CD44 Deficiency Attenuates Chronic Murine Ileitis COLM B. COLLINS,* JOHNSON HO, ‡ THEODORE E. WILSON,* JOSHUA D. WERMERS,* JOSÉ L. TLAXCA, § MICHAEL B. LAWRENCE, § MICHAEL SOLGA, JOANNE LANNIGAN, and JESÚS RIVERA–NIEVES* *Mucosal Inflammation Program, Division of Gastroenterology, University of Colorado Health Sciences Center, Denver, Colorado, ‡ Digestive Health Center of Excellence, § Department of Biomedical Engineering, and Flow Cytometry Core Facility, University of Virginia Health Sciences Center, Charlottesville, Virginia Background & Aims: Lymphocyte recruitment to sites of inflammation requires the sequential engage- ment of adhesion molecules and chemokine recep- tors. In the current studies we analyzed the role of CD44 for the development of chronic small-intestinal inflammatory infiltrates in vivo. Methods: By using a tumor necrosis factor (TNF)-driven model of chronic ileitis (ie, B6.129P-TNF AU-rich element [ARE] ) that recapit- ulates many features of Crohn’s disease, we noticed dynamic changes in the expression and functional state of CD44 and its ligand hyaluronan via enzyme- linked immunosorbent assay, real-time reverse-tran- scription polymerase chain reaction, immunohisto- chemistry, and flow cytometry. In addition, we assessed the role of lymphocyte populations during induction of ileitis through adoptive transfer studies, and generated CD44-deficient TNFARE mice to as- sess the role of CD44 for development of ileitis. Results: Soluble hyaluronan levels and expression of hyaluronan synthase-1 were increased in TNFARE mice. This coincided with increased expression of CD44 (including variant 7) and reactivity towards hyaluronan on CD4 T cells. CD44 was spatially co- localized with the gut-homing integrin 4 7 , spatially linking lymphocyte rolling with arrest. These cells had an effector phenotype because they lacked L- selectin and a higher proportion in diseased mice produced TNF and interleukin-2 compared with wild-type littermates. Lastly, CD4 but not CD8 T cells conferred ileitis to RAG / recipients and de- ficiency of one or both alleles of the CD44 gene resulted in attenuation of the severity of ileitis in TNFARE mice. Conclusions: Our findings sup- port an important role of CD44 expressed by CD4 and CD8 for development of ileitis mediated by TNF overproduction. U lcerative colitis (UC) is strictly a colonic disease, whereas Crohn’s disease (CD) involves the small bowel in two thirds of patients. 1 One explanation for this difference is that the recirculating effector/memory cell pool expresses distinct repertoires of adhesion molecules and chemokine receptors that drive differential traffic between the small and large bowels. 2 Our knowledge of the specific molecules that mediate small-intestinal hom- ing during chronic ileitis is limited, in part because of the lack of animal models that develop chronic inflammation localized to the terminal ileum. 3 The tumor necrosis factor (TNF)AU-rich element (ARE) model is 1 of only 2 mouse inflammatory bowel disease (IBD) models that develop chronic inflammation localized to the ileum, reminiscent of human CD in its histologic features and the pivotal role played by TNF in its pathogenesis. Targeted deletion of 69 base pairs within the AU-rich region of the TNF gene resulted in increased messenger RNA (mRNA) stability, increased TNF protein synthesis, and systemic levels of TNF. In addition, these mice develop deforming arthritis, similar to human rheumatoid arthritis, in both heterozygous and homozygous mice. 4 Interestingly, both CD and rheu- matoid arthritis respond therapeutically to TNF antago- nists (eg, infliximab, adalimumab, and certolizumab), supporting the relevance of this model for the investiga- tion of pathogenic mechanisms that mediate the human diseases. 5 TNF overproduction increases the expression of CD44 on CD8 lymphocytes; 6 however, within the CD8 T-cell population there are both effector and regulatory arms, defined by the presence or absence of CD44 or CD103 on the cell surface. 7 In the present study we investigated whether the inflammatory process affects the levels of hyaluronan (HA) or the enzymes that mediate its synthe- sis (ie, hyaluronan synthases [HAS]). We analyzed the proportion of CD4 T cells that express CD44 during early and established disease and examined potential integrins that support arrest in CD44-expressing lym- phocytes. We additionally investigated the capacity of CD4 and CD8 T cells to adoptively transfer ileitis to RAG / recipients and assessed the functional role of CD44 in ileitis by developing TNFARE mice deficient for CD44 on the cell surface. Abbreviations used in this paper: HA, hyaluronan; HAS, hyaluronan synthase; MFI, mean fluorescence intensity; MLN, mesenteric lymph node; PCR, polymerase chain reaction; SBDS, similarity bright detail quantification; SEM, standard error of the mean; TNF, tumor necrosis factor; WT, wild type. © 2008 by the AGA Institute 0016-5085/08/$34.00 doi:10.1053/j.gastro.2008.08.053 BASIC– ALIMENTARY TRACT GASTROENTEROLOGY 2008;135:1993–2002

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

C
CM
*E
BsmtCitiudslscaiasRhmChllhspwcfirTpaT
Ubdpab
GASTROENTEROLOGY 2008;135:1993–2002
D44 Deficiency Attenuates Chronic Murine Ileitis
OLM B. COLLINS,* JOHNSON HO,‡ THEODORE E. WILSON,* JOSHUA D. WERMERS,* JOSÉ L. TLAXCA,§
ICHAEL B. LAWRENCE,§ MICHAEL SOLGA,� JOANNE LANNIGAN,� and JESÚS RIVERA–NIEVES*
Mucosal Inflammation Program, Division of Gastroenterology, University of Colorado Health Sciences Center, Denver, Colorado, ‡Digestive Health Center of
xcellence, §Department of Biomedical Engineering, and �Flow Cytometry Core Facility, University of Virginia Health Sciences Center, Charlottesville, Virginiatill
(dlhiwiTatamnstd
opdtwhspeipCRCf
snqf
BA
SIC–
ALI
MEN
TARY
TRA
CT
ackground & Aims: Lymphocyte recruitment toites of inflammation requires the sequential engage-
ent of adhesion molecules and chemokine recep-ors. In the current studies we analyzed the role ofD44 for the development of chronic small-intestinal
nflammatory infiltrates in vivo. Methods: By using aumor necrosis factor (TNF)-driven model of chronicleitis (ie, B6.129P-TNF�AU-rich element [ARE]) that recapit-lates many features of Crohn’s disease, we noticedynamic changes in the expression and functionaltate of CD44 and its ligand hyaluronan via enzyme-inked immunosorbent assay, real-time reverse-tran-cription polymerase chain reaction, immunohisto-hemistry, and flow cytometry. In addition, wessessed the role of lymphocyte populations duringnduction of ileitis through adoptive transfer studies,nd generated CD44-deficient TNF�ARE mice to as-ess the role of CD44 for development of ileitis.esults: Soluble hyaluronan levels and expression ofyaluronan synthase-1 were increased in TNF�AREice. This coincided with increased expression ofD44 (including variant 7) and reactivity towardsyaluronan on CD4� T cells. CD44 was spatially co-
ocalized with the gut-homing integrin �4�7, spatiallyinking lymphocyte rolling with arrest. These cellsad an effector phenotype because they lacked L-electin and a higher proportion in diseased miceroduced TNF and interleukin-2 compared withild-type littermates. Lastly, CD4� but not CD8� T
ells conferred ileitis to RAG�/� recipients and de-ciency of one or both alleles of the CD44 geneesulted in attenuation of the severity of ileitis inNF�ARE mice. Conclusions: Our findings sup-ort an important role of CD44 expressed by CD4�
nd CD8� for development of ileitis mediated byNF overproduction.
lcerative colitis (UC) is strictly a colonic disease,whereas Crohn’s disease (CD) involves the small
owel in two thirds of patients.1 One explanation for thisifference is that the recirculating effector/memory cellool expresses distinct repertoires of adhesion moleculesnd chemokine receptors that drive differential traffic
etween the small and large bowels.2 Our knowledge ofhe specific molecules that mediate small-intestinal hom-ng during chronic ileitis is limited, in part because of theack of animal models that develop chronic inflammationocalized to the terminal ileum.3
The tumor necrosis factor (TNF)�AU-rich elementARE) model is 1 of only 2 mouse inflammatory bowelisease (IBD) models that develop chronic inflammation
ocalized to the ileum, reminiscent of human CD in itsistologic features and the pivotal role played by TNF in
ts pathogenesis. Targeted deletion of 69 base pairsithin the AU-rich region of the TNF gene resulted in
ncreased messenger RNA (mRNA) stability, increasedNF protein synthesis, and systemic levels of TNF. Inddition, these mice develop deforming arthritis, similaro human rheumatoid arthritis, in both heterozygousnd homozygous mice.4 Interestingly, both CD and rheu-atoid arthritis respond therapeutically to TNF antago-
ists (eg, infliximab, adalimumab, and certolizumab),upporting the relevance of this model for the investiga-ion of pathogenic mechanisms that mediate the humaniseases.5
TNF overproduction increases the expression of CD44n CD8� lymphocytes;6 however, within the CD8� T-cellopulation there are both effector and regulatory arms,efined by the presence or absence of CD44 or CD103 onhe cell surface.7 In the present study we investigatedhether the inflammatory process affects the levels ofyaluronan (HA) or the enzymes that mediate its synthe-is (ie, hyaluronan synthases [HAS]). We analyzed theroportion of CD4� T cells that express CD44 duringarly and established disease and examined potentialntegrins that support arrest in CD44-expressing lym-hocytes. We additionally investigated the capacity ofD4� and CD8� T cells to adoptively transfer ileitis toAG�/� recipients and assessed the functional role ofD44 in ileitis by developing TNF�ARE mice deficient
or CD44 on the cell surface.
Abbreviations used in this paper: HA, hyaluronan; HAS, hyaluronanynthase; MFI, mean fluorescence intensity; MLN, mesenteric lymphode; PCR, polymerase chain reaction; SBDS, similarity bright detailuantification; SEM, standard error of the mean; TNF, tumor necrosisactor; WT, wild type.
© 2008 by the AGA Institute0016-5085/08/$34.00
doi:10.1053/j.gastro.2008.08.053

wTaMATTiCoao
tc�ef
fsmt
�mpowt(
tCw(rvu(Em
BTmptpcedCrTCd(sct
t1wwc
csmsMhwrw(wplflsam1m
wf(
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1994 COLLINS ET AL GASTROENTEROLOGY Vol. 135, No. 6
Materials and MethodsMiceThe B6.129S-Tnftm2Gkl/Jarn strain (MGI: 3720980)
as generated by continuous backcrosses betweenNF�ARE mice on mixed genetic background (ie, C57BL6nd 129S6)4 to C57BL6/J mice as previously described.7
ice were kept under specific pathogen-free conditions.ll progeny were either heterozygous (TNF�ARE/�,NF�ARE) or carried no mutated alleles, wild-type (WT).he latter were used as the noninflamed controls. CD44-,
ntegrin �7-, and L-selectin– deficient mice on the57BL6/J background were obtained from Jackson Lab-ratories (Bar Harbor, ME). All animal procedures werepproved by the institutional committees for animal usef the Universities of Virginia and Colorado.
Detection of Serum HyaluronanConcentration by Enzyme-LinkedImmunosorbent AssaySerum from 30-week-old TNF�ARE and WT lit-
ermates was separated by centrifugation from red bloodells, snap-frozen in liquid nitrogen, and stored at80°C. Serum hyaluronan levels were determined by
nzyme-linked immunosorbent assay as per the manu-acturer’s instructions (Corgenix, Westminster, CO).
Tissue Collection and Histologic AnalysesThe distal ilea (10 cm) were fixed in 10% buffered
ormalin, embedded, cut into 3- to 5-�m sections, andtained with H&E. Histologic assessment of ileal inflam-
ation was performed using a standardized semiquanti-ative scoring system, as described previously.8
ImmunohistochemistryTerminal ilea were snap-frozen and sectioned (5
m) on a cryostat (Microm HM505N, Walldorf, Ger-any), incubated with biotinylated hyaluronan binding
rotein (US Biological, Swampscott, MA) in the presencer absence of soluble hyaluronan (Sigma, St. Louis, MO),hich served as a specificity control, followed by incuba-
ion with horseradish-peroxidase–labeled streptavidinBD Biosciences, San José, CA).
Real-Time Reverse-Transcription PolymeraseChain ReactionTotal RNA was isolated from homogenized ileal
issue using the RNeasy Mini Kit (Qiagen, Valencia,A) and converted to complementary DNA (cDNA)ith the GeneAmp RNA polymerase chain reaction
PCR) kit (Applied Biosystems, Foster City, CA) usingandom hexamers (0.75 �g of total RNA, final reactionolume 20 �L). cDNA was quantified by real-time PCRsing an ABI PRISM 7000 Sequence Detection System
Applied Biosystems). Primers (1.25 �L TaqMan Genexpression Assays: HAS-1, mm00468496_m1, HAS-2,
m00515089_m1, HAS-3, mm00515091_m1; Applied riosystems), 4 �L of first-strand cDNA, 12.5 �L ofaqMan 2� PCR master mix, and 7.25 �L of water wereixed together for each reaction. Each PCR reaction was
erformed in triplicate. Thermocycling conditions for theargets were as follows: 50°C for 2 minutes (iTaq DNAolymerase activation), 95°C for 10 minutes and 40 cy-les of 95°C for 15 seconds, and 60°C for 1 minute. Inach sample, 18S RNA expression was measured as en-ogenous control. The reaction volume (10 �L) forD44v7 contained iQ SYBR Green PCR Supermix (Bio-
ad, Hercules, CA), primer pairs (forward 5=CAGCCTGT-GGGTTGGTATT 3, reverse 5=CAGACTCAGGAGC-CACAAC 3=), and cDNA template. Samples were run inuplicate for 1 cycle (95°C for 10 s) followed by 40 cycles
95°C for 15 s, 60°C for 1 min). Fluorescence was mea-ured after each cycle. The ratio of mRNA expression wasalculated by the ��cycle threshold method (User Bulle-in no. 2; Applied Biosystems).
Lymphocyte IsolationSingle-cell suspensions were obtained by passing
he spleen or mesenteric lymph node (MLN) through a00-�m cell-strainer. Lamina propria mononuclear cellsere isolated as previously described.9 Red blood cellsere lysed by a 15-minute incubation in ammonium
hloride lysing reagent (BD PharM Lyse; BD Biosciences).
Adhesion Assay Under FlowFreshly isolated cells isolated from MLN were
ultured in complete medium (RPMI with 10% fetal calferum, 2 mmol/L L-glutamine, 100 U penicillin, 100
g/mL streptomycin, 10 mmol/L HEPES, 1 mmol/Lodium pyruvate, nonessential amino acids, 1 � 105
2ME) in the presence of anti-CD3 (5 �L/mL) for 24ours at 37°C. Polystyrene dishes were coated overnightith 500 �L of 2.5 mg/mL soluble rooster comb hyalu-
onan (Sigma). Nonspecific adhesion sites were blockedith 0.5% Tween-20 in Hank’s Balanced Salt Solution
HBBS) for 1 hour. Cell suspensions (2 � 106 cells/mL)ere drawn into the chamber at 37°C using a syringeump (Harvard Apparatus, South Natick, MA). Physio-
ogic flow conditions were produced in a 35-mm parallelow chamber (Glyco Tech, Rockville, MD) at a sheartress of 2 dyn/cm2. Equilibrium was attained and inter-ctions were monitored using an inverted phase-contrasticroscope (Diaphot–TMD; Nikon, Garden City, NY) at
0� magnification. Rolling cells were counted in 8 orore fields of view, changing fields every 25 seconds.
Flow Cytometry and Intracellular CytokineStainingCells from indicated compartments were incubated
ith antibodies against the following: mouse CD4 (GK1.5)or gating, CD44 (IM7), L-selectin (MEL-14), CD103M290), and integrin �7 (M293; BD Biosciences) or their
espective isotype controls. Additional controls included
cCiaCa(JF(
i3sa�fiw(wacwp
liocfioSS
icBtftbtlrCCvT5t
im
2w(t
tiincP
chiu1uedT
dpw(hfi
fltWrb(3rfh
BA
SIC–
ALI
MEN
TARY
TRA
CT
December 2008 CD44 REQUIREMENT IN CHRONIC MURINE ILEITIS 1995
ells isolated from respective organs of mice deficient forD44, L-selectin, and integrin �7. Separate cytokine stain-
ng was performed using the BD Cytofix/Cytoperm kits per the manufacturer’s instructions (BD Biosciences).ells were fixed with 2% paraformaldehyde and 4 –5 colornalyses were performed using the FACS Calibur systemBeckton–Dickinson Immunocytometry Systems, Sanosé, CA, modified by Cytek Development, Fremont, CA).urther analyses were performed using FLOWJo software
Tree Star Inc, Ashland, OR).
Multispectral Imaging Flow CytometryCells isolated from MLN of TNF�ARE mice were
ncubated with anti-integrin antibodies for 15 minutes at7°C to allow for capping of the integrins, washed, re-uspended in staining buffer, incubated with antibodiesgainst CD44 (IM7), integrin CD103 (M290), or integrin4�7 (DATK32) for 20 minutes at 4°C, then washed andxed with 2% paraformaldehyde. Images (10,000 events)ere acquired on the Imagestream imaging cytometer
Amnis, Seattle, WA). Unstained cells and cells stainedith a single antibody/fluorophore combination werecquired without bright field illumination and used forompensation. Spectral compensation and data analysisere performed using the IDEAS image analysis softwareackage (Amnis).The similarity bright detail score (SBDS) was calcu-
ated in a 3-step process: (1) determination of the open-ng residue image for each channel image; (2) calculationf a non-mean normalized Pearson correlation coeffi-ient; and (3) log transformation of the correlation coef-cient to give roughly Gaussian distributions, the meanf which is the SBDS.10,11 Poorly colocalized events haveBDS values of 1 to 2, whereas colocalized events haveBDS values of 3 or greater.
Induction of Ileitis by Transfer of T-CellSubsets to Recombination-ActivatingGene�/� MiceCD4� T cells from spleen of TNF�ARE mice were
solated by positive selection with anti-mouse CD4 mi-robeads as per the manufacturer’s instructions (Miltenyiiotec, Auburn, CA). Preliminary studies, which compared
he ability of positively or negatively selected CD4� to trans-er ileitis, showed that the selection method did not affecthe severity or time course of the resultant disease in recom-ination-activating gene (RAG)�/� recipients. The flow-hrough then was stained with fluorescein isothiocyanate–abeled anti-mouse CD8 antibody, phycoerythrin-labeledat anti-mouse CD103 and APC-labeled rat anti-mouseD44. Unfractionated CD8�, CD8�/CD44�, and CD8�/D103� subsets were separated using a fluorescence-acti-ated cell sorter Vantage SE Diva system (BD Biosciences).-cell fractions (�97% pure) (5 � 105) were suspended in00 �L of phosphate-buffered saline and injected intraperi-
oneally into 6-week-old female RAG�/� recipients. Their clea were harvested after 6 weeks and the severity of inflam-ation was assessed as described.8
StatisticsStatistical analyses were performed using the
-tailed Student t test or 2-way analysis of variance. Dataere expressed as mean � standard error of the mean
SEM). Statistical significance was set at a P value of lesshan .05.
ResultsIncreased Expression of Hyaluronan andHyaluronan Synthase-1 in TNF�ARE MiceWe compared the levels of hyaluronan in serum,
he expression of hyaluronan synthases, and the reactiv-ty of hyaluronan binding protein within the terminalleum. Levels of soluble hyaluronan were increased sig-ificantly in 30-week-old TNF�ARE mice with ileitisompared with WT mice (822 � 121 vs 410 � 51 ng/mL;� .01; Figure 1A).To try to identify the enzyme(s) responsible for in-
reased hyaluronan levels, we assessed the expression ofyaluronan synthases 1–3 (HAS-1, -2, and -3) (Figure 1B)
n ileum. Expression of HAS-1, but not HAS-2 or -3, wasp-regulated significantly during peak and late disease in0- and 30-week-old mice compared with WT mice (Fig-re 1B), because the severity of ileitis progressively wors-ns (Figure 1C). By contrast, HAS-3 appeared to decreaseuring late disease, yet the differences between WT andNF�ARE did not reach statistical significance.
Hyaluronan Is Expressed on Crypt Cells andInfiltrating Leukocytes in the Ileum ofTNF�ARE MiceAs shown in Figure 1D, hyaluronan localized pre-
ominantly to the crypt regions (Figure 1D, inset middleanel) and to the surface of infiltrating leukocytes, whichere more numerous in inflamed ileum of TNF�ARE mice
middle panel) than in WT mice (top panel). Pre-incubation ofyaluronan binding protein with soluble hyaluronan con-rmed signal specificity (Figure 1D, bottom).
Increased Reactivity and Rolling Flux Fractionsby CD4� T Cells in TNF�ARE MiceTo determine whether chronic inflammation in-
uenced the functional state of CD44 in vivo, we assayedhe ability of CD4� T cells freshly isolated from MLN of
T mice and TNF�ARE mice to bind fluorescent hyalu-onan. Significantly more T cells from TNF�ARE miceound fluorescent hyaluronan, both within the CD44high
45 � 5 vs 31 � 2; P � .01) and within the CD44low (23 �vs 7 � 4; P � .01; Figure 2A and B). Flow chamber
olling studies confirmed that a higher percentage of cellsrom TNF�ARE mice show rolling interactions on ayaluronan-coated flow chamber compared with WT
ontrols (53.2% � 7% increase; Figure 2C).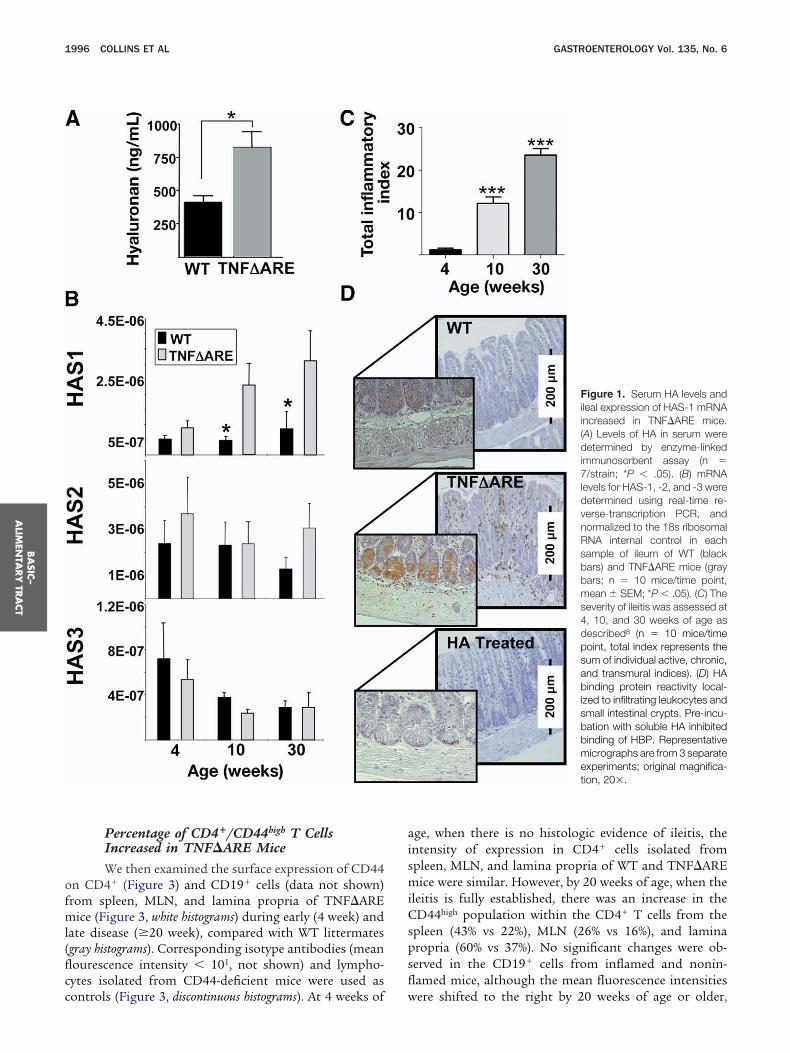
ofml(flcc
aismiCspsfl
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1996 COLLINS ET AL GASTROENTEROLOGY Vol. 135, No. 6
Percentage of CD4�/CD44high T CellsIncreased in TNF�ARE MiceWe then examined the surface expression of CD44
n CD4� (Figure 3) and CD19� cells (data not shown)rom spleen, MLN, and lamina propria of TNF�ARE
ice (Figure 3, white histograms) during early (4 week) andate disease (�20 week), compared with WT littermatesgray histograms). Corresponding isotype antibodies (meanourescence intensity � 101, not shown) and lympho-ytes isolated from CD44-deficient mice were used as
ontrols (Figure 3, discontinuous histograms). At 4 weeks of wge, when there is no histologic evidence of ileitis, thentensity of expression in CD4� cells isolated frompleen, MLN, and lamina propria of WT and TNF�ARE
ice were similar. However, by 20 weeks of age, when theleitis is fully established, there was an increase in theD44high population within the CD4� T cells from the
pleen (43% vs 22%), MLN (26% vs 16%), and laminaropria (60% vs 37%). No significant changes were ob-erved in the CD19� cells from inflamed and nonin-amed mice, although the mean fluorescence intensities
Figure 1. Serum HA levels andileal expression of HAS-1 mRNAincreased in TNF�ARE mice.(A) Levels of HA in serum weredetermined by enzyme-linkedimmunosorbent assay (n �7/strain; *P � .05). (B) mRNAlevels for HAS-1, -2, and -3 weredetermined using real-time re-verse-transcription PCR, andnormalized to the 18s ribosomalRNA internal control in eachsample of ileum of WT (blackbars) and TNF�ARE mice (graybars; n � 10 mice/time point,mean � SEM; *P � .05). (C) Theseverity of ileitis was assessed at4, 10, and 30 weeks of age asdescribed8 (n � 10 mice/timepoint, total index represents thesum of individual active, chronic,and transmural indices). (D) HAbinding protein reactivity local-ized to infiltrating leukocytes andsmall intestinal crypts. Pre-incu-bation with soluble HA inhibitedbinding of HBP. Representativemicrographs are from 3 separateexperiments; original magnifica-tion, 20�.
ere shifted to the right by 20 weeks of age or older,

c(
ca
t21s(
FWwac m 3
FspcclwaCcflclangsto4(dvnhntcT21*
BA
SIC–
ALI
MEN
TARY
TRA
CT
December 2008 CD44 REQUIREMENT IN CHRONIC MURINE ILEITIS 1997
onsistent with an age-related increase in expressiondata not shown).
When the absolute numbers of CD44high cells werealculated by correcting the percentages of expressiongainst the cellularity of spleen and MLN, an increase in
igure 2. Enhanced HA binding and rolling flux fraction of CD4� in TNT littermates were incubated with fluorescein isothiocyanate (FITC)-labas used as specificity control (data not shown). Cells were gated on ft 20 weeks of age. (C) Physiologic flow conditions were produced usinells in 8 or more fields of view (pooled mean � SEM rolling fraction fro
igure 3. Expansion of CD44high
ubset within CD4� T cells withrogression of ileitis in indicatedompartments of TNF�ARE miceompared with age-matched WT
ittermates. (A–C) Lymphocytesere incubated with anti-CD4,nd allophycocyanin- or PE-y5.5–labeled anti-CD44 mono-lonal antibodies and analyzed byow cytometry using CD44-defi-ient lymphocytes (discontinuous
ine) from the respective organsnd isotype antibody (MFI � 101,ot shown) as controls. Cells wereated on forward scatter, sidecatter, and indicated popula-ions. Representative histogramsbtained from 3–6 mice/group atand 20 or more weeks of age.
D) mRNA levels for CD44v7 wereetermined using real-time re-erse-transcription PCR, andormalized to the glyceralde-yde-3-phosphate dehydroge-ase (GAPDH) mRNA internal con-rol in each sample of ileum andolon of WT (gray bars) andNF�ARE mice (white bars) at0 or more weeks of age (n �0 mice/strain, mean � SEM;
P � .05).he frequency of CD4� CD44high cells occurs in MLN of0-week-old and older TNF�ARE mice (WT � 2.7 � 0.6 �06 vs TNF�ARE � 6.8 � 0.8 � 106; P � .01). Within thepleen the differences did not reach statistical significanceWT � 15.4 � 2.4 � 106 vs TNF�ARE � 16.8 � 3.3 �
E mice. (A and B) CD4� T cells from the MLN of TNF�ARE mice andA (HA-FITC) and analyzed by flow cytometry. Unconjugated soluble HA
d scatter, side scatter, and CD4. Representative data are from 4 micew chamber at 2 dyn/cm2. Rolling interactions were analyzed using 107
separate experiments; P � .05).
F�AReled Horwarg a flo

1a
dl(th
3hwtci
s
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1998 COLLINS ET AL GASTROENTEROLOGY Vol. 135, No. 6
06) because the cellularity of spleen decreases duringdvanced disease.
Expression of CD44 Variant Exon v7 mRNAIs Increased in Inflamed Ileum of TNF�AREMiceThe mRNA levels for variant exon 7 (CD44v7) were
etermined in the inflamed total ileum and noninflamed co-onic tissues from 20-week-old and older TNF�ARE micewhite bars) and WT (gray bars) using real-time reverse-transcrip-ion PCR, normalized to the glyceraldehyde-3-phosphate de-ydrogenase mRNA internal control in each sample (Figure
D). The expression of CD44v7 mRNA was significantlyigher in the inflamed ilea of TNF�ARE mice comparedith that of normal WT controls (gray bars). By contrast,
here was no difference in the expression within uninflamedolonic tissues of TNF�ARE and WT mice, supportingnduction within the ileum by inflammatory mediators.
Most CD44� T Cells Co-Express �7 Integrins,Which Colocalize With CD44 on theLymphocyte SurfaceWe investigated whether the expanded CD44�
ubset in TNF�ARE mice co-expressed �7 integrins and
Figure 4. CD4� T cells co-ex-pressed integrin �7 and CD44,which colocalized with integrin�4�7 on the cell surface. (A andB) Lymphocytes isolated fromMLN of TNF�ARE mice and WTsiblings were incubated with in-dicated antibodies and fluores-cein isothiocyanate (FITC)-labeledHA, and analyzed by flow cytom-etry. Cells were gated on for-ward scatter, side scatter, andCD4 using CD44- and integrin�7-deficient lymphocytes, andisotype antibodies (MFI � 101,not shown) as controls. Repre-sentative data were obtainedfrom 4 mice at 20 or more weeksof age run in duplicate. (C and D)Multispectral imaging flow cy-tometry suggested colocaliza-tion of CD44 and integrin �4�7
on the cell surface but not withCD103, as confirmed by SBDSdetermination (right panels; me-dian � SEM � 3.3 � 0.5 vs 0.5
� 0.4).
w(TgmwosaDsoadT(a(ncSt
pWTmTs.3
CtCaniTr
uffdTrtCRiTct
gfBtbshaPCa
sieCcrr
Fpma1pt
BA
SIC–
ALI
MEN
TARY
TRA
CT
December 2008 CD44 REQUIREMENT IN CHRONIC MURINE ILEITIS 1999
hether the gut-homing �7� T cells bind hyaluronan
Figure 4A and B). Most CD4� T cells in MLN of WT andNF�ARE mice expressed intermediate levels of �7 inte-rins (MFI, �102) and these �7
intermediate cells comprisedost of the cells that bind hyaluronan. Because �7 pairsith either integrins �4 or CD103, we investigated whichf the � subunits (ie, �4, CD103) is co-expressed on theurface of CD44-expressing cells by probing cells with anntibody that recognizes the �4�7 heterodimer (ie, cloneATK32) or CD103. Binding was analyzed using multi-
pectral imaging flow cytometry. The fluorescent imagesbtained (Figure 4C) suggested colocalization of CD44nd �4�7 on the cell surface. Data then were compared toetermine the degree of colocalization (ie, high SBDS).10
he median SBDS between �4�7 and CD44 was 3.3 � 0.5minimum, 2; maximum, 4.3), whereas between CD103nd CD44 was 0.5 � 0.4 (minimum, 0.2; maximum, 2.2)Figure 4D). The high score (�3) suggests that CD44 wasot only co-expressed with �4�7, but also spatially colo-alized on the cell surface. By contrast, the low medianBDS between CD103 and CD44 excluded colocaliza-ion.
Expanded CD4� Population Show Effector/Memory PhenotypeWe evaluated the phenotype of the CD4� T-cell
opulation from the spleen and MLN of TNF�ARE andT controls. The expanded CD44 population in
NF�ARE mice showed predominance of an effectoremory phenotype (CD44high/L-selectinlow, Figure 5).he percentages of effector CD4� cells increased in the
pleen (WT � 19% � 8% vs TNF�ARE � 30% � 7%; P �01) and in the MLN (WT � 15% � 3% vs TNF�ARE �1% � 9%; P � .01) of TNF�ARE mice.
igure 5. Expansion of effector CD4� T cells in TNF�ARE mice. Lym-hocytes from indicated compartments were incubated with indicatedonoclonal antibodies and analyzed by flow cytometry using CD44-
nd L-selectin–deficient lymphocytes and isotype antibodies (MFI �01, not shown) as controls. Representative CD44 and L-selectin ex-ression of cells gated on forward scatter, side scatter, and CD4 ob-
aained from 4 mice at 20 or more weeks of age.
Interleukin-2 and TNF Production Increasesin CD4�/CD44high T Cells From TNF�AREMice
To begin to understand the role of the CD4�/D44� subset in the disease process we assessed its cy-
okine profile. We found an increase in number ofD44high T cells that produce TNF in spleen (27% vs 18%)nd MLN (16% vs 9%) (Figure 6, left panels). Similarly, theumber of CD44high T cells that produce interleukin-2
ncreased in spleen (18% vs 13%) and MLN (12% vs 7%) ofNF�ARE mice compared with WT littermates (Figure 6,
ight panels).
CD4� T Cells Adoptively Transfer Ileitis toRAG�/� MiceTo assess the relative contribution of T-cell pop-
lations to intestinal inflammation, we adoptively trans-erred unfractionated CD4� or CD8� T cells isolatedrom spleen and MLN of TNF�ARE mice into immuno-eficient RAG�/� mice. Transfer of CD4� but not CD8�
cells induced ileitis in RAG�/� mice (Figure 7). Weecently described that the CD8�/CD103� subset con-ains regulatory T cells,7 thus we fractionated the 2 majorD8� subsets and transferred them individually intoAG�/� mice. CD8�/CD44� T cells were unable to
nduce ileitis even when the regulatory CD8�/CD103�
-cell subset was depleted.7 These findings suggest aritical effector role for the CD4� T-cell population forhe induction of ileitis.
CD44 Deficiency Results in Dose-DependentAttenuation of Ileitis in TNF�ARE MiceTo assess the role of CD44 in vivo in the patho-
enesis of ileitis, we generated TNF�ARE mice deficientor one or both alleles of the CD44 gene (Figure 8).linded histologic examination of the severity of ileitis in
he resultant progeny revealed considerable differencesetween inflammatory indices in ileal tissues from CD44-ufficient (�/�) mice (8.4 � 1.2, n � 35) to CD44eterozygous (�/�) mice (5.4 � 1.8; P � 0.01; n � 20)nd CD44-deficient (�/�) TNF�ARE mice (1.2 � 0.7;� .0001; n � 16). Partial or complete deficiency for
D44 in sibling mice that did not carry the TNF�AREllele did not develop ileitis or colitis (not shown).
DiscussionA prior study in the TNF�ARE mouse model of CD
howed that the CD8/CD44high T-cell population played anmportant role in disease development.6 The current studyxtends those observations to additionally show that CD4/D44� effectors also are important players. These CD4� T
ells have an effector surface phenotype, show enhancedeactivity to HA, produced TNF and interleukin-2, and wereequired for the induction of ileitis in RAG�/� mice. We
dditionally show that the synthesis of the CD44 ligand
hHTCg
eHmodw
sddgHlms
srh
FTmCCsfsaB2�omcgm
BA
SIC–
ALIM
ENTA
RY
TRA
CT
2000 COLLINS ET AL GASTROENTEROLOGY Vol. 135, No. 6
yaluronan is increased locally and in serum, and that theAS-1 enzyme is also increased in ileum of TNF�ARE mice.his increase is associated with increased expression ofD44 on the CD4�, which spatially colocalizes with inte-rin �4�7 on the cell surface.
Increased HA has been reported in human IBD.12 How-ver, the specific HAS isoform responsible for increasedA in the intestine has not been described. In TNF�AREice the expression of HAS-1 correlated with progression
f disease, suggesting that this enzyme may play a roleuring the progression of the disease. This is consistentith data from synoviocytes in which proinflammatory
igure 7. CD4� but not CD8�
cells transfer ileitis to RAG�/�
ice. (A) CD4�, unfractionatedD8, CD8/CD44�, or CD8/D103� cells isolated frompleen and MLN were trans-erred into RAG�/� mice and theeverity of ileitis was assessedfter 6 weeks, as described inurns et al8 (mean � SEM fromexperiments, n � 7/group; *P.05). (B) Representative histol-
gy of terminal ileum of RAG�/�
ice receiving vehicle or CD4� Tells. Representative micro-raphs are shown (H&E; original
agnification, 10�).timuli such as interleukin-1� and prostaglandin E2 in-uced expression of HAS-1.13,14 Another cytokine with aual role for the expression of HAS is transformingrowth factor-�, which stimulates HAS-1 yet inhibitsAS-3.15 We recently described a regulatory T-cell popu-
ation that produces transforming growth factor-�7 anday in part explain the trend towards decreased expres-
ion of HAS-3 observed during late disease.The localization of HA to leukocytes infiltrating the
mall-intestinal lamina propria is in agreement with aecent study.16 It is proposed that HA binding proteinas increased reactivity towards HA bound to CD44,
Figure 6. The proportion ofCD4�/CD44high T cells that pro-duce TNF and interleukin (IL)-2increased in TNF�ARE mice.Lymphocytes from indicated com-partments of WT or TNF�AREmice were stimulated with phorbolmyristate acetate/ionomycin, in-cubated with indicated antibod-ies, and analyzed by flow cytom-etry. Cells were gated on forwardscatter, side scatter, and CD4 us-ing CD44-deficient lymphocytesfor surface staining and isotypeantibody for intracellular stainingcontrols (MFI � 101, not shown).Representative density plots wereobtained from 3 experiments us-ing 4 mice per strain at 20 or moreweeks of age.

mebislweima
samfstfwit
omhh
mttwctermtCiTH
TaatCpsna
omcapTiiCMt�ohmifbaisl
terfiCds
Fmshmsm
BA
SIC–
ALI
MEN
TARY
TRA
CT
December 2008 CD44 REQUIREMENT IN CHRONIC MURINE ILEITIS 2001
ore than to free hyaluronate. Because crypt cells alsoxpress CD44, these cells were also reactive towards HAinding protein, similar to what was reported previously
n the large intestine.17 To a lesser degree we also ob-erved reactivity within the muscularis propria, particu-arly in inflamed TNF�ARE mice. This is in agreementith a prior study in which viral infection increased HA
xpression by smooth muscle cells.18 CD44/hyaluronannteractions thus may in part mediate the transmural
igration of effector (CD44�) lymphocytes in these mice,shared feature of this model and CD.Prior studies have shown that the total surface expres-
ion of CD44 does not necessarily correlate with thebility of the molecule to bind HA.19 Indeed, althoughany cell lines constitutively express activated CD44, its
unctional state in vivo is tightly regulated.20 This is noturprising because CD44 –HA interactions are ubiqui-ously relevant for cellular processes. We show that theraction of cells that carried CD44 in an activated stateas higher in TNF�ARE than WT littermates, support-
ng modulation of the reactivity of CD44 by inflamma-ory mediators in vivo.
CD44 is widely expressed on hematopoietic cells, but alson endothelium, reflecting its versatility as an adhesionolecule.21 Although many alternatively spliced isoforms
ave been described (CD44v1–10), the most common is the
igure 8. CD44 deficiency attenuates ileitis. (A) Active, chronic, trans-ural, and total inflammatory indices from ilea of 8-week-old CD44-
ufficient TNF�ARE mice (�/�; n � 35) were compared with miceeterozygous for CD44 (�/�; n � 21) and CD44-deficient TNF�AREice (�/�; n � 16). Attenuation of ileitis correlated with surface expres-
ion of CD44. Mean � SEM, *P � .05, ***P � .0001. (B) Representativeicrographs (H&E; original magnification, 10�).
ematopoietic or standard (CD44H or s), which is the C
ajor form expressed on leukocytes.22 A prior study showedhat the expression of CD44 was greatly increased withinhe CD8� population from spleen of TNF�ARE mice.6 Yet,hen we additionally analyzed the expression of CD44 on
ells originating from the MLN, we detected expansion ofhe CD44high subset also within the CD4�. Because thisxpansion is observed in mice at the peak of the diseaseather than during early disease and is not present in age-
atched WT littermates, these changes are attributable tohe chronic inflammatory process, supporting a role forD44-expressing CD4� T cells in chronic ileitis. Our find-
ngs are in agreement with work by other groups in whichh-1- and Th-2-polarized CD4� T cells use CD44 to roll onA.16,23
Among the CD44 isoforms, variant v7 is expressed byh-1–polarized T cells and macrophages. Immunoblock-de of this variant improved trinitrobenzene sulphoniccid colitis24 and CD44v6/v7-deficient mice were pro-ected from trinitrobenzene sulfonic acid. In agreement,D44v6/v7/interleukin-10 double-deficient mice wererotected from colitis.25 In our studies, increased expres-ion of CD44v7 in the inflamed ileum, but not in theormal colon, of TNF�ARE mice suggests that this vari-nt may additionally be important in ileitis.
A growing body of evidence suggests that there may beverlap in the function of molecules initially believed toediate exclusively the early steps of the adhesion cas-
ade (ie, tethering/rolling) with the later steps (ie, firmdhesion). Indeed, Nandi et al26 have shown that there ishysical association between CD44 and �4�1 (VLA-4).his novel finding spatially links the molecules involved
n rolling and arrest on the surface of the migrating celln an unprecedented manner.27 An association betweenD44 and the �4�7 integrin has not been described.ultispectral imaging flow cytometry suggested that
here is a similar physical association between CD44 and4�7, but not between CD44 and CD103, the expressionf which appear to be mutually exclusive. Prior studiesave suggested that �4�1 and �4�7 integrin not onlyediate arrest, but also the initial tethering and rolling
nteractions.28 Our findings allow us to propose that theoundation for those prior observations might have beenased on the close physical association between CD44 withny of the �4 integrins: �4�1 in the periphery or �4�7 in thentestine. Thus, molecules involved during early and latetages of the adhesion cascade appear to be physically co-ocalized on the cell surface.
Further analyses revealed that the CD44high cells withinhe CD4� population lacked L-selectin, indicative of anffector memory subset (CD44high/L-selectinlow), whichecirculate to inflammatory sites while limiting their traf-c through secondary lymphoid organs.29 The CD8/D44high T-cell population plays an important role inisease development in TNF�ARE mice.6 In this study wehow that the CD4�, which included CD44intermediates and
D44high, but not the CD8�, T cells from TNF�ARE mice
aRi
tsilfitT�ocdv
1
1
1
1
1
1
1
1
1
1
2
2
2
2
2
2
2
2
2
2
mRa
C
SD
BA
SIC–
ALIM
ENTA
RY
TRA
CT
2002 COLLINS ET AL GASTROENTEROLOGY Vol. 135, No. 6
doptively transferred ileitis into immunodeficientAG�/� mice, supporting a role for the CD4� cells dur-
ng induction of ileitis.A role for CD44 in ileitis was highlighted further by
he finding that decreased density of CD44 on the cellurface correlated with the level of attenuation ofleitis in TNF�ARE mice, suggesting that the effectikely is related to an important role for this moleculeor the trafficking of pathogenic leukocytes to theleum. Although the CD8� T cells originally werehought to be the critical effectors in ileitis ofNF�ARE mice based on the attenuating effect of2-microglobulin deficiency on the severity of ileitis,ur studies additionally suggest that CD4�/CD44� Tells also represent an important effector subset for theevelopment of the disease. CD44 thus may be a rele-ant therapeutic target in IBD.
References
1. Fiocchi C. Inflammatory bowel disease: etiology and pathogene-sis. Gastroenterology 1998;115:182–205.
2. Kunkel EJ, Butcher EC. Chemokines and the tissue-specific mi-gration of lymphocytes. Immunity 2002;16:1–4.
3. Strober W, Fuss IJ, Blumberg RS. The immunology of mucosalmodels of inflammation. Annu Rev Immunol 2002;20:495–549.
4. Kontoyiannis D, Pasparakis M, Pizarro TT, et al. Impaired on/offregulation of TNF biosynthesis in mice lacking TNF AU-rich ele-ments: implications for joint and gut-associated immunopatholo-gies. Immunity 1999;10:387–398.
5. Sandborn WJ, Targan SR. Biologic therapy of inflammatory boweldisease. Gastroenterology 2002;122:1592–1608.
6. Kontoyiannis D, Boulougouris G, Manoloukos M, et al. Geneticdissection of the cellular pathways and signaling mechanisms inmodeled tumor necrosis factor-induced Crohn’s-like inflammatorybowel disease. J Exp Med 2002;196:1563–1574.
7. Ho J, Kurtz CC, Naganuma M, et al. A CD8�/CD103high T cellsubset regulates TNF-mediated chronic murine ileitis. J Immunol2008;180:2573–2580.
8. Burns RC, Rivera-Nieves J, Moskaluk CA, et al. Antibody blockadeof ICAM-1 and VCAM-1 ameliorates inflammation in the SAMP-1/Yit adoptive transfer model of Crohn’s disease in mice. Gastro-enterology 2001;121:1428–1436.
9. Bamias G, Martin C, Mishina M, et al. Proinflammatory effects ofTH2 cytokines in a murine model of chronic small intestinalinflammation. Gastroenterology 2005;128:654–666.
0. George TC, Fanning SL, Fitzgeral-Bocarsly P, et al. Quantitativemeasurement of nuclear translocation events using similarityanalysis of multispectral cellular images obtained in flow. J Im-munol Methods 2006;311:117–129.
1. Beum PV, Lindorfer MA, Hall BE, et al. Quantitative analysis ofprotein co-localization on B cells opsonized with rituximab andcomplement using the ImageStream multispectral imaging flowcytometer. J Immunol Methods 2006;317:90–99.
2. de la Motte CA, Hascall VC, Drazba J, et al. Mononuclear leuko-cytes bind to specific hyaluronan structures on colon mucosalsmooth muscle cells treated with polyinosinic acid:polycytidylicacid: inter-alpha-trypsin inhibitor is crucial to structure and func-tion. Am J Pathol 2003;163:121–133.
3. Stuhlmeier KM. Prostaglandin E2: a potent activator of hyaluro-nan synthase 1 in type-B-synoviocytes. Biochim Biophys Acta
2007;1770:121–129. D4. Kao JJ. The NF-kappaB inhibitor pyrrolidine dithiocarbamateblocks IL-1beta induced hyaluronan synthase 1 (HAS1) mRNAtranscription, pointing at NF-kappaB dependence of the geneHAS1. Exp Gerontol 2006;41:641–647.
5. Stuhlmeier KM, Pollaschek C. Differential effect of transforminggrowth factor beta (TGF-beta) on the genes encoding hyaluronansynthases and utilization of the p38 MAPK pathway in TGF-beta-induced hyaluronan synthase 1 activation. J Biol Chem 2004;279:8753–8760.
6. Bonder CS, Clark SR, Norman MU, et al. Use of CD44 by CD4�Th1 and Th2 lymphocytes to roll and adhere. Blood 2006;107:4798–4806.
7. Alho AM, Underhill CB. The hyaluronate receptor is preferentiallyexpressed on proliferating epithelial cells. J Cell Biol 1989;108:1557–1565.
8. de La Motte CA, Hascall VC, Calabro A, et al. Mononuclearleukocytes preferentially bind via CD44 to hyaluronan on humanintestinal mucosal smooth muscle cells after virus infection ortreatment with poly(I.C). J Biol Chem 1999;274:30747–30755.
9. Siegelman MH, DeGrendele HC, Estess P. Activation and inter-action of CD44 and hyaluronan in immunological systems. J Leu-koc Biol 1999;66:315–321.
0. Lesley J, Howes N, Perschl A, et al. Hyaluronan binding functionof CD44 is transiently activated on T cells during an in vivoimmune response. J Exp Med 1994;180:383–387.
1. Koopman G, Taher TE, Mazzucchelli I, et al. CD44 isoforms,including the CD44 V3 variant, are expressed on endothelium,suggesting a role for CD44 in the immobilization of growth factorsand the regulation of the local immune response. Biochem Bio-phys Res Commun 1998;245:172–176.
2. Screaton GR, Bell MV, Jackson DG, et al. Genomic structure ofDNA encoding the lymphocyte homing receptor CD44 reveals atleast 12 alternatively spliced exons. Proc Natl Acad Sci U S A1992;89:12160–12164.
3. Siegelman MH, Stanescu D, Estess P. The CD44-initiated path-way of T-cell extravasation uses VLA-4 but not LFA-1 for firmadhesion. J Clin Invest 2000;105:683–691.
4. Wittig B, Schwarzler C, Fohr N, et al. Curative treatment of anexperimentally induced colitis by a CD44 variant V7-specific an-tibody. J Immunol 1998;161:1069–1073.
5. Wittig BM, Johansson B, Zoller M, et al. Abrogation of experimentalcolitis correlates with increased apoptosis in mice deficient forCD44 variant exon 7 (CD44v7). J Exp Med 2000;191:2053–2064.
6. Nandi A, Estess P, Siegelman M. Bimolecular complex betweenrolling and firm adhesion receptors required for cell arrest; CD44association with VLA-4 in T cell extravasation. Immunity 2004;20:455–465.
7. Dustin M. A supercode for inflammation. Immunity 2004;20:361–362.
8. Berlin C, Bargatze RF, Campbell JJ, et al. alpha 4 integrinsmediate lymphocyte attachment and rolling under physiologicflow. Cell 1995;80:413–422.
9. von Andrian UH, Mackay CR. T-cell function and migration. Twosides of the same coin. N Engl J Med 2000;343:1020–1034.
Received December 18, 2007. Accepted August 28, 2008.Address reprint requests to: Jesús Rivera-Nieves, MD, Mucosal Inflam-ation Program, University of Colorado Health Sciences Center, BRB,oom 742A, 4200 E. 9th Avenue, B146, Denver, Colorado 80206. e-mailddress: [email protected]; fax: (303) 315-5711.
The authors thank Sharon Hoang, Anthony Bruce, Oscar Castañón-ervantes, and Megan R. Ferguson for technical assistance.The authors disclose the following: Supported by US Public Health
ervice/National Institutes of Health grants DK067254 andK073280 (J. R.-N.) and by the University of Virginia Silvio Conte
igestive Health Research Center (grant DK56703).
Related Documents











