APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Sept. 2011, p. 6215–6224 Vol. 77, No. 17 0099-2240/11/$12.00 doi:10.1128/AEM.00662-11 Copyright © 2011, American Society for Microbiology. All Rights Reserved. Cd-Specific Mutants of Mercury-Sensing Regulatory Protein MerR, Generated by Directed Evolution † Kaisa M. Hakkila, 1,2 * Pia A. Nikander, 1,2 Sini M. Junttila, 1 ‡ Urpo J. Lamminma ¨ki, 1 and Marko P. Virta 2 * Department of Biochemistry and Food Chemistry, University of Turku, FI-20014 Turku, Finland, 1 and Department of Food and Environmental Sciences, University of Helsinki, P.O. Box 56, FI-00014 Helsinki, Finland 2 Received 23 March 2011/Accepted 28 June 2011 The mercury-sensing regulatory protein, MerR (Tn21), which regulates mercury resistance operons in Gram-negative bacteria, was subjected to directed evolution in an effort to generate a MerR mutant that responds to Cd but not Hg. Oligonucleotide-directed mutagenesis was used to introduce random mutations into the key metal-binding regions of MerR. The effects of these mutations were assessed using a vector in which MerR controlled the expression of green fluorescent protein (GFP) and luciferase via the mer operator/ promoter. An Escherichia coli cell library was screened by fluorescence-activated cell sorting, using a fluores- cence-based dual screening strategy that selected for MerR mutants that showed GFP repression when cells were induced with Hg but GFP activation in the presence of Cd. Two Cd-responsive MerR mutants with decreased responses toward Hg were identified through the first mutagenesis/selection round. These mutants were used for a second mutagenesis/selection round, which yielded eight Cd-specific mutants that had no significant response to Hg, Zn, or the other tested metal(loid)s. Seven of the eight Cd-specific MerR mutants showed repressor activities equal to that of wild-type (wt) MerR. These Cd-specific mutants harbored multiple mutations (12 to 22) in MerR, indicating that the alteration of metal specificity with maintenance of repressor function was due to the combined effect of many mutations rather than just a few amino acid changes. The amino acid changes were studied by alignment against the sequences of MerR and other metal-responsive MerR family proteins. The analysis indicated that the generated Cd-specific MerR mutants appear to be unique among the MerR family members characterized to date. It is fundamentally important that we understand the inter- actions of metal ions in biological systems, as estimates indi- cate that 40% of all known proteins contain metal ions (25). The relationship between the structure of a protein and its binding specificity toward different metal ions has been the subject of active research but is still not well understood (10, 25). In bacteria, the metalloregulatory proteins play crucial roles in discriminating among metal ions, controlling the cellular levels of essential metals, and exporting/detoxifying metal(loid)s (17). The members of the metal-responsive MerR protein family are transcriptional activators that can be grouped on the basis of their metal-sensing preferences for Hg (MerR), Cu (CueR), Cd (CadR), Pb (PbrR), Zn (ZntR), and Au (GolS) (6, 21, 22). The archetypal and best-studied MerR family member is the mercuric ion-sensing protein, MerR, which was initially identified to be a regulator of the mercury resistance operons found in transposons Tn501 and Tn21 of the Gram-negative bacteria Pseudomonas aeruginosa and Shi- gella flexneri, respectively (2, 5, 26, 37). MerR functions as a homodimer and binds to the dyad operator (MerOP), the gene for which is located between 10 and 35 promoter elements of the mercury resistance operon (20). In the absence of Hg, MerR prevents transcription initiation by bending DNA (1) and forming an inactive complex with RNA polymerase (15, 20). In vitro, MerR binds one Hg ion per dimer; this provokes an allosteric change in the MerR protein that triggers unwind- ing of the spacer DNA, allowing transcription initiation of the mer operon (27). Cd and Zn can also activate the mer operon, but only when they are present at 100- and 1,000-fold higher concentrations, respectively, than the effective concentration of Hg (31). The three-dimensional (3D) structure of MerR has not yet been solved, but the crystal structures of ZntR and CueR (8), together with genetic, biochemical, and structural studies of MerR, have suggested that the MerR protein com- prises an N-terminal helix-turn-helix DNA-binding domain (residues Ile10 to Arg29), a C-terminal Hg-binding domain (residues Cys82 to Cys126) containing a long helical region (Cys82 to Cys117), and the so-called coupling domain (Lys30- His81), which lies between the first two domains and is pro- posed to mediate inductive signaling from the metal-binding domain to the DNA-binding domain (33, 35, 38). Residues 80 through 128 have been shown to be necessary and sufficient for stable dimer formation and high-affinity Hg binding (43). The MerR homodimer binds Hg via three cysteine thiols that are conserved among all MerR proteins (19, 33, 40, 42). In the present study, we constructed mutant libraries where metal-binding regions of MerR were randomized and * Corresponding author. Mailing address for Kaisa M. Hakkila: De- partment of Biochemistry and Food Chemistry, Molecular Plant Biol- ogy, University of Turku, Luonnontieteidentalo II, 2. krs, FI-20014 Turku, Finland. Phone: 358 2 333 5798. Fax: 358 2 333 8075. E-mail: kaihak@utu.fi. Mailing address for Marko P. Virta: Department of Food and Environmental Sciences, Microbiology, University of Helsinki, P.O. Box 56, FI-00014 Helsinki, Finland. Phone: 358 9 191 57586. Fax: 358 9 191 59322. E-mail: marko.virta@helsinki.fi. ‡ Present address: Turku Centre for Biotechnology, Tykisto ¨katu 6, FI-20520 Turku, Finland. † Supplemental material for this article may be found at http://aem .asm.org/. Published ahead of print on 15 July 2011. 6215

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Sept. 2011, p. 6215–6224 Vol. 77, No. 170099-2240/11/$12.00 doi:10.1128/AEM.00662-11Copyright © 2011, American Society for Microbiology. All Rights Reserved.
Cd-Specific Mutants of Mercury-Sensing Regulatory Protein MerR,Generated by Directed Evolution�†
Kaisa M. Hakkila,1,2* Pia A. Nikander,1,2 Sini M. Junttila,1‡Urpo J. Lamminmaki,1 and Marko P. Virta2*
Department of Biochemistry and Food Chemistry, University of Turku, FI-20014 Turku, Finland,1 and Department ofFood and Environmental Sciences, University of Helsinki, P.O. Box 56, FI-00014 Helsinki, Finland2
Received 23 March 2011/Accepted 28 June 2011
The mercury-sensing regulatory protein, MerR (Tn21), which regulates mercury resistance operons inGram-negative bacteria, was subjected to directed evolution in an effort to generate a MerR mutant thatresponds to Cd but not Hg. Oligonucleotide-directed mutagenesis was used to introduce random mutationsinto the key metal-binding regions of MerR. The effects of these mutations were assessed using a vector inwhich MerR controlled the expression of green fluorescent protein (GFP) and luciferase via the mer operator/promoter. An Escherichia coli cell library was screened by fluorescence-activated cell sorting, using a fluores-cence-based dual screening strategy that selected for MerR mutants that showed GFP repression when cellswere induced with Hg but GFP activation in the presence of Cd. Two Cd-responsive MerR mutants withdecreased responses toward Hg were identified through the first mutagenesis/selection round. These mutantswere used for a second mutagenesis/selection round, which yielded eight Cd-specific mutants that had nosignificant response to Hg, Zn, or the other tested metal(loid)s. Seven of the eight Cd-specific MerR mutantsshowed repressor activities equal to that of wild-type (wt) MerR. These Cd-specific mutants harbored multiplemutations (12 to 22) in MerR, indicating that the alteration of metal specificity with maintenance of repressorfunction was due to the combined effect of many mutations rather than just a few amino acid changes. Theamino acid changes were studied by alignment against the sequences of MerR and other metal-responsiveMerR family proteins. The analysis indicated that the generated Cd-specific MerR mutants appear to beunique among the MerR family members characterized to date.
It is fundamentally important that we understand the inter-actions of metal ions in biological systems, as estimates indi-cate that 40% of all known proteins contain metal ions (25).The relationship between the structure of a protein and itsbinding specificity toward different metal ions has been thesubject of active research but is still not well understood (10,25). In bacteria, the metalloregulatory proteins play crucialroles in discriminating among metal ions, controlling thecellular levels of essential metals, and exporting/detoxifyingmetal(loid)s (17). The members of the metal-responsive MerRprotein family are transcriptional activators that can begrouped on the basis of their metal-sensing preferences for Hg(MerR), Cu (CueR), Cd (CadR), Pb (PbrR), Zn (ZntR), andAu (GolS) (6, 21, 22). The archetypal and best-studied MerRfamily member is the mercuric ion-sensing protein, MerR,which was initially identified to be a regulator of the mercuryresistance operons found in transposons Tn501 and Tn21 of
the Gram-negative bacteria Pseudomonas aeruginosa and Shi-gella flexneri, respectively (2, 5, 26, 37). MerR functions as ahomodimer and binds to the dyad operator (MerOP), the genefor which is located between �10 and �35 promoter elementsof the mercury resistance operon (20). In the absence of Hg,MerR prevents transcription initiation by bending DNA (1)and forming an inactive complex with RNA polymerase (15,20). In vitro, MerR binds one Hg ion per dimer; this provokesan allosteric change in the MerR protein that triggers unwind-ing of the spacer DNA, allowing transcription initiation of themer operon (27). Cd and Zn can also activate the mer operon,but only when they are present at 100- and 1,000-fold higherconcentrations, respectively, than the effective concentrationof Hg (31). The three-dimensional (3D) structure of MerR hasnot yet been solved, but the crystal structures of ZntR andCueR (8), together with genetic, biochemical, and structuralstudies of MerR, have suggested that the MerR protein com-prises an N-terminal helix-turn-helix DNA-binding domain(residues Ile10 to Arg29), a C-terminal Hg-binding domain(residues Cys82 to Cys126) containing a long helical region(Cys82 to Cys117), and the so-called coupling domain (Lys30-His81), which lies between the first two domains and is pro-posed to mediate inductive signaling from the metal-bindingdomain to the DNA-binding domain (33, 35, 38). Residues 80through 128 have been shown to be necessary and sufficient forstable dimer formation and high-affinity Hg binding (43). TheMerR homodimer binds Hg via three cysteine thiols that areconserved among all MerR proteins (19, 33, 40, 42).
In the present study, we constructed mutant librarieswhere metal-binding regions of MerR were randomized and
* Corresponding author. Mailing address for Kaisa M. Hakkila: De-partment of Biochemistry and Food Chemistry, Molecular Plant Biol-ogy, University of Turku, Luonnontieteidentalo II, 2. krs, FI-20014Turku, Finland. Phone: 358 2 333 5798. Fax: 358 2 333 8075. E-mail:[email protected]. Mailing address for Marko P. Virta: Department ofFood and Environmental Sciences, Microbiology, University ofHelsinki, P.O. Box 56, FI-00014 Helsinki, Finland. Phone: 358 9 19157586. Fax: 358 9 191 59322. E-mail: [email protected].
‡ Present address: Turku Centre for Biotechnology, Tykistokatu 6,FI-20520 Turku, Finland.
† Supplemental material for this article may be found at http://aem.asm.org/.
� Published ahead of print on 15 July 2011.
6215

screened the libraries for those mutants that were activatedby Cd but not Hg. In the screening vector, a reporter geneencoding GFPmut2 (12) was placed under the control ofMerOP, and either Hg or Cd was used to induce the mutantMerR library. On the basis of high or low green fluorescenceintensity, rounds of fluorescence-activated cell sorting(FACS) were performed to enrich Cd-specific MerR mu-tants. As a result, we report on MerR mutants with a strictspecificity for a single metal, Cd, showing that it is possibleto remove the activity of MerR for its preferred metal whilemaintaining the protein’s wild-type (wt) level repressor/ac-tivator functions.
MATERIALS AND METHODS
Construction of mutant merR library. Escherichia coli XL1-Blue was used asthe host for DNA cloning and library construction. The screening vector pKH100contained gfpmut2 (12) and the firefly luciferase gene (luc) (13) under the controlof MerOP. Details of the bacterial strain and construction of the screening vectorare presented in the supplemental material.
Site-directed mutagenesis of mer was done using two different mutagenizingoligonucleotides (Fig. 1; see Table S1 in the supplemental material). In the firstmethod, certain bases of oligonucleotides were doped with other bases, with theproportions designed to yield 79% of the original bases and 7% of each of theother bases. In the second method, selected positions of oligonucleotides wererandomized with equal representation of the desired bases. Some of these mu-tagenized sites of oligonucleotides could potentially encode any amino acid,while some positions were limited to certain amino acids (Fig. 1; see Table S2 inthe supplemental material). The utilized oligonucleotides were commerciallysynthesized by TAG Copenhagen or Oligomer. Details of the molecular cloningof the merR libraries are presented in supplemental material.
Four parallel electroporations were performed to transform each merR libraryinto XL1-Blue competent cells. After 1 h recovery in SOC medium (34), trans-formed cell cultures were paired and pooled. Small samples were plated on Luriaagar (LA) plates supplemented with 100 �g ml�1 ampicillin (Amp) and 12.5 �gml�1 tetracycline (Tet), and colonies were counted to determine the number ofplasmid-carrying cells and the corresponding library diversity. The remainingbacteria were centrifuged at 600 � g for 15 min at 4°C and washed twice withHMM medium (23) supplemented with 0.5% (wt/vol) hydrolyzed casein, 1 �gml�1 thiamine, 200 �g ml�1 Amp, and 12.5 �g ml�1 Tet. The washed cells werediluted to an optical density at 600 nm (OD600) of 0.5 with the same HMMmedium and cultivated to an OD600 of 0.6 to 0.7. During cultivation, ampicillinwas added hourly to maintain plasmid selection.
FIG. 1. Amino acids targeted in each MerR mutant library and their positions in the model of MerR. (a) The schematic shows all six � helicesof MerR (colored as described for panel b) and the positions of the restriction enzymes used during library construction. Helices �3, �4, �5, and�6 are underlined in the presented MerR sequence. The locations of the Cd-responsive single mutations and a double mutation (G79S and M106I[in boldface]) described by Caguiat et al. (7) are shown above the MerR sequence. The numbers represent the different mutational strategies usedin library construction: 1, randomized amino acids; 2, doped amino acids; and 3, amino acids with limited changes. The parental amino acidsequences are shown as Xs. (b) A model of MerR generated using the crystal structure of CueR (8) as a template. The ribbon diagram shows oneprotomer in color and the other in gray. The residues targeted in our mutagenesis strategy are shown in red on the colored protomer and inraspberry on the gray protomer. Residues C82, K99, M106, C117, and C126 are presented in stick format in both protomers.
6216 HAKKILA ET AL. APPL. ENVIRON. MICROBIOL.
Induction of merR library and flow cytometric cell sorting. Induction of merR-dependent gfpmut2 expression was performed in 1 ml containing 100 �l of 1 �10�5 or 5 � 10�5 M HgCl2 (�99.5%; Riedel-de Haen) or 1 � 10�5 or 1 � 10�4
M CdCl2 (99%; Riedel-de Haen) and 900 �l of freshly cultivated transformantcells (OD600, 0.6 to 0.7). The concentrations of metals in the various experimentsare shown in Table 1. After 2 h of metal induction, the cells were pelleted andwashed twice with an equal volume of ice-cold phosphate-buffered saline (PBS;137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 2 mM KH2PO4, pH 7.4). Washedbacteria were stored overnight at 4°C, and each merR library was suspended with1 ml of PBS.
Flow cytometric sorting was performed with an argon laser (488 nm) in eithera FACSVantage SE or a FACSCalibur (BDBiosciences) FACS apparatus on thebasis of the technique described by Valdivia and Falkow (41) and Tang et al.(39). Bacteria were detected by forward and side scatter and green fluorescenceusing a 530/30-nm band-pass filter. The GFPmut2 used in this study has anexcitation maximum at 481 nm and an emission maximum at 507 nm (12). Sidescatter, forward scatter, and fluorescence data were collected with logarithmicamplification. The sorting parameters were set to discriminate on the basis offluorescence intensity. The sorting mechanism of the FACSCalibur apparatusdiffered from that of the FACSVantage SE apparatus, in that the FACSCalibursystem used a mechanical catcher tube to sort cells, while the FACSVantagesystem was a jet-in-air-type cell sorter. Approximately 5,000 events/s were ana-lyzed with the FACSVantage SE apparatus. To obtain a 99% purity of sortedcells, the analysis rate of the FACSCalibur apparatus was set to not exceed 300cells/s, as recommended (14); however, the actual sorting rates depended on theproperties of the library and sorting gate used in each experiment.
The first sorting round was done with the higher-throughput FACSVantageflow cytometer, since the aim was to interrogate 10 times more cells than thenumber of initial transformed colonies (i.e., to sample each transformant clone�10 times) (Table 1). Subsequent sorting rounds involved the lower-throughputFACSCalibur apparatus, since the first sort considerably reduced the library size.After the FACSVantage sort, cells were plated on LA supplemented with 100 �gml�1 Amp and 12.5 �g ml�1 Tet. After being sorted with the FACSCaliburapparatus, the sorted cells were in 300 to 500 ml of sheath fluid (sterile PBS). Thecells were recovered using 45-�m-pore-size membrane filters in plastic filterholders (Schleicher & Schuell MicroScience GmbH), each filter was placed on anLA plate supplemented with 100 �g ml�1 Amp and 12.5 �g ml�1 Tet, and thecells were grown overnight on filter membranes. After each sorting experiment,approximately 30 to 52 colonies were further tested as described below. Lowviability of the sorted cells was a problem throughout the study; the reasons forthis are discussed in the supplemental material.
Preliminary testing of MerR mutants in vivo. Isolated MerR mutants weregrown on LA plates supplemented with 100 �g ml�1 Amp and 12.5 �g ml�1 Tet,and single colonies derived from sorted cells were separately inoculated to 2 mlof HMM medium. After cells were cultivated for 24 h at 37°C with shaking at 250rpm, luminescence was measured using a microplate luminometer (LabsystemsLuminoskan; Thermo Electron Corporation). Nonluminescent cultures werediscarded. Cell density was observed by visual inspection (OD600, higher thanapproximately 0.3), and 10 ml of HMM medium was added to cultures that weredetermined to have a high cell density. Precultures with low cell densities wereused directly. On white 96-well microtiter plates (Thermo Electron Corpora-tion), 50 �l of standard metal solution (3 � 10�8, 1 � 10�7, and 3 � 10�7 M forHgCl2; 1 � 10�7, 3 � 10�7, 1 � 10�6, and 3 � 10�6 M for CdCl2) or water wasmixed with 50 �l of bacterial suspension per well. The plates were incubated at
37°C for 2 h, and then 100 �l of 0.5 mM D-luciferin (in 0.1 M sodium citratebuffer, pH 5.0) was added to each reaction mixture (18). The samples wereincubated for 5 min at room temperature and then analyzed with a Luminoskanplate luminometer (Thermo Electron Corporation). Mutants showing alteredresponses to the inducing metals were sequenced.
Characterization of MerR mutants in vivo. For further in vivo testing of themetal specificities of the various MerR mutants, MerR mutants were grownovernight in HMM medium, diluted to an OD600 of 0.02 with HMM medium,and cultivated until the OD600 reached 0.10. For luminescence measurement, theculture was then diluted to an OD600 of 0.05, and measurements were performedas described above using HgCl2 and CdCl2 concentrations ranging from 6 �10�10 to 2 � 10�5 M. MerR mutants were also tested with the following metals:Pb(CH3COO)2 (�99.5%), CuSO4 � 5H2O (99%), and AgNO3 (99%) (from J. T.Baker); ZnCl2 (�98%) and C4H4KO7Sb � 0.5 H2O (99 to 103%) (from FlukaChemica); NaAsO2 (92%), Na2HAsO4 � 7H2O (100.3%), AlCl3 (99%), andK2Cr2O7 (99%) (from Sigma-Aldrich); and CoCl2 (analytical grade; Riedel deHaen).
The metal-dependent luminescence responses of the MerR mutants were fit bynonlinear regression to a sigmoidal dose-response curve (variable slope) usingthe Prism software package (GraphPad Software). The quality of fit was ex-pressed as an r-squared (R2) value. The effective concentration of the metal thatprovoked the half-maximum response (the 50% effective concentration [EC50]),the activation efficiency (NLmax; which described the maximum luminescenceresponse with respect to normalized luminescence, i.e., the maximum lumines-cence signal divided by the minimum luminescence signal), and the Hill coeffi-cient (which described the steepness of the curve and indicated the sensitivity ofthe response [31]) were determined. The background luminescence value, whichwas obtained from uninduced cells and reflected the repressor activity of theMerR mutant, was expressed as relative light units (RLUs). The limit of deter-mination (LOD) was calculated using the equation, (XB � 3SDB)/XB, where XB
is the mean value of background luminescence and SDB is the standard devia-tion.
Construction of homology model of MerR and sequence alignments of mu-tants against MerR family members. A 3D model of MerR was generated usingthe crystal structure of CueR (Protein Data Bank entry 1QO5 [8]) as a template.Details of the homology modeling of MerR (Tn21) are described in the supple-mental material.
Metalloregulators of the MerR family proteins were classified on the basis oftheir metal preferences (8). Various numbers of sequences were selected fromdifferent species and used in multiple-sequence alignments (136 for MerR[Hg2�], 92 for CadR and PbrR [Cd2�/Pb2�], 64 for ZntR [Zn2�], and 72 forCueR, HmrR, PmtR, and GolS [Cu�, Ag�, and Au�]). The sequence alignmentswere done with Clustal W in BioEdit sequence analysis program (Tom Hall, IbisBiosciences, Carlsbad, CA). See Fig. S4 to S7 in the supplemental material forthe multiple-sequence alignments.
RESULTS AND DISCUSSION
First mutagenesis round, library A. As a crystal structure isnot yet available for MerR, we used homology modeling topredict the structure of MerR on the basis of that of CueR (8)and then used the predicted structure of MerR to design a
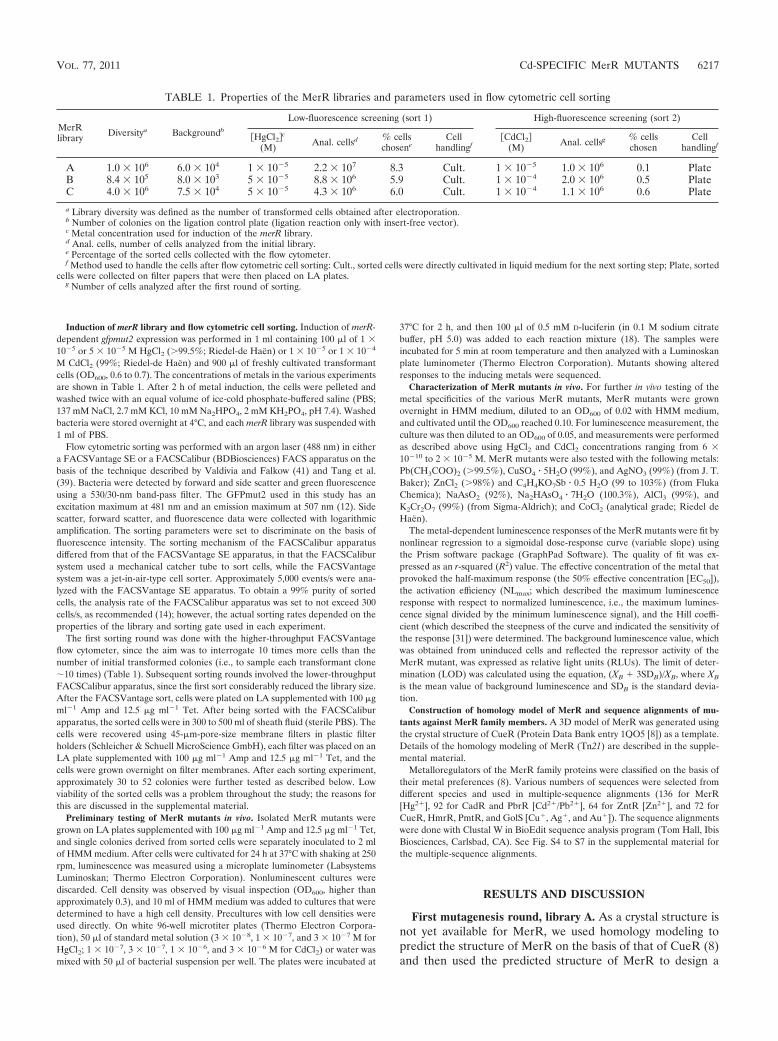
TABLE 1. Properties of the MerR libraries and parameters used in flow cytometric cell sorting
MerRlibrary Diversitya Backgroundb
Low-fluorescence screening (sort 1) High-fluorescence screening (sort 2)
�HgCl2c
(M) Anal. cellsd % cellschosene
Cellhandlingf
�CdCl2(M) Anal. cellsg % cells
chosenCell
handlingf
A 1.0 � 106 6.0 � 104 1 � 10�5 2.2 � 107 8.3 Cult. 1 � 10�5 1.0 � 106 0.1 PlateB 8.4 � 105 8.0 � 103 5 � 10�5 8.8 � 106 5.9 Cult. 1 � 10�4 2.0 � 106 0.5 PlateC 4.0 � 106 7.5 � 104 5 � 10�5 4.3 � 106 6.0 Cult. 1 � 10�4 1.1 � 106 0.6 Plate
a Library diversity was defined as the number of transformed cells obtained after electroporation.b Number of colonies on the ligation control plate (ligation reaction only with insert-free vector).c Metal concentration used for induction of the merR library.d Anal. cells, number of cells analyzed from the initial library.e Percentage of the sorted cells collected with the flow cytometer.f Method used to handle the cells after flow cytometric cell sorting: Cult., sorted cells were directly cultivated in liquid medium for the next sorting step; Plate, sorted
cells were collected on filter papers that were then placed on LA plates.g Number of cells analyzed after the first round of sorting.
VOL. 77, 2011 Cd-SPECIFIC MerR MUTANTS 6217

mutagenesis strategy that avoided the introduction of aminoacid changes that could reduce the functionality of the result-ing mutant library (Fig. 1). Library A was constructed usingrandomized oligonucleotides in which six amino acids (His81,Glu83, Glu84, Ser87, Cys115, and Ala116) were fully random-ized using an NNS scheme (where N is G, A, T, or C and S isG or S) around the metal-binding domain and near Cys82,Cys117, and Cys126, without actually mutating these essentialmetal-binding cysteines (19, 33, 35) (Fig. 1; see Table S2 in thesupplemental material). According to our structural model,these positions were predicted to tolerate mutations withoutdistorting the protein’s tertiary structure. In addition, sevenother amino acids specifically mutated in library A, Ala85,Ser86, Val114, His118, Ala119, Arg120, and Lys121, were po-tentially changed to the amino acids listed in Table S2 in thesupplemental material. The DNA-binding domain was not tar-geted for mutagenesis in any of the constructed MerR mutantlibraries, because two previous studies had indicated that theC-terminal metal-binding domain is essential for the metalselectivity of MerR family proteins (3, 9).
The mutated PCR products of mer were cloned into thescreening vector to construct a system where each MerR mu-tant controlled the expression of GFPmut2 (12) via MerOP. Inthe first selection round, we induced library A with HgCl2 andselected the bacteria representing the lowest 8% of intensity ofthe green fluorescent signals (Table 1; see Fig. S1 in the sup-plemental material). This selection state was intended to ex-clude MerR mutants that were capable of responding toHgCl2. Furthermore, the self-circularizing vector can drive ex-pression of the reporter genes (gfpmut2 and luc) even in theabsence of MerR; however, this background expression ishigher than that seen in the presence of MerR mutants thathave maintained their repressor activities. Therefore, the first-round sorting of MerR mutants was also expected to excludebacteria harboring only insert-free vectors. Sorted cells wereplated, and the expression of the luciferase reporter gene(which was expressed after gfpmut2 was expressed in thescreening vector) was used to screen for Cd-specific pheno-types among the sorted cells (Fig. 2; see Fig. S1 in the supple-mental material). The cells could be divided into five differentphenotypes, namely, MerR mutants having (i) a Cd-specificresponse, (ii) a Cd/Hg response, (iii) an Hg-specific response,(iv) continuous luminescence but no Cd or Hg response, and(v) no luminescence (Fig. 2). Four out of 32 selected MerRmutants showed specific responses toward Cd after the firstround of the low-fluorescence sorting strategy (Fig. 2). Ap-proximately 70% of the sorted cells showed constitutive lumi-nescence, probably because they were not metal inducible andhad repressor activity, which translates to constitutive fluores-cence. Although the activator MerR mutants were excluded bythe first round of negative sorting, the non-metal-induciblerepressor mutants were not (expression data are not shown).
For the second sorting step, the sorted mutants after the firstscreening round were cultivated and induced with CdCl2, andthen the brightest 0.1% of bacteria was enriched. The sortedMerR mutants theoretically either (i) had maintained theirrepressor function and could be transcriptionally activated onlyby CdCl2 or (ii) had maintained their repressor function andcould be transcriptionally activated by CdCl2 and other metals,potentially including HgCl2, at concentrations greater than the
concentration used in the first sorting round. Four out of 60MerR mutants showed Cd-specific responses after completionof the preliminary testing protocol (Fig. 2).
All of the MerR mutants that showed Cd-specific responses(i.e., a Cd-specific response or a Cd/Hg-specific response) inpreliminary testing and had distinctive DNA sequences werethen characterized for their responses to other metal(loid)s(Hg, Cd, Zn, Cu, Ag, Co, Cr, As, Sb, and Al), using subtoxic totoxic metal(loid) concentrations and a luciferase reporter geneassay. None of the tested mutants were completely Cd specific.Instead, they had lower-than-wt responses toward Hg. Onlytwo mutants, A_D23 and A_D24, were more sensitive to Cdthan Hg (Table 2 and Fig. 3). Mutants A_D23 and A_D24 had1,600- and 3,600-fold less sensitive Hg responses, respectively,than the wt, whereas the Cd response remained unchanged inboth mutants. MerR mutant A_D23 also responded weaklytoward antimony and arsenic. In contrast, MerR mutantA_D24 responded only to Hg and Cd. In mutants A_D23 andA_D24, 10 and 13 amino acids, respectively, were changed inand near the metal-binding domain of MerR (Fig. 4).
Second mutagenesis round, libraries B and C. Cd-respon-sive MerR mutants A_D23 and A_D24 were used for a secondmutation round that generated libraries B and C (Fig. 1; seeFig. S1 in the supplemental material). To expand the mutatedregion of A_D24, we used doped oligonucleotides containing79% of the original bases and 7% of each of the other bases forthe construction of library B. Eighteen amino acids were tar-geted in MerR mutant A_D24; they were located in helix 4, inthe loop prior to helix �5, and from Gly122 of the metal-binding loop to the end of helix �6. For the construction of
FIG. 2. Properties of the cells sorted from libraries A, B, and Cafter the first and second rounds of FACS. Sorted cells were prelimi-narily tested with a luciferase assay. The letters given after the name ofthe library (A, B, and C) represent the sorting experiments, D repre-sents the dimmest bacteria that were sorted from Hg-induced cells,and B represents the brightest bacteria that were sorted from Cd-induced cells. The data compare the percentages that each value con-tributed to the total. The total number of tested colonies is given as anumber at the top of each column. The number of mutants sequencedis shown under the bar representing each sorting step. The percentageof mutants having identical sequences is shown in parentheses.
6218 HAKKILA ET AL. APPL. ENVIRON. MICROBIOL.

library C, residues Lys99 and Met106 were fully randomizedusing both MerR mutants A_D23 and A_D24 as parents (Fig.1). The library construction technique allowed formation ofchimeras from A_D23 and A_D24, thereby further increasingthe diversity of library C. Lys99 and Met106 were selected formutagenesis, as they reside in the fifth long alpha helix, whichconstitutes the protomer interface, and these residues havebeen shown to play roles in the Cd response of MerR (7). Theflow cytometric selection and testing strategies used for librar-ies B and C were the same as those described for library A(Table 1; see Fig. S1 in the supplemental material).
The FACS-based dual screening strategy yielded 29% and79% Cd-specific mutants from library B after the first (low-fluorescence) and the second (high-fluorescence) screeningsteps, respectively, and 17% and 31% Cd-specific mutantsfrom library C after the first and the second screening steps,respectively (Fig. 2). Sequencing of the selected mutants re-vealed that some had identical genotypes, indicating that li-brary diversity decreased during the selection process (Fig. 2).
Properties of Cd-specific MerR mutants. Characterizationof the MerR mutants obtained from libraries B and C for theirresponses toward other metal(loid)s revealed that we had gen-
erated eight Cd-specific MerR mutants (Fig. 3). None of theeight Cd-specific mutants responded significantly to any othermetal(loid). Two of the mutants (B_DB10 and B_DB17) hadstrictly Cd-specific responses, while five (B_D20, B_DB4,B_DB26, B_DB27, and B_DB29) showed barely detectableresponses to the highest nontoxic concentration of HgCl2.MerR mutants B_DB26 and C_DB21 also showed weak re-sponses toward high concentrations of ZnCl2 (Fig. 3). TheCd-specific mutants from library B harbored 2 to 9 mutationsin addition to the 13 parental ones in clone A_D24, while thesingle Cd-specific mutant from library C, C_DB21, had 12mutations in total (Fig. 4). None of the Cd-specific mutantsshowed a higher Cd sensitivity than that of wt MerR, perhapssuggesting that the high CdCl2 concentration used for libraryinduction did not favor the selection of more sensitive Cd-responsive MerR mutants. It is also possible that as the Cysresidues were not mutated in MerR and Hg has a strongeraffinity to thiolates than Cd, these could pose difficulties to geta higher affinity for Cd in comparison to the wt with Hg.
To study the repressor activity of the mutants, we con-structed a reporter plasmid with deletion of the MerR regula-tory protein (merR; Table 2 and Fig. 3b) and compared the
TABLE 2. Characteristics of wt MerR and the MerR mutants obtained from the first and second mutagenesis rounds
MerR mutant ormutation round and
MerR mutanta
Metalresponse EC50 (M) NLmax
Background luminescencec
(RLU � SDB)Hill
coefficient R2
wt merR Hg 2.0 � 10�9 18 0.12 � 0.01 (1.3) 2.5 0.930Cd 2.2 � 10�6 18 2.2 0.869
merR Hg/Cd NRd NR 0.53 � 0.11 NR NR
Mutation round 1A_D23 Hgb 3.2 � 10�6 3.7 0.14 � 0.07 (2.5) 1.7e 0.816
Cdb 4.9 � 10�6 7.8 1.2e 0.892Sb 7.9 � 10�5 5.4 3.7 0.941Asb 1.1 � 10�3 2.5 1.0e 0.948
A_D24 Hgb 7.2 � 10�6 6.8 0.20 � 0.13 (3.0) 1.2e 0.839Cd 4.2 � 10�6 18 1.3e 0.810
Mutation round 2B_D20 Hg NDf 1.8 0.20 � 0.03 (1.5) NR NR
Cd 2.7 � 10�6 7.4 1.3e 0.912B_DB4 Hg ND 2.4 0.18 � 0.02 (1.3) ND ND
Cd 3.8 � 10�6 9.0 0.6e 0.942B_DB10 Cd 1.6 � 10�5 10 0.84 � 0.20 (1.7) 1.7e 0.944B_DB17 Cd 3.7 � 10�6 15 0.10 � 0.04 (2.2) 1.6 0.955B_DB26 Hg 1.4 � 10�5 4.2 0.09 � 0.02 (1.7) ND ND
Cd 9.8 � 10�7 20 1.7 0.698Zn 3.2 � 10�5 5.2 15 0.566
B_DB27 Hg ND 1.6 0.11 � 0.01 (1.3) ND NDCd 3.9 � 10�6 16 1.9 0.988
B_DB29 Hg ND 2.6 0.13 � 0.06 (2.4) ND NDCd 2.3 � 10�6 11 1.6 0.882
C_DB21 Cd 2.7 � 10�6 25 0.11 � 0.01 (1.3) 1.3e 0.857Zn 5.0 � 10�5 3.8 4.1 0.734
a In the mutant name, the initial character is the library name and the second and third characters after the underscore indicate the sorting criteria (D, dimmest;B, brightest) through one or two sorting rounds. For example, A_D23 is from library A and was obtained from the first round of collection of the dimmest bacteria.
b Due to metal toxicity, luminescence decreased in the presence of concentrations above 1 � 10�5 M Hg and 1 � 10�4 M Cd, As, or Sb. The EC50 was calculatedon the basis of the highest luminescence observed; we assumed that the curve fitted a nonlinear regression and followed a sigmoidal model.
c The background luminescence value. Values are the mean (XB) from two to nine independent measurements � the standard deviation (SDB). The LOD is shownin parentheses. NLmax values equal to or above the LOD were taken as representing a significant response toward the given metal.
d NR, no response.e The Hill coefficient was extrapolated on the basis of the beginning of the curve.f ND, not determined.
VOL. 77, 2011 Cd-SPECIFIC MerR MUTANTS 6219

uninduced luminescence level from the merR strain withthose from the wt and mutant MerR reporter strains. Theexpression of firefly luciferase from the merR strain was �4-fold higher than that from the wt merR strain, while repressionfrom merR mutant B_DB10 was �7-fold higher than that fromthe wt merR strain (Table 2). All of the other Cd-specificmutants had repressions that ranged from wt levels to a max-imum 1.7-fold decrease in repression.
MerR responds ultrasensitively (a response that rises morerapidly than the concentration increase of the inducer wouldpredict) to its inducing metals both in vivo (11) and in vitro(31). Ultrasensitivity, which is expressed as the Hill coefficient(e.g., it is �2 for wt MerR) (31), results from Hg binding to the
MerR-RNA polymerase complex (1, 31). We found that theHill coefficients were from 0.6 to 1.9 for all of the Cd-respon-sive MerR mutants, potentially indicating that this repressor-to-activator transition is less efficient in the mutants than in wtMerR. Notably, MerR mutants B_DB17, B_DB26, andB_DB27 had the highest Hill coefficients among the mutants(from 1.6 to 1.9) and also showed wt-level repressor activities(background luminescence from 0.09 to 0.11, where that forthe wt was 0.12) and NLmaxs (from 15 to 20, where that for thewt was 18) (Table 2). Mutants B_D20, B_DB4, and C_DB21had the lowest Hill coefficients (from 0.6 to 1.3).
Caguiat et al. (7) identified 11 Cd-responsive MerR mutants,all of which had single or double mutations largely located
FIG. 3. Response curves of wt MerR and the Cd-specific MerR mutants found after the first and second mutation rounds. (a) wt MerR. (b)Vector with wt MerR deleted from the reporter gene plasmid. The curve shows continuous expression of the firefly luciferase gene from MerOP.The MerR mutants obtained after the first mutagenesis round were A_D23 (c) and A_D24 (d), and those obtained after the second mutagenesisround were B_D20 (e), B_DB4 (f), B_DB10 (g), B_DB17 (h), B_DB26 (i), B_DB27 (j), B_DB29 (k), and C_DB21 (l). The responses to HgCl2and CdCl2 are shown for wt MerR and the MerR mutants; the detected responses to other metals are shown, while negative results are omitted.
6220 HAKKILA ET AL. APPL. ENVIRON. MICROBIOL.

close to the metal-binding pocket. None of these identifiedCd-responsive mutants were Cd specific (7) but had retainedthe normal wt Hg responses and were defective in repression,indicating that they had relaxed inducer requirements ratherthan altered metal specificities. The metal sensitivities of theseCd-responsive mutants were not measured. These mutants alsoseemed to have gain-of-function mutations, as their Cd re-sponses were severalfold higher than the wt Cd response. OurA_D24 and B_DB10 mutants showed higher signals toward Cdthan wt MerR (Fig. 3), although the sensitivity was not higherthan that of wt MerR. Therefore, our Cd-specific MerR mu-tants could be classified as loss-of-function mutants, in thattheir mercury sensitivities had been decreased by at least10,000-fold. Comparison of the characteristics between ourmutants and the mutants of Caguiat et al. (7) is relativelydifficult, as there were differences between our protocol andtheir protocol and the experimental procedures; e.g., a differ-ence in cell amounts and/or cultivation medium can affectmetal toxicity. A sequence comparison of the previously re-ported Cd-responsive and Cd-specific MerR mutants (7) withthose reported herein revealed that there was only one muta-tion common to both studies: mutation S125P in B_DB10 ofthe present study (Fig. 1 and 4).
Role of K99 and M106 in Cd-specific response. The Cd-specific mutant C_DB21 arose through the combination ofmutants A_D23 and A_D24 (Fig. 4). In addition, C_DB21 hadadditional mutations in residues K99R and M106L; these mu-tations modified it in the direction of the CadR proteins, whichhave highly conserved Arg and Leu residues in the correspond-ing positions (Table 3; see Fig. S5 in the supplemental mate-rial). The Cd-responsive (but nonspecific) MerR mutantC_DB10 arose from A_D23 and also had additional mutations,K99R and M106L (see Fig. S2 and S3 in the supplementalmaterial). The metal specificity of C_DB10 was not signifi-cantly different from that of the parental mutant, A_D23, ex-cept that A_D23 was 10-fold less sensitive to Sb. This indicatesthat mutations K99R and M106L did not induce significantchanges in the metal specificity of MerR. Our findings furthersuggest that the Cd specificity of C_DB21 was based on theformation of chimeras from A_D23 and A_D24 rather thanmutations K99R and M106L in the dimerization helix.
The other Cd-responsive MerR mutant, C_DB36, was de-scended from A_D24 and had the additional mutations K99E
and M106F (see Fig. S2 and S3 in the supplemental material)in the dimerization helix. The metal response of this mutantdiffered from that of parental strain A_D24 as follows:C_DB36 was repressor deficient, was at least 10-fold less sen-sitive toward Hg, and, additionally, responded to Sb and As. Along helix (�5), the so-called coiled coil region of MerR, formsfive heptads, each with a, b, c, d, e, f, and g positions indicatingamino acid locations; notably, K99 and M106 are both in dpositions (36). In the study of Caguiat et al. (7), 4 of 13randomly selected or screened Cd(II)-responsive MerR mu-tants had mutations of K99 or M106. On the basis of this, theauthors suggested that interface positions could be involved inthe metal-specific response of MerR. Furthermore, it has beenobserved that in natural coiled coils having hydrophobic resi-dues, the d position forms the helix interface between twosubunits, and the presence of polar and charged residues deepwithin the hydrophobic core of coiled coils can affect a pro-tein’s behavior (36). In the model of MerR, K99 and K99� arefound in very close proximity to one another (Fig. 1b), andpolar substitutions of these residues have been associated withdecreased repressor activity in vivo (7). It has also been pro-posed that during metal-binding-provoked movement, M106interacts with the L63� residue of the other protomer’s helix�4; this could cause a shift toward the DNA-binding domain.In the absence of metal, in contrast, M106 may stabilize DNAbinding (36). Therefore, the two moderately radical substitu-tions (K99E and M106F) in C_DB36 could affect the protein’sDNA-binding properties and metal specificity in a mannerunlike that seen for C_DB21 and C_DB10, which had lessradical mutations (K99R and M106L).
Fine adjustment of Cd selectivity in MerR mutants. Cd-specific mutant B_DB17 (Fig. 3) had 18 amino acid changes(Fig. 4). Mutant B_DB34 had the same amino acid changes,with the exception of the Leu-to-Phe mutation at residue 73(see Fig. S3 in the supplemental material), which is situated inthe coupling domain at the end of helix �4. The metal re-sponses of these mutants were different. B_DB34 was Cd re-sponsive but not Cd specific, as it strongly responded to Hg andnoticeably responded to several other metals (Sb, Zn, Cr, andAs) (see Fig. S2 in the supplemental material). In addition, itwas repressor deficient, giving a clearly higher backgroundsignal than B_DB17 in the absence of metal. This shows that asingle relatively conserved type of amino acid change could
FIG. 4. Sequencing results from the MerR mutants beginning at residue L67. In the sequence of wt MerR, helices �4, �5, and �6 areunderlined. The metal-binding cysteines are marked with black arrows above the wt MerR sequence.
VOL. 77, 2011 Cd-SPECIFIC MerR MUTANTS 6221

have a significant effect on the MerR metal response. Mecha-nistically, it is notable that the side chain of residue 73 is buriedwithin the metal coordination site, meaning that the structureof the coordination site is likely to be affected by the replace-
ment of Leu with the larger Phe residue. It is tempting tospeculate that the properties of the metals in question could beresponsible for the mutation-induced change in the metal re-sponse, as the ionic radius of Cd(II) is 103 pm, while that of
TABLE 3. Sequence alignments between Cd-specific MerR mutants and MerR family members presented in the supplemental material
Mutation or region and mutationa Mutant(s)
% of mutations in sequence alignment
MerR(136)b
CadR/PbrR(92)
ZntR(64)
CueR/HmrR/PmtR/GolS (72)
E72A B_DB4 7.4 32.6 0.0 5.4E72D B_DB17/26 5.9 0.0 6.3 16.2L73I B_DB10 0.0 0.0 0.0 0.0L73F B_DB17 0.0 0.0 1.6 4.1L74W B_DB4 0.0 0.0 0.0 0.0R75W B_DB4/10 0.0 0.0 0.0 0.0L76I B_DB17/27/29 2.2 0.0 96.9 0.0L76V B_DB10 12.5 1.1 0.0 0.0
Loop from residues D77 to H81c
D77E B_D20/D_DB27 28.7D77C B_DB4 0.0D77Y B_DB10 0.0D78E B_DB17 5.9G79Y B_D20 0.0T80S B_DB4 5.1T80N B_DB26 0.0T80A B_DB29 10.3H81G All 1.5E83V All 0.7 0.0 1.6 1.4E84G All 1.5 13.0 0.0 0.0A85H All 0.0 0.0 0.0 0.0S86D All 0.0 0.0 0.0 0.0S87A All 14.7 48.9 31.3 45.9K99R C_DB21 14.0 95.7 20.3 55.4M106L C_DB21 16.9 100.0 1.6 16.2V114T B_D20, B_DB4/10/17/26/27/29 1.5 0.0 1.6 0.0C115G B_D20, B_DB4/10/17/26/27/29 3.7 2.2 0.0 1.4C115Q C_DB21 14.7 16.3 3.1 4.1A116M B_D20, B_DB4/10/17/26/27/29 0.0 0.0 0.0 0.0A116P C_DB21 0.0 0.0 0.0 0.0
Metal-binding loop fromresidues H118 to S125d
H118N All B mutants 2.9A119L All B mutants 1.5A119P C_DB21 0.0R120P All B mutants 3.7K121P All 4.4G122A B_D20 4.4G122R B_DB4 2.2N123K B_DB10 2.2N123Y B_DB29 0.0V124F B_D20 0.7V124Y B_DB27 6.6S125F B_DB4 0.0S125P B_DB10 0.0L128M B_D20 3.7 0.0 0.0 0.0L128F B_DB4 0.0 0.0 0.0 0.0I129V B_DB4 2.9 22.8 0.0 0.0A130R B_DB17 0.0 7.6 0.0 4.1A130E B_DB27 21.3 5.4 71.9 18.9S131T B_D20 19.1 0.0 7.8 4.1
a Numbering of the residues is based on that of MerR from Tn21. Highly conserved amino acids of MerR proteins are highlighted in boldface.b Data in parentheses are the number of sequences used in the multiple-sequence alignment.c The loop lengths of the MerR-like proteins were from 5 to 7 residues. CadR/PbrR, ZntR, and CueR/HmrR/PmtR/GolS were 2, 3, and 2 residues longer than MerR
(Tn21), respectively. CadR/PbrR and CueR/HmrR/PmtR/GolS loop sequences contained conserved P and R, respectively. After the loop of CueR/HmrR/PmtR/GolS,there is an S instead of metal-binding C82. H and P residues were present in the loops of ZntR-like proteins.
d The motifs in the metal-binding loop were CysX8Cys for MerR, CysX5–9Cys for CadR/PbrR, CysX9Cys (which has C and H residues involved in Zn binding) forZntR, and CysXGlyX4Asp/GluCys (in which there is a high degree of conservation of the D following the G) for CueR/HmrR/PmtR/GolS.
6222 HAKKILA ET AL. APPL. ENVIRON. MICROBIOL.

Hg(II) is 112 pm. Alternatively, the altered metal specificitycould be related to the coordination preferences of the metals,with Cd(II) being more stably bound by the mutants.
Since protein tertiary structure preferences dictate the finalgeometry of a bound metal (16, 30), it seems logical to surmisethat Cd-specific MerR mutants having a large number ofamino acid changes could coordinate Cd(II) differently than wtMerR, with some of the mutations being responsible for mak-ing small, Hg-excluding changes to the metal-binding pocket. Itis also possible that the coordination of a metal and subsequentspecific conformational change or steric hindrance are respon-sible for metal specificity, as concluded by Rensing (32). Thebasis for selective metal ion recognition by MerR, which bindsHg highly selectively over Cd or Zn, is not yet fully understood(31), even though we do know the essential role of cysteines inHg binding (19, 33, 35). It is also noteworthy that metal ionrecognition preferences differ even between two Cd-responsiveCadR proteins from Pseudomonas putida and Pseudomonasaeruginosa (at its cognate promoter PcadA) with the inductionprofiles Cd �� Pb � Zn and Cd �� Zn � Hg, respectively (4,24). Giedroc and Arunkumar (17) have suggested that rela-tively subtle modifications in the metal-binding pocket couldlead to the evolution of MerR family regulatory proteins, e.g.,CadR and PbrR, which preferentially detect Cd and Pb overZn, respectively, or MerR, which preferentially detects Hgover Cd. The X-ray crystallographic structures of two MerRfamily members, CueR and ZntR (3, 8, 29), indicate that themetal receptor sites are buried in a loop at the dimer interface,which holds the key factors for metal ion selectivity. The mech-anisms of metal coordination can differ among the variousmembers of the family. For example, ZntR binds four zincatoms per dimer, and each Zn(II) atom is bound in a tetrahe-dral coordination environment (8, 28). In contrast, CueR bindstwo Cu(I) or Ag(I) ions and forms linear two-coordinate ge-ometries in both cases (8), whereas Hg(II)-MerR forms anunusual tricoordinated complex (40), even though the pre-ferred coordination geometry for Hg(II) is two ligands. Cd(II)and Zn(II) compete for MerR binding, and they may eitherform a trigonal thiolate complex or adopt higher coordinations(40). Unfortunately, the coordination geometries of Cd(II) arenot yet known for wt MerR. In the future, it could prove veryuseful to examine the coordination environment of all theCd-specific mutants described in this work.
Evolutionary aspects of Cd-specific MerR mutants. In orderto determine whether the mutations of our Cd-specific MerRvariants exist in nature, the amino acids from Asp68 to the endof the protein were aligned with the equivalent sequences ofMerR family proteins from different species: MerR, CadR/PbrR, ZntR, and CueR/HmrR/PmtR/GolS (see Fig. S4 to S7in the supplemental material). Table 3 shows an amino acidsequence comparison of our Cd-specific mutants against thevarious MerR family proteins. This analysis excluded the loopstructures, which were compared only among the MerR-likeproteins, as the loop lengths differed significantly among theother MerR family proteins. Amino acid changes were classi-fied into three categories: new mutations, where a mutation didnot exist in any other of the aligned sequences; low-frequencymutations, where a mutation was found in less than 10% ofsequences; and existing mutations, where a mutation wasfound at the aligned position in more than 10% of the se-
quences. Mutations found from the Cd-specific MerR mutantsharbored 37% new mutations compared to MerR proteins and53 to 58% compared to other family members. CadR/PbrRproteins had 73% and the other family members had 80 to 85%of mutations that were new or low-frequency mutations. Themutations were largely derived from parental sequence A_D24(nine in library B and five in library C), as library A was quiterandomly mutated. All of the Cd-specific mutants from librar-ies B and C also had one to five additional new mutations intheir sequences in comparison to MerR proteins. Further-more, it is interesting that the largest amount of mutations(27% were existing mutations) occurred in the aligned se-quences of CadR/PbrR proteins compared to other familymembers (15 to 18%). Thus, such changes can favor a Cd-specific response. Also, our analysis of these mutations re-vealed that the structure of MerR appears to tolerate radicalamino acid changes fairly well, especially near and within themetal-binding loop structures.
Among the conserved amino acids of MerR, we targeted 10for mutagenesis. The total number of amino acids changes (18to 22) was higher in the Cd-specific mutants that had mutationsin 4 or 5 of the conserved amino acids (B_D20, B_DB4,B_DB10, and B_DB17) than in those having mutations in only2 or 3 of the conserved amino acids (B_DB26, B_D27,B_DB29, and C_DB21, which had 12 to 17 amino acidchanges) (Table 3). This suggests that the conserved residuesare important to the structure of MerR, and additional muta-tions were needed to compensate for the structural and func-tional changes induced by mutations in these conserved resi-dues. We also found that the amino acid residues after the last� helix (�6) did not seem to be critical for the functionality ofMerR proteins, as Cd-specific mutant B_DB4 had a stop codonat position Ser131. Our sequence alignment also supportedthis observation, as many of the MerR-like proteins had tails ofonly one to three amino acids after helix �6 (see Fig. S4 in thesupplemental material).
We believe that the Cd-specific MerR mutants describedherein are unique among the MerR family members charac-terized to date. There appear to be few similarities in the loopstructures of the Cd-specific mutants and other MerR familyproteins, and the involved amino acid residues vary widely.However, the Cd-specific mutants and the other MerR familymetalloregulatory proteins do have one interesting feature incommon: they all have a Pro(s) in the two loop structures ofthe binding region, whereas parental MerR (Tn21) and severalother MerR proteins do not. Our Cd-specific MerR mutantshad two to three Pro residues in their metal-binding loops.Similarly, the CadR proteins have a highly conserved Pro in theloop of the N-terminal end of the dimerization helix or in themetal-binding loop (Table 3; see Fig. S5 in the supplementalmaterial), while ZntR and CueR each have a Pro in one oftheir loop structures. Because Pro has a five-memberedpyrrolidine ring that connects the � carbon to the preceding amidenitrogen, it imposes rigid constraints on the conformation ofthe peptide backbone. The introduction of two to three Proresidues within an eight-residue metal-binding loop couldtherefore conceivably cause significant changes in loop confor-mation and dynamics, resulting in a mutant with altered metal-binding properties (i.e., Cd specificity). The metal-binding loop(CysX8Cys) of the MerR proteins is followed by a proline
VOL. 77, 2011 Cd-SPECIFIC MerR MUTANTS 6223

(Pro127) instead of the Gly (or Ala) seen in the CadR proteins(see Fig. S4 and S5 in supplemental material). Interestingly,even though this Pro was targeted by our mutagenesis proce-dure, none of the Cd-specific MerR mutants had mutations inthis residue. This, along with its conserved nature, indicatesthat this particular Pro is likely to play an important role in themetal-binding functionality of MerR.
ACKNOWLEDGMENTS
The work was financially supported by the Academy of Finland(grants 201677 and 207258 to Marko P. Virta).
We are grateful for the assistance of P. Terho and M. Korkeamaki(Cell Imagining Core facility of Turku). We also thank Taina Tyystjarvifor critical reading of the manuscript.
REFERENCES
1. Ansari, A. Z., M. L. Chael, and T. V. O’Halloran. 1992. Allosteric under-winding of DNA is a critical step in positive control of transcription byHg-MerR. Nature 355:87–89.
2. Barrineau, P., et al. 1984. The DNA sequence of the mercury resistanceoperon of the IncFII plasmid NR1. J. Mol. Appl. Genet. 2:601–619.
3. Brocklehurst, K. R., et al. 1999. ZntR is a Zn(II)-responsive MerR-liketranscriptional regulator of zntA in Escherichia coli. Mol. Microbiol. 31:893–902.
4. Brocklehurst, K. R., S. J. Megit, and A. P. Morby. 2003. Characterisation ofCadR from Pseudomonas aeruginosa: a Cd(II)-responsive MerR homologue.Biochem. Biophys. Res. Commun. 308:234–239.
5. Brown, N. L., S. J. Ford, R. D. Pridmore, and D. C. Fritzinger. 1983.Nucleotide-sequence of a gene from the Pseudomonas transposon Tn501encoding mercuric reductase. Biochemistry 22:4089–4095.
6. Brown, N. L., J. V. Stoyanov, S. P. Kidd, and J. L. Hobman. 2003. The MerRfamily of transcriptional regulators. FEMS Microbiol. Rev. 27:145–163.
7. Caguiat, J. J., A. L. Watson, and A. O. Summers. 1999. Cd(II)-responsiveand constitutive mutants implicate a novel domain in MerR. J. Bacteriol.181:3462–3471.
8. Changela, A., et al. 2003. Molecular basis of metal-ion selectivity andzeptomolar sensitivity by CueR. Science 301:1383–1387.
9. Checa, S. K., et al. 2007. Bacterial sensing of and resistance to gold salts.Mol. Microbiol. 63:1307–1318.
10. Chen, P. R., and C. He. 2008. Selective recognition of metal ions by metal-loregulatory proteins. Curr. Opin. Chem. Biol. 12:214–221.
11. Condee, C. W., and A. O. Summers. 1992. A mer-lux transcriptional fusionfor real-time examination of in vivo gene expression kinetics and promoterresponse to altered superhelicity. J. Bacteriol. 174:8094–8101.
12. Cormack, B. P., R. H. Valdivia, and S. Falkow. 1996. FACS-optimizedmutants of the green fluorescent protein (GFP). Gene 173:33–38.
13. deWet, J. R., K. W. Wood, and M. De Luca. 1985. Cloning of firefly luciferasecDNA and the expression of active luciferse in Escherichia coli. Proc. Natl.Acad. Sci. U. S. A. 80:7870–7873.
14. Ferrari, B. C., G. Oregaard, and S. J. Sorensen. 2004. Recovery of GFP-labeled bacteria for culturing and molecular analysis after cell sorting usinga benchtop flow cytometer. Microb. Ecol. 48:239–245.
15. Franz, B., and T. V. O. O’Halloran. 1990. DNA distortion accompaniestranscriptional activation by the metal-responsive gene-regulatory proteinMerR. Biochemistry 29:4747–4751.
16. Ghosh, D., K. H. Lee, B. Demeler, and V. L. Pecoraro. 2005. Linear free-energy analysis of mercury(II) and cadmium(II) binding to three-strandedcoiled coils. Biochemistry 44:10732–10740.
17. Giedroc, D. P., and A. I. Arunkumar. 2007. Metal sensor proteins: nature’smetalloregulated allosteric switches. Dalton Trans. 29:3107–3120.
18. Hakkila, K., M. Maksimow, M. Karp, and M. Virta. 2002. Reporter geneslucFF, luxCDABE, gfp, and dsred have different characteristics in whole-cellbacterial sensors. Anal. Biochem. 301:235–242.
19. Helmann, J. D., B. T. Ballard, and C. T. Walsh. 1990. The MerR metallo-regulatory protein binds mercuric ion as a tricoordinate, metal-bridgeddimer. Science 247:946–948.
20. Heltzel, A., D. Gambill, W. J. Jackson, P. A. Totis, and A. O. Summers. 1987.Overexpression and DNA-binding properties of the mer-encoded regulatoryprotein from plasmid NR1 (Tn21). J. Bacteriol. 169:3379–3384.
21. Hobman, J. L. 2007. MerR family transcription activators: similar designs,different specificities. Mol. Microbiol. 63:1275–1278.
22. Hobman, J. L., J. Wilkie, and N. L. Brown. 2005. A design for life: prokary-otic metal-binding MerR family regulators. Biometals 18:429–436.
23. LaRossa, R. A., D. R. Smulski, and T. K. Van Dyk. 1995. Interaction of leadnitrate and cadmium chloride with Escherichia coli K-12 and Salmonellatyphimurium global regulatory mutants. J. Ind. Microbiol. 14:252–258.
24. Lee, S. W., E. Glickmann, and D. A. Cooksey. 2001. Chromosomal locus forcadmium resistance in Pseudomonas putida consisting of a cadmium-trans-porting ATPase and a MerR family response regulator. Appl. Environ.Microbiol. 67:1437–1444.
25. Lovell, T., F. Himo, W. Han, and L. Noodleman. 2003. Density functionalmethods applied to metalloenzymes. Coordin. Chem. Rev. 238:211–232.
26. Misra, T. K., et al. 1984. Mercuric ion-resistance operons of plasmid R100and transposon Tn501: the beginning of the operon including the regulatoryregion and the first two structural genes. Proc. Natl. Acad. Sci. U. S. A.81:5975–5979.
27. O’Halloran, T. V., B. Frantz, M. K. Shin, D. M. Ralston, and J. G. Wright.1989. The MerR heavy metal receptor mediates positive activation in atopologically novel transcription complex. Cell 56:119–129.
28. Outten, C. E., F. W. Outten, and T. V. O’Halloran. 1999. DNA distortionmechanism for transcriptional activation by ZntR, a Zn(II)-responsive MerRhomologue in Escherichia coli. J. Biol. Chem. 274:37517–37524.
29. Outten, F. W., C. E. Outten, J. Hale, and T. V. O’Halloran. 2000. Transcrip-tional activation of an Escherichia coli copper efflux regulon by the chromo-somal MerR homologue, CueR. J. Biol. Chem. 275:31024–31029.
30. Pennella, M. A., and D. P. Giedroc. 2005. Structural determinants of metalselectivity in prokaryotic metal-responsive transcriptional regulators. Bio-metals 18:413–428.
31. Ralston, D. M., and T. V. O’Halloran. 1990. Ultrasensitivity and heavy-metalselectivity of the allosterically modulated MerR transcription complex. Proc.Natl. Acad. Sci. U. S. A. 87:3846–3850.
32. Rensing, C. 2005. Form and function in metal-dependent transcriptionalregulation: dawn of the enlightenment. J. Bacteriol. 187:3909–3912.
33. Ross, W., S. J. Park, and A. O. Summers. 1989. Genetic analysis of tran-scriptional activation and repression in the Tn21 mer operon. J. Bacteriol.171:4009–4018.
34. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, NY.
35. Shewchuk, L. M., et al. 1989. Transcriptional switching by the MerR protein:activation and repression mutants implicate distinct DNA and mercury(II)binding domains. Biochemistry 28:2340–2344.
36. Song, L. Y., Q. Teng, R. S. Phillips, J. M. Brewer, and A. O. Summers. 2007.19F-NMR reveals metal and operator-induced allostery in MerR. J. Mol.Biol. 371:79–92.
37. Stanisich, V. A., P. M. Bennett, and M. H. Richmond. 1977. Characterizationof a translocation unit encoding resistance to mercuric ions that occurs on anon conjugative plasmid in Pseudomonas aeruginosa. J. Bacteriol. 129:1227–1233.
38. Summers, A. O. 1992. Untwist and shout: a heavy metal-responsive tran-scriptional regulator. J. Bacteriol. 174:3097–3101.
39. Tang, S.-Y., H. Fazelinia, and P. C. Cirino. 2008. AraC regulatory proteinmutants with altered effector specificity. J. Am. Chem. Soc. 130:5267–5271.
40. Utschig, L. M., J. W. Bryson, and T. V. O’Halloran. 1995. Mercury-199 NMRof the metal receptor site in MerR and its protein-DNA complex. Science268:380–385.
41. Valdivia, R. H., and S. Falkow. 1996. Bacterial genetics by flow cytometry:rapid isolation of Salmonella typhimurium acid-inducible promoters by dif-ferential fluorescence induction. Mol. Microbiol. 22:367–378.
42. Wright, J. G., H. T. Tsang, J. E. Pennerhahn, and T. V. Ohalloran. 1990.Coordination chemistry of the Hg-MerR metalloregulatory protein: evi-dence for a novel tridentate Hg-cysteine receptor site. J. Am. Chem. Soc.112:2434–2435.
43. Zeng, Q., C. Stalhandske, M. C. Anderson, R. A. Scott, and A. O. Summers.1998. The core metal-recognition domain of MerR. Biochemistry 37:15885–15895.
6224 HAKKILA ET AL. APPL. ENVIRON. MICROBIOL.
Related Documents











