of April 21, 2016. This information is current as Formation T Cell Motility and Immunological Synapse CD80 Interactions Control Regulatory - CD28 Dustin and David C. Parker Timothy J. Thauland, Yoshinobu Koguchi, Michael L. http://www.jimmunol.org/content/193/12/5894 doi: 10.4049/jimmunol.1401752 October 2014; 2014; 193:5894-5903; Prepublished online 29 J Immunol Material Supplementary 2.DCSupplemental.html http://www.jimmunol.org/content/suppl/2014/10/29/jimmunol.140175 References http://www.jimmunol.org/content/193/12/5894.full#ref-list-1 , 22 of which you can access for free at: cites 54 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved. Copyright © 2014 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on April 21, 2016 http://www.jimmunol.org/ Downloaded from by guest on April 21, 2016 http://www.jimmunol.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

of April 21, 2016.This information is current as
FormationT Cell Motility and Immunological Synapse
CD80 Interactions Control Regulatory−CD28
Dustin and David C. ParkerTimothy J. Thauland, Yoshinobu Koguchi, Michael L.
http://www.jimmunol.org/content/193/12/5894doi: 10.4049/jimmunol.1401752October 2014;
2014; 193:5894-5903; Prepublished online 29J Immunol
MaterialSupplementary
2.DCSupplemental.htmlhttp://www.jimmunol.org/content/suppl/2014/10/29/jimmunol.140175
Referenceshttp://www.jimmunol.org/content/193/12/5894.full#ref-list-1
, 22 of which you can access for free at: cites 54 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2014 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on April 21, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on April 21, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
CD28–CD80 Interactions Control Regulatory T Cell Motilityand Immunological Synapse Formation
Timothy J. Thauland,* Yoshinobu Koguchi,*,1 Michael L. Dustin,†,‡ and David C. Parker*
Regulatory T cells (Tregs) are essential for tolerance to self and environmental Ags, acting in part by downmodulating costimulatory
molecules on the surface of dendritic cells (DCs) and altering naive CD4 T cell–DC interactions. In this study, we show that Tregs
form stable conjugates with DCs before, but not after, they decrease surface expression of the costimulatory molecule CD80 on the
DCs. We use supported planar bilayers to show that Tregs dramatically slow down but maintain a highly polarized and motile
phenotype after recognizing Ag in the absence of costimulation. These motile cells are characterized by distinct accumulations of
LFA-1–ICAM-1 in the lamella and TCR-MHC in the uropod, consistent with a motile immunological synapse or “kinapse.”
However, in the presence of high, but not low, concentrations of CD80, Tregs form stationary, symmetrical synapses. Using
blocking Abs, we show that, whereas CTLA-4 is required for CD80 downmodulation, CD28–CD80 interactions are critical for
modulating Treg motility in the presence of Ag. Taken together, these results support the hypothesis that Tregs are tuned to alter
their motility depending on costimulatory signals. The Journal of Immunology, 2014, 193: 5894–5903.
Foxp3+ CD4 regulatory T cells (Tregs) are critical forcontrolling immune responses and promoting tolerance toautoantigens and commensal gut microflora (1, 2). The
transcription factor Foxp3 is necessary for Treg development andfunction (3–5), and it controls the expression of hundreds of genes,several of which, including Ctla4, are crucial for Treg developmentor function (2). Indeed, CTLA-4 is required for proper Treg func-tioning, as specific ablation in Foxp3+ cells results in severe auto-immunity (6). The costimulatory molecule CD28 is required forTreg development and survival (7, 8). In addition to thymus-derivedor natural Tregs (nTregs), Foxp3+ Tregs can be induced from pe-ripheral CD4 T cells both in vitro and in vivo (9).Tregs use a variety of processes, involving both contact-
dependent and cytokine-mediated mechanisms, to suppress im-mune reactions in vitro and in vivo (10, 11). Several reports have
shown that Tregs can downmodulate expression of CD80 andCD86 on dendritic cells (DCs) (6, 12–15), a process that involvestransendocytosis (16) and depends on PKC-h (17). Down-modulation of CD80/86 on DCs requires expression of CTLA-4by Tregs, and occurs in an Ag- and LFA-1–ICAM-1-dependentmanner (14, 15). CD80 downmodulation is observed even whennTregs and DCs are cocultured in the presence of LPS, IFN-g, orother potent maturing stimuli, indicating that CTLA-4–mediatedregulation of DCs by nTregs is a powerful mechanism for alteringDC function (15). In vivo imaging experiments have demonstratedthat in the presence of Ag, Tregs prevent sustained naive T cell–DC interactions (18, 19) and induce dysfunction in CTLs in tumortissue (20). Tregs were not observed directly interacting withnaive T cells or CTLs in these experiments, and instead formedAg-dependent, unstable tethering interactions with DCs coincid-ing with marked downregulation of CD80 and CD86 (20). Inexperiments with TCR transgenic Tregs and naive CD4 T cellscocultured with peptide-pulsed splenic DCs, Tregs were highlymotile, swarmed around Ag-loaded DCs, and outcompeted thenaive cells for space around the DCs (15). This swarming phe-notype, possibly in conjunction with CTLA-4–mediated down-modulation of CD80/86, could explain the in vivo imaging resultsdescribed above. Owing to the importance of Tregs in modulatingthe surface phenotype of DCs and the sparseness of studies ex-amining the immunological synapse (IS) formed by Tregs, thefactors governing Treg–DC interactions are of interest.In this study, we tested the hypothesis that costimulatory mol-
ecules on the surface of APCs are responsible for determining Tregmotility and IS structure. Given that Tregs are capable of down-modulating CD80 molecules on the surface of DCs, and thatCD28–CD80 interactions can modulate synapse structure (21), weexamined the role that CD80 plays in Treg–DC IS formation. Weshow that in vitro–generated Tregs, similar to nTregs, are capableof downmodulating CD80 on the surface of DCs, and that alter-ation of the DC surface phenotype affects subsequent Treg–DCinteractions. Using supported planar bilayers as artificial APCs,we demonstrate that Tregs slow down, but are still highly motile,when they recognize Ag in the absence of costimulation. How-ever, the presence of high, but not low, levels of CD80 is sufficientto induce the formation of symmetrical, nonmotile IS by Tregs.
*Department of Molecular Microbiology and Immunology, Oregon Health & ScienceUniversity, Portland, OR 97239; †Department of Pathology, New York UniversitySchool of Medicine, New York, NY 10016; and ‡Kennedy Institute of Rheumatology,Nuffield Department of Orthopaedics, Rheumatology, and Musculoskeletal Sciences,University of Oxford, Oxford OX3 7FY, United Kingdom
1Current address: Earle A. Chiles Research Institute, Robert W. Franz CancerResearch Center, Providence Cancer Center, Portland, OR.
ORCIDs: 0000-0003-4983-6389 (M.L.D.); 0000-0003-4456-2788 (D.C.P.).
Received for publication July 10, 2014. Accepted for publication October 8, 2014.
This work was supported by National Institutes of Health Grants R01 AI050823 (toD.C.P.), R01 AI092080 (to D.C.P.), and R01 AI043542 (to M.L.D.) and by a Well-come Trust Principal Research Fellowship (to M.L.D.). T.J.T. was supported byNational Institute for Allergy and Infectious Diseases Grants T32 AI007472 andT32 AI078903.
Address correspondence and reprint requests to Dr. David C. Parker, Department ofMolecular Microbiology and Immunology, L220, Oregon Health & Science Univer-sity, 3181 SW Sam Jackson Park Road, Portland, OR 97239 or Dr. Timothy J.Thauland at the current address: Division of Immunology, Allergy, and Rheumatol-ogy, Department of Pediatrics, Stanford University, 300 Pasteur Drive, Grant Build-ing Room S374, Stanford, CA 94305. E-mail addresses: [email protected] (D.C.P.)or [email protected] (T.J.T.)
The online version of this article contains supplemental material.
Abbreviations used in this article: BMDC, bone marrow–derived DC; DC, dendriticcell; DIC, differential interference contrast; IS, immunological synapse; MCC, mothcytochrome c peptide; MSD, mean square displacement; nTreg, natural regulatoryT cell; pMHC, peptide-MHC; Treg, regulatory T cell.
Copyright� 2014 by TheAmericanAssociation of Immunologists, Inc. 0022-1767/14/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1401752
by guest on April 21, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

We suggest a model wherein highly motile Tregs are constantlyscanning the surface of APCs, only stopping when the APC isactivated and displaying high levels of costimulatory activity. Thisbehavior may allow Tregs specific for self and environmental Agsto efficiently downmodulate costimulatory molecules on inap-propriately activated cells while ignoring resting cells displayingthose Ags.
Materials and MethodsAnimals
Heterozygous AD10 TCR transgenic mice on a B10.BR background,specific for pigeon cytochrome c 88–104 and reactive against moth cyto-chrome c (MCC) 88–103 (22), were provided by S. Hedrick (University ofCalifornia at San Diego, La Jolla, CA) by way of P. Marrack (NationalJewish Medical Center, Denver, CO). B6.Cg-Foxp3tm2Tch/J (Foxp3-GFP)mice were obtained from The Jackson Laboratory. Male AD10+ mice werebred to homozygous female Foxp3-GFP mice and the AD10+ progenywere used as a source of TCR transgenic GFP+ nTregs. All mice werehoused in specific pathogen-free conditions and used in accordance withNational Institutes of Health guidelines under an animal protocol approvedby the Oregon Health & Science University Institutional Animal Care andUse Committee.
Abs
The Abs used for flow cytometry were as follows: anti-CD4Alexa Fluor 488(GK1.5; eBioscience), anti-CD4 PerCP (RM4-5; BioLegend), anti-CD152PE (UC10-4B9; BioLegend), anti-Foxp3 Alexa Fluor 647 (150D; Bio-Legend), anti-CD11c FITC and PE (HL3; BD Biosciences), anti-CD25PerCP (PC61; BioLegend), and anti-CD80 FITC and PerCP-Cy5.5 (16-10A1; BD Biosciences).
DC culture
Bone marrow–derived DCs (BMDCs) were cultured as described (23).Briefly, bone marrow cells were cultured for 9 d in bacteriological Petridishes in complete RPMI 1640 media supplemented with GM-CSF su-pernatant (final concentration, 20 ng/ml). On day 9 of culture, immatureDCs were plated in LabTek II eight-well chambers (no. 1.5; Nunc) or six-well plates in fresh media with 20 ng/ml GM-CSF and 1 mg/ml LPS.
In vitro Treg polarization
CD4+ cells and B cells were purified from AD10 and B10.BR spleen cellsuspensions, respectively, using EasySep immunomagnetic negative se-lection (StemCell Technologies). B cells (5 3 106) and CD4+ cells (2.5 3106) were cultured in 1 ml complete RPMI 1640 media in six-well plateswith 2.5 mM MCC peptide, 20 ng/ml TGF-b, 100 U/ml IL-2, and 10 nMall-trans retinoic acid. On days 2 and 3, 1 ml media supplemented with100 U/ml IL-2 was added to the cultures. To confirm that T cells werepolarized to an Treg phenotype, cells were fixed, permeabilized, andstained for CD4, CTLA-4, and Foxp3. Fixation and permeabilizationreagents were from BioLegend. Cells were used on day 4.
CD80 downmodulation assay
Day 9 BMDCs (1.6 3 105) were seeded onto 12-well plates and treatedLPS as described above. One day later, the indicated numbers of day 4Tregs were added per well in the presence or absence of 2.5 mm MCC. Insome experiments, Fab fragments of anti-CD28 (E18; gift from ThomasH€unig) (24) or anti–CTLA-4 (UC10-4F10-11; Bio X Cell) were added at100 mg/ml. The Fab fragments were generated with a Pierce Fab prepa-ration kit (Thermo Scientific). As a control, Tregs were not added to somewells. Twenty-four hours later, the cells were harvested and stained for30 min on ice for CD4, CD11c, and CD80. All Abs were used at 1:200.EDTA (1 mM) was included in the FACS buffer to discourage continuedinteractions. Samples were collected on a BD FACSCalibur with CellQuestsoftware and analyzed with FlowJo (Tree Star).
Imaging Treg–DC interactions
Day 9 BMDCs (3 3 104) were seeded onto coverslips in eight-wellchambers and treated with LPS as described above. One day later, 1.5 3105 CFSE-loaded, day 4 Tregs were added to the wells. Imaging com-menced as soon as Tregs were added to the wells. Differential interferencecontrast (DIC) and fluorescent images were obtained every minute for 2 h.All imaging was conducted at 37˚C with 5% CO2. Wide-field imaging wasperformed with an Applied Precision DeltaVision system using an
Olympus 320/0.75 numerical aperture Plan Apo objective. This systemincluded an Applied Precision chassis with a motorized XYZ stage,WeatherStation environmental chamber, Olympus IX71 inverted fluores-cent microscope, xenon lamp, and CoolSnap HQ2 camera. The DeltaVisionSoftWorx software package was used for image acquisition.
Supported planar bilayer experiments
GPI-linked forms of Oregon Green 488 labeled I-Ek (200 molecules/mm2)and Cy5-labeled ICAM-1 (300 molecules/mm2) were incorporated intodioleoylphosphatidylcholine bilayers exactly as described (25, 26). Forsome experiments, GPI-linked CD80 was incorporated into bilayers at 20,40, or 200 mm2 (27). These bilayers were supported on a coverslip ina Bioptechs flow cell and were loaded with 100 mM MCC or hemoglobinpeptide (GKKVITAFNEGLK) in a PBS/citrate buffer (pH 4.5) for 24 h at37˚C (25). T cells (107) in 1 ml HBS buffer with 1% BSA were injectedonto bilayers at 37˚C. In some experiments, anti-CD28 and/or anti–CTLA-4 Fab fragments were added to the T cells at 50 or 100 mg/ml 10 min priorto injection onto the bilayers. Images were acquired every minute for 30–60 min on the DeltaVision system using an Olympus 360/1.42 numericalaperture Plan Apo objective.
Imaris 6.3 (Bitplane) was used to track cells interacting with the bilayer.To ensure that only cells productively interacting with the bilayer wereanalyzed, Cy5 fluorescence (ICAM-1 accumulation) was tracked auto-matically with the Spots tool in Surpass mode. In situations where theICAM-1 signal was ambiguous, DIC images were examined to determinewhether the cell was flattened against the bilayer. DIC images were used totrack cells interacting with bilayers loaded with irrelevant peptide, as therewas only sparse ICAM-1 accumulation under these conditions.
Scoring T cell–bilayer interactions
Cells were scored as nonmotile when they moved one cell diameter or lessduring the entire imaging session. Cells were considered to have reformedan IS when they stopped forward progress, lost their uropod and ICAM-1arc, and remained nonmotile for at least 10 min. Cells were considered tohave broken symmetry when theywere nonmotile for 10 min or more beforegaining motility and moving more than one cell diameter. The vast majorityof cells were symmetrical when first contacting the bilayer, but this statewastransient (much less than 10 min) and cells that subsequently became motilewere not considered to have broken symmetry. Cells were scored as havinga uropod anchor when the uropod of a motile cell was attached to the sameplace for at least 10 min, causing the cell to pivot.
ResultsTregs transition from stable to motile contacts with DCs,concomitant with downmodulation of costimulatory molecules
We set out to study the IS formed between Tregs and APCs. Togenerate a population of Ag-specific cells for use in imagingexperiments, Tregs were induced in vitro by culturing purified CD4T cells from AD10 TCR transgenic mice with naive B cells in thepresence of MCC peptide, TGF-b, IL-2, and all-trans retinoic acid,as previously described (28). After 4 d of culture under Tregconditions, the cells were almost completely polarized to a Tregphenotype with high levels of Foxp3 and CTLA-4 (Fig. 1A).Day 4 Tregs were loaded with CFSE, introduced to peptide-loadedBMDCs, and imaged over time.The initial interactions between Tregs and BMDCs were very
stable, with the vast majority of Tregs flattening against BMDCsand maintaining a rounded, unpolarized shape for the duration ofthe experiment (Fig. 1B, Supplemental Video 1). However, after24 h of coculture, the Tregs became highly motile and formedlarge, swarming clusters around the BMDCs (Fig. 1C). To furtherinvestigate this change in motility, we added fresh Tregs labeledwith CFSE to the Treg-BMDC cocultures and observed theirbehavior. Surprisingly, the freshly added Tregs also displayeda motile, swarming phenotype. As shown in Fig. 1D andSupplemental Video 2, the freshly added Tregs adopted a polar-ized shape and crawled along the surface of the BMDCs. Thetendency of freshly added cells to form motile, rather than stable,contacts indicates a change in the BMDCs rather than a change inthe Tregs following 24 h of culture together.
The Journal of Immunology 5895
by guest on April 21, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

nTregs have been shown to cluster around DCs after a 12-hincubation, and this phenotype was associated with down-modulation of CD80 and CD86 on the DC surface (15). To de-termine whether changes in the surface phenotype of the DCscould be responsible for the altered motility of Tregs after a 24-hcoculture, we conducted assays to measure downmodulation ofCD80. Incubating Tregs with BMDCs resulted in Ag-specificdownmodulation of CD80 at both 1:1 and 5:1 Treg/BMDC ratios(Fig. 1E, 1F). In agreement with published work (15), Fab frag-ments of an anti–CTLA-4 Ab completely inhibited CD80 down-modulation, even at a 10:1 Treg/BMDC ratio (Fig. 1G). Treatmentwith Fab fragments of an anti-CD28 Ab known to efficiently blockCD28–CD80 interactions (24) resulted in a decrease in the effi-ciency of CD80 downmodulation, but the effect was relativelyminor compared with anti–CTLA-4 treatment (Fig. 1H).
CD80 modulates IS formation in Tregs
Although qualitatively informative, the Treg–DC interactions werenot conducive to precise measurements of Treg motility because
the DCs in these experiments were often highly motile. To care-fully measure the interactions between Tregs and APCs in twodimensions, we used supported planar bilayers containing GPI-linked, fluorescently labeled ICAM-1 and MHC class II (I-Ek)loaded with MCC peptide (25).Day 4 Tregs were injected onto the supported planar bilayers and
examined for the distribution of peptide-MHC (pMHC)–TCR andICAM-1–LFA-1 interactions at the Treg–APC interface. Althoughthe cells fluxed calcium immediately upon touching the bilayers(data not shown), we did not observe symmetrical IS with well-defined supramolecular activation clusters, in agreement with arecent report (Fig. 2A) (29). Instead, the Tregs interacting withthe bilayer had the polarized morphology of motile cells, witha well-defined uropod. An arc of ICAM-1 was located in the mid-cell region (Fig. 2B), as previously described for motile T cells(30). The strongest accumulation of pMHC-TCR was in the uro-pod of the crawling cells (Fig. 2B), consistent with a phenotypeseen for CD8 T cell blasts (31). As shown in Fig. 2C, the Tregsbroke radial symmetry and formed motile synapses almost im-
FIGURE 1. Tregs modify DC phenotype, concomitant with a change in the stability of Treg–DC interactions. (A) TCR transgenic CD4 T cells were
cultured under Treg-polarizing conditions and expressed high levels of Foxp3 and CTLA-4 at day 4 poststimulation. Naive B10.BR CD4 T cells are shown
for comparison. (B) Tregs were labeled with CFSE and introduced to BMDCs loaded with MCC peptide. Most Treg–DC interactions were stable. For
further examples of stable Treg–DC contacts, see Supplemental Video 1. (C) Twenty-four hours after addition, the Tregs were highly motile and swarmed
around BMDCs. (D) Fresh, CFSE-labeled Tregs were added to BMDCs 24 h after the addition of unlabeled Tregs. These freshly added Tregs had a po-
larized phenotype and crawled along the surface of BMDCs. The still images shown in (D) are from Supplemental Video 2. (E–H) Tregs were incubated
with BMDCs in the presence or absence of MCC peptide, and the BMDCs were assayed at 24 h for surface levels of CD80. The T cell/DC ratios were 1:1
(E), 5:1 (F), or 10:1 (G and H). Fab fragments of anti–CTLA-4 (G) or anti-CD28 (H) were included in the cultures at 100 mg/ml. Results are representative of
three (B–D) or six (E–H) independent experiments. Scale bars, 10 mm.
5896 COSTIMULATION CONTROLS Treg MOTILITY
by guest on April 21, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

mediately upon interacting with the bilayers. Motile IS, or“kinapses,” have been previously reported in naive T cells in vitro(32) and in vivo (33).Our examination of Treg–DC interactions suggested that the
level of costimulatory molecules on the APC surface couldmodulate Treg behavior. Therefore, we examined IS and kinapseformation in the presence of 0, 40, and 200 molecules/mm2 GPI-linked CD80 in the bilayers. In the presence of low levels of CD80(40 molecules/mm2), the Tregs predominantly formed motilekinapses, similar to the phenotype observed in the absence ofCD80 (Fig. 3A, 3B, Supplemental Videos 3, 4) . However, whenTregs were introduced to bilayers containing 200 molecules/mm2
CD80, their movement was drastically reduced, with some of theTregs forming stable IS (Fig. 3C, Supplemental Video 5). Analysisof the motility of Tregs on bilayers containing varying amounts ofCD80 showed that high levels of CD80 significantly decreased themean square displacement (MSD) of Tregs over time (Fig. 3D,3E). As shown in Fig. 3F, the average speed of Tregs was sig-nificantly reduced in the presence of high levels of CD80. Becausechanges in the centroid of relatively nonmotile cells can inflatespeed measurements, we also measured the total displacement ofeach cell and divided by track duration. This measurement yieldedvalues close to 0 for nonmotile cells and showed that Tregs aredramatically less motile when introduced to bilayers with highlevels of CD80 (Fig. 3G). We also noted a decrease in thestraightness of Treg tracks in the presence of high levels of CD80(Fig. 3H).Although the fraction of cells that maintained symmetrical,
nonmotile interactions with the planar bilayers for the entirety ofa 45- to 60-min experiment was 3-fold higher in the presence of 200molecules/mm2 CD80, this result did not quite meet the standardfor statistical significance (Fig. 4A). However, significantly fewercells were motile for the entire experiment in the presence of highlevels of CD80 (Fig. 4B). These results indicate that in addition toslowing the motility of Tregs, high levels of CD80 also inducedstable IS formation. As shown in Fig. 2C, Tregs that formedkinapses broke radial symmetry almost immediately upon con-tacting the bilayers. However, we observed that a fraction of thecells regained symmetry and formed a stable IS during the courseof the experiment (Fig. 4C). The percentage of Tregs displayingthis behavior was dramatically increased in the presence of highlevels of CD80 (Fig. 4D).
In the absence of CD80, the vast majority of Tregs moved freelyacross the bilayer, but the addition of CD80 caused some motilecells to become anchored to the substrate via their uropods,frustrating the forward motion of the cells and causing them to turnin circles (Fig. 4E). This behavior was seen at both concentrationsof CD80, but was especially prominent when high levels wereused (Fig. 4F).It has been shown that TCR transgenic mice contain a pop-
ulation of nTregs that are most likely derived from cells expressingendogenous TCR a-chains in addition to transgenic a- andb-chains (34). Indeed, we found that a fraction of naive CD4+
cells from AD10 mice expressed Foxp3 (Fig. 5A). To confirmthat our findings were generalizable to nTregs, we crossedFoxp3-GFP mice to our AD10 TCR transgenic mice and intro-duced purified CD4 cells to bilayers (Fig. 5B, 5C). Unlike acti-vated Tregs, a significant fraction of the nTregs was sessileregardless of CD80 concentration (Fig. 5E), possibly becausethese cells are relatively metabolically inactive compared within vitro–generated Tregs. Nevertheless, the presence on CD80in the bilayers resulted in a significant decrease in motility(Fig. 5D–F). Interestingly, naive non-Tregs were much less mo-tile than Tregs in the absence of CD80. These results demonstratethat nTregs behave similarly to in vitro–induced Tregs in thepresence and absence of CD80.
Treg motility decreases dramatically in the presence of Ag
Tregs interacting with bilayers accumulated pMHC-TCR in thecontact zone (Fig. 2B) and fluxed intracellular calcium (data notshown), but they were almost always continuously motile overthe entire imaging experiment (Fig. 4B). This led us to askwhether there were any changes in Treg motility upon Ag rec-ognition, or whether motility was only altered in the presence ofhigh levels of CD80. Therefore, we introduced AD10+ Tregs tobilayers containing ICAM-1 and MHC loaded with either cognatepeptide (MCC) or an irrelevant peptide (hemoglobin). As shownin Fig. 6A, in the absence of cognate Ag, Tregs moved rapidlyacross the bilayers with only transient accumulations of ICAM-1,and at no time were pMHC-TCR clusters apparent. Analysis ofthe tracks made by Tregs in the absence of Ag showed that Tregsscanning the surface of an APC lacking cognate Ag move muchfaster and straighter than did Tregs that have recognized Ag(Fig. 6B–F). Thus, Tregs are highly motile when scanning the
FIGURE 2. Tregs form immunological kinapses on supported planar bilayers containing pMHC and ICAM-1. (A) Representative field of Tregs inter-
acting with bilayers containing pMHC and ICAM-1. DIC (left) and fluorescence (right) images with pMHC (green) and ICAM-1 (red) are shown. (B)
Representative image of a motile Treg with ICAM-1 accumulated in the midbody of the cell and pMHC accumulation in the uropod. (C) Representative
time-lapse images of the initial Treg–bilayer contact showing pMHC (green) and ICAM-1 (red). Tregs displayed a polarized phenotype rapidly upon
contact with the bilayer. Images are representative of seven independent experiments. Scale bars, 10 mm.
The Journal of Immunology 5897
by guest on April 21, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

surface with ICAM and MHC without specific Ag, slow consid-erably in the presence of Ag, and slow even more, often to thepoint of stopping, in the presence of Ag and high levels of co-stimulation.
CD28–CD80 interactions control Treg motility
To dissect the mechanism by which CD80 modulates Treg motility,we incubated cells with Fab Abs against CD28 or CTLA-4 andintroduced them to bilayers containing pMHC, ICAM-1, and highlevels of CD80. As shown in Fig. 7A, blockade of CD28 resulted inhighly motile Tregs in the presence of CD80. In contrast, many ofthe Tregs treated with anti–CTLA-4 formed relatively nonmotilecontacts with the bilayer or had their uropods anchored (Fig. 7B).We analyzed the tracks of Tregs loaded onto bilayers withoutCD80 or with CD80 and either anti-CD28, anti–CTLA-4, or noAb. Tregs treated with anti-CD28 were much more motile thancells in the no Ab control condition and were almost as motile ascells loaded onto bilayers lacking CD80 (Fig. 7C–G). Treatmentwith anti–CTLA-4 did not have an appreciable effect on motilityby most measures, as 10 min MSD displacement/duration andspeed were not different from the no Ab control (Fig. 7C–E).Treatment with anti–CTLA-4 did result in somewhat straightertracks, although the significance of this finding is unclear (Fig. 7G).
Taken together, these data show that CD28–CD80 interactions controlthe motility of Tregs.
DiscussionIn this study, we propose a model wherein Treg–DC interactionsare modulated by relative levels of costimulatory molecules. In theabsence of Ag recognition, Tregs rapidly migrate over the surfaceof APCs, scanning for their cognate Ag. Upon Ag recognition,Tregs form either relatively slow-moving kinapses or nonmotile ISdepending on the levels of costimulatory molecules. In the pres-ence of high levels of costimulation, symmetrical ISs are favored.After levels of costimulatory molecules are reduced, kinapseformation is favored and the Treg starts migrating again. Thus, byalternating between kinapses and stable ISs, Tregs could effi-ciently contact and modulate levels of costimulatory molecules orotherwise suppress inappropriately activated DCs.Our results are consistent with a recent study showing that
nTregs interacting with supported planar bilayers containing 90molecules/mm2 CD80 form unstable ISs, but because CD80 wasnot tested at higher concentrations in that study, the profoundeffect that CD80 has on Treg motility was not discovered (29). Incontrast to our study, human Tregs were observed to formhyperstable ISs compared with T effector cells (35). This dis-
FIGURE 3. High concentrations of CD80 reduce the motility of Tregs. Tregs were introduced to bilayers containing pMHC, ICAM-1 and 0 (A), 40 (B),
or 200 (C) molecules/mm2 CD80. Time-coded tracks (blue to white from the beginning to the end of the experiment) are overlaid on ICAM-1 images. Scale
bars, 10 mm. The tracks shown are representative of four (A), two (B), or six (C) independent experiments. Supplemental Videos 3, 4, and 5 are time-lapse
videos of Tregs interacting with bilayers containing 0, 40, and 200 molecules/mm2 CD80, respectively. (D) MSD over time is plotted for three densities of
CD80, and the MSD at 10 min is shown (E). The average speed (F), displacement/duration (G), and straightness (track displacement/length) (H) of Treg
tracks on bilayers with 0, 40, or 200 molecules/mm2 CD80 are shown. Results (D–H) are from one representative experiment of three with n = 38, 52, and 41
for 0, 40, and 200 molecules/mm2 CD80, respectively. Error bars represent SEM. One-way ANOVAs followed by Tukey HSD tests were used to determine
p values. A p value .0.05 was considered not significant.
5898 COSTIMULATION CONTROLS Treg MOTILITY
by guest on April 21, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

crepancy could be due to a species difference, but is probably bestexplained by differences in the TCR stimulus provided to theTregs. Zanin-Zhorov et al. (35) introduced T cells to supportedplanar bilayers containing anti-CD3ε. It is likely that the presenceof such a strong and qualitatively different TCR stimulus obviatesthe need for CD80 to induce stopping of Tregs, masking the dif-ferences in motility we noted when using saturating concen-trations of a physiological TCR ligand. However, note that CD252
non-Tregs displayed rapid symmetry breaking and motility in thesame conditions, demonstrating that anti-CD3 does not generatea durable stop signal for all human T cells.How does signaling through CD28 alter Treg motility? Cross-
linking CD28 results in Lck-mediated phosphorylation of theYMNMmotif and recruitment of PI3K and Grb2 (36). The activityof PI3K leads to activation of Akt, whereas Grb2 recruits theguanine nucleotide exchange factor Vav1, resulting in activationof the small GTPases Rac1 and Cdc42, which in turn activateWAVE2 and WASp, respectively, ultimately leading to Arp2/3-mediated actin remodeling (37). Additionally, a proline-rich do-main of CD28 is known to recruit several kinases downstream ofthe TCR, including Lck and PKC-u (36). We hypothesize thatTregs have a relatively higher threshold for activation than doconventional T cells, which allows them to monitor their envi-ronment without being inappropriately activated, even in the faceof ubiquitously expressed, low-affinity cognate Ags. Under thisscenario, Tregs would require a robust signal through CD28 beforetransitioning from motile kinapses to stable synapses. In support ofa model where CD28 signaling through PI3K is critical for Tregfunction, it has been shown that Tregs expressing an inactive formof the PI3K catalytic subunit, p110d, are poor suppressors and failto prevent inflammation in a colitis model (38). Strong CD28
crosslinking may simply augment TCR-induced signaling and/orcytoskeletal remodeling, or it could provide a unique signal. Asimilar phenomenon has recently been observed in thymocytes, inwhich CD28-dependent actin remodeling was required for maxi-mal activation after TCR triggering (39).An annular ring of CTLA-4 surrounding the central supramo-
lecular activation cluster characterizes the IS formed by inducedTregs interacting with bilayers containing CD80 (40). This denseaccumulation of CTLA-4 may increase the efficiency of costim-ulatory molecule downmodulation. Thus, the reduced motility andincreased IS formation we observed in the presence of high levelsof costimulation may serve two purposes: 1) to increase the dwelltime of a Treg on a particular activated APC, and 2) to enhance theefficiency of downmodulation. The IS functions as a platform forthe polarized secretion of effector molecules toward the APC (41,42). Therefore, it is possible that additional Treg functions, in-cluding Ag-specific delivery of perforin (43, 44) or IL-10 (42),may require the formation of a stable IS.CTLA-4 ligation has been reported to increase motility of T cells
both in vitro and in vivo (45–47). Because CTLA-4 has a muchhigher affinity for CD80 than CD28 (48), it follows that highlevels of CD80 are needed to trigger the CD28-mediated stopsignal we see in our experiments. We suggest that the abundantexpression of CTLA-4 on Tregs tunes these cells to be highlymotile unless they recognize Ag in the context of large amountsof costimulation. Conventional CD4 T cells, unlike self-reactiveTregs that are constantly seeing Ag, are able to efficiently scanAPCs without this mechanism. In disagreement with our model,a recent report showed that activated CD4+CD25+ cells fromCD282/2 mice did not migrate differently in lymph node slicesfrom CD4+CD25+ cells from CD28+/+ mice (49). The apparent
FIGURE 4. Treg–APC interactions have an altered phenotype in the presence of CD80. The percentage of Tregs that were either nonmotile (A) or
continuously motile (B) during the entire length of the experiment is shown. (C) An example of a cell transitioning from a motile phenotype to a stable IS.
DIC (top) and fluorescence (bottom) images with pMHC (green) and ICAM-1 (red) are shown. Scale bar, 10 mm. (D) The percentage of cells that reformed
IS over the length of an experiment. (E) An example of a cell attached to the bilayer via its uropod. The cell’s track is overlaid on DIC images. Scale bar,
10 mm. (F) The percentage of cells that were anchored by their uropods over the course of an experiment. Data in (A), (B), (D), and (F) show the mean of the
fraction of cells displaying a given phenotype from two (40 molecules/mm2 CD80) or three (0 and 200 molecules/mm2 CD80) independent experiments
with n = 115, 87, and 129 for 0, 40, and 200 molecules/mm2 CD80, respectively. Error bars represent SEM. One-way ANOVAs followed by Tukey HSD
tests were used to determine p values. A p value .0.05 was considered not significant.
The Journal of Immunology 5899
by guest on April 21, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

disparity between these results and ours could be explained by thefact that the absence of CD28 has a profound effect on the de-velopment and homeostasis of Tregs (8). In the absence of CD28,the small number of surviving peripheral Tregs may have com-pensatory mechanisms for activation and motility.Mice that have CD28 specifically ablated in Foxp3+ cells de-
velop severe multiorgan autoimmunity (50). We suggest thatefficient downmodulation of costimulatory molecules on acti-vated APCs presenting self-Ag may be a CD28-dependentfunction that is missing in these mice. This hypothesis leads tothe prediction that blocking CD28 will result in less efficientdownmodulation, and we did see a modest decrease in down-modulation in the presence of anti-CD28 (Fig. 1H). Alternatively,CD28-dependent arrest and symmetrical synapse formation maybe required for other Treg suppressive functions, as mentionedabove.Mice deficient in the actin regulatory protein WASp develop
chronic colitis, and Tregs from these mice are poor suppressorsboth in vitro and in vivo (51–53). WASp is also required forT cells to maintain IS symmetry (30). Although defective mi-gration to lymphoid tissue likely plays a role in the phenotype
described for WASp-deficient Tregs, defective IS formation mayalso be important. Tregs incapable of IS formation could also bedeficient in CD80/86 downmodulation. Thus, constant surveillanceand modulation of DCs (via IS-dependent downmodulation ofcostimulatory molecules) may be required to prevent autoimmunedisease. Consistent with this idea, Tang et al. (19) have demon-strated that nTregs from BDC2.5 TCR Tg mice (which have TCRsspecific for an Ag associated with pancreatic b cells) are motilein the pancreatic lymph node when transferred into NOD mice.However, when the same nTregs are transferred into NOD.CD282/2 mice lacking Tregs, they are nonmotile (19). Given theresults presented in the present study, it is likely that the DCs inthe Treg-deficient animals had relatively high levels of CD80/86compared with animals containing endogenous Tregs, therebyaltering the behavior of the transferred Tregs. Higher levels ofcostimulatory molecules on DCs in the pancreatic lymph node ofthese already diabetes-prone animals could partially explain whyNODmice lacking Tregs develop diabetes more quickly than do wild-type mice (54).To maintain tolerance and immune homeostasis, Tregs need to
monitor the activation state of all the DCs in the body. Our finding
FIGURE 5. CD80 reduces the motility of nTregs on supported planar bilayers. (A) Naive CD4 cells from B10.BR or AD10 mice were assayed for the
expression of CD25 and Foxp3. (B and C) Naive AD10 mice CD4 cells were purified from the spleens of AD10 3 Foxp3-GFP mice and introduced to
supported planar bilayers containing pMHC, ICAM-1, and 0 (B) or 200 (C) molecules/mm2 CD80. Time-coded tracks (blue to white from the beginning to
the end of the experiment) are overlaid on ICAM-1 images. The movements of both nTregs (GFP+) and naive CD4 cells (GFP2) were followed over time,
but only the nTreg tracks are shown. Scale bars, 10 mm. (D) MSD over time is plotted for nTregs and naive CD4 cells on bilayers lacking CD80 and on
bilayers with 200 molecules/mm2 CD80. The MSD at 10 min and the fraction of cells with an MSD of .500 um2 are shown (E and F). The results are
pooled from three experiments with n = 48, 40, 73, and 77 for nTreg/no CD80, nTreg/CD80 200 molecules/mm2, naive/no CD80, and naive/CD80
200 molecules/mm2, respectively. Error bars represent SEM (E) or 95% confidence interval (F). The p values were determined with a one-way ANOVA
followed by a Tukey HSD test (E) or a Fisher exact test (F).
5900 COSTIMULATION CONTROLS Treg MOTILITY
by guest on April 21, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
that Ag can slow Treg migration may help explain how Tregsaccumulate where their Ags are displayed. High CD80/CD86 levelsthen alter Treg behavior further to enable Ag-specific delivery of
suppressive signals. The functional consequences of regulation ofTreg motility and synapse formation by CD28 will be a fertile fieldof investigation in future experiments.
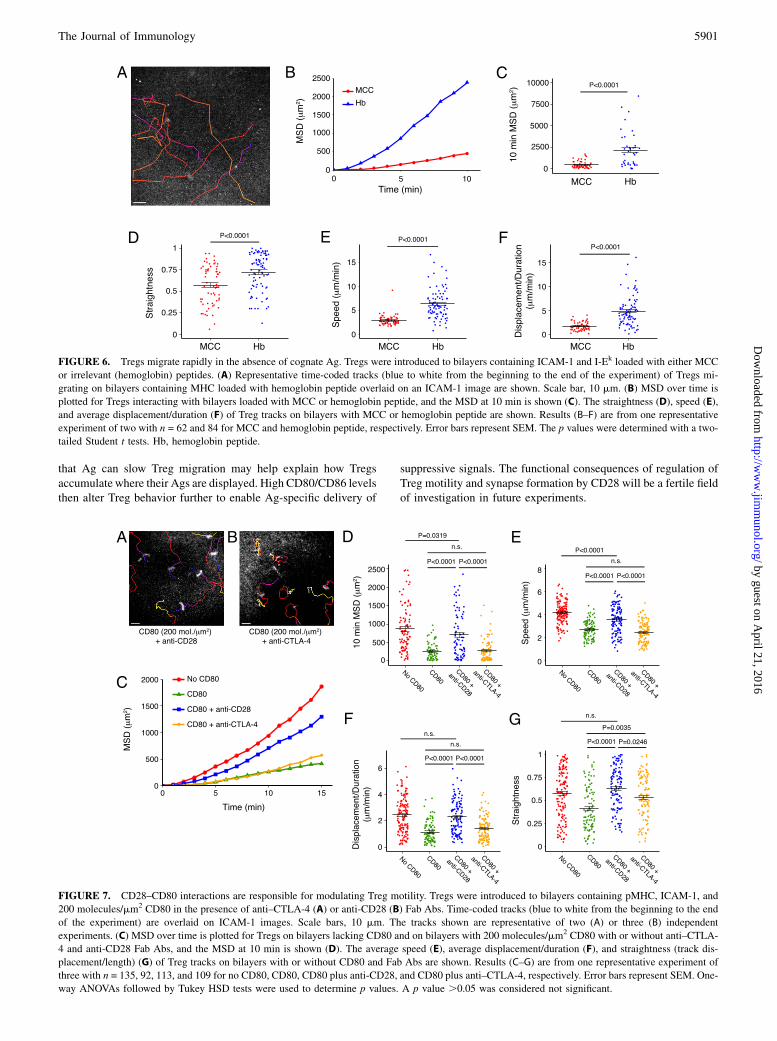
FIGURE 6. Tregs migrate rapidly in the absence of cognate Ag. Tregs were introduced to bilayers containing ICAM-1 and I-Ek loaded with either MCC
or irrelevant (hemoglobin) peptides. (A) Representative time-coded tracks (blue to white from the beginning to the end of the experiment) of Tregs mi-
grating on bilayers containing MHC loaded with hemoglobin peptide overlaid on an ICAM-1 image are shown. Scale bar, 10 mm. (B) MSD over time is
plotted for Tregs interacting with bilayers loaded with MCC or hemoglobin peptide, and the MSD at 10 min is shown (C). The straightness (D), speed (E),
and average displacement/duration (F) of Treg tracks on bilayers with MCC or hemoglobin peptide are shown. Results (B–F) are from one representative
experiment of two with n = 62 and 84 for MCC and hemoglobin peptide, respectively. Error bars represent SEM. The p values were determined with a two-
tailed Student t tests. Hb, hemoglobin peptide.
FIGURE 7. CD28–CD80 interactions are responsible for modulating Treg motility. Tregs were introduced to bilayers containing pMHC, ICAM-1, and
200 molecules/mm2 CD80 in the presence of anti–CTLA-4 (A) or anti-CD28 (B) Fab Abs. Time-coded tracks (blue to white from the beginning to the end
of the experiment) are overlaid on ICAM-1 images. Scale bars, 10 mm. The tracks shown are representative of two (A) or three (B) independent
experiments. (C) MSD over time is plotted for Tregs on bilayers lacking CD80 and on bilayers with 200 molecules/mm2 CD80 with or without anti–CTLA-
4 and anti-CD28 Fab Abs, and the MSD at 10 min is shown (D). The average speed (E), average displacement/duration (F), and straightness (track dis-
placement/length) (G) of Treg tracks on bilayers with or without CD80 and Fab Abs are shown. Results (C–G) are from one representative experiment of
three with n = 135, 92, 113, and 109 for no CD80, CD80, CD80 plus anti-CD28, and CD80 plus anti–CTLA-4, respectively. Error bars represent SEM. One-
way ANOVAs followed by Tukey HSD tests were used to determine p values. A p value .0.05 was considered not significant.
The Journal of Immunology 5901
by guest on April 21, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

AcknowledgmentsWe thank members of the Parker laboratory for helpful discussions, Thomas
H€unig for the anti-CD28 mAb (clone E18), and V.K. Thomas for the
purified GPI-anchored CD80.
DisclosuresThe authors have no financial conflicts of interest.
References1. Belkaid, Y., and K. Tarbell. 2009. Regulatory T cells in the control of host-
microorganism interactions. Annu. Rev. Immunol. 27: 551–589.2. Sakaguchi, S., T. Yamaguchi, T. Nomura, and M. Ono. 2008. Regulatory T cells
and immune tolerance. Cell 133: 775–787.3. Fontenot, J. D., M. A. Gavin, and A. Y. Rudensky. 2003. Foxp3 programs the
development and function of CD4+CD25+ regulatory T cells. Nat. Immunol. 4:330–336.
4. Khattri, R., T. Cox, S. A. Yasayko, and F. Ramsdell. 2003. An essential role forScurfin in CD4+CD25+ T regulatory cells. Nat. Immunol. 4: 337–342.
5. Hori, S., T. Nomura, and S. Sakaguchi. 2003. Control of regulatory T cell de-velopment by the transcription factor Foxp3. Science 299: 1057–1061.
6. Wing, K., Y. Onishi, P. Prieto-Martin, T. Yamaguchi, M. Miyara, Z. Fehervari,T. Nomura, and S. Sakaguchi. 2008. CTLA-4 control over Foxp3+ regulatoryT cell function. Science 322: 271–275.
7. Tai, X., M. Cowan, L. Feigenbaum, and A. Singer. 2005. CD28 costimulation ofdeveloping thymocytes induces Foxp3 expression and regulatory T cell differ-entiation independently of interleukin 2. Nat. Immunol. 6: 152–162.
8. Tang, Q., K. J. Henriksen, E. K. Boden, A. J. Tooley, J. Ye, S. K. Subudhi,X. X. Zheng, T. B. Strom, and J. A. Bluestone. 2003. Cutting edge: CD28controls peripheral homeostasis of CD4+CD25+ regulatory T cells. J. Immunol.171: 3348–3352.
9. Horwitz, D. A., S. G. Zheng, and J. D. Gray. 2008. Natural and TGF-b-inducedFoxp3+CD4+ CD25+ regulatory T cells are not mirror images of each other.Trends Immunol. 29: 429–435.
10. Vignali, D. A., L. W. Collison, and C. J. Workman. 2008. How regulatory T cellswork. Nat. Rev. Immunol. 8: 523–532.
11. Tang, Q., and J. A. Bluestone. 2008. The Foxp3+ regulatory T cell: a jack of alltrades, master of regulation. Nat. Immunol. 9: 239–244.
12. Cederbom, L., H. Hall, and F. Ivars. 2000. CD4+CD25+ regulatory T cells down-regulate co-stimulatory molecules on antigen-presenting cells. Eur. J. Immunol.30: 1538–1543.
13. Misra, N., J. Bayry, S. Lacroix-Desmazes, M. D. Kazatchkine, andS. V. Kaveri. 2004. Cutting edge: human CD4+CD25+ T cells restrain thematuration and antigen-presenting function of dendritic cells. J. Immunol.172: 4676–4680.
14. Oderup, C., L. Cederbom, A. Makowska, C. M. Cilio, and F. Ivars. 2006. Cy-totoxic T lymphocyte antigen-4-dependent down-modulation of costimulatorymolecules on dendritic cells in CD4+ CD25+ regulatory T-cell-mediated sup-pression. Immunology 118: 240–249.
15. Onishi, Y., Z. Fehervari, T. Yamaguchi, and S. Sakaguchi. 2008. Foxp3+
natural regulatory T cells preferentially form aggregates on dendritic cellsin vitro and actively inhibit their maturation. Proc. Natl. Acad. Sci. USA 105:10113–10118.
16. Qureshi, O. S., Y. Zheng, K. Nakamura, K. Attridge, C. Manzotti,E. M. Schmidt, J. Baker, L. E. Jeffery, S. Kaur, Z. Briggs, et al. 2011. Trans-endocytosis of CD80 and CD86: a molecular basis for the cell-extrinsic functionof CTLA-4. Science 332: 600–603.
17. Kong, K. F., G. Fu, Y. Zhang, T. Yokosuka, J. Casas, A. J. Canonigo-Balancio,S. Becart, G. Kim, J. R. Yates, III, M. Kronenberg, et al. 2014. Protein kinaseC-h controls CTLA-4-mediated regulatory T cell function. Nat. Immunol. 15: 465–472.
18. Tadokoro, C. E., G. Shakhar, S. Shen, Y. Ding, A. C. Lino, A. Maraver,J. J. Lafaille, and M. L. Dustin. 2006. Regulatory T cells inhibit stable contactsbetween CD4+ T cells and dendritic cells in vivo. J. Exp. Med. 203: 505–511.
19. Tang, Q., J. Y. Adams, A. J. Tooley, M. Bi, B. T. Fife, P. Serra, P. Santamaria,R. M. Locksley, M. F. Krummel, and J. A. Bluestone. 2006. Visualizing regu-latory T cell control of autoimmune responses in nonobese diabetic mice. Nat.Immunol. 7: 83–92.
20. Bauer, C. A., E. Y. Kim, F. Marangoni, E. Carrizosa, N. M. Claudio, andT. R. Mempel. 2014. Dynamic Treg interactions with intratumoral APCs pro-mote local CTL dysfunction. J. Clin. Invest. 124: 2425–2440.
21. Wetzel, S. A., T. W. McKeithan, and D. C. Parker. 2002. Live-cell dynamics andthe role of costimulation in immunological synapse formation. J. Immunol. 169:6092–6101.
22. Kaye, J., N. J. Vasquez, and S. M. Hedrick. 1992. Involvement of the sameregion of the T cell antigen receptor in thymic selection and foreign peptiderecognition. J. Immunol. 148: 3342–3353.
23. Lutz, M. B., N. Kukutsch, A. L. Ogilvie, S. Rossner, F. Koch, N. Romani, andG. Schuler. 1999. An advanced culture method for generating large quantities ofhighly pure dendritic cells from mouse bone marrow. J. Immunol. Methods 223:77–92.
24. Beyersdorf, N., X. Ding, G. Blank, K. M. Dennehy, T. Kerkau, and T. H€unig.2008. Protection from graft-versus-host disease with a novel B7 binding site-specific mouse anti-mouse CD28 monoclonal antibody. Blood 112: 4328–4336.
25. Grakoui, A., S. K. Bromley, C. Sumen, M. M. Davis, A. S. Shaw, P. M. Allen,and M. L. Dustin. 1999. The immunological synapse: a molecular machinecontrolling T cell activation. Science 285: 221–227.
26. Thauland, T. J., Y. Koguchi, S. A. Wetzel, M. L. Dustin, and D. C. Parker. 2008.Th1 and Th2 cells form morphologically distinct immunological synapses. J.Immunol. 181: 393–399.
27. Bromley, S. K., A. Iaboni, S. J. Davis, A. Whitty, J. M. Green, A. S. Shaw,A. Weiss, and M. L. Dustin. 2001. The immunological synapse and CD28-CD80interactions. Nat. Immunol. 2: 1159–1166.
28. Benson, M. J., K. Pino-Lagos, M. Rosemblatt, and R. J. Noelle. 2007. All-transretinoic acid mediates enhanced T reg cell growth, differentiation, and guthoming in the face of high levels of co-stimulation. J. Exp. Med. 204: 1765–1774.
29. Tomiyama, T., Y. Ueda, T. Katakai, N. Kondo, K. Okazaki, and T. Kinashi. 2013.Antigen-specific suppression and immunological synapse formation by regula-tory T cells require the Mst1 kinase. PLoS ONE 8: e73874.
30. Sims, T. N., T. J. Soos, H. S. Xenias, B. Dubin-Thaler, J. M. Hofman, J. C. Waite,T. O. Cameron, V. K. Thomas, R. Varma, C. H. Wiggins, et al. 2007. Opposingeffects of PKCu and WASp on symmetry breaking and relocation of the im-munological synapse. Cell 129: 773–785.
31. Beemiller, P., J. Jacobelli, and M. F. Krummel. 2012. Integration of the move-ment of signaling microclusters with cellular motility in immunological syn-apses. Nat. Immunol. 13: 787–795.
32. Gunzer, M., A. Schafer, S. Borgmann, S. Grabbe, K. S. Zanker, E. B. Brocker,E. Kampgen, and P. Friedl. 2000. Antigen presentation in extracellular matrix:interactions of T cells with dendritic cells are dynamic, short lived, and se-quential. Immunity 13: 323–332.
33. Mempel, T. R., S. E. Henrickson, and U. H. Von Andrian. 2004. T-cell primingby dendritic cells in lymph nodes occurs in three distinct phases. Nature 427:154–159.
34. Itoh, M., T. Takahashi, N. Sakaguchi, Y. Kuniyasu, J. Shimizu, F. Otsuka, andS. Sakaguchi. 1999. Thymus and autoimmunity: production of CD25+CD4+
naturally anergic and suppressive T cells as a key function of the thymus inmaintaining immunologic self-tolerance. J. Immunol. 162: 5317–5326.
35. Zanin-Zhorov, A., Y. Ding, S. Kumari, M. Attur, K. L. Hippen, M. Brown,B. R. Blazar, S. B. Abramson, J. J. Lafaille, and M. L. Dustin. 2010. Proteinkinase C-u mediates negative feedback on regulatory T cell function. Science328: 372–376.
36. Rudd, C. E., A. Taylor, and H. Schneider. 2009. CD28 and CTLA-4 coreceptorexpression and signal transduction. Immunol. Rev. 229: 12–26.
37. Burkhardt, J. K., E. Carrizosa, and M. H. Shaffer. 2008. The actin cytoskeleton inT cell activation. Annu. Rev. Immunol. 26: 233–259.
38. Patton, D. T., O. A. Garden, W. P. Pearce, L. E. Clough, C. R. Monk, E. Leung,W. C. Rowan, S. Sancho, L. S. Walker, B. Vanhaesebroeck, and K. Okkenhaug.2006. Cutting edge: the phosphoinositide 3-kinase p110d is critical for thefunction of CD4+CD25+Foxp3+ regulatory T cells. J. Immunol. 177: 6598–6602.
39. Tan, Y. X., B. N. Manz, T. S. Freedman, C. Zhang, K. M. Shokat, and A. Weiss.2014. Inhibition of the kinase Csk in thymocytes reveals a requirement for actinremodeling in the initiation of full TCR signaling. Nat. Immunol. 15: 186–194.
40. Yokosuka, T., W. Kobayashi, M. Takamatsu, K. Sakata-Sogawa, H. Zeng,A. Hashimoto-Tane, H. Yagita, M. Tokunaga, and T. Saito. 2010. Spatiotemporalbasis of CTLA-4 costimulatory molecule-mediated negative regulation of T cellactivation. Immunity 33: 326–339.
41. Griffiths, G. M., A. Tsun, and J. C. Stinchcombe. 2010. The immunologicalsynapse: a focal point for endocytosis and exocytosis. J. Cell Biol. 189: 399–406.
42. Huse, M., B. F. Lillemeier, M. S. Kuhns, D. S. Chen, and M. M. Davis. 2006.T cells use two directionally distinct pathways for cytokine secretion. Nat.Immunol. 7: 247–255.
43. Zhao, D. M., A. M. Thornton, R. J. DiPaolo, and E. M. Shevach. 2006. ActivatedCD4+CD25+ T cells selectively kill B lymphocytes. Blood 107: 3925–3932.
44. Boissonnas, A., A. Scholer-Dahirel, V. Simon-Blancal, L. Pace, F. Valet,A. Kissenpfennig, T. Sparwasser, B. Malissen, L. Fetler, and S. Amigorena.2010. Foxp3+ T cells induce perforin-dependent dendritic cell death in tumor-draining lymph nodes. Immunity 32: 266–278.
45. Schneider, H., J. Downey, A. Smith, B. H. Zinselmeyer, C. Rush, J. M. Brewer,B. Wei, N. Hogg, P. Garside, and C. E. Rudd. 2006. Reversal of the TCR stopsignal by CTLA-4. Science 313: 1972–1975.
46. Schneider, H., X. Smith, H. Liu, G. Bismuth, and C. E. Rudd. 2008. CTLA-4disrupts ZAP70 microcluster formation with reduced T cell/APC dwell timesand calcium mobilization. Eur. J. Immunol. 38: 40–47.
47. Ruocco, M. G., K. A. Pilones, N. Kawashima, M. Cammer, J. Huang, J. S. Babb,M. Liu, S. C. Formenti, M. L. Dustin, and S. Demaria. 2012. Suppressing T cellmotility induced by anti-CTLA-4 monotherapy improves antitumor effects. J.Clin. Invest. 122: 3718–3730.
48. Collins, A. V., D. W. Brodie, R. J. Gilbert, A. Iaboni, R. Manso-Sancho,B. Walse, D. I. Stuart, P. A. van der Merwe, and S. J. Davis. 2002. The inter-action properties of costimulatory molecules revisited. Immunity 17: 201–210.
49. Lu, Y., H. Schneider, and C. E. Rudd. 2012. Murine regulatory T cells differfrom conventional T cells in resisting the CTLA-4 reversal of TCR stop-signal.Blood 120: 4560–4570.
50. Zhang, R., A. Huynh, G. Whitcher, J. Chang, J. S. Maltzman, and L. A. Turka.2013. An obligate cell-intrinsic function for CD28 in Tregs. J. Clin. Invest. 123:580–593.
51. Maillard, M. H., V. Cotta-de-Almeida, F. Takeshima, D. D. Nguyen, P. Michetti,C. Nagler, A. K. Bhan, and S. B. Snapper. 2007. The Wiskott-Aldrich syndromeprotein is required for the function of CD4+CD25+Foxp3+ regulatory T cells. J.Exp. Med. 204: 381–391.
5902 COSTIMULATION CONTROLS Treg MOTILITY
by guest on April 21, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

52. Marangoni, F., S. Trifari, S. Scaramuzza, C. Panaroni, S. Martino,L. D. Notarangelo, Z. Baz, A. Metin, F. Cattaneo, A. Villa, et al. 2007. WASPregulates suppressor activity of human and murine CD4+CD25+FOXP3+ naturalregulatory T cells. J. Exp. Med. 204: 369–380.
53. Snapper, S. B., F. S. Rosen, E. Mizoguchi, P. Cohen, W. Khan, C. H. Liu,T. L. Hagemann, S. P. Kwan, R. Ferrini, L. Davidson, et al. 1998. Wiskott-
Aldrich syndrome protein-deficient mice reveal a role for WASP in T but notB cell activation. Immunity 9: 81–91.
54. Salomon, B., D. J. Lenschow, L. Rhee, N. Ashourian, B. Singh, A. Sharpe, andJ. A. Bluestone. 2000. B7/CD28 costimulation is essential for the homeostasis ofthe CD4+CD25+ immunoregulatory T cells that control autoimmune diabetes.Immunity 12: 431–440.
The Journal of Immunology 5903
by guest on April 21, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
Related Documents











