ION CHANNELS, RECEPTORS AND TRANSPORTERS Cationic uremic toxins affect human renal proximal tubule cell functioning through interaction with the organic cation transporter Carolien M. S. Schophuizen & Martijn J. Wilmer & Jitske Jansen & Lena Gustavsson & Constanze Hilgendorf & Joost G. J. Hoenderop & Lambert P. van den Heuvel & Rosalinde Masereeuw Received: 3 January 2013 / Revised: 31 May 2013 / Accepted: 2 June 2013 / Published online: 29 June 2013 # Springer-Verlag Berlin Heidelberg 2013 Abstract Several organic cations, such as guanidino com- pounds and polyamines, have been found to accumulate in plasma of patients with kidney failure due to inadequate renal clearance. Here, we studied the interaction of cationic uremic toxins with renal organic cation transport in a condi- tionally immortalized human proximal tubule epithelial cell line (ciPTEC). Transporter activity was measured and validated in cell suspensions by studying uptake of the fluorescent sub- strate 4-(4-(dimethylamino)styryl)-N-methylpyridinium-iodide (ASP + ). Subsequently, the inhibitory potencies of the cationic uremic toxins, cadaverine, putrescine, spermine and spermidine (polyamines), acrolein (polyamine breakdown product), guani- dine, and methylguanidine (guanidino compounds) were deter- mined. Concentration-dependent inhibition of ASP + uptake by TPA, cimetidine, quinidine, and metformin confirmed function- al endogenous organic cation transporter 2 (OCT2) expression in ciPTEC. All uremic toxins tested inhibited ASP + uptake, of which acrolein required the lowest concentration to provoke a half-maximal inhibition (IC 50 =44±2 μM). A Dixon plot was constructed for acrolein using three independent inhibition curves with 10, 20, or 30 μM ASP + , which demonstrated competitive or mixed type of interaction (K i =93 ± 16 μM). Exposing the cells to a mixture of cationic uremic toxins resulted in a more potent and biphasic inhibitory response curve, indicating complex interactions between the toxins and ASP + uptake. In conclusion, ciPTEC proves a suitable model to study cationic xenobiotic interactions. Inhibition of cellular uptake transport was demonstrated for several The manuscript was written through contributions of all authors. All authors have given approval to the final version of the manuscript. Carolien M.S. Schophuizen, Martijn J. Wilmer, Lambert P. van den Heuvel, and Rosalinde Masereeuw contributed equally to this work. Electronic supplementary material The online version of this article (doi:10.1007/s00424-013-1307-z) contains supplementary material, which is available to authorized users. C. M. S. Schophuizen : L. P. van den Heuvel Department of Pediatric Nephrology, Radboud University Nijmegen Medical Centre, 6525 GA Nijmegen, The Netherlands M. J. Wilmer : J. Jansen : R. Masereeuw (*) Department of Pharmacology and Toxicology (149), Nijmegen Centre for Molecular Life Sciences/Institute for Genetic and Metabolic Disease, Radboud University Nijmegen Medical Centre, P.O. Box 9101, 6500 HB Nijmegen, The Netherlands e-mail: [email protected] J. Jansen : J. G. J. Hoenderop Department of Physiology, Radboud University Nijmegen Medical Centre, 6525 GA Nijmegen, The Netherlands L. Gustavsson Department of Laboratory Medicine (Malmö), Lund University, Skåne University Hospital, 205 02 Malmö, Sweden C. Hilgendorf AstraZeneca R&D, Innovative Medicines, Global DMPK, 431 83 Mölndal, Sweden L. P. van den Heuvel Department of Pediatrics, Catholic University Leuven, 3000 Leuven, Belgium Pflugers Arch - Eur J Physiol (2013) 465:1701–1714 DOI 10.1007/s00424-013-1307-z

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ION CHANNELS, RECEPTORS AND TRANSPORTERS
Cationic uremic toxins affect human renal proximal tubulecell functioning through interaction with the organiccation transporter
Carolien M. S. Schophuizen & Martijn J. Wilmer & Jitske Jansen &
Lena Gustavsson & Constanze Hilgendorf & Joost G. J. Hoenderop &
Lambert P. van den Heuvel & Rosalinde Masereeuw
Received: 3 January 2013 /Revised: 31 May 2013 /Accepted: 2 June 2013 /Published online: 29 June 2013# Springer-Verlag Berlin Heidelberg 2013
Abstract Several organic cations, such as guanidino com-pounds and polyamines, have been found to accumulate inplasma of patients with kidney failure due to inadequaterenal clearance. Here, we studied the interaction of cationicuremic toxins with renal organic cation transport in a condi-tionally immortalized human proximal tubule epithelial cellline (ciPTEC). Transporter activity was measured and validatedin cell suspensions by studying uptake of the fluorescent sub-strate 4-(4-(dimethylamino)styryl)-N-methylpyridinium-iodide(ASP+). Subsequently, the inhibitory potencies of the cationicuremic toxins, cadaverine, putrescine, spermine and spermidine(polyamines), acrolein (polyamine breakdown product), guani-dine, and methylguanidine (guanidino compounds) were deter-mined. Concentration-dependent inhibition of ASP+ uptake by
TPA, cimetidine, quinidine, and metformin confirmed function-al endogenous organic cation transporter 2 (OCT2) expressionin ciPTEC. All uremic toxins tested inhibited ASP+ uptake, ofwhich acrolein required the lowest concentration to provoke ahalf-maximal inhibition (IC50=44±2 μM). A Dixon plot wasconstructed for acrolein using three independent inhibitioncurves with 10, 20, or 30 μM ASP+, which demonstratedcompetitive or mixed type of interaction (Ki=93 ± 16 μM).Exposing the cells to a mixture of cationic uremic toxinsresulted in a more potent and biphasic inhibitory responsecurve, indicating complex interactions between the toxinsand ASP+ uptake. In conclusion, ciPTEC proves a suitablemodel to study cationic xenobiotic interactions. Inhibitionof cellular uptake transport was demonstrated for several
The manuscript was written through contributions of all authors. Allauthors have given approval to the final version of the manuscript.
Carolien M.S. Schophuizen, Martijn J. Wilmer, Lambert P. van denHeuvel, and Rosalinde Masereeuw contributed equally to this work.
Electronic supplementary material The online version of this article(doi:10.1007/s00424-013-1307-z) contains supplementary material,which is available to authorized users.
C. M. S. Schophuizen : L. P. van den HeuvelDepartment of Pediatric Nephrology, Radboud UniversityNijmegen Medical Centre, 6525 GA Nijmegen, The Netherlands
M. J. Wilmer : J. Jansen : R. Masereeuw (*)Department of Pharmacology and Toxicology (149),Nijmegen Centre for Molecular Life Sciences/Institute for Geneticand Metabolic Disease, Radboud University Nijmegen MedicalCentre, P.O. Box 9101, 6500 HB Nijmegen, The Netherlandse-mail: [email protected]
J. Jansen : J. G. J. HoenderopDepartment of Physiology, Radboud University Nijmegen MedicalCentre, 6525 GA Nijmegen, The Netherlands
L. GustavssonDepartment of Laboratory Medicine (Malmö), Lund University,Skåne University Hospital, 205 02 Malmö, Sweden
C. HilgendorfAstraZeneca R&D, Innovative Medicines, Global DMPK,431 83 Mölndal, Sweden
L. P. van den HeuvelDepartment of Pediatrics, Catholic University Leuven,3000 Leuven, Belgium
Pflugers Arch - Eur J Physiol (2013) 465:1701–1714DOI 10.1007/s00424-013-1307-z

uremic toxins, which might indicate a possible role in kidneydisease progression during uremia.
Keywords Human proximal tubule cell . OCT . Uremictoxin . Polyamines . Guanidine . Acrolein
AbbreviationsASP+ 4-(4-(Dimethylamino)styryl)-N-
methylpyridinium-iodideciPTEC Conditionally immortalized proximal tubule
epithelial cellsCKD Chronic kidney diseaseDMEM Dulbecco's modified eagle mediumESRD End-stage renal diseaseFCS Fetal calf serumIC50 Half-maximal inhibitory concentrationMATE Multidrug and toxic compound extrusion proteinMTT (3-(4,5-Dimethylthiazol-2-yl)-2,5-
diphenyltetrazolium bromideOCT Organic cation transporterRLU Relative light unitsTPA Tetrapentylammonium
Introduction
In patients suffering from end-stage renal disease (ESRD),kidney function is severely impaired. As a consequence,waste products from normal metabolism are insufficientlycleared from the circulation, resulting in accumulation ofuremic retention solutes in the patient [19, 42]. Although inESRD soluble xenobiotics can be cleared by dialysis, theclearance of small lipid soluble and/or protein bound toxinsusing currently available techniques is insufficient [24]. Theaccumulation of these uremic toxins contributes to the de-velopment of uremic syndrome, resulting in increased mor-bidity and mortality [23]. In addition, drug disposition isaffected in ESRD patients due to altered pharmacokinetics.Renal failure has been found to affect both renal and non-renal toxin elimination routes [25]. Downregulation of var-ious renal and hepatic drug influx transporters, such asorganic anion transporters, p-glycoprotein, multidrug resis-tance proteins, and OCT2, has been observed in ERSD [30,39]. Insights in the mechanism of uremic toxin clearance inhumans could aid us in the search for new therapeutic pos-sibilities for patients on dialysis.
The human kidney expresses transporters involved in ure-mic toxin clearance. A system involved in renal tubular uptakeare the OCT proteins, for which three isoforms (OCT1-3)have been identified [28].The OCT2 (SLC22A2) is abun-dantly expressed in the proximal tubule of human kidney
and is considered essential for the removal of potentiallytoxic cationic compounds from the circulation, includingdrugs such as cimetidine (H2 receptor antagonist), metformin(antidiabetic), and various antibiotics and antihistamines [18,48]. These compounds can cause toxicity via drug–drug in-teractions due to competitive inhibition caused by the limitedtransport capacity of OCT2 [9]. Furthermore, OCT2 is adeterminant in cisplatin-induced nephrotoxicity due to its rolein drug uptake into renal proximal tubular cells [3]. Next toOCT2, the OCT1 (SLC22A1) and 3 (SLC22A3) isoforms arealso involved in cation uptake. While the expression of OCT1andOCT2 is mainly restricted to the liver and kidney, OCT3 ismore widely expressed in various tissues [28]. In normalfunctioning kidneys, the first step in the urinary excretion ofcationic uremic toxins might involve renal tubular cell uptakemediated by OCTs. Indeed, in renal failure several organiccations, such as guanidino compounds and polyamines thatare derived from amino acids metabolism, have been found toaccumulate in plasma of uremic patients [6].
The guanidino compounds guanidine and methylguanidineare part of a large group of structural metabolites of arginine.Guanidino compounds are small, water-soluble cations andnot bound to plasma proteins. Despite this, their dialyticremoval is insufficient [8]. The accumulation of guanidineand methylguanidine is known to affect hematopoietic cellfunctioning and can induce neuronal and cardiovascular ad-verse events [4, 10, 11, 22].
The polyamines, cadaverine, putrescine, spermine, sper-midine, and their breakdown product acrolein, are all proteinbound solutes and therefore not freely filtered by the glo-merulus. As a result, these solutes are insufficiently clearedthrough dialysis in ESRD [6]. These polyamines are formedduring cellular lysine, arginine, and ornithine catabolism. Inhumans, serum levels of free polyamines are relatively lowas the polyamines are predominantly localized intracellularly[40]. Spermine, spermidine, putrescine, and cadaverine arereported to inhibit erythropoiesis and erythroid colony for-mation in a dose-dependent manner [20, 31]. High levels ofputrescine were reported to inhibit cell growth in vitro and toalter cytoplasmic, mitochondrial, and nuclear membranestructures [38].
In the present study, we examined the interaction of theseclasses of cationic uremic retention solutes with OCT-mediated transport in a human conditionally immortalizedproximal tubule epithelial cell model (ciPTEC) [46]. To thisend, we characterized the interaction of known substratesfor OCT2 and studied the inhibitory potency of theguanidino compounds and polyamines on OCT-mediatedtransport. Furthermore, the cytotoxic potential of the cat-ionic uremic retention solutes was evaluated, as well as theinteraction of a cationic uremic toxin mixture, mimickinguremic conditions.
1702 Pflugers Arch - Eur J Physiol (2013) 465:1701–1714

Materials and methods
Chemicals
4-(4-(Dimethylamino)styryl)-N-methylpyridinium iodide(ASP+) was purchased from Invitrogen (Eugene, OR,USA). Tetrapentylammoniumchloride (TPA), metformin,quinidine, cimetidine hydrochloride, spermine, spermidine,cadaverine, putrescine dihydrochloride, acrolein, guanidinehydrochloride, methylguanidine hydrochloride, insulin, trans-ferrin, selenium, tri-iodothyronine, hydrocortisone, and epi-dermal growth factor were purchased from Sigma-Aldrich Co.(Zwijndrecht, the Netherlands).
Cell culture
The immortalized human proximal tubule epithelial cell lineciPTEC was developed as previously described [46].ciPTEC were cultured in Dulbecco's modified eagle mediumDMEM-HAM’s F12 (Lonza, Basel, Switzerland) containing10 % v/v fetal calf serum (FCS) (Greiner Bio-One, Alphena/d Rijn, the Netherlands), 5 μg/ml insulin, 5 μg/ml trans-ferrin, 5 ng/ml selenium, 36 ng/ml hydrocortisone, 10 ng/mlepidermal growth factor, and 40 pg/ml tri-iodothyronine.Cells were cultured in the absence of phenol red and antibi-otics. Culture of ciPTEC was done for up to 40 passages, forwhich proximal tubular characteristics remained unaltered[46]. Cells were seeded at 30 % density in tissue cultureflasks and cultured for 24 h at 33 °C 5 % v/v CO2, followedby maturation of the cells at 37 °C 5 % v/v CO2 for 7 daysprior to experiments.
RNA expression of OCT1, 2, and 3 transporters in ciPTEC
Total RNAwas isolated from proliferating and matured cellsof interest using TRIzol (Life Technologies Europe BV) andchloroform extraction according to the manufacturers’ proto-col. The Omniscript RT kit (Qiagen, Venlo, The Netherlands)was used to synthesize cDNA. The mRNA expressionlevels were detected using gene-specific primer-probe setsobtained from Applied Biosystems® [OCT1 (SLC22A1)Hs00427552_m1, OCT2 (SLC22A2) hs01010723_m1, andOCT3 (SLC22A3) Hs01009568_m1; Applied Biosystems,CA, USA] and TaqMan Universal PCR Master Mix(Applied Biosystems). The quantitative PCR reactions wereperformed using the CFX96 Real Time PCR system (Bio-RadLaboratories, Veenendaal, The Netherlands), and data wereanalyzed using the CFX ManagerTM software (Bio-RadLaboratories). Data were normalized to mRNA expressionlevels of the reference gene GAPDH. Data were expressedas fold increase compared to proliferating ciPTEC.
Polyacrylamide gel electrophoresis and Western blotting
Cellular homogenates of matured ciPTEC were obtainedby scraping cells off a 75-cm2 tissue culture flask by using arubber policeman and lysis in 400 μl RIPA buffer containing1 % Igepal CA630, 0.5 % Na–deoxycholate, 0.1 % SDS,0.01 % phenylmethane sulfonylfluoride, 3 % aprotinin, and1 mM Na–orthovanadate. Western blotting was done usingreduced 12 % sodium dodecyl sulfate polyacrylamidegel electrophoresis (SDS–PAGE) and transferred onto apolyvinylidene difluoride membrane (Immobilon; Millipore,Bedford, MA, USA). Expression of OCT2 in cell homoge-nates was analyzed using a rabbit anti-OCT2 (1:500; AlphaDiagnostics, San Antonio, TX, USA), followed by a goat-anti-rabbit–HRP conjugate (DAKO). As a loading control,the household gene rabbit anti-β-Actin was used (1:10,000;Alpha Diagnostics). Human kidney homogenate in RIPAbuffer was used as a control.
OCT-mediated ASP+ transport
OCT activity was measured in freshly harvested maturedcell suspensions by addition of the fluorescent OCT sub-strate ASP+ in HEPES–TRIS [Hepes–Tris (10 mM), NaCl(132 mM), KCl (4.2 mM), CaCl2 (1 mM), MgCl2 (1 mM),and D-glucose (5.5 mM), pH 7.4] buffer at 37 °C, as de-scribed before [45, 46]. The use of this suspension assayallows for high-throughput compound screening. We com-pared the ASP+ uptake by ciPTEC using this suspensionassay at 37 °C and 4 °C in order to investigate the feasibilityof this method as cation influx screening assay (results aregiven in Online Resource 1). To validate the cation influxmeasurement further in ciPTEC, cells were pre-incubatedfor 30 min with compounds known for their OCT2 inhibi-tory effects—tetrapentylammonium (TPA), cimetidine, met-formin, quinidine, or verapamil at a concentration range of5–25,000 μM—prior to co-incubation with ASP+ (25 μM)for an additional 20 min. Exposure to various uremictoxins—spermine, spermidine, cadaverine, putrescine, acrole-in, guanidine, and methylguanidine—was performed accord-ingly within a concentration range of 0.5–5000 μM. Thefluorescence intensity was monitored on a Cytofluor 4000Fluorescence/Bioluminescence Reader (excitation 485 nm,emission 590 nm) for the complete incubation period. ASP+
uptake was calculated by subtracting the initial fluorescence att=0 from the endpoint. Uptake was linear for at least up to60 min (results are given in Online Resource 2). If significantmaximal inhibition of ASP+ uptake was measured, the inhibi-tion constant (Ki) was determined by performing the uptakewith ASP+ at 10, 20, and 30 μM in addition to the respectiveinhibitor at the concentration ranges indicated.
Pflugers Arch - Eur J Physiol (2013) 465:1701–1714 1703
Additionally, we tested the effect of a combination ofuremic toxins on ASP+ uptake, which represents more close-ly the situation in uremic patients. To mimic uremic condi-tions, a mixture of toxins was used comparable to 1,000, 100,10, 1, 0.1, and 0.01 times the uremic plasma concentrationsreported in literature (Table 2; spermidine 0.67 μM, spermine0.09 μM, cadaverine 0.21 μM, putrescine 0.88 μM, acrolein1.42 μM, guanidine 2.18 μM, and methylguanidine 7.66 μM)with which cells were pre-incubated for 30 min followed byASP+ uptake measurements as described.
MTT assay
An (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bro-mide (MTT) cell proliferation assay was performed to measurecellular toxicity of the compounds tested on the ciPTEC [26].In short, cells were seeded onto a 96-well plate at 30 %confluence. After maturation at 37 °C for 7 days, the culturemediumwas removed and the cells were exposed for 30 min touremic toxins dissolved in culture medium at the concentration
range used in the uptake assays. The medium was changed to5 mg/ml MTT in PBS and the plate was incubated for 4 h at37 °C. The intracellular accumulated precipitate was solubi-lized in 200 μl dimethylsulfoxide. The absorbance was readusing a Dynatech MR 580 plate reader using a wavelength of570 nm from which background was subtracted.
Data analysis
Nonlinear regression analysis was performed using GraphPadPrism version 5.00 for Windows. A sigmoidal or biphasicdose–response model was applied, depending on best fit (de-termined using the extra sum-of-squares F test). For the ure-mic toxins, a constraint was applied limiting the top of thecurve to a maximum of 100 %. To enable detection of partialinhibition, no constraint was applied to the bottom of thecurve. The Ki values were calculated by determining the in-tersections from three separate experiments. Lines of best fitwere determined by least square linear regression analysis.Transport inhibition studies were performed in triplicate and
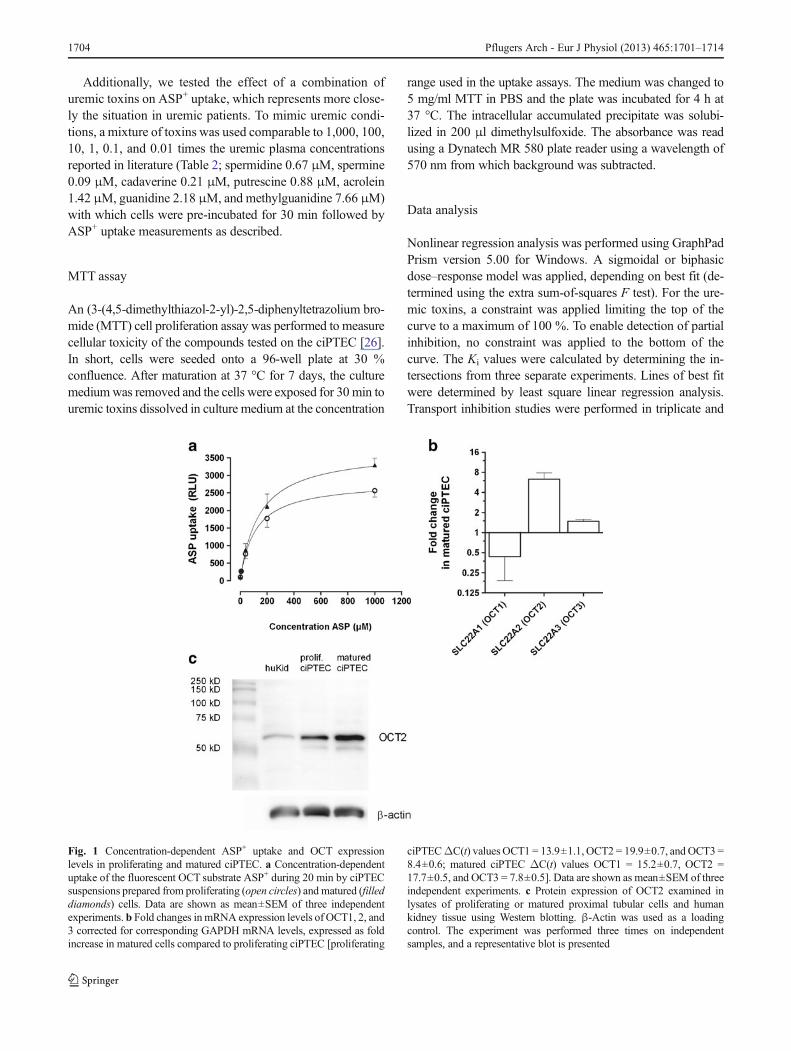
Fig. 1 Concentration-dependent ASP+ uptake and OCT expressionlevels in proliferating and matured ciPTEC. a Concentration-dependentuptake of the fluorescent OCT substrate ASP+ during 20 min by ciPTECsuspensions prepared from proliferating (open circles) and matured (filleddiamonds) cells. Data are shown as mean±SEM of three independentexperiments. b Fold changes inmRNA expression levels of OCT1, 2, and3 corrected for corresponding GAPDH mRNA levels, expressed as foldincrease in matured cells compared to proliferating ciPTEC [proliferating
ciPTECΔC(t) valuesOCT1 = 13.9±1.1, OCT2 = 19.9±0.7, and OCT3 =8.4±0.6; matured ciPTEC ΔC(t) values OCT1 = 15.2±0.7, OCT2 =17.7±0.5, and OCT3 = 7.8±0.5]. Data are shown as mean±SEM of threeindependent experiments. c Protein expression of OCT2 examined inlysates of proliferating or matured proximal tubular cells and humankidney tissue using Western blotting. β-Actin was used as a loadingcontrol. The experiment was performed three times on independentsamples, and a representative blot is presented
1704 Pflugers Arch - Eur J Physiol (2013) 465:1701–1714
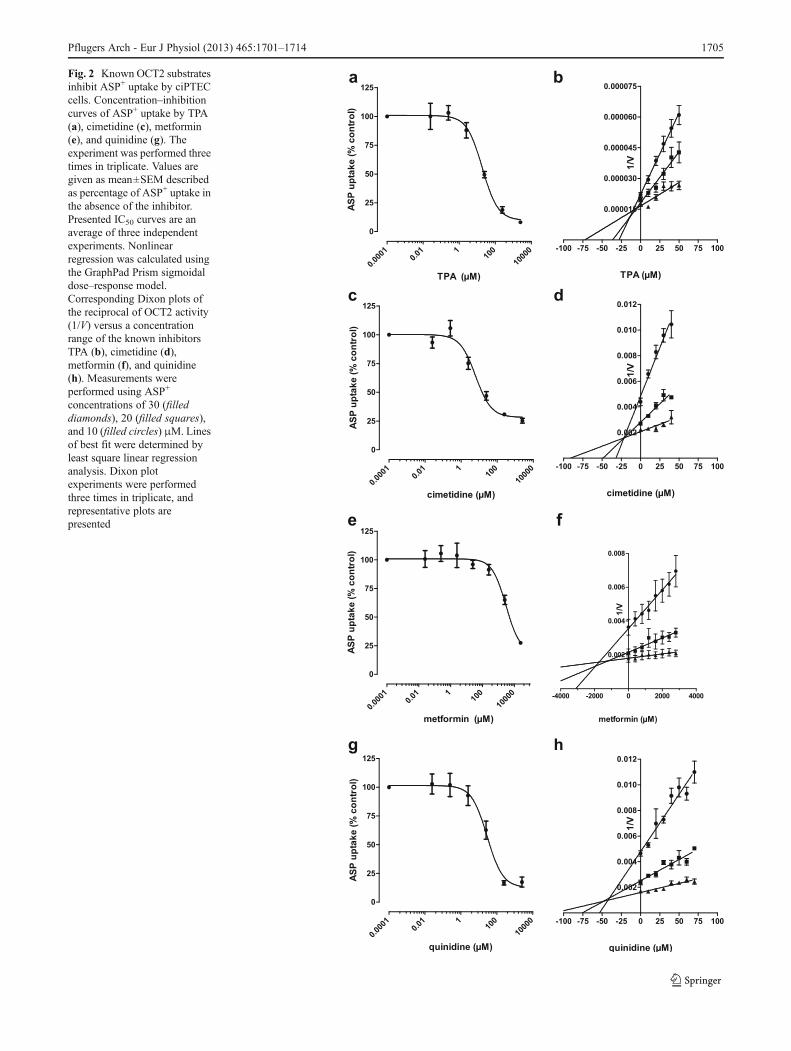
Fig. 2 Known OCT2 substratesinhibit ASP+ uptake by ciPTECcells. Concentration–inhibitioncurves of ASP+ uptake by TPA(a), cimetidine (c), metformin(e), and quinidine (g). Theexperiment was performed threetimes in triplicate. Values aregiven as mean±SEM describedas percentage of ASP+ uptake inthe absence of the inhibitor.Presented IC50 curves are anaverage of three independentexperiments. Nonlinearregression was calculated usingthe GraphPad Prism sigmoidaldose–response model.Corresponding Dixon plots ofthe reciprocal of OCT2 activity(1/V) versus a concentrationrange of the known inhibitorsTPA (b), cimetidine (d),metformin (f), and quinidine(h). Measurements wereperformed using ASP+
concentrations of 30 (filleddiamonds), 20 (filled squares),and 10 (filled circles) μM. Linesof best fit were determined byleast square linear regressionanalysis. Dixon plotexperiments were performedthree times in triplicate, andrepresentative plots arepresented
Pflugers Arch - Eur J Physiol (2013) 465:1701–1714 1705

repeated at least three times. From the MTT assay, viabilitywas expressed as a percentage of the values obtained fromuntreated control cells within the same experiment. MTTassays were performed in triplicate and repeated at leastthree times.
Statistics were performed using GraphPad Prism 5.00 viaan unpaired Student’s t test or one-way analysis of variancefollowed by Dunnet’s multiple comparison test where appro-priate. Differences between groups were considered to bestatistically significant when P <0.05.
Results
ASP+ uptake by ciPTEC is concentration dependentand sensitive to inhibition by TPA, cimetidine, metformin,and quinidine
Wilmer et al. recently showed functional expression of theOCT2 transporter in the ciPTEC model [46]. To validatefurther the ciPTEC line as a human kidney cell model suit-able to study cation transport, the uptake of the substrateASP+ was evaluated using a fluorescence-based method [36,45]. Figure 1 demonstrates the concentration-dependentASP+ uptake and OCT expression levels in proliferatingand matured ciPTEC. A 6.3±1.4-fold (mean±SEM) increasein OCT2 mRNA and elevated protein expression levels wereobserved in matured ciPTEC compared to proliferating cellsat 33 °C, while changes less than 2-fold were observed forOCT1 and OCT3. The increased expression for OCT2 wasfurther demonstrated on a protein level by Western blotting.Furthermore, ASP+ uptake increased with 23±5 % in fullymatured ciPTEC, with an apparent Km of 139±37 μM formatured ciPTEC versus an apparent Km of 110±28 μM in
Table 1 Apparent IC50 and Ki values for the inhibition of ASP+ uptake
by known OCT inhibitors
Known inhibitors IC50±SEM (μM) Ki±SEM (μM)
TPA 16±2 13±2
Metformin 3,954±1,245 4,271±521
Cimetidine 8±2 36±4
Quinidine 30±5 40±5
Verapamil 10±4 n.d.
n.d. not determined
Fig. 3 Chemical structures andcharges of the uremic toxinsstudied
1706 Pflugers Arch - Eur J Physiol (2013) 465:1701–1714

Fig. 4 Cationic uremic toxinsinhibit ASP+ uptake by ciPTECcells. Concentration–inhibitioncurves of ASP+ uptake by thepolyamines; spermidine (a),spermine (b), putrescine (c),cadaverine (d), and theguanidine compounds;guanidine (e) andmethylguanidine (f). In (g), theASP+ uptake is shown in thepresence of acrolein, ametabolite of spermine andspermidine. In (h), indoxylsulfate, an anionic uremic toxin,was used as a negative control.The experiment was performedthree times in triplicate. Valuesare described as percentage ofASP+ uptake in the absence ofthe inhibitor and given asmean±SEM. Nonlinearregression was calculated usingthe GraphPad Prism sigmoidaldose–response model. Foracrolein the IC50±SEM valuewas estimated at 44±2 μM. Thesymbol # indicates the detectionof a significant decrease inviability for theseconcentrations, as determinedby the MTT assay. For 5 mMcadaverine, guanidine, andacrolein, a residual cell viabilityof 87±4 %, 86±11 %, and79±10 % was observed,respectively
Pflugers Arch - Eur J Physiol (2013) 465:1701–1714 1707

proliferating ciPTEC. Taken together, these findings suggestASP uptake in matured ciPTEC is mainly influenced byOCT2.
The ciPTEC line was validated further by examining theeffects of known substrates and/or inhibitors of the trans-porter, namely, TPA, cimetidine, metformin, quinidine, andverapamil (not shown), on ASP+ uptake. The results showthat the drugs tested effectively inhibited the uptake of ASP+
by the ciPTEC line in a dose-dependent manner (Fig. 2a, c, e,and g). The calculated half-maximal inhibitory concentrationvalues (IC50) are given in Table 1. To investigate the mode ofinteraction, Dixon plots were constructed from the resultinginhibition curves (Fig. 2b, d, f, and h), which were analyzedby linear regression. These plots were consistent with acompetitive or mixed-type inhibition for all compounds test-ed; the Ki values calculated are also presented in Table 1.Additional ASP+ uptake experiments without pre-incubationof the substrates and/or inhibitors of OCT2 did not alter theobserved inhibition curves or IC50 values (data not shown).
Uremic toxins inhibit OCT-mediated transport
Cation transporters in the kidney are not only associated withexogenous xenobiotic transport (e.g., drugs) but could deter-mine metabolic waste secretion as well. Therefore, sevenuremic toxins were selected to study their effect on OCT-mediated transport in ciPTEC. Their chemical structures andcorresponding electrical charges at pH 7.4 are presented inFig. 3. The selection was based on the fact that these cationiccompounds are protein bound or compartmentalized in pa-tients with renal failure, and accumulate systemically due toinsufficient clearance with regular dialysis therapy [5].
The effect of the different toxins on ASP+ uptake ispresented in Fig. 4. The uremic toxins tested inhibitedASP+ uptake dose dependently. The IC50 value±SEM foracrolein was 44±2 μM. The inhibition data of the othercompounds did not allow for calculation of accurate IC50
values since the lower plateau of inhibition was not reached.The well-characterized anionic uremic toxin, indoxyl sulfate[29], was used to confirm the specificity of the interaction.As expected, this toxin did not affect ASP+ uptake in ciPTEC(Fig. 4h). Spermine, spermidine, and cadaverine (Fig. 4a, b,and d, respectively) inhibited uptake significantly only at thehighest concentration tested (5 mM). Of all uremic toxinstested, acrolein, the end product of both spermine andspermidine metabolism, was the most potent (Fig. 4g). Incontrast, another polyamine, putrescine, was the least potentcationic uremic toxin (Fig. 4c). Furthermore, both guanidineand methylguanidine inhibited ASP+ transport significantly,of which guanidine showed a higher potency (Fig. 4e–f,respectively).
For acrolein, the mode of interaction of this toxin with thetransporter was studied in more detail. To this end, inhibition
was determined using three different ASP+ concentrationsand a Dixon plot was constructed from the individual inhi-bition curves, which was analyzed by linear regression(Fig. 5). The Dixon plot is based on the acrolein concentra-tions within the linear part of the inhibition curve (200, 150,100, 50, 25, and 0 μM). In agreement with the known OCT2inhibitors and substrates, inhibition of ASP+ uptake by acro-lein was consistent with a competitive or mixed type ofinteraction, and a Ki value of 93±16 μM was calculated.
High concentrations of cadaverine, guanidine, and acroleincan affect cellular viability
The cytotoxic potential of the uremic toxins was investigatedby measuring ciPTEC viability after a 30-min exposureperiod, corresponding to the exposure time during theOCT2 uptake assay. Exposure to spermine, spermidine, pu-trescine, and methylguanidine did not result in reduced cellviability. For the other uremic toxins, a reduction in cellviability was observed only at a concentration of 5 mM(represented by # in Fig. 4d, e, and g). For cadaverine, areduction in viability of 13±4 % was observed when ex-posed to 5 mM, guanidine resulted in 14±11 % reduction inviability, and for acrolein viability was reduced with21±10 %. It should be noted that no effect on viability wasobserved at concentrations below 5 mM.
A uremic mixture of toxins inhibits OCT-mediated transport
To determine the relevance of the observed inhibitory effectsof uremic toxins on ASP+-mediated transport, we determinedthe transport function in ciPTEC incubated with a mix oftoxins, mimicking more closely the situation in patients with
Fig. 5 Dixon plot of the reciprocal of OCT2 activity (1/V) versus aconcentration range of acrolein. Measurements were done using ASP+
concentrations of 10, 20, and 30 μM. The experiment was performedthree times in triplicate. Lines of best fit were determined by leastsquare linear regression analysis; Ki = 93±16
1708 Pflugers Arch - Eur J Physiol (2013) 465:1701–1714

renal dysfunction (Fig. 6). The polyamine, acrolein, and gua-nidine mixture was prepared at 1,000, 100, 10, 1, 0.1, and 0.01times the free uremic plasma concentrations as reported inliterature (Table 2).
No inhibition of ASP+ uptake was observed after exposingthe cells to a mixture of the polyamines (spermidine,spermine, cadaverine, and putrescine; Fig. 6a). When acroleinwas added to the mixture, a large inhibition of ASP uptakewas observed only at 1,000-fold the uremic concentration
(Fig. 6b). Exposure of ciPTEC to a mixture of only theguanidino compounds (methylguanidine and guanidine) in-duced a dose-dependent inhibition of ASP+ uptake (Fig. 6c).The observed transport inhibition at 1× the uremic concentra-tion was 19 %±6. With acrolein added to this mixture, inhi-bition did not increase any further (Fig. 6d).
Interestingly, after exposing the ciPTEC to a mixture of allexamined cationic uremic toxins, a biphasic inhibition curvewas observed (Fig. 6f). The estimated IC50 values of the first
Fig. 6 A mixture of cationicuremic toxins inhibit ASP+
uptake by ciPTEC cells atphysiologically relevantconcentrations. Concentration–inhibition curves of ASP+
uptake by the followingmixtures: polyamine mix(a); polyamines and acrolein(b); guanidino mix (c);guanidino compounds andacrolein (d); polyamines andguanidino compounds(e); polyamines, acrolein, andguanidino compounds (f). Toxinconcentrations were taken at1,000, 100, 10, 1, 0.1, and 0.01times the uremic plasmaconcentrations reported inliterature (see Table 2, inμM—spermidine 0.67,spermine 0.09, cadaverine 0.21,putrescine 0.88, acrolein 1.42,guanidine 2.18, andmethylguanidine 7.66). Valuesare given as mean±SEM anddescribed as percentage ofASP+ uptake in the absence ofthe inhibition mixture.Nonlinear regression wascalculated using the GraphPadPrism sigmoidal or biphasicdose–response model. Allexperiments were performed atleast three times in triplicate
Pflugers Arch - Eur J Physiol (2013) 465:1701–1714 1709
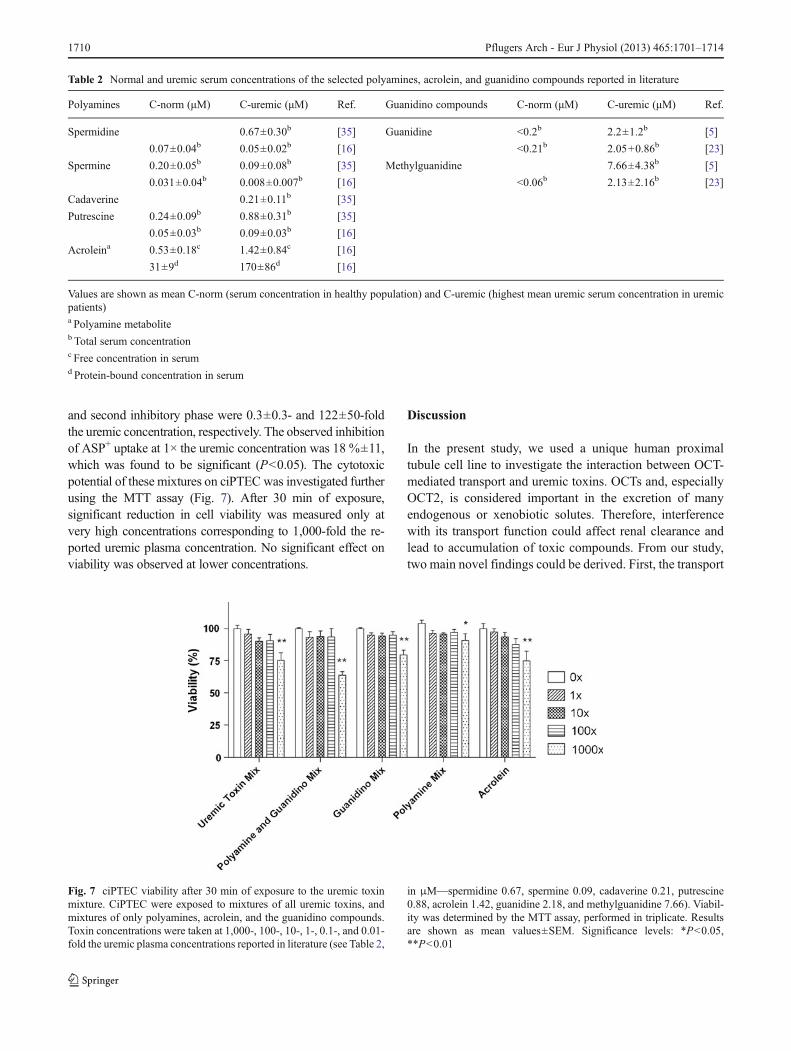
and second inhibitory phase were 0.3±0.3- and 122±50-foldthe uremic concentration, respectively. The observed inhibitionof ASP+ uptake at 1× the uremic concentration was 18 %±11,which was found to be significant (P<0.05). The cytotoxicpotential of these mixtures on ciPTEC was investigated furtherusing the MTT assay (Fig. 7). After 30 min of exposure,significant reduction in cell viability was measured only atvery high concentrations corresponding to 1,000-fold the re-ported uremic plasma concentration. No significant effect onviability was observed at lower concentrations.
Discussion
In the present study, we used a unique human proximaltubule cell line to investigate the interaction between OCT-mediated transport and uremic toxins. OCTs and, especiallyOCT2, is considered important in the excretion of manyendogenous or xenobiotic solutes. Therefore, interferencewith its transport function could affect renal clearance andlead to accumulation of toxic compounds. From our study,two main novel findings could be derived. First, the transport
Table 2 Normal and uremic serum concentrations of the selected polyamines, acrolein, and guanidino compounds reported in literature
Polyamines C-norm (μM) C-uremic (μM) Ref. Guanidino compounds C-norm (μM) C-uremic (μM) Ref.
Spermidine 0.67±0.30b [35] Guanidine <0.2b 2.2±1.2b [5]
0.07±0.04b 0.05±0.02b [16] <0.21b 2.05+0.86b [23]
Spermine 0.20±0.05b 0.09±0.08b [35] Methylguanidine 7.66±4.38b [5]
0.031±0.04b 0.008±0.007b [16] <0.06b 2.13±2.16b [23]
Cadaverine 0.21±0.11b [35]
Putrescine 0.24±0.09b 0.88±0.31b [35]
0.05±0.03b 0.09±0.03b [16]
Acroleina 0.53±0.18c 1.42±0.84c [16]
31±9d 170±86d [16]
Values are shown as mean C-norm (serum concentration in healthy population) and C-uremic (highest mean uremic serum concentration in uremicpatients)a Polyamine metaboliteb Total serum concentrationc Free concentration in serumd Protein-bound concentration in serum
Fig. 7 ciPTEC viability after 30 min of exposure to the uremic toxinmixture. CiPTEC were exposed to mixtures of all uremic toxins, andmixtures of only polyamines, acrolein, and the guanidino compounds.Toxin concentrations were taken at 1,000-, 100-, 10-, 1-, 0.1-, and 0.01-fold the uremic plasma concentrations reported in literature (see Table 2,
in μM—spermidine 0.67, spermine 0.09, cadaverine 0.21, putrescine0.88, acrolein 1.42, guanidine 2.18, and methylguanidine 7.66). Viabil-ity was determined by the MTT assay, performed in triplicate. Resultsare shown as mean values±SEM. Significance levels: *P<0.05,**P<0.01
1710 Pflugers Arch - Eur J Physiol (2013) 465:1701–1714

of and inhibition by known substrates and inhibitors could beconfirmed in our human ciPTEC model endogenously ex-pressing OCT2. Secondly, cationic uremic toxins were foundto inhibit cellular ASP+ uptake. Such interaction may con-tribute to the progression of renal failure, as active tubularsecretion is essential for the removal of cationic uremictoxins while OCTs transport capacity is finite.
Knowledge of the clearance of cationic uremic retentionsolutes in humans is limited, but the human ciPTEC cellmodel can be a valuable tool to acquire mechanistic insightsin renal cationic toxin uptake. In general, the availability ofsuitable human kidney cell models is scarce. A drawback ofmany human cell models available is their restricted endog-enous transporter expression levels and functionality [1, 16].Moreover, primary renal cell cultures that are also often usedin xenobiotic uptake studies quickly go into senescence [1].With regard to cation transporters, our ciPTEC model is nothampered by these limitations. These immortalized humancells endogenously express functional OCT2 in concert withessential organic cation transporters, such as OCT1, OCT3,p-glycoprotein (ABCB1) [46], breast cancer resistance pro-tein (ABCG2), and multidrug and toxic compound extrusionMATE proteins (SLC47) (unpublished data). The co-expressionof drug transporters at both the apical and basolateral site willallow for the development of transcellular transport assays.These assays will increase our understanding of renal excretionmechanisms, superior to the study of single transporters in overexpression models.
The observed increase in the Vmax for ASP+ uptake inmatured ciPTEC corresponds to an increased mRNA andprotein expression of OCT2. Therefore, we expect OCT2to be the dominant cation uptake transporter for cationicuremic toxins in ciPTEC. Furthermore, inhibition of ASP+
uptake by the known substrates and inhibitors TPA, cimeti-dine, metformin, and quinidine confirms functionality ofOCT-mediated uptake in ciPTEC. Though some of the testedcompounds can also function as substrates for OCT1 andOCT3, the apparent Ki values are in line with the valuesreported for other systems in which only OCT2 was over-expressed [12, 27].
In the present study, the observed Ki values for the knowninhibitors sometimes exceeded the measured IC50 values,although generally the differences are small. The variationin the numbers obtained could be influenced by, amongothers, the number of binding sites of the OCT transporters[21, 44]. In the case of cimetidine, the highest concentrationsof inhibitor (≥50 μM) might have caused slight saturation ofthe system, thereby influencing the Ki values.
This investigation demonstrates that cationic uremic toxinscan interfere with ASP+ uptake. The observed inhibitory effectof the polyamines and guanidino compounds indicate thatthese compounds may be able to affect uptake and hamperthe clearance of xenobiotics, including exogenous toxins or
drugs. The inhibition of ASP+ transport by methylguanidineand guanidine observed here is comparable to the findingsof Kimura et al. who reported the interference of variousguanidino-compounds with OCT2 using a human embryonickidney cell line (HEK293) over-expressing the transporter[17]. Using the same expression system, Winter et al. identi-fied putrescine as a substrate for OCT2 [47]. In agreement,our findings indicate that high concentrations of putrescineinhibit ASP+ transport. Furthermore, Winter et al. observedthat putrescine, spermidine, spermine, and guanidine par-tially inhibited the accumulation of agmatine, an OCT2 andMATE substrate, in hMATE-HEK cells. Although theysuggested that these compounds might influence bidirec-tional agmatine transport by hMATE1 [47], our findingsargue that interference with OCT transporters by the poly-amines might also hamper the cellular influx of these sub-strates and thereby limit the excretion and clearance of organiccations. Of course, since ASP+ is a substrate for the MATEtransporters as well, these efflux transporters could haveinfluenced our measurements [2]. However, any effect ofthese efflux transporters on the uptake of ASP+ would haveresulted in an underestimation of maximal uptake capacities inciPTEC. Future research should therefore be directed to trans-epithelial transport studies in order to clarify the overall effectof the polyamines and guanidine compounds on trans-tubularorganic cation transport. In Fig. 8, the proposed mechanism ofcationic toxin clearance in the proximal tubule is depicted.
Next to polyamines and guanidino compounds, we iden-tified acrolein (a breakdown product of spermine andspermidine [35, 37]) as an inhibitor of ASP+ uptake. Out of
Fig. 8 Scheme illustrating the proposed mechanisms of transport as-sociated with organic cation (OC+) and cationic uremic toxin (UT+)secretion by renal proximal tubule cells. Arrows indicate the direction ofnet substrate transport that occurs under physiological conditions. Solidlines depict what are believed to be the principal pathways of substratetransport; dotted lines indicate pathways that are believed to be ofsecondary importance. In vivo, organic cation transporters (OCTs)and the OATP4C1 transporter facilitate the uptake of cationic com-pounds into the cell. The MATE1/2-K and OCTN1/2 transportersfacilitate the exchange of luminal H+ for intracellular OC+. Further-more, P-gp and BCRP mediate efflux of a wide variety of substrates.Cationic uremic toxin transport is presumed to be comparable to puta-tive excretion of organic cations in the kidney. Furthermore, based onour findings accumulation of cationic uremic toxins in the blood mayresult in inhibition of OC+ uptake into the proximal tubule cell
Pflugers Arch - Eur J Physiol (2013) 465:1701–1714 1711

the tested inhibitors, acrolein required the lowest exposureconcentration to provoke a half-maximal inhibition. Its Ki
value lies within the range of acrolein levels that werereported in the plasma of uremic patients [35]. However,because we did not examine the effect of protein bindingon ASP+ uptake, we cannot speculate on the OCT interactionin ESRD, during which systemic protein binding of drugsand endogenous compounds can be altered [32, 33, 43].
Previously, Igarashi et al. showed that acrolein reducedcell growth due to cytotoxicity. Their findings suggested thataccumulation of both free and protein bound acrolein inplasma of chronic kidney disease (CKD) patients could besufficient to cause cell damage [14, 34]. Indeed, we alsoobserved decreased viability of cells exposed to acrolein;however, this was detected only at high concentrations.Therefore, we can exclude that the inhibitory effect of acroleinon ASP+ uptake is related to its cytotoxic effect. Additionally,the Dixon plot of acrolein inhibition indicated a competitive ormixed-type inhibition as the mode of interaction. Thus, it ismost likely that acrolein has a direct effect on the binding sitethat is essential for transport or functions as a substrate for thistransporter. Furthermore, we have shown that exposure toseveral compounds, including acrolein, did not lead to maxi-mal inhibition of ASP+ uptake. The presence of OCT isoformsmay explain the sub-maximal inhibition by the uremic toxins.To elucidate the exact mechanism, further subtype-specificstudies are required, for example by testing acrolein uptakein HEK cells over-expressing one specific transporter.
The systemic accumulation of uremic toxins in CKDpatients is known to negatively influence the health of thepatient and contribute to the progression of renal failure.Furthermore, interactions between uremic toxins and livertransporters, and interactions with transport proteins at theblood–brain barrier have been described [13, 39]. However,to date little data are available on the exact cationic uremictoxin levels in patients. Reports on polyamine serum valuesin patients are not always consistent, as was recentlyreviewed by Duranton et al. [7]. For instance, whereas earlierpapers reported increased spermidine levels in CKD [41],more recent publications measured decreased values in pa-tients suffering from renal failure [7, 15]. Possibly, thisdiscrepancy is due to improvements in dialysis techniquesor to methodological changes in quantification techniques.
The present findings indicate that the concentrations ofindividual uremic toxins that led to inhibition of OCT trans-port most likely exceed the systemic levels of uremic toxins inpatients and might therefore have low clinical relevance.Nevertheless, it should be noted that in uremia, a mixture ofuremic toxins is present, and the combination of retentionsolutes can have cumulative, or even synergistic, effects oncation transport in the proximal tubule. Our results showedthat exposure to a mixture of toxins for 30 min can cause up to18 % inhibition of ASP uptake in vitro when concentrations
are used that reflect the situation in uremic patients.Addition of the toxins as a mixture resulted in a strongereffect as compared to the separate exposures to these toxins.Interestingly, the biphasic dose–response curve was ob-served only when a combination of guanidine compounds,polyamines, and acrolein was studied. The biphasic natureof the uremic toxins mixture suggests different modes of actionof the individual compounds or may be related to specificinteraction with different subtypes of OCT transporters.
In conclusion, OCT-mediated organic cation transport canbe reliably studied in the ciPTEC cell model. The exposureto cationic uremic toxins was found to negatively affectsubstrate uptake in proximal tubule cells. Our findings indi-cate that the accumulation of uremic toxins may hamper theclearance of OCT substrates, which can also be an importantfactor in kidney disease progression.
Acknowledgment This work was supported by the BioMedical Ma-terials Institute (BMM, Project P3.01 BioKid); the Dutch Ministry ofEconomic Affairs, Agriculture and Innovation; the Nierstichting Neder-land; and the Netherlands Institute for Regenerative Medicine (NIRM,grant no. FES0908). M.J.G. Wilmer and R. Masereeuw were supportedby a collaboration research grant from AstraZeneca (grant no. A10-0324). J. Hoenderop was supported by an EURYI award.
Ethical standards The experiments described in this article complywith the current laws of the country in which they were performed (theNetherlands).
Conflict of interest The authors declare that they have no conflict ofinterest.
References
1. Ahlin G, Hilgendorf C, Karlsson J, Szigyarto CA, Uhlen M,Artursson P (2009) Endogenous gene and protein expression ofdrug-transporting proteins in cell lines routinely used in drug dis-covery programs. Drug Metab Dispos 37:2275–2283. doi:10.1124/dmd.109.028654
2. Astorga B, Ekins S, Morales M, Wright SH (2012) Moleculardeterminants of ligand selectivity for the human multidrug andtoxin extruder proteins mate1 and mate2-k. J Pharmacol Exp Ther341:743–755. doi:10.1124/jpet.112.191577
3. Ciarimboli G, Ludwig T, Lang D, Pavenstadt H, Koepsell H,Piechota HJ, Haier J, Jaehde U, Zisowsky J, Schlatter E (2005)Cisplatin nephrotoxicity is critically mediated via the human organiccation transporter 2. Am J Pathol 167:1477–1484. doi:10.1016/S0002-9440(10)61234-5
4. D'Hooge R, De Deyn PP, Van de Vijver G, Antoons G, Raes A, VanBogaert PP (1999) Uraemic guanidino compounds inhibit gamma-aminobutyric acid-evoked whole cell currents in mouse spinal cordneurones. Neurosci Lett 265:83–86. doi:10.1016/S0304-3940(99)00190-1
5. De Deyn PP, Robitaille P, Vanasse M, Qureshi IA, Marescau B(1995) Serum guanidino compound levels in uremic pediatric pa-tients treated with hemodialysis or continuous cycle peritonealdialysis. Correlations between nerve conduction velocities and
1712 Pflugers Arch - Eur J Physiol (2013) 465:1701–1714

altered guanidino compound concentrations. Nephrol 69:411–417.doi:10.1159/000188511
6. Dhondt A, Vanholder R, Van Biesen W, Lameire N (2000) Theremoval of uremic toxins. Kidney Int Suppl 76:S47–S59.doi:10.1046/j.1523-1755.2000.07606.x
7. Duranton F, Cohen G, De Smet R, Rodriguez M, Jankowski J,Vanholder R, Argiles A (2012) Normal and pathologic concentra-tions of uremic toxins. J Am Soc Nephrol 23:1258–1270.doi:10.1681/ASN.2011121175
8. Eloot S, van Biesen W, Dhondt A, de Smet R, Marescau B, DeDeyn PP, Verdonck P, Vanholder R (2009) Impact of increasinghaemodialysis frequency versus haemodialysis duration on remov-al of urea and guanidino compounds: a kinetic analysis. NephrolDial Transplant 24:2225–2232. doi:10.1093/ndt/gfp059
9. Fujita T, Urban TJ, Leabman MK, Fujita K, Giacomini KM (2006)Transport of drugs in the kidney by the human organic cationtransporter, oct2 and its genetic variants. J Pharm Sci 95:25–36.doi:10.1002/jps.20536
10. Giovannetti S, Cioni L, Balestri PL, Biagnini M (1968) Evidencethat guanidines and some related compounds cause haemolysis inchronic uraemia. Clin Sci 34:141–148http://www.ncbi.nlm.nih.gov/pubmed/5640680
11. Glorieux GL, Dhondt AW, Jacobs P, Van Langeraert J, LameireNH, De Deyn PP, Vanholder RC (2004) In vitro study of thepotential role of guanidines in leukocyte functions related to ath-erogenesis and infection. Kidney Int 65:2184–2192. doi:10.1111/j.1523-1755.2004.00631.x
12. Gorboulev V, Ulzheimer JC, Akhoundova A, Ulzheimer-Teuber I,Karbach U, Quester S, Baumann C, Lang F, Busch AE, Koepsell H(1997) Cloning and characterization of two human polyspecificorganic cation transporters. DNA Cell Biol 16:871–881. doi:10.1089/dna.1997.16.871
13. Hosoya K, Tachikawa M (2011) Roles of organic anion/cationtransporters at the blood–brain and blood–cerebrospinal fluid bar-riers involving uremic toxins. Clin Exp Nephrol 15:478–485.doi:10.1007/s10157-011-0460-y
14. Igarashi K, Kashiwagi K (2010) Modulation of cellular function bypolyamines. Int J Biochem Cell Biol 42:39–51. doi:10.1016/j.biocel.2009.07.009
15. Igarashi K, Ueda S, Yoshida K, Kashiwagi K (2006) Polyamines inrenal failure. Amino Acids 31:477–483. doi:10.1007/s00726-006-0264-7
16. Jenkinson SE, Chung GW, van Loon E, Bakar NS, Dalzell AM,Brown CD (2012) The limitations of renal epithelial cell line hk-2as a model of drug transporter expression and function in the prox-imal tubule. Pflugers Arch 464:601–611. doi:10.1007/s00424-012-1163-2
17. Kimura N, Masuda S, Katsura T, Inui K (2009) Transport ofguanidine compounds by human organic cation transporters, hoct1and hoct2. Biochem Pharmacol 77:1429–1436. doi:10.1016/j.bcp.2009.01.010
18. Kimura N, Okuda M, Inui K (2005) Metformin transport by renalbasolateral organic cation transporter hoct2. Pharm Res 22:255–259. doi:10.1007/s11095-004-1193-3
19. Krieter DH, Hackl A, Rodriguez A, Chenine L, Moragues HL,Lemke HD, Wanner C, Canaud B (2010) Protein-bound uraemictoxin removal in haemodialysis and post-dilution haemodiafiltration.Nephrol Dial Transplant 25:212–218. doi:10.1093/ndt/gfp437
20. Kushner D, Beckman B, Nguyen L, Chen S, Della Santina C,Husserl F, Rice J, Fisher JW (1991) Polyamines in the anemia ofend-stage renal disease. Kidney Int 39:725–732. doi:10.1038/ki.1991.88
21. Lumen AA, Acharya P, Polli JW, Ayrton A, Ellens H, Bentz J(2010) If the KI is defined by the free energy of binding toP-glycoprotein, which kinetic parameters define the IC50 for theMadin–Darby canine kidney II cell line overexpressing human
multidrug resistance 1 confluent cell monolayer? Drug MetabDispos 38:260–269. doi:10.1124/dmd.109.029843
22. MacAllister RJ, Whitley GS, Vallance P (1994) Effects ofguanidino and uremic compounds on nitric oxide pathways.Kidney Int 45:737–742. doi:10.1038/ki.1994.98
23. Meyer TW, Hostetter TH (2007) Uremia. N Engl J Med 357:1316–1325. doi:10.1056/NEJMra071313
24. Meyer TW, Walther JL, Pagtalunan ME, Martinez AW,Torkamani A, Fong PD, Recht NS, Robertson CR, Hostetter TH(2005) The clearance of protein-bound solutes by hemofiltrationand hemodiafiltration. Kidney Int 68:867–877. doi:10.1111/j.1523-1755.2005.00469.x
25. Momper JD, Venkataramanan R, Nolin TD (2010) Nonrenal drugclearance in CKD: searching for the path less traveled. Adv ChronicKidney Dis 17:384–391. doi:10.1053/j.ackd.2010.05.009
26. Mosmann T (1983) Rapid colorimetric assay for cellular growth andsurvival: application to proliferation and cytotoxicity assays. JImmunol Methods 65:55–63http://www.ncbi.nlm.nih.gov/pubmed/6606682
27. Motohashi H, Uwai Y, Hiramoto K, Okuda M, Inui K (2004)Different transport properties between famotidine and cimetidineby human renal organic ion transporters (slc22a). Eur J Pharmacol503:25–30. doi:10.1016/j.ejphar.2004.09.032
28. Nies AT, Koepsell H, Damme K, Schwab M (2011) Organic cationtransporters (OCTs, MATEs), in vitro and in vivo evidence for theimportance in drug therapy. Handb Exp Pharmacol:105–167.doi:10.1007/978-3-642-14541-4_3
29. Niwa T (2010) Uremic toxicity of indoxyl sulfate. Nagoya J MedSci 72:1–11. doi:10.1159/000057433
30. Nolin TD (2008) Altered nonrenal drug clearance in ESRD.Curr Opin Nephrol Hypertens 17:555–559. doi:10.1097/MNH.0b013e3283136732
31. Radtke HW, Rege AB, LaMarche MB, Bartos D, Bartos F,Campbell RA, Fisher JW (1981) Identification of spermine as aninhibitor of erythropoiesis in patients with chronic renal failure. JClin Invest 67:1623–1629. doi:10.1172/JCI110197
32. Reidenberg MM, Drayer DE (1984) Alteration of drug–proteinbinding in renal disease. Clin Pharmacokinet 9(Suppl 1):18–26.doi:10.2165/00003088-198400091-00003
33. Rowland Yeo K, Aarabi M, Jamei M, Rostami-Hodjegan A(2011) Modeling and predicting drug pharmacokinetics in patientswith renal impairment. Expert Rev Clin Pharmacol 4:261–274.doi:10.1586/ecp.10.143
34. Sakata K, Kashiwagi K, Sharmin S, Ueda S, Igarashi K (2003)Acrolein produced from polyamines as one of the uraemic toxins.Biochem Soc Trans 31:371–374. doi:10.1042/BST0310371
35. Sakata K, Kashiwagi K, Sharmin S, Ueda S, Irie Y, MurotaniN, Igarashi K (2003) Increase in putrescine, amine oxidase,and acrolein in plasma of renal failure patients. BiochemBiophys Res Commun 305:143–149. doi:10.1016/S0006-291x(03)00716-2
36. Schlatter E, Monnich V, Cetinkaya I, Mehrens T, Ciarimboli G,Hirsch JR, Popp C, Koepsell H (2002) The organic cation trans-porters rOCT1 and hOCT2 are inhibited by cGMP. J Membr Biol189:237–244. doi:10.1007/s00232-002-1023-7
37. Sharmin S, Sakata K, Kashiwagi K, Ueda S, Iwasaki S, ShirahataA, Igarashi K (2001) Polyamine cytotoxicity in the presence ofbovine serum amine oxidase. Biochem Biophys Res Commun282:228–235. doi:10.1006/bbrc.2001.4569
38. Stabellini G, Mariani G, Pezzetti F, Calastrini C (1997) Directinhibitory effect of uremic toxins and polyamines on proliferationof VERO culture cells. Exp Mol Pathol 64:147–155. doi:10.1006/exmp.1997.2215
39. Sun H, Frassetto L, Benet LZ (2006) Effects of renal failure on drugtransport and metabolism. Pharmacol Ther 109:1–11. doi:10.1016/j.pharmthera.2005.05.010
Pflugers Arch - Eur J Physiol (2013) 465:1701–1714 1713

40. Tadolini B, Hakim G, Orlandini G, Casti A (1986) Intracellularlocation of polyamines associated to red blood cells. BiochemBiophys Res Commun 134:1365–1371. doi:10.1016/0006-291X(86)90400-6
41. Takagi T, Chung TG, Saito A (1983) Determination of polyaminesin hydrolysates of uremic plasma by high-performance cation-exchange column chromatography. J Chromatogr 272:279–285,<Go to ISI>://A1983QE07100007
42. Vanholder R, De Smet R (1999) Pathophysiologic effects of uremicretention solutes. J Am Soc Nephrol 10:1815–1823
43. Vanholder R, Van Landschoot N, De Smet R, Schoots A,Ringoir S (1988) Drug protein binding in chronic renal fail-ure: evaluation of nine drugs. Kidney Int 33:996–1004. doi:10.1038/ki.1988.99
44. Volk C, Gorboulev V, Budiman T, Nagel G, Koepsell H (2003)Different affinities of inhibitors to the outwardly and inwardlydirected substrate binding site of organic cation transporter 2. MolPharmacol 64:1037–1047. doi:10.1124/mol.64.5.1037
45. Wilde S, Schlatter E, Koepsell H, Edemir B, Reuter S, PavenstadtH, Neugebauer U, Schroter R, Brast S, Ciarimboli G (2009)Calmodulin-associated post-translational regulation of rat organiccation transporter 2 in the kidney is gender dependent. Cell MolLife Sci 66:1729–1740. doi:10.1007/s00018-009-9145-z
46. Wilmer MJ, Saleem MA, Masereeuw R, Ni L, van der Velden TJ,Russel FG, Mathieson PW, Monnens LA, van den Heuvel LP,Levtchenko EN (2010) Novel conditionally immortalized humanproximal tubule cell line expressing functional influx and effluxtransporters. Cell Tissue Res 339:449–457. doi:10.1007/s00441-009-0882-y
47. Winter TN, Elmquist WF, Fairbanks CA (2011) Oct2 and mate1provide bidirectional agmatine transport. Mol Pharm 8:133–142.doi:10.1021/mp100180a
48. Zolk O, Solbach TF, Konig J, Fromm MF (2009) Structural de-terminants of inhibitor interaction with the human organic cationtransporter oct2 (slc22a2). Naunyn Schmiedebergs Arch Pharmacol379:337–348. doi:10.1007/s00210-008-0369-5
1714 Pflugers Arch - Eur J Physiol (2013) 465:1701–1714
Related Documents











