Cytometry Part B (Clinical Cytometry) 84B:207–217 (2013) CD38 and CD157: A Long Journey from Activation Markers to Multifunctional Molecules Valeria Quarona, 1 Gianluca Zaccarello, 1 Antonella Chillemi, 1 Enrico Brunetti, 1 Vijay Kumar Singh, 2 Enza Ferrero, 1,3 Ada Funaro, 1,3 Alberto L. Horenstein, 1,3 and Fabio Malavasi 1,3,4 * 1 Department of Medical Sciences, Laboratory of Immunogenetics, University of Torino Medical School, TORINO, Italy 2 Cancer Genomics Laboratory, Fondazione Edo ed Elvo Tempia, BIELLA, Italy 3 Centro di Ricerca in Medicina Sperimentale (CeRMS), University of Torino Medical School, Torino, Italy 4 Transplantation Immunology Service, Citta ` della Salute Hospital, TORINO, Italy CD38 (also known as T10) was identified in the late 1970s in the course of pioneering work carried out at the Dana-Farber Cancer Center (Boston, MA) that focused on the identification of surface mole- cules involved in antigen recognition. CD38 was initially found on thymocytes and T lymphocytes, but today we know that the molecule is found throughout the immune system, although its expression levels vary. Because of this, CD38 was considered an ‘‘activation marker,’’ a term still popular in routine flow cytometry. This review summarizes the findings obtained from different approaches, which led to CD38 being re-defined as a multifunctional molecule. CD38 and its homologue CD157 (BST-1), contiguous gene duplicates on human chromosome 4 (4p15), are part of a gene family encoding products that mod- ulate the social life of cells by means of bidirectional signals. Both CD38 and CD157 play dual roles as receptors and ectoenzymes, endowed with complex activities related to signaling and cell homeostasis. The structure-function analysis presented here is intended to give clinical scientists and flow cytomet- rists a background knowledge of these molecules. The link between CD38/CD157 and human diseases will be explored here in the context of chronic lymphocytic leukemia, myeloma and ovarian carcinoma, although other disease associations are also known. Thus CD38 and CD157 have evolved from simple leukocyte activation markers to multifunctional molecules involved in health and disease. Future tasks will be to explore their potential as targets for in vivo therapeutic interventions and as regulators of the immune response. V C 2013 International Clinical Cytometry Society Key terms: leukocyte markers; tissue distribution; cell receptor; ectoenzymes; calcium homeostasis How to cite this article: Quarona V, Zaccarello G, Chillemi A, Brunetti E, Singh VK, Ferrero E, Funaro A, Horen- stein AL, Malavasi F. CD38 and CD157: A Long Journey from Activation Markers to Multifunctional Molecules. Cytometry Part B 2013; 84B: 207–217. CD38, also known as T10, epitomizes a story shared by several other molecules, identified within a project aimed at probing the cell surface of human leukocytes using murine monoclonal antibodies (mAbs), reagents which became available in the late 1970s. Both probes and targets were unknown, prompting scientists to devise original strategies in order to identify the struc- ture and functions of the target molecules. The Leuko- cyte Workshop adopted a strategy that proved successful in the early days of HLA studies: when two or more mAbs reacted with the same target, they were said Grant sponsor: Associazione Italiana Ricerca Cancro (AIRC); Grant numbers: 11602, AIRC 5x1000 (partial); Grant sponsor: PRIN (Ministry of Education, University, and Innovation, Rome, Italy), from FIRB (Fondo per gli Investimenti della Ricerca di Base, Rome, Italy), from ex-60% Project (University of Torino). *Correspondence to: Fabio Malavasi, Lab of Immunogenetics, Uni- versity of Torino Medical School, Via Santena 19, 10126, Torino, Italy. E-mail: [email protected] Received 17 January 2013; Revision 6 March 2013; Accepted 21 March 2013 Published online 10 April 2013 in Wiley Online Library (wileyonlinelibrary.com). DOI: 10.1002/cyto.b.21092 Review Article V C 2013 International Clinical Cytometry Society

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Cytometry Part B (Clinical Cytometry) 84B:207–217 (2013)
CD38 and CD157: A Long Journey fromActivation Markers to Multifunctional Molecules
Valeria Quarona,1 Gianluca Zaccarello,1 Antonella Chillemi,1 Enrico Brunetti,1
Vijay Kumar Singh,2 Enza Ferrero,1,3 Ada Funaro,1,3 Alberto L. Horenstein,1,3 andFabio Malavasi1,3,4*
1Department of Medical Sciences, Laboratory of Immunogenetics, University of Torino Medical School,TORINO, Italy
2Cancer Genomics Laboratory, Fondazione Edo ed Elvo Tempia, BIELLA, Italy3Centro di Ricerca in Medicina Sperimentale (CeRMS), University of Torino Medical School, Torino, Italy
4Transplantation Immunology Service, Citta della Salute Hospital, TORINO, Italy
CD38 (also known as T10) was identified in the late 1970s in the course of pioneering work carriedout at the Dana-Farber Cancer Center (Boston, MA) that focused on the identification of surface mole-cules involved in antigen recognition. CD38 was initially found on thymocytes and T lymphocytes, buttoday we know that the molecule is found throughout the immune system, although its expression levelsvary. Because of this, CD38 was considered an ‘‘activation marker,’’ a term still popular in routine flowcytometry. This review summarizes the findings obtained from different approaches, which led to CD38being re-defined as a multifunctional molecule. CD38 and its homologue CD157 (BST-1), contiguousgene duplicates on human chromosome 4 (4p15), are part of a gene family encoding products that mod-ulate the social life of cells by means of bidirectional signals. Both CD38 and CD157 play dual roles asreceptors and ectoenzymes, endowed with complex activities related to signaling and cell homeostasis.The structure-function analysis presented here is intended to give clinical scientists and flow cytomet-rists a background knowledge of these molecules. The link between CD38/CD157 and human diseaseswill be explored here in the context of chronic lymphocytic leukemia, myeloma and ovarian carcinoma,although other disease associations are also known. Thus CD38 and CD157 have evolved from simpleleukocyte activation markers to multifunctional molecules involved in health and disease. Future taskswill be to explore their potential as targets for in vivo therapeutic interventions and as regulators of theimmune response. VC 2013 International Clinical Cytometry Society
Key terms: leukocyte markers; tissue distribution; cell receptor; ectoenzymes; calcium homeostasis
How to cite this article: Quarona V, Zaccarello G, Chillemi A, Brunetti E, Singh VK, Ferrero E, Funaro A, Horen-stein AL, Malavasi F. CD38 and CD157: A Long Journey from Activation Markers to Multifunctional Molecules.Cytometry Part B 2013; 84B: 207–217.
CD38, also known as T10, epitomizes a story sharedby several other molecules, identified within a projectaimed at probing the cell surface of human leukocytesusing murine monoclonal antibodies (mAbs), reagentswhich became available in the late 1970s. Both probesand targets were unknown, prompting scientists todevise original strategies in order to identify the struc-ture and functions of the target molecules. The Leuko-cyte Workshop adopted a strategy that provedsuccessful in the early days of HLA studies: when two ormore mAbs reacted with the same target, they were said
Grant sponsor: Associazione Italiana Ricerca Cancro (AIRC); Grantnumbers: 11602, AIRC 5x1000 (partial); Grant sponsor: PRIN(Ministry of Education, University, and Innovation, Rome, Italy), fromFIRB (Fondo per gli Investimenti della Ricerca di Base, Rome, Italy),from ex-60% Project (University of Torino).
*Correspondence to: Fabio Malavasi, Lab of Immunogenetics, Uni-versity of Torino Medical School, Via Santena 19, 10126, Torino,Italy.
E-mail: [email protected] 17 January 2013; Revision 6 March 2013; Accepted 21
March 2013Published online 10 April 2013 in Wiley Online Library
(wileyonlinelibrary.com).DOI: 10.1002/cyto.b.21092
Review Article
VC 2013 International Clinical Cytometry Society

to form a cluster centered on an unknown molecule.This approach (Cluster Designation, CD) made it possi-ble to identify an amazing number of surface moleculesin a limited number of years. However, it took manymore years to identify the functions exerted in vivo bythe majority of these molecules, an effort still far frombeing concluded.
CD38 was identified during the pioneering work ofE.L. Reinherz (Dana-Farber Cancer Center, Boston, MA)(1), focused on the identification of membrane mole-cules involved in antigen recognition. The fact thatthose studies were performed in a medical institutioninfluenced the first part of CD380s life history. The mole-cule was found in thymocytes and in activated normallymphocytes, and also in selected leukemias and myelo-mas (2). Its initial deployment in clinical diagnosis gaveCD38 the label of ‘‘activation marker,’’ a term still quitepopular in routine flow cytometry.
This review will attempt to summarize and update thefindings obtained from different areas of research, whichled to the redefinition of CD38 as a multifunctional mol-ecule. We now know that CD38 is joined by CD157(also referred to as bone marrow stromal cell antigen 1,BST-1) in being part of a gene family coding for productsthat modulate the social life of cells by means of bidirec-
tional signals (from outside to inside and vice versa).Besides being receptors, the same molecules act also asectoenzymes, endowed with complex activities.
A complete analysis of the structure and functions ofCD38 and CD157 in humans is intended to give clinicalscientists and flow cytometrists access to backgroundknowledge which is usually found only within the realmof basic science (3). The hope is that this set of informa-tion may improve the reading and comprehension ofresults obtained daily in clinics.
THE CD38 GENE FAMILY
CD38 and CD157 genes are located on human chro-mosome 4 (4p15) (4,5) (Fig. 1). Over 98% of the 70 kbCD38 gene is represented by intronic sequences. Thegene encompasses 8 exons: the largest, exon 1, deter-mines the intracytoplasmic and transmembrane regionsand the 33 membrane-proximal amino acids of theextracellular region (6,7). CD38 expression appears tobe under quite complex multilayered transcriptional reg-ulation. The first layer of control lies in the promoterregion characterized by the lack of a TATA box and thepresence of a CpG island (6), a methylation-controlledregion more frequently associated with housekeepingthan tissue-specific genes. A second layer of control is
FIG. 1. Schematic representation of human chromosome 4, with attention to CD38 (and its regulatory elements) and CD157. The right side showsthe schematic features of membrane CD38 and CD157 proteins.
208 QUARONA ET AL.
Cytometry Part B: Clinical Cytometry

likely to be upstream of the CpG island, where potentialbinding site for transcription factors (e.g., T cell tran-scription factor-1, nuclear factor for interleukin-6) lie(6), while a third level of gene control lies within thefirst intron of the CD38 gene. The 50-end of the intron 1contains responsive elements for retinoic acid (8) andperoxisome proliferator-activated receptor c (PPARc)(9). Intron 1 is also the location of a single nucleotidepolymorphism (SNP) (10) that binds the transcriptionfactor E2A (11) and whose variants are differentiallyexpressed in pathology (vide infra).
The CD157 gene extends for over 35 kb, very closeto its paralogue CD38 and consists of nine exons (4,12).CD38 and CD157 are highly conserved, as demonstratedby the fact that exons 2-8 are similar in length and main-tain the same phase of intron insertion (13). The50-flanking region of the human CD157 gene shows con-sensus sequences for interferon responsive elements(c-IREs and ISRE-like), for nuclear transcription factors(E2A, AP2, AP3, PEA3, C/EBP, CREB, Sp1), for cytokine-responsive factors (NF-IL-6, NF-jB), and for p53 (4). Thisgene also lacks a TATA box, suggesting multiple tran-scription start sites (4,14). All these elements suggestthat the CD157 gene may be upregulated by eventssuch as DNA damage, inflammation and infection,whereas NF-IL-6 and NF-jB binding sites may explainthe increased expression of the CD157 molecule inpatients affected by rheumatoid arthritis (12).
TISSUE DISTRIBUTION OF CD38 AND CD157
CD38 expression was first observed on thymocytesand T lymphocytes (15). Today, CD38 is considered vir-tually ubiquitous, at least in the immune system, butwith variable expression levels. Table 1 lists the main tis-sues and cells where CD38 and CD157 are detectable.Because CD157 joined the family relatively later, data onits distribution is still limited.
Flow cytometric analyses indicate that human CD157is constitutively expressed by myeloid cells in peripheralblood mononuclear cells (PBMCs). The molecule is alsoexpressed by synovial, vascular endothelial and folliculardendritic cells (16). Moreover, CD157 is also present onother cell types and tissues, such as dermal fibroblasts,human mast cells from lung, uterus, foreskin, and perito-neal mesothelial cells, among others (17–21). CD157was recently reported to be expressed in Paneth cells,where it mediates the effects of calorie restriction andrapamycin on murine intestinal stem cell function (22).
CD38 AS A RECEPTOR
An initial function attributed to CD38 was the regula-tion of activation and proliferation of human T lympho-cytes. Early functional studies of unidentified moleculeswere pursued by monitoring the effects induced by theengagement of different domains of the target by a panelof specific mAbs. The rationale was that effects inducedby ligation with a surrogate mAb would mimick thoseinduced by a ligand still unknown at the time. CD38engagement was followed by the activation of selected
PBMC populations (23). The identification of a first puta-tive ligand was obtained by exploiting the observationthat human T lymphocytes tended to adhere to endothe-lial cells (24). Experiments blocking this adhesion con-cluded that CD31 [also known as PECAM-1, a memberof the immunoglobulin (Ig) superfamily crucial to leuko-cyte adhesion and transmigration (25)] was a non-sub-strate ligand for CD38. It was later demonstrated thatCD38/CD31 interactions trigger the same signaling cas-cade and recapitulate the biological events observedusing agonistic mAbs (26,27). The interplay betweenCD38 and CD31 is crucial for leukocyte migration
Table 1Distribution of CD38 and CD157 in normal human tissues
CD38 CD157
LymphoidBlood T cells (precursors,
activated)Neutrophils
B cells (precursors,activated)
Eosinophils
Myeloid cells (mono-cytes, macrophages,dendritic cells)
Basophils
NK cells MonocytesErythrocytes MacrophagesPlatelets Plasmocytoid
dendritic cellsCord Blood T and B lymphocytes,
monocytesBone Marrow Precursors B cells precursors
Plasma cells Myeloid precursorsStromal cells
Thymus Cortical thymocytesLymph nodes Germinal center B
cellsFollicular dendritic
cellsSpleen Reticular cells
(white pulp)Non-lymphoidLiver Fetal B cell
progenitorsVessels Mast cellsBrain Purkinje cells
Cultured astrocytesCerebellum
Eye CorneaRetinal gangliar cells
Prostate Epithelial cellsGut Intraepithelial
lymphocytesBrush border, epi-
thelial cells ofvilli
Lamina proprialymphocytes
Peyer’s patches
Stromal cells incryptopatches
Isolated lymphoidfollicles
Peritoneum Mesothelial cellsMacrophages/
peritonealexudates
Pancreas b-cells a and b-cellsMuscle Sarcolemma (smooth
and striatedmuscle)
Bone OsteoclastsKidney Glomeruli Collecting tubuli
CD38 AND CD157 209
Cytometry Part B: Clinical Cytometry

through the endothelium (28). The CD38/CD31 cross-talk has been extensively analyzed in a number of differ-ent environments ranging from T to B, NK, and myeloidcells, in normal and pathological conditions (29). CD38-mediated signals are regulated at distinct levels: the firstlevel concerns the ultrastructural organization of themolecule, which exists both in monomeric (2) and di-meric (30) (or multimeric) type I forms (31). A flip-flopmechanism of membrane positioning has been recentlyproposed, with a type III form of CD38 displaying itscatalytic site in the cytoplasm (32). The second level isbased on the dynamic localization of CD38 in lipidmicrodomains within the plasma membrane. Lateralassociations with other proteins, which vary accordingto the cell lineage, determine a third level of control.Lipid raft localization and association with professionalsignaling complexes are pre-requisites for signals medi-ated through CD38 (33).
CD157 AS A RECEPTOR
The receptor and signaling features of CD157 havealso been investigated by using agonistic mAbs to mimicputative ligand(s). By doing so, it has been demonstratedthat CD157 ligation induces tyrosine phosphorylation ofa 130 kDa protein, identified as focal adhesion kinase(FAK), in myeloid cell lines (34,35), and that CD157engagement regulates Ca2þ homeostasis and mediatessuperoxide (O2�) production in the human myelomono-cytic U937 line (36). Accumulating evidence indicatesthat CD157 is a key player in the control of leukocyteadhesion, migration and diapedesis (37,38). In this con-text, CD157 behaves as a receptor by establishing lateralinteractions with other transmembrane molecules, thusovercoming its structural limitation (i.e., of being a GPI-anchored molecule) and acquiring the ability to trans-duce signals (13). In more detail, CD157 interacts withb1 and b2 integrins, and Ab-induced cross-linking ofCD157 promotes relocation of these complexes into de-tergent-resistant membrane domains (21). Moreover,CD157 effectively contributes to the integrin-driven sig-naling network, which is critical during leukocyte trans-migration, leading to optimal phosphorylation oftyrosine kinase receptors and activation of PI3K andMAPK signaling cascades (39).
CD38 AND CD157 ENZYMATIC FUNCTIONS
The enzymatic functions of the two proteins wereinvestigated after the enzyme ADP-ribosyl cyclase(ADPRC, purified from the mollusk Aplysia californica)was observed to display a striking similarity in proteinsequence with human CD38 (40). CD38 is a multifunc-tional enzyme that catalyzes the synthesis of cyclic ADP-ribose (cADPR) from nicotinamide adenine dinucleotide(NADþ ) and also mediates the hydrolysis of cADPR toADPR (41–44). In acidic conditions, CD38 catalyzes thegeneration of nicotinic acid adenine dinucleotide phos-phate (NAADP) from nicotinamide adenine dinucleotidephosphate (NADPþ ) (45). cADPR, ADPR, and NAADPbind different receptors and channels involved in the
regulation of cytoplasmic Ca2þ fluxes, activating signal-ing pathways critical for several biological processes[e.g., lymphocyte proliferation (46,47), cardiac (48) andintestinal longitudinal muscle contraction (49), glucose-induced insulin release in pancreas (50)]. The role ofCD38 and of its products in regulating a wide range ofphysiological functions is indicated by the multipledefects revealed in CD38 knock-out (KO) mice. Theseinclude impairment of neutrophil chemotaxis, defectiveoxytocin (OT) release, and aberrant social behavior (51).
The recombinant soluble extracellular domain ofCD38 mediates ADP-ribosylation of cysteine residues ofseveral proteins, including CD38 itself (52). CD38 is alsomodified by ecto-ADP-ribosyltransferases (ARTs). Argi-nine ADP-ribosylation results in inactivation of both cy-clase and hydrolase activities, whereas cysteine ADP-ribosylation leads only to inhibition of the hydrolase ac-tivity. Arginine ADP-ribosylation causes a decrease in in-tracellular cADPR and a subsequent decrease in Ca2þ
influx, with consequent death of activated T lympho-cytes (53). Moreover, CD38 is the major NAD glycohy-drolase (NADase) in mammalian cells, regulatingintracellular NADþ levels. CD38 thus modulates the ac-tivity of sirtuins, intracellular NADþ -dependent deacety-lases implicated in ageing, cell protection and energymetabolism (54,55).
CD157 also cleaves extracellular NADþ , generatingcADPR and ADPR (13,36). However, the catalytic effi-ciency of CD157 in generating cADPR is even lowerthan that of CD38 (13).
The products derived from NADþ cleavage operatedby CD38 or CD157 can also act as extracellular immuno-modifiers (56). Emerging data indicate that these prod-ucts may operate outside the cell as paracrine factors(57). Moreover, the catalytic reactions generate sub-strates for ARTs and poly-ADP-ribose polymerases(PARPs) involved in cell signaling, DNA repair and apo-ptosis (56).
CD38 is also found in soluble form in normal andpathological fluids (58) and in exosomes, which aremembrane vesicles secreted by B cells and likely a com-ponent of an intercellular communication network (59).
CD38 AND DISEASE
Chronic Lymphocytic Leukemia (CLL). CLL is acommon adult leukemia which results from the accumu-lation of small B (CD19þ /CD5þ /CD23þ ) lymphocytesin blood, bone marrow (BM), lymph nodes (LN), andother lymphoid tissues (60). The latter districts repre-sent permissive niches, where lymphocytes can prolifer-ate in response to microenvironmental signals (61).Elevated expression of CD38 in CLL cells is generallyassociated with advanced disease stage, higher incidenceof lymphadenopathy, high-risk cytogenetics, shorter lym-phocyte doubling time, shorter time to first treatmentand poorer response to therapy. Besides being a prog-nostic marker, CD38 is a component of a molecular
210 QUARONA ET AL.
Cytometry Part B: Clinical Cytometry

network which delivers growth and survival signals toCLL cells (62). CD38 acts as a receptor in leukemic cellsand its signals are mediated by ZAP70, another negativeprognostic factor for the disease and a limiting factor forCD38-mediated activation (33,63). CD38 can work inassociation with chemokines and their receptors, mainlyCXCL12/CXCR4, influencing the migratory responsesand contributing to the recirculation of neoplastic cellsfrom blood to lymphoid organs (64) and with specificadhesion molecules belonging to the integrin family(65,66). CD38 engagement by means of the ligand CD31(or surrogate agonistic mAb) triggers the activation of anintracellular signaling pathway and these signals increasechemotaxis as well as proliferation of neoplastic B cells.Together, this evidence contributes to the current viewof CLL as a chronic disease in which the host’s microen-vironment promotes leukemic cell growth and also con-trols the sequential acquisition and accumulation ofgenetic alterations (62).
Multiple Myeloma (MM). MM is a malignancy char-acterized by accumulation of monoclonal plasma cells inBM, a high concentration of monoclonal Igs in serumand urine, and lytic bone lesions. The proliferation ofneoplastic plasma cells in MM interferes with the normalproduction of blood cells, while the monoclonal Igimpairs humoral immunity.
There are several issues suggesting that CD38 playssignificant role(s) in MM. First, CD38 is expressed byplasma cells (normal and tumoral) at top levels of cellsurface density. Secondly, experiments in murine modelsshowed that CD38 is a key regulator of OT levels in bio-logical fluids while OT is also released by human osteo-blasts (67). A receptor for OT (OTR) is detectable onmyeloma cells and derived lines (Malavasi F., unpub-lished results, 2012). CD38 is expressed by osteoblastsand osteoclasts, where it implements signals leading toIL-6 release and inhibition of bone resorption (68,69).
In light of these considerations, plasma cells (andtheir malignant counterpart) and bone niches are goodtesting grounds for assessing the presence of a connec-tion between ectoenzymes and neuropeptides (70). Thesystem is closed, and nucleosides represent additionalsignals to those led by cytokines/chemokines and otherconventional regulators: ATP and NADþ operating inloco may complement the physiological regulatory sys-tems of plasma cells.
Acute Promyelocytic Leukemia (APL). APL is aunique subtype of acute leukemia, which causes anarrest of leukocyte differentiation at the promyelocytestage. Retinoic acid (RA) is included in therapeuticalprotocols for its ability to induce the differentiation ofleukemic cells into mature granulocytes. This therapymay be associated with retinoic acid syndrome (RAS), aclinical manifestation characterized by fever, dyspnea,pulmonary edema and infiltrates (71). Normal granulo-cytes are CD38-, while RA-treated APL cells express highamounts of the molecule (72,73). The aberrant expres-sion of CD38 on leukemic cells enhances their propen-sity to interact with CD31, expressed by lung
endothelial cells, resulting in a local production ofinflammatory cytokines, apoptosis of endothelial cellsand eventually contributing to the development of RAS(71).
CD157 AND DISEASE
Ovarian Cancer. Leukocyte extravasation is a pro-cess which shares similarities with metastatic infiltrationto secondary organs. CD157 is expressed by mesothelialcells, which share biological properties and embryologi-cal origin with ovarian surface epithelial cells (18). Aworking hypothesis was that CD157 might be expressedby epithelial ovarian cancer cells, guiding interactionsbetween tumor cells and mesothelium. If confirmed,CD157 could be involved in the control of ovarian can-cer dissemination. This hypothesis was independentlysupported by a report that BST-1/CD157 was amonggenes that were differentially expressed in primary cul-tures of epithelial ovarian cancer cells when comparedto their normal counterparts (74). The results obtainedhave confirmed the hypothesis: indeed, CD157 isexpressed by >90% of epithelial ovarian cancers and isinvolved in the interactions between epithelial ovariancancer cells, extracellular matrix proteins and mesothe-lial cells. All these steps ultimately control migration oftumor cells and invasion of surrounding tissues. Highexpression of CD157 in human ovarian cancers is associ-ated with clinical aggressiveness, confirming the role ofCD157 as an independent prognostic factor of tumorrelapse shortly after surgery (75). The functional contri-bution of CD157 to the progression of epithelial ovariancancer relies on its ability to switch on a differentiationprogram, which allows neoplastic cells to overcome therules of epithelial tissue architecture, turning it towardsa more mesenchymal state. The outcome of clinical rele-vance in clinics is that these events boost the malignantprogression of the disease (76).
Parkinson’s Disease. The BST-1/CD157 gene hasrecently been associated with Parkinson’s disease.Indeed, BST-1 SNPs rs11931532, rs12645693,rs4698412, and rs4538475 were identified as risk factorsin sporadic late-onset Parkinson’s disease in a Japanesegenome-wide association (GWA) study. rs4538475showed the strongest association (77). The associationbetween BST-1 (rs4698412 SNP) and Parkinson’s diseasewas confirmed in the European population (78), even ifnot present in a Northern Han Chinese population (79).A meta-analysis of GWA studies performed on a North-ern American and European population indicates thatBST-1 did surpass the threshold for genome-wide signifi-cance (80). A conclusion is that ethnicity significantlyinfluences the association between BST-1/CD157 locusand Parkinson’s disease.
ROLE OF THE CD38 FAMILY IN OTHER HUMAN DISEASES
Other human diseases showing correlation with CD38or CD157 are reviewed in (13). The polymorphismsassociated with genetic susceptibility to CLL have also
CD38 AND CD157 211
Cytometry Part B: Clinical Cytometry

been studied in other diseases, including systemic lupuserythematosus (SLE), where the CC genotype increasessusceptibility to and the GC genotype confers protectionfrom discoid rash development (81).
High ratios of CD8þ /CD38þ T lymphocytes predictdisease progression in HIV-infected adults (82). HIVinfection is clinically characterized by increased expres-sion of CD38, apparently not directly caused by theinfection (83). Highly active anti-retroviral therapy(HAART), the standard treatment in both pediatric andadult HIV infection, leads to viral suppression withdecreased CD38 expression, increased CD4þ T lympho-cyte counts, recovery of immune functions againstopportunistic infections and a good clinical outcome inmost patients.
Recent studies have combined traditional diagnostictests (viral load, CD4þ T lymphocyte counts, and immu-nophenotyping) with an evaluation of CD38 activation.A study involving small sample of patients has shownthat (i) CD38 expression levels in CD8þ T lymphocytesand (ii) lymphoproliferative assays in response tomycotic antigens provide additional parameters for mon-itoring patients with discordant immune-virologicalresponses to HAART (84).
CD38 expression has been also matter of technologi-cal debate, in terms of mAb used [see the discrepancyreported (3)] or protocols adopted. To this end, con-certed efforts are being made to standardize CD38 pro-tocols in order to obtain comparable evaluation of themarker molecule in HIV-infected patients. A recent pa-per provided a platform based on flow cytometry withfluorosphere MFI values, color compensation along withbiological controls which assure technologically stableanalysis. This standardized protocols have been adoptedby the laboratories of South African National HealthService for the evaluation of the CD8 þ /CD38þ ratiosfor monitoring HIV patients (85).
CD38 is also reported as target of autoantibodies intype 1 and type 2 diabetes mellitus (86,87), as well as inSLE (Pavon et al., 2013 in press).
Results from CD38 KO mice have highlighted a rolefor CD38 in the release of OT from the neurohypophysis(51). The clinical diagnosis of a deficit in short-termsocial memory in these mice has drawn the attention tohuman conditions sharing this feature, such as autismspectrum disorder (ASD). Indeed, some polymorphismsof the CD38 gene (rs6449197, rs3796863, andrs1800561) are associated with ASD (88,89). Beside this,it has recently been reported that a quantitative trait ofCD38 expression correlates with social functions in ASD(90). The study was performed by analyzing CD38expression in lymphoblastoid cell lines derived fromPBMCs of ASD patients, which has been showed to belower than CD38 expression in similar lines obtainedfrom the parents. A possible explanation is that reducedcell surface expression negatively influences the enzy-matic performances of CD38 in ASD, leading to a mal-function of the CD38/OT axis in this disorder (51,91). Afurther step of the study investigated the role of reti-
noids in upregulating cell surface CD38, potentially sug-gesting a new therapeutic approach (92,93).
EMERGING WORKING HYPOTHESIS
From a phylogenetic perspective, the human CD38
and BST-1 genes share gene structure (6). A similarintron/exon organization is also shared by their respec-tive orthologs in mouse, chicken and frog, and by theADPRCs from Aplysia, Schistosoma mansoni and thepurple sea urchin (Strongylocentrotus purpuratus)(NCBI and Ensembl genome databases). Thus theADPRCs are believed to derive from a common ancestralgene. As the common taxonomic denominator of theADPRC-bearing species is that they all belong to thebilateria (i.e., animals with front/back and left/right sym-metry), this suggests that the origin of the ADPRCs datesback to the last common bilaterian ancestor, about 555million years ago (mya).
Read from an applied perspective, the evidencederived from a phylogenetic analysis of the CD38 familyprompts the hypothesis that early CD38/CD157 precur-sors were components of innate immunity, and thattheir passage to the surface of immune cells evolved inparallel with the transition to adaptive immunity (Fig.2). We hypothesized that CD38 (CD157 having not beenconsidered in this context yet) might be part of a circuitgenerating activation or suppression of immuneresponses according to the environment. CD38 wouldalso be a component of one of the multiple strategiesadopted by tumoral cells to fool the immune system.The enzymes CD39 (ectonucleoside triphosphatediphosphohydrolase 1) and CD73 (ecto-50 nucleotidase)govern a metabolic pathway leading to the generation ofadenosine (ADO) and likely to immunosuppressionwhen ADO is taken up by specific receptors expressedby lymphocytes. This pathway is flanked by differentmechanisms guided by NADþ , which is consumed bythe CD38/CD157 ectoenzymes and—in some systems—by ART2. Endogenous signals released during physiologi-cal or pathological conditions may contribute to alarmthe innate immune system, accompanied by the produc-tion of pro-inflammatory cytokines. In addition, extracel-lular NADþ may influence the immune system byaltering the balance between activation and suppressionled by specific lymphocyte subsets in different districtsand organs.
CD38 has recently been associated with functionsexerted by regulatory T cells (Tregs) in murine models.High CD38 espression in Foxp3þ /CD4þ T cell popula-tions correlates with extremely powerful modulatoryproperties of CD4þ regulatory T lymphocytes (94). Fur-thermore, CD38 is part of the Treg transcriptional signa-ture (95,96). The role of CD38 has been confirmed inCD38 KO mice, where NADþ influences the survival,phenotype and function of Treg cells and providesproof-of-principle that acting on the ART2/P2X7 systemmay be a new strategy for manipulating these cells invivo (97). A recent report states that CD8þ /CD38high
T lymphocytes have strong immunosuppressive
212 QUARONA ET AL.
Cytometry Part B: Clinical Cytometry
capabilities in vitro and in vivo. This subset possesses aregulatory potential that could work together with theinnate immune response and control immune homeosta-sis during inflammation (98).
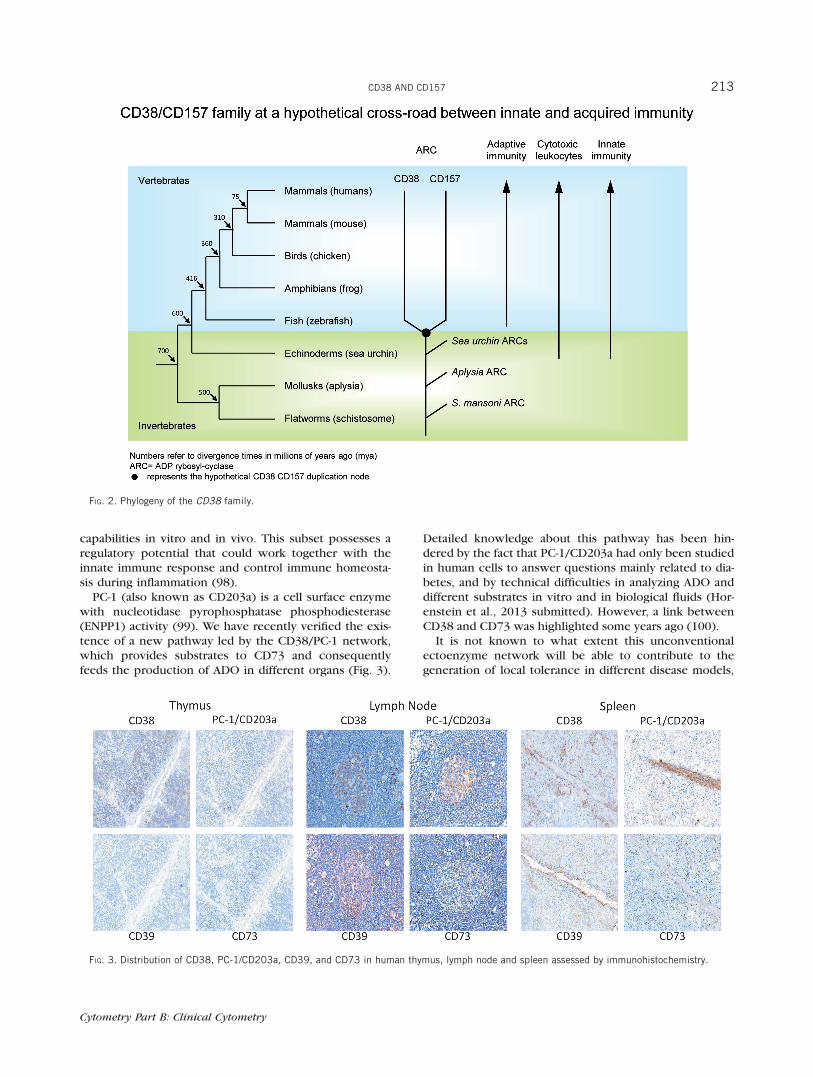
PC-1 (also known as CD203a) is a cell surface enzymewith nucleotidase pyrophosphatase phosphodiesterase(ENPP1) activity (99). We have recently verified the exis-tence of a new pathway led by the CD38/PC-1 network,which provides substrates to CD73 and consequentlyfeeds the production of ADO in different organs (Fig. 3).
Detailed knowledge about this pathway has been hin-dered by the fact that PC-1/CD203a had only been studiedin human cells to answer questions mainly related to dia-betes, and by technical difficulties in analyzing ADO anddifferent substrates in vitro and in biological fluids (Hor-enstein et al., 2013 submitted). However, a link betweenCD38 and CD73 was highlighted some years ago (100).
It is not known to what extent this unconventionalectoenzyme network will be able to contribute to thegeneration of local tolerance in different disease models,
FIG. 3. Distribution of CD38, PC-1/CD203a, CD39, and CD73 in human thymus, lymph node and spleen assessed by immunohistochemistry.
FIG. 2. Phylogeny of the CD38 family.
CD38 AND CD157 213
Cytometry Part B: Clinical Cytometry
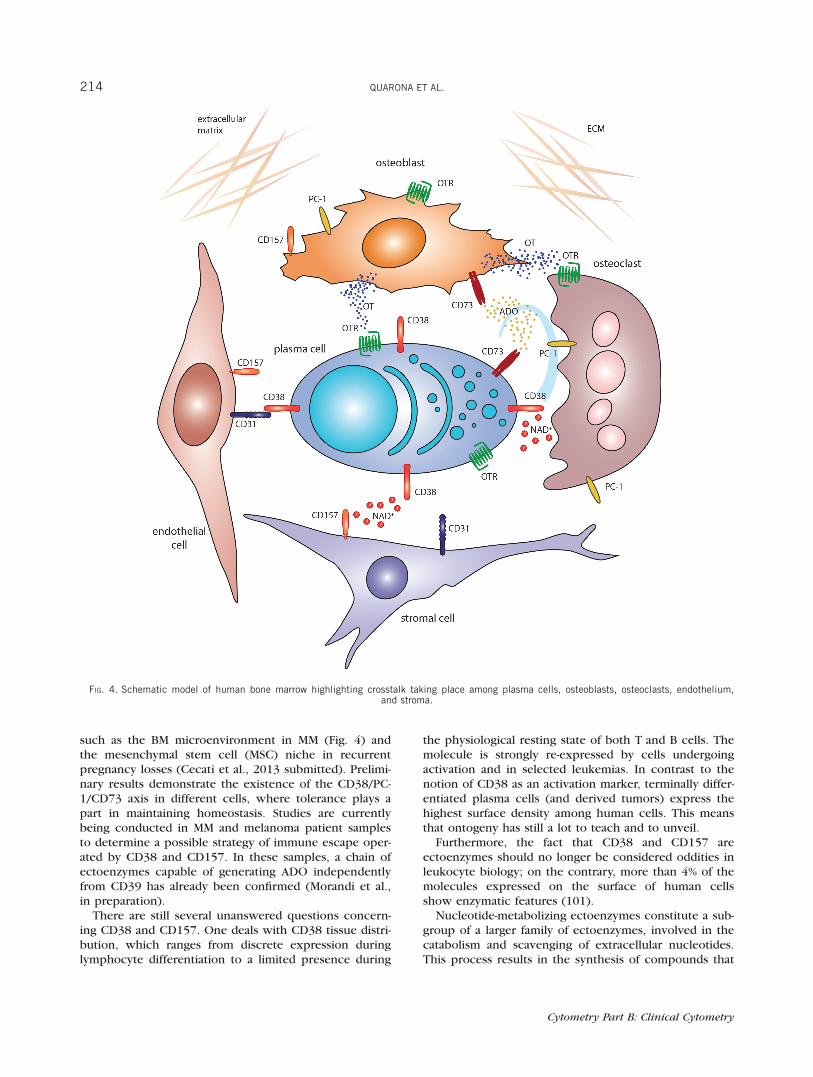
such as the BM microenvironment in MM (Fig. 4) andthe mesenchymal stem cell (MSC) niche in recurrentpregnancy losses (Cecati et al., 2013 submitted). Prelimi-nary results demonstrate the existence of the CD38/PC-1/CD73 axis in different cells, where tolerance plays apart in maintaining homeostasis. Studies are currentlybeing conducted in MM and melanoma patient samplesto determine a possible strategy of immune escape oper-ated by CD38 and CD157. In these samples, a chain ofectoenzymes capable of generating ADO independentlyfrom CD39 has already been confirmed (Morandi et al.,in preparation).
There are still several unanswered questions concern-ing CD38 and CD157. One deals with CD38 tissue distri-bution, which ranges from discrete expression duringlymphocyte differentiation to a limited presence during
the physiological resting state of both T and B cells. Themolecule is strongly re-expressed by cells undergoingactivation and in selected leukemias. In contrast to thenotion of CD38 as an activation marker, terminally differ-entiated plasma cells (and derived tumors) express thehighest surface density among human cells. This meansthat ontogeny has still a lot to teach and to unveil.
Furthermore, the fact that CD38 and CD157 areectoenzymes should no longer be considered oddities inleukocyte biology; on the contrary, more than 4% of themolecules expressed on the surface of human cellsshow enzymatic features (101).
Nucleotide-metabolizing ectoenzymes constitute a sub-group of a larger family of ectoenzymes, involved in thecatabolism and scavenging of extracellular nucleotides.This process results in the synthesis of compounds that
FIG. 4. Schematic model of human bone marrow highlighting crosstalk taking place among plasma cells, osteoblasts, osteoclasts, endothelium,and stroma.
214 QUARONA ET AL.
Cytometry Part B: Clinical Cytometry

play critical roles in cell homeostasis and metabolism,and not simply nucleotide recycling. Initially it wasthought that nucleotide-metabolizing ectoenzymeswould operate in environments containing only traceamounts of the substrate and that the final productwould be used prevalently inside the cell. This view haslater been revised, and it is now known that substratesand final products are not topologically confined to oneside or other of the plasma membrane.
The author’s laboratory tackled these issues by (i) fo-cusing exclusively on human model, (ii) using the ontog-eny and phylogeny of these molecules as a source ofphysiological clues, and (iii) trying to infer informationfrom disease, the best experiments performed by natureitself.
The aim of this paper was to review old and newfacts enriching the field related to the CD38 family prod-ucts and on the role of ectoenzymes in general. Eventhough interpretations are kept to a minimum, the per-spective of this Laboratory has inevitably pervaded thevision of the gene family.
In conclusion, CD38 and CD157 have shifted frombeing considered simple activation markers of leukocytepopulations to being recognized as molecules exertingmultiple functions in health and disease. A last dauntingtask is now to demonstrate that these surface structuresmay become appropriate targets for in vivo therapeuticinterventions (Chillemi et al., 2013 submitted) and regu-lators of the immune response (Horenstein et al., 2013submitted).
ACKNOWLEDGMENTS
Valeria Quarona and Antonella Chillemi are studentsof the PhD Program ‘‘Biomedical Sciences and Oncol-ogy’’, University of Torino. Andrea Zito provided much-appreciated technical assistance. The FondazioneRicerca Medicina Sperimentale (FIRMS) provided valua-ble assistance and support to this research project.
LITERATURE CITED
1. Reinherz EL, Kung PC, Goldstein G, Levey RH, Schlossman SF.Discrete stages of human intrathymic differentiation: Analysis ofnormal thymocytes and leukemic lymphoblasts of T-cell lineage.Proc Natl Acad Sci USA 1980;77:1588–1592.
2. Terhorst C, van Agthoven A, LeClair K, Snow P, Reinherz E,Schlossman S. Biochemical studies of the human thymocyte cell-surface antigens T6, T9 and T10. Cell 1981;23:771–780.
3. Maecker HT, McCoy JP, Nussenblatt R. Standardizing immunophe-notyping for the Human Immunology Project. Nat Rev Immunol2012;12:191–200.
4. Muraoka O, Tanaka H, Itoh M, Ishihara K, Hirano T. Genomicstructure of human BST-1. Immunol Lett 1996;54:1–4.
5. Katz F, Povey S, Parkar M, Schneider C, Sutherland R, Stanley K,Solomon E, Greaves M. Chromosome assignment of monoclonalantibody-defined determinants on human leukemic cells. Eur JImmunol 1983;13:1008–1013.
6. Ferrero E, Malavasi F. Human CD38, a leukocyte receptor andectoenzyme, is a member of a novel eukaryotic gene family of nic-otinamide adenine dinucleotideþ -converting enzymes: Extensivestructural homology with the genes for murine bone marrow stro-mal cell antigen 1 and aplysian ADP-ribosyl cyclase. J Immunol1997;159:3858–3865.
7. Ferrero E, Saccucci F, Malavasi F. The making of a leukocyte re-ceptor: Origin, genes and regulation of human CD38 and relatedmolecules. Chem Immunol 2000;75:1–19.
8. Kishimoto H, Hoshino S, Ohori M, Kontani K, Nishina H, SuzawaM, Kato S, Katada T. Molecular mechanism of human CD38 geneexpression by retinoic acid. Identification of retinoic acidresponse element in the first intron. J Biol Chem 1998;273:15429–15434.
9. Song EK, Lee YR, Kim YR, Yeom JH, Yoo CH, Kim HK, Park HM,Kang HS, Kim JS, Kim UH, et al. NAADP mediates insulin-stimu-lated glucose uptake and insulin sensitization by PPARgamma inadipocytes. Cell Rep 2012;2:1607–1619.
10. Ferrero E, Saccucci F, Malavasi F. The human CD38 gene: Poly-morphism, CpG island, and linkage to the CD157 (BST-1) gene.Immunogenetics 1999;49:597–604.
11. Saborit-Villarroya I, Vaisitti T, Rossi D, D’Arena G, Gaidano G, Mala-vasi F, Deaglio S. E2A is a transcriptional regulator of CD38expression in chronic lymphocytic leukemia. Leukemia 2011;25:479–488.
12. Ortolan E, Vacca P, Capobianco A, Armando E, Crivellin F, Horen-stein A, Malavasi F. CD157, the Janus of CD38 but with a uniquepersonality. Cell Biochem Funct 2002;20:309–322.
13. Malavasi F, Deaglio S, Funaro A, Ferrero E, Horenstein AL, OrtolanE, Vaisitti T, Aydin S. Evolution and function of the ADP ribosylcyclase/CD38 gene family in physiology and pathology. PhysiolRev 2008;88:841–886.
14. Dong C, Willerford D, Alt FW, Cooper MD. Genomic organizationand chromosomal localization of the mouse Bp3 gene, a memberof the CD38/ADP-ribosyl cyclase family. Immunogenetics 1996;45:35–43.
15. Bhan AK, Reinherz EL, Poppema S, McCluskey RT, Schlossman SF.Location of T cell and major histocompatibility complex antigensin the human thymus. J Exp Med 1980;152:771–782.
16. Hernandez-Campo PM, Almeida J, Sanchez ML, Malvezzi M,Orfao A. Normal patterns of expression of glycosylphosphatidyli-nositol-anchored proteins on different subsets of peripheralblood cells: A frame of reference for the diagnosis of paroxysmalnocturnal hemoglobinuria. Cytometry B Clin Cytom B 2006;70:71–81.
17. Ghannadan M, Baghestanian M, Wimazal F, Eisenmenger M, LatalD, Kargul G, Walchshofer S, Sillaber C, Lechner K, Valent P. Phe-notypic characterization of human skin mast cells by combinedstaining with toluidine blue and CD antibodies. J Invest Dermatol1998;111:689–695.
18. Ross JA, Ansell I, Hjelle JT, Anderson JD, Miller-Hjelle MA, DobbieJW. Phenotypic mapping of human mesothelial cells. Adv PeritDial 1998;14:25–30.
19. Shimaoka Y, Attrep JF, Hirano T, Ishihara K, Suzuki R, Toyosaki T,Ochi T, Lipsky PE. Nurse-like cells from bone marrow and syno-vium of patients with rheumatoid arthritis promote survival andenhance function of human B cells. J Clin Invest 1998;102:606–618.
20. Wimazal F, Ghannadan M, Muller MR, End A, Willheim M, Meidlin-ger P, Schernthaner GH, Jordan JH, Hagen W, Agis H, et al. Expres-sion of homing receptors and related molecules on human mastcells and basophils: A comparative analysis using multi-color flowcytometry and toluidine blue/immunofluorescence staining techni-ques. Tissue Antigens 1999;54:499–507.
21. Lavagno L, Ferrero E, Ortolan E, Malavasi F, Funaro A. CD157 ispart of a supramolecular complex with CD11b/CD18 on thehuman neutrophil cell surface. J Biol Regul Homeost Agents2007;21:5–11.
22. Yilmaz OH, Katajisto P, Lamming DW, Gultekin Y, Bauer-Rowe KE,Sengupta S, Birsoy K, Dursun A, Yilmaz VO, Selig M, et al.mTORC1 in the Paneth cell niche couples intestinal stem-cellfunction to calorie intake. Nature 2012;486:490–495.
23. Alessio M, Roggero S, Funaro A, De Monte LB, Peruzzi L, GeunaM, Malavasi F. CD38 molecule: Structural and biochemical analysison human T lymphocytes, thymocytes, and plasma cells.J Immunol 1990;145:878–884.
24. Dianzani U, Malavasi F. Lymphocyte adhesion to endothelium.Crit Rev Immunol 1995;15:167–200.
25. Newman PJ. Switched at birth: A new family for PECAM-1. J ClinInvest 1999;103:5–9.
26. Deaglio S, Dianzani U, Horenstein AL, Fernandez JE, vanKooten C, Bragardo M, Funaro A, Garbarino G, Di Virgilio F,Banchereau J, et al. Human CD38 ligand. A 120-KDA protein pre-dominantly expressed on endothelial cells. J Immunol 1996;156:727–734.
27. Horenstein AL, Stockinger H, Imhof BA, Malavasi F. CD38 bindingto human myeloid cells is mediated by mouse and human CD31.Biochem J 1998;330 (Part 3):1129–1135.
28. Deaglio S, Morra M, Mallone R, Ausiello CM, Prager E,Garbarino G, Dianzani U, Stockinger H, Malavasi F. Human CD38
CD38 AND CD157 215
Cytometry Part B: Clinical Cytometry

(ADP-ribosyl cyclase) is a counter-receptor of CD31, an Ig super-family member. J Immunol 1998;160:395–402.
29. Deaglio S, Mallone R, Baj G, Arnulfo A, Surico N, Dianzani U,Mehta K, Malavasi F. CD38/CD31, a receptor/ligand system rulingadhesion and signaling in human leukocytes. Chem Immunol2000;75:99–120.
30. Mallone R, Ferrua S, Morra M, Zocchi E, Mehta K, Notarangelo LD,Malavasi F. Characterization of a CD38-like 78-kilodalton solubleprotein released from B cell lines derived from patients with X-linked agammaglobulinemia. J Clin Invest 1998;101:2821–2830.
31. Hara-Yokoyama M, Kukimoto-Niino M, Terasawa K, Harumiya S,Podyma-Inoue KA, Hino N, Sakamoto K, Itoh S, Hashii N, Hiruta Y,et al. Tetrameric interaction of the ectoenzyme CD38 on the cellsurface enables its catalytic and raft-association activities. Structure2012;20:1585–1595.
32. Zhao YJ, Lam CM, Lee HC. The membrane-bound enzyme CD38exists in two opposing orientations. Sci Signal 2012;5:ra67.
33. Deaglio S, Vaisitti T, Billington R, Bergui L, Omede P, GenazzaniAA, Malavasi F. CD38/CD19: A lipid raft-dependent signaling com-plex in human B cells. Blood 2007;109:5390–5398.
34. Hussain AM, Lee HC, Chang CF. Functional expression of secretedmouse BST-1 in yeast. Protein Expr Purif 1998;12:133–137.
35. Okuyama Y, Ishihara K, Kimura N, Hirata Y, Sato K, Itoh M, OkLB, Hirano T. Human BST- 1 expressed on myeloid cells functionsas a receptor molecule. Biochem Biophys Res Commun 1996;228:838–845.
36. Ishihara K, Hirano T. BST- 1/CD157 regulates the humoral immuneresponses in vivo. Chem Immunol 2000;75:235–255.
37. Funaro A, Ortolan E, Ferranti B, Gargiulo L, Notaro R, Luzzatto L,Malavasi F. CD157 is an important mediator of neutrophil adhe-sion and migration. Blood 2004;104:4269–4278.
38. Ortolan E, Tibaldi EV, Ferranti B, Lavagno L, Garbarino G, Notaro R,Luzzatto L, Malavasi F, Funaro A. CD157 plays a pivotal rolein neutrophil transendothelial migration. Blood 2006;108:4214–4222.
39. Lo Buono N, Parrotta R, Morone S, Bovino P, Nacci G, Ortolan E,Horenstein AL, Inzhutova A, Ferrero E, Funaro A. The CD157-integrin partnership controls transendothelial migration and adhe-sion of human monocytes. J Biol Chem 2011;286:18681–18691.
40. States DJ, Walseth TF, Lee HC. Similarities in amino acid sequen-ces of Aplysia ADP-ribosyl cyclase and human lymphocyte antigenCD38. Trends Biochem Sci 1992;17:495.
41. Howard M, Grimaldi JC, Bazan JF, Lund FE, Santos-Argumedo L,Parkhouse RM, Walseth TF, Lee HC. Formation and hydrolysis ofcyclic ADP-ribose catalyzed by lymphocyte antigen CD38. Science1993;262:1056–1059.
42. Kim H, Jacobson EL, Jacobson MK. Synthesis and degradation ofcyclic ADP-ribose by NAD glycohydrolases. Science 1993;261:1330–1333.
43. Zocchi E, Franco L, Guida L, Calder L, De Flora A. Self-aggregationof purified and membrane-bound erythrocyte CD38 induces exten-sive decrease of its ADP-ribosyl cyclase activity. FEBS Lett 1995;359:35–40.
44. Takasawa S, Tohgo A, Noguchi N, Koguma T, Nata K, Sugimoto T,Yonekura H, Okamoto H. Synthesis and hydrolysis of cyclic ADP-ribose by human leukocyte antigen CD38 and inhibition of the hy-drolysis by ATP. J Biol Chem 1993;268:26052–26054.
45. Aarhus R, Graeff RM, Dickey DM, Walseth TF, Lee HC. ADP-ribo-syl cyclase and CD38 catalyze the synthesis of a calcium-mobiliz-ing metabolite from NADP. J Biol Chem 1995;270:30327–30333.
46. Guse AH, da Silva CP, Berg I, Skapenko AL, Weber K, Heyer P,Hohenegger M, Ashamu GA, Schulze-Koops H, Potter BV, et al.Regulation of calcium signalling in T lymphocytes by the secondmessenger cyclic ADP-ribose. Nature 1999;398:70–73.
47. Morra M, Zubiaur M, Terhorst C, Sancho J, Malavasi F. CD38 isfunctionally dependent on the TCR/CD3 complex in human Tcells. FASEB J 1998;12:581–592.
48. Meszaros LG, Bak J, Chu A. Cyclic ADP-ribose as an endogenousregulator of the non-skeletal type ryanodine receptorCa2þ channel. Nature 1993;364:76–79.
49. Kuemmerle JF, Makhlouf GM. Agonist-stimulated cyclic ADP ribose.Endogenous modulator of Ca(2þ )-induced Ca2þ release in intesti-nal longitudinal muscle. J Biol Chem 1995;270:25488–25494.
50. Takasawa S, Nata K, Yonekura H, Okamoto H. Cyclic ADP-ribosein insulin secretion from pancreatic beta cells. Science 1993;259:370–373.
51. Jin D, Liu HX, Hirai H, Torashima T, Nagai T, Lopatina O, ShnayderNA, Yamada K, Noda M, Seike T, et al. CD38 is critical for socialbehaviour by regulating oxytocin secretion. Nature 2007;446:41–45.
52. Grimaldi JC, Balasubramanian S, Kabra NH, Shanafelt A, Bazan JF,Zurawski G, Howard MC. CD38-mediated ribosylation of proteins.J Immunol 1995;155:811–817.
53. Han MK, Cho YS, Kim YS, Yim CY, Kim UH. Interaction of twoclasses of ADP-ribose transfer reactions in immune signaling.J Biol Chem 2000;275:20799–20805.
54. Malavasi F, Deaglio S, Zaccarello G, Horenstein AL, Chillemi A,Audrito V, Serra S, Gandione M, Zitella A, Tizzani A. The hiddenlife of NADþ -consuming ectoenzymes in the endocrine system.J Mol Endocrinol 2010;45:183–191.
55. Audrito V, Vaisitti T, Rossi D, Gottardi D, D’Arena G, Laurenti L,Gaidano G, Malavasi F, Deaglio S. Nicotinamide blocks prolifera-tion and induces apoptosis of chronic lymphocytic leukemia cellsthrough activation of the p53/miR-34a/SIRT1 tumor suppressornetwork. Cancer Res 2011;71:4473–4483.
56. Haag F, Adriouch S, Brass A, Jung C, Moller S, Scheuplein F,Bannas P, Seman M, Koch-Nolte F. Extracellular NAD and ATP:Partners in immune cell modulation. Purinergic Signal 2007;3:71–81.
57. Moreschi I, Bruzzone S, Bodrato N, Usai C, Guida L, Nicholas RA,Kassack MU, Zocchi E, De Flora A. NAADPþ is an agonist of thehuman P2Y11 purinergic receptor. Cell Calcium 2008;43:344–355.
58. Funaro A, Horenstein AL, Calosso L, Morra M, Tarocco RP, FrancoL, De Flora A, Malavasi F. Identification and characterization of anactive soluble form of human CD38 in normal and pathologicalfluids. Int Immunol 1996;8:1643–1650.
59. Zumaquero E, Munoz P, Cobo M, Lucena G, Pavon EJ, Martin A,Navarro P, Garcia-Perez A, Ariza-Veguillas A, Malavasi F, et al.Exosomes from human lymphoblastoid B cells express enzymati-cally active CD38 that is associated with signaling complexescontaining CD81, Hsc-70 and Lyn. Exp Cell Res 2010;316:2692–2706.
60. Chiorazzi N, Ferrarini M. Cellular origin(s) of chronic lymphocyticleukemia: Cautionary notes and additional considerations and pos-sibilities. Blood 2011;117:1781–1791.
61. Chiorazzi N. Implications of new prognostic markers in chroniclymphocytic leukemia. Hematol Am Soc Hematol Educ Program2012;2012:76–87.
62. Malavasi F, Deaglio S, Damle R, Cutrona G, Ferrarini M, ChiorazziN. CD38 and chronic lymphocytic leukemia: A decade later.Blood 2011;118:3470–3478.
63. Deaglio S, Capobianco A, Bergui L, Durig J, Morabito F, DuhrsenU, Malavasi F. CD38 is a signaling molecule in B-cell chronic lym-phocytic leukemia cells. Blood 2003;102:2146–2155.
64. Vaisitti T, Aydin S, Rossi D, Cottino F, Bergui L, D’Arena G, BonelloL, Horenstein AL, Brennan P, Pepper C, et al. CD38 increasesCXCL12-mediated signals and homing of chronic lymphocytic leu-kemia cells. Leukemia 2010;24:958–969.
65. Zucchetto A, Benedetti D, Tripodo C, Bomben R, Dal Bo M,Marconi D, Bossi F, Lorenzon D, Degan M, Rossi FM, et al. CD38/CD31, the CCL3 and CCL4 chemokines, and CD49d/vascular celladhesion molecule-1 are interchained by sequential events sustain-ing chronic lymphocytic leukemia cell survival. Cancer Res 2009;69:4001–4009.
66. Zucchetto A, Vaisitti T, Benedetti D, Tissino E, Bertagnolo V, RossiD, Bomben R, Dal Bo M, Del Principe MI, Gorgone A, et al. TheCD49d/CD29 complex is physically and functionally associatedwith CD38 in B-cell chronic lymphocytic leukemia cells. Leukemia2012;26:1301–1312.
67. Colaianni G, Di Benedetto A, Zhu LL, Tamma R, Li J, Greco G,Peng Y, Dell’Endice S, Zhu G, Cuscito C, et al. Regulated produc-tion of the pituitary hormone oxytocin from murine and humanosteoblasts. Biochem Biophys Res Commun 2011;411:512–515.
68. Iqbal J, Zaidi M. Extracellular NADþmetabolism modulates osteo-clastogenesis. Biochem Biophys Res Commun 2006;349:533–539.
69. Colaianni G, Sun L, Di Benedetto A, Tamma R, Zhu LL, Cao J,Grano M, Yuen T, Colucci S, Cuscito C, et al. Bone marrow oxyto-cin mediates the anabolic action of estrogen on the skeleton.J Biol Chem 2012;287:29159–29167.
70. Krause DS, Scadden DT, Preffer FI. The hematopoietic stem cellniche—Home for friend and foe? Cytometry B Clin Cytom 2013;84B:7–20.
71. Gao Y, Camacho LH, Mehta K. Retinoic acid-induced CD38 anti-gen promotes leukemia cells attachment and interferon-gamma/interleukin-1beta-dependent apoptosis of endothelial cells: Implica-tions in the etiology of retinoic acid syndrome. Leuk Res 2007;31:455–463.
72. Drach J, McQueen T, Engel H, Andreeff M, Robertson KA, CollinsSJ, Malavasi F, Mehta K. Retinoic acid-induced expression of
216 QUARONA ET AL.
Cytometry Part B: Clinical Cytometry

CD38 antigen in myeloid cells is mediated through retinoic acidreceptor-alpha. Cancer Res 1994;54:1746–1752.
73. Mehta K, Shahid U, Malavasi F. Human CD38, a cell-surface pro-tein with multiple functions. FASEB J 1996;10:1408–1417.
74. Le Page C, Sanceau J, Drapier JC, Wietzerbin J. Inhibitors of ADP-ribosylation impair inducible nitric oxide synthase gene transcrip-tion through inhibition of NF kappa B activation. Biochem Bio-phys Res Commun 1998;243:451–457.
75. Ortolan E, Arisio R, Morone S, Bovino P, Lo-Buono N, Nacci G, Par-rotta R, Katsaros D, Rapa I, Migliaretti G, et al. Functional role andprognostic significance of CD157 in ovarian carcinoma. J NatlCancer Inst 2010;102:1160–1177.
76. Morone S, Lo-Buono N, Parrotta R, Giacomino A, Nacci G, BruscoA, Larionov A, Ostano P, Mello-Grand M, Chiorino G, et al. Overex-pression of CD157 contributes to epithelial ovarian cancer pro-gression by promoting mesenchymal differentiation. PLoS One2012;7:e43649.
77. Satake W, Nakabayashi Y, Mizuta I, Hirota Y, Ito C, Kubo M,Kawaguchi T, Tsunoda T, Watanabe M, Takeda A, et al. Genome-wide association study identifies common variants at four loci asgenetic risk factors for Parkinson’s disease. Nat Genet 2009;41:1303–1307.
78. Saad M, Lesage S, Saint-Pierre A, Corvol JC, Zelenika D, LambertJC, Vidailhet M, Mellick GD, Lohmann E, Durif F, et al. Genome-wide association study confirms BST1 and suggests a locus on12q24 as the risk loci for Parkinson’s disease in the European pop-ulation. Hum Mol Genet 2011;20:615–627.
79. Zhu LH, Luo XG, Zhou YS, Li FR, Yang YC, Ren Y, Pang H. Lackof association between three single nucleotide polymorphisms inthe PARK9, PARK15, and BST1 genes and Parkinson’s disease inthe northern Han Chinese population. Chin Med J (Engl) 2012;125:588–592.
80. Nalls MA, Plagnol V, Hernandez DG, Sharma M, Sheerin UM, SaadM, Simon-Sanchez J, Schulte C, Lesage S, Sveinbjornsdottir S, et al.Imputation of sequence variants for identification of genetic risksfor Parkinson’s disease: A meta-analysis of genome-wide associa-tion studies. Lancet 2011;377:641–649.
81. Gonzalez-Escribano MF, Aguilar F, Torres B, Sanchez-Roman J,Nunez-Roldan A. CD38 polymorphisms in Spanish patients withsystemic lupus erythematosus. Hum Immunol 2004;65:660–664.
82. Giorgi JV, Ho HN, Hirji K, Chou CC, Hultin LE, O’Rourke S, ParkL, Margolick JB, Ferbas J, Phair JP. CD8þ lymphocyte activation athuman immunodeficiency virus type 1 seroconversion: Develop-ment of HLA-DRþCD38- CD8þ cells is associated with subse-quent stable CD4þ cell levels. The Multicenter AIDS Cohort StudyGroup. J Infect Dis 1994;170:775–781.
83. Savarino A, Bottarel F, Malavasi F, Dianzani U. Role of CD38 inHIV-1 infection: An epiphenomenon of T-cell activation or anactive player in virus/host interactions? Aids 2000;14:1079–1089.
84. Rosso R, Fenoglio D, Terranova MP, Lantieri F, Risso D, Pontali E,Setti M, Cossarizza A, Ravetti JL, Viscoli C, et al. Relevance ofCD38 expression on CD8 T cells to evaluate antiretroviral therapyresponse in HIV-1-infected youths. Scand J Immunol 2010;71:45–51.
85. Coetzee LM, Tay SS, Lawrie D, Janossy G, Glencross DK. Fromresearch tool to routine test: CD38 monitoring in HIV patients.Cytometry B Clin Cytom 2009;76B:375–384.
86. Mallone R, Ortolan E, Pinach S, Volante M, Zanone MM, Bruno G,Baj G, Lohmann T, Cavallo-Perin P, Malavasi F. Anti-CD38 autoanti-
bodies: Characterisation in new-onset type I diabetes and latentautoimmune diabetes of the adult (LADA) and comparison withother islet autoantibodies. Diabetologia 2002;45:1667–1677.
87. Mallone R, Ortolan E, Baj G, Funaro A, Giunti S, Lillaz E, SaccucciF, Cassader M, Cavallo-Perin P, Malavasi F. Autoantibody responseto CD38 in Caucasian patients with type 1 and type 2 diabetes:Immunological and genetic characterization. Diabetes 2001;50:752–762.
88. Lerer E, Levi S, Israel S, Yaari M, Nemanov L, Mankuta D, Nurit Y,Ebstein RP. Low CD38 expression in lymphoblastoid cells andhaplotypes are both associated with autism in a family-basedstudy. Autism Res 2010;3:293–302.
89. Munesue T, Yokoyama S, Nakamura K, Anitha A, Yamada K, Haya-shi K, Asaka T, Liu HX, Jin D, Koizumi K, et al. Two genetic var-iants of CD38 in subjects with autism spectrum disorder andcontrols. Neurosci Res 2010;67:181–191.
90. Riebold M, Mankuta D, Lerer E, Israel S, Zhong S, Nemanov L,Monakhov MV, Levi S, Yirmiya N, Yaari M, et al. All-trans retinoicacid upregulates reduced CD38 transcription in lymphoblastoidcell lines from Autism spectrum disorder. Mol Med 2011;17:799–806.
91. Salmina AB, Lopatina O, Ekimova MV, Mikhutkina SV, Higashida H.CD38/cyclic ADP-ribose system: A new player for oxytocin secre-tion and regulation of social behaviour. J Neuroendocrinol 2010;22:380–392.
92. Malavasi F. Editorial: CD38 and retinoids: A step toward a cure.J Leukoc Biol 2011;90:217–219.
93. Ebstein RP, Mankuta D, Yirmiya N, Malavasi F. Are retinoids poten-tial therapeutic agents in disorders of social cognition includingautism? FEBS Lett 2011;585:1529–1536.
94. Patton DT, Wilson MD, Rowan WC, Soond DR, Okkenhaug K.The PI3K p110delta regulates expression of CD38 on regulatory Tcells. PLoS One 2011;6:e17359.
95. Blair PA, Norena LY, Flores-Borja F, Rawlings DJ, Isenberg DA,Ehrenstein MR, Mauri C. CD19(þ)CD24(hi)CD38(hi) B cells ex-hibit regulatory capacity in healthy individuals but are functionallyimpaired in systemic Lupus Erythematosus patients. Immunity2010;32:129–140.
96. Yu A, Zhu L, Altman NH, Malek TR. A low interleukin-2 receptorsignaling threshold supports the development and homeostasis ofT regulatory cells. Immunity 2009;30:204–217.
97. Hubert S, Rissiek B, Klages K, Huehn J, Sparwasser T, Haag F,Koch-Nolte F, Boyer O, Seman M, Adriouch S. Extracellular NADþshapes the Foxp3þ regulatory T cell compartment through theART2-P2X7 pathway. J Exp Med 2010;207:2561–2568.
98. Bahri R, Bollinger A, Bollinger T, Orinska Z, Bulfone-Paus S. Ecto-nucleotidase CD38 demarcates regulatory, memory-like CD8(þ) Tcells with IFN-gamma-mediated suppressor activities. PLoS One2012;7:e45234.
99. Goding JW, Terkeltaub R, Maurice M, Deterre P, Sali A, Belli SI.Ecto-phosphodiesterase/pyrophosphatase of lymphocytes andnon-lymphoid cells: Structure and function of the PC-1 family.Immunol Rev 1998;161:11–26.
100. Massaia M, Perrin L, Bianchi A, Ruedi J, Attisano C, Altieri D,Rijkers GT, Thompson LF. Human T cell activation. Synergy be-tween CD73 (ecto-5’-nucleotidase) and signals delivered throughCD3 and CD2 molecules. J Immunol 1990;145:1664–1674.
101. Goding JW, Howard MC. Ecto-enzymes of lymphoid cells. Immu-nol Rev 1998;161:5–10.
Cytometry Part B: Clinical Cytometry
CD38 AND CD157 217
Related Documents











