Vaccine 30 (2012) 254–264 Contents lists available at SciVerse ScienceDirect Vaccine j ourna l ho me pag e: www.elsevier.com/locate/vaccine Cationic liposome–DNA complexes (CLDC) adjuvant enhances the immunogenicity and cross-protective efficacy of a pre-pandemic influenza A H5N1 vaccine in mice Libo Dong a,1 , Feng Liu a,1 , Jeffery Fairman b , David K. Hong c , David B. Lewis c , Thomas Monath d , John F. Warner d , Jessica A. Belser a , Jenish Patel a , Kathy Hancock a , Jacqueline M. Katz a , Xiuhua Lu a,∗ a Centers for Disease Control and Prevention, Atlanta, GA, USA b Colby Pharmaceutical Company, Menlo Park, CA, USA c Department of Pediatrics, Interdepartmental Program in Immunology, and Institute for Immunity, Transplantation, and Infection, Stanford University, Stanford, CA, USA d Juvaris BioTherapeutics, Inc., Burlingame, CA, USA a r t i c l e i n f o Article history: Received 27 July 2011 Received in revised form 27 October 2011 Accepted 31 October 2011 Available online 12 November 2011 Keywords: Influenza A H5N1 virus Adjuvant CLDC Cross-clade protection Dose-sparing Antibody responses CD4 + T-cell immunity a b s t r a c t The development of pre-pandemic influenza A H5N1 vaccines that confer both antigen-sparing and cross- clade protection are a high priority given the limited worldwide capacity for influenza vaccine production, and the antigenic and genetic heterogeneity of circulating H5N1 viruses. The inclusion of potent adjuvants in vaccine formulations may achieve both of these aims. Here we show that the addition of JVRS-100, an adjuvant consisting of cationic liposome–DNA complexes (CLDC) to a clade 1-derived H5N1 split vaccine induced significantly higher virus-specific antibody than unadjuvanted formulations, with a >30- fold dose-sparing effect and induction of increased antigen-specific CD4 + T-cell responses in mice. All mice that received one dose of adjuvanted vaccine and subsequent H5N1 viral challenges exhibited mild illness, lower lung viral titers, undetectable spleen and brain viral titers, and 100% survival after either homologous clade 1 or heterologous clade 2 H5N1 viral challenges, whereas unadjuvanted vaccine recipients showed significantly increased weight loss, viral titers, and mortality. The protective immunity induced by JVRS-100 adjuvanted H5N1 vaccine was shown to last for over one year without significant waning. Thus, JVRS-100 adjuvanted H5N1 vaccine elicited enhanced humoral and T-cell responses, dose- sparing, and cross-clade protection in mice. CLDC holds promise as an adjuvant for human pre-pandemic inactivated H5N1 vaccines. © 2011 Elsevier Ltd. All rights reserved. 1. Introduction Influenza pandemics are unpredictable in their timing and severity, but even relatively mild pandemic strains such as the 2009 H1N1 place a burden on public health systems and result in increased morbidity and mortality in age groups not usually at risk for complications with seasonal influenza [1–4]. The 1918 pandemic caused by an avian-descended H1N1 virus killed an esti- mated 50 million people or more globally [4]. The 1957 H2N2, 1968 H3N2, and 2009 H1N1 pandemics were the result of the emer- gence of novel reassortant viruses containing genomic segments from human seasonal influenza A strains and either avian or, in the case of 2009 H1N1, both avian and swine origin viruses [5,6], which Disclaimer: The findings and conclusions in this report are those of the authors and do not necessarily represent the views of the funding agency. ∗ Corresponding author. Tel.: +1 4046395445. E-mail address: [email protected] (X. Lu). 1 These authors contributed equally to this work. rapidly spread in susceptible human populations lacking neutraliz- ing antibodies to the novel strains [7,8]. The 2009 H1N1 pandemic has not diminished the possibility that another pandemic strain will emerge from animal sources to challenge the public health infras- tructure. Highly pathogenic avian influenza (HPAI) H5N1 virus remains of particular concern, as this virus continues to circulate in wild and domestic avian populations in Asia, the Middle East, parts of Africa and Europe, and is capable of causing sporadic human disease with high mortality [9–11]. Since 1997, H5N1 viruses have diversified both genetically and antigenically into multiple clades and subclades (clades 0–9). Viruses from clades 0, 1, 2.1, 2.2, 2.3, and 7 have caused human disease [9,10]. Although both clade 1 and clade 2 viruses continue to cause outbreaks in poultry and sporadic human infections in 2010 and 2011, the majority of recent human infections have been caused by clade 2 viruses [11]. Asymptomatic infection appears to be rare even in populations with relatively high levels of exposure to infected poultry [12], and hence the majority of humans remain serologically naïve to the H5 subtype. The inability of H5N1 viruses to spread efficiently among humans remains a major barrier for the 0264-410X/$ – see front matter © 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.vaccine.2011.10.103

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

CiH
LJa
b
c
d
a
ARRAA
KIACCDAC
1
s2iapmHgfc
a
0d
Vaccine 30 (2012) 254– 264
Contents lists available at SciVerse ScienceDirect
Vaccine
j ourna l ho me pag e: www.elsev ier .com/ locate /vacc ine
ationic liposome–DNA complexes (CLDC) adjuvant enhances themmunogenicity and cross-protective efficacy of a pre-pandemic influenza A5N1 vaccine in mice�
ibo Donga,1, Feng Liua,1, Jeffery Fairmanb, David K. Hongc, David B. Lewisc, Thomas Monathd,ohn F. Warnerd, Jessica A. Belsera, Jenish Patela, Kathy Hancocka, Jacqueline M. Katza, Xiuhua Lua,∗
Centers for Disease Control and Prevention, Atlanta, GA, USAColby Pharmaceutical Company, Menlo Park, CA, USADepartment of Pediatrics, Interdepartmental Program in Immunology, and Institute for Immunity, Transplantation, and Infection, Stanford University, Stanford, CA, USAJuvaris BioTherapeutics, Inc., Burlingame, CA, USA
r t i c l e i n f o
rticle history:eceived 27 July 2011eceived in revised form 27 October 2011ccepted 31 October 2011vailable online 12 November 2011
eywords:nfluenza A H5N1 virusdjuvantLDC
a b s t r a c t
The development of pre-pandemic influenza A H5N1 vaccines that confer both antigen-sparing and cross-clade protection are a high priority given the limited worldwide capacity for influenza vaccine production,and the antigenic and genetic heterogeneity of circulating H5N1 viruses. The inclusion of potent adjuvantsin vaccine formulations may achieve both of these aims. Here we show that the addition of JVRS-100,an adjuvant consisting of cationic liposome–DNA complexes (CLDC) to a clade 1-derived H5N1 splitvaccine induced significantly higher virus-specific antibody than unadjuvanted formulations, with a >30-fold dose-sparing effect and induction of increased antigen-specific CD4+ T-cell responses in mice. Allmice that received one dose of adjuvanted vaccine and subsequent H5N1 viral challenges exhibitedmild illness, lower lung viral titers, undetectable spleen and brain viral titers, and 100% survival after
ross-clade protectionose-sparingntibody responsesD4+ T-cell immunity
either homologous clade 1 or heterologous clade 2 H5N1 viral challenges, whereas unadjuvanted vaccinerecipients showed significantly increased weight loss, viral titers, and mortality. The protective immunityinduced by JVRS-100 adjuvanted H5N1 vaccine was shown to last for over one year without significantwaning. Thus, JVRS-100 adjuvanted H5N1 vaccine elicited enhanced humoral and T-cell responses, dose-sparing, and cross-clade protection in mice. CLDC holds promise as an adjuvant for human pre-pandemic
s.
inactivated H5N1 vaccine. Introduction
Influenza pandemics are unpredictable in their timing andeverity, but even relatively mild pandemic strains such as the009 H1N1 place a burden on public health systems and result
n increased morbidity and mortality in age groups not usuallyt risk for complications with seasonal influenza [1–4]. The 1918andemic caused by an avian-descended H1N1 virus killed an esti-ated 50 million people or more globally [4]. The 1957 H2N2, 19683N2, and 2009 H1N1 pandemics were the result of the emer-
ence of novel reassortant viruses containing genomic segmentsrom human seasonal influenza A strains and either avian or, in thease of 2009 H1N1, both avian and swine origin viruses [5,6], which� Disclaimer: The findings and conclusions in this report are those of the authorsnd do not necessarily represent the views of the funding agency.∗ Corresponding author. Tel.: +1 4046395445.
E-mail address: [email protected] (X. Lu).1 These authors contributed equally to this work.
264-410X/$ – see front matter © 2011 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2011.10.103
© 2011 Elsevier Ltd. All rights reserved.
rapidly spread in susceptible human populations lacking neutraliz-ing antibodies to the novel strains [7,8]. The 2009 H1N1 pandemichas not diminished the possibility that another pandemic strain willemerge from animal sources to challenge the public health infras-tructure. Highly pathogenic avian influenza (HPAI) H5N1 virusremains of particular concern, as this virus continues to circulate inwild and domestic avian populations in Asia, the Middle East, partsof Africa and Europe, and is capable of causing sporadic humandisease with high mortality [9–11].
Since 1997, H5N1 viruses have diversified both geneticallyand antigenically into multiple clades and subclades (clades 0–9).Viruses from clades 0, 1, 2.1, 2.2, 2.3, and 7 have caused humandisease [9,10]. Although both clade 1 and clade 2 viruses continueto cause outbreaks in poultry and sporadic human infections in2010 and 2011, the majority of recent human infections have beencaused by clade 2 viruses [11]. Asymptomatic infection appears to
be rare even in populations with relatively high levels of exposureto infected poultry [12], and hence the majority of humans remainserologically naïve to the H5 subtype. The inability of H5N1 virusesto spread efficiently among humans remains a major barrier for the
cine 3
io2a
ahvjltaioevadt
imaatiadehacp
cgTacsC3sIuewTpssTbort
2
2
A(se
L. Dong et al. / Vac
nitiation of a pandemic [13,14]. Nevertheless, the rapid evolutionf H5N1 viruses and their cocirculation with human seasonal and009 H1N1 pandemic strains raise concerns that H5N1 virus maycquire genetic changes to overcome this barrier.
Since 1999, human clinical studies have evaluated the safetynd immunogenicity of pre-pandemic H5 subtype vaccines. Thereas been consistently poor immunogenicity of unadjuvanted H5accines [15–20]. Two 90 �g of hemagglutinin (HA) doses of unad-uvanted split H5N1 vaccine were required to provide an acceptableevel of seroconversion in healthy adults based on hemagglutina-ion inhibition (HAI) antibody titers [20]. Aluminum salt (alum)djuvanted split H5N1 vaccines have shown modest increases inmmunogenicity over unadjuvanted vaccines in humans, but with-ut substantial dose-sparing [21]. The squalene-based oil-in-watermulsions (such as MF59, AF03, and AS03) combined with H5N1accines increased the magnitude of the antibody response in anntigen dose-sparing manner against both the homologous andrifted H5N1 viruses in humans, but also required two immuniza-ions [22–26].
The mouse model has been extensively used for studyingmmune responses to influenza vaccination and infection. Accu-
ulating evidence from this model suggests that both magnitudend quality of immune responses are important for protectiongainst influenza infection [27–29]. Further studies have shownhat Th1/IgG2a responses are superior to Th2/IgG1 responsesn providing protection against influenza infections caused byntigenic drifted or shifted viruses in mice [29–31]. A possiblerawback of squalene-based MF59 or alum is that they primarilynhance an IgG1 antibody response through a Th2 pathway andave little effect on Th1 responses in mice [27,32]. Thus use ofn adjuvant promoting Th1 responses may result in humoral andell-mediated immune responses that may result in better cross-rotection against influenza disease in humans.
JVRS-100 is comprised of novel cationic liposome–DNAomplexes (CLDC) under development for enhancing the immuno-enicity of vaccines against infectious diseases and cancer [33–37].he DNA component of CLDC consists of the empty vector pMB75.6,
double-stranded plasmid DNA (pDNA), which contains a humanytomegalovirus enhancer/promoter, but lacks a gene insert down-tream of the promoter [38]. The cationic liposome component ofLDC consists of DOTIM (octadecenoyloxy[ethyl-2-heptadecenyl--hydroxyethyl] chloride) and cholesterol [38]. CLDC administeredystemically resulted in robust Th1 cytokines and predominantgG2a antibody responses in mice [31,35]. Pre-clinical studiessing JVRS-100 in mice and non-human primates demonstratednhanced humoral and/or T-cell responses and cross-protectionhen JVRS-100 was added to split seasonal influenza vaccines [36].
his is the first study to investigate JVRS-100 adjuvant activity for are-pandemic H5N1 vaccine. Here, we demonstrate that JVRS-100ignificantly improves immunogenicity to a clade 1-derived H5N1plit vaccine, resulting in enhanced cross-clade antibody and CD4+
-cell responses, and cross-protection in mice. The protective anti-odies induced by adjuvanted vaccine were shown to last for overne year without significant waning. The addition of JVRS-100 alsoesulted in significant antigen-sparing, a property that may extendhe limited supply of vaccine in the event of a pandemic.
. Materials and methods
.1. Viruses
HPAI H5N1 viruses used in this study were
/Vietnam/1203/2004 (VN1203, clade 1), A/Turkey/15/2006TK15, clade 2.2), and A/Anhui/1/2005 (AH1, clade 2.3). Virustocks were propagated in the allantoic cavity of 10 day-oldmbryonated chicken eggs at 37 ◦C for 26 h, and then were
0 (2012) 254– 264 255
aliquoted and stored at −70 ◦C until used. Inactivated H5N1viruses were prepared by mixing virus with �-propiolactone (BPL)(Sigma–Aldrich, St. Louis, MO, USA) at a final concentration of 0.1%at 4 ◦C overnight. Inactivation was confirmed by the absence ofvirus growth after two passages in embryonated chicken eggs.Fifty percent egg-infectious dose (EID50) titers were determinedby serial titration of virus in eggs [39]. Fifty percent lethal dose(LD50) titers were determined in mice as previously described [39].
2.2. Preparation of JVRS-100 adjuvanted H5N1 vaccine
JVRS-100 (Colby Pharmaceutical Company, Menlo Park, CA) wasmanufactured from pDNA (pMB75.6) with DOTIM/cholesterol lipo-somes as previously described [38]. The adjuvant was provided as alyophilized powder, stored at 2–8 ◦C and reconstituted prior to useby the addition of sterile water. Inactivated split egg-based VN1203clade 1 H5N1 vaccine was obtained from BEI Resources (Manassas,VA, USA). The vaccine was manufactured by Sanofi Pasteur. Vaccinewas diluted in 5% dextrose in water and then mixed with JVRS-100prior to immunization.
2.3. Immunization of mice
All animals used in this study were cared for in accordancewith the guidelines of the Committee on Care and Use of Labo-ratory Animals (Institute of Laboratory Animal Resources, NationalResearch Council) under an animal use protocol approved by CDC’sInstitutional Animal Care and Use Committee. Female BALB/c mice,6–8 weeks old (Jackson Laboratories, Bar Harbor, MA, USA) wereused in all experiments. Groups of mice were anesthetized withan intraperitoneal (i.p.) injection of 0.2 ml of Avertin (2% 2,2,2-tribromoethanol and tertiary amyl alcohol as a diluent) and wereinjected intramuscularly (i.m.) with various doses of vaccine withor without 10 �g of JVRS-100 or 50 �l of 2% of aluminum hydroxide(Alhydrogel; Superfos Biosectors, Kvistgaard, Denmark) as previ-ously described [39]. A group of mice was injected with JVRS-100alone as a negative control. The left and right hind limbs of micewere injected i.m. with 50 �l each of vaccine (total 0.1 ml) to induceoptimal immune responses in mice, as previously described [39].Mice for T cell studies received two doses one week apart.
2.4. Serological assays
Sera from blood collected from the orbital plexus weretreated with receptor-destroying enzyme from Vibrio cholerae(Denka-Seiken, Tokyo, Japan) before testing for the presence ofvirus-specific antibodies. The HAI assay was performed using 4hemagglutinating units (HAU) of BPL-inactivated H5N1 virus and1% horse red blood cells [40]. Titers of neutralizing antibody weredetermined as previously described [41]. IgG and IgG subclassresponses to H5N1 virus HA were detected by an enzyme-linkedimmunosorbent assay (ELISA) as previously described [42] exceptthat 1.0 �g/ml of purified baculovirus-expressed recombinant HA(rHA) from VN1203 virus (Protein Sciences Corporation, Meriden,CT, USA) was used to coat plates. The ELISA end-point titers wereexpressed as the highest dilution that yielded an optical density(OD) greater than two times mean OD + standard deviation (SD) ofsimilarly diluted negative control samples.
2.5. Flow cytometric analysis of influenza-specific T-cell responses
Mice were vaccinated i.m. with two doses of H5N1 vaccine
equivalent to 0.3 �g HA/mouse alone or adjuvanted with either50 �l of 2% alum or 10 �g of JVRS-100 on days 0 and 7. Spleno-cytes, harvested 10 days after vaccine boost, were stimulated with40 HAU/ml of BPL-inactivated wild-type VN1203, TK15, or AH1
2 cine 3
vtCBDwIy(m
2
asacodaVta
2
tc
3
3H
aBv1ttobVcaaabacdivHoa1II1rJc
56 L. Dong et al. / Vac
irus for 24 h, with 25 �g/ml of brefeldin-A (Sigma) added forhe last 5 h of incubation. Cells were stained with CD3-APC-Cy7,D4-PE-Cy5, and CD69-PE-Cy7 monoclonal antibodies (mAbs) (BDiosciences, Sunnyvale, CA, USA) and LIVE/DEAD® Fixable Aquaead Cell Stain (Invitrogen). Cells were fixed and permeabilizedith CytoPerm/CytoFix solution (BD Biosciences), and stained with
FN-�-PE mAb at 4 ◦C for 30 min, followed by flow cytometric anal-sis using a LSRII instrument (BD Biosciences). FlowJo softwareTreestar, Ashland, OR, USA) was used for analysis of flow cyto-
etric data.
.6. Infection of mice
Mice were anesthetized with 0.2 ml of Avertin administered i.p.,nd 50 �l of infectious virus diluted in sterile phosphate-bufferedaline was inoculated by the intranasal (i.n.) route. Intranasaldministration of this large volume in mice results in a pulmonaryhallenge [43]. Mice were infected with 1000 LD50 of VN1203, TK15,r AH1 virus. Four mice in each group were euthanized 3 or 6ays post-infection (p.i). Lung and spleen or brain was collectednd titered in eggs for infectious virus as previously described [39].irus end-point titers are expressed as log10 EID50/ml. The six to
en remaining mice in each group were observed daily for sicknessnd weight loss for 21 days p.i.
.7. Statistical analysis
Statistical significance of the data was determined by using two-ailed Student’s t-test, Fisher’s exact test or Kaplan–Meier survivalurves and log rank tests to calculate p values.
. Results
.1. Immunogenicity and dose-sparing of JVRS-100 adjuvanted5N1 vaccine
To evaluate whether JVRS-100 could enhance immunogenicitynd lead to dose-sparing effects on an H5N1 vaccine, groups of tenALB/c mice were immunized i.m. with one dose of split VN1203accine equivalent to 1.0, 0.3, 0.1, or 0.03 �g of HA/mouse with0 �g of JVRS-100. Another group of mice was immunized withhe same doses of vaccine, but without adjuvant. A negative con-rol group of mice received JVRS-100 alone. The adjuvant activityf JVRS-100, when co-administered with vaccine, was determinedy measuring the serum antibody responses to the homologousN1203 virus (Fig. 1). Sera were collected three weeks after vac-ination and tested by HAI, ELISA, and microneutralization (MN)ssays. Mice receiving JVRS-100 alone did not exhibit any HAIntibodies (Fig. 1A). Only 11 of 40 (28%) mice immunized withny dose of vaccine without adjuvant had detectable HAI anti-ody titers, and none had titers of ≥40, an HAI titer previouslyssociated with protection of mice from lethal disease followinghallenges with HPAI H5N1 viruses [39]. The administration of anyose of vaccine with JVRS-100 resulted in 100% of mice respond-
ng with greatly elevated HAI titers compared to mice receivingaccine alone (p < 0.01). All mice that received 0.1, 0.3 or 1.0 �g ofA plus JVRS-100 achieved HAI titers of ≥40 against the homol-gous virus. Sera were also tested by ELISA for HA-specific IgGntibody responses (Fig. 1B). As expected, mice receiving JVRS-00 alone did not have any detectable IgG antibody. Low levels of
gG were detected in sera of mice receiving vaccine alone, but thegG levels were greatly enhanced by the co-administration of JVRS-
00 (p < 0.01). JVRS-100 also substantially enhanced MN antibodyesponses at all antigen doses (Fig. 1C). These results indicate thatVRS-100 significantly enhanced immunogenicity of VN1203 vac-ine in mice. Administration of a decreasing amount of vaccine with0 (2012) 254– 264
JVRS-100 resulted in at least a 30-fold dose-sparing effect based onHAI, IgG, and MN antibody responses.
3.2. Cross-reactive HAI antibody responses of JVRS-100adjuvanted H5N1 vaccine
Because clade 2 viruses have caused a majority of humaninfections since 2005 [11], we next determined whether co-administration of JVRS-100 with a clade 1 vaccine could augmentcross-reactive HAI antibody responses to clade 2 viruses. Sera fromsix mice per group were collected two months after vaccinationwith one dose of clade 1 vaccine with or without JVRS-100, andwere tested for HAI antibodies to both the homologous clade 1virus (VN1203, Fig. 2A) and heterologous clade 2 viruses, TK15clade 2.2 (Fig. 2B) and AH1 clade 2.3 (Fig. 2C). Although vac-cine alone induced low levels of HAI antibody to the homologousvirus, this vaccine failed to induce significant cross-reactive HAIantibody responses to either clade 2 virus. In contrast, JVRS-100adjuvanted vaccine not only significantly enhanced HAI antibodyresponses against the homologous virus (p < 0.01), but also signif-icantly enhanced cross-reactive HAI antibody responses againstthe clade 2.2 virus at all antigen doses (p < 0.01), and againstclade 2.3 virus at antigen doses of 0.3 and 1.0 �g of HA/mouse(p < 0.01).
3.3. Protective and cross-protective efficacy of JVRS-100adjuvanted H5N1 vaccine
To evaluate protective efficacy, mice were immunized i.m.with one dose of VN1203 vaccine equivalent to 0.3 �g HA/mousewith 10 �g JVRS-100 adjuvant, the lowest vaccine dose thatshowed significantly enhanced HAI antibody responses in the pres-ence of JVRS-100 to both clade 1 and clade 2 viruses (Fig. 2).Mice receiving either vaccine alone or JVRS-100 alone wereused as controls. Sera were collected before challenge withH5N1 viruses and tested for HAI antibodies (Fig. 3). Similar tothe above experiment, formulation of vaccine with JVRS-100,compared with vaccine alone, not only significantly enhancedHAI antibody responses against the homologous virus (Fig. 3A,p < 0.01), but also significantly enhanced cross-reactive HAI anti-body responses against TK15 (Fig. 3B, p < 0.01) and AH1 viruses(Fig. 3C, p < 0.01).
Three months after vaccination, mice were infected with 1000LD50 of VN1203, TK15, or AH1 virus. Each group was then dividedinto two subgroups that were either monitored for weight lossand death for 21 days (n = 6 or 7), or were euthanized on day 3p.i. to determine the amount of infectious virus in the lungs andspleens (n = 4). Day 3 p.i. was chosen to evaluate protection frominfection because naive mice were shown previously to have sub-stantial titers of virus in both the lungs and spleens at this timepoint [39].
Protective efficacy against the homologous VN1203 virus wasaugmented by JVRS-100. As shown in Fig. 3A, all control micereceiving JVRS-100 alone died 6–8 days p.i. with high titers ofvirus in the lungs and spleens. Mice receiving vaccine withoutJVRS-100 did not show substantial virus reduction in the lungscompared with control mice (p = 0.49); but showed greater than100-fold virus reduction in the spleens (p = 0.08). Five of sevenmice receiving vaccine without adjuvant died 7–10 days p.i.;two surviving mice showed over 20% body weight loss. In con-trast, mice receiving vaccine plus JVRS-100 had over 10-fold lessvirus in their lungs compared with mice receiving vaccine alone
(p < 0.05); and virus was not detected in the spleen of these mice.The formulation of vaccine with JVRS-100 resulted in a reduced(>100-fold) virus titer in the spleen compared with mice receivingvaccine alone, although this did not achieve statistical significance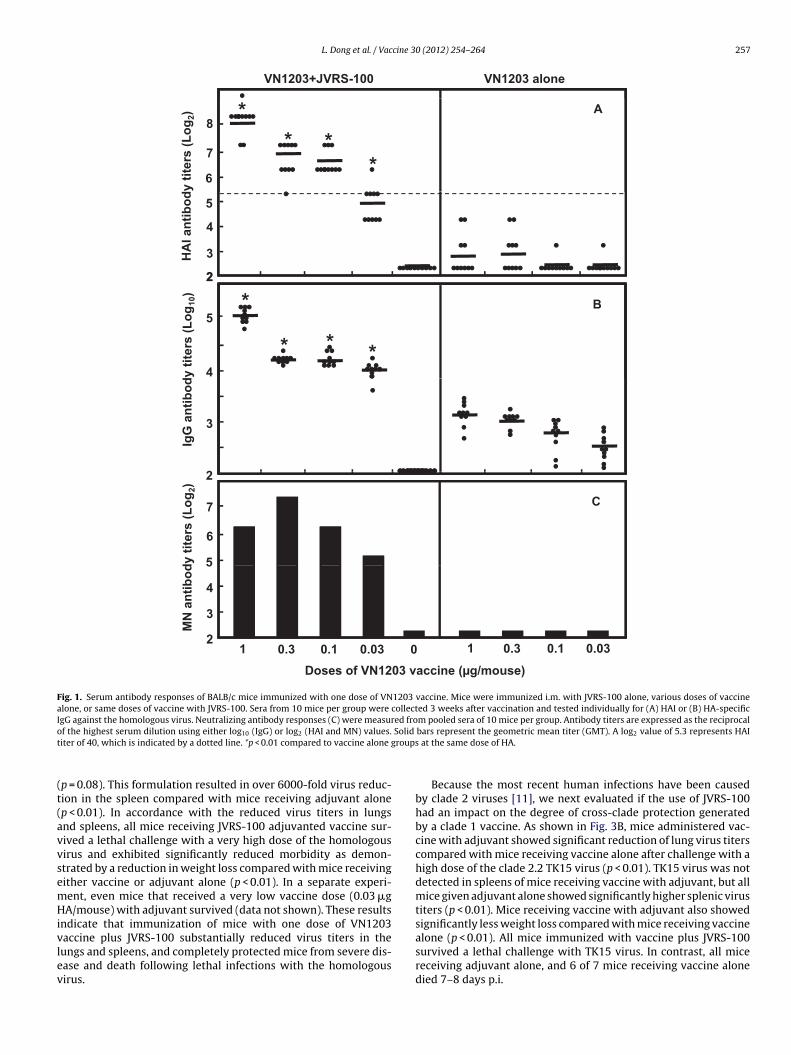
L. Dong et al. / Vaccine 30 (2012) 254– 264 257
VN1203 alone VN1203+JVRS-100
2)6
7
8
***
* A
2
HA
I ant
ibod
y tit
ers
(Log
2
3
4
510
)
4
5*
* * *
B
IgG
ant
ibod
y tit
ers
(Log
3
2)
6
7 C
2
MN
ant
ibod
y tit
ers
(Log
2
3
4
5
1
Doses of VN1203 vaccine (µg/mouse)1 0.3 0.1 0.03 0 0.3 0.1 0.03
Fig. 1. Serum antibody responses of BALB/c mice immunized with one dose of VN1203 vaccine. Mice were immunized i.m. with JVRS-100 alone, various doses of vaccinealone, or same doses of vaccine with JVRS-100. Sera from 10 mice per group were collected 3 weeks after vaccination and tested individually for (A) HAI or (B) HA-specificIgG against the homologous virus. Neutralizing antibody responses (C) were measured from pooled sera of 10 mice per group. Antibody titers are expressed as the reciprocalo Solid
t roups
(t(avvsemHivlev
f the highest serum dilution using either log10 (IgG) or log2 (HAI and MN) values.iter of 40, which is indicated by a dotted line. *p < 0.01 compared to vaccine alone g
p = 0.08). This formulation resulted in over 6000-fold virus reduc-ion in the spleen compared with mice receiving adjuvant alonep < 0.01). In accordance with the reduced virus titers in lungsnd spleens, all mice receiving JVRS-100 adjuvanted vaccine sur-ived a lethal challenge with a very high dose of the homologousirus and exhibited significantly reduced morbidity as demon-trated by a reduction in weight loss compared with mice receivingither vaccine or adjuvant alone (p < 0.01). In a separate experi-ent, even mice that received a very low vaccine dose (0.03 �gA/mouse) with adjuvant survived (data not shown). These results
ndicate that immunization of mice with one dose of VN1203
accine plus JVRS-100 substantially reduced virus titers in theungs and spleens, and completely protected mice from severe dis-ase and death following lethal infections with the homologousirus.bars represent the geometric mean titer (GMT). A log2 value of 5.3 represents HAI at the same dose of HA.
Because the most recent human infections have been causedby clade 2 viruses [11], we next evaluated if the use of JVRS-100had an impact on the degree of cross-clade protection generatedby a clade 1 vaccine. As shown in Fig. 3B, mice administered vac-cine with adjuvant showed significant reduction of lung virus titerscompared with mice receiving vaccine alone after challenge with ahigh dose of the clade 2.2 TK15 virus (p < 0.01). TK15 virus was notdetected in spleens of mice receiving vaccine with adjuvant, but allmice given adjuvant alone showed significantly higher splenic virustiters (p < 0.01). Mice receiving vaccine with adjuvant also showedsignificantly less weight loss compared with mice receiving vaccine
alone (p < 0.01). All mice immunized with vaccine plus JVRS-100survived a lethal challenge with TK15 virus. In contrast, all micereceiving adjuvant alone, and 6 of 7 mice receiving vaccine alonedied 7–8 days p.i.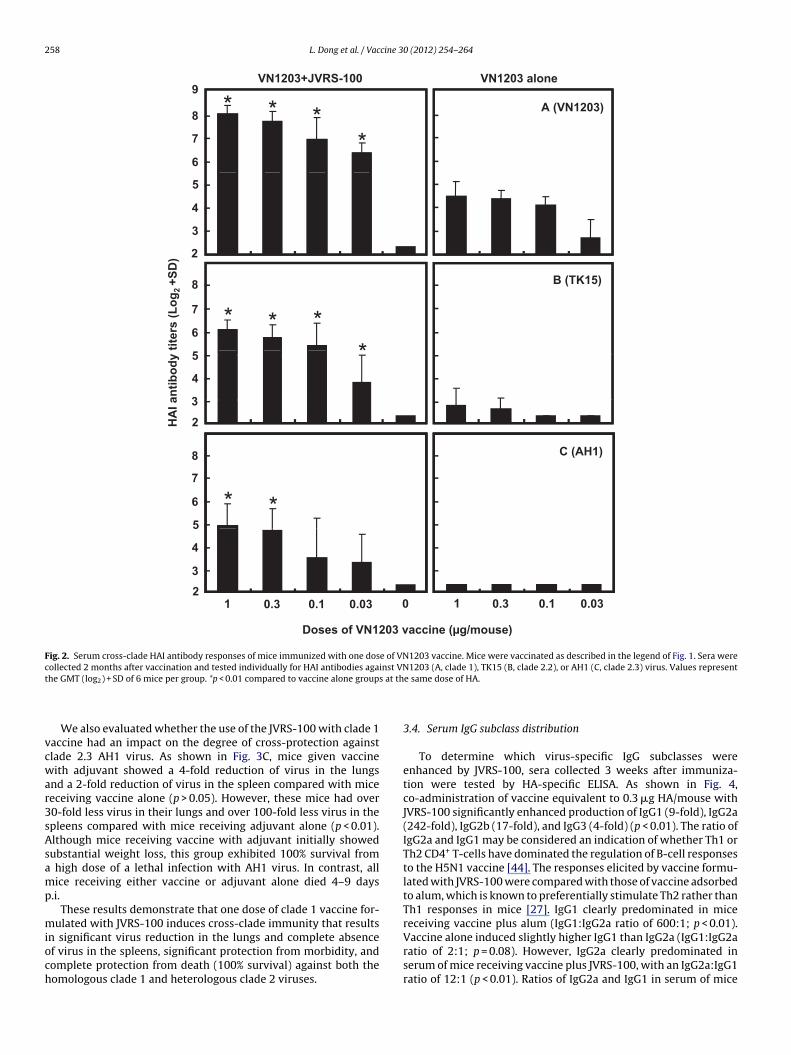
258 L. Dong et al. / Vaccine 30 (2012) 254– 264
VN1203 alone VN1203+JVRS-100
6
7
8
9A (VN1203)* * *
*
2
3
4
52
+SD
)
6
7
8 B (TK15)
****
HA
I ant
ibod
y tit
ers
(Log
2
3
4
5
6
7
8 C (AH1)
* *
23
4
5
1 0.03 0.1 0.31 0.03 0.1 0.3 0
Doses of VN1203 vaccine (µg/mouse)
Fig. 2. Serum cross-clade HAI antibody responses of mice immunized with one dose of VN1203 vaccine. Mice were vaccinated as described in the legend of Fig. 1. Sera werecollected 2 months after vaccination and tested individually for HAI antibodies against VN1203 (A, clade 1), TK15 (B, clade 2.2), or AH1 (C, clade 2.3) virus. Values representt s at th
vcwar3sAsamp
mioch
he GMT (log2) + SD of 6 mice per group. *p < 0.01 compared to vaccine alone group
We also evaluated whether the use of the JVRS-100 with clade 1accine had an impact on the degree of cross-protection againstlade 2.3 AH1 virus. As shown in Fig. 3C, mice given vaccineith adjuvant showed a 4-fold reduction of virus in the lungs
nd a 2-fold reduction of virus in the spleen compared with miceeceiving vaccine alone (p > 0.05). However, these mice had over0-fold less virus in their lungs and over 100-fold less virus in thepleens compared with mice receiving adjuvant alone (p < 0.01).lthough mice receiving vaccine with adjuvant initially showedubstantial weight loss, this group exhibited 100% survival from
high dose of a lethal infection with AH1 virus. In contrast, allice receiving either vaccine or adjuvant alone died 4–9 days
.i.These results demonstrate that one dose of clade 1 vaccine for-
ulated with JVRS-100 induces cross-clade immunity that results
n significant virus reduction in the lungs and complete absencef virus in the spleens, significant protection from morbidity, andomplete protection from death (100% survival) against both theomologous clade 1 and heterologous clade 2 viruses.e same dose of HA.
3.4. Serum IgG subclass distribution
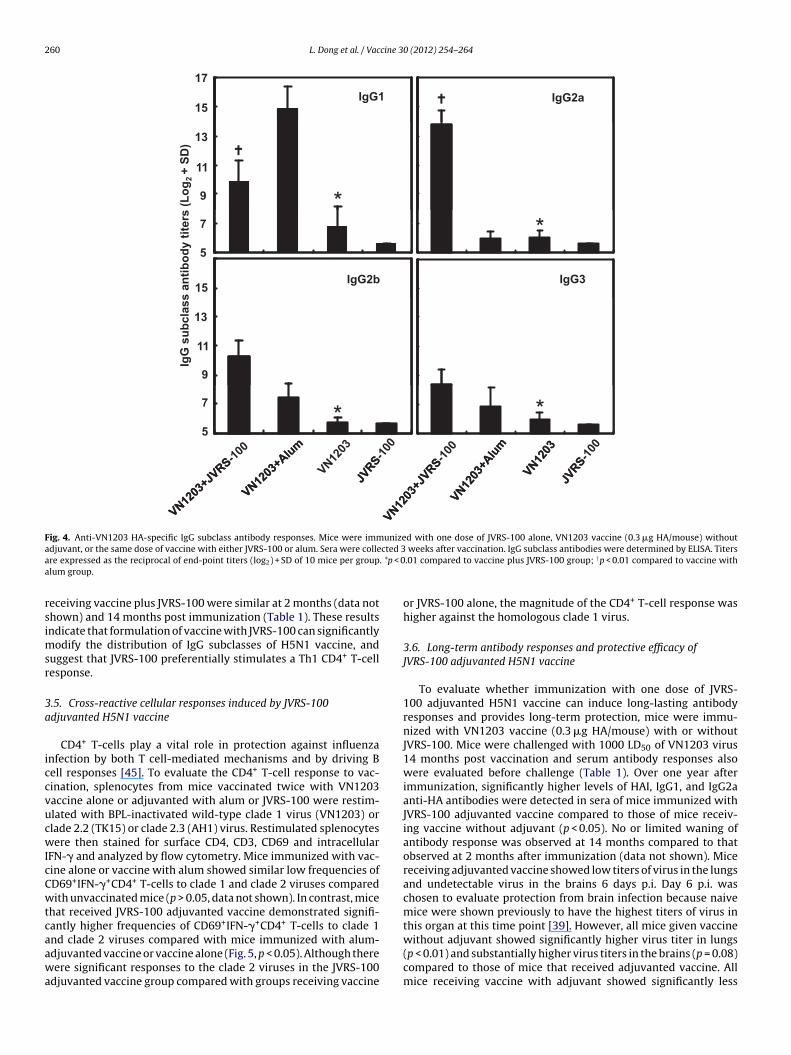
To determine which virus-specific IgG subclasses wereenhanced by JVRS-100, sera collected 3 weeks after immuniza-tion were tested by HA-specific ELISA. As shown in Fig. 4,co-administration of vaccine equivalent to 0.3 �g HA/mouse withJVRS-100 significantly enhanced production of IgG1 (9-fold), IgG2a(242-fold), IgG2b (17-fold), and IgG3 (4-fold) (p < 0.01). The ratio ofIgG2a and IgG1 may be considered an indication of whether Th1 orTh2 CD4+ T-cells have dominated the regulation of B-cell responsesto the H5N1 vaccine [44]. The responses elicited by vaccine formu-lated with JVRS-100 were compared with those of vaccine adsorbedto alum, which is known to preferentially stimulate Th2 rather thanTh1 responses in mice [27]. IgG1 clearly predominated in micereceiving vaccine plus alum (IgG1:IgG2a ratio of 600:1; p < 0.01).
Vaccine alone induced slightly higher IgG1 than IgG2a (IgG1:IgG2aratio of 2:1; p = 0.08). However, IgG2a clearly predominated inserum of mice receiving vaccine plus JVRS-100, with an IgG2a:IgG1ratio of 12:1 (p < 0.01). Ratios of IgG2a and IgG1 in serum of mice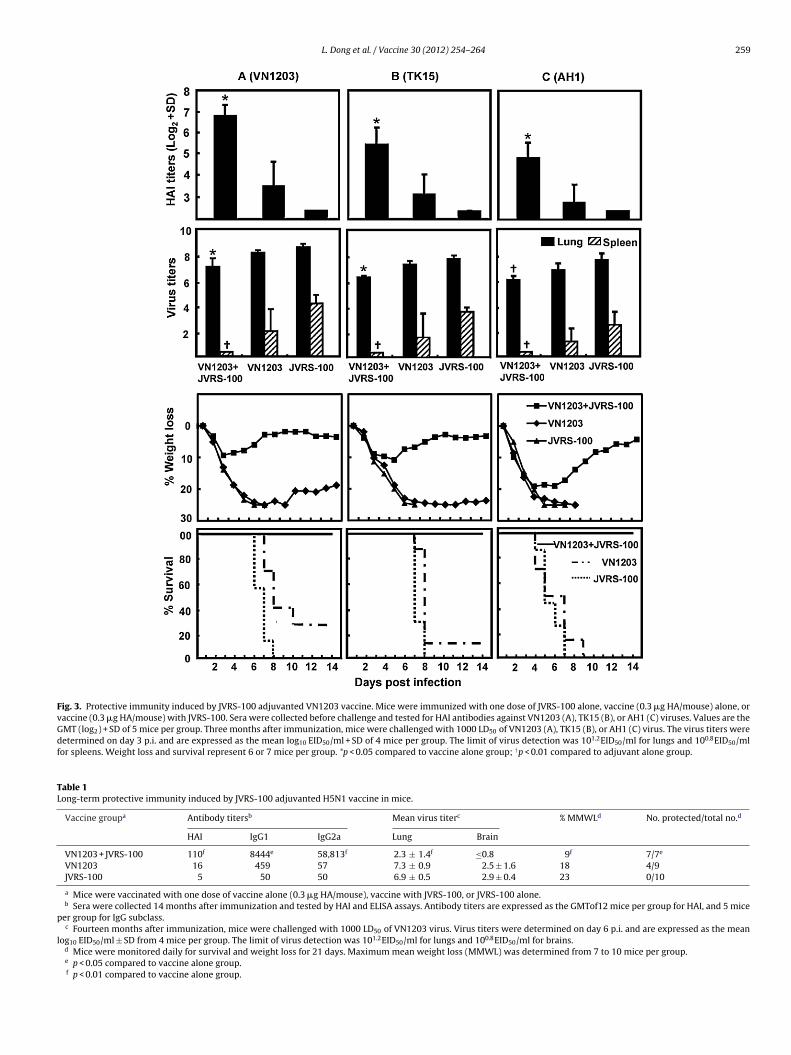
L. Dong et al. / Vaccine 30 (2012) 254– 264 259
Fig. 3. Protective immunity induced by JVRS-100 adjuvanted VN1203 vaccine. Mice were immunized with one dose of JVRS-100 alone, vaccine (0.3 �g HA/mouse) alone, orvaccine (0.3 �g HA/mouse) with JVRS-100. Sera were collected before challenge and tested for HAI antibodies against VN1203 (A), TK15 (B), or AH1 (C) viruses. Values are theGMT (log2) + SD of 5 mice per group. Three months after immunization, mice were challenged with 1000 LD50 of VN1203 (A), TK15 (B), or AH1 (C) virus. The virus titers weredetermined on day 3 p.i. and are expressed as the mean log10 EID50/ml + SD of 4 mice per group. The limit of virus detection was 101.2EID50/ml for lungs and 100.8EID50/mlfor spleens. Weight loss and survival represent 6 or 7 mice per group. *p < 0.05 compared to vaccine alone group; †p < 0.01 compared to adjuvant alone group.
Table 1Long-term protective immunity induced by JVRS-100 adjuvanted H5N1 vaccine in mice.
Vaccine groupa Antibody titersb Mean virus titerc % MMWLd No. protected/total no.d
HAI IgG1 IgG2a Lung Brain
VN1203 + JVRS-100 110f 8444e 58,813f 2.3 ± 1.4f ≤0.8 9f 7/7e
VN1203 16 459 57 7.3 ± 0.9 2.5 ± 1.6 18 4/9JVRS-100 5 50 50 6.9 ± 0.5 2.9 ± 0.4 23 0/10
a Mice were vaccinated with one dose of vaccine alone (0.3 �g HA/mouse), vaccine with JVRS-100, or JVRS-100 alone.b Sera were collected 14 months after immunization and tested by HAI and ELISA assays. Antibody titers are expressed as the GMTof12 mice per group for HAI, and 5 mice
per group for IgG subclass.c Fourteen months after immunization, mice were challenged with 1000 LD50 of VN1203 virus. Virus titers were determined on day 6 p.i. and are expressed as the mean
log10 EID50/ml ± SD from 4 mice per group. The limit of virus detection was 101.2EID50/ml for lungs and 100.8EID50/ml for brains.d Mice were monitored daily for survival and weight loss for 21 days. Maximum mean weight loss (MMWL) was determined from 7 to 10 mice per group.e p < 0.05 compared to vaccine alone group.f p < 0.01 compared to vaccine alone group.
260 L. Dong et al. / Vaccine 30 (2012) 254– 264
15
17IgG2aIgG1
2 +
SD)
9
11
13
*7
5
15IgG2b IgG3
*
IgG
sub
clas
s an
tibod
y tit
ers
(Log
9
11
13
7
5**
Fig. 4. Anti-VN1203 HA-specific IgG subclass antibody responses. Mice were immunized with one dose of JVRS-100 alone, VN1203 vaccine (0.3 �g HA/mouse) withouta cted 3a . *p < 0a
rsimsr
3a
iccvucwIcCwtcaawa
djuvant, or the same dose of vaccine with either JVRS-100 or alum. Sera were collere expressed as the reciprocal of end-point titers (log2) + SD of 10 mice per grouplum group.
eceiving vaccine plus JVRS-100 were similar at 2 months (data nothown) and 14 months post immunization (Table 1). These resultsndicate that formulation of vaccine with JVRS-100 can significantly
odify the distribution of IgG subclasses of H5N1 vaccine, anduggest that JVRS-100 preferentially stimulates a Th1 CD4+ T-cellesponse.
.5. Cross-reactive cellular responses induced by JVRS-100djuvanted H5N1 vaccine
CD4+ T-cells play a vital role in protection against influenzanfection by both T cell-mediated mechanisms and by driving Bell responses [45]. To evaluate the CD4+ T-cell response to vac-ination, splenocytes from mice vaccinated twice with VN1203accine alone or adjuvanted with alum or JVRS-100 were restim-lated with BPL-inactivated wild-type clade 1 virus (VN1203) orlade 2.2 (TK15) or clade 2.3 (AH1) virus. Restimulated splenocytesere then stained for surface CD4, CD3, CD69 and intracellular
FN-� and analyzed by flow cytometry. Mice immunized with vac-ine alone or vaccine with alum showed similar low frequencies ofD69+IFN-�+CD4+ T-cells to clade 1 and clade 2 viruses comparedith unvaccinated mice (p > 0.05, data not shown). In contrast, mice
hat received JVRS-100 adjuvanted vaccine demonstrated signifi-antly higher frequencies of CD69+IFN-�+CD4+ T-cells to clade 1
nd clade 2 viruses compared with mice immunized with alum-djuvanted vaccine or vaccine alone (Fig. 5, p < 0.05). Although thereere significant responses to the clade 2 viruses in the JVRS-100djuvanted vaccine group compared with groups receiving vaccine
weeks after vaccination. IgG subclass antibodies were determined by ELISA. Titers.01 compared to vaccine plus JVRS-100 group; †p < 0.01 compared to vaccine with
or JVRS-100 alone, the magnitude of the CD4+ T-cell response washigher against the homologous clade 1 virus.
3.6. Long-term antibody responses and protective efficacy ofJVRS-100 adjuvanted H5N1 vaccine
To evaluate whether immunization with one dose of JVRS-100 adjuvanted H5N1 vaccine can induce long-lasting antibodyresponses and provides long-term protection, mice were immu-nized with VN1203 vaccine (0.3 �g HA/mouse) with or withoutJVRS-100. Mice were challenged with 1000 LD50 of VN1203 virus14 months post vaccination and serum antibody responses alsowere evaluated before challenge (Table 1). Over one year afterimmunization, significantly higher levels of HAI, IgG1, and IgG2aanti-HA antibodies were detected in sera of mice immunized withJVRS-100 adjuvanted vaccine compared to those of mice receiv-ing vaccine without adjuvant (p < 0.05). No or limited waning ofantibody response was observed at 14 months compared to thatobserved at 2 months after immunization (data not shown). Micereceiving adjuvanted vaccine showed low titers of virus in the lungsand undetectable virus in the brains 6 days p.i. Day 6 p.i. waschosen to evaluate protection from brain infection because naivemice were shown previously to have the highest titers of virus inthis organ at this time point [39]. However, all mice given vaccine
without adjuvant showed significantly higher virus titer in lungs(p < 0.01) and substantially higher virus titers in the brains (p = 0.08)compared to those of mice that received adjuvanted vaccine. Allmice receiving vaccine with adjuvant showed significantly less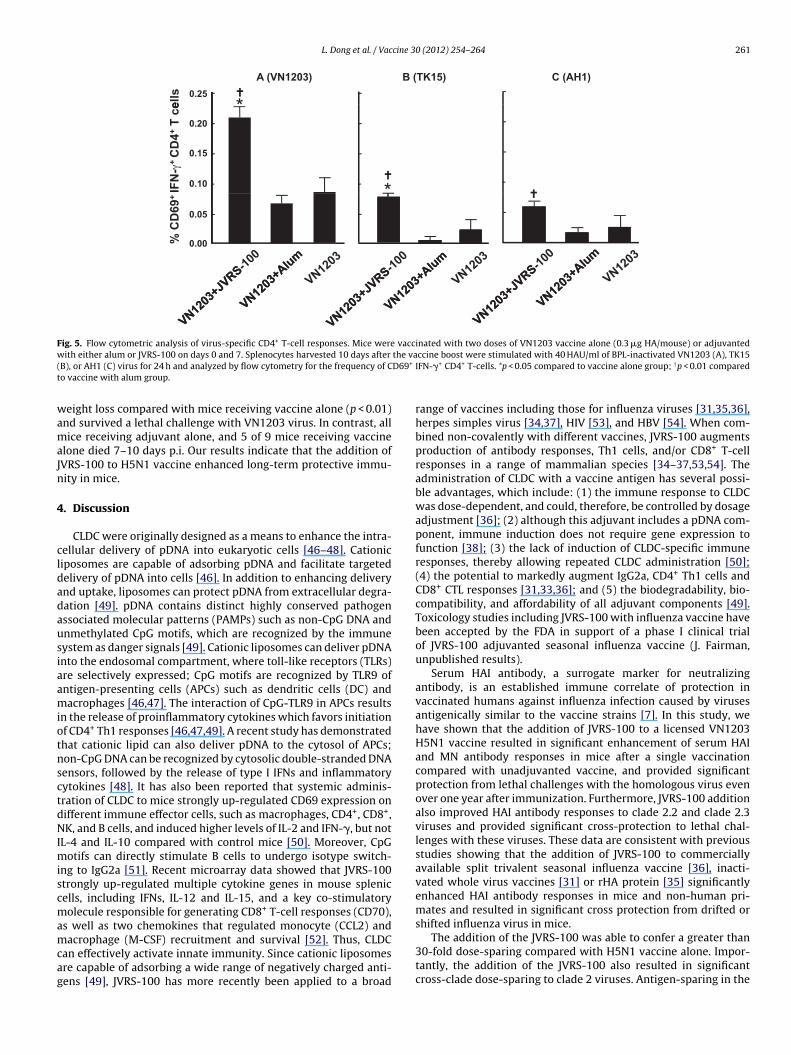
L. Dong et al. / Vaccine 30 (2012) 254– 264 261
0.25
ells
A (VN1203) B (TK15 ) C (AH1 )
*
0.10
0.15
0.20
IFN
- γ+
CD
4+T
ce *
*
0.00
0.05
% C
D69
+
Fig. 5. Flow cytometric analysis of virus-specific CD4+ T-cell responses. Mice were vaccinated with two doses of VN1203 vaccine alone (0.3 �g HA/mouse) or adjuvantedw the va( D69+
t
wamaJn
4
cldadausiaamiotnsctdNImiscmamcag
ith either alum or JVRS-100 on days 0 and 7. Splenocytes harvested 10 days after
B), or AH1 (C) virus for 24 h and analyzed by flow cytometry for the frequency of Co vaccine with alum group.
eight loss compared with mice receiving vaccine alone (p < 0.01)nd survived a lethal challenge with VN1203 virus. In contrast, allice receiving adjuvant alone, and 5 of 9 mice receiving vaccine
lone died 7–10 days p.i. Our results indicate that the addition ofVRS-100 to H5N1 vaccine enhanced long-term protective immu-ity in mice.
. Discussion
CLDC were originally designed as a means to enhance the intra-ellular delivery of pDNA into eukaryotic cells [46–48]. Cationiciposomes are capable of adsorbing pDNA and facilitate targetedelivery of pDNA into cells [46]. In addition to enhancing deliverynd uptake, liposomes can protect pDNA from extracellular degra-ation [49]. pDNA contains distinct highly conserved pathogenssociated molecular patterns (PAMPs) such as non-CpG DNA andnmethylated CpG motifs, which are recognized by the immuneystem as danger signals [49]. Cationic liposomes can deliver pDNAnto the endosomal compartment, where toll-like receptors (TLRs)re selectively expressed; CpG motifs are recognized by TLR9 ofntigen-presenting cells (APCs) such as dendritic cells (DC) andacrophages [46,47]. The interaction of CpG-TLR9 in APCs results
n the release of proinflammatory cytokines which favors initiationf CD4+ Th1 responses [46,47,49]. A recent study has demonstratedhat cationic lipid can also deliver pDNA to the cytosol of APCs;on-CpG DNA can be recognized by cytosolic double-stranded DNAensors, followed by the release of type I IFNs and inflammatoryytokines [48]. It has also been reported that systemic adminis-ration of CLDC to mice strongly up-regulated CD69 expression onifferent immune effector cells, such as macrophages, CD4+, CD8+,K, and B cells, and induced higher levels of IL-2 and IFN-�, but not
L-4 and IL-10 compared with control mice [50]. Moreover, CpGotifs can directly stimulate B cells to undergo isotype switch-
ng to IgG2a [51]. Recent microarray data showed that JVRS-100trongly up-regulated multiple cytokine genes in mouse splenicells, including IFNs, IL-12 and IL-15, and a key co-stimulatoryolecule responsible for generating CD8+ T-cell responses (CD70),
s well as two chemokines that regulated monocyte (CCL2) and
acrophage (M-CSF) recruitment and survival [52]. Thus, CLDCan effectively activate innate immunity. Since cationic liposomesre capable of adsorbing a wide range of negatively charged anti-ens [49], JVRS-100 has more recently been applied to a broad
ccine boost were stimulated with 40 HAU/ml of BPL-inactivated VN1203 (A), TK15IFN-�+ CD4+ T-cells. *p < 0.05 compared to vaccine alone group; †p < 0.01 compared
range of vaccines including those for influenza viruses [31,35,36],herpes simples virus [34,37], HIV [53], and HBV [54]. When com-bined non-covalently with different vaccines, JVRS-100 augmentsproduction of antibody responses, Th1 cells, and/or CD8+ T-cellresponses in a range of mammalian species [34–37,53,54]. Theadministration of CLDC with a vaccine antigen has several possi-ble advantages, which include: (1) the immune response to CLDCwas dose-dependent, and could, therefore, be controlled by dosageadjustment [36]; (2) although this adjuvant includes a pDNA com-ponent, immune induction does not require gene expression tofunction [38]; (3) the lack of induction of CLDC-specific immuneresponses, thereby allowing repeated CLDC administration [50];(4) the potential to markedly augment IgG2a, CD4+ Th1 cells andCD8+ CTL responses [31,33,36]; and (5) the biodegradability, bio-compatibility, and affordability of all adjuvant components [49].Toxicology studies including JVRS-100 with influenza vaccine havebeen accepted by the FDA in support of a phase I clinical trialof JVRS-100 adjuvanted seasonal influenza vaccine (J. Fairman,unpublished results).
Serum HAI antibody, a surrogate marker for neutralizingantibody, is an established immune correlate of protection invaccinated humans against influenza infection caused by virusesantigenically similar to the vaccine strains [7]. In this study, wehave shown that the addition of JVRS-100 to a licensed VN1203H5N1 vaccine resulted in significant enhancement of serum HAIand MN antibody responses in mice after a single vaccinationcompared with unadjuvanted vaccine, and provided significantprotection from lethal challenges with the homologous virus evenover one year after immunization. Furthermore, JVRS-100 additionalso improved HAI antibody responses to clade 2.2 and clade 2.3viruses and provided significant cross-protection to lethal chal-lenges with these viruses. These data are consistent with previousstudies showing that the addition of JVRS-100 to commerciallyavailable split trivalent seasonal influenza vaccine [36], inacti-vated whole virus vaccines [31] or rHA protein [35] significantlyenhanced HAI antibody responses in mice and non-human pri-mates and resulted in significant cross protection from drifted orshifted influenza virus in mice.
The addition of the JVRS-100 was able to confer a greater than30-fold dose-sparing compared with H5N1 vaccine alone. Impor-tantly, the addition of the JVRS-100 also resulted in significantcross-clade dose-sparing to clade 2 viruses. Antigen-sparing in the

2 cine 3
cmet
icdhtvdnHmvwwo[riiOcoca
casvTpcrcocwwgeiTPgdtiscr
mHalswatiCT
62 L. Dong et al. / Vac
ontext of a pandemic strain that exhibits a reduced antigenicatch to stockpiled vaccines would be highly advantageous in the
arly stages of a pandemic, since a well-matched vaccine wouldake months to prepare.
Given the importance of both antibody and T-cell responsesn the control of influenza infection, the most optimal vac-ine/adjuvant combination for protection against antigenicallyrifted and shifted viruses would substantially enhance bothumoral and T-cell immunity [55,56]. Perhaps the most impor-ant consideration for deciding what adjuvant to use in influenzaaccine is based on the type of CD4+ T-cell response required toirect the ensuing ideal immune response [57]. It is, therefore,oteworthy that the immune response to JVRS-100 adjuvanted5N1 vaccine demonstrated enhanced CD4+ Th1 responses to bothatched and drifted H5N1 viruses compared to either unadju-
anted or alum adjuvanted vaccine. Furthermore, JVRS-100 mixedith seasonal influenza vaccines enhanced CD4+ Th1 responses,hich exhibited multi-functional characteristics (i.e. co-expression
f IFN-�, IL-2, and TNF-�) in mice and/or non-human primates36]. The ability of JVRS-100 to induce an enhanced Th1/IFN-�esponse is important for development of a pre-pandemic vaccine;t may also be important for the development of an optimal seasonalnfluenza vaccine for the elderly with impaired Th1 immunity [58].n the other hand, squalene-based MF59 adjuvanted influenza vac-ine primarily enhances Th2/IgG1 responses and has little effectn Th1 responses in mice [32]. The data of the type of CD4+ Tells responses enhanced by the squalene-based AF03 and AS03djuvants in mice are lacking.
Cationic liposomes containing TLR9 CpG-containing oligonu-leotides have been shown to effectively cross-prime functionallyctive CD8+ T-cell responses [33]. Formulation of either split sea-onal influenza vaccine or inactivated seasonal whole influenzairus vaccines with JVRS-100 resulted in the induction of CD8+
-cell responses in mice and non-human primates [31,36]. In theresent study, we failed to detect CD8+ T-cell responses in vac-inated mice using BPL-inactivated wild-type H5N1 viruses toestimulate splenocytes (data not shown). Live H5N1 viruses, whichould be directly presented or cross-presented more effectivelyn MHC Class I, could not be used for restimulation due to safetyoncerns. In addition, the H5N1 vaccine strain used in this studyas generated by reverse genetics using the surface genes fromild-type H5N1 virus and internal genes derived from PR8, a high-
rowth H1N1 donor strain for vaccine production in embryonatedggs. The viral nucleoprotein (NP), M1, and PB1 proteins are signif-cant sources of epitopes inducing H5N1 virus cross-reactive CD8+
-cell responses [56]. Comparing amino acid sequences betweenR8 and VN1203 virus, NP and M1 genes were 93% identical; PB1ene was 96% identical, with considerable conservation of knownominant CTL epitopes for BALB/c mice. However, not all protec-ive epitopes are known, and even a single mutation can abolish thenteraction between the TCR and the CTL epitope [59]. Thus, furthertudies will be necessary to determine if split or whole H5N1 vac-ines made from wild-type virus adjuvanted with JVRS-100 wouldesult in the induction of CD8+ T-cell responses.
The addition of JVRS-100 to vaccine significantly decreased theorbidity and mortality associated with both matched and drifted5N1 virus challenges. One consistent observation with JVRS-100djuvanted vaccine was that the lung viral titers, following chal-enges with high doses of clade 1 or clade 2.2 or 2.3 virus, wereignificantly reduced and the viral load in the spleens and brainsas below the limit of detection. These results suggest that the
ddition of JVRS-100 to H5N1 vaccine increased the level of protec-
ive immunity sufficiently to prevent systemic infection and severenfluenza disease, even when a very high challenge dose was used.omparison of the HA1 sequence of VN1203 with those of eitherK15 or AH1 virus demonstrated a similar level of identity, 95.2%0 (2012) 254– 264
and 94.6% amino acid identity, respectively. Nevertheless, JVRS-100 adjuvanted clade 1 vaccine provided better cross-protectionagainst clade 2.2 virus than clade 2.3 virus, which is consistent withcross-reactive HAI and cross-reactive CD4+ T-cell response.
Because IgG isotypes differ in their ability to opsonize antigensand lyse infected cells, controlled induction of a specific isotypeis desirable for the design of effective vaccines [60,61]. Previousstudies in mice have shown that vaccination strategies that induceboth influenza virus-specific IgG1 and IgG2a antibodies are supe-rior to those in which IgG1 predominates [28]. IgG2a antibodiesnot only neutralize influenza viruses [62], but also interact veryefficiently with complement components [60] and activating Fcreceptors [61], which contributes to viral clearance by activation ofthe complement system, stimulation of antibody-dependent cell-mediated cytotoxicity (ADCC), and clearance of opsonized virus bymacrophages [60,63,64]. Even anti-HA IgG2a mAbs that displayedvery low neutralizing activity, cured a pulmonary influenza virusinfection in SCID mice [65]. Although IgG1 can neutralize viralparticles [62], IgG1 neither activates complement efficiently norhas a high affinity for Fc receptors [60,61]. Thus Th2/IgG1 doesnot contribute to viral clearance [29,30]. Split seasonal and pre-pandemic H5N1 vaccines stimulate mainly an IgG1 response inBALB/c mice (Fig. 4, [36]). In contrast, JVRS-100 enhanced both IgG1and IgG2a responses to influenza vaccine, but primarily enhancedIgG2a responses. This marked skewing of the antibody responsetowards IgG2a in BALB/c mice, is also was observed in C57BL/6Jmice, in which the functionally equivalent IgG2c rather than IgG2ais expressed [31]. In humans, IgG1 and IgG4 correspond to IgG2a/2cand IgG1 in the mouse [66]. High levels of Th1/IFN-� production andTLR 9 activation may both promote a switch to IgG2a [31,36,51].IgG1 has been shown to be particularly resistant to proteolysiscompared to IgG2a [67], which may enhance its stability on therespiratory epithelial surfaces facilitating a role in protection frompulmonary infection. However, IgG1 alone was shown only to pro-tect mice from a modest influenza virus challenge dose but notfrom a high dose challenge [27]. In contrast, both IgG1 and IgG2asubclasses have strong neutralizing capacity [62], which would pro-vide protection from systemic infection. Here, we found that highlevels of IgG2a and modest levels of IgG1 induced by JVRS-100 adju-vanted H5N1 vaccine could completely prevent systemic infection,but provided only limited protection from pulmonary infection,when a very high challenge dose was used.
In summary, our results indicate that JVRS-100 significantlyimproves H5N1 vaccine immunogenicity, resulting in enhancedantibody responses, CD4+ Th1 responses and protective efficaciesagainst both clade 1 and clade 2 viruses in mice. The addition ofJVRS-100 to the split H5N1 vaccine results in antigen-sparing, aproperty that may extend the limited supply of vaccine in the eventof a pandemic. H7 and H9 subtype viruses are also targets of pre-pandemic vaccines [7] which like H5 vaccines, showed reducedimmunogenicity in humans [68,69]. The ability of CLDC to enhanceavian influenza vaccine immunogenicity in humans is not known,but the adjuvant activity of CLDC is not species or antigen specific[34–37,53,54]. Therefore, JVRS-100 warrants further investigationas a potential adjuvant for use in human vaccines against theseavian influenza viruses with pandemic potential.
Acknowledgements
We thank the national public health laboratories of Vietnam,Turkey, and China for providing the human influenza A H5N1virus isolates. We thank Dr. Terry Tumpey for animal manage-
ment support. This work was supported in part by NIAID grant1U01AI074512-1 (to J. Fairman and D.B. Lewis). This work also wassupported in part by an appointment to the Research Participa-tion Program at the Centers for Diseases Control and Prevention
cine 3
ato
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
L. Dong et al. / Vac
dministered by the Oak Ridge Institute for Science and Educationhrough an interagency agreement between the U.S. Departmentf Energy and CDC (to F. Liu).
eferences
[1] Lewis DB. Avian flu to human influenza. Annu Rev Med 2006;57:139–54.[2] Viboud C, Miller M, Olson D, Osterholm M, Simonsen L. Preliminary estimates
of mortality and years of life lost associated with the 2009 A/H1N1 Pandemic inthe US and comparison with past influenza seasons. PLoS Curr 2010:RRN1153.
[3] Estimates of deaths associated with seasonal influenza – United States,1976–2007. MMWR Morb Mortal Wkly Rep 2010;59(33):1057–62.
[4] Taubenberger JK, Morens DM. Influenza: the once and future pandemic. PublicHealth Rep 2010;125(Suppl. 3):16–26.
[5] Garten RJ, Davis CT, Russell CA, Shu B, Lindstrom S, Balish A, et al. Antigenicand genetic characteristics of swine-origin 2009 A(H1N1) influenza virusescirculating in humans. Science 2009;325(5937):197–201.
[6] Kawaoka Y, Krauss S, Webster RG. Avian-to-human transmission of thePB1 gene of influenza A viruses in the 1957 and 1968 pandemics. J Virol1989;63(11):4603–8.
[7] Fiore AE, Uyeki TM, Broder K, Finelli L, Euler GL, Singleton JA, et al. Preven-tion and control of influenza with vaccines: recommendations of the AdvisoryCommittee on Immunization Practices (ACIP), 2010. MMWR Recomm Rep2010;59(RR-8):1–62.
[8] Hancock K, Veguilla V, Lu X, Zhong W, Butler EN, Sun H, et al. Cross-reactiveantibody responses to the 2009 pandemic H1N1 influenza virus. N Engl J Med2009;361(20):1945–52.
[9] Abdel-Ghafar AN, Chotpitayasunondh T, Gao Z, Hayden FG, Nguyen DH, de JongMD, et al. Update on avian influenza A (H5N1) virus infection in humans. N EnglJ Med 2008;358(3):261–73.
10] Uyeki TM. Human infection with highly pathogenic avian influenza A (H5N1)virus: review of clinical issues. Clin Infect Dis 2009;49(2):279–90.
11] WHO. Cumulative number of confirmed human cases of avian influenzaA/(H5N1) reported to WHO; 2011. Available from: Http://www.who.int/csr/disease/avian influenza/country/cases table 2011 06 22/en/index.
12] Katz JM, Veguilla V, Belser JA, Maines TR, Van Hoeven N, Pappas C, et al. Thepublic health impact of avian influenza viruses. Poult Sci 2009;88(4):872–9.
13] Cinatl Jr J, Michaelis M, Doerr HW. The threat of avian influenza A (H5N1). Part I:epidemiologic concerns and virulence determinants. Med Microbiol Immunol2007;196(4):181–90.
14] Neumann G, Kawaoka Y. Host range restriction and pathogenicity in the contextof influenza pandemic. Emerg Infect Dis 2006;12(6):881–6.
15] Sambhara S, Poland GA. H5N1 Avian influenza: preventive and therapeuticstrategies against a pandemic. Annu Rev Med 2010;61:187–98.
16] Prieto-Lara E, Llanos-Mendez A. Safety and immunogenicity of prepandemicH5N1 influenza vaccines: a systematic review of the literature. Vaccine2010;28(26):4328–34.
17] Girard MP, Katz J, Pervikov Y, Palkonyay L, Kieny MP. Report of the 6thmeeting on the evaluation of pandemic influenza vaccines in clinical trialsWorld Health Organization, Geneva, Switzerland, 17–18 February 2010. Vac-cine 2010;28(42):6811–20.
18] Bresson JL, Perronne C, Launay O, Gerdil C, Saville M, Wood J, et al. Safety andimmunogenicity of an inactivated split-virion influenza A/Vietnam/1194/2004(H5N1) vaccine: phase I randomised trial. Lancet 2006;367(9523):1657–64.
19] Nicholson KG, Colegate AE, Podda A, Stephenson I, Wood J, Ypma E, et al.Safety and antigenicity of non-adjuvanted and MF59-adjuvanted influenzaA/Duck/Singapore/97 (H5N3) vaccine: a randomised trial of two potential vac-cines against H5N1 influenza. Lancet 2001;357(9272):1937–43.
20] Treanor JJ, Campbell JD, Zangwill KM, Rowe T, Wolff M. Safety and immuno-genicity of an inactivated subvirion influenza A (H5N1) vaccine. N Engl J Med2006;354(13):1343–51.
21] WHO. Tables on the Clinical trials of pandemic influenza prototype vac-cines; 2010. Available from: http://www.who.int/vaccine research/diseases/influenza/flu trials tables/en/index.html.
22] Leroux-Roels G. Prepandemic H5N1 influenza vaccine adjuvanted withAS03: a review of the pre-clinical and clinical data. Expert Opin Biol Ther2009;9(8):1057–71.
23] Chu DW, Hwang SJ, Lim FS, Oh HM, Thongcharoen P, Yang PC, et al. Immuno-genicity and tolerability of an AS03(A)-adjuvanted prepandemic influenzavaccine: a phase III study in a large population of Asian adults. Vaccine2009;27(52):7428–35.
24] Levie K, Leroux-Roels I, Hoppenbrouwers K, Kervyn AD, Vandermeulen C,Forgus S, et al. An adjuvanted, low-dose, pandemic influenza A (H5N1) vaccinecandidate is safe, immunogenic, and induces cross-reactive immune responsesin healthy adults. J Infect Dis 2008;198(5):642–9.
25] Leroux-Roels I, Bernhard R, Gerard P, Drame M, Hanon E, Leroux-Roels G. Broadclade 2 cross-reactive immunity induced by an adjuvanted clade 1 rH5N1 pan-demic influenza vaccine. PLoS One 2008;3(2):e1665.
26] Bernstein DI, Edwards KM, Dekker CL, Belshe R, Talbot HK, Graham IL, et al.Effects of adjuvants on the safety and immunogenicity of an avian influenzaH5N1 vaccine in adults. J Infect Dis 2008;197(5):667–75.
27] Bungener L, Geeraedts F, Ter Veer W, Medema J, Wilschut J, HuckriedeA. Alum boosts TH2-type antibody responses to whole-inactivated virus
[
0 (2012) 254– 264 263
influenza vaccine in mice but does not confer superior protection. Vaccine2008;26(19):2350–9.
28] Huber VC, McKeon RM, Brackin MN, Miller LA, Keating R, Brown SA, et al. Dis-tinct contributions of vaccine-induced immunoglobulin G1 (IgG1) and IgG2aantibodies to protective immunity against influenza. Clin Vaccine Immunol2006;13(9):981–90.
29] Moran TM, Park H, Fernandez-Sesma A, Schulman JL. Th2 responses to inacti-vated influenza virus can Be converted to Th1 responses and facilitate recoveryfrom heterosubtypic virus infection. J Infect Dis 1999;180(3):579–85.
30] Graham MB, Braciale VL, Braciale TJ. Influenza virus-specific CD4+ T helper type2T lymphocytes do not promote recovery from experimental virus infection. JExp Med 1994;180(4):1273–82.
31] Hong DK, Chang S, Botham CM, Giffon TD, Fairman J, Lewis DB. Cationiclipid/DNA complex-adjuvanted influenza A virus vaccination induces robustcross-protective immunity. J Virol 2010;84(24):12691–702.
32] Valensi JP, Carlson JR, Van Nest GA. Systemic cytokine profiles in BALB/c miceimmunized with trivalent influenza vaccine containing MF59 oil emulsion andother advanced adjuvants. J Immunol 1994;153(9):4029–39.
33] Zaks K, Jordan M, Guth A, Sellins K, Kedl R, Izzo A, et al. Efficient immuniza-tion and cross-priming by vaccine adjuvants containing TLR3 or TLR9 agonistscomplexed to cationic liposomes. J Immunol 2006;176(12):7335–45.
34] Bernstein DI, Cardin RD, Bravo FJ, Strasser JE, Farley N, Chalk C, et al. Potent adju-vant activity of cationic liposome–DNA complexes for genital herpes vaccines.Clin Vaccine Immunol 2009;16(5):699–705.
35] Chang S, Warner J, Liang L, Fairman J. A novel vaccine adjuvant for recombinantflu antigens. Biologicals 2009;37(3):141–7.
36] Lay M, Callejo B, Chang S, Hong DK, Lewis DB, Carroll TD, et al. Cationic lipid/DNAcomplexes (JVRS-100) combined with influenza vaccine (Fluzone) increasesantibody response, cellular immunity, and antigenically drifted protection.Vaccine 2009;27(29):3811–20.
37] Bernstein DI, Farley N, Bravo FJ, Earwood J, McNeal M, Fairman J, et al. Theadjuvant CLDC increases protection of a herpes simplex type 2 glycoprotein Dvaccine in guinea pigs. Vaccine 2010;28(21):3748–53.
38] Gowen BB, Fairman J, Smee DF, Wong MH, Jung KH, Pace AM, et al. Protectiveimmunity against acute phleboviral infection elicited through immunostimu-latory cationic liposome–DNA complexes. Antiviral Res 2006;69(3):165–72.
39] Lu X, Tumpey TM, Morken T, Zaki SR, Cox NJ, Katz JM. A mouse model forthe evaluation of pathogenesis and immunity to influenza A (H5N1) virusesisolated from humans. J Virol 1999;73(7):5903–11.
40] Stephenson I, Wood JM, Nicholson KG, Charlett A, Zambon MC. Detectionof anti-H5 responses in human sera by HI using horse erythrocytes fol-lowing MF59-adjuvanted influenza A/Duck/Singapore/97 vaccine. Virus Res2004;103(1–2):91–5.
41] Rowe T, Abernathy RA, Hu-Primmer J, Thompson WW, Lu X, Lim W, et al. Detec-tion of antibody to avian influenza A (H5N1) virus in human serum by using acombination of serologic assays. J Clin Microbiol 1999;37(4):937–43.
42] Katz JM, Lu X, Young SA, Galphin JC. Adjuvant activity of the heat-labileenterotoxin from enterotoxigenic Escherichia coli for oral administration ofinactivated influenza virus vaccine. J Infect Dis 1997;175(2):352–63.
43] Larson EW, Dominik JW, Rowberg AH, Higbee GA. Influenza virus populationdynamics in the respiratory tract of experimentally infected mice. Infect Immun1976;13(2):438–47.
44] Mosmann TR, Coffman RL. TH1 and TH2 cells: different patterns of lym-phokine secretion lead to different functional properties. Annu Rev Immunol1989;7:145–73.
45] Brown DM, Roman E, Swain SL. CD4 T cell responses to influenza infection.Semin Immunol 2004;16(3):171–7.
46] Yasuda K, Ogawa Y, Yamane I, Nishikawa M, Takakura Y. Macrophage activa-tion by a DNA/cationic liposome complex requires endosomal acidification andTLR9-dependent and -independent pathways. J Leukoc Biol 2005;77(1):71–9.
47] Yoshinaga T, Yasuda K, Ogawa Y, Nishikawa M, Takakura Y. DNA and its cationiclipid complexes induce CpG motif-dependent activation of murine dendriticcells. Immunology 2007;120(3):295–302.
48] Yasuda S, Yoshida H, Nishikawa M, Takakura Y. Comparison of the typeof liposome involving cytokine production induced by non-CpG Lipoplex inmacrophages. Mol Pharm 2010;7(2):533–42.
49] Nordly P, Madsen HB, Nielsen HM, Foged C. Status and future prospects of lipid-based particulate delivery systems as vaccine adjuvants and their combinationwith immunostimulators. Expert Opin Drug Deliv 2009;6(7):657–72.
50] Dow SW, Fradkin LG, Liggitt DH, Willson AP, Heath TD, Potter TA. Lipid-DNAcomplexes induce potent activation of innate immune responses and antitumoractivity when administered intravenously. J Immunol 1999;163(3):1552–61.
51] Jegerlehner A, Maurer P, Bessa J, Hinton HJ, Kopf M, Bachmann MF. TLR9 sig-naling in B cells determines class switch recombination to IgG2a. J Immunol2007;178(4):2415–20.
52] Gowen BB, Fairman J, Dow S, Troyer R, Wong MH, Jung KH, et al. Prophylaxiswith cationic liposome–DNA complexes protects hamsters from phlebovi-ral disease: importance of liposomal delivery and CpG motifs. Antiviral Res2009;81(1):37–46.
53] Fairman J, Moore J, Lemieux M, Van Rompay K, Geng Y, Warner J, et al. Enhancedin vivo immunogenicity of SIV vaccine candidates with cationic liposome–DNA
complexes in a rhesus macaque pilot study. Hum Vaccin 2009;5(3):141–50.54] Cote PJ, Butler SD, George AL, Fairman J, Gerin JL, Tennant BC, et al.Rapid immunity to vaccination with woodchuck hepatitis virus surfaceantigen using cationic liposome–DNA complexes as adjuvant. J Med Virol2009;81(10):1760–72.

2 cine 3
[
[
[
[
[
[
[
[
[
[
[
[
[
[
2009;27(13):1889–97.
64 L. Dong et al. / Vac
55] Droebner K, Haasbach E, Fuchs C, Weinzierl AO, Stevanovic S, Buttner M, et al.Antibodies and CD4(+) T-cells mediate cross-protection against H5N1 influenzavirus infection in mice after vaccination with a low pathogenic H5N2 strain.Vaccine 2008;26(52):6965–74.
56] Brown LE, Kelso A. Prospects for an influenza vaccine that induces cross-protective cytotoxic T lymphocytes. Immunol Cell Biol 2009;87(4):300–8.
57] McKee AS, MacLeod MK, Kappler JW, Marrack P. Immune mechanisms of pro-tection: can adjuvants rise to the challenge? BMC Biol 2010;8:37.
58] McElhaney JE. The unmet need in the elderly: designing new influenza vaccinesfor older adults. Vaccine 2005;23(Suppl. 1):S10–25.
59] Zhong W, Liu F, Dong L, Lu X, Hancock K, Reinherz EL, et al. Significant impactof sequence variations in the nucleoprotein on CD8T cell-mediated cross-protection against influenza A virus infections. PLoS One 2010;5(5):e10583.
60] Feng JQ, Mozdzanowska K, Gerhard W. Complement component C1q enhancesthe biological activity of influenza virus hemagglutinin-specific antibodiesdepending on their fine antigen specificity and heavy-chain isotype. J Virol2002;76(3):1369–78.
61] Nimmerjahn F, Ravetch JV. Divergent immunoglobulin g subclass activitythrough selective Fc receptor binding. Science 2005;310(5753):1510–2.
62] Gerhard W, Mozdzanowska K, Furchner M, Washko G, Maiese K. Role of theB-cell response in recovery of mice from primary influenza virus infection.Immunol Rev 1997, October;159:95–103.
[
0 (2012) 254– 264
63] Nimmerjahn F, Ravetch JV. Fcgamma receptors as regulators of immuneresponses. Nat Rev Immunol 2008;8(1):34–47.
64] Jegerlehner A, Schmitz N, Storni T, Bachmann MF. Influenza A vaccine basedon the extracellular domain of M2: weak protection mediated via antibody-dependent NK cell activity. J Immunol 2004;172(9):5598–605.
65] Mozdzanowska K, Furchner M, Washko G, Mozdzanowski J, Gerhard W. A pul-monary influenza virus infection in SCID mice can be cured by treatment withhemagglutinin-specific antibodies that display very low virus-neutralizingactivity in vitro. J Virol 1997;71(6):4347–55.
66] Lundgren M, Persson U, Larsson P, Magnusson C, Smith CI, Hammarstrom L,et al. Interleukin 4 induces synthesis of IgE and IgG4 in human B cells. Eur JImmunol 1989;19(7):1311–5.
67] Parham P. On the fragmentation of monoclonal IgG1, IgG2a, and IgG2b fromBALB/c mice. J Immunol 1983;131(6):2895–902.
68] Cox RJ, Madhun AS, Hauge S, Sjursen H, Major D, Kuhne M, et al. A phaseI clinical trial of a PER.C6 cell grown influenza H7 virus vaccine. Vaccine
69] Nicholson KG, Thompson CI, Klap JM, Wood JM, Batham S, Newman RW, et al.Safety and immunogenicity of whole-virus, alum-adjuvanted whole-virus,virosomal, and whole-virus intradermal influenza A/H9N2 vaccine formula-tions. Vaccine 2009;28(1):171–8.
Related Documents



![Liposome [GoR]](https://static.cupdf.com/doc/110x72/54f49f044a795997318b4927/liposome-gor.jpg)







