ISSN: 1524-4563 Copyright © 2009 American Heart Association. All rights reserved. Print ISSN: 0194-911X. Online 72514 Hypertension is published by the American Heart Association. 7272 Greenville Avenue, Dallas, TX DOI: 10.1161/HYPERTENSIONAHA.109.134338 published online Sep 14, 2009; Hypertension Robin A. Felder John J. Gildea, Jonathan A. Israel, Andrew K. Johnson, Jin Zhang, Pedro A. Jose and Renal Proximal Tubule Cells Caveolin-1 and Dopamine-Mediated Internalization of NaKATPase in Human http://hyper.ahajournals.org/cgi/content/full/HYPERTENSIONAHA.109.134338/DC1 Data Supplement (unedited) at: http://hyper.ahajournals.org located on the World Wide Web at: The online version of this article, along with updated information and services, is http://www.lww.com/reprints Reprints: Information about reprints can be found online at [email protected] 410-528-8550. E-mail: Fax: Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters http://hyper.ahajournals.org/subscriptions/ Subscriptions: Information about subscribing to Hypertension is online at by on May 11, 2011 hyper.ahajournals.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ISSN: 1524-4563 Copyright © 2009 American Heart Association. All rights reserved. Print ISSN: 0194-911X. Online
72514Hypertension is published by the American Heart Association. 7272 Greenville Avenue, Dallas, TX
DOI: 10.1161/HYPERTENSIONAHA.109.134338 published online Sep 14, 2009; HypertensionRobin A. Felder
John J. Gildea, Jonathan A. Israel, Andrew K. Johnson, Jin Zhang, Pedro A. Jose and Renal Proximal Tubule Cells
Caveolin-1 and Dopamine-Mediated Internalization of NaKATPase in Human
http://hyper.ahajournals.org/cgi/content/full/HYPERTENSIONAHA.109.134338/DC1Data Supplement (unedited) at:
http://hyper.ahajournals.org
located on the World Wide Web at: The online version of this article, along with updated information and services, is
http://www.lww.com/reprintsReprints: Information about reprints can be found online at
[email protected]. E-mail:
Fax:Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters
http://hyper.ahajournals.org/subscriptions/Subscriptions: Information about subscribing to Hypertension is online at
by on May 11, 2011 hyper.ahajournals.orgDownloaded from

Caveolin-1 and Dopamine-Mediated Internalization ofNaKATPase in Human Renal Proximal Tubule Cells
John J. Gildea, Jonathan A. Israel, Andrew K. Johnson, Jin Zhang, Pedro A. Jose, Robin A. Felder
Abstract—In moderate sodium-replete states, dopamine 1–like receptors (D1R/D5R) are responsible for regulating �50%of renal sodium excretion. This is partly mediated by internalization and inactivation of NaKATPase, when associatedwith adapter protein 2. We used dopaminergic stimulation via fenoldopam (D1-like receptor agonist) to study theinteraction among D1-like receptors, caveolin-1 (CAV1), and the G protein–coupled receptor kinase type 4 in culturedhuman renal proximal tubule cells (RPTCs). We compared 2 groups of RPTCs, 1 of cell lines that were isolated fromnormal subjects (nRPTCs) and a second group of cell lines that have D1-like receptors that are uncoupled (uncoupledRPTCs) from adenylyl cyclase second messengers. In nRPTCs, fenoldopam increased the plasma membrane expressionof D1R (10.0-fold) and CAV1 (1.3-fold) and markedly decreased G protein–coupled receptor kinase type 4 by 94�8%;no effects were seen in uncoupled RPTCs. Fenoldopam also increased the association of adapter protein 2 andNaKATPase by 53�9% in nRPTCs but not in uncoupled RPTCs. When CAV1 expression was reduced by 86.0�8.5%using small interfering RNA, restimulation of the D1-like receptors with fenoldopam in nRPTCs resulted in only a7�9% increase in association between adapter protein 2 and NaKATPase. Basal CAV1 expression and association withG protein–coupled receptor kinase type 4 was decreased in uncoupled RPTCs (58�5% decrease in association) relativeto nRPTCs. We conclude that the scaffolding protein CAV1 is necessary for the association of D1-like receptors withG protein–coupled receptor kinase type 4 and the adapter protein 2–associated reduction in plasma membraneNaKATPase. (Hypertension. 2009;54:00-00.)
Key Words: dopamine receptors � caveolin � proximal tubule � hypertension � GRK4 � NaKATPase � FRET
The mechanisms by which renal dopamine, in concert withother sodium regulator pathways, regulates �50% of
renal sodium excretion are not completely understood. Intra-renal administration of a dopamine 1 (D1)-like receptorantagonist (SCH23390) decreased sodium excretion by 57%in conscious uninephrectomized dogs1,2 and in anesthetizedvolume-loaded rats.3,4 Disruption of D1-like receptor (D1Ror D5R) genes in mice increases salt-sensitive bloodpressure.5 Dopamine and D1-like receptor agonists arenatriuretic in experimental animals6 and humans,7,8 andecopipam, a selective D1-like receptor antagonist, in-creases blood pressure in humans9 (presumably by pre-venting sodium excretion).
Dopamine inhibits sodium transport in several segments ofthe nephron, causing increased sodium excretion.5 In the renalproximal tubule (RPT), dopamine inhibits the sodium/hydro-gen exchanger 3 (NHE3)10,11 and NaKATPase.12,13 The de-crease in sodium/hydrogen exchanger 310 and NaKATPase14
from the plasma membrane involves binding to adapterprotein 2 (AP2), followed by endocytosis. Dopaminestimulation and/or sodium concentration associate AP2with NaKATPase, cause NaKATPase translocation from the
plasma membrane into early and late endosomes, and de-crease rubidium uptake, an index of NaKATPase activity inopossum and rodent cells.15–18 However, the effect of dopa-mine on NaKATPase internalization, activity, and associationwith AP2 has not been shown in human RPT cells (RPTCs).There may be species differences in the regulation of sodiumtransport. For example, in humans, the � subunit of NaKATPaseexpressed in the kidney is more sensitive to the inhibition byouabain compared with rodents.19,20
Caveolin, especially caveolin-1 (CAV1), has been shownto be important in the NaKATPase internalization pro-cess.21,22 Caveolins, localized in lipid rafts of plasma mem-branes, tether and regulate signaling complexes into func-tional units (eg, G protein–coupled receptors).23,24 We havereported that fenoldopam (FEN; D1-like receptor agonist)stimulates caveolin 2 (CAV2) and D1R association in humanembryonic kidney 293 cells, which do not express CAV1.25
Furthermore, FEN-stimulated membrane D1R recruitmentand cAMP accumulation were greater in membranes withCAV2-containing lipid rafts than in those depleted of CAV2with antisense oligonucleotides or in nonlipid rafts withoutCAV2.26
Received April 14, 2009; first decision May 8, 2009; revision accepted August 18, 2009.From the Department of Pathology (J.J.G., J.A.I., A.K.J., R.A.F.), University of Virginia Health System, Charlottesville, Va; Children’s National
Medical Center (P.A.J.), Center for Molecular Physiology, Department of Pediatrics, George Washington University, Washington, DC; Department ofPharmacology and Molecular Sciences, Johns Hopkins School of Medicine (J.Z.), Baltimore, Md.
Correspondence to Robin A. Felder, University of Virginia, PO Box 801400, Charlottesville, VA 22908. E-mail [email protected]© 2009 American Heart Association, Inc.
Hypertension is available at http://hyper.ahajournals.org DOI: 10.1161/HYPERTENSIONAHA.109.134338
1 by on May 11, 2011 hyper.ahajournals.orgDownloaded from

G protein–coupled receptor kinase 4 (GRK4) is 1 of 7members of the GRK family and has been shown to bespecifically located in RPTCs, where it constitutively phos-phorylates and desensitizes D1R in specific plasma membranemicrodomains.27,28 However, the role of the interactionamong GRK4, CAV1, and D1-like receptors and NaKATPaseinternalization in human RPTCs has not been described. Wetested the hypothesis that CAV1, interacting with GRK4, isnecessary for D1R-mediated inhibition of NaKATPaseactivity in human RPTCs through binding with AP2.Because D1R signaling is impaired in hypertensive humansand rats,6,8,23,29,30 this interaction may be aberrant inessential hypertension.
Materials and MethodsPlease see the online Data Supplement at http://hyper.ahajournals.org for the Expanded Methods section.
Cell LinesWe have generated RPTCs from institutional review board–approved normal human subjects (nRPTCs) and RPTCs fromsubjects in which D1R is uncoupled from adenylyl cyclase stimula-tion (uRPTCs)28,31–33 (see details in the online Data SupplementMethods section), as described previously.27,28,31,34–36
Determination of cAMP AccumulationcAMP was measured both by a commercial ELISA (CaymanChemical) and an intracellular real-time kinetic assay, which in-volves transfecting RPTCs with a plasmid containing a novelfluorescence resonance energy transfer (FRET) cAMP sensor(ICUE3) according to the method of Violin et al.37 Details of bothmethods are outlined in the online Data Supplement.
Measurement of Plasma Membrane and Total CellProtein ExpressionDetergent-free plasma membrane sheet isolation was performedusing sulfo-NHS-SS-biotin, as reported previously.35 Total celllysates (20 �g; 4°C) were loaded per lane for electrophoresis andimmunoblotting.35 The proteins of interest were detected using rabbitpolyclonal antibodies to CAV1 (1:500 dilution; BD Biosciences),D1R,26 and GRK4 (1:200 dilution; Santa Cruz; sc-13079), followedby a goat antirabbit infrared dye (IR Dye 800; Li-Cor) secondaryantibody and imaged on an Odyssey infrared imaging system (Li-Cor).
Measurement of the Association of GRK4and CAV1GRK4 and CAV1 association was measured by coimmunoprecipi-tation of 1 mg of cellular protein in 1 mL of lysis buffer (M-PER;Pierce) and 2 �g of CAV1 monoclonal antibody (BD Biosciences).Detection used rabbit polyclonal anti-GRK4 (1:200 dilution; SantaCruz; sc-13079), followed by goat antirabbit IR Dye 800 secondaryantibody. Details are provided in the online Data Supplement.
Small Interfering RNA to CAV1 and GRK4CAV1 small interfering RNA (siRNA; target sequence: 5�CCGCAT-CAACTTGCAGAAA3� and scrambled control: 5�CCGAACTGT-TCGACACAAA3�)38 was designed and ordered prehybridized(Sigma-Genosys). The most effective GRK4 siRNA (target se-quence: 5�AATACAAAGAGAAAGTCAA3� and scrambled con-trol: 5�AGAAGATAAGAACAATAAC3�) was chosen among 10candidate target sequences by Western blotting. Details of transfec-tion are provided in the online Data Supplement.
NaKATPase Internalization AssayMeasurement of NaKATPase internalization with AP2 was adaptedfrom previously published techniques,15,13 which showed that the
internalization of and decrease in NaKATPase activity are associatedwith AP2. Monensin was shown to be necessary to measure theeffect of dopamine.17,18 Details are provided in the online DataSupplement.
NaKATPase Confocal Microscopy and PlasmaMembrane LocalizationPlasma membrane–localized NaKATPase expression was performedon RPTCs using similar treatments as for the NaKATPase experi-ments, with subsequent fixation and staining with a monoclonalantibody to NaKATPase. Details are outlined in the online DataSupplement.
NaKATPase ActivityWe have established a sodium efflux assay that measures NaKATPaseactivity in cultured cells as ouabain-sensitive reduction in sodiumexport. Details are included in the online Data Supplement.
Statistical AnalysisThe data are expressed as mean�SE. Comparisons within and amonggroups were made by repeated-measures or factorial ANOVA, respec-tively, followed by Holm-Sidak or Duncan test. A t test was used for2-group comparisons. A P�0.05 was considered significant.
ResultsOur cell lines were well characterized for their RPT ori-gin.27,28 We measured cAMP in 2 nRPTC and 2 uRPTC celllines, determined by the response to fenoldopam (FEN;1 �mol/L; 30 minutes) or dimethyl sulfoxide (DMSO)vehicle control. We compared our ELISA with a novelFRET–based method (Figure 1). Figure 1A shows the �2-fold increase in cAMP accumulation (ELISA) induced byFEN in nRPTCs (but not in uRPTCs) that was blocked by theD1-like receptor antagonist, LE300 (10 �mol/L), indicatingthat the stimulatory effect was via D1-like receptors. A cAMPFRET biosensor, ICUE3,37 showed that D1-like stimulationwith FEN caused a significant rise in the intracellular cAMPlevel in nRPTCs, which reached a plateau by 7 minutes(Figure 1B). Minimal effect was seen in uRPTCs, confirmingour previous reports with older methods.28,32
We next studied the plasma membrane expression of D1R,GRK4, and CAV1 in nRPTCs and uRPTCs using Westernblotting techniques (Figure 2). Figure 2A shows that D1Rplasma membrane recruitment was increased (10.73�1.70-fold; P�0.05; n�4) over DMSO vehicle (VEH) control afterFEN (1 �mol/L; 30 minutes) stimulation in nRPTCs. InuRPTCs, there was no significant increase in FEN-inducedD1R recruitment to the plasma membrane. Basal levels ofplasma membrane GRK4 expression were lower in uRPTCsthan in nRPTCs (58.4�8% decrease; P�0.05; n�4). Figure2B shows that FEN stimulation decreased plasma membraneGRK4 abundance in nRPTCs (94�8.0; P�0.05; n�4) buthad no effect in uRPTCs. CAV1 basal plasma membraneexpression was 40.0% (�7.0%; P�0.05; n�4) lower inuRPTCs than in nRPTCs and increased (1.3�0.09-fold;P�0.05; n�4) after FEN stimulation only in nRPTCs (Figure2C). Figure 2D shows �-tubulin and Ponceau S loadingcontrols.
To prove that the reduced recruitment of D1R to the plasmamembrane in uRPTCs with FEN stimulation was because ofGRK4, we silenced GRK4 gene expression with GRK4siRNA (supplemental Figure S1, available in the online Data
2 Hypertension November 2009
by on May 11, 2011 hyper.ahajournals.orgDownloaded from
Supplement). The addition of GRK4-specific siRNA reducesGRK4 expression by 70.6�5.6 (P�0.05; n�3) by Westernblotting (Figure S1A). We have reported that, in uRPTCs,GRK4 is constitutively active, and the D1R is phosphorylated,desensitized, and internalized.31,34 When siRNA to GRK4was added to uRPTCs and compared with scrambled siRNA(SCR) and DMSO VEH controls, the FEN-mediated(1 �mol/L for 30 minutes) D1R plasma membrane expressionwas partially restored (P�0.05; n�6; Figure S1B).
The total cellular CAV1 expression level was increased by47�16% (P�0.05; n�8) with FEN stimulation (1 �mol/L; 4hours) when compared with DMSO VEH (Figure 3). Angio-tensin (ANG) II stimulation (10 nM; 4 hours) reduced CAV1expression by 67�8% (P�0.05; n�8).
Because CAV1 is a known negative regulator of GRKs,39 westudied the association of GRK4 and CAV1 in nRPTCs anduRPTCs with coimmunoprecipitation experiments. CAV1 wasimmunoprecipitated using a monoclonal anti-CAV1 antibodyand immunoblotted with GRK4 rabbit polyclonal antibody(Figure 4). A 62-kDa immunoreactive band was observedcorresponding with the predicted size of GRK4. CAV1/GRK4coimmunoprecipitation was 58�5% less in uRPTCs comparedwith nRPTCs (P�0.05, t test; n�4 per group).
The inverse of this experiment is shown in Figure S2, wherewe immunoprecipitated GRK4 and detected CAV1 and GRK4.There was a 55.6�14.1% decrease in uRPTCs compared withnRPTCs (P�0.05; n�4), further validating the difference inGRK4/CAV1 association between the cell groups.
Treatment with �-methyl cyclodextrin (�MCD; 2 mmol/L; 1hour), an agent that reduces cholesterol and CAV1 in the plasmamembrane, decreased CAV1 membrane expression in nRPTCs(52�4%; P�0.05 versus PBS VEH, nRPTCs; ANOVA, Holm-Sidak test, n�8) but not in uRPTCs (Figure 5). The depletion ofplasma membrane CAV1 by �MCD reduced CAV1 expressionin nRPTCs to the basal levels seen in uRPTCs. Basal plasmamembrane expression of CAV1 was lower in uRPTCs by56�11% (P�0.05; n�8) relative to nRPTCs.
We used another approach to reduce CAV1 expression.siRNA to CAV1 reduced total cellular CAV1 expression by86�8.5% (P�0.05, t test; n�8) in nRPTCs and 89�4%(P�0.05, t test; n�8) in uRPTCs compared with SCR-transfected control cells (Figure S3). Total cellular expressionof CAV1 was once again lower (25.2�3.0%; P�0.05; n�8)in uRPTCs than in nRPTCs.
Internalization of NaKATPase was measured by immuno-precipitating AP2 with a rabbit polyclonal antibody andWestern blotting with NaKATPase and AP2 monoclonalantibodies, producing single bands of the correct molecularweight for both AP2 and NaKATPase (Figure S4). Becausethere was only a single band for both proteins, furtheranalysis was continued by immuno-dot blot (Figure 6). FENstimulated the AP2-mediated NaKATPase internalizationin nRPTCs (�), a response that was not seen in uRPTCs (o).In nRPTCs, there was a 53�9% (P�0.05; n�6) increase inassociation between AP2 and NaKATPase with FEN(1 �mol/L for 30 minutes). This effect was completelyblocked by �MCD or siRNA to CAV1. Control SCR had noeffect on FEN-stimulated internalization of NaKATPase innRPTCs. In uRPTCs, FEN had no effect on AP2-NaKATPase coimmunoprecipitation and was not affected byCAV1 siRNA or CAV1 SCR. Other controls used were�MCD alone, CAV1 siRNA alone, CAV SCR alone, ortreatment without monensin; these controls had no effect onthe basal association of AP2 and NaKATPase (Figure S5).Controls without monensin showed no change in cell-surfaceNaKATPase expression after FEN stimulation in nRPTCs inFigure S6. Thus, an increase in intracellular sodium concen-tration is necessary for NaKATPase internalization.
Confocal images of NaKATPase displayed a predomi-nantly plasma membrane expression pattern in control (VEH)cells, which was reduced after FEN stimulation when anincrease in cytoplasmic accumulation was seen (P�0.05,t test; n�3; Figure S7). uRPTCs displayed an identicalexpression pattern as control nRPTCs but displayed no
0
1
2
3
4
5
6
7
8
9
nRPTCCON
nRPTCFEN
nRPTCFENLE300
uRPTCCON
uRPTCFEN
uRPTCFEN
LE300
pmol
escA
MP/
mg
prot
ein
*
#
A
0.8
0.9
1.0
1.1
1.2
1.3
1.4
1.5
0 2 4 6 8 10MINUTES
CFP
/YFP
nRPTC
uRPTC
FEN
B
Figure 1. Comparison of FEN-stimulated coupling efficiency with adenylyl cyclase in nRPTCs and uRPTCs. A, cAMP accumulationusing ELISA, in nRPTCs or uRPTCs after stimulation for 30 minutes with either DMSO vehicle control (CON), 1 �mol/L of FEN, or FENwith 10 �mol/L of LE300, a D1-like receptor antagonist (FEN LE300; *P�0.05 vs others; n�6 per group). B, Intracellular cAMP accumu-lation in real time using a cAMP fluorescence resonance energy transfer biosensor, ICUE3, measured in nRPTCs and uRPTCs. FEN(1 �mol/L; marked by an arrow at 1 minute) increased intracellular cAMP in nRPTCs vs uRPTCs. Data are from 3 separate wells of 10cells per well, imaged simultaneously. Graph is representative of 3 replicate experiments.
Gildea et al Dopamine Receptor Internalization of NaKATPase 3
by on May 11, 2011 hyper.ahajournals.orgDownloaded from
measurable internalization on FEN stimulation (data notshown).
We measured NaKATPase activity as ouabain-inhibitable(P�0.01; n�3 versus VEH) sodium efflux (Figure S8). FENreduced the sodium efflux in nRPTCs by 50.2�4.6% versusVEH (P�0.01; n�3), which was CAV1 dependent, becauseit was blocked by �MCD (P�0.01; n�3). FEN had no effectin uRPTCs.
DiscussionThese current studies demonstrate a CAV1 dependence onthe D1R-like receptor coupling to NaKATPase in humanRPTCs. The current study, along with our previous report,26
indicates that regulation of the D1R by CAV1 and/or CAV2may depend on the surrounding microenvironment (lipids andassociating receptors) in human RPTCs versus human em-bryonic kidney 293 cells.
CAV1 can also directly interact with GRKs 1, 2, and 5 inA431, NIH-3T3, and COS-1 cells,39 but a CAV1 interactionwith GRK4 has not been reported. In human embryonic kidney293 cells, GRK2 physically interacts with CAV226,39–41; CAV1is not expressed in human embryonic kidney 293 cells.22 Thus,our demonstration of GRK4 coimmunoprecipitation with CAV1is also novel, although not surprising. The lower GRK4/CAV1association in uRPTCs is in keeping with the negative modula-tory role of CAV1 on GRK4; GRK4 is constitutively active inuRPTCs.27
Our experiments demonstrate that CAV1 is also involvedwith GRK4 in the proper recycling, resensitization, anddesensitization of the D1R.42 In contrast, in uRPTCs, basalmembrane expression of CAV1 is decreased compared withnRPTCs. In uRPTCs, short-term FEN stimulation does notincrease CAV1 plasma membrane protein expression, GRK4plasma membrane protein does not change, and the D1R isnot recruited to the plasma membrane. These studies suggest
GRK4D1R
CAV1
62 kDa75 kDa
24 kDa
Fold
Indu
ctio
n (R
LU)
0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
nRPTC VEH nRPTC FEN uRPTC VEH uRPTC FEN0
2
4
6
8
10
12
14
nRPTC VEH nRPTC FEN uRPTC VEH uRPTC FEN
0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
nRPTC VEH nRPTC FEN uRPTC VEH uRPTC FEN
*
Fold
Indu
ctio
n (R
LU)
Fold
Indu
ctio
n (R
LU)
*
*
*
*
βTub 51 kDa
nRPT
C V
EH
nRPT
C F
EN
uRPT
C V
EH
uRPT
C F
EN
Ponceau SStain
BA
C D
Figure 2. Western blot analysis showing the effect of DMSO VEH or FEN (1 �mol/L for 30 minutes) on plasma membrane expressionof D1R, GRK4, and CAV1 in nRPTCs and uRPTCs. The results from representative Western blots are shown above each bar graph(*P�0.05 vs nRPTC VEH; n�4 for each graph). A, Increase in D1R plasma membrane expression after FEN (1 �mol/L for 30 minutes) innRPTCs but not in uRPTCs. B, Decrease in GRK4 plasma membrane expression after FEN (1 �mol/L for 30 minutes) in nRPTCs butnot in uRPTCs. The basal expression of plasma membrane GRK4 was lower in uRPTCs than in nRPTCs. C, Increase in CAV1 plasmamembrane expression after FEN stimulation seen in nRPTCs but not in uRPTCs. The basal expression of CAV1 was lower in uRPTCsvs nRPTCs (*P�0.05 vs VEH, nRPTCs). D, Loading controls of �-tubulin (�-Tub) and Ponceau S staining.
4 Hypertension November 2009
by on May 11, 2011 hyper.ahajournals.orgDownloaded from
that genetic defects in GRK4 that interrupt its associationwith CAV1 lead to desensitization and impaired function ofthe D1R. The role of ANG II in this process is currentlyunknown, although it is possible that the decreased expres-sion of CAV1 may be attributed to the lack of negativeregulation of the uncoupled D1R on the ANG II type 1receptor.43 The expression of the ANG II type 1 receptor isnot different in the nRPTC and uRPTC cell lines examined(data not shown). However, we have demonstrated that ANGII type 1 receptor expression and function are increased inmice overexpressing human GRK4 142V and 486V genes,relative to mice overexpressing human GRK4 wild-typegenes.44,45
In genetic hypertension, the ability of dopamine andD1-like receptor agonists to increase sodium excretion is
impaired. RPTs from hypertensive rats have an impairedability to decrease sodium transport (eg, sodium/hydrogenexchanger 3,46–49 NaHCO3 cotransporter,50 Cl/HCO3 ex-changer,51 and NaKATPase6,52–56). Dopamine inhibits sodiumtransporter activity, in part by increasing its internalization,an action that is also impaired in hypertension.16,57 Anincreased activity of NaKATPase in RPTs has been shown inseveral rodent genetic models of hypertension.56,58 Our stud-ies presented here in human RPTCs demonstrate that thescaffolding protein CAV1 is a mediator in the association ofthe D1R with its kinase, GRK4, and its role in AP2-associatedreduction in plasma membrane NaKATPase. Ultimately, thetransregulation of CAV1, D1R, and GRK4 will influencesodium transport. We are studying NaKATPase and sodium/hydrogen exchanger 3–mediated Na� flux, which will bereported in a future article.
*
*
0
50
100
150
VEH FEN AngII
CA
V1 (%
of C
ontr
ol)
VEH FEN AngII
24 kDaCAV1
200
βTub 51 kDa
Figure 3. Western blot analysis of total cellular expression ofCAV1 in nRPTCs treated with VEH, FEN, or ANG II. FEN(1 �mol/L; 4 hours) increased whereas ANG II (10 nM; 4 hours)decreased the total cell expression of CAV1 (*P�0.05 vs others,ANOVA, Holm-Sidak test; n�8 per group). �-Tubulin (�-Tub)shows even loading of proteins.
0
50
100
nRPTC uRPTC
GR
K4
(% n
RPT
C)
*
GRK4
CAV1
62 kDa
24 kDa
Figure 4. Association of CAV1 with GRK4 in nRPTCs anduRPTCs. CAV1 and GRK4 association was measured as theamount of immunoreactive GRK4 by Western blot of immuno-precipitated CAV1 (shown in top insert) using a monoclonal anti-body. Immunoreactive GRK4 associated with CAV1 wasdecreased in uRPTCs vs nRPTCs (*P�0.05 vs nRPTC, t test;n�4 per group).
Mem
bran
e C
AV1
(%
nR
PTC
VEH
)
0
50
100
nRPTCVEH
nRPTCβMCD
nRPTCVEH
uRPTC βMCD
*24 kDa
Figure 5. Effect of �MCD on the plasma membrane CAV1 expres-sion in nRPTCs and uRPTCs. �MCD (2 mmol/L; 1 hour), a choles-terol-reducing agent, decreased CAV1 expression in nRPTCs butnot in uRPTCs. Membrane CAV1 was reduced in nRPTCs by�MCD to the same level of CAV1 expression in uRPTCs (*P�0.05vs nRPTC PBS VEH, ANOVA, Holm-Sidak test; n�6 per group).
0
50
100
150
200
βMCD+
FEN
IP N
aKA
TPas
e (%
VEH
) nRPTCuRPTC
* #
VEH+
FEN
VEH+
VEH
CAV1siRNA
FEN
CAV1SCR
FEN+ +
Figure 6. AP2-mediated internalization of NaKATPase. Immuno-precipitated AP2 followed by NaKATPase dot blot is expressedas percentage of VEH in nRPTCs (�) and uRPTCs (o). FEN(1 �mol/L for 30 minutes) stimulated the AP2-mediatedNaKATPase internalization in nRPTCs but not in uRPTCs. TheFEN effect in nRPTCs was blocked by �MCD or CAV1 siRNA(*P�0.05 �MCD FEN vs VEH FEN in nRPTC, n�6 per group;#P�0.05 CAV1 siRNA FEN vs SCR FEN in nRPTCs, n�6 pergroup).
Gildea et al Dopamine Receptor Internalization of NaKATPase 5
by on May 11, 2011 hyper.ahajournals.orgDownloaded from

PerspectivesThere is a growing appreciation for the role of the dopami-nergic system in regulating sodium balance and, hence, bloodpressure. Genetic variants in a protein, GRK4, have beenlinked to essential hypertension and salt sensitivity in bothhumans and rodent models for these conditions. In thesestudies, we showed that CAV1, a principal membrane scaf-folding protein, is involved in the organization of the D1R-like receptor signaling pathway and intracellular sodium.These studies further decipher the mechanisms of howdopamine, GRK4, and CAV1 control the sodium pump. Theubiquitous expression of CAV1 may limit its usefulness as apotential therapeutic target. However, selective targeting ofGRK4, which regulates only a few G protein–coupled recep-tors in a limited number of organs, may be a new usefulantihypertensive therapeutic approach.
AcknowledgmentsWe thank Helen E. McGrath for editorial assistance with this article.
Sources of FundingThis work was supported by National Institutes of Health grantsHL074940, HL23081, DK39308, HL68686, and HL092196.
DisclosuresR.A.F. and P.A.J. were awarded a US Patent (No. 6 660 474) on“GRK Mutants in Essential Hypertension,” which has been assignedto Hypogen, Inc.
References1. Siragy HM, Felder RA, Howell NE, Chevalier RL, Peach MJ, Carey RM.
Intrarenal dopamine acts at the dopamine-1 receptor to control renalfunction. J Hypertens. 1988;6(suppl):S479–S481.
2. Siragy HM, Felder RA, Howell NL, Chevalier RL, Peach MJ, Carey RM.Intrarenal dopamine-1 receptors control renal function. Trans Assoc AmPhysicians. 1988;101:288–291.
3. Jose PA, Asico LD, Eisner GM, Pocchiari F, Semeraro C, Felder RA.Effects of costimulation of dopamine D1- and D2-like receptors on renalfunction. Am J Physiol. 1998;275:R986–R994.
4. Hedge SS, Ricci A, Amenta F, Lokhandwala MF. Evidence from func-tional and autoradiographic studies for the presence of tubulardopamine-1 receptors and their involvement in the renal effects offenoldopam. J Pharmacol Exp Ther. 1989;251:1237–1245.
5. Zeng C, Armando I, Luo Y, Eisner GM, Felder RA, Jose PA. Dysregu-lation of dopamine-dependent mechanisms as a determinant of hyper-tension: studies in dopamine receptor knockout mice. Am J Physiol HeartCirc Physiol. 2008;294:H551–H569.
6. Chen CJ, Lokhandwala MF. An impairment of renal tubular DA-1receptor function as the causative factor for diminished natriuresis tovolume expansion in spontaneously hypertensive rats. Clin ExpHypertens A. 1992;14:615–628.
7. Olsen NV. Effects of dopamine on renal haemodynamics tubular functionand sodium excretion in normal humans. Dan Med Bull. 1998;45:282–297.
8. O’Connell DP, Ragsdale NV, Boyd DG, Felder RA, Carey RM. Differ-ential human renal tubular responses to dopamine type 1 receptor stim-ulation are determined by blood pressure status. Hypertension. 1997;29:115–122.
9. Haney M, Ward AS, Foltin RW, Fischman MW. Effects of ecopipam, aselective dopamine D1 antagonist, on smoked cocaine self-administrationby humans. Psychopharmacology (Berl). 2001;155:330–337.
10. Hu MC, Fan L, Crowder LA, Karim-Jimenez Z, Murer H, Moe OW.Dopamine acutely stimulates Na�/H� exchanger (NHE3) endocytosisvia clathrin-coated vesicles: dependence on protein kinase A-mediatedNHE3 phosphorylation. J Biol Chem. 2001;276:26906–26915.
11. Felder CC, Campbell T, Albrecht F, Jose PA. Dopamine inhibitsNa(�)-H� exchanger activity in renal BBMV by stimulation of adenyl-ate cyclase. Am J Physiol. 1990;259:F297–F303.
12. Trivedi M, Narkar VA, Hussain T, Lokhandwala MF. Dopamine recruitsD1A receptors to Na-K-ATPase-rich caveolar plasma membranes in ratrenal proximal tubules. Am J Physiol Renal Physiol. 2004;287:F921–F931.
13. Efendiev R, Das-Panja K, Cinelli AR, Bertorello AM, Pedemonte CH.Localization of intracellular compartments that exchange Na,K-ATPasemolecules with the plasma membrane in a hormone-dependent manner.Br J Pharmacol. 2007;151:1006–1013.
14. Pedemonte CH, Efendiev R, Bertorello AM. Inhibition of Na,K-ATPaseby dopamine in proximal tubule epithelial cells. Semin Nephrol. 2005;25:322–327.
15. Yudowski GA, Efendiev R, Pedemonte CH, Katz AI, Berggren PO,Bertorello AM. Phosphoinositide-3 kinase binds to a proline-rich motif inthe Na�, K�-ATPase alpha subunit and regulates its trafficking. ProcNatl Acad Sci U S A. 2000;97:6556–6561.
16. Efendiev R, Budu CE, Cinelli AR, Bertorello AM, Pedemonte CH.Intracellular Na� regulates dopamine and angiotensin II receptors avail-ability at the plasma membrane and their cellular responses in renalepithelia. J Biol Chem. 2003;278:28719–28726.
17. Efendiev R, Krmar RT, Ogimoto G, Zwiller J, Tripodi G, Katz AI,Bianchi G, Pedemonte CH, Bertorello AM. Hypertension-linked mutationin the adducin alpha-subunit leads to higher AP2-mu2 phosphorylationand impaired Na�,K�-ATPase trafficking in response to GPCR signalsand intracellular sodium. Circ Res. 2004;95:1100–1108.
18. Cinelli AR, Efendiev R, Pedemonte CH. Trafficking of Na-K-ATPaseand dopamine receptor molecules induced by changes in intracellularsodium concentration of renal epithelial cells. Am J Physiol RenalPhysiol. 2008;295:F1117–F1125.
19. Dostanic-Larson I, Lorenz JN, Van Huysse JW, Neumann JC, MoseleyAE, Lingrel JB. Physiological role of the alpha1- and alpha2-isoforms ofthe Na�-K�-ATPase and biological significance of their cardiac gly-coside binding site. Am J Physiol Regul Integr Comp Physiol. 2006;290:R524–R528.
20. Li Z, Rodriguez-Iturbe B, Ni Z, Shahkarami A, Sepassi L, Vaziri ND.Effect of hereditary obesity on renal expressions of NO synthase,caveolin-1, AKt, guanylate cyclase, and calmodulin. Kidney Int. 2005;68:2766–2772.
21. Liu J, Liang M, Liu L, Malhotra D, Xie Z, Shapiro JI. Ouabain-inducedendocytosis of the plasmalemmal Na/K-ATPase in LLC-PK1 cellsrequires caveolin-1. Kidney Int. 2005;67:1844–1854.
22. Liu J, Shapiro JI. Regulation of sodium pump endocytosis by car-diotonic steroids: molecular mechanisms and physiological impli-cations. Pathophysiology. 2007;14:171–181.
23. Lisanti MP, Scherer PE, Vidugiriene J, Tang Z, Hermanowski-Vosatka A,Tu YH, Cook RF, Sargiacomo M. Characterization of caveolin-richmembrane domains isolated from an endothelial-rich source: implicationsfor human disease. J Cell Biol. 1994;126:111–126.
24. Patel HH, Murray F, Insel PA. Caveolae as organizers of pharmacolog-ically relevant signal transduction molecules. Annu Rev PharmacolToxicol. 2008;48:359–391.
25. Yu P, Yang Z, Asico L, Hopfer U, Felder R, Jose P. Redistribution ofcaveolin-2 in the response to D1-like receptor stimulation in immortalizedrenal proximal tubule cells. Mol Biol Cell. 2000;11:551.
26. Yu P, Yang Z, Jones JE, Wang Z, Owens SA, Mueller SC, Felder RA,Jose PA. D1 dopamine receptor signaling involves caveolin-2 inHEK-293 cells. Kidney Int. 2004;66:2167–2180.
27. Felder RA, Jose PA. Mechanisms of disease: the role of GRK4 in theetiology of essential hypertension and salt sensitivity. Nat Clin PractNephrol. 2006;2:637–650.
28. Watanabe H, Xu J, Bengra C, Jose PA, Felder RA. Desensitization ofhuman renal D1 dopamine receptors by G protein-coupled receptor kinase4. Kidney Int. 2002;62:790–798.
29. Felder RA, Seikaly MG, Cody P, Eisner GM, Jose PA. Attenuated renalresponse to dopaminergic drugs in spontaneously hypertensive rats.Hypertension. 1990;15:560–569.
30. Lokhandwala MF, Hussain T. Defective renal dopamine D1-like receptorsignal transduction in obese hypertensive rats. Acta Physiol Scand. 2000;168:251–255.
31. Felder RA, Sanada H, Xu J, Yu PY, Wang Z, Watanabe H, Asico LD,Wang W, Zheng S, Yamaguchi I, Williams SM, Gainer J, Brown NJ,Hazen-Martin D, Wong LJ, Robillard JE, Carey RM, Eisner GM, JosePA. G protein-coupled receptor kinase 4 gene variants in human essentialhypertension. Proc Natl Acad Sci U S A. 2002;99:3872–3877.
6 Hypertension November 2009
by on May 11, 2011 hyper.ahajournals.orgDownloaded from

32. Kinoshita S, Sidhu A, Felder RA. Defective dopamine-1 receptor ade-nylate cyclase coupling in the proximal convoluted tubule from thespontaneously hypertensive rat. J Clin Invest. 1989;84:1849–1856.
33. Felder RA, Eisner GM, Jose PA. D1 dopamine receptor signalling defectin spontaneous hypertension. Acta Physiol Scand. 2000;168:245–250.
34. Sanada H, Jose PA, Hazen-Martin D, Yu PY, Xu J, Bruns DE, Phipps J,Carey RM, Felder RA. Dopamine-1 receptor coupling defect in renalproximal tubule cells in hypertension. Hypertension. 1999;33:1036–1042.
35. Gildea JJ, Wang X, Jose PA, Felder RA. Differential D1 and D5 receptorregulation and degradation of the angiotensin type 1 receptor. Hypertension.2008;51:360–366.
36. Han W, Li H, Villar VA, Pascua AM, Dajani MI, Wang X, Natarajan A,Quinn MT, Felder RA, Jose PA, Yu P. Lipid rafts keep NADPH oxidasein the inactive state in human renal proximal tubule cells. Hypertension.2008;51:481–487.
37. Violin JD, DiPilato LM, Yildirim N, Elston TC, Zhang J, Lefkowitz RJ.Beta2-adrenergic receptor signaling and desensitization elucidated byquantitative modeling of real time cAMP dynamics. J Biol Chem. 2008;283:2949–2961.
38. Gilad LA, Schwartz B. Association of estrogen receptor beta withplasma-membrane caveola components: implication in control of vitaminD receptor. J Mol Endocrinol. 2007;38:603–618.
39. Carman CV, Lisanti MP, Benovic JL. Regulation of G protein-coupledreceptor kinases by caveolin. J Biol Chem. 1999;274:8858–8864.
40. Grilo A, Fernandez ML, Beltran M, Ramirez-Lorca R, Gonzalez MA,Royo JL, Gutierrez-Tous R, Moron FJ, Couto C, Serrano-Rios M, SaezME, Ruiz A, Real LM. Genetic analysis of CAV1 gene in hypertensionand metabolic syndrome. Thromb Haemost. 2006;95:696–701.
41. Ribas C, Penela P, Murga C, Salcedo A, Garcia-Hoz C, Jurado-Pueyo M,Aymerich I, Mayor F Jr. The G protein-coupled receptor kinase (GRK)interactome: role of GRKs in GPCR regulation and signaling. BiochimBiophys Acta. 2007;1768:913–922.
42. Gildea JJ, Yatabe J, Sasaki M, Jose PA, Felder RA. G-protein coupledreceptor kinase 4 (GRK4) polymorphisms block receptor recruitment tocell membranes. Hypertension. 2006;48:e85.
43. Zeng C, Luo Y, Asico LD, Hopfer U, Eisner GM, Felder RA, Jose PA.Perturbation of D1 dopamine and AT1 receptor interaction in sponta-neously hypertensive rats. Hypertension. 2003;42:787–792.
44. Wang Z, Asico LD, Escano CS, Felder RA, Jose PA. Human G protein-coupled receptor kinase type 4 g (hGRK4g) wild-type prevents saltsensitivity while its variant, hGRK4g486V, promotes salt sensitivity intransgenic mice: role of genetic background. Hypertension. 2006;48:e27.
45. Wang Z, Zeng C, Asico L, Felder RA, Jose PA. The role of GRK4gA142Vin the dysfunction of renal AT1 and D1 dopamine receptor in genetichypertension. Circulation. 2005;112:187.
46. Albrecht FE, Drago J, Felder RA, Printz MP, Eisner GM, Robillard JE,Sibley DR, Westphal HJ, Jose PA. Role of the D1A dopamine receptor inthe pathogenesis of genetic hypertension. J Clin Invest. 1996;97:2283–2288.
47. Li XX, Xu J, Zheng S, Albrecht FE, Robillard JE, Eisner GM, Jose PA.D(1) dopamine receptor regulation of NHE3 during development inspontaneously hypertensive rats. Am J Physiol Regul Integr CompPhysiol. 2001;280:R1650–R1656.
48. Xu J, Li XX, Albrecht FE, Hopfer U, Carey RM, Jose PA. Dopamine(1)receptor, G(salpha), and Na(�)-H(�) exchanger interactions in thekidney in hypertension. Hypertension. 2000;36:395–399.
49. Gesek FA, Schoolwerth AC. Hormone responses of proximal Na(�)-H�exchanger in spontaneously hypertensive rats. Am J Physiol. 1991;261:F526–F536.
50. Kunimi M, Seki G, Hara C, Taniguchi S, Uwatoko S, Goto A, Kimura S,Fujita T. Dopamine inhibits renal Na�:HCO3- cotransporter in rabbitsand normotensive rats but not in spontaneously hypertensive rats. KidneyInt. 2000;57:534–543.
51. Pedrosa R, Jose PA, Soares-da-Silva P. Defective D1-like receptor-mediated inhibition of the Cl-/HCO3- exchanger in immortalized SHRproximal tubular epithelial cells. Am J Physiol Renal Physiol. 2004;286:F1120–F1126.
52. Nishi A, Eklof AC, Bertorello AM, Aperia A. Dopamine regulation ofrenal Na�,K(�)-ATPase activity is lacking in Dahl salt-sensitive rats.Hypertension. 1993;21:767–771.
53. Hussain T, Lokhandwala MF. Renal dopamine DA1 receptor couplingwith G(S) and G(q/11) proteins in spontaneously hypertensive rats. Am JPhysiol. 1997;272:F339–F346.
54. Chen C, Beach RE, Lokhandwala MF. Dopamine fails to inhibit renaltubular sodium pump in hypertensive rats. Hypertension. 1993;21:364–372.
55. Lucas-Teixeira VA, Vieira-Coelho MA, Serrao P, Pestana M, Soares-da-Silva P. Salt intake and sensitivity of intestinal and renal Na�-K�atpase to inhibition by dopamine in spontaneous hypertensive andWistar-Kyoto rats. Clin Exp Hypertens. 2000;22:455–469.
56. Debska-Slizien A, Ho P, Drangova R, Baines AD. Endogenous dopamineregulates phosphate reabsorption but not NaK-ATPase in spontaneouslyhypertensive rat kidneys. J Am Soc Nephrol. 1994;5:1125–1132.
57. Efendiev R, Chen Z, Krmar RT, Uhles S, Katz AI, Pedemonte CH,Bertorello AM. The 14–3-3 protein translates the NA�,K�-ATPase{alpha}1-subunit phosphorylation signal into binding and activation ofphosphoinositide 3-kinase during endocytosis. J Biol Chem. 2005;280:16272–16277.
58. Hinojos CA, Doris PA. Altered subcellular distribution of Na�,K�-ATPase in proximal tubules in young spontaneously hypertensive rats.Hypertension. 2004;44:95–100.
Gildea et al Dopamine Receptor Internalization of NaKATPase 7
by on May 11, 2011 hyper.ahajournals.orgDownloaded from

#134388 ONLINE SUPPLEMENT
Caveolin-1 and Dopamine Mediated Internalization of NaKATPase in Human Renal Proximal Tubule Cells
1John J. Gildea, 1Jonathan A. Israel, 1Andrew K. Johnson, 3Jin Zhang, 2Pedro A. Jose, and 1Robin A. Felder
1The University of Virginia Health System, Department of Pathology, Charlottesville, VA 2Children’s National Medical Center, Center for Molecular Physiology, Department of Pediatrics, George Washington University, Washington, DC 3Johns Hopkins School of Medicine, Baltimore, MD. Short title: Dopamine receptor internalization of NaKATPase Correspondence to be sent to: Robin A. Felder, Ph.D. The University of Virginia P.O. Box 801400 Charlottesville, VA 22908 [email protected] Ph 434-466-1131 Fax 434-924-5718
by on May 11, 2011 hyper.ahajournals.orgDownloaded from

Expanded Materials and Methods Cell Lines: University of Virginia Institutional Review Board (IRB)-approved normal tissue from nephrectomized kidneys was used to isolate RPTC verified by the following criteria: cAMP accumulation upon parathyroid hormone stimulation; positive plasma membrane binding of lotus tetragonobolus agglutinin; expression of γ-glutamyl transpeptidase, aminopeptidase N and NHE3 (1). The hybridoma producing monoclonal antibody 452 against CD13 (aminopeptidase N) was kindly donated by Dr. Meenhard Herlyn. We also verified the lack of expression of Tamm-Horsfall protein, sodium potassium 2Cl co-transporter, sodium chloride transporter, and epithelial sodium channel, which are markers of potentially contaminating tubule cells from the thick ascending limb of Henle, distal convoluted tubule and collecting duct. Determination of cAMP accumulation: ELISA: nRPTC and uRPTC were grown to 80% confluence, serum- starved overnight, and then washed in phosphate-buffered saline (PBS) with calcium and magnesium. 1 mM isobutyl-1-methyl-xanthine (to inhibit phosphodiesterase activity) was added along with the D1-like receptor agonist fenoldopam (1 μM) or vehicle control (dimethyl sulfoxide, DMSO) and incubated for 30 minutes at 37oC. The D1-like receptor antagonist LE300 (10 μM) was added 10 minutes prior to the addition of fenoldopam, to show D1-like receptor specificity of the measured end points. Commercial ELISA kit (Cayman Chemical) was performed according to manufacturer’s specifications. ICUE3: The optimized transfection protocol for human RPTC was found to be a square wave pulse with 250 mV, 2000 uF capacitance, 1000 ohms resistance, 15 ms duration, two sets of 2 pulses, and 0.1 ms between pulses using 5x105 cells per well and 0.4 μg/well plasmid. These settings produced between 60% and 90% transfection efficiencies depending on the cell line, passage number, and confluence of the cells at the time of harvest. Transfected cells were plated in 96-well collagen coated glass-bottomed plates (Matrical, Fisher Scientific), and CFP/YFP ratiometric imaging was performed following fenoldopam (1 uM) treatment. An increase in cAMP decreases the association of CFP and YFP producing an increase in the normalized CFP/YFP ratio. Images were acquired from 10 cells per well, 3 wells per group, on an Olympus IX81 spinning disk multi-well automated microscope and analyzed using the ratiometric imaging module of Slidebook 4.2. Measurement of the association of GRK4 and CAV1: GRK4 and CAV1 association was measured by co-immunoprecipitation. Cells were lysed in M-PER (Pierce), and 1 mg protein in 1 ml lysis buffer was immunoprecipitated by adding 2 μg CAV1 monoclonal antibody (BD Biosciences) for two hours. Protein A/G agarose (Pierce) was subsequently added, and then incubated for 30 minutes. The beads were washed three times in lysis buffer and protein was eluted from the column with lithium dodecyl sulfate (LDS) loading buffer with reducing agent (Invitrogen) and western-blotted as above, using rabbit polyclonal anti-GRK4 (1:200 dilution, Santa Cruz, sc-13079). The HA antigen is not present in these cells and therefore the 12CA5 monoclonal anti-HA antibody (gift from Tanya Watts University of Toronto) was used as a negative control. The association was also shown by immunoprecipitation with monoclonal GRK4 antibody conjugated to agarose (Santa Cruz, sc-9985AC) and detected with rabbit polyclonal CAV1 followed by goat anti-rabbit IR Dye 800 secondary antibody and imaged on Odyssey infrared imaging system. SiRNA to CAV1 and GRK4: Cells were grown in six-well plates (60% confluence), and 50% fresh complete media changed 2 hours before transfection. siRNA (50 nM) against CAV1 and GRK4, or scrambled control siRNA, was incubated with 6 μl oligofectamine (Invitrogen) in Optimem media (Invitrogen) and added to the cells for 4 hours, then FBS (fetal bovine serum) was added to a 2% final concentration and incubated overnight. The next day the cell culture media were switched to growth factor-free media with 0.5%
by on May 11, 2011 hyper.ahajournals.orgDownloaded from

FBS for another 24 hours before lysing for western blotting or using in the NaKATPase internalization assay. NaKATPase Internalization Assay: RPTC were growth factor-starved overnight in 0.5% FBS, then monensin (5 µM) was added for 30 minutes to increase the intracellular sodium concentration to 20 mM. β-methyl cyclodextrin (βMCD, 2 mM) was added to deplete membrane cholesterol and CAV1 (2, 3). Two µg rabbit anti-AP2 antibody (α-adaptin, Santa Cruz, sc-10761) or a non-specific rabbit IgG (Sigma) were added to 500 µg lysate in 1 ml lysis buffer (2 hours). Protein A sepharose was incubated for 30 minutes with the lysate, washed 3X in lysis buffer and eluted with LDS loading buffer with reducing agent (Invitrogen). The eluate was immuno dot-blotted for NaKATPase using a 1:50 dilution of hybridoma culture supernatant (clone A5, Developmental Studies Hybridoma Bank). Non-specific rabbit IgG did not show any signal above background when examined by western blot (Figure S4). The NaKATPase signal was normalized to the amount of AP2 pulled down using a replicate well probed with a mouse anti-AP2 monoclonal antibody (α-adaptin, Santa Cruz, sc-17771). Following the addition of both NaKATPase and AP2 mouse monoclonal antibodies, goat anti-mouse IR Dye 800 secondary antibody was added, and the cells were imaged using an Odyssey infrared imaging system. NaKATPase confocal microscopy and plasma membrane localization: Human RPTC were plated on 25 mm collagen-coated glass coverslips in a 6 well dish. Treatments were the same as for the NaKATPase internalization assay, except that after the 30 minute treatment period, the cells were washed with PBS three times and fixed with 4% paraformaldehyde in PBS for 5 minutes, washed with PBS, and residual paraformaldehyde quenched with 50 mM glycine for 5 minutes. Cells were washed, then permeabilized with PBS containing 0.2% Triton X 100 for 5 minutes, and blocked with 1% BSA (bovine serum albumin) and 5% normal goat serum in PBS with 0.2% Tween 20 for 1 hour. NaKATPase α-1 monoclonal (clone C464.6, Millipore) culture supernatant was diluted 1:500 in blocking buffer and incubated overnight at 4oC. Cells were washed in PBS with 0.2% Tween 20 and secondary goat anti-mouse Alexa 488 was diluted 1:500 and incubated in blocking buffer for one hour at room temperature, then washed three times in PBS and mounted in Fluoromount G (Chemicon). Images were acquired with a Zeiss 510 Meta confocal microscope using a 60X water immersion objective at 1 micron z-axis intervals for a total z-axis height of 10 microns. A mid z-axis plane was taken at 5 microns from the most basal plane in both VEH control and FEN (1 μM, 30 min.) All images were acquired using the exact same settings, which were set such that the secondary only control showed no staining and the VEH control displayed strong plasma membrane specific staining. Plasma membrane localized NaKATPase expression was quantified using the ImageJ version 1.39o image processing software program. The confocal image and the DIC image were opened in two separate windows and the “sync windows” macro (courtesy of Patrick Kelly) was run. A 5 μ box was selected and placed at the plasma membrane chosen using the DIC image, and the integrated intensity of the confocal image was measured. The box was then moved 10 μ toward the cell interior and the integrated intensity re-measured. This was repeated 20X around the perimeter of each cell, and 10 cells per well quantified. Each cell was studied three times. The plasma membrane to cytoplasm ratio was the average surface intensity divided by the average cytoplasmic intensity. Background intensity was measured from a cell free region and subtracted prior to calculating the intensity ratio. Sodium Efflux Assay: RPTC were cultured in 96-well glass bottom collagen coated Matrical plates (Spokane, WA) at 37˚C until they reached 80% confluence. Cells were serum-starved overnight prior to loading with a sodium ion indicator, sodium benzofuran isophthalate (SBFI, 5μM) (Molecular Probes, Eugene, OR) with Pluronic 127 for 2 hours in PBS with calcium and magnesium. Cells were allowed to recover at
by on May 11, 2011 hyper.ahajournals.orgDownloaded from

37oC in serum free media for 30 minutes, then washed twice with room temperature potassium free HEPES media (20 mM HEPES pH 7.4, 130 mM NaCl, 1 mM CaCl, 1 mM MgCl) and incubated at room temperature for 30 minutes to raise the internal sodium concentration. VEH, FEN, βMCD or OUB (Ouabain, a cardiac glycoside, known to bind to and inhibit NaKATPase) were added to respective wells for 10 minutes and placed on the stage of an automated microscope for time lapse multiwell ratiometric imaging. Directly before imaging, EIPA (10 uM final concentration) and KCl (2.7 mM final concentration) were added to all wells as 10X stocks, and then imaged for 20 minutes. EIPA (5-(N-ethyl-N-isopropyl) amiloride) is a selective inhibitor of the Na+/H+ exchanger 3 (NHE3), which is the exclusive sodium hydrogen exchanger in human RPTC. Each measurement shown is the change in 340/380 ratio from time 0 in 3 separate well with 20 cells per well measured. Microscopy: Images were collected on an Olympus IX-81 inverted epifluorescence microscope with a UV-specific 20X UAPO objective and Hamamatsu ER CCD camera. Slidebook software package 4.2 (Intelligent Imaging Innovations, Inc., Denver, CO) integrated all of the hardware as well as provided for image acquisition and processing. An automated XY stage and piezo-Z allowed us to precisely capture images from multiple wells of a 96-well microplate under the same conditions. We can simultaneously capture time lapse data from up to 3 fields of view per well, in 12 separate wells per experiment.
by on May 11, 2011 hyper.ahajournals.orgDownloaded from

References
1. Sanada H, Jose PA, Hazen-Martin D, Yu PY, Xu J, Bruns DE, Phipps J, Carey RM, Felder RA. Dopamine-1 receptor coupling defect in renal proximal tubule cells in hypertension. Hypertension.. 1999; 33:1036-1042.
2. Han W, Li H, Villar VA, Pascua AM, Dajani MI, Wang X, Aruna Natarajan; Quinn MT,
Felder RA, Jose PA, Yu P. Lipid rafts keep NADPH oxidase in the inactive state in human renal proximal tubule cells. Hypertension. 2008; 51: 481-487.
3. Linder AE, Thakali KM, Thompson JM, Watts SW , Webb RC, Leite R. Methyl-beta-cyclodextrin prevents angiotensin II-induced tachyphylactic contractile responses in rat aorta. J Pharmacol Exp Ther. 2007; 323: 78-84.
by on May 11, 2011 hyper.ahajournals.orgDownloaded from
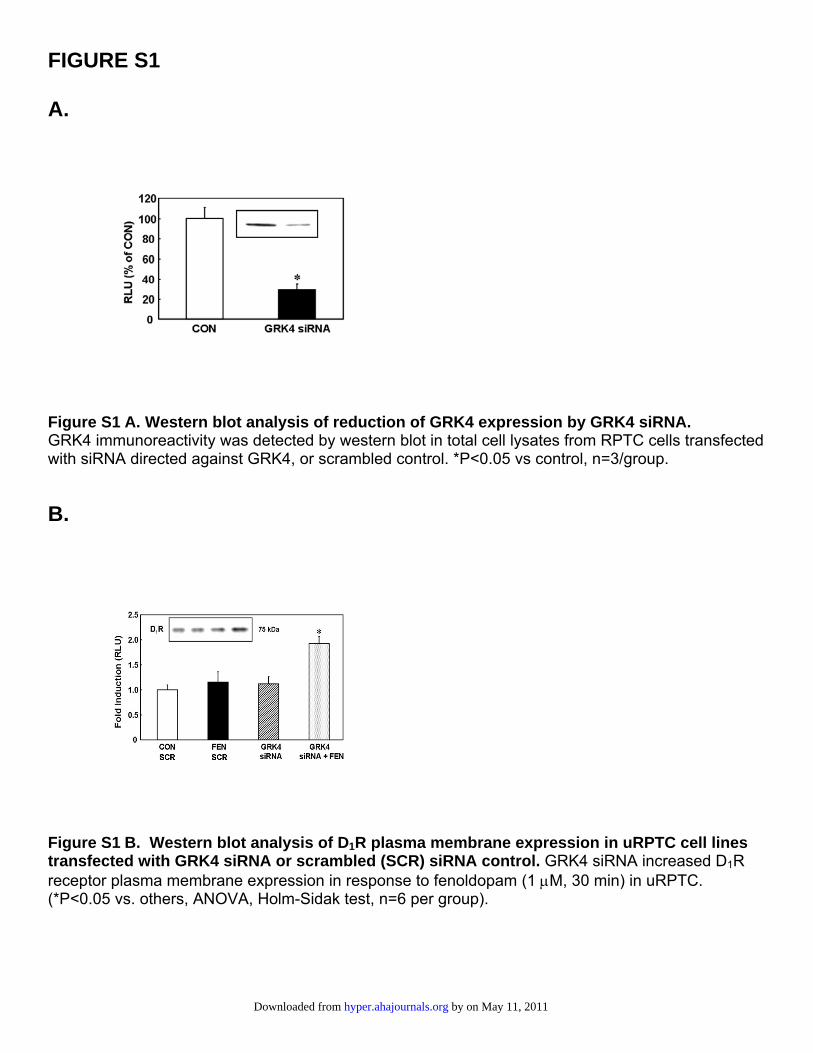
FIGURE S1 A.
Figure S1 A. Western blot analysis of reduction of GRK4 expression by GRK4 siRNA. GRK4 immunoreactivity was detected by western blot in total cell lysates from RPTC cells transfected with siRNA directed against GRK4, or scrambled control. *P<0.05 vs control, n=3/group.
B.
Figure S1 B. Western blot analysis of D1R plasma membrane expression in uRPTC cell lines transfected with GRK4 siRNA or scrambled (SCR) siRNA control. GRK4 siRNA increased D1R receptor plasma membrane expression in response to fenoldopam (1 μM, 30 min) in uRPTC. (*P<0.05 vs. others, ANOVA, Holm-Sidak test, n=6 per group).
by on May 11, 2011 hyper.ahajournals.orgDownloaded from
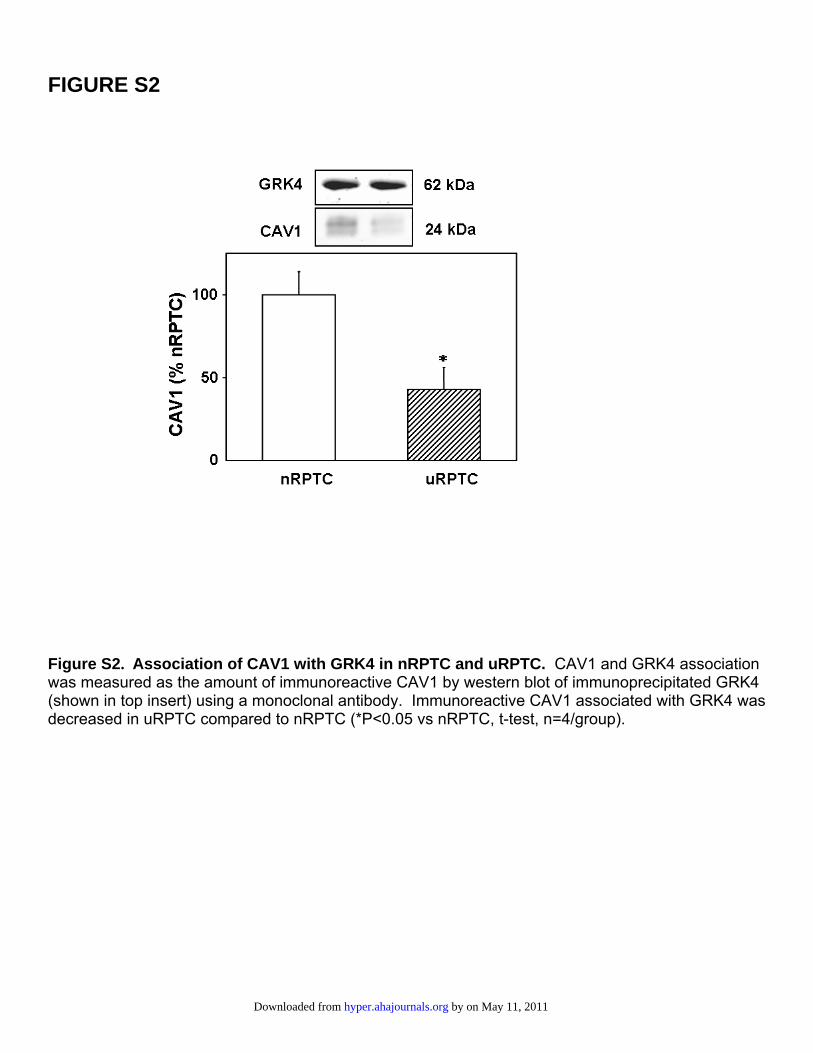
FIGURE S2
Figure S2. Association of CAV1 with GRK4 in nRPTC and uRPTC. CAV1 and GRK4 association was measured as the amount of immunoreactive CAV1 by western blot of immunoprecipitated GRK4 (shown in top insert) using a monoclonal antibody. Immunoreactive CAV1 associated with GRK4 was decreased in uRPTC compared to nRPTC (*P<0.05 vs nRPTC, t-test, n=4/group).
by on May 11, 2011 hyper.ahajournals.orgDownloaded from
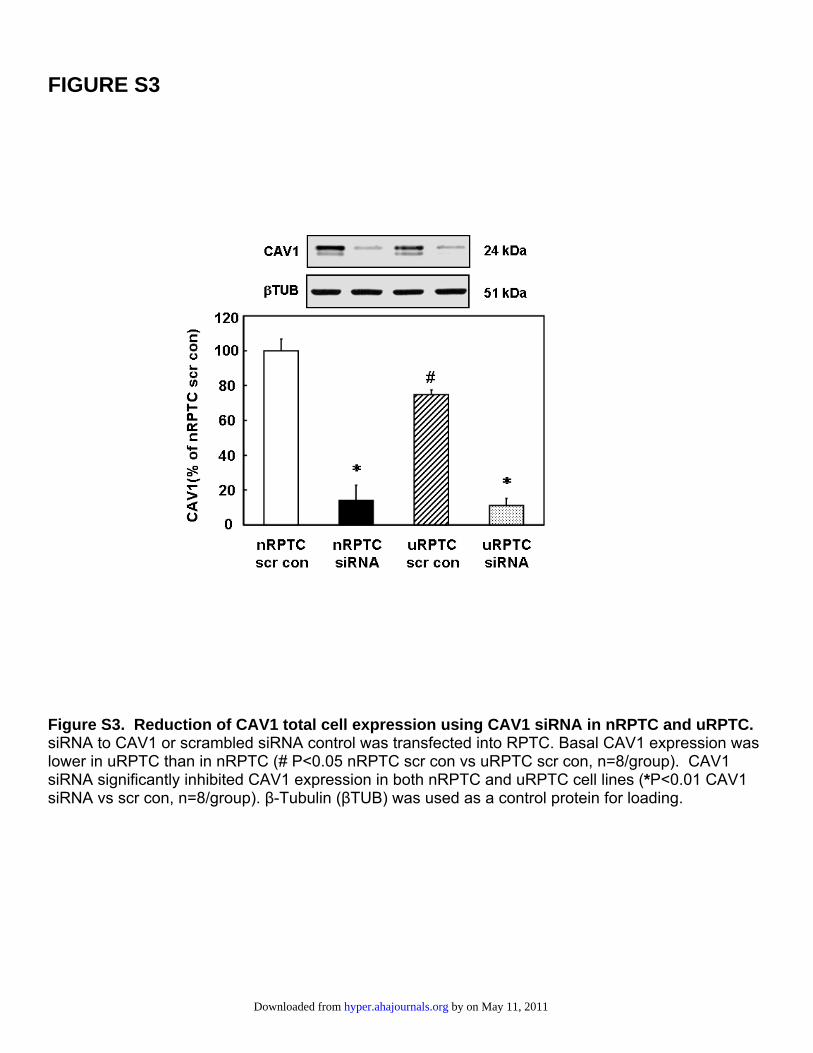
FIGURE S3
Figure S3. Reduction of CAV1 total cell expression using CAV1 siRNA in nRPTC and uRPTC. siRNA to CAV1 or scrambled siRNA control was transfected into RPTC. Basal CAV1 expression was lower in uRPTC than in nRPTC (# P<0.05 nRPTC scr con vs uRPTC scr con, n=8/group). CAV1 siRNA significantly inhibited CAV1 expression in both nRPTC and uRPTC cell lines (*P<0.01 CAV1 siRNA vs scr con, n=8/group). β-Tubulin (βTUB) was used as a control protein for loading.
by on May 11, 2011 hyper.ahajournals.orgDownloaded from
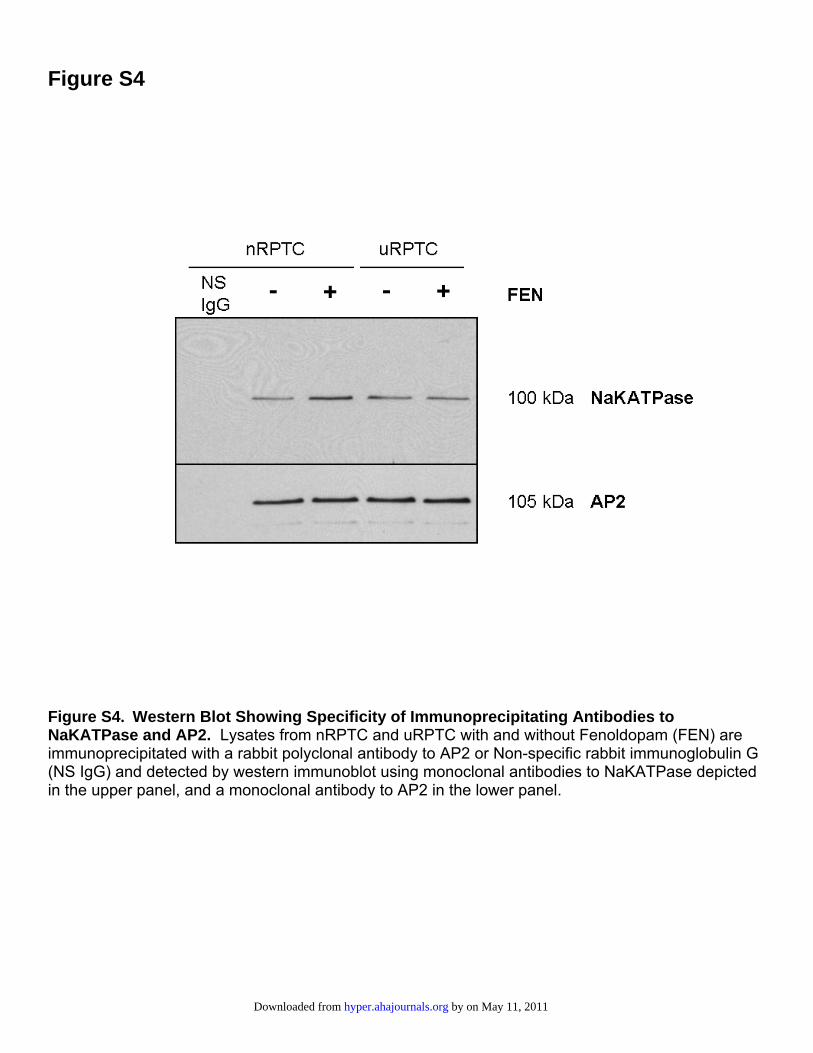
Figure S4
Figure S4. Western Blot Showing Specificity of Immunoprecipitating Antibodies to NaKATPase and AP2. Lysates from nRPTC and uRPTC with and without Fenoldopam (FEN) are immunoprecipitated with a rabbit polyclonal antibody to AP2 or Non-specific rabbit immunoglobulin G (NS IgG) and detected by western immunoblot using monoclonal antibodies to NaKATPase depicted in the upper panel, and a monoclonal antibody to AP2 in the lower panel.
by on May 11, 2011 hyper.ahajournals.orgDownloaded from
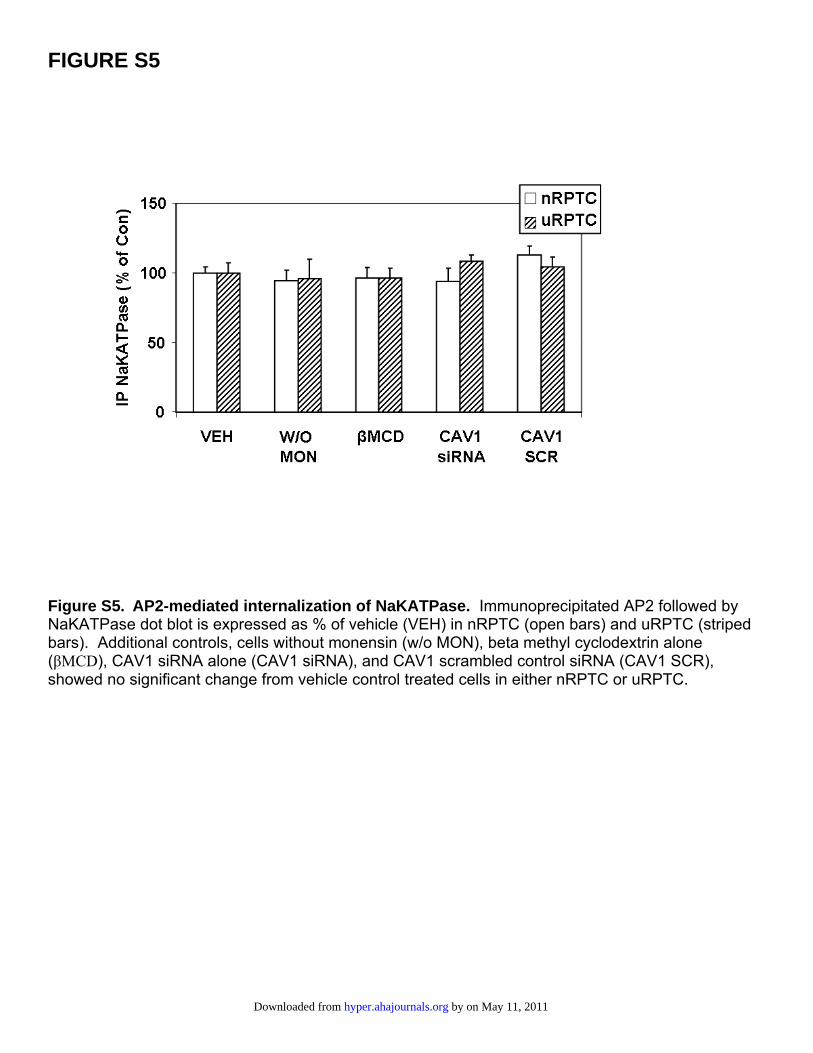
FIGURE S5
Figure S5. AP2-mediated internalization of NaKATPase. Immunoprecipitated AP2 followed by NaKATPase dot blot is expressed as % of vehicle (VEH) in nRPTC (open bars) and uRPTC (striped bars). Additional controls, cells without monensin (w/o MON), beta methyl cyclodextrin alone (βMCD), CAV1 siRNA alone (CAV1 siRNA), and CAV1 scrambled control siRNA (CAV1 SCR), showed no significant change from vehicle control treated cells in either nRPTC or uRPTC.
by on May 11, 2011 hyper.ahajournals.orgDownloaded from
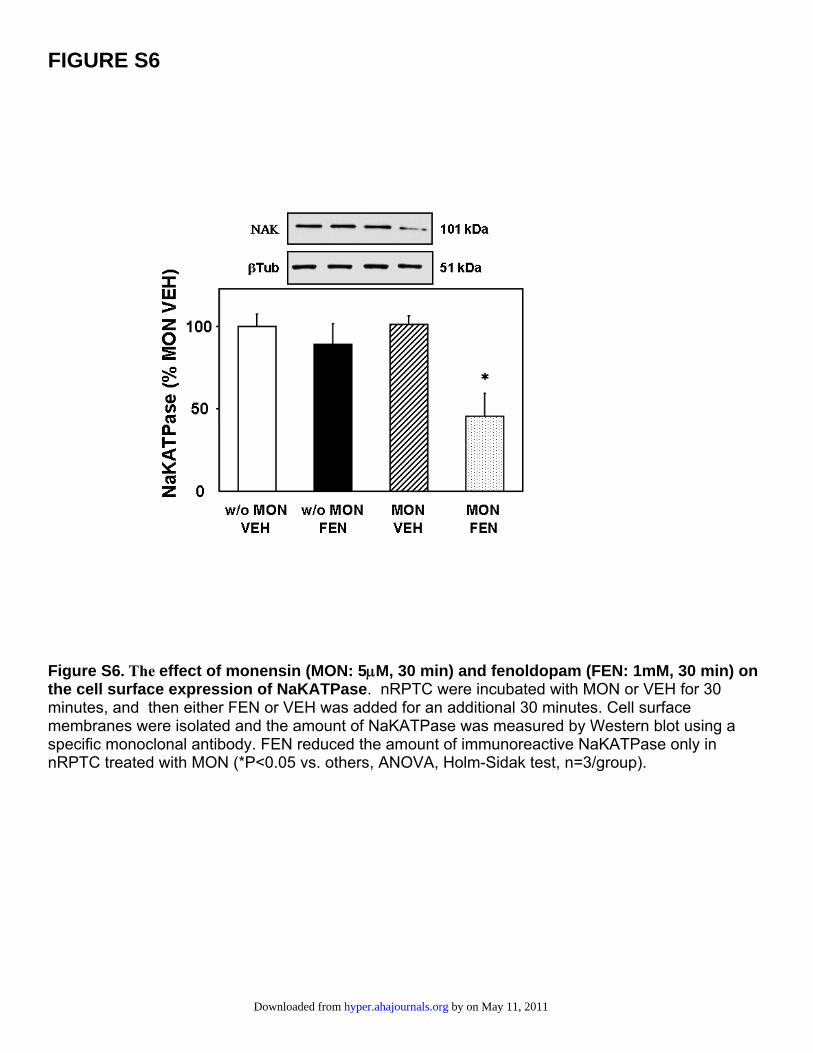
FIGURE S6
Figure S6. The effect of monensin (MON: 5μM, 30 min) and fenoldopam (FEN: 1mM, 30 min) on the cell surface expression of NaKATPase. nRPTC were incubated with MON or VEH for 30 minutes, and then either FEN or VEH was added for an additional 30 minutes. Cell surface membranes were isolated and the amount of NaKATPase was measured by Western blot using a specific monoclonal antibody. FEN reduced the amount of immunoreactive NaKATPase only in nRPTC treated with MON (*P<0.05 vs. others, ANOVA, Holm-Sidak test, n=3/group).
by on May 11, 2011 hyper.ahajournals.orgDownloaded from
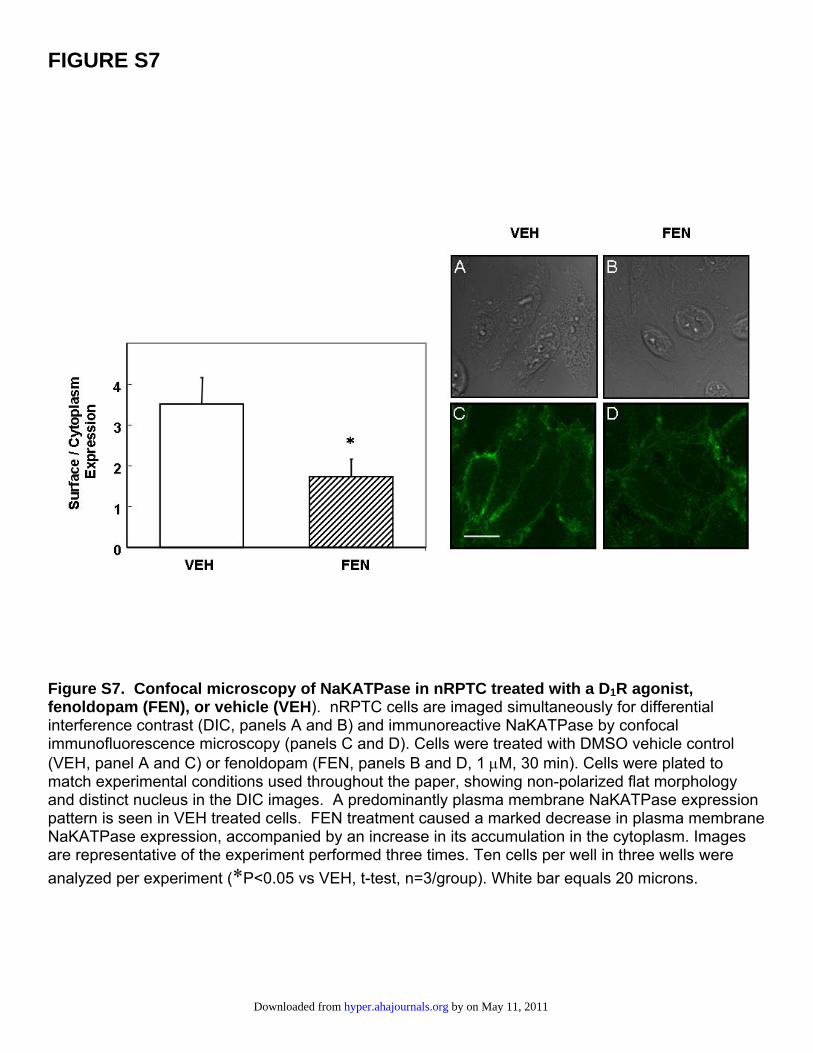
FIGURE S7
Figure S7. Confocal microscopy of NaKATPase in nRPTC treated with a D1R agonist, fenoldopam (FEN), or vehicle (VEH). nRPTC cells are imaged simultaneously for differential interference contrast (DIC, panels A and B) and immunoreactive NaKATPase by confocal immunofluorescence microscopy (panels C and D). Cells were treated with DMSO vehicle control (VEH, panel A and C) or fenoldopam (FEN, panels B and D, 1 μM, 30 min). Cells were plated to match experimental conditions used throughout the paper, showing non-polarized flat morphology and distinct nucleus in the DIC images. A predominantly plasma membrane NaKATPase expression pattern is seen in VEH treated cells. FEN treatment caused a marked decrease in plasma membrane NaKATPase expression, accompanied by an increase in its accumulation in the cytoplasm. Images are representative of the experiment performed three times. Ten cells per well in three wells were analyzed per experiment (*P<0.05 vs VEH, t-test, n=3/group). White bar equals 20 microns.
by on May 11, 2011 hyper.ahajournals.orgDownloaded from
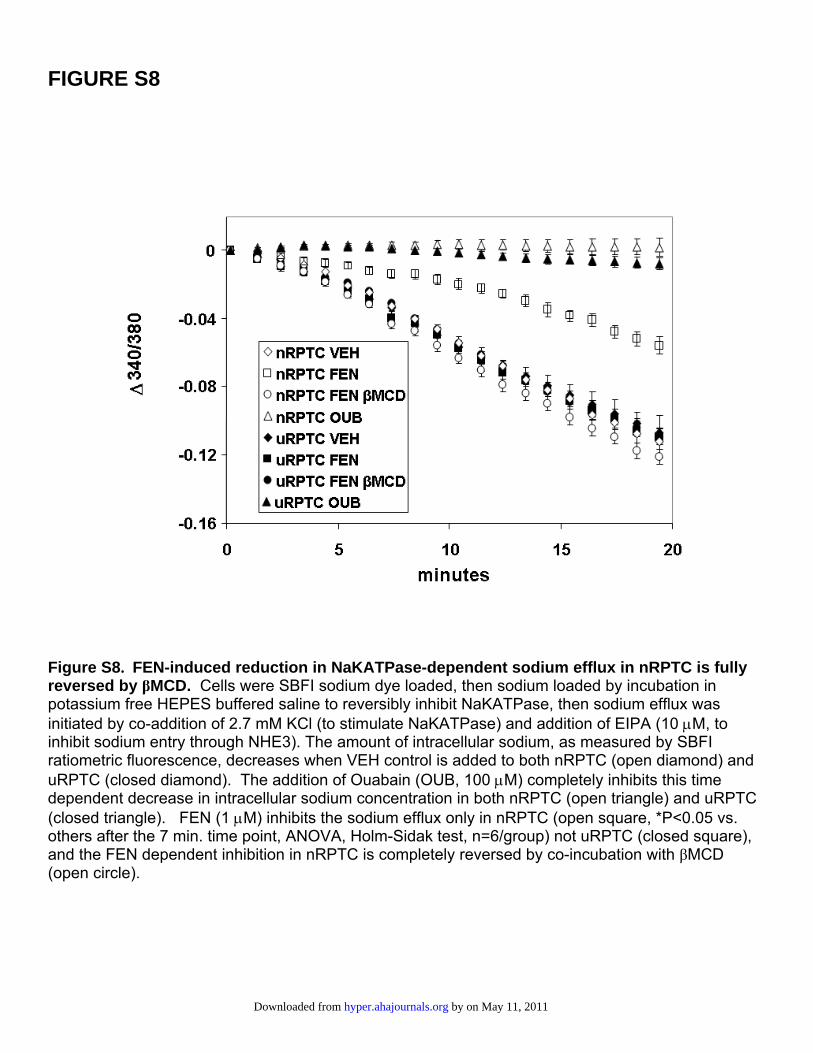
FIGURE S8
Figure S8. FEN-induced reduction in NaKATPase-dependent sodium efflux in nRPTC is fully reversed by βMCD. Cells were SBFI sodium dye loaded, then sodium loaded by incubation in potassium free HEPES buffered saline to reversibly inhibit NaKATPase, then sodium efflux was initiated by co-addition of 2.7 mM KCl (to stimulate NaKATPase) and addition of EIPA (10 μM, to inhibit sodium entry through NHE3). The amount of intracellular sodium, as measured by SBFI ratiometric fluorescence, decreases when VEH control is added to both nRPTC (open diamond) and uRPTC (closed diamond). The addition of Ouabain (OUB, 100 μM) completely inhibits this time dependent decrease in intracellular sodium concentration in both nRPTC (open triangle) and uRPTC (closed triangle). FEN (1 μM) inhibits the sodium efflux only in nRPTC (open square, *P<0.05 vs. others after the 7 min. time point, ANOVA, Holm-Sidak test, n=6/group) not uRPTC (closed square), and the FEN dependent inhibition in nRPTC is completely reversed by co-incubation with βMCD (open circle).
by on May 11, 2011 hyper.ahajournals.orgDownloaded from
Related Documents











