ofsecretory vesicles.
Materials and MethodsAbs and reagents
The anti-influenza hemagglutinin tag epitope (HA) mAb 12CA5 conju-gated to FITC or rhodamine isothiocyanate (TRITC) were from RocheMolecular Biochemicals (Indianapolis, IN). The 16B12 anti-HA from
*Program of Signal Transduction, †Cell Analysis and Histology Facility, and ‡Gly-cobiology Program, Cancer Research Center, The Burnham Institute, La Jolla, CA92037; and §Department of Clinical Chemistry, Uppsala University Hospital, Upp-sala, Sweden
Received for publication August 30, 2001. Accepted for publication February27, 2002.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by a fellowship from the Swedish Cancer Foundation (toA.G.-W.); and Grants CA48737 (to M.F.), AI35603 (to T.M.), AI41481 (to T.M.),and AI40552 (to T.M.) from the National Institutes of Health.2 Current address: Gene Therapy Inc., San Diego, CA 92121.3 Current address: Department of Medical Microbiology, Malmo University Hospital,Lund University, SE-205 02 Malmo, Sweden.4 Address correspondence and reprint requests to Dr. Tomas Mustelin, Program ofSignal Transduction, Cancer Research Center, The Burnham Institute, 10901 NorthTorrey Pines Road, La Jolla, CA 92037. E-mail address: [email protected] Abbreviations used in this paper: PTPase, protein tyrosine phosphatase; HA,hemagglutinin tag epitope; RBL, rat basophilic leukemia; TRITC, rhodamineisothiocyanate.
Copyright © 2002 by The American Association of Immunologists 0022-1767/02/$02.00
by guest on February 6, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
BAbCO (Richmond, CA) was used for immunoblotting. Anti-carboxypep-tidase E was from Transduction Laboratories (Lexington, KY). FITC-con-jugated rabbit anti-IL-2 was from Alexis (San Diego, CA). Goat anti-rabbitIgG (F(ab)2)-FITC was from Pierce (Rockford, IL). Anti-Lamp-1 (931B)antiserum, anti-Lamp-2 (932B), and anti-TGN46/51 polyclonal Abs havebeen described (19, 20). Anti-chromogranin antisera were from M. Strids-berg (21). A polyclonal Ab against PTP-MEG2 was generated against aGST N-terminal MEG2 fusion protein as described (22). Another anti-serum was raised against a synthetic peptide corresponding to the last 30 aaof PTP-MEG2 conjugated to keyhole limpet hemocyanin. A third anti-serum was raised against the internal peptide HSYRETRRKEGIVK, cor-responding to aa 66–79 of human PTP-MEG2.
Plasmid construction
The cDNA for PTP-MEG2 was kindly provided by P. Majerus (Washing-ton University, St. Louis, MO). It was subcloned in to the pEF/HA vector(23), which adds an HA to the N terminus of the cloned insert. The cata-lytically inactive mutant PTP-MEG2-C515S was generated using theQuickChange site-directed mutagenesis kit (Stratagene, La Jolla, CA). Themutation was verified by nucleotide sequencing.
Cells and transient transfection
PBLs were isolated from buffy coats from healthy volunteers by gradientcentrifugation. The cells were cultured in RPMI medium with 10% heat-inactivated FCS, 10 �g/ml PHA, and 100 U/ml of IL-2 for 72 h before use.Rat basophilic leukemia (RBL), MCP5 mast cells, and Jurkat cells werekept at logarithmic growth in RPMI supplemented with 10% heat-inacti-vated FCS, L-glutamine, and antibiotics. Transient transfections were con-ducted by electroporation as described before (24–27). Electroporationconditions typically contained 20 � 106 cells and a total of 1–10 �g ofplasmid DNA, and in each transfection the DNA amount was kept constant
by the addition of empty vector. Cells were used for experiments 48 h aftertransfection.
Immunoblots and immunoprecipitation
Immunoprecipitation was performed as before (24–28). Immunoblots weredeveloped by the ECL technique (ECL kit; Amersham Pharmacia Biotech,Little Chalfont, U.K.) according to the manufacturer’s instructions.
Confocal microscopy
Double immunofluorescence staining was done as before (17). Briefly,cells were washed in PBS and fixed in freshly made 3.7% formaldehyde.Fixed cells were permeabilized with 0.05% saponin, 0.5% BSA in PBS for10 min at room temperature, and then incubated with primary and second-ary Ab diluted in the same buffer for 1 h each at room temperature. Afterthree washes with PBS, the cells were mounted onto glass slides andviewed under a confocal laser scanning microscopy MRC-1024 (Bio-Rad,Hercules, CA). A differential interference contrast image was also taken ofmost cells.
Electron microscopy
Cells were fixed in 3.7% paraformaldehyde and 0.2% glutaraldehyde inPHEM buffer (60 mM PIPES, 25 mM HEPES (pH 6.9), 10 mM EGTA, 2mM Mg2SO4, and 0.02% NaN3) for 45 min at room temperature. Cellswere washed three times with 0.002% glycine in PHEM buffer, and werepostfixed in 1% (w/v) osmium tetroxide in 0.2 M cacodylate (pH 7.2),washed in distilled water, and dehydrated successively in ethanol. Dehy-drated pellets were infused with propylene oxide and embedded in Epon812. Ultrathin sections were obtained on a Reichert-Jung microtome, anddouble stained with uranyl acetate and lead citrate (29). Specimens wereexamined in a Hitachi H-600 electron microscope (Hitachi, Tokyo, Japan)at 75 kV with magnifications ranging from �3,000 to �100,000.
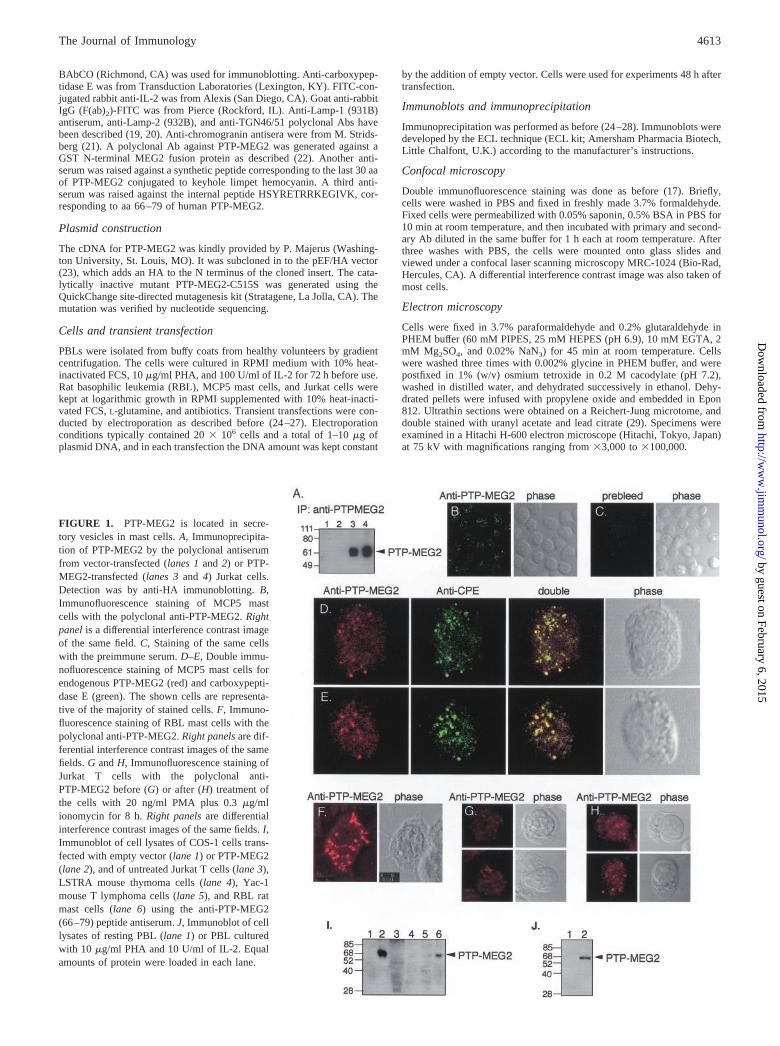
FIGURE 1. PTP-MEG2 is located in secre-tory vesicles in mast cells. A, Immunoprecipita-tion of PTP-MEG2 by the polyclonal antiserumfrom vector-transfected (lanes 1 and 2) or PTP-MEG2-transfected (lanes 3 and 4) Jurkat cells.Detection was by anti-HA immunoblotting. B,Immunofluorescence staining of MCP5 mastcells with the polyclonal anti-PTP-MEG2. Rightpanel is a differential interference contrast imageof the same field. C, Staining of the same cellswith the preimmune serum. D–E, Double immu-nofluorescence staining of MCP5 mast cells forendogenous PTP-MEG2 (red) and carboxypepti-dase E (green). The shown cells are representa-tive of the majority of stained cells. F, Immuno-fluorescence staining of RBL mast cells with thepolyclonal anti-PTP-MEG2. Right panels are dif-ferential interference contrast images of the samefields. G and H, Immunofluorescence staining ofJurkat T cells with the polyclonal anti-PTP-MEG2 before (G) or after (H) treatment ofthe cells with 20 ng/ml PMA plus 0.3 �g/mlionomycin for 8 h. Right panels are differentialinterference contrast images of the same fields. I,Immunoblot of cell lysates of COS-1 cells trans-fected with empty vector (lane 1) or PTP-MEG2(lane 2), and of untreated Jurkat T cells (lane 3),LSTRA mouse thymoma cells (lane 4), Yac-1mouse T lymphoma cells (lane 5), and RBL ratmast cells (lane 6) using the anti-PTP-MEG2(66–79) peptide antiserum. J, Immunoblot of celllysates of resting PBL (lane 1) or PBL culturedwith 10 �g/ml PHA and 10 U/ml of IL-2. Equalamounts of protein were loaded in each lane.
4613The Journal of Immunology
by guest on February 6, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
IL-2 secretion assay
A total of 20 � 106 Jurkat T cells cotransfected with PTP-MEG2 or emptyvector and green fluorescent protein were cultured for 48 h, and then 2 �106 green fluorescent cells were sorted in a BD Biosciences FACStar flu-orescence-activated cell sorter with a 488-nm argon laser (BD Biosciences,Mountain View, CA) to �95% purity. A total of 0.33 � 106 sorted cells intriplicate were stimulated in 100 �l of medium with 50 ng/ml PMA plus0.3 �g/ml ionomycin for 12 h, and 20 �l of the supernatant used formeasurement of the amount of IL-2 using an ELISA kit (Roche MolecularBiochemicals). Results are given as picograms per milliliter of secretedIL-2 (by 66 � 103 cells).
ResultsEndogenous PTP-MEG2 is located on secretory vesicles in mastcells
A polyclonal antiserum was raised against the N terminus of hu-man PTP-MEG2. This antiserum immunoprecipitated PTP-MEG2(Fig. 1A) and reacted well with transfected PTP-MEG2 in immu-nofluorescence staining (Fig. 2, A–D). Indirect immunofluores-cence staining of MCP5 (Fig. 1B) or RBL (Fig. 1F) mast cells gaverise to a granular cytoplasmic staining. The preimmune serum didnot react with these structures (Fig. 1C). Double immunofluores-cence staining with Abs against carboxypeptidase E, an enzymetypically located in secretory vesicles (30) where it processes pro-hormones by cleaving C-terminal arginine-containing motifs (31,32), showed a great deal of colocalization (Fig. 1, D and E), sug-gesting that PTP-MEG2 is primarily located on secretory vesicles.In agreement with this notion, the antiserum reacted weakly withresting Jurkat T cells (Fig. 1G), which have very few secretoryvesicles, but considerably better with cells stimulated for 8 h withphorbol ester and ionomycin (Fig. 1H). By immunoblotting, lowlevels of PTP-MEG2 were detected in T cell lines and better levelsin RBL mast cells (Fig. 1I). Expression of PTP-MEG2 was alsoinduced in normal T lymphocytes by polyclonal activation withPHA and IL-2 (Fig. 1J).
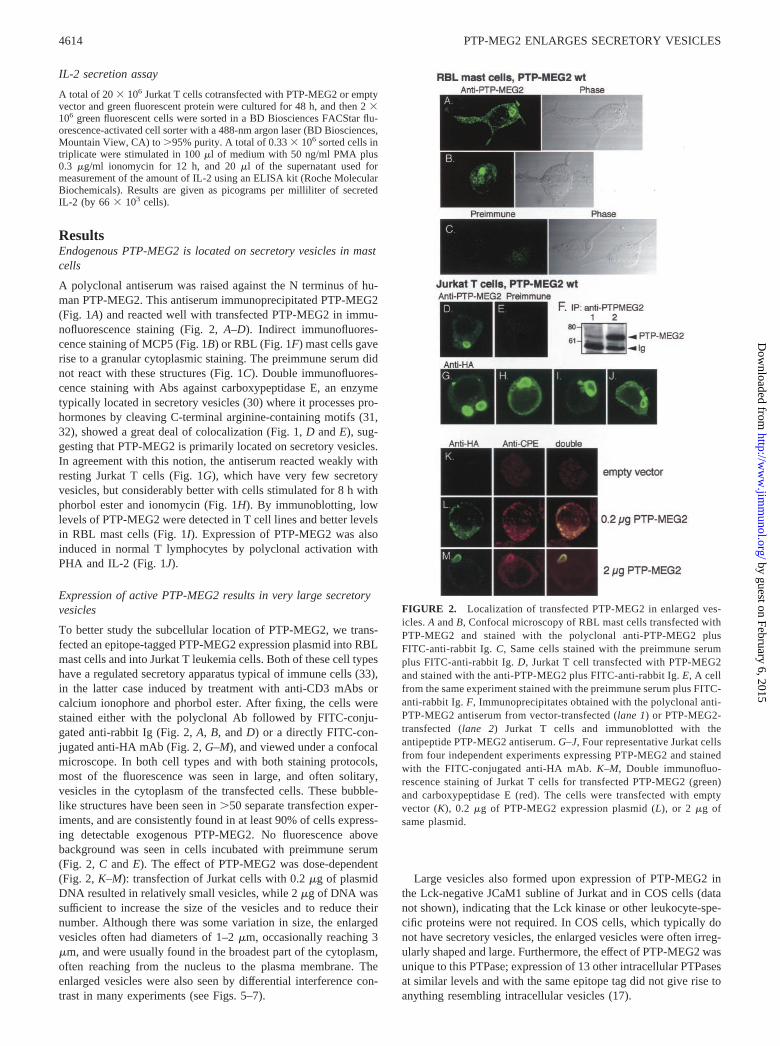
Expression of active PTP-MEG2 results in very large secretoryvesicles
To better study the subcellular location of PTP-MEG2, we trans-fected an epitope-tagged PTP-MEG2 expression plasmid into RBLmast cells and into Jurkat T leukemia cells. Both of these cell typeshave a regulated secretory apparatus typical of immune cells (33),in the latter case induced by treatment with anti-CD3 mAbs orcalcium ionophore and phorbol ester. After fixing, the cells werestained either with the polyclonal Ab followed by FITC-conju-gated anti-rabbit Ig (Fig. 2, A, B, and D) or a directly FITC-con-jugated anti-HA mAb (Fig. 2, G–M), and viewed under a confocalmicroscope. In both cell types and with both staining protocols,most of the fluorescence was seen in large, and often solitary,vesicles in the cytoplasm of the transfected cells. These bubble-like structures have been seen in �50 separate transfection exper-iments, and are consistently found in at least 90% of cells express-ing detectable exogenous PTP-MEG2. No fluorescence abovebackground was seen in cells incubated with preimmune serum(Fig. 2, C and E). The effect of PTP-MEG2 was dose-dependent(Fig. 2, K–M): transfection of Jurkat cells with 0.2 �g of plasmidDNA resulted in relatively small vesicles, while 2 �g of DNA wassufficient to increase the size of the vesicles and to reduce theirnumber. Although there was some variation in size, the enlargedvesicles often had diameters of 1–2 �m, occasionally reaching 3�m, and were usually found in the broadest part of the cytoplasm,often reaching from the nucleus to the plasma membrane. Theenlarged vesicles were also seen by differential interference con-trast in many experiments (see Figs. 5–7).
Large vesicles also formed upon expression of PTP-MEG2 inthe Lck-negative JCaM1 subline of Jurkat and in COS cells (datanot shown), indicating that the Lck kinase or other leukocyte-spe-cific proteins were not required. In COS cells, which typically donot have secretory vesicles, the enlarged vesicles were often irreg-ularly shaped and large. Furthermore, the effect of PTP-MEG2 wasunique to this PTPase; expression of 13 other intracellular PTPasesat similar levels and with the same epitope tag did not give rise toanything resembling intracellular vesicles (17).
FIGURE 2. Localization of transfected PTP-MEG2 in enlarged ves-icles. A and B, Confocal microscopy of RBL mast cells transfected withPTP-MEG2 and stained with the polyclonal anti-PTP-MEG2 plusFITC-anti-rabbit Ig. C, Same cells stained with the preimmune serumplus FITC-anti-rabbit Ig. D, Jurkat T cell transfected with PTP-MEG2and stained with the anti-PTP-MEG2 plus FITC-anti-rabbit Ig. E, A cellfrom the same experiment stained with the preimmune serum plus FITC-anti-rabbit Ig. F, Immunoprecipitates obtained with the polyclonal anti-PTP-MEG2 antiserum from vector-transfected (lane 1) or PTP-MEG2-transfected (lane 2) Jurkat T cells and immunoblotted with theantipeptide PTP-MEG2 antiserum. G–J, Four representative Jurkat cellsfrom four independent experiments expressing PTP-MEG2 and stainedwith the FITC-conjugated anti-HA mAb. K–M, Double immunofluo-rescence staining of Jurkat T cells for transfected PTP-MEG2 (green)and carboxypeptidase E (red). The cells were transfected with emptyvector (K), 0.2 �g of PTP-MEG2 expression plasmid (L), or 2 �g ofsame plasmid.
4614 PTP-MEG2 ENLARGES SECRETORY VESICLES
by guest on February 6, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
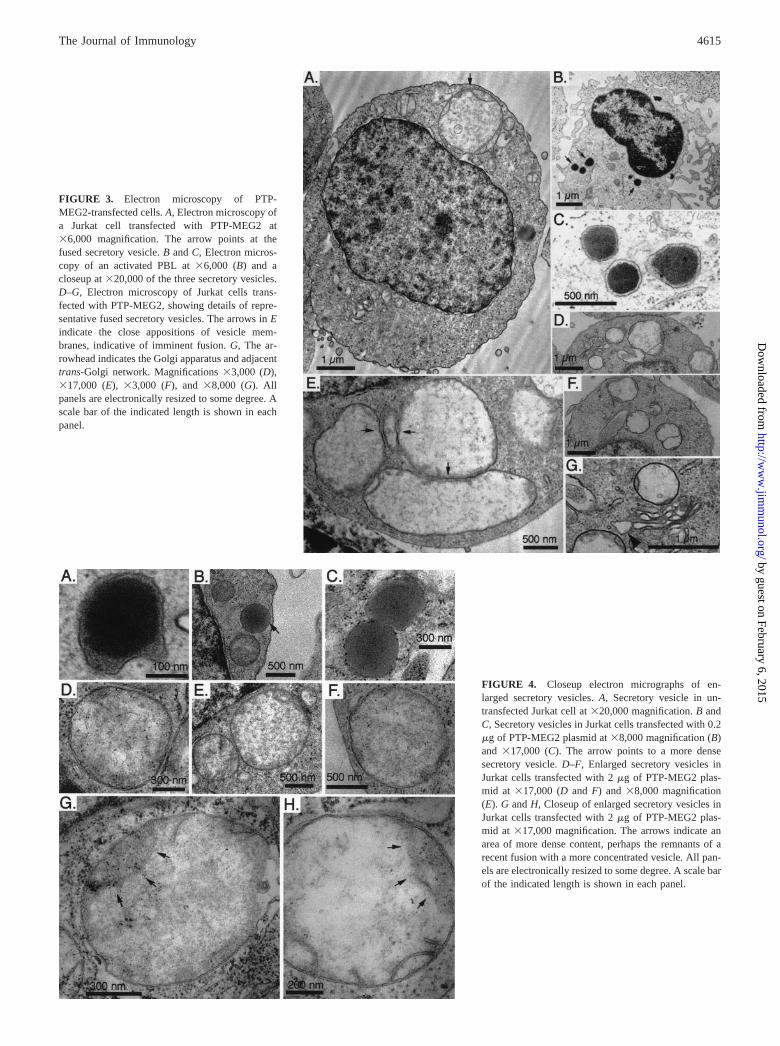
FIGURE 3. Electron microscopy of PTP-MEG2-transfected cells. A, Electron microscopy ofa Jurkat cell transfected with PTP-MEG2 at�6,000 magnification. The arrow points at thefused secretory vesicle. B and C, Electron micros-copy of an activated PBL at �6,000 (B) and acloseup at �20,000 of the three secretory vesicles.D–G, Electron microscopy of Jurkat cells trans-fected with PTP-MEG2, showing details of repre-sentative fused secretory vesicles. The arrows in Eindicate the close appositions of vesicle mem-branes, indicative of imminent fusion. G, The ar-rowhead indicates the Golgi apparatus and adjacenttrans-Golgi network. Magnifications �3,000 (D),�17,000 (E), �3,000 (F), and �8,000 (G). Allpanels are electronically resized to some degree. Ascale bar of the indicated length is shown in eachpanel.
FIGURE 4. Closeup electron micrographs of en-larged secretory vesicles. A, Secretory vesicle in un-transfected Jurkat cell at �20,000 magnification. B andC, Secretory vesicles in Jurkat cells transfected with 0.2�g of PTP-MEG2 plasmid at �8,000 magnification (B)and �17,000 (C). The arrow points to a more densesecretory vesicle. D–F, Enlarged secretory vesicles inJurkat cells transfected with 2 �g of PTP-MEG2 plas-mid at �17,000 (D and F) and �8,000 magnification(E). G and H, Closeup of enlarged secretory vesicles inJurkat cells transfected with 2 �g of PTP-MEG2 plas-mid at �17,000 magnification. The arrows indicate anarea of more dense content, perhaps the remnants of arecent fusion with a more concentrated vesicle. All pan-els are electronically resized to some degree. A scale barof the indicated length is shown in each panel.
4615The Journal of Immunology
by guest on February 6, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
Ultrastructure of PTP-MEG2-induced large secretory vesicles
Electron microscopy of Jurkat cells transfected with PTP-MEG2showed that the enlarged vesicles were membrane-enclosed vesi-cles with an amorphous content typical of secretory vesicles (Figs.3 and 4). In cells with very large vesicles, the amorphous contentwas diluted and dispersed throughout the lumen, while in smallervesicles it remained more densely packed. For comparison, Fig. 3Bshows an electron micrograph of an activated blood T lymphocytewith normal secretory vesicles with diameters of 200–300 nm(Fig. 3C).
The very large vesicles seen in Jurkat T cells expressing PTP-MEG2 were frequently adjacent to each other, and they were oftenattached to each other through a portion of their enclosing mem-brane (Fig. 3E). In addition, many vesicles contained signs of re-cent fusion with other vesicles in the form of rings inside theenclosing membrane (Figs. 3, A, D, and F, and 4, G and H). Oncloser inspection, these rings consisted of the amorphous secretoryvesicle contents (Fig. 4, G and H), not lipid bilayer. The enlargedvesicles were often found adjacent to the trans-Golgi cisternae(Fig. 3G). Taken together, these electron microscopy studies sug-gest that PTP-MEG2 causes a striking increase in fusion and/orlack of fission within the secretory vesicle compartment, resultingin giant vesicles with diluted contents.
Lack of markers for other organelles in the fused secretoryvesicles
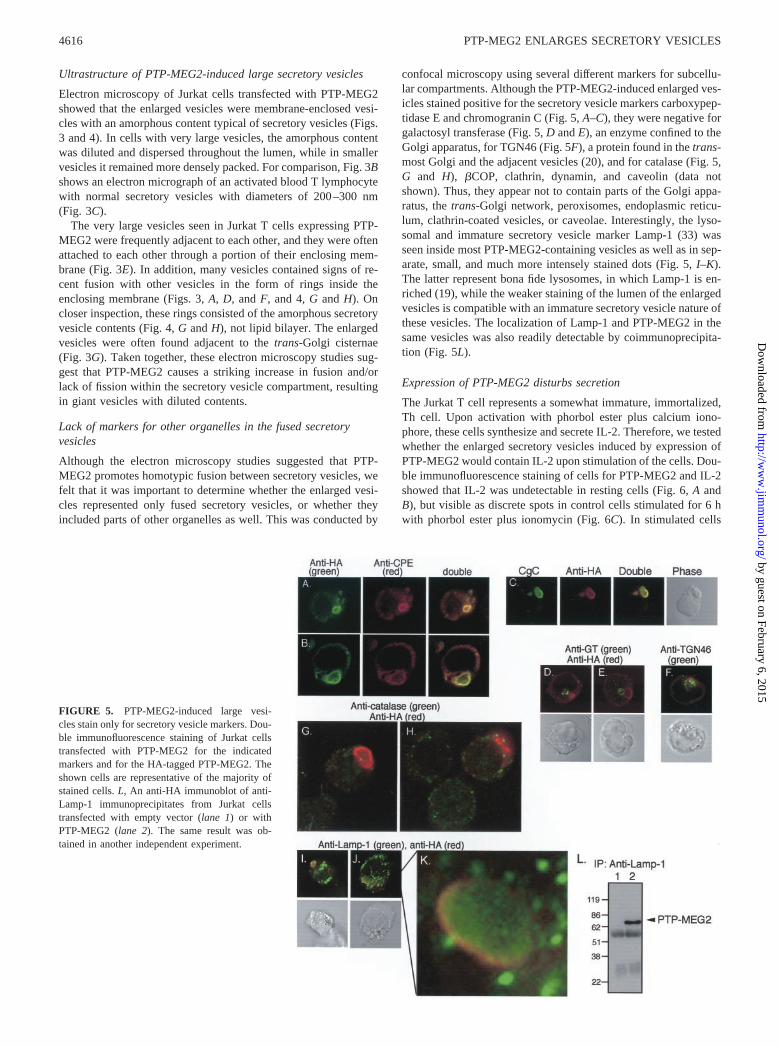
Although the electron microscopy studies suggested that PTP-MEG2 promotes homotypic fusion between secretory vesicles, wefelt that it was important to determine whether the enlarged vesi-cles represented only fused secretory vesicles, or whether theyincluded parts of other organelles as well. This was conducted by
confocal microscopy using several different markers for subcellu-lar compartments. Although the PTP-MEG2-induced enlarged ves-icles stained positive for the secretory vesicle markers carboxypep-tidase E and chromogranin C (Fig. 5, A–C), they were negative forgalactosyl transferase (Fig. 5, D and E), an enzyme confined to theGolgi apparatus, for TGN46 (Fig. 5F), a protein found in the trans-most Golgi and the adjacent vesicles (20), and for catalase (Fig. 5,G and H), �COP, clathrin, dynamin, and caveolin (data notshown). Thus, they appear not to contain parts of the Golgi appa-ratus, the trans-Golgi network, peroxisomes, endoplasmic reticu-lum, clathrin-coated vesicles, or caveolae. Interestingly, the lyso-somal and immature secretory vesicle marker Lamp-1 (33) wasseen inside most PTP-MEG2-containing vesicles as well as in sep-arate, small, and much more intensely stained dots (Fig. 5, I–K).The latter represent bona fide lysosomes, in which Lamp-1 is en-riched (19), while the weaker staining of the lumen of the enlargedvesicles is compatible with an immature secretory vesicle nature ofthese vesicles. The localization of Lamp-1 and PTP-MEG2 in thesame vesicles was also readily detectable by coimmunoprecipita-tion (Fig. 5L).
Expression of PTP-MEG2 disturbs secretion
The Jurkat T cell represents a somewhat immature, immortalized,Th cell. Upon activation with phorbol ester plus calcium iono-phore, these cells synthesize and secrete IL-2. Therefore, we testedwhether the enlarged secretory vesicles induced by expression ofPTP-MEG2 would contain IL-2 upon stimulation of the cells. Dou-ble immunofluorescence staining of cells for PTP-MEG2 and IL-2showed that IL-2 was undetectable in resting cells (Fig. 6, A andB), but visible as discrete spots in control cells stimulated for 6 hwith phorbol ester plus ionomycin (Fig. 6C). In stimulated cells
FIGURE 5. PTP-MEG2-induced large vesi-cles stain only for secretory vesicle markers. Dou-ble immunofluorescence staining of Jurkat cellstransfected with PTP-MEG2 for the indicatedmarkers and for the HA-tagged PTP-MEG2. Theshown cells are representative of the majority ofstained cells. L, An anti-HA immunoblot of anti-Lamp-1 immunoprecipitates from Jurkat cellstransfected with empty vector (lane 1) or withPTP-MEG2 (lane 2). The same result was ob-tained in another independent experiment.
4616 PTP-MEG2 ENLARGES SECRETORY VESICLES
by guest on February 6, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
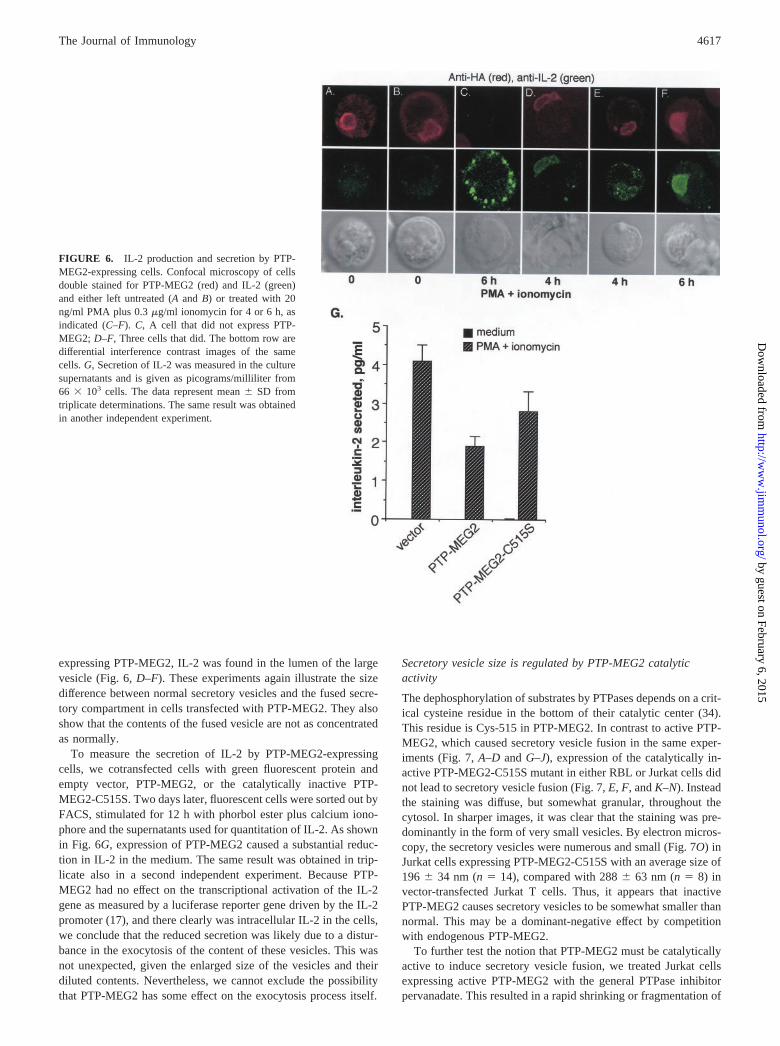
expressing PTP-MEG2, IL-2 was found in the lumen of the largevesicle (Fig. 6, D–F). These experiments again illustrate the sizedifference between normal secretory vesicles and the fused secre-tory compartment in cells transfected with PTP-MEG2. They alsoshow that the contents of the fused vesicle are not as concentratedas normally.
To measure the secretion of IL-2 by PTP-MEG2-expressingcells, we cotransfected cells with green fluorescent protein andempty vector, PTP-MEG2, or the catalytically inactive PTP-MEG2-C515S. Two days later, fluorescent cells were sorted out byFACS, stimulated for 12 h with phorbol ester plus calcium iono-phore and the supernatants used for quantitation of IL-2. As shownin Fig. 6G, expression of PTP-MEG2 caused a substantial reduc-tion in IL-2 in the medium. The same result was obtained in trip-licate also in a second independent experiment. Because PTP-MEG2 had no effect on the transcriptional activation of the IL-2gene as measured by a luciferase reporter gene driven by the IL-2promoter (17), and there clearly was intracellular IL-2 in the cells,we conclude that the reduced secretion was likely due to a distur-bance in the exocytosis of the content of these vesicles. This wasnot unexpected, given the enlarged size of the vesicles and theirdiluted contents. Nevertheless, we cannot exclude the possibilitythat PTP-MEG2 has some effect on the exocytosis process itself.
Secretory vesicle size is regulated by PTP-MEG2 catalyticactivity
The dephosphorylation of substrates by PTPases depends on a crit-ical cysteine residue in the bottom of their catalytic center (34).This residue is Cys-515 in PTP-MEG2. In contrast to active PTP-MEG2, which caused secretory vesicle fusion in the same exper-iments (Fig. 7, A–D and G–J), expression of the catalytically in-active PTP-MEG2-C515S mutant in either RBL or Jurkat cells didnot lead to secretory vesicle fusion (Fig. 7, E, F, and K–N). Insteadthe staining was diffuse, but somewhat granular, throughout thecytosol. In sharper images, it was clear that the staining was pre-dominantly in the form of very small vesicles. By electron micros-copy, the secretory vesicles were numerous and small (Fig. 7O) inJurkat cells expressing PTP-MEG2-C515S with an average size of196 � 34 nm (n � 14), compared with 288 � 63 nm (n � 8) invector-transfected Jurkat T cells. Thus, it appears that inactivePTP-MEG2 causes secretory vesicles to be somewhat smaller thannormal. This may be a dominant-negative effect by competitionwith endogenous PTP-MEG2.
To further test the notion that PTP-MEG2 must be catalyticallyactive to induce secretory vesicle fusion, we treated Jurkat cellsexpressing active PTP-MEG2 with the general PTPase inhibitorpervanadate. This resulted in a rapid shrinking or fragmentation of
FIGURE 6. IL-2 production and secretion by PTP-MEG2-expressing cells. Confocal microscopy of cellsdouble stained for PTP-MEG2 (red) and IL-2 (green)and either left untreated (A and B) or treated with 20ng/ml PMA plus 0.3 �g/ml ionomycin for 4 or 6 h, asindicated (C–F). C, A cell that did not express PTP-MEG2; D–F, Three cells that did. The bottom row aredifferential interference contrast images of the samecells. G, Secretion of IL-2 was measured in the culturesupernatants and is given as picograms/milliliter from66 � 103 cells. The data represent mean � SD fromtriplicate determinations. The same result was obtainedin another independent experiment.
4617The Journal of Immunology
by guest on February 6, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
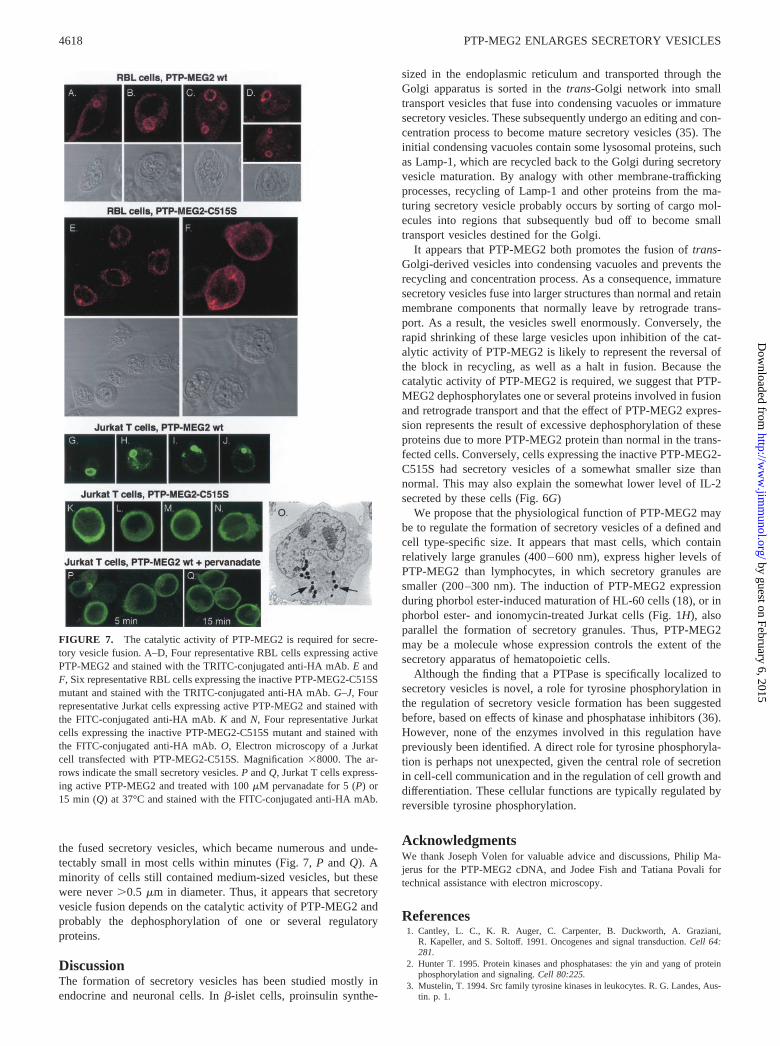
the fused secretory vesicles, which became numerous and unde-tectably small in most cells within minutes (Fig. 7, P and Q). Aminority of cells still contained medium-sized vesicles, but thesewere never �0.5 �m in diameter. Thus, it appears that secretoryvesicle fusion depends on the catalytic activity of PTP-MEG2 andprobably the dephosphorylation of one or several regulatoryproteins.
DiscussionThe formation of secretory vesicles has been studied mostly inendocrine and neuronal cells. In �-islet cells, proinsulin synthe-
sized in the endoplasmic reticulum and transported through theGolgi apparatus is sorted in the trans-Golgi network into smalltransport vesicles that fuse into condensing vacuoles or immaturesecretory vesicles. These subsequently undergo an editing and con-centration process to become mature secretory vesicles (35). Theinitial condensing vacuoles contain some lysosomal proteins, suchas Lamp-1, which are recycled back to the Golgi during secretoryvesicle maturation. By analogy with other membrane-traffickingprocesses, recycling of Lamp-1 and other proteins from the ma-turing secretory vesicle probably occurs by sorting of cargo mol-ecules into regions that subsequently bud off to become smalltransport vesicles destined for the Golgi.
It appears that PTP-MEG2 both promotes the fusion of trans-Golgi-derived vesicles into condensing vacuoles and prevents therecycling and concentration process. As a consequence, immaturesecretory vesicles fuse into larger structures than normal and retainmembrane components that normally leave by retrograde trans-port. As a result, the vesicles swell enormously. Conversely, therapid shrinking of these large vesicles upon inhibition of the cat-alytic activity of PTP-MEG2 is likely to represent the reversal ofthe block in recycling, as well as a halt in fusion. Because thecatalytic activity of PTP-MEG2 is required, we suggest that PTP-MEG2 dephosphorylates one or several proteins involved in fusionand retrograde transport and that the effect of PTP-MEG2 expres-sion represents the result of excessive dephosphorylation of theseproteins due to more PTP-MEG2 protein than normal in the trans-fected cells. Conversely, cells expressing the inactive PTP-MEG2-C515S had secretory vesicles of a somewhat smaller size thannormal. This may also explain the somewhat lower level of IL-2secreted by these cells (Fig. 6G)
We propose that the physiological function of PTP-MEG2 maybe to regulate the formation of secretory vesicles of a defined andcell type-specific size. It appears that mast cells, which containrelatively large granules (400–600 nm), express higher levels ofPTP-MEG2 than lymphocytes, in which secretory granules aresmaller (200–300 nm). The induction of PTP-MEG2 expressionduring phorbol ester-induced maturation of HL-60 cells (18), or inphorbol ester- and ionomycin-treated Jurkat cells (Fig. 1H), alsoparallel the formation of secretory granules. Thus, PTP-MEG2may be a molecule whose expression controls the extent of thesecretory apparatus of hematopoietic cells.
Although the finding that a PTPase is specifically localized tosecretory vesicles is novel, a role for tyrosine phosphorylation inthe regulation of secretory vesicle formation has been suggestedbefore, based on effects of kinase and phosphatase inhibitors (36).However, none of the enzymes involved in this regulation havepreviously been identified. A direct role for tyrosine phosphoryla-tion is perhaps not unexpected, given the central role of secretionin cell-cell communication and in the regulation of cell growth anddifferentiation. These cellular functions are typically regulated byreversible tyrosine phosphorylation.
AcknowledgmentsWe thank Joseph Volen for valuable advice and discussions, Philip Ma-jerus for the PTP-MEG2 cDNA, and Jodee Fish and Tatiana Povali fortechnical assistance with electron microscopy.
References1. Cantley, L. C., K. R. Auger, C. Carpenter, B. Duckworth, A. Graziani,
R. Kapeller, and S. Soltoff. 1991. Oncogenes and signal transduction. Cell 64:281.
2. Hunter T. 1995. Protein kinases and phosphatases: the yin and yang of proteinphosphorylation and signaling. Cell 80:225.
3. Mustelin, T. 1994. Src family tyrosine kinases in leukocytes. R. G. Landes, Aus-tin. p. 1.
FIGURE 7. The catalytic activity of PTP-MEG2 is required for secre-tory vesicle fusion. A–D, Four representative RBL cells expressing activePTP-MEG2 and stained with the TRITC-conjugated anti-HA mAb. E andF, Six representative RBL cells expressing the inactive PTP-MEG2-C515Smutant and stained with the TRITC-conjugated anti-HA mAb. G–J, Fourrepresentative Jurkat cells expressing active PTP-MEG2 and stained withthe FITC-conjugated anti-HA mAb. K and N, Four representative Jurkatcells expressing the inactive PTP-MEG2-C515S mutant and stained withthe FITC-conjugated anti-HA mAb. O, Electron microscopy of a Jurkatcell transfected with PTP-MEG2-C515S. Magnification �8000. The ar-rows indicate the small secretory vesicles. P and Q, Jurkat T cells express-ing active PTP-MEG2 and treated with 100 �M pervanadate for 5 (P) or15 min (Q) at 37°C and stained with the FITC-conjugated anti-HA mAb.
4618 PTP-MEG2 ENLARGES SECRETORY VESICLES
by guest on February 6, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

4. Molina, T. J., K. Kishihara, D. P. Siderowski, W. van Ewijk, A. Narendran,E. Timms, A. Wakeham, C. J. Paige, K.-U. Hartman, A. Veillette, et al. 1992.Profound block in thymocyte development in mice lacking p56lck. Nature 357:161.
5. Arpaia, E., M. Shahar, H. Dadi, A. Cohen, and C. M. Roifman. 1994. DefectiveT cell receptor signaling and CD8� thymic selection in humans lacking Zap-70kinase. Cell 76:947.
6. Croce, C. M., and P. C. Nowell. 1985. Molecular basis of human B cell neoplasia.Blood 65:1.
7. Tycko, B., S. D. Smith, and J. Sklar. 1991. Chromosomal translocations joiningLCK and TCRB loci in human T cell leukemia. J. Exp. Med. 174:867.
8. Fargnoli, M.-C., R. L. Edelson, C. L. Berger, S. Chimenti, C. Couture,T. Mustelin, and R. Halaban. 1997. TCR diminished signaling in cutaneous T-celllymphoma is associated with decreased kinase activities of Syk and membrane-associated Csk. Leukemia 11:1338.
9. Tonks, N. K., and B. G. Neel. 1996. From form to function: signaling by proteintyrosine phosphatases. Cell 87:365.
10. Mustelin, T., J. Brockdorff, A. Gjorloff-Wingren, P. Tailor, S. Han, X. Wang, andM. Saxena. 1998. T cell activation: the coming of the phosphatases. Front. Biosci.3:1060.
11. Gu, M., I. Warshawsky, and P. W. Majerus. 1992. Cloning and expression of acytosolic megakaryocyte protein-tyrosine-phosphatase with sequence homologyto retinaldehyde-binding protein and yeast SEC14p. Proc. Natl. Acad. Sci. USA89:2980.
12. Del Vecchio, R. L., and N. K. Tonks. 1994. Characterization of two structurallyrelated Xenopus laevis protein tyrosine phosphatases with homology to lipid-binding proteins. J. Biol. Chem. 269:19639.
13. Sato, Y., H. Arai, A. Miyata, S. Tokita, K. Yamamoto, T. Tanabe, and K. Inoue.1993. Primary structure of �-tocopherol transfer protein from rat liver: homologywith cellular retinaldehyde-binding protein. J. Biol. Chem. 268:17705.
14. Merkulova, M. I., S. G. Andreeva, T. M. Shuvaeva, S. V. Novoselov,I. V. Peshenko, M. F. Bystrova, V. I. Novoselov, E. E. Fesenko, andV. M. Lipkin. 1999. A novel 45 kDa secretory protein from rat olfactory epithe-lium: primary structure and localisation. FEBS Lett. 450:126.
15. Chinen, K., E. Takahashi, and Y. Nakamura. 1996. Isolation and mapping of ahuman gene (SEC14L), partially homologous to yeast SEC14, that contains avariable number of tandem repeats (VNTR) site in its 3� untranslated region.Cytogenet. Cell Genet. 73:218.
16. Aravind, L., A. F. Neuwald, and C. P. Ponting. 1999. Sec14p-like domains inNF1 and Dbl-like proteins indicate lipid regulation of Ras and Rho signaling.Curr. Biol. 9:195.
17. Gjorloff-Wingren, A., P. Oh, M. Saxena, S. Han, X. Wang, S. Williams,J. Schnitzer, and T. Mustelin. 2000. Subcellular localization of intracellular pro-tein tyrosine phosphatases in T cells. Eur. J. Immunol. 30:2412.
18. Seimiya, H., T. Sawabe, M. Toho, and T. Tsuro. 1995. Phorbol ester-resistantmonoblastoid leukemia cells with a functional mitogen-activated protein kinasecascade but without responsive protein tyrosine phosphatases. Oncogene11:2047.
19. Carlsson, S. R., J. Roth, F. Piller, and M. Fukuda. 1988. Isolation and charac-terization of human lysosomal membrane glycoproteins, h-lamp-1 and h-lamp-2.J. Biol. Chem. 263:18911.
20. Kain, R., K. Angata, D. Kerjaschki, and M. Fukuda. 1998. Molecular cloning andexpression of a novel human trans-Golgi network glycoprotein, TGN51, thatcontains multiple tyrosine-containing motifs. J. Biol. Chem. 273:981.
21. Stridsberg, M., K. Oberg, Q. Li, U. Engstrom, and G. Lundqvist. 1995. Mea-surement of chromogranin A, chromogranin B (secretogranin 1), chromogranin C(secretogranin 11) and pancreastatin in plasma and urine of patients with carci-noid tumours. J. Endocrinol. 144:49.
22. Saxena, M., S. Williams, J. Gilman, and T. Mustelin. 1998. Negative regulationof T cell antigen receptor signaling by hematopoietic tyrosine phosphatase(HePTP). J. Biol. Chem. 273:15340.
23. von Willebrand, M., T. Jascur, N. Bonnefoy Berard, H. Yano, A. Altman,Y. Matsuda, and T. Mustelin. 1996. Inhibition of phosphatidylinositol 3-kinaseblocks TCR/CD3-induced activation of the mitogen-activated kinase Erk2. Eur.J. Biochem. 235:828.
24. Couture, C., Z. Songyang, T. Jascur, S. Williams, P. Tailor, L. C. Cantley, andT. Mustelin. 1996. Regulation of the Lck SH2 domain by tyrosine phosphoryla-tion. J. Biol. Chem. 271:24880.
25. Jascur, T., J. Gilman, and T. Mustelin. 1997. Involvement of phosphatidylinositol3-kinase in NFAT activation in T cells. J. Biol. Chem. 272:14483.
26. Saxena, M., S. Williams, J. Brockdorff, J. Gilman, and T. Mustelin. 1999. Inhi-bition of T cell signaling by MAP kinase-targeted hematopoietic tyrosine phos-phatase (HePTP). J. Biol. Chem. 274:11693.
27. Saxena, M., S. Williams, K. Tasken, and T. Mustelin. 1999. Crosstalk betweencAMP-dependent kinase and MAP kinase through hematopoietic protein tyrosinephosphatase (HePTP). Nat. Cell Biol. 1:305.
28. Couture, C., G. Baier, A. Altman, and T. Mustelin. 1994. p56lck-independentactivation and tyrosine phosphorylation of p72syk by TCR/CD3 stimulation. Proc.Natl. Acad. Sci. USA 91:5301.
29. Reynolds, E. S. 1963. The use of lead citrate at high pH as an electronopaquestain in electron microscopy. J. Cell Biol. 17:208.
30. Cool, D. R., E. Normant, F. Shen, H. C. Chen, L. Pannell, Y. Zhang, andY. P. Loh. 1997. Carboxypeptidase E is a regulated secretory pathway sortingreceptor: genetic obliteration leads to endocrine disorders in Cpe(fat) mice. Cell88:73.
31. Manser, E., D. Fernandez, L. Loo, P. Y. Goh, C. Monfries, C. Hall, and L. Lim.1990. Human carboxypeptidase E: isolation and characterization of the cDNA,sequence conservation, expression and processing in vitro. Biochem. J. 267:517.
32. Naggert, J. K., L. D. Fricker, O. Varmalov, P. M. Nishina, Y. Rouille,D. F. Steiner, R. J. Carroll, B. J. Paigen, and E. H. Leiter. 1995. Hyperproinsu-linaemia in obese fat/fat mice associated with a carboxypeptidase E mutationwhich reduces enzyme activity. Nat. Genet. 10:135.
33. Stinchcombe, J. C., and G. M. Griffiths. 1999. Regulated secretion from hema-topoietic cells. J. Cell Biol. 147:1.
34. Barford, D., A. J. Flint, and N. K. Tonks. 1994. Crystal structure of humanprotein tyrosine phosphatase 1B. Science 263:1397.
35. Huang, X. F., and P. Arvan. 1994. Formation of the insulin-containing secretorygranule core occurs within immature �-granules. J. Biol. Chem. 269:20838.
36. Austin, C. D., and D. Shields. 1996. Formation of nascent secretory vesicles fromthe trans-Golgi network of endocrine cells is inhibited by tyrosine kinase andphosphatase inhibitors. J. Cell Biol. 135:1471.
4619The Journal of Immunology
by guest on February 6, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
Related Documents











