Cell Reports Report Bisphosphonates Target B Cells to Enhance Humoral Immune Responses Elena Tonti, 1,2 Nereida Jime ´ nez de Oya, 1 Gabriele Galliverti, 1 E. Ashley Moseman, 2 Pietro Di Lucia, 1 Angelo Amabile, 1 Stefano Sammicheli, 1 Marco De Giovanni, 1 Laura Sironi, 1,4 Nicolas Chevrier, 3 Giovanni Sitia, 1 Luigi Gennari, 5 Luca G. Guidotti, 1,6 Ulrich H. von Andrian, 2, * and Matteo Iannacone 1,2,7, * 1 Division of Immunology, Transplantation and Infectious Diseases, San Raffaele Scientific Institute, 20132 Milan, Italy 2 Department of Microbiology and Immunobiology, Harvard Medical School, Boston, MA 02115, USA 3 Harvard University, FAS Center for Systems Biology, Cambridge, MA 02138, USA 4 Department of Physics, University of Milano Bicocca, 20126 Milan, Italy 5 Department of Medicine, Surgery and Neurosciences, University of Siena, 53100 Siena, Italy 6 Department of Immunology and Microbial Sciences, The Scripps Research Institute, La Jolla, CA 92037, USA 7 Vita-Salute San Raffaele University, 20132 Milan, Italy *Correspondence: [email protected] (U.H.v.A.), [email protected] (M.I.) http://dx.doi.org/10.1016/j.celrep.2013.09.004 This is an open-access article distributed under the terms of the Creative Commons Attribution-NonCommercial-No Derivative Works License, which permits non-commercial use, distribution, and reproduction in any medium, provided the original author and source are credited. SUMMARY Bisphosphonates are a class of drugs that are widely used to inhibit loss of bone mass in patients. We show here that the administration of clinically rele- vant doses of bisphosphonates in mice increases antibody responses to live and inactive viruses, pro- teins, haptens, and existing commercial vaccine for- mulations. Bisphosphonates exert this adjuvant-like activity in the absence of CD4 + and gd T cells, neutro- phils, or dendritic cells, and their effect does not rely on local macrophage depletion, Toll-like receptor signaling, or the inflammasome. Rather, bisphospho- nates target directly B cells and enhance B cell expansion and antibody production upon antigen encounter. These data establish bisphosphonates as an additional class of adjuvants that boost humoral immune responses. INTRODUCTION Bisphosphonates (BPs) are small-molecule inhibitors of bone resorption that are clinically approved for the treatment of skel- etal diseases such as osteoporosis and Paget’s disease of bone (Favus, 2010; Scott and Gershon, 1970); these compounds represent a large family of drugs that include first-generation clodronate (CLD) and etidronate (ETD) and nitrogen-containing alendronate (ALD), pamidronate (PMD), zoledronate (ZLD), and neridronate (NRD). Upon liposome encapsulation, BPs like CLD have been widely used to experimentally deplete tissue- resident phagocytes in rodents (Moseman et al., 2012; Van Rooijen and Sanders, 1994). A few studies in BP-treated mice unexpectedly noted increased antigen (Ag)-specific humoral immune responses (Gonzalez et al., 2010; Iannacone et al., 2010; Norton et al., 2011); herein, we set out to systematically dissect the mechanistic basis for this activity. RESULTS AND DISCUSSION As the majority of the above-mentioned studies utilized subcuta- neously administered BP-encapsulated liposomes prior to local viral challenge, we initially chose the same experimental setup to ask whether liposome encapsulation is required to increase anti- body (Ab) responses. To this end, footpads of C57BL/6 mice were injected with PBS, PBS liposomes (PBS-Lip), CLD lipo- somes (CLD-Lip), or CLD prior to infection in the same footpad with vesicular stomatitis virus (VSV), a prototypic cytopathic virus that induces an early T-independent immunoglobulin M (IgM) response followed by a T-dependent immunoglobulin G (IgG) response (Hangartner et al., 2006). When compared to PBS-injected mice, mice that received CLD exhibited up to 100-fold higher neutralizing antibody (nAb) titers toward VSV (Figure 1A), and this occurred when CLD was administered either prior to or concomitantly with the Ag (Figure S1A). Importantly, free CLD was as effective as CLD-Lip (Figure 1A), it exhibited a dose-dependent effect (Figure S1B), and its adjuvant activity was shared by other BPs that are currently in clinical use, including ETD, PMD, and ALD (Figure 1B). Subcutaneously administered CLD also increased Ab titers against inactive VSV, soluble proteins (ovalbumin [OVA]), haptens (NP-CGG), and the adjuvant-containing formulation Engerix-B (an approved vaccine against hepatitis B virus) (Figures 1C–1F); similar results were observed when CLD was administered intramuscularly along with the hemoaggluti- nin/neuroaminidase subunits of the human influenza virus A/NewCaledonia/20/99 (H 1 N 1 ; Figure 1G). CLD treatment increased both neutralizing IgM and IgG responses against VSV (Figures S1C–S1E) without altering the subtype of Ag-specific IgG induced upon immunization (Fig- ure S1F), and this correlated with the total number of CD138 + Cell Reports 5, 1–8, October 31, 2013 ª2013 The Authors 1 Please cite this article in press as: Tonti et al., Bisphosphonates Target B Cells to Enhance Humoral Immune Responses, Cell Reports (2013), http:// dx.doi.org/10.1016/j.celrep.2013.09.004

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Please cite this article in press as: Tonti et al., Bisphosphonates Target B Cells to Enhance Humoral Immune Responses, Cell Reports (2013), http://dx.doi.org/10.1016/j.celrep.2013.09.004
Cell Reports
Report
Bisphosphonates Target B Cellsto Enhance Humoral Immune ResponsesElena Tonti,1,2 Nereida Jimenez de Oya,1 Gabriele Galliverti,1 E. Ashley Moseman,2 Pietro Di Lucia,1 Angelo Amabile,1
Stefano Sammicheli,1 Marco De Giovanni,1 Laura Sironi,1,4 Nicolas Chevrier,3 Giovanni Sitia,1 Luigi Gennari,5
Luca G. Guidotti,1,6 Ulrich H. von Andrian,2,* and Matteo Iannacone1,2,7,*1Division of Immunology, Transplantation and Infectious Diseases, San Raffaele Scientific Institute, 20132 Milan, Italy2Department of Microbiology and Immunobiology, Harvard Medical School, Boston, MA 02115, USA3Harvard University, FAS Center for Systems Biology, Cambridge, MA 02138, USA4Department of Physics, University of Milano Bicocca, 20126 Milan, Italy5Department of Medicine, Surgery and Neurosciences, University of Siena, 53100 Siena, Italy6Department of Immunology and Microbial Sciences, The Scripps Research Institute, La Jolla, CA 92037, USA7Vita-Salute San Raffaele University, 20132 Milan, Italy
*Correspondence: [email protected] (U.H.v.A.), [email protected] (M.I.)
http://dx.doi.org/10.1016/j.celrep.2013.09.004
This is an open-access article distributed under the terms of the Creative Commons Attribution-NonCommercial-No Derivative WorksLicense, which permits non-commercial use, distribution, and reproduction in any medium, provided the original author and source are
credited.
SUMMARY
Bisphosphonates are a class of drugs that are widelyused to inhibit loss of bone mass in patients. Weshow here that the administration of clinically rele-vant doses of bisphosphonates in mice increasesantibody responses to live and inactive viruses, pro-teins, haptens, and existing commercial vaccine for-mulations. Bisphosphonates exert this adjuvant-likeactivity in the absence of CD4+ and gd T cells, neutro-phils, or dendritic cells, and their effect does not relyon local macrophage depletion, Toll-like receptorsignaling, or the inflammasome. Rather, bisphospho-nates target directly B cells and enhance B cellexpansion and antibody production upon antigenencounter. These data establish bisphosphonatesas an additional class of adjuvants that boosthumoral immune responses.
INTRODUCTION
Bisphosphonates (BPs) are small-molecule inhibitors of bone
resorption that are clinically approved for the treatment of skel-
etal diseases such as osteoporosis and Paget’s disease of
bone (Favus, 2010; Scott andGershon, 1970); these compounds
represent a large family of drugs that include first-generation
clodronate (CLD) and etidronate (ETD) and nitrogen-containing
alendronate (ALD), pamidronate (PMD), zoledronate (ZLD), and
neridronate (NRD). Upon liposome encapsulation, BPs like
CLD have been widely used to experimentally deplete tissue-
resident phagocytes in rodents (Moseman et al., 2012; Van
Rooijen and Sanders, 1994). A few studies in BP-treated mice
unexpectedly noted increased antigen (Ag)-specific humoral
immune responses (Gonzalez et al., 2010; Iannacone et al.,
2010; Norton et al., 2011); herein, we set out to systematically
dissect the mechanistic basis for this activity.
RESULTS AND DISCUSSION
As the majority of the above-mentioned studies utilized subcuta-
neously administered BP-encapsulated liposomes prior to local
viral challenge, we initially chose the same experimental setup to
ask whether liposome encapsulation is required to increase anti-
body (Ab) responses. To this end, footpads of C57BL/6 mice
were injected with PBS, PBS liposomes (PBS-Lip), CLD lipo-
somes (CLD-Lip), or CLD prior to infection in the same footpad
with vesicular stomatitis virus (VSV), a prototypic cytopathic
virus that induces an early T-independent immunoglobulin M
(IgM) response followed by a T-dependent immunoglobulin G
(IgG) response (Hangartner et al., 2006). When compared to
PBS-injected mice, mice that received CLD exhibited up to
100-fold higher neutralizing antibody (nAb) titers toward VSV
(Figure 1A), and this occurredwhenCLDwas administered either
prior to or concomitantly with the Ag (Figure S1A). Importantly,
free CLD was as effective as CLD-Lip (Figure 1A), it exhibited a
dose-dependent effect (Figure S1B), and its adjuvant activity
was shared by other BPs that are currently in clinical use,
including ETD, PMD, and ALD (Figure 1B).
Subcutaneously administered CLD also increased Ab titers
against inactive VSV, soluble proteins (ovalbumin [OVA]),
haptens (NP-CGG), and the adjuvant-containing formulation
Engerix-B (an approved vaccine against hepatitis B virus)
(Figures 1C–1F); similar results were observed when CLD
was administered intramuscularly along with the hemoaggluti-
nin/neuroaminidase subunits of the human influenza virus
A/NewCaledonia/20/99 (H1N1; Figure 1G).
CLD treatment increased both neutralizing IgM and IgG
responses against VSV (Figures S1C–S1E) without altering the
subtype of Ag-specific IgG induced upon immunization (Fig-
ure S1F), and this correlated with the total number of CD138+
Cell Reports 5, 1–8, October 31, 2013 ª2013 The Authors 1
25Days after VSV
CLD CLD-Lip
PBS-Lip PBS
103
104
105
106
0 5 10 15 20
Neu
traliz
ing
Ab
titer
102
101
* **
*** ***
*** *** ***
*** *** ***
A B **
** **
103
104
102
105
101
Neu
traliz
ing
Ab
titer
(d7)
106
PBS CLD ETD PMD ALD
**
PBS CLD
102
103
104
101
OVA
OVA
-spe
cific
IgG
tite
r (d1
4) ***
PBS CLD
102
103
104
105
NP
-spe
cific
IgG
tite
r (d1
4)
NP-CGG
101
**
PBS CLD
H1N
1-sp
ecifi
c Ig
G ti
ter (
d14)
102
103
101
H1N1
104
**
HB
sAb
(mIU
/ml)
(d21
)
PBS CLD
102
103
101
Engerix-B
104 **
PBS PFA-VSV PFA-VSV+CLD
0 2010Days after VSV
0
25
50
75
100
Sur
viva
l (%
)
Neu
traliz
ing
Ab
titer
(d12
5)
PBS CLD
103
104
102
105
101
106
**
Neu
traliz
ing
Ab
titer
(d14
)
PBS CLD
102
103
101
PFA-VSV 104
***
C
D E F
G H I
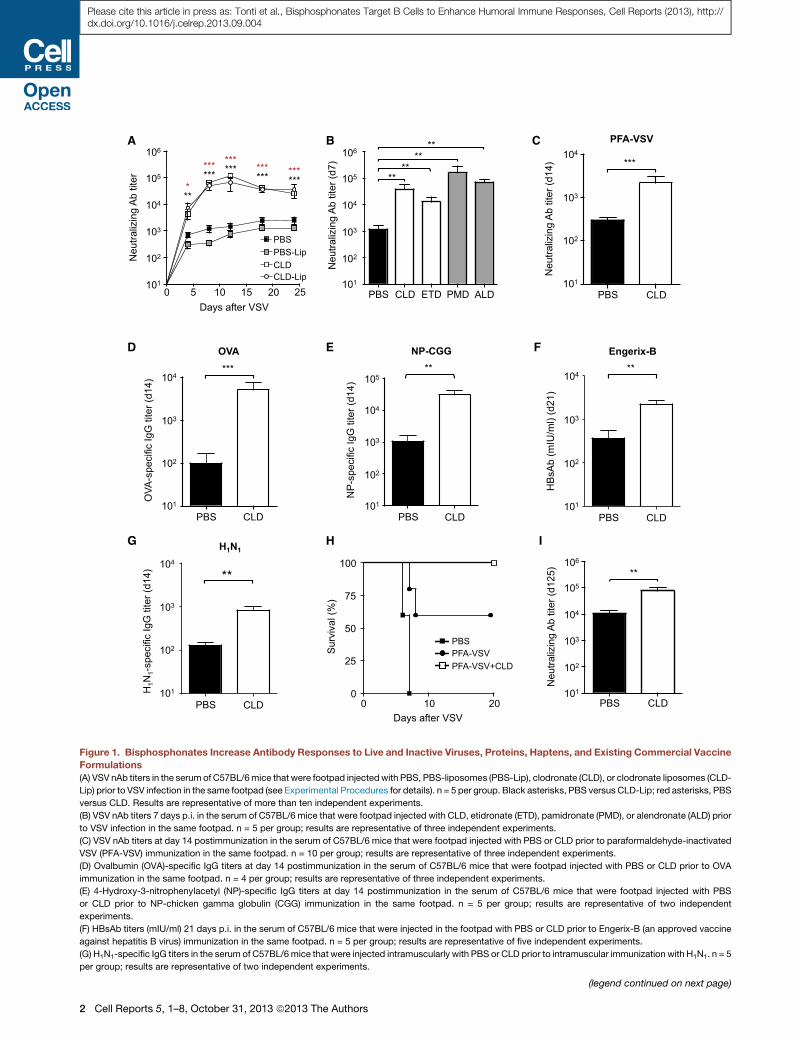
Figure 1. Bisphosphonates Increase Antibody Responses to Live and Inactive Viruses, Proteins, Haptens, and Existing Commercial Vaccine
Formulations
(A) VSV nAb titers in the serum of C57BL/6mice that were footpad injectedwith PBS, PBS-liposomes (PBS-Lip), clodronate (CLD), or clodronate liposomes (CLD-
Lip) prior to VSV infection in the same footpad (see Experimental Procedures for details). n = 5 per group. Black asterisks, PBS versus CLD-Lip; red asterisks, PBS
versus CLD. Results are representative of more than ten independent experiments.
(B) VSV nAb titers 7 days p.i. in the serum of C57BL/6 mice that were footpad injected with CLD, etidronate (ETD), pamidronate (PMD), or alendronate (ALD) prior
to VSV infection in the same footpad. n = 5 per group; results are representative of three independent experiments.
(C) VSV nAb titers at day 14 postimmunization in the serum of C57BL/6 mice that were footpad injected with PBS or CLD prior to paraformaldehyde-inactivated
VSV (PFA-VSV) immunization in the same footpad. n = 10 per group; results are representative of three independent experiments.
(D) Ovalbumin (OVA)-specific IgG titers at day 14 postimmunization in the serum of C57BL/6 mice that were footpad injected with PBS or CLD prior to OVA
immunization in the same footpad. n = 4 per group; results are representative of three independent experiments.
(E) 4-Hydroxy-3-nitrophenylacetyl (NP)-specific IgG titers at day 14 postimmunization in the serum of C57BL/6 mice that were footpad injected with PBS
or CLD prior to NP-chicken gamma globulin (CGG) immunization in the same footpad. n = 5 per group; results are representative of two independent
experiments.
(F) HBsAb titers (mIU/ml) 21 days p.i. in the serum of C57BL/6 mice that were injected in the footpad with PBS or CLD prior to Engerix-B (an approved vaccine
against hepatitis B virus) immunization in the same footpad. n = 5 per group; results are representative of five independent experiments.
(G) H1N1-specific IgG titers in the serum of C57BL/6mice that were injected intramuscularly with PBS or CLD prior to intramuscular immunization with H1N1. n = 5
per group; results are representative of two independent experiments.
(legend continued on next page)
Please cite this article in press as: Tonti et al., Bisphosphonates Target B Cells to Enhance Humoral Immune Responses, Cell Reports (2013), http://dx.doi.org/10.1016/j.celrep.2013.09.004
2 Cell Reports 5, 1–8, October 31, 2013 ª2013 The Authors
A
103
104
102
101
Neu
traliz
ing
IgM
tite
r
0 5 10 15 20Days after VSV
CD40L-/-+PBS CD40L-/-+CLD
**
***
***
***
D.L.
C
103
101
104
102
105
4 80Days after VSV
*** ***
*** ***
CLD PBS
α-Gr-1 α-Gr-1+CLD
103
104
102
105
0 5 10 15 20 25Days after VSV
101
***
Neu
traliz
ing
Ab
titer
** *** ***
*** *** ***
**
WT+PBS WT+CLD TCRδ-/-+PBSTCRδ-/-+ CLD
E
PBS CLD DT+PBSDT+CLD
0 4 8 12 16Days after VSV
103
104
102
105
101
Neu
traliz
ing
Ab
titer
N
eutra
lizin
g A
b tit
er
*** *** *** ***
*** *
103
101
104
102
Neu
traliz
ing
Ab
titer
(d14
)
105
** ** **
B
D
F
D.L.
103
104
102
101
Neu
traliz
ing
IgM
tite
r
0 5 10 15 20Days after VSV
* ***
***
***
MHC-II-/-+PBS MHC-II-/-+CLD
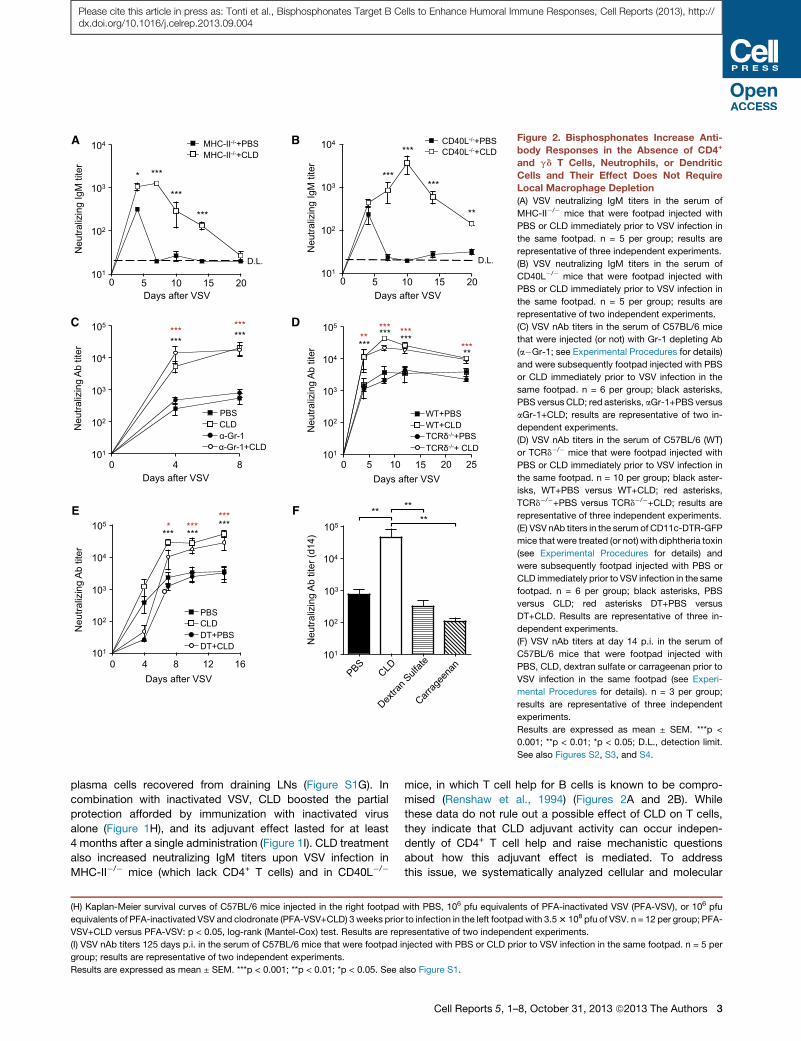
Figure 2. Bisphosphonates Increase Anti-
body Responses in the Absence of CD4+
and gd T Cells, Neutrophils, or Dendritic
Cells and Their Effect Does Not Require
Local Macrophage Depletion
(A) VSV neutralizing IgM titers in the serum of
MHC-II�/� mice that were footpad injected with
PBS or CLD immediately prior to VSV infection in
the same footpad. n = 5 per group; results are
representative of three independent experiments.
(B) VSV neutralizing IgM titers in the serum of
CD40L�/� mice that were footpad injected with
PBS or CLD immediately prior to VSV infection in
the same footpad. n = 5 per group; results are
representative of two independent experiments.
(C) VSV nAb titers in the serum of C57BL/6 mice
that were injected (or not) with Gr-1 depleting Ab
(a�Gr-1; see Experimental Procedures for details)
and were subsequently footpad injected with PBS
or CLD immediately prior to VSV infection in the
same footpad. n = 6 per group; black asterisks,
PBS versus CLD; red asterisks, aGr-1+PBS versus
aGr-1+CLD; results are representative of two in-
dependent experiments.
(D) VSV nAb titers in the serum of C57BL/6 (WT)
or TCRd�/� mice that were footpad injected with
PBS or CLD immediately prior to VSV infection in
the same footpad. n = 10 per group; black aster-
isks, WT+PBS versus WT+CLD; red asterisks,
TCRd�/�+PBS versus TCRd�/�+CLD; results are
representative of three independent experiments.
(E) VSV nAb titers in the serum of CD11c-DTR-GFP
mice that were treated (or not) with diphtheria toxin
(see Experimental Procedures for details) and
were subsequently footpad injected with PBS or
CLD immediately prior to VSV infection in the same
footpad. n = 6 per group; black asterisks, PBS
versus CLD; red asterisks DT+PBS versus
DT+CLD. Results are representative of three in-
dependent experiments.
(F) VSV nAb titers at day 14 p.i. in the serum of
C57BL/6 mice that were footpad injected with
PBS, CLD, dextran sulfate or carrageenan prior to
VSV infection in the same footpad (see Experi-
mental Procedures for details). n = 3 per group;
results are representative of three independent
experiments.
Results are expressed as mean ± SEM. ***p <
0.001; **p < 0.01; *p < 0.05; D.L., detection limit.
See also Figures S2, S3, and S4.
Please cite this article in press as: Tonti et al., Bisphosphonates Target B Cells to Enhance Humoral Immune Responses, Cell Reports (2013), http://dx.doi.org/10.1016/j.celrep.2013.09.004
plasma cells recovered from draining LNs (Figure S1G). In
combination with inactivated VSV, CLD boosted the partial
protection afforded by immunization with inactivated virus
alone (Figure 1H), and its adjuvant effect lasted for at least
4 months after a single administration (Figure 1I). CLD treatment
also increased neutralizing IgM titers upon VSV infection in
MHC-II�/� mice (which lack CD4+ T cells) and in CD40L�/�
(H) Kaplan-Meier survival curves of C57BL/6 mice injected in the right footpad
equivalents of PFA-inactivated VSV and clodronate (PFA-VSV+CLD) 3 weeks prio
VSV+CLD versus PFA-VSV: p < 0.05, log-rank (Mantel-Cox) test. Results are rep
(I) VSV nAb titers 125 days p.i. in the serum of C57BL/6 mice that were footpad i
group; results are representative of two independent experiments.
Results are expressed as mean ± SEM. ***p < 0.001; **p < 0.01; *p < 0.05. See a
mice, in which T cell help for B cells is known to be compro-
mised (Renshaw et al., 1994) (Figures 2A and 2B). While
these data do not rule out a possible effect of CLD on T cells,
they indicate that CLD adjuvant activity can occur indepen-
dently of CD4+ T cell help and raise mechanistic questions
about how this adjuvant effect is mediated. To address
this issue, we systematically analyzed cellular and molecular
with PBS, 106 pfu equivalents of PFA-inactivated VSV (PFA-VSV), or 106 pfu
r to infection in the left footpad with 3.53 108 pfu of VSV. n = 12 per group; PFA-
resentative of two independent experiments.
njected with PBS or CLD prior to VSV infection in the same footpad. n = 5 per
lso Figure S1.
Cell Reports 5, 1–8, October 31, 2013 ª2013 The Authors 3

A
103
104
102
101
Neu
traliz
ing
Ab
titer
0 4 8 12 16Days after VSV
PBS CLD
**
*
D.L.
PBS CLD
VI1
0Yen
B c
ells
(x10
4 )
2
0
4
6
8
10 *
VSV
B c
ells
(x10
6 )
0.5
0
1
1.5
2
2.5
PBS VSV+ CLD
**
***
***
103
104
102
105
0 5 10 15 20Days after VSV
101
WT+PBS WT+CLD MyD88-/-TRIF-/-+PBS MyD88-/-TRIF-/-+CLD
**
*** ***
*
*** ***
Neu
traliz
ing
Ab
titer
* **
103
101
104
102
Neu
traliz
ing
Ab
titer
105
0 5 10 15 20 25
WT+PBS WT+CLD Caspase-1-/-+PBSCaspase-1-/-+CLD
Days after VSV
*** ***
*** *
*** ***
*** *
B C D
E F G
I H
0 1 30 90 180 3600 1 30 90 180 360Days after ZLD
*
*** 1400
900
1000
1100
1200
1300
hum
an Ig
G (m
g/dl
) **
103
104
102
105
0 2 6 8 10Weeks after immunization
101
** ***
*
OVA
-spe
cific
IgG
tite
r
106
4 12
D.L.
* *
** *
*** **
**
Vehicle CLD CT CT+CLD
0 2 6 8 10Weeks after immunization
4101
103
102
Vehicle CLD CT CT+CLD
OVA
-spe
cific
IgA
titer
*
*** ***
*** ***
** ***
*** ***
D.L.
J
0 1 30 90 180 360Days after NRD
* *
1400
900
1000
1100
1200
1300
hum
an Ig
G (m
g/dl
)
Gene sets up-regulated in CLD
p-value 10-5 10-6
Extracellular region (GO:0005576)Immunoglobulin complex, circulating (GO:0042571)Humoral immune response (GO:0006959)Extracellular space (GO:0005615)Extracellular region part (GO:0044421)
0h 8h TERM
Figure 3. Bisphosphonates Directly Target B Cells and Enhance B Cell Expansion and Antibody Production upon Antigen EncounterIndependently of Toll-like Receptor Signaling or the Inflammasome
(A) Gene enrichment analysis of clodronate-dependent genes from microarray measurements of whole lymph node. Shown are the Gene Ontology (GO) terms
(rows) statistically enriched among the 156 genes with at least a 2-fold increase in expression at 0 and 8 hr (columns) after VSV infection in clodronate (CLD)-
treatedmice compared to control (PBS). Enriched GO term categories include BP (biological process) and CC (cellular component). Green: p value < 10�6; white:
p value < 10�4; gray: no enrichment. See Experimental Procedures for details.
(B) VSV nAb titers in the serum of DHLMP2A mice that were intravenously transferred with PBS- or CLD-treated VI10Yen B cells 24 hr prior to footpad VSV
infection. n = 6 per group; results are representative of two independent experiments.
(C) Quantification of the total number of VI10Yen B cells recovered from the spleen of the mice described in (B) 8 days after VSV.
(D) Quantification of the total number of cells recovered 48 hr upon in vitro culture of purified VI10Yen B cells with PBS, VSV, or 10 nM clodronate and VSV.
(E) VSV nAb titers in the serum of C57BL/6 (WT) or MyD88�/�/Trif�/� mice that were footpad injected with PBS or CLD immediately prior to VSV infection in the
same footpad. n = 4 per group; black asterisks, WT + PBS versus WT + CLD; red asterisks MyD88�/�/Trif�/�+PBS versus MyD88�/�/Trif�/�+CLD; results are
representative of three independent experiments.
(legend continued on next page)
Please cite this article in press as: Tonti et al., Bisphosphonates Target B Cells to Enhance Humoral Immune Responses, Cell Reports (2013), http://dx.doi.org/10.1016/j.celrep.2013.09.004
4 Cell Reports 5, 1–8, October 31, 2013 ª2013 The Authors

Please cite this article in press as: Tonti et al., Bisphosphonates Target B Cells to Enhance Humoral Immune Responses, Cell Reports (2013), http://dx.doi.org/10.1016/j.celrep.2013.09.004
changes induced by BPs at the site of injection and at the level
of the draining LN.
First, we examined the BP injection site. In agreement with
previously published data (Iannacone et al., 2010; Norton et al.,
2011), footpad injection of CLD alone induced a local inflamma-
tory infiltrate composed mostly of Gr-1+ neutrophils and inflam-
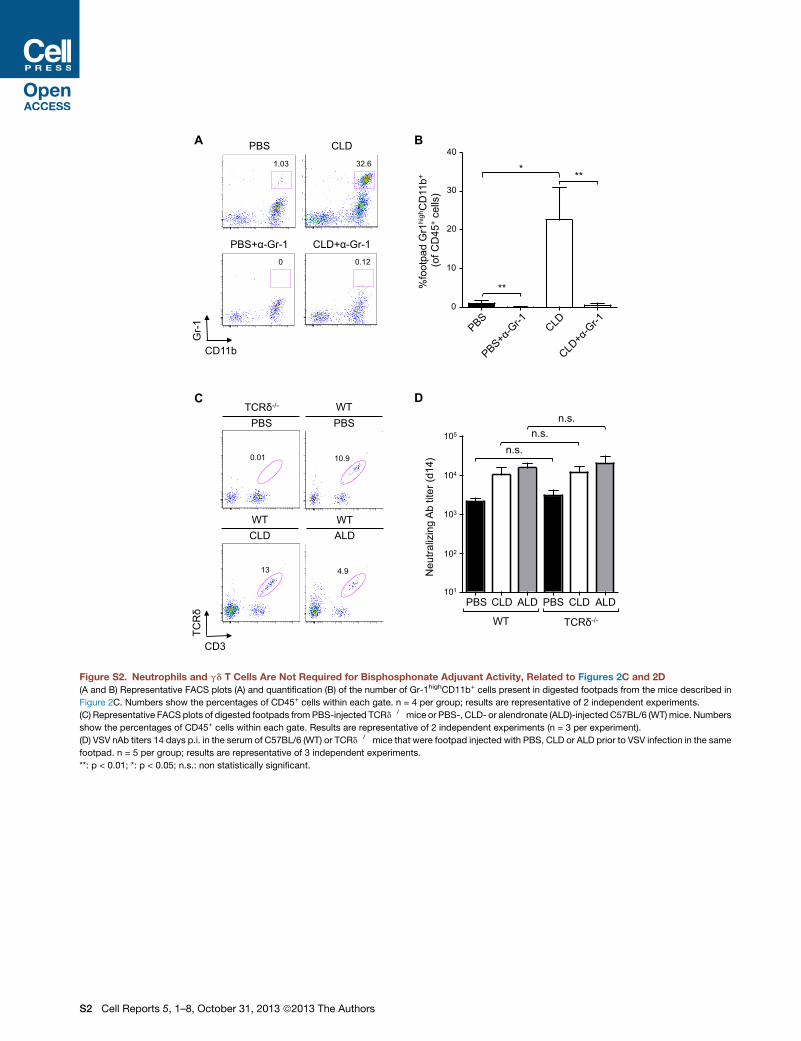
matory monocytes (Figure S2A). Elimination of this infiltrate by
systemic anti-Gr-1 treatment (Figures S2A and S2B), however,
did not prevent CLD from increasing nAb titers upon VSV infec-
tion (Figure 2C), indicating that CLD adjuvant activity does not
require Gr-1+ leukocytes.
Next, we considered the role of two additional footpad-resi-
dent cell types: gd T cells and conventional dendritic cells
(DCs). gd T cells can provide help to B cells (Pao et al., 1996;
Wen et al., 1996) and are reportedly activated by CLD (Schilbach
et al., 2001) or ALD (Thompson et al., 2010). Footpad CLD or ALD
injection did not increase the local number of gd T cells (Fig-
ure S2C), and VSV infection of TCRd�/� mice (Itohara et al.,
1993) revealed that CLD and ALD adjuvant activity is maintained
in the absence of these cells (Figures 2D and S2D). We then
reasoned that BPs might act on DCs, because DCs can present
Ag to B cells (Qi et al., 2006) and most known adjuvants are
thought to function by activating DCs (Coffman et al., 2010).
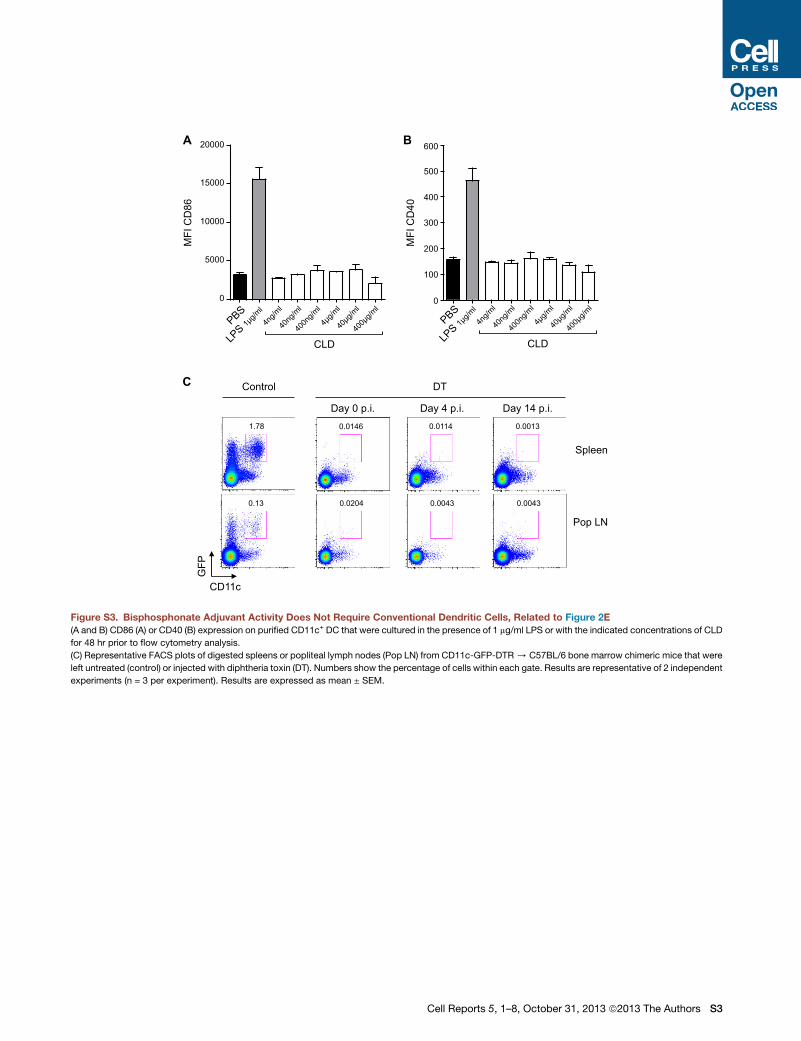
Incubating DCs with CLD in vitro did not alter expression of acti-
vation markers, such as CD86 and CD40 (Figures S3A and S3B),
and the systemic elimination of these cells in vivo (Figure S3C)
did not significantly inhibit CLD adjuvant activity. Indeed, CLD
administration in DC-depleted mice challenged with VSV re-
sulted in nAb titers that, by day 7 postinfection (p.i.), were com-
parable to those observed in DC-competent animals subjected
to the same procedures (Figure 2E). It is worth noting that, inde-
pendently of CLD treatment, DC depletion caused a transient
reduction in nAb titers at day 4 p.i., consistent with a role of
DC in early B cell activation (Qi et al., 2006; Scandella et al.,
2007). Overall, these data indicate that CLD adjuvant activity
does not require conventional DCs.
We next analyzed the draining LN. The only quantitative
change in LN cellular composition detectable upon CLD treat-
ment was the depletion of CD169+ macrophages lining the sub-
capsular and medullary sinuses (Figures S4A and S4B). We
found it surprising that BPs increase Ab responses despite
depleting subcapsular sinus macrophages, because these cells
were recently identified as critical Ag-presenting cells for B cell
responses (Carrasco and Batista, 2007; Junt et al., 2007; Phan
et al., 2007). To clarify these seemingly contradictory observa-
tions, we sought alternative strategies to deplete LN macro-
phages independently of BP administration. These strategies
(F) VSV nAb titers in the serum of C57BL/6 (WT) or caspase-1�/�mice that were in
same footpad. n = 10 per group; black asterisks, WT+PBS versus WT+CLD; red
sentative of three independent experiments.
(G) Ovalbumin (OVA)-specific IgG titers in the serum of C57BL/6mice that were im
(CT), or OVA + cholera toxin + CLD (CT + CLD). n = 5 per group; results are repr
(H) OVA-specific IgA titers in the feces of the mice described in (G).
(I) Total IgG in the sera of patients immediately prior to or at the indicated time poin
Table S2 for patient characteristics.
(J) Total IgG in the sera of patients immediately prior to or at the indicated time po
for patient characteristics.
Results are expressed as mean ± SEM. ***p < 0.001; **p < 0.01; *p < 0.05. D.L.,
(footpad dextran sulfate or carrageenan injections in wild-type
[WT] mice or footpad diphtheria toxin injection in CD11c-DTR
mice) effectively depleted LN macrophages (Figure S4C; Ianna-
cone et al., 2010), but they failed to increase nAb titers upon VSV
infection (Figures 2F and S4D). This indicates that LN macro-
phage depletion per se does not increase humoral immune
responses and suggests that CLD adjuvant activity does not
rely on LN macrophages.
To investigate the molecular basis for CLD adjuvant effect, we
next performed genome-wide mRNA profile analysis of draining
LNs from mice treated with either PBS or CLD and sacrificed
before (0 hr) or 8 hr after VSV infection (Figure S5; Table S1).
This analysis showed that a relatively small number of genes
were regulated by CLD treatment (Figures 3A, S6, S7, S8, S9,
and S10; Table S1). Most of the downregulated genes upon
CLD treatment were macrophage specific (Figures S6, S7, and
S8; Table S1), in line with the observation that CLD treatment
depleted LN macrophages (Figure S4). CLD treatment also
downregulated type I interferon (IFN-I) genes induced upon
VSV infection (Figures S7 and S8; Table S1). This is consistent
with previously published data showing that LN subcapsular
sinus macrophages are a major source of IFN-I during this infec-
tion and their depletion inhibits IFN-I gene expression (Ianna-
cone et al., 2010). Reduced IFN-I gene expression could have
also resulted from plasmacytoid DC (pDC) dysfunction, as these
cells represent an additional source of IFN-I in this system and
they are not depleted upon BP treatment (Iannacone et al.,
2010). The CLD-dependent reduced expression of the pDC-spe-
cific marker Siglec-H (Blasius et al., 2006) is also suggestive of
pDC dysfunction (Figures S6 and S7; Table S1). It is noteworthy,
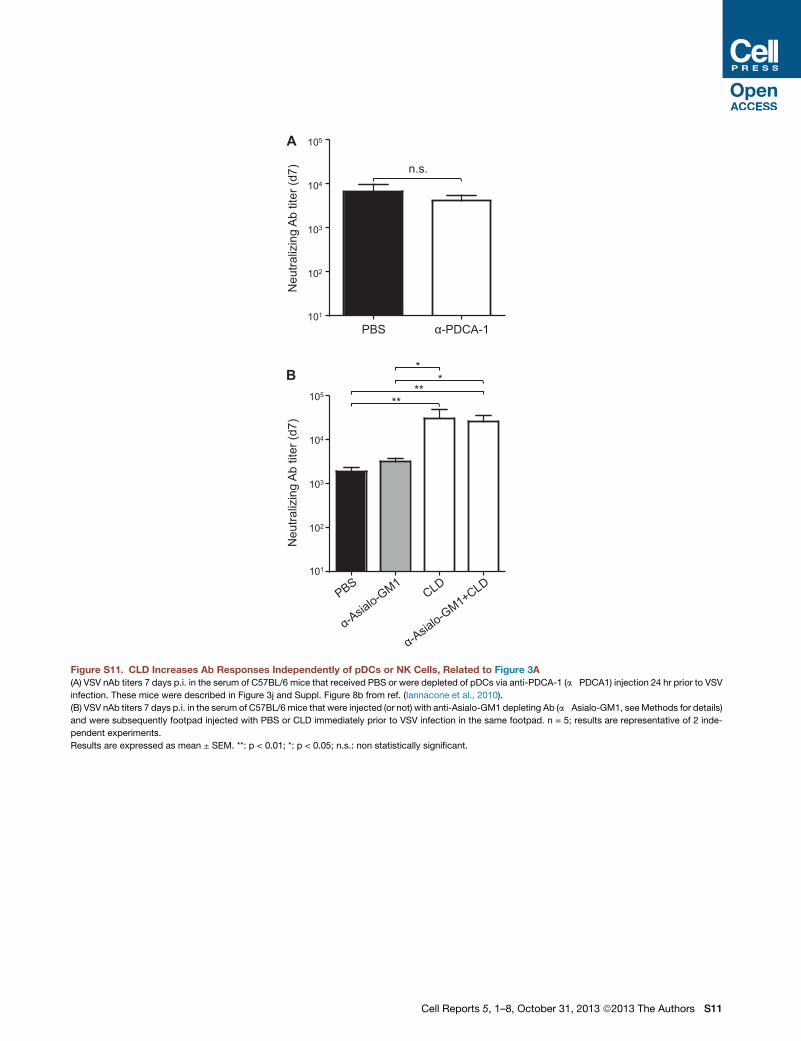
however, that depletion of pDCs (which reduced IFN-I by�50%;
Iannacone et al., 2010) failed to increase Ab responses (Fig-
ure S11A), suggesting that CLD adjuvant activity does not rely
on pDCs.We also noted that some natural killer (NK) cell-specific
transcripts were reduced upon CLD treatment (Figures S6 and
S7; Table S1). Depleting these cells did not alter the capacity
of CLD to increase nAb responses upon VSV infection (Fig-
ure S11B), arguing against a role for NK cells in the adjuvant
activity of BPs.
Next, we examined genes that were upregulated by CLD
treatment. CLD induced several neutrophil-specific transcripts
(Figures 3A, S9, and S10; Table S1), in keeping with the ability
of BPs to recruit and activate these cells (Norton et al., 2011;
2012). Data previously described in Figure 2C, however,
ruled out a role for neutrophils in BP adjuvant activity. Of note,
CLD upregulated a number of B cell-specific transcripts and
gene sets associated with B cell function (Figures 3A, S9,
jected in the footpad with PBS or CLD immediately prior to VSV infection in the
asterisks, caspase-1�/�+PBS versus caspase-1�/�+CLD; results are repre-
munized orally with OVA alone (vehicle), OVA + CLD (CLD), OVA + cholera toxin
esentative of two independent experiments.
ts after a single intravenous infusion of zoledronate (ZLD) is shown. n = 11; see
ints after a single intravenous infusion of neridronate (NRD). n = 9; see Table S2
detection limit. See also Figures S5, S6, S7, S8, S9, S10, and S11.
Cell Reports 5, 1–8, October 31, 2013 ª2013 The Authors 5

Please cite this article in press as: Tonti et al., Bisphosphonates Target B Cells to Enhance Humoral Immune Responses, Cell Reports (2013), http://dx.doi.org/10.1016/j.celrep.2013.09.004
and S10; Table S1). When compared to controls, these B cell-
related transcripts were higher in CLD-treated animals before
(Figure S9) and after (Figure S10) VSV infection, suggesting
that CLD might directly impact B cells. To directly test this pos-
sibility, we isolated B cells from VI10Yen mice (which express a
VSV-specific B cell receptor), and treated them with PBS or
CLD in vitro prior to adoptive transfer into DHLMP2A mice
(which are devoid of surface-expressed and secreted Abs). In
this experimental setup - where Abs can be produced only by
the transferred B cells—VSV infection led to nAb titers that
were higher in mice that received CLD-treated, rather than
PBS-treated, VI10Yen B cells (Figure 3B); this correlated with
the capacity of CLD to promote Ag-specific B cell expansion
both in vivo and in vitro (Figures 3C and 3D). Taken together,
these results indicate that CLD directly targets B cells to
enhance their expansion and their Ab production upon Ag
encounter. Although the molecular pathways by which CLD
and other BPs impact B cell function remain to be determined,
it is worth noting that CLD adjuvanticity did not involve the two
known signaling pathways through which most other adjuvants
are thought to act (Coffman et al., 2010) (the Toll-like receptors
and the inflammasome pathway, which rely on Myd88/TRIF and
caspase-1 for signaling, respectively). Indeed, VSV nAb titers in
CLD-treated Myd88�/�/TRIF�/� and caspase-1�/� mice were
indistinguishable from those of similarly treated WT mice (Fig-
ures 3E and 3F).
Since BPs are often taken orally by patients, we next sought
to evaluate whether these small molecules act as adjuvants
upon oral immunization in mice. As expected (Pierre et al.,
1992), the oral administration of OVA alone failed to induce a
detectable OVA-specific Ab response (Figures 3G and 3H).
By contrast, the concomitant oral administration of OVA
and CLD induced OVA-specific serum IgG and fecal immuno-
globulin A (IgA) responses (Figures 3G and 3H). Furthermore,
CLD was able to increase the OVA-specific Ab response
afforded when OVA was administered together with the
mucosal adjuvant cholera toxin (Pierre et al., 1992) (Figures
3G and 3H). These data indicate that CLD is an effective oral
adjuvant capable of increasing both systemic and mucosal
Ab responses.
Finally, we tested whether the nitrogen-containing BPs ZLD
and NRD increase Ab responses in a cohort of patients affected
by osteoporosis or Paget disease of bone (Table S2). Notably, a
transient but significant increase in total serum IgG levels was
detected for up to 3 months upon a single intravenous infusion
of BPs (Figures 3I and 3J). Although the impact of BPs on Ag-
specific Ab responses remains to be determined, this observa-
tion suggests that these drugs enhance B cell responses in
humans.
In conclusion, these data establish BPs as adjuvants that
target B cells to increase humoral immune responses. Transla-
tional evaluation of these compounds in humans should be rela-
tively straightforward, because BPs, unlike all other adjuvants,
are already widely used in the clinic as stand-alone drugs with
excellent safety profiles. Thus, BPs could be readily combined
with both existing and newly developed vaccines, especially in
settings where immune responses to a vaccine alone are weak
or where Ags are in short supply.
6 Cell Reports 5, 1–8, October 31, 2013 ª2013 The Authors
EXPERIMENTAL PROCEDURES
Mice
C57BL/6 and Balb/c mice were purchased from Charles River or The Jackson
Laboratory. CD11c- GFP-DTR [B6.FVB-Tg(Itgax-DTR/EGFP)57Lan/J] mice
were purchased from The Jackson Laboratory. MHC-II�/� (B6.129-H2-
Ab1tm1Doi/DoiOrl) mice were provided by P. Dellabona (San Raffaele
Scientific Institute). TCRd�/� (B6.129P2-Tcrdtm1Mom/J) mice were provided
by W. Havran (The Scripps Research Institute). MyD88�/�/TrifLps2/Lps2 mice
were provided by B. Beutler (The Scripps Research Institute). Caspase-1�/�
(B6.129S2-Casp1tm1sesh) and CD40L�/� (B6.129S2-Cd40tm1Imx/J) mice were
obtained through the Swiss Immunological Mutant Mouse Repository (Zurich,
Switzerland). DHLMP2A mice (Casola et al., 2004) were originally provided by
K. Rajewsky (Harvard Medical School) and bred five generations against
C57BL/6 mice. VI10Yenmice (Hangartner et al., 2003) were originally provided
by R. M. Zinkernagel and H. Hengartner. Bone marrow chimeras were gener-
ated by irradiation of C57BL/6 mice with 1,300 rad in split doses and reconsti-
tution with CD11c-DTR-GFP bone marrow; mice were allowed to reconstitute
for at least 8 weeks prior to use. Mice were housed under specific pathogen-
free conditions and used at 6–8 weeks of age. All experimental animal proce-
dures were approved by the Institutional Animal Committees of San Raffaele
Scientific Institute, Harvard Medical School, and The Scripps Research
Institute.
Bisphosphonate Treatment
CLD, ALD, PMD, and ETD were obtained from Sigma, dissolved in PBS, and
injected in the footpad in a volume of 20 ml. The injected amount for each
bisphosphonate was 2 mg, unless otherwise indicated. In the experiments
described in Figure 1A, 30 ml of clodronate liposomes (CLD-Lip) or PBS lipo-
somes (PBS-Lip, both provided by N. Van Rooijen) were injected in the footpad
7 days before antigen administration.
Infections and Immunizations
Mice were infected with 104 plaque-forming units (pfu) of VSV serotype Indiana
(VSV-IND), 104 pfu of VSV serotype New Jersey (VSV-NJ), or 106 pfu of
VSVeGFP (Iannacone et al., 2010). Alternatively, mice were immunized with
106 pfu equivalent of PFA-inactivated VSV (Bachmann et al., 1993). Viruses
were dissolved in 20 ml of PBS and injected into the footpad. In survival exper-
iments, mice were challenged with 3.5 3 108 pfu of VSV in the left footpad
3 weeks after PFA-VSV immunization in the right footpad. Other antigens
used for footpad immunization experiments included 4-hydroxy-3-nitro-
phenyl-chicken gamma globulin (NP-CGG, 50 mg/dose; Biosearch Technolo-
gies), ovalbumin (OVA, 100 mg/dose; Sigma), and Engerix-B (20 ml/dose given
twice 2 weeks apart; GlaxoSmithKline). Hemoagglutinin/neuroaminidase sub-
units from the human influenza virus A/NewCaledonia/20/99 (H1N1, 3 mg/dose;
provided by P. Dellabona) was injected in the anterior tibialis muscle.
For experiments involving oral immunization, mice were given vehicle or
10 mg of CLD 24 hr prior to receiving 1 mg of OVA with or without 10 mg of
cholera toxin. All treatments were administered via gavage in 0.2 ml of antacid
buffer (3% sodium bicarbonate) on days 0, 7, and 21. For the measurement of
mucosal IgA, fresh fecal pellets were collected and immediately frozen
at �20�C. Before analysis, fecal pellets were weighed and dissolved in prote-
ase inhibitor solution at a concentration of 200 mg/ml, as described previously
(Lauterslager et al., 2001).
All infectious work was performed in designated BL-2 workspaces in accor-
dance with institutional guidelines. Mice were retro-orbitally bled at the indi-
cated time points for Ag-specific Abs and measured by endpoint ELISA (Galli
et al., 2007; Tonti et al., 2012), VSV neutralization assay (Iannacone et al.,
2010), or with an HBsAb detection kit (Diagnostic Bioprobes) in accordance
with the manufacturer’s instructions. For determination of VSV-neutralizing
IgG titers, serawere incubatedwith equal volumes of 0.1M2-mercaptoethanol
in PBS for 1 hr at room temperature before dilution (Scott and Gershon, 1970).
Confocal Microscopy
Confocal microscopy analysis of popliteal LNs was performed as described
previously (Moseman et al., 2012). Sections were stained with eFluor450-
conjugated anti-B220 (RA3-6B2, eBioscience), FITC-conjugated anti-CD169

Please cite this article in press as: Tonti et al., Bisphosphonates Target B Cells to Enhance Humoral Immune Responses, Cell Reports (2013), http://dx.doi.org/10.1016/j.celrep.2013.09.004
(3D6.112, AbD Serotec), and Alexa Fluor 647-conjugated anti-TCRb (H57-597,
BioLegend).
In Vivo Depletion of Neutrophils, DCs, LN Macrophages, pDCs, and
NK Cells
Neutrophils were depleted by intraperitoneal injection of 100 mg of Gr-1 Abs
(RB6-8C5, BioXCell) every 2 days, beginning 3 days prior to immunization.
DCs were depleted from CD11c-GFP-DTR / C57BL/6 bone marrow chi-
meric mice by injecting diphtheria toxin (DT, Sigma) in the footpad (50 ng) and
in the peritoneum (500 ng) every 2 days, starting the day prior to immunization.
Popliteal LN macrophages were depleted by injecting 2 mg of carrageenan
(Sigma) in the footpad every 2 days, starting 5 days prior to immunization.
Alternatively, LN macrophages were depleted by footpad injection of 1 mg
of dextran sulfate (Sigma) 5 days prior to immunization. Serum from CD11c-
DTR-GFP mice (described in Figure S3 in Iannacone et al., 2010), where LN
macrophages are depleted by a single footpad injection of DT (4 ng) 6 days
before VSV infection, were assessed for VSV neutralizing Ab titers 4 days after
VSV infection.
Serum from the mice described in Figures 3J and S8b of Iannacone et al.
(2010) (where pDCs were depleted by intravenous injection of 500 mg of
PDCA-1 depleting Ab [JF05-1C2.4.1, Miltenyi Biotec] 24 hr prior to VSV infec-
tion) were assessed for VSV neutralizing Ab titers 7 days after VSV infection.
NK cells were depleted by intravenous injection of Asialo-GM1 Abs
(Cedarlane) every 2 days beginning 3 days prior to VSV infection, as described
previously (Tonti et al., 2012).
All depletions were confirmed by flow cytometry and/or confocal
microscopy.
Tissue Digestion and Flow Cytometry
Single-cell suspensions of LNs, spleens, and footpads were generated as
described in Iannacone et al. (2010). All flow cytometry analyses were per-
formed in fluorescence-activated cell sorting buffer containing PBS with
2 mM EDTA and 2% FBS on a FACS CANTO (BD Pharmingen) and analyzed
with FlowJo software (Tree Star).
Abs used included FITC-conjugated anti-CD169 (3D6.112, AbD Serotec),
Alexa Fluor 488-conjugated and PE-Cy7-conjugated anti-B220 (RA3-6B2,
BioLegend), PE-conjugated anti-TCRd (GL3, BioLegend), and anti-CD86
(GL-1, BioLegend), PE-Cy7-conjugated anti-CD45 (30-F11, BioLegend), PB-
conjugated anti-Ly6C (HK1.4, BioLegend), APC-Cy7-conjugated anti-F4/80
(BM8, BioLegend), Alexa Fluor 647-conjugated anti-CD8 (53-6.7, BioLegend),
biotinylated anti-CD11b (M1/70, BioLegend), PE-conjugated anti-CD138
(281-2, BioLegend), PE-Cy7-conjugated anti-CD3 (145-2C11, BioLegend),
PE-Cy5-conjugated anti-NK1.1 (PK136, BioLegend), PerCP-Cy5.5-conju-
gated anti-Gr1 (RB6-8C5), APC-conjugated anti-CD40 (HM40-3, BD
Pharmingen), Alexa Fluor 488-conjugated anti-igG (A-11023, Invitrogen), PE-
Cy7-conjugated anti-CD11c (N418, eBioscience), eFluor450-conjugated
anti-CD4 (RM4-5, eBioscience). The idiotypic Ab 35.61 for detection of the
VI10 BCR in VI10Yenmice (Hangartner et al., 2003) was produced from hybrid-
oma supernatants in accordance with standard methods.
Isolation and Activation of DCs and B Cells
Splenic DCs were isolated from C57BL/6 mice that were injected subcutane-
ously 11–14 days earlier with 43 106 Flt3-ligand-secreting B16 tumor cells, as
described previously (Cavanagh et al., 2005). CD11c+ DCs were purified by
positive selection with anti-CD11c microbeads (>95% CD11c+, Miltenyi) and
cultured in the presence of 1 mg/ml LPS (E. coli 0.26:B6, Sigma) or with the indi-
cated concentrations of CLD for 48 hr prior to flow cytometry analysis.
Naive B cells from spleens of VI10Yen mice were negatively selected by
magnetic isolation with CD43 beads (Miltenyi) as described previously (Junt
et al., 2007). The purity was 98% as determined by CD19 surface staining.
B cells (106 cells/ml) were cultured in RPMI 1640 media (Lonza) supplemented
with 10% fetal bovine serum, 50 mM 2-ME (Sigma), 10 mM HEPES (Lonza),
2 mM L-glutamine (Lonza), and 100 U/ml penicillin/streptomycin with or
without 10 nM CLD. After 4 hr, VI10Yen B cells were harvested, washed three
times, and either intravenously injected into DHLMP2A recipients (107 cells/
mouse) 24 hr prior to footpad VSV infection (Figures 3B and 3C) or cultured
for 48 hr in the presence of VSV at a multiplicity of infection of 1 (Figure 3D).
Gene Expression Profiling
Whole lymph nodes were lysed in 600 ml of QIAzol (QIAGEN) reagent using the
TissueLyser II. Total RNA was extracted following the miRNeasy kit’s proce-
dure (QIAGEN), and sample quality was tested on a 2100 Bioanalyzer (Agilent).
RNAwas reverse transcribedwith the High Capacity cDNAReverse Transcrip-
tion kit (Applied Biosystems). Real time quantitative PCR reactions were per-
formed on the LightCycler 480 system (Roche) with FastStart Universal
SYBR Green Master Mix (Roche). Every reaction was run in triplicate and
GAPDH levels were used as an endogenous control for normalization. For
oligonucleotide microarray hybridization, 1 mg of RNA were labeled, frag-
mented, and hybridized to an Affymetrix Mouse Genome 430A 2.0 Array. After
scanning, the expression value for each gene was calculated with RMA
(Robust Multi-Array) normalization using R. The average intensity difference
values were normalized across the sample set. Probe sets that were absent
in all samples according to Affymetrix flags were removed. All values below
40 were floored to 40. Only probe sets that changed in two biological dupli-
cates by 2-fold or more were analyzed further in this study: (1) regulated genes
were hierarchically clustered using the software Gene-E (http://www.
broadinstitute.org/cancer/software/GENE-E/), and (2) functional enrichment
of GO terms (BP, biological processes; CC, cellular components) and KEGG
pathway enrichment analysis were performed using DAVID (http://david.
abcc.ncifcrf.gov) (Huang et al., 2009).
Patients
Total serum IgG levels were assessed in 20 patients affected by osteoporosis
or Paget disease of bone (age range 52–78 years; Table S2) that received a sin-
gle intravenous injection of 5 mg zoledronate (Aclasta, Novartis Pharmaceuti-
cals) or 200 mg neridronate (Nerixia, Abiogen Pharma). Serum samples were
obtained before and 1, 30, 90, 180, and 360 days after BP infusion. IgG levels
were quantified from stored samples by an immunodiffusion technique using
NOR Partigen immunoplates purchased from Siemens (Siemens Healthcare
Diagnostics). All experiments involving human subjects were approved by
the institutional review board of the University of Siena and performed after
obtaining informed consent from all subjects.
Statistical Analyses
Results are expressed as mean ± SEM. All statistical analyses were performed
in Prism (GraphPad Software). Means between two groups were compared
with a two-tailed t test. Means among three or more groups were compared
with one-way or two-way analysis of variance with Bonferroni’s post hoc
test. Kaplan-Meier survival curves were compared with the log-rank (Mantel-
Cox) test.
ACCESSION NUMBERS
The microarray data discussed in this publication have been deposited in
NCBI’s Gene Expression Omnibus and are accessible through GEO Series
accession number GSE50403.
SUPPLEMENTAL INFORMATION
Supplemental Information includes eleven figures and two tables and can be
foundwith this article online at http://dx.doi.org/10.1016/j.celrep.2013.09.004.
AUTHOR CONTRIBUTIONS
E.T., L.G.G., U.H.v.A., andM.I. designed the study; E.T., N.J.d.O, G.G., E.A.M.,
P.D.L., A.A., S.S., M.D.G., L.S., and M.I. performed experiments; E.T. and M.I.
analyzed the data; N.C. performed and analyzed themicroarray data; L.G. per-
formed measurements in human subjects; G.S. gave conceptual advice; and
E.T., L.G.G., U.H.v.A. and M.I. wrote the manuscript.
ACKNOWLEDGMENTS
We thank T. Cataudella, A. Fiocchi, B. Fiore, D. Covarello, and M. Mainetti for
technical support; R. Serra for secretarial assistance; P. Dellabona (San
Cell Reports 5, 1–8, October 31, 2013 ª2013 The Authors 7

Please cite this article in press as: Tonti et al., Bisphosphonates Target B Cells to Enhance Humoral Immune Responses, Cell Reports (2013), http://dx.doi.org/10.1016/j.celrep.2013.09.004
Raffaele Scientific Institute) for providing MHC-II�/� mice; W. Havran (The
Scripps Research Institute) for providing TCRd�/� mice; B. Beutler (The
Scripps Research Institute) for providing MyD88�/�/TrifLps2/Lps2 mice; S. Whe-
lan (Harvard Medical School) for providing VSV and VSVeGFP; N. van Rooijen
for providing clodronate liposomes; F. Benvenuti (International Centre for Ge-
netic Engineering and Biotechnology) for providing Flt3-ligand-secreting B16
tumor cells; S. Cenci, C. Scielzo, P. Ghia, and the members of the Iannacone,
Guidotti and von Andrian laboratories for helpful discussions; and F.V. Chisari,
A. Mondino, P. Dellabona, R. Pardi, and Z.M. Ruggeri for critical reading of the
manuscript. This work was supported by ERC grants 281648 (to M.I.) and
250219 (to L.G.G.); NIH grants AI40696 (to L.G.G.) and AI078897 and
AI069259 (to U.H.v.A.); Italian Association for Cancer Research (AIRC) grant
9965 (to M.I.); and a career development award from the Giovanni Armen-
ise-Harvard Foundation (to M.I.).
Received: July 18, 2013
Revised: September 3, 2013
Accepted: September 4, 2013
Published: October 10, 2013
REFERENCES
Bachmann, M.F., Kundig, T.M., Kalberer, C.P., Hengartner, H., and Zinkerna-
gel, R.M. (1993). Formalin inactivation of vesicular stomatitis virus impairs
T-cell- but not T-help-independent B-cell responses. J. Virol. 67, 3917–3922.
Blasius, A.L., Cella, M., Maldonado, J., Takai, T., and Colonna, M. (2006).
Siglec-H is an IPC-specific receptor that modulates type I IFN secretion
through DAP12. Blood 107, 2474–2476.
Carrasco, Y.R., and Batista, F.D. (2007). B cells acquire particulate antigen in a
macrophage-rich area at the boundary between the follicle and the subcapsu-
lar sinus of the lymph node. Immunity 27, 160–171.
Casola, S., Otipoby, K.L., Alimzhanov, M., Humme, S., Uyttersprot, N., Kutok,
J.L., Carroll, M.C., and Rajewsky, K. (2004). B cell receptor signal strength de-
termines B cell fate. Nat. Immunol. 5, 317–327.
Cavanagh, L.L., Bonasio, R., Mazo, I.B., Halin, C., Cheng, G., van der Velden,
A.W.M., Cariappa, A., Chase, C., Russell, P., Starnbach, M.N., et al. (2005).
Activation of bone marrow-resident memory T cells by circulating, antigen-
bearing dendritic cells. Nat. Immunol. 6, 1029–1037.
Coffman, R.L., Sher, A., and Seder, R.A. (2010). Vaccine adjuvants: putting
innate immunity to work. Immunity 33, 492–503.
Favus, M.J. (2010). Bisphosphonates for osteoporosis. N. Engl. J. Med. 363,
2027–2035.
Galli, G., Pittoni, P., Tonti, E., Malzone, C., Uematsu, Y., Tortoli, M., Maione, D.,
Volpini, G., Finco, O., Nuti, S., et al. (2007). Invariant NKT cells sustain specific
B cell responses and memory. Proc. Natl. Acad. Sci. USA 104, 3984–3989.
Gonzalez, S.F., Lukacs-Kornek, V., Kuligowski, M.P., Pitcher, L.A., Degn, S.E.,
Kim, Y.-A., Cloninger, M.J., Martinez-Pomares, L., Gordon, S., Turley, S.J.,
and Carroll, M.C. (2010). Capture of influenza by medullary dendritic cells
via SIGN-R1 is essential for humoral immunity in draining lymph nodes. Nat.
Immunol. 11, 427–434.
Hangartner, L., Senn, B.M., Ledermann, B., Kalinke, U., Seiler, P., Bucher, E.,
Zellweger, R.M., Fink, K., Odermatt, B., Burki, K., et al. (2003). Antiviral immune
responses in gene-targeted mice expressing the immunoglobulin heavy chain
of virus-neutralizing antibodies. Proc. Natl. Acad. Sci. USA 100, 12883–12888.
Hangartner, L., Zinkernagel, R.M., and Hengartner, H. (2006). Antiviral anti-
body responses: the two extremes of a wide spectrum. Nat. Rev. Immunol.
6, 231–243.
Huang, W., Sherman, B.T., and Lempicki, R.A. (2009). Systematic and integra-
tive analysis of large gene lists using DAVID bioinformatics resources. Nat.
Protoc. 4, 44–57.
Iannacone, M., Moseman, E.A., Tonti, E., Bosurgi, L., Junt, T., Henrickson,
S.E., Whelan, S.P., Guidotti, L.G., and von Andrian, U.H. (2010). Subcapsular
sinusmacrophages prevent CNS invasion on peripheral infection with a neuro-
tropic virus. Nature 465, 1079–1083.
8 Cell Reports 5, 1–8, October 31, 2013 ª2013 The Authors
Itohara, S., Mombaerts, P., Lafaille, J., Iacomini, J., Nelson, A., Clarke, A.R.,
Hooper, M.L., Farr, A., and Tonegawa, S. (1993). T cell receptor d gene mutant
mice: independent generation of a b T cells and programmed rearrangements
of g d TCR genes. Cell 72, 337–348.
Junt, T., Moseman, E.A., Iannacone, M., Massberg, S., Lang, P.A., Boes, M.,
Fink, K., Henrickson, S.E., Shayakhmetov, D.M., Di Paolo, N.C., et al. (2007).
Subcapsular sinus macrophages in lymph nodes clear lymph-borne viruses
and present them to antiviral B cells. Nature 450, 110–114.
Lauterslager, T.G., Florack, D.E., van der Wal, T.J., Molthoff, J.W., Langeveld,
J.P., Bosch, D., Boersma, W.J., and Hilgers, L.A. (2001). Oral immunisation
of naive and primed animals with transgenic potato tubers expressing LT-B.
Vaccine 19, 2749–2755.
Moseman, E.A., Iannacone, M., Bosurgi, L., Tonti, E., Chevrier, N., Tumanov,
A., Fu, Y.-X., Hacohen, N., and von Andrian, U.H. (2012). B cell maintenance of
subcapsular sinus macrophages protects against a fatal viral infection inde-
pendent of adaptive immunity. Immunity 36, 415–426.
Norton, J.T., Hayashi, T., Crain, B., Corr, M., and Carson, D.A. (2011). Role of
IL-1 receptor-associated kinase-M (IRAK-M) in priming of immune and inflam-
matory responses by nitrogen bisphosphonates. Proc. Natl. Acad. Sci. USA
108, 11163–11168.
Norton, J.T., Hayashi, T., Crain, B., Cho, J.S., Miller, L.S., Corr, M., andCarson,
D.A. (2012). Cutting edge: nitrogen bisphosphonate-induced inflammation is
dependent upon mast cells and IL-1. J. Immunol. 188, 2977–2980.
Pao, W., Wen, L., Smith, A.L., Gulbranson-Judge, A., Zheng, B., Kelsoe, G.,
MacLennan, I.C.M., Owen, M.J., and Hayday, A.C. (1996). g d T cell help of
B cells is induced by repeated parasitic infection, in the absence of other
T cells. Curr. Biol. 6, 1317–1325.
Phan, T.G., Grigorova, I., Okada, T., and Cyster, J.G. (2007). Subcapsular
encounter and complement-dependent transport of immune complexes by
lymph node B cells. Nat. Immunol. 8, 992–1000.
Pierre, P., Denis, O., Bazin, H., MbongoloMbella, E., and Vaerman, J.P. (1992).
Modulation of oral tolerance to ovalbumin by cholera toxin and its B subunit.
Eur. J. Immunol. 22, 3179–3182.
Qi, H., Egen, J.G., Huang, A.Y.C., and Germain, R.N. (2006). Extrafollicular
activation of lymph node B cells by antigen-bearing dendritic cells. Science
312, 1672–1676.
Renshaw, B.R., Fanslow, W.C., 3rd, Armitage, R.J., Campbell, K.A., Liggitt, D.,
Wright, B., Davison, B.L., and Maliszewski, C.R. (1994). Humoral immune re-
sponses in CD40 ligand-deficient mice. J. Exp. Med. 180, 1889–1900.
Scandella, E., Fink, K., Junt, T., Senn, B.M., Lattmann, E., Forster, R., Hengart-
ner, H., and Ludewig, B. (2007). Dendritic cell-independent B cell activation
during acute virus infection: a role for early CCR7-driven B-T helper cell collab-
oration. J. Immunol. 178, 1468–1476.
Schilbach, K., Geiselhart, A., and Handgretinger, R. (2001). Induction of
proliferation and augmented cytotoxicity of gammadelta T lymphocytes by
bisphosphonate clodronate. Blood 97, 2917–2918.
Scott, D.W., and Gershon, R.K. (1970). Determination of total and merecapto-
thanol-resistant antibody in the same serum sample. Clin. Exp. Immunol. 6,
313–316.
Thompson, K., Roelofs, A.J., Jauhiainen, M., Monkkonen, H., Monkkonen, J.,
and Rogers, M.J. (2010). Activation of gd T cells by bisphosphonates. Adv.
Exp. Med. Biol. 658, 11–20.
Tonti, E., Fedeli, M., Napolitano, A., Iannacone,M., von Andrian, U.H., Guidotti,
L.G., Abrignani, S., Casorati, G., and Dellabona, P. (2012). Follicular helper
NKT cells induce limited B cell responses and germinal center formation in
the absence of CD4(+) T cell help. J. Immunol. 188, 3217–3222.
Van Rooijen, N., and Sanders, A. (1994). Liposomemediated depletion ofmac-
rophages: mechanism of action, preparation of liposomes and applications.
J. Immunol. Methods 174, 83–93.
Wen, L., Pao, W., Wong, F.S., Peng, Q., Craft, J., Zheng, B., Kelsoe, G., Dia-
nda, L., Owen, M.J., and Hayday, A.C. (1996). Germinal center formation,
immunoglobulin class switching, and autoantibody production driven by
‘‘non alpha/beta’’ T cells. J. Exp. Med. 183, 2271–2282.

Supplemental Information
103
104
102
105
101 0
Neu
traliz
ing
Ig ti
ter
4 8 12 20Days after VSV
2816 24
PBS CLD
*
*** *** **
* **
103
104
102
105
101
Neu
traliz
ing
IgM
tite
r
0 4 8 12 20Days after VSV
2816 24
PBS CLD
** **
***
*
0 4 8 12 20Days after VSV
2816 24
103
104
102
105
101 N
eutra
lizin
g Ig
G ti
ter
PBS CLD
**
** *** **
C D E
103
104
102
105
101
PBS CLD
***
VS
V-sp
ecifi
c A
b tit
er (d
7)
D.L.
IgG1 IgG2c
0 2 4 6 8 100
5
10
15
CD
138+
B22
0+/P
op L
N (x
104 )
Days after VSV
PBS CLD
F G
PBS CLD day -3, -1 CLD day 0
103
104
102
105
101 0 4 8 12 20
Days after VSV28
Neu
traliz
ing
Ab
titer
16 24
***
***
** ***
** **
*** *** **
**
A
PBS
***
103
104
102
105
101
Neu
traliz
ing
Ab
titer
(d7)
CLD
*** B
Figure S1. Bisphosphonates Increase Antibody Responses to Live and Inactive Viruses, Proteins, Haptens and Existing Commercial Vaccine
Formulations, Related to Figure 1
(A) VSV nAb titers in the serum of C57BL/6mice that were footpad injected with CLD 3 days and 1 day (day�3,�1) or immediately (day 0) prior to VSV infection in
the same footpad. n = 5 per group. Black asterisks, PBS versus CLD day�3,�1; red asterisks PBS versus CLD day 0; results are representative of 3 independent
experiments.
(B) VSV nAb titers 7 days p.i. in the serum of C57BL/6 mice that were footpad injected with PBS or with the indicated doses of CLD prior to VSV infection in the
same footpad. n = 5 per group; results are representative of 2 independent experiments.
(C) VSV nAb titers in the serum of C57BL/6 mice that were footpad injected with PBS or CLD prior to infection in the same footpad with VSV. n = 5 per group;
results are representative of 3 independent experiments.
(D) Neutralizing IgM titers in the serum of the same mice described in (a).
(E) Neutralizing IgG titers of the same mice described in (a).
(F) VSV-specific IgG1 and IgG2c titers 7 day p.i. in the serum of C57BL/6 mice that were footpad injected with PBS or CLD prior to VSV infection in the same
footpad. n = 5 per group; results are representative of 2 independent experiments.
(G) Total number of CD138+ cells recovered from draining popliteal lymph nodes (Pop LN) that were pooled from 3 C57BL/6 mice footpad injected with PBS or
CLD prior to VSV infection in the same footpad. Results are representative of 2 independent experiments.
Results are expressed as mean ± SEM. ***: p < 0.001; **: p < 0.01; *: p < 0.05; D.L.: detection limit.
Cell Reports 5, 1–8, October 31, 2013 ª2013 The Authors S1
CD11b
Gr-
1
2 3 4
PBS 1.03
0
2
3
4
5
CLD 32.6
2 3 4 5
PBS+α-Gr-10
2 3
CLD+α-Gr-10.12
A
0
10
20
30
40
%fo
otpa
d G
r1hi
ghC
D11
b+
(of C
D45
+ ce
lls)
**
** *
B
PBS
CLD ALD
PBS
4.864.9
0.0740.01
TCRδ-/- WT
10.9
13
C
CD3
TCR
δ
WT WT
Neu
traliz
ing
Ab
titer
(d14
)
103
104
102
105
101
PBS CLD ALD PBS CLD ALD
WT TCRδ-/-
n.s. n.s.
n.s.
D
Figure S2. Neutrophils and gd T Cells Are Not Required for Bisphosphonate Adjuvant Activity, Related to Figures 2C and 2D
(A and B) Representative FACS plots (A) and quantification (B) of the number of Gr-1highCD11b+ cells present in digested footpads from the mice described in
Figure 2C. Numbers show the percentages of CD45+ cells within each gate. n = 4 per group; results are representative of 2 independent experiments.
(C) Representative FACS plots of digested footpads fromPBS-injected TCRd�/�mice or PBS-, CLD- or alendronate (ALD)-injected C57BL/6 (WT)mice. Numbers
show the percentages of CD45+ cells within each gate. Results are representative of 2 independent experiments (n = 3 per experiment).
(D) VSV nAb titers 14 days p.i. in the serum of C57BL/6 (WT) or TCRd�/� mice that were footpad injected with PBS, CLD or ALD prior to VSV infection in the same
footpad. n = 5 per group; results are representative of 3 independent experiments.
**: p < 0.01; *: p < 0.05; n.s.: non statistically significant.
S2 Cell Reports 5, 1–8, October 31, 2013 ª2013 The Authors
20000
15000
10000
5000
0
MFI
CD
86 400
300
200
100
0
MFI
CD
40
500
600
CLD CLD
Control
1.78 0.0146 0.0114 0.0013
DT
Day 0 p.i. Day 4 p.i. Day 14 p.i.
Spleen
0.13 0.0204 0.0043 0.0043
Pop LN
CD11c
GFP
A B
C
Figure S3. Bisphosphonate Adjuvant Activity Does Not Require Conventional Dendritic Cells, Related to Figure 2E
(A and B) CD86 (A) or CD40 (B) expression on purified CD11c+ DC that were cultured in the presence of 1 mg/ml LPS or with the indicated concentrations of CLD
for 48 hr prior to flow cytometry analysis.
(C) Representative FACS plots of digested spleens or popliteal lymph nodes (Pop LN) from CD11c-GFP-DTR/ C57BL/6 bone marrow chimeric mice that were
left untreated (control) or injected with diphtheria toxin (DT). Numbers show the percentage of cells within each gate. Results are representative of 2 independent
experiments (n = 3 per experiment). Results are expressed as mean ± SEM.
Cell Reports 5, 1–8, October 31, 2013 ª2013 The Authors S3

B PBS CLD B220 TCRβ CD169 B220 TCRβ CD169
100μm 100μm
Pop
LN
cel
ls (x
105 )
5
10
15
20
25
0 Total B T
A CLD
1
2
3
4
0
Pop
LN
cel
ls (x
105 )
* PBS
CD169
CD
11b
1.25 0.0866
0.0916 0.0966
PBS CLD
dextran sulfate carrageenan
1.25 0.0866
0.0916 0.0966
control DT
Neu
traliz
ing
Ab
titer
(d4)
103
104
102
105
101
n.s.
C D
Figure S4. Lymph Node Macrophage Depletion Per Se Does Not Increase Humoral Immune Responses, Related to Figure 2F
(A) Cellular subset frequency in popliteal lymph nodes (Pop LN) from C57BL/6 mice that were treated with PBS or CLD. n = 3 per group; results are representative
of 3 independent experiments.
(B) Representative confocal micrographs of Pop LN from C57BL/6 mice that were treated with PBS or CLD. Sections were stained with anti-B220 (gray), anti-
TCRb (red) and anti-CD169 (green) to identify B cells, T cells and macrophages, respectively. Scale bars represent 100 mm. Results are representative of 2
independent experiments (n = 3 per experiment).
(C) Representative FACS plots of digested popliteal lymph nodes from C57BL/6 mice that were treated with PBS, CLD, dextran sulfate or carrageenan. Numbers
show the percentage of cells within each gate. Results are representative of 2 independent experiments (n = 3 per experiment).
(D) VSV nAb titers 4 days p.i. in the serum of CD11c-GFP-DTR mice that were left untreated (control) or were depleted of CD169+ LN macrophages through a
single footpad injection of diphtheria toxin (DT) 6 days prior to VSV infection. These mice were described in Figure S3 from ref. (Iannacone et al., 2010).
Results are expressed as mean ± SEM. *: p < 0.05; n.s.: non statistically significant.
S4 Cell Reports 5, 1–8, October 31, 2013 ª2013 The Authors

PBS PBS CLD CLD VSV 8h
Row max
Row min
Figure S5. mRNA Expression Profiles of Differentially Expressed Genes in Whole Lymph Node, Related to Figure 3A
Shown are expression profiles for 4761 differentially expressed genes (rows) at 0 and 8 hr after VSV infection inmice treatedwith CLD or PBS as control (columns).
Shown are genes with at least a 2-fold change in expression between treatments in both duplicate arrays (each column corresponds to a replicate). Values from
multiple probe sets targeting the same gene were collapsed and gene-expression profiles were hierarchically clustered. Results are expressed as mean ± SEM.
Cell Reports 5, 1–8, October 31, 2013 ª2013 The Authors S5

Gene Symbol Fold change ImmGenCcl8 6.55Cd209f 5.58Retn 4.49Rya3 4.16Angptl4 3.54LOC100048885 3.54Gpr34 3.23Wasf1 2.85Car3 2.68Thrsp 2.53Ccl12 2.58Eps8l1 2.39Adipoq 2.38Dab2 2.36Cfh 2.29Clec10a 2.27Fcna 2.18Col12a1 2.12Mcoln3 2.28 B cellMcoln2 2.89 B cellP2ry12 3.95 Dendritic cellCd209d 5.41 Dendritic cellP2ry14 2.36 Dendritic cellIl22ra2 8.31 Dendritic cellCd209e 2.91 Dendritic cellSiglech 2.74 Dendritic cellHpgd 2.12 Dendritic cellEtv5 3.41 γδ T cellCpa3 3.49 γδ T cellAbca9 2.36 Macroph/monoScarb1 2.01 Macroph/monoF13a1 8.11 Macroph/monoFcgr1 2.69 Macroph/monoCpt1a 2.01 Macroph/monoCd209b 14.24 Macroph/monoFolr2 14.13 Macroph/monoCbr2 9.34 Macroph/monoTimd4 4.44 Macroph/monoMrc1 4.09 Macroph/monoMcpt4 3.55 Macroph/monoSlc15a2 3.44 Macroph/monoMarco 2.43 Macroph/monoHmox1 2.42 Macroph/monoCsf1r 2.34 Macroph/monoEdnrb 2.36 Macroph/monoFrmd4b 2.22 Macroph/monoMs4a8a 2.28 Macroph/monoTrf 2.24 Macroph/monoLyve1 2.15 Macroph/monoP2ry13 2.15 Macroph/monoCadm1 2.1 Macroph/monoRgl1 2.09 Macroph/monoGfra2 2.09 Macroph/monoCysltr1 2.03 Macroph/monoFabp4 2.02 Macroph/monoNinj1 2 Macroph/monoC1qa 2 Macroph/monoCd300ld 2.23 NeutrophilItgam 2.21 NeutrophilDdx60 2.18 NeutrophilCma1 4.56 NK cell
PBS CLD
Row max
Row min
Figure S6. mRNA Expression Profiles of Genes Decreased upon CLD Treatment inWhole Lymph Node before Infection, Related to Figure 3A
Shown are expression profiles for 62 differentially expressed genes (rows) with at least a 2-fold decrease in expression in CLD-treatedmice compared to controls
(PBS). The right-most column indicates the cell types in which each gene is specifically expressed based on the Immunological Genome (ImmGen) database.
Results are expressed as mean ± SEM.
S6 Cell Reports 5, 1–8, October 31, 2013 ª2013 The Authors
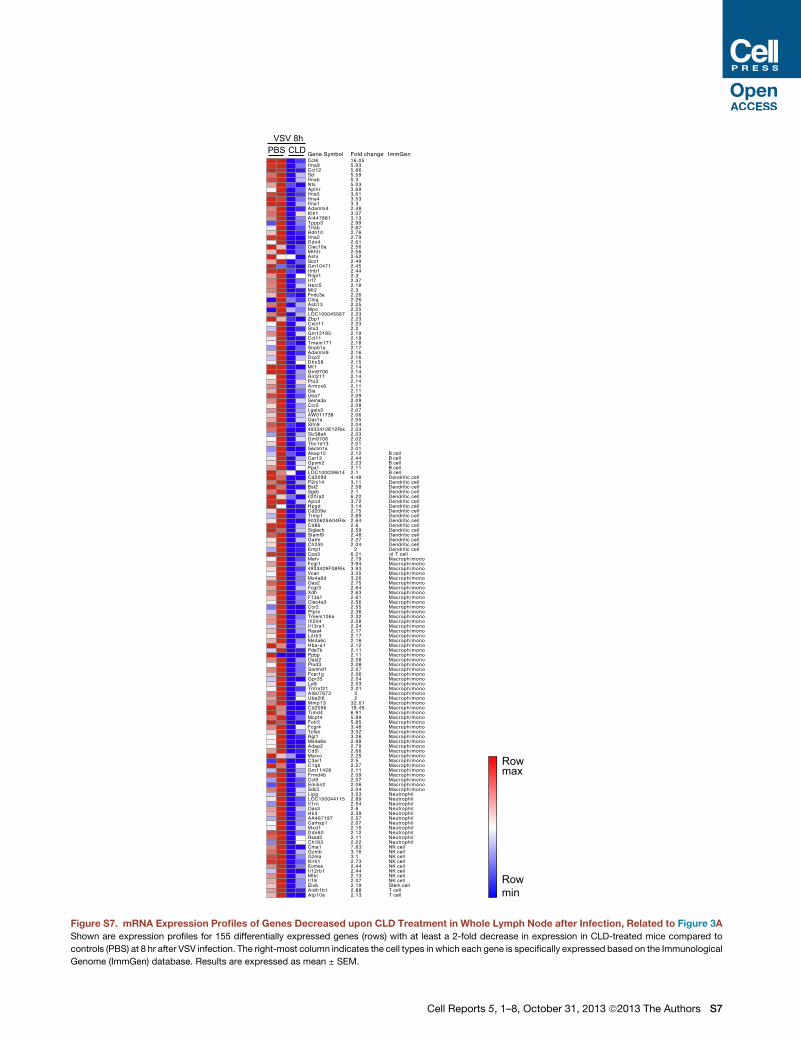
Gene Symbol Fold change ImmGenCcl8 16.05Ifna9 5.93Ccl12 5.86Sct 5.59Ifnab 5.3Nts 5.03Aplnr 3.69Ifna5 3.61Ifna4 3.53Ifna1 3.3Adamts4 2.48Klk1 3.07AI447881 3.13Tppp3 2.99Tifab 2.67Rdh10 2.76Ifna2 2.79Ddx4 2.61Clec10a 2.56Mthfr 2.56Astx 2.52Sco1 2.49Gm10471 2.45Ifnb1 2.44Rilpl1 2.3Irf7 2.37Herc5 2.18Mt2 2.3Fndc3a 2.26Ctsg 2.26Asb13 2.25Mpo 2.25LOC100045567 2.23Zbp1 2.23Cxcl11 2.23Stx3 2.2Gm12185 2.19Ccl11 2.19Tmem171 2.18Sirpb1a 2.17Adamts9 2.16Dcp2 2.16Dhx58 2.15Mt1 2.14Gm9706 2.14Rnf217 2.14Ptx3 2.14Armcx6 2.11Gla 2.11Uba7 2.09Sema3a 2.09Ccr5 2.08Lgals3 2.07AW011738 2.06Oas1a 2.05Slfn9 2.044933412E12Rik 2.03Slc38a4 2.03Gm9706 2.02Tbc1d13 2.01Sectm1a 2.01Akap12 2.12 B cellCar13 2.44 B cellGpsm2 2.23 B cellPpa1 2.11 B cellLOC100039614 2.1 B cellCd209d 4.48 Dendritic cellP2ry14 3.11 Dendritic cellBst2 2.58 Dendritic cellSgcb 2.1 Dendritic cellIl22ra2 6.22 Dendritic cellApod 3.72 Dendritic cellHpgd 3.14 Dendritic cellCd209e 2.75 Dendritic cellTimp1 2.69 Dendritic cell9030625A04Rik 2.64 Dendritic cellCd86 2.6 Dendritic cellSiglech 2.59 Dendritic cellSlamf9 2.48 Dendritic cellGatm 2.27 Dendritic cellCh25h 2.04 Dendritic cellEmp1 2 Dendritic cellCpa3 6.21 γδ T cellMefv 2.79 Macroph/monoFcgr1 3.94 Macroph/mono4933429F08Rik 3.93 Macroph/monoVcan 3.25 Macroph/monoMs4a6d 3.26 Macroph/monoOas2 2.75 Macroph/monoFcgr3 2.64 Macroph/monoXdh 2.63 Macroph/monoF13a1 2.61 Macroph/monoClec4a3 2.56 Macroph/monoCcr2 2.55 Macroph/monoPtpro 2.36 Macroph/monoTmem106a 2.32 Macroph/monoIfi204 2.28 Macroph/monoIl13ra1 2.24 Macroph/monoRasa4 2.17 Macroph/monoLilrb3 2.17 Macroph/monoMs4a6c 2.16 Macroph/monoHba-a1 2.12 Macroph/monoPde7b 2.11 Macroph/monoPpbp 2.11 Macroph/monoOasl2 2.08 Macroph/monoPlod3 2.08 Macroph/monoSamhd1 2.07 Macroph/monoFcer1g 2.06 Macroph/monoGpr35 2.04 Macroph/monoLy6i 2.03 Macroph/monoTnfrsf21 2.01 Macroph/monoAI607873 2 Macroph/monoUbe2l6 2 Macroph/monoMmp13 32.01 Macroph/monoCd209b 18.45 Macroph/monoTimd4 6.91 Macroph/monoMcpt4 5.89 Macroph/monoFolr2 5.85 Macroph/monoFcgr4 3.48 Macroph/monoTcfec 3.32 Macroph/monoRgl1 3.26 Macroph/monoMs4a8a 2.98 Macroph/monoAdap2 2.79 Macroph/monoCd5l 2.66 Macroph/monoMarco 2.25 Macroph/monoC3ar1 2.5 Macroph/monoC1qb 2.27 Macroph/monoGm11428 2.11 Macroph/monoFrmd4b 2.09 Macroph/monoCcl9 2.07 Macroph/monoEmilin2 2.06 Macroph/monoSdc3 2.04 Macroph/monoLipg 3.03 NeutrophilLOC100044115 2.89 NeutrophilIl1rn 2.54 NeutrophilOas3 2.6 NeutrophilHk3 2.39 NeutrophilAA467197 2.57 NeutrophilCarhsp1 2.07 NeutrophilMxd1 2.15 NeutrophilDdx60 2.12 NeutrophilRsad2 2.11 NeutrophilChi3l3 2.02 NeutrophilCma1 7.63 NK cellGzmb 3.16 NK cellGzma 3.1 NK cellKlrk1 2.73 NK cellEomes 2.44 NK cellIl12rb1 2.44 NK cellMlkl 2.13 NK cellIl18 2.07 NK cellEtv6 2.19 Stem cellAldh1b1 2.88 T cellAtp10a 2.13 T cell
PBSVSV 8h
CLDVSV 8hPBS CLD
VSV 8h
Row max
Row min
Figure S7. mRNA Expression Profiles of Genes Decreased upon CLD Treatment in Whole Lymph Node after Infection, Related to Figure 3A
Shown are expression profiles for 155 differentially expressed genes (rows) with at least a 2-fold decrease in expression in CLD-treated mice compared to
controls (PBS) at 8 hr after VSV infection. The right-most column indicates the cell types in which each gene is specifically expressed based on the Immunological
Genome (ImmGen) database. Results are expressed as mean ± SEM.
Cell Reports 5, 1–8, October 31, 2013 ª2013 The Authors S7
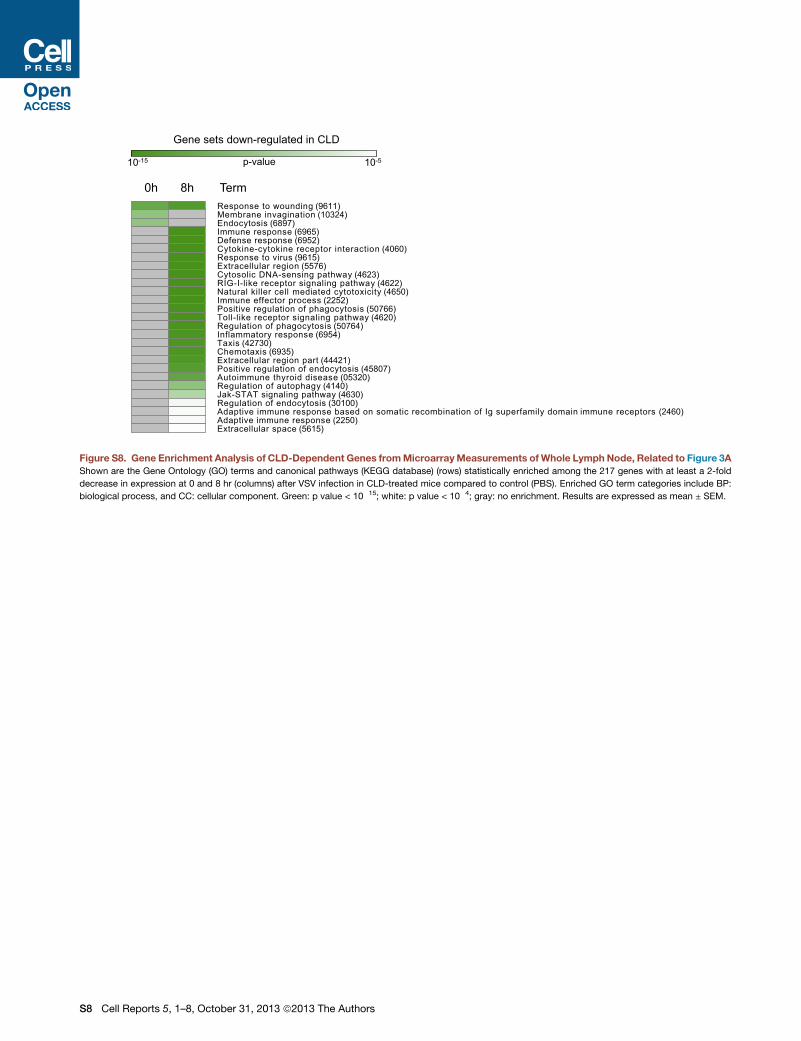
Gene sets down-regulated in CLD
p-value
0h 8h Term Response to wounding (9611)Membrane invagination (10324)Endocytosis (6897)Immune response (6965)Defense response (6952)Cytokine-cytokine receptor interaction (4060)Response to virus (9615)Extracellular region (5576)Cytosolic DNA-sensing pathway (4623)RIG-I-like receptor signaling pathway (4622)Natural killer cell mediated cytotoxicity (4650)Immune effector process (2252)Positive regulation of phagocytosis (50766)Toll-like receptor signaling pathway (4620)Regulation of phagocytosis (50764)Inflammatory response (6954)Taxis (42730)Chemotaxis (6935)Extracellular region part (44421)Positive regulation of endocytosis (45807)Autoimmune thyroid disease (05320)Regulation of autophagy (4140)Jak-STAT signaling pathway (4630)Regulation of endocytosis (30100)Adaptive immune response based on somatic recombination of Ig superfamily domain immune receptors (2460)Adaptive immune response (2250)Extracellular space (5615)
10-5 10-15
Figure S8. Gene Enrichment Analysis of CLD-Dependent Genes fromMicroarrayMeasurements ofWhole Lymph Node, Related to Figure 3A
Shown are the Gene Ontology (GO) terms and canonical pathways (KEGG database) (rows) statistically enriched among the 217 genes with at least a 2-fold
decrease in expression at 0 and 8 hr (columns) after VSV infection in CLD-treated mice compared to control (PBS). Enriched GO term categories include BP:
biological process, and CC: cellular component. Green: p value < 10�15; white: p value < 10�4; gray: no enrichment. Results are expressed as mean ± SEM.
S8 Cell Reports 5, 1–8, October 31, 2013 ª2013 The Authors
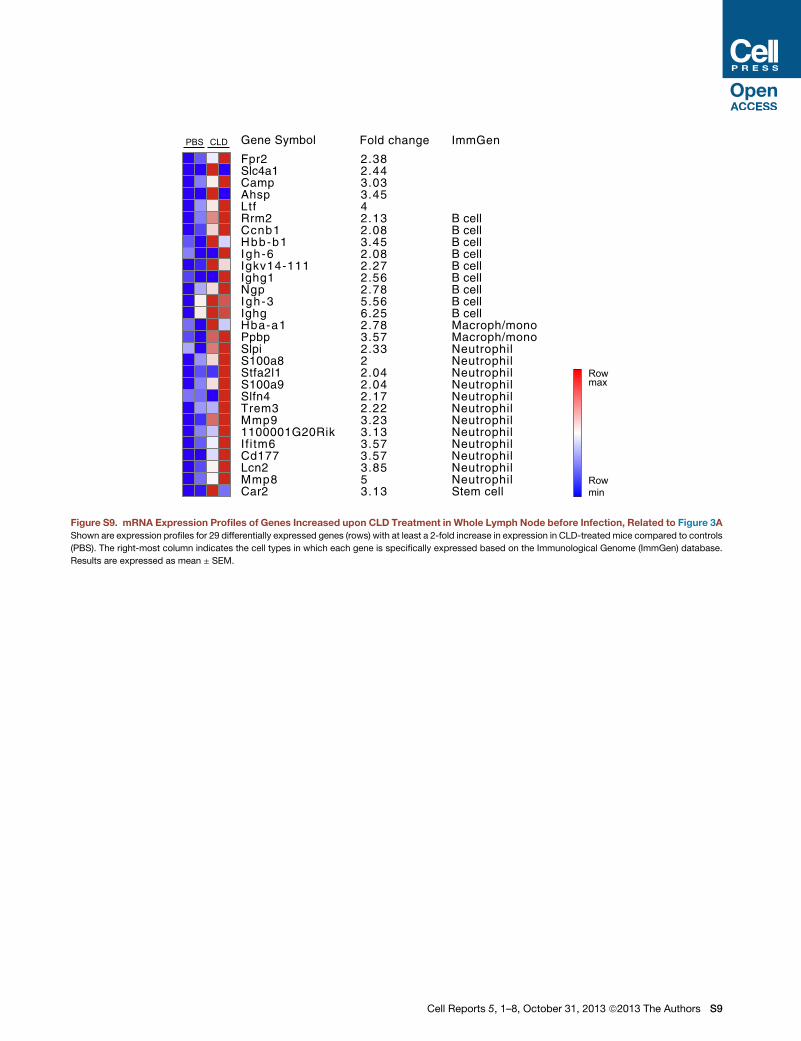
Gene Symbol Fold change ImmGen
Fpr2 2.38Slc4a1 2.44Camp 3.03Ahsp 3.45Ltf 4Rrm2 2.13 B cellCcnb1 2.08 B cellHbb-b1 3.45 B cellIgh-6 2.08 B cellIgkv14-111 2.27 B cellIghg1 2.56 B cellNgp 2.78 B cellIgh-3 5.56 B cellIghg 6.25 B cellHba-a1 2.78 Macroph/monoPpbp 3.57 Macroph/monoSlpi 2.33 NeutrophilS100a8 2 NeutrophilStfa2l1 2.04 NeutrophilS100a9 2.04 NeutrophilSlfn4 2.17 NeutrophilTrem3 2.22 NeutrophilMmp9 3.23 Neutrophil1100001G20Rik 3.13 NeutrophilI f i tm6 3.57 NeutrophilCd177 3.57 NeutrophilLcn2 3.85 NeutrophilMmp8 5 NeutrophilCar2 3.13 Stem cell
PBS CLD
Row max
Row min
Figure S9. mRNA Expression Profiles of Genes Increased upon CLD Treatment in Whole Lymph Node before Infection, Related to Figure 3A
Shown are expression profiles for 29 differentially expressed genes (rows) with at least a 2-fold increase in expression in CLD-treated mice compared to controls
(PBS). The right-most column indicates the cell types in which each gene is specifically expressed based on the Immunological Genome (ImmGen) database.
Results are expressed as mean ± SEM.
Cell Reports 5, 1–8, October 31, 2013 ª2013 The Authors S9

Gene Symbol Fold change ImmGen2310035P21Rik 2Selm 2Fhl1 2Chpt1 2.04Cilp 2.04Mmd 2.04Cdh2 2.08Cygb 2.083110080E11Rik 2.082510009E07Rik 2.08Higd1c 2.13Naa35 2.13Fmo1 2.13Prss23 2.13Cyp1b1 2.13Lum 2.17Vegfc 2.17Ebf3 2.17Maob 2.17Dclk1 2.179430047G12Rik 2.17Dsc2 2.17Evi5 2.17Parm1 2.17Il7 2.224930426D05Rik 2.22Fbxl7 2.22Nedd4 2.22Mxd4 2.27Fam168a 2.335830408C22Rik 2.33Irx3 2.332410066E13Rik 2.33Bnip3 2.33Synpo2 2.38Bok 2.38Arrb1 2.38Stard4 2.38AI256396 2.44Thrsp 3.334931406C07Rik 2.44Pdgfra 2.44LOC100047565 2.44Meox2 2.63Hist1h1c 2.5Plin4 2.63Timp4 2.63Vsig10 2.63Scara5 2.7Cyp26b1 2.86Aoc3 2.86Ghr 2.94Abca8a 2.94Odz3 3.03Retn 3.03Gdf10 3.33Inmt 3.45Atp1a2 3.45Cfd 3.7Car3 3.7Gm7202 3.85BB144871 3.85Adipoq 4.17Cxcl5 4.35Cyp2e1 7.14Lpl 2.22 B cellCcna2 2.27 B cellBmf 2.08 B cellNuak1 2.13 B cellCpm 2 B cellArhgef12 2.08 B cellCyp51 2.27 B cellFcer2a 2.38 B cellCiita 2.04 B cellCd55 2.17 B cellTmem204 2.08 B cellSrpk3 2.13 B cellCdc25b 2.13 B cellIgj 2.27 B cellProx1 2.44 B cellIgk-V28 2.63 B cellIgh-6 11.11 B cellIghg 7.69 B cellIghg1 8.33 B cellIgh-3 14.29 B cellZmynd15 2.17 Dendritic cellPard6g 2.08 Dendritic cellGria3 2.22 Dendritic cellPxdn 2 Dendritic cellCyp4b1 2 Dendritic cellGstm2 2.04 Dendritic cellAno1 2.04 Dendritic cellDsg2 2.04 Dendritic cellSlc36a2 3.13 Dendritic cellAdh1 2.04 γδ T cellC1ql3 2.17 γδ T cellSlc40a1 2.94 Macroph/monoB230120H23Rik 2.04 Macroph/monoIl13ra2 2 Macroph/monoMyl9 2.04 Macroph/monoNfia 2.08 Macroph/monoFbxo32 2.04 Macroph/monoKifap3 2.04 Macroph/monoSlco2b1 2.33 Macroph/monoTrim2 2.13 Macroph/monoTtc3 2.08 Macroph/monoDhrs3 2.22 Macroph/monoIdh1 2.27 Macroph/monoKitl 2.27 Macroph/monoCd36 2.56 Macroph/monoFabp4 2.56 Macroph/monoPon3 2.7 Macroph/monoSlc15a2 1 0 Macroph/monoGclm 2.04 NeutrophilSlpi 2.63 NeutrophilMmp9 3.33 NeutrophilAcsl1 2.56 NeutrophilCes3 4.17 NeutrophilNrarp 2.17 NK cellKlhl4 2.22 NK cellA930038C07Rik 2.63 NK cellMpdz 2.08 Stem cellMettl7a1 2.44 Stem cellSlc22a3 2.78 Stem cellSdpr 3.23 Stem cellSerpinf1 2.04 Stem cellHmgcs2 2.7 T cell
PBSVSV 8h
CLDVSV 8h
Row max
Row min
PBS CLD VSV 8h
Figure S10. mRNA Expression Profiles of Genes Increased upon CLD Treatment in Whole Lymph Node after Infection, Related to Figure 3A
Shown are expression profiles for 127 differentially expressed genes (rows) with at least a 2-fold increase in expression in CLD-treatedmice compared to controls
(PBS) at 8 hr after VSV infection. The right-most column indicates the cell types in which each gene is specifically expressed based on the Immunological Genome
(ImmGen) database. Results are expressed as mean ± SEM.
S10 Cell Reports 5, 1–8, October 31, 2013 ª2013 The Authors
Neu
traliz
ing
Ab
titer
(d7)
103
104
102
105
101
**
* *
** B
Neu
traliz
ing
Ab
titer
(d7)
103
104
102
105
101
A
PBS α-PDCA-1
n.s.
Figure S11. CLD Increases Ab Responses Independently of pDCs or NK Cells, Related to Figure 3A(A) VSV nAb titers 7 days p.i. in the serum of C57BL/6 mice that received PBS or were depleted of pDCs via anti-PDCA-1 (a�PDCA1) injection 24 hr prior to VSV
infection. These mice were described in Figure 3j and Suppl. Figure 8b from ref. (Iannacone et al., 2010).
(B) VSV nAb titers 7 days p.i. in the serum of C57BL/6 mice that were injected (or not) with anti-Asialo-GM1 depleting Ab (a�Asialo-GM1, seeMethods for details)
and were subsequently footpad injected with PBS or CLD immediately prior to VSV infection in the same footpad. n = 5; results are representative of 2 inde-
pendent experiments.
Results are expressed as mean ± SEM. **: p < 0.01; *: p < 0.05; n.s.: non statistically significant.
Cell Reports 5, 1–8, October 31, 2013 ª2013 The Authors S11
Related Documents











