Cathodoluminescence tools provide clues to depositional history in Miocene and Pliocene mammalian teeth Loïc Ségalen a,c, ⁎, Marc de Rafélis a , Julia A. Lee-Thorp b,c , Anne-France Maurer a , Maurice Renard a a Université Pierre et Marie Curie-Paris06, JE 2477 Biominéralisations et Paléoenvironnements, Paris, F-75005 France b Department of Archaeological Sciences, University of Bradford, Bradford BD7 1DP, United Kingdom c Department of Archaeology, University of Cape Town, Private Bag, Rondebosch 7701, South Africa ABSTRACT ARTICLE INFO Article history: Accepted 26 March 2008 Keywords: Cathodoluminescence Fossil teeth REE Namibia Kenya Miocene Pliocene This paper presents the results of a study which used cathodoluminescence (CL) observations (optical images and SEM spectra) as a tool for assessing diagenetic processes during fossilisation of continental mammalian teeth. The study was performed on fossil teeth of large mammals (Gomphotheriidae, Rhinocerotidae and Hippopotamidae) from Namibia and from Kenya, with ages from the Lower and Middle Miocene to Pliocene. The fossil sites are distinguished by their environmental conditions. They include floodplain deposits, fluvial deposits of the Proto-Orange River and aeolianite deposits in Namibia, and fluvio-lacustrine deposits of different ages in Kenya. Optical CL provides information about the sites' environments and diagenetic processes related to those environments: a blue intrinsic luminescence for the aeolianite deposits and purple to yellowish for floodplain or fluvial deposits. The CL-spectra of these samples show sharp emission bands, which are classically related to late (recent) incorporation of rare- earth elements (REE) substituting in the Ca 2+ sites. High-resolution mapping of the distribution of these REE shows a gradient from the enamel interior (low concentrations) to the outer enamel margin (high concentrations). © 2008 Elsevier B.V. All rights reserved. 1. Introduction Biochemical data, including trace elements (e.g. Sr/Ca, Ba/Ca) and stable light isotope ratios retrieved from fossil vertebrate teeth and bones are now frequently used to reconstruct past diets and envi- ronments, and the biology of extinct species (i.e. Sillen, 1992; Gilbert et al., 1994; Sillen and Lee-Thorp, 1994; Burton and Wright, 1995; Balter et al., 2001; Balter et al., 2002; Sponheimer and Lee-Thorp, 2006). In application of those methods one has to be careful about what exactly is being analysed. Fossilisation processes can cause recrystallisation into different biominerals (depending on conditions), and incorporation of foreign ions can occur via a number of processes including solid state exchange in addition to recrystallisation. For all these reasons it is helpful to find out about the pathway(s) of fossili- sation in each particular case, and for each site. Hence it is important to understand the depositional, taphonomic and fossilisation history of the biogenic materials. In this study we used REE concentrations as a means of ascertaining the burial and depositional history and sub- sequent alteration of fossil teeth. The mineral component of teeth and bones is composed of bio- logical apatite, a member of the apatite series [Ca 5 (PO 4 ,CO 3 ) 3 (F,OH,Cl)]. Bone and dentine apatite are distinct from enamel: they are highly substituted (up to 6 wt.% CO 3 ) compared to enamel (about 3 wt.% CO 3 ). Other differences are found in their structures. Bone and dentine apatite show assemblages of minute crystallites (LeGeros, 1981, 1991; Elliott, 2002), whereas enamel apatite crystallites are larger and arranged in prisms (LeGeros, 1981, 1991; Elliott, 2002). Several studies have shown a rapid postmortem increase in bone crystallinity as a result of loss of the organic phase (Tuross et al., 1989; Person et al., 1995; Sillen and Parkington, 1996; Trueman et al., 2004). Rapid alteration and degradation of calcified tissues can also be related to combination of heat, moisture and bacterial actions. Diage- netic processes occur along several pathways depending on burial environments and conditions (Sponheimer and Lee-Thorp, 1999; Lee- Thorp and Sponheimer, 2003; Schoeninger et al., 2003; Farlow and Argast, 2006) that include precipitation and/or recrystallisation of phosphate compounds, authigenic carbonate, or other minerals, ionic exchanges and rearrangements of phosphate or carbonate in the primary structure (based on FTIR or XRD analysis). Incorporation of foreign ions in the crystal lattice can also take place in several ways. For instance, based on direct chemical measurement, several au- thors have noted formation of fluoroapatite following F - substitution for OH - (e.g. Bryant et al., 1994). Rare earth elements (REE) and trace elements (e.g. Na 2+ , Ba 2+ , Fe 2+ , Mg 2+ , Pb 2+ , Sr 2+ , Zn 2+ ) also substitute for Palaeogeography, Palaeoclimatology, Palaeoecology 266 (2008) 246–253 ⁎ Corresponding author. Laboratoire Biominéralisation et Paléoenvironnements, JE 2477, Université Pierre et Marie Curie, Case postale 116, 4 place Jussieu, 75 252 Paris cedex 05, France. E-mail address: [email protected] (L. Ségalen). 0031-0182/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.palaeo.2008.03.035 Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Palaeogeography, Palaeoclimatology, Palaeoecology 266 (2008) 246–253
Contents lists available at ScienceDirect
Palaeogeography, Palaeoclimatology, Palaeoecology
j ourna l homepage: www.e lsev ie r.com/ locate /pa laeo
Cathodoluminescence tools provide clues to depositional history in Miocene andPliocene mammalian teeth
Loïc Ségalen a,c,⁎, Marc de Rafélis a, Julia A. Lee-Thorp b,c, Anne-France Maurer a, Maurice Renard a
a Université Pierre et Marie Curie-Paris06, JE 2477 Biominéralisations et Paléoenvironnements, Paris, F-75005 Franceb Department of Archaeological Sciences, University of Bradford, Bradford BD7 1DP, United Kingdomc Department of Archaeology, University of Cape Town, Private Bag, Rondebosch 7701, South Africa
⁎ Corresponding author. Laboratoire Biominéralisatio2477, Université Pierre et Marie Curie, Case postale 116cedex 05, France.
E-mail address: [email protected] (L. Ségalen).
0031-0182/$ – see front matter © 2008 Elsevier B.V. Aldoi:10.1016/j.palaeo.2008.03.035
A B S T R A C T
A R T I C L E I N F OArticle history:
This paper presents the re Accepted 26 March 2008Keywords:CathodoluminescenceFossil teethREENamibiaKenyaMiocenePliocene
sults of a study which used cathodoluminescence (CL) observations (opticalimages and SEM spectra) as a tool for assessing diagenetic processes during fossilisation of continentalmammalian teeth. The study was performed on fossil teeth of large mammals (Gomphotheriidae,Rhinocerotidae and Hippopotamidae) from Namibia and from Kenya, with ages from the Lower and MiddleMiocene to Pliocene. The fossil sites are distinguished by their environmental conditions. They includefloodplain deposits, fluvial deposits of the Proto-Orange River and aeolianite deposits in Namibia, andfluvio-lacustrine deposits of different ages in Kenya. Optical CL provides information about the sites'environments and diagenetic processes related to those environments: a blue intrinsic luminescence forthe aeolianite deposits and purple to yellowish for floodplain or fluvial deposits. The CL-spectra of thesesamples show sharp emission bands, which are classically related to late (recent) incorporation of rare-earth elements (REE) substituting in the Ca2+ sites. High-resolution mapping of the distribution of theseREE shows a gradient from the enamel interior (low concentrations) to the outer enamel margin (highconcentrations).
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
Biochemical data, including trace elements (e.g. Sr/Ca, Ba/Ca) andstable light isotope ratios retrieved from fossil vertebrate teeth andbones are now frequently used to reconstruct past diets and envi-ronments, and the biology of extinct species (i.e. Sillen, 1992; Gilbertet al., 1994; Sillen and Lee-Thorp, 1994; Burton and Wright, 1995;Balter et al., 2001; Balter et al., 2002; Sponheimer and Lee-Thorp,2006). In application of those methods one has to be careful aboutwhat exactly is being analysed. Fossilisation processes can causerecrystallisation into different biominerals (depending on conditions),and incorporation of foreign ions can occur via a number of processesincluding solid state exchange in addition to recrystallisation. For allthese reasons it is helpful to find out about the pathway(s) of fossili-sation in each particular case, and for each site. Hence it is importantto understand the depositional, taphonomic and fossilisation historyof the biogenic materials. In this study we used REE concentrations asa means of ascertaining the burial and depositional history and sub-sequent alteration of fossil teeth.
n et Paléoenvironnements, JE, 4 place Jussieu, 75 252 Paris
l rights reserved.
The mineral component of teeth and bones is composed of bio-logical apatite, amember of the apatite series [Ca5(PO4,CO3)3(F,OH,Cl)].Bone and dentine apatite are distinct from enamel: they are highlysubstituted (up to 6 wt.% CO3) compared to enamel (about 3 wt.% CO3).Other differences are found in their structures. Bone and dentineapatite show assemblages of minute crystallites (LeGeros, 1981, 1991;Elliott, 2002), whereas enamel apatite crystallites are larger andarranged in prisms (LeGeros, 1981, 1991; Elliott, 2002).
Several studies have shown a rapid postmortem increase in bonecrystallinity as a result of loss of the organic phase (Tuross et al., 1989;Person et al., 1995; Sillen and Parkington, 1996; Trueman et al., 2004).Rapid alteration and degradation of calcified tissues can also berelated to combination of heat, moisture and bacterial actions. Diage-netic processes occur along several pathways depending on burialenvironments and conditions (Sponheimer and Lee-Thorp, 1999; Lee-Thorp and Sponheimer, 2003; Schoeninger et al., 2003; Farlow andArgast, 2006) that include precipitation and/or recrystallisation ofphosphate compounds, authigenic carbonate, or other minerals, ionicexchanges and rearrangements of phosphate or carbonate in theprimary structure (based on FTIR or XRD analysis). Incorporationof foreign ions in the crystal lattice can also take place in several ways.For instance, based on direct chemical measurement, several au-thors have noted formation of fluoroapatite following F− substitutionfor OH− (e.g. Bryant et al., 1994). Rare earth elements (REE) and traceelements (e.g. Na2+, Ba2+, Fe2+, Mg2+, Pb2+, Sr2+, Zn2+) also substitute for
247L. Ségalen et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 266 (2008) 246–253
Ca2+ in Ca sites (I and/or II) at higher levels compared with modernteeth and bone (Sillen, 1986; Williams and Henderson, 1997;Sponheimer and Lee-Thorp, 2006). In particular, Kohn et al. (1999)have shown that enhanced concentrations of Fe, Mn, Si, Al, Ba andMn–Fe oxyhydroxides are most closely related to chemical alterationof the apatite mineral and/or the precipitation of secondary minerals.All these changes can evolve rapidly, depend on the burial and depo-sitional environment and can modify the original biomineral statesignificantly and hence, possibly, the isotopic and chemical composi-tion of fossil remains (Kohn et al., 1999; Trueman and Tuross, 2002;Schoeninger et al., 2003; Sponheimer and Lee-Thorp, 2006).
It is generally recognised that bone structure is more sensitive thanenamel to the diagenesis (e.g., Ayliffe et al., 1994; Wang and Cerling,1994; Lee-Thorp and Sponheimer, 2003; Sponheimer and Lee-Thorp,2006) but some studies have pointed out that enamel is not neces-sarily immune, particularly for trace elements and REE incorporation(e.g., Kohn et al., 1999). Williams and Henderson (1997) showed thatfossil bone apatite concentrates the REE during fossilisation, and theirincorporation is related to interaction with groundwater and whetherequilibrium between the bone and the groundwater was attained(Trueman and Tuross, 2002). Very little is known about REE pene-tration of enamel (and dentine), hence patterns of REE take-up mightprovide clues to processes of uptake in this denser, less adsorbtivematerial and possibly throw light on patterns of recrystallisation andmineral change during fossilisation. We would expect to see thatpatterns follow somewhat different pathways under different deposi-tional conditions, and understanding these patterns might in turnprovide clues to burial environments and history. This informationcould prove useful in cases where such information is, for one reasonor another, not available.
To characterise the distribution of these contaminants in the latticestructure of biominerals, cathodoluminescence (bothoptical images andSEM spectra) may be useful (e.g. Habermann et al., 2000; Barbin et al.,2001; Balter et al., 2002; Schoeninger et al., 2003). Therefore, the goal ofthis study was to develop a simple method using a combination of CL-photographs (to map the relative REE/trace element distributions) andthe CL-spectra (to quantify REE analyses in some fossil samples), andhence, to examine the late incorporation and distribution of theseelements. We tested this approach to assess the state of preservation ofsamples collected from different African regions (Southern and EastAfrica), which are characterised by different depositional contexts (aridto humid, and hydrothermal environments).
Table 1List of Namibian samples analysed and Kenyan specimens studied in this paper
Samplesnumber
Namibian specimen (Southern Namib)
Specimen Age Sites Environment
EF 6 Gomphotheriidae 18–20 (Ma) Elisabethfeld Floodplain depositsAR Rhinocerotidae 17.5 (Ma) Arrisdrift Fluviatil
(Proto-Orange)AG Gomphotheriidae 17.5 (Ma) Arrisdrift Fluviatil
(Proto-Orange)RO Gomphotheriidae 10–12 (Ma) Rooilepel Aeolianites
Samplesnumber
Kenyan specimen (Tugen Hills)
Specimen Age Sites Environment
LS A1 Hippopotamidae Mabaget Fm(4 Ma)
Sagatia Fluvio-lacustine
LS 97 Hippopotamidae Mabaget Fm(4 Ma)
Kingetabkwony Fluvio-lacustine
LS 22 Gomphotheriidae Lukeino Fm(6 Ma)
Kapcheberek Fluvio-lacustine
LS 89 Hippopotamidae Lukeino Fm(6 Ma)
Kapcheberek Fluvio-lacustine
2. Materials and methods
2.1. Namibian samples
The first group comes from the Namibian faunal sites collected bythe Namibian Palaeontology Expedition (Pickford et al., 1995, 1996;Pickford and Senut, 2000, 2003). Teeth were collected from threelocalities in the southern Namib and differ in age and environmentaldeposits (sub-humid to arid environments). The first site, Elisabeth-feld, is close to Luderitz (Table 1, Fig.1) and consists offluvialfloodplaindeposits (Faunal Set late P0/basal PI, ~20–19 Ma; Pickford and Senut,2000). The second site, Arrisdrift, consists of fluviatile deposits accu-mulated in a palaeochannel of the Proto-Orange River (Table 1, Fig. 1)and this site is correlated to Faunal association Set PIIIa (~17–17.5 Ma;Hendey, 1978; Pickford and Senut, 2003). The last site is Rooilepel,close to the Orange River, and consists of aeolianite deposits (palaeo-barkane remains: Ségalen et al., 2004) and correlated to Faunal asso-ciation Set PV/PVI, (~10–12 Ma).
2.2. Kenyan samples
The second group of samples comes from the Tugen Hills zone inKenyawhere remains of the homininOrrorin tugenensiswere discoveredby the Kenya Palaeontology Expedition (Senut et al., 2001). The Kap-cheberek site is related to the Lukeino Fm (~6 Ma) and the King-etabkwony and Sagatia site to the Mabaget Fm (~4 Ma). These sites areinterpreted as fluvio-lacustrine sedimentary deposits (Pickford andSenut, 2001) (Table 1, Fig. 1).
2.3. Sample preparation and mineralogy
CL observations of fossil teeth were performed on thin sectionstaken along the vertical axial plane of each tooth fragment (eitherexclusively enamel, or for some samples, through both the enamel andthe dentine). The samples were selected for thick enamel (~1 mm to~8 mm) to provide a reasonable amount of working material. Thinsections (30 µm) were polished and carbon-coated before CL-spectro-scopy analysis.
Bulk enamel and bulk dentine samples were analysed by X-RayDiffraction where possible. All samples are bioapatite and no othermineral species were observed. The state of crystallinity was alsochecked following the protocol proposed by Person et al. (1995, 1996).The enamel and dentine samples show an average Crystallinity Index(CI) of about 1 and 0.5 respectively, regardless of the site's origin. Theincrease in crystallinity suggests that the samples experienced approx-imately the samedegree of postmortemcrystallisation, independentofage and burial environments.
2.4. CL-microscopy and spectroscopy of biological apatite
Cathodoluminescence refers to photon emission in the visible rangeresulting from excitation of a sample by high-energy electrons. Mn2+ isthe main activator for luminescence phenomena, and REE also con-tributewhen they are located in the Ca sites of apatite (Gaft et al., 1996).This method has been mostly used to identify diagenetic phases andalteration zones in sediments (Machel and Burton, 1991; Machel et al.,1991; de Rafélis et al., 2000), and less commonly in biological mineralssuch as shell (Barbin, 2000), conodonts (Habermann et al., 2000),bone (Williams and Henderson, 1997; Balter et al., 2002) or enamel(Götze et al., 2001; Schoeninger et al., 2003).Without any activators (i.e.trace elements and REE) a dull luminescence (dark bluish to light bluecolour around 400 nm) is observed that results from natural defects inthe crystal lattice and is interpreted as primary, or intrinsic, lumines-cence (Götze et al., 2001; Schoeninger et al., 2003).When substitution ofMn2+ occurs in the Ca sites, a yellow to bright orange luminescence isobserved with a broad emission at 565 nm (site Ca I) and 585 nm (site
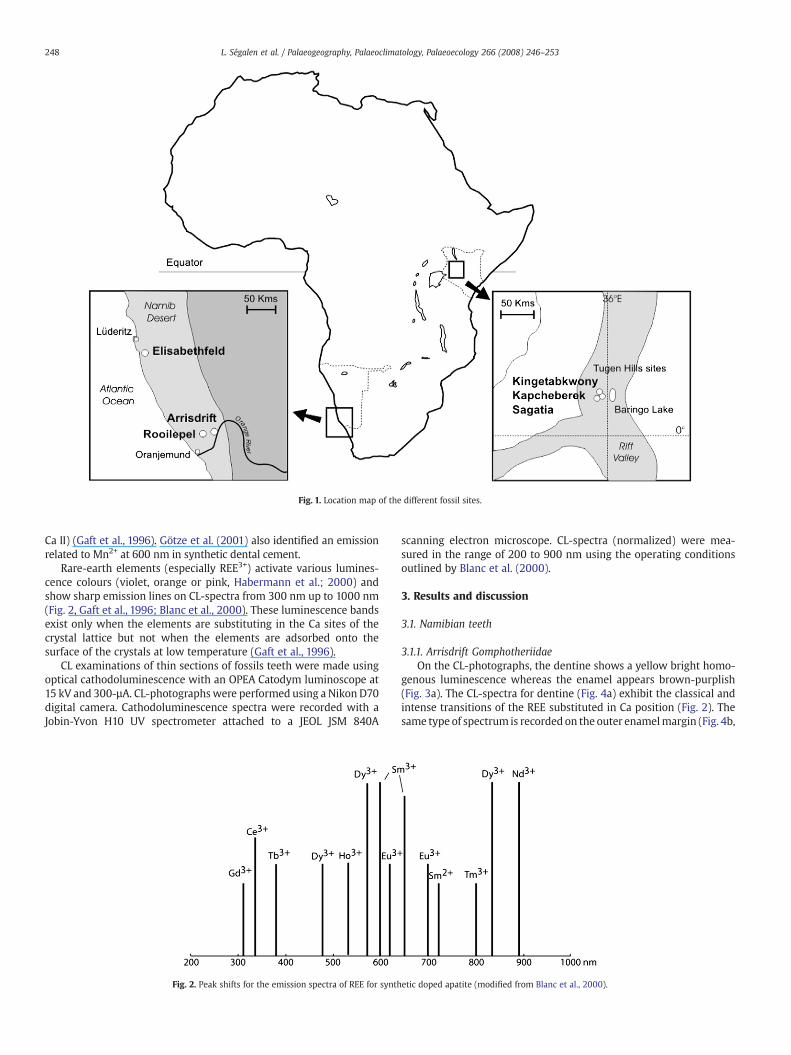
Fig. 1. Location map of the different fossil sites.
248 L. Ségalen et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 266 (2008) 246–253
Ca II) (Gaft et al., 1996). Götze et al. (2001) also identified an emissionrelated to Mn2+ at 600 nm in synthetic dental cement.
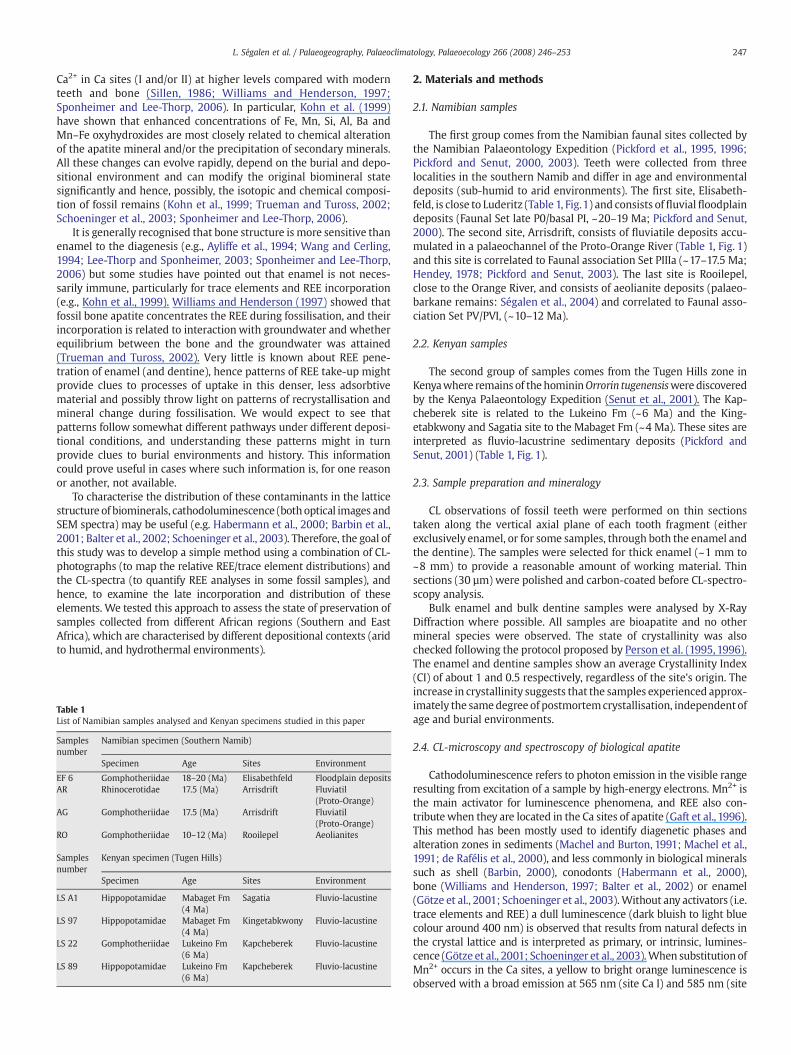
Rare-earth elements (especially REE3+) activate various lumines-cence colours (violet, orange or pink, Habermann et al.; 2000) andshow sharp emission lines on CL-spectra from 300 nm up to 1000 nm(Fig. 2, Gaft et al., 1996; Blanc et al., 2000). These luminescence bandsexist only when the elements are substituting in the Ca sites of thecrystal lattice but not when the elements are adsorbed onto thesurface of the crystals at low temperature (Gaft et al., 1996).
CL examinations of thin sections of fossils teeth were made usingoptical cathodoluminescence with an OPEA Catodym luminoscope at15 kV and 300-µA. CL-photographswere performed using a Nikon D70digital camera. Cathodoluminescence spectra were recorded with aJobin-Yvon H10 UV spectrometer attached to a JEOL JSM 840A
Fig. 2. Peak shifts for the emission spectra of REE for synth
scanning electron microscope. CL-spectra (normalized) were mea-sured in the range of 200 to 900 nm using the operating conditionsoutlined by Blanc et al. (2000).
3. Results and discussion
3.1. Namibian teeth
3.1.1. Arrisdrift GomphotheriidaeOn the CL-photographs, the dentine shows a yellow bright homo-
genous luminescence whereas the enamel appears brown-purplish(Fig. 3a). The CL-spectra for dentine (Fig. 4a) exhibit the classical andintense transitions of the REE substituted in Ca position (Fig. 2). Thesame type of spectrum is recorded on the outer enamelmargin (Fig. 4b,
etic doped apatite (modified from Blanc et al., 2000).

Fig. 3. Binocular (natural light) and CL-photographs of teeth from different fossil localities (a to d: Namibia; e to h: Kenya): a=Gomphoteriidae from Arrisdrift; b=Rhinocerotidae —
Arrisdrift; c=Gomphoteriidae — Elisabethfeld; d=Gomphoteriidae — Rooilepel; e=Hippopotamidae — Sagatia; f=Hippopotamidae — Kingetabkwony; g=Hippopotamidae —
Kapcheberek; h=Gomphoteriidae — Kapcheberek. D = dentine and E = enamel. Scale bar=0.5 cm. Crosses and associated numbers indicate the location of CL-spectra analysis.
249L. Ségalen et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 266 (2008) 246–253

Fig. 4. CL-spectra of Namibian tooth samples. Wavelength (nm) is plotted on the X-axis and relative intensity (counts) on the Y-axis. For precise location of each measurement see Fig. 3.
250 L. Ségalen et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 266 (2008) 246–253
AG 1) but those on the central region of the enamel (Fig. 4b, AG 2 to 4)show different emissions: intrinsic at 400 nm and at 600 nm activatedby the Mn2+. The AG 3 CL-spectra show a higher emission at 600 nmthan the AG 2 and AG 4 CL-spectrum (Fig. 4b). These variations arelikely related to differential distribution of the Mn2+ concentrationswithin the enamel (CL-intensity being proportional to the content inactivators Amieux, 1982; de Rafélis et al., 2000).
3.1.2. Arrisdrift RhinocerotidaeAs a general observation, the CL colours are identical to those
observed on Gomphotheriidae coming from the same site (dull yellowfor the dentine and brown to purple for the enamel, Fig. 3b). Thesimilarity is also found on the CL-spectra of enamel with low inten-sities for the internal enamel zone (Fig. 4c, AR 4, 5 and 6) corres-
ponding to the intrinsic luminescence (400 nm) and to Mn2+. REEactivate luminescence in the outer enamel margin (Fig. 4d, AR 7). Thespectra for dentine (Fig. 4e, AR 1 to 3) show different emissions:intrinsic luminescence (400 nm) andMn2+ (600 nm) for the inner partand poorly resolved REE activation (Fig. 4e, AR 1) on the outer dentinemargin. Due to the break, we assume the dentinewas not protected byenamel and was therefore in direct contact with the sediment.
3.1.3. Elisabethfeld GomphotheriidaeThe Gomphotheriidae teeth (enamel only) show similar CL colours
to the Arrisdrift specimens (Fig. 3c). In the same way, the CL-spectrashow a Mn2+-activated luminescence in the enamel (Fig. 4f, EF6-2 toEF6-4) expected for the outer margin (Fig. 4f, EF6-1) where theluminescence is clearly related to REE activators. We also analysed the

Fig. 5. CL-spectra of Kenyan tooth samples. Wavelength (nm) is plotted on the X-axis and relative intensity (counts) on the Y-axis. For precise location of each measurement see Fig. 3.
251L. Ségalen et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 266 (2008) 246–253
cement of a fracture which shows a bright orange luminescenceassociated with a 620 nm emission (Fig. 4g, EF6-5). This emission ischaracteristic of the Mn2+activator in calcite, suggesting that thecement is calcitic. SEM-EDS analysis confirmed the occurrence of thissecondary mineral.
3.1.4. Rooilepel GomphotheriidaeThe CL-photograph shows a homogeneous dark-blue luminescence
in all the enamel except on the external surface where a yellow fringedevelops (Fig. 3d). CL-spectra (RO 1 to 3, Fig. 4h) show a very lowluminescence identified as an intrinsic emission (400 nm) and Mn2+
(600 nm). Spectra of the outer enamelmargin (Fig. 4i, RO 4 and 5) show
the progressive incorporation of the REE from the outer fringe (Figs. 3dand 4i, RO 5) to the inner enamel part (Figs. 3d and 4i, RO 4).
3.2. Kenyan teeth
3.2.1. Sagatia HippopotamidaeThe luminescence of the tooth of Sagatia has low intensity. The
colours for both enamel and dentine are beige to brown (Fig. 3e).Compared to the dentine, the enamel presents many non-luminescentzones. CL-spectra are similar in the two tissues and show very high CLemissions characterising an activation of Mn2+ (Fig. 5a and b, LS A1-1to LS A1-5). No REE have been detected with the CL-SEM (Fig. 5a).

Table 2Example of REE concentration (ICP-MS analysis) in two Gomphotheriidae enamelsamples
REE (ppm) Ce Dy Gd Nd Sm
Arrisdrift 7 245 120 545 99.9Rooilepel 0.26 1.03 1.26 7.73 1.13
252 L. Ségalen et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 266 (2008) 246–253
3.2.2. Kingetabkwony HippopotamidaeThe CL-photograph of thedentine of Hippopotamidae tooth from the
Kingetabkwony site appears homogenous with bright yellow to orangecolours (Fig. 3f). In contrast, the enamel shows irregular luminescencewith grey (even non-luminescent) to yellow-green colour (Fig. 3f).
CL-spectra of enamel (Fig. 5c, LS97-1 to 3) and of dentine (Fig. 5d,LS97-4 and 5) are quite similar with characteristic intrinsic emission(400 nm) andMn2+ activated luminescence, with a dentine CL-intensityat 590 nm more intense compared to the enamel. In the enamel, theseintensities are not systematically related to the CL-photograph. Forexample, the bright colour (yellow) does not always correspond to thehighest luminescence intensities (Fig. 5c, LS 97-1 to LS 97-3).
3.2.3. Kapcheberek HippopotamidaeCL-photographs of the enamel and the dentine are easily distin-
guished by dark brown and yellow bright colours, respectively (Fig. 3g).The fractures are identifiable because of their non-luminescence(Fig. 3g). The outer enamel margin reveals (see detail on Fig. 3g) abright pink zonation, which corresponds to REE-activations (Figs. 3gand 5e, LS89-5) and differs from the rest of the enamel where theluminescence is induced by theMn2+ activator (Figs. 3g and 5e, LS89-4).CL-spectra of the dentine show that the luminescence phenomenon isalso related to the Mn2+ activator (Fig. 5f, LS89-2 and LS89-3) exceptfor the external fringe where luminescence is REE-activated (Fig. 5f,LS89-1).
3.2.4. Kapcheberek GomphotheriidaeThe tooth presents a thin zonewhere the dentine is preservedwith
a yellow colour is observed (Fig. 3h). The luminescence is related toMn2+ activator and low occurrence of REE (Fig. 5g, LS22-6). CL image ofenamel shows a colour zonation (beige–orange luminescence in theinner enamel zone to blue/dark brown towards the outer enamelmargin; Fig. 3h). In CL-spectra LS22-1 and LS22-2 (Fig. 5h), a blue/darkbrown colour is observed, suggesting a very dull Mn2+-luminescence,while CL-spectra LS22-3 (Fig. 5h) (beige/orange colour) display theusual intrinsic band (400 nm) associated with a similar emission at620 nm probably due to Mn2+ in small amount of calcite (less than 1%of the total sample). We also focused the analysis on two particularareas inside the enamel (Fig. 5i, LS22-4 and 5). CL-spectra indicate thatthese relative bright luminescences are due to Mn2+ in the enamellattice.
4. Discussion
Generally, the dentine of all the samples is characterised by ahomogeneous luminescence (yellow to brown colours) where onlythe emission intensity changes from one site or specimen to anotherone. From the CL-spectra, these colours can be related to Mn2+ incor-poration in the bioapatite lattice or to a diagenetic incorporation ofREE in large quantities. The results obtained with the enamel seemmore complex as suggested by the large spectrum of colours ob-served: blue, purple, yellow, pink, grey, brown. Nevertheless, theseluminescences are (as the dentine) always associated with the occur-rence of the Mn2+ or REE in the crystal lattice.
For the Namibian teeth, some coherent patterns appear, mainlyrelated to the type of environment. For Rhinocerotidae and Gompho-theriidae teeth, purple luminescence corresponds to Mn-rich teethdeposited in water-rich sediments (floodplain or fluvial), whereas the
blue luminescence (mainly intrinsic) corresponds to arid environ-ments (aeolianites). Such patterning is less clear for Kenyan teeth,probably because all samples come from similar taphonomic contexts(fluvio-lacustrine deposits) although they reveal many different CLcolours. These differences could be linked to the diversity of hydro-thermal and volcanic activity in the Rift valley, which producesgroundwaters with variable chemical compositions (Darling et al.,1996; Renaut et al., 2002).
In detail, evidence for REE incorporation in enamel always appearson the outer fringes. To confirm this incorporation, we used ICP-MSanalysis to check the REE concentrations in two different enamelsamples chosen because of the difference on the CL-spectra: AR-7(Fig. 4d, REE-activated luminescence) and RO 1 to 3 (Fig. 4h, intrinsicand Mn2+ luminescence). The results show that REE concentrations ofsample AR are two orders of magnitude greater than sample RO(Table 2). Williams and Henderson (1997) indicated that maximumconcentration of REE was systematically found in the sediment-bonesurface and that these elements in fossil bones are incorporatedthrough interaction with groundwater (and not enclosing sediment)during diagenesis. Our results obtained for the Arrisdrift (fluviatiledeposits) and Rooilepel (arid environment) sites are consistent withthis suggestion.
This study demonstrates that cathodoluminescence provides a use-ful window for evaluating diagenesis of fossil biological apatites, espe-cially teeth. CL-spectroscopy shows late incorporation of REE. Inparticular, the scale of resolution of this technique makes it possible todefine and to map precisely the thickness of the contaminated zoneduring burial modifications since the REE are present only on the ex-ternal margins. This method also permits distinction between differentkinds of burial environment. The broader importance is that this couldbe useful in establishing depositional histories of skeletal elements/teeth either where this is not known (for instance, where they occur insecondarycontexts) or because the environmenthas changeddrasticallybetween the death and the present time.
Acknowledgements
The authors gratefully acknowledge funding support from theGeological Survey of Namibia, the Namdeb Diamond Corporation (Pty)Ltd, De Beers Africa Exploration, the National Monuments Council ofthe Republic of Namibia, the French Embassy in Namibia, the MuséumNational d'Histoire Naturelle of Paris, the University Pierre & MarieCurie of Paris, the CNRS (ECLIPSE), the Community Museum of Kenya,the National Research Foundation of South Africa, the Palaeoanthro-pology Scientific Trust, and the University of Cape Town. We thank M.Lemoine for producing the thin sections and P. Blanc for his help andcomments on this paper. We thank V. Barbin and an anonymousreviewer for their comments. LS would like to thank Judith Sealy andJulia Lee-Thorp for their support during his postdoctoral fellowship atUCT.
References
Ayliffe, L.K., Chivas, A.R., Leakey, M.G., 1994. The retention of primary oxygen isotopecompositions of fossil elephant skeletal phosphate. Geochim. Cosmochim. Acta 58,5291–5298.
Amieux, P., 1982. La cathodoluminescence: méthode d'étude sédimentologique descarbonates. Bull. Centre Rech. Explor. Prod. Elf-Aquitaine 6, 437–483.
Balter, V., Person, A., Labourdette, N., Drucker, D., Renard, M., Vandermeersch, B., 2001.Were Neandertalians essentially carnivores? Sr and Ba preliminary results of themammalian palaeobiocoenoses of Saint-Césaire. C. R. Acad. Sci. Paris 332, 59–65.
Balter, V., Bocherens, H., Person, A., Labourdette, N., Renard, M., Vandermeersch, B., 2002.Ecological andphysiological variability of Sr/Ba and Ba/Ca inmammals ofWest Europeanmid-Würmian food webs. Palaeogeogr. Palaeoclimatol. Palaeoecol. 186, 127–143.
Barbin, V., 2000. Cathodoluminescence of carbonate shells: biochemical vs diageneticprocess. In: Page, M., Barbin, V., Blanc, P., Ohnenstetter, D. (Eds.), Cathodolumines-cence in Geosciences. Springer, Berlin, pp. 303–329.
Barbin, V., Ramseyer, K., Debenay, J.P., Schein, E., Roux, M., Decrouez, 2001. Catho-doluminescence of recent biogenic carbonates: an environmental and ontogenicfingerprint. Geol. Mag. 128, 19–26.

253L. Ségalen et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 266 (2008) 246–253
Blanc, Ph., Baumer, A., Cesbron, F., Ohnenstteter, D., Panczer, G., Rémond, G., 2000.Systematic cathodoluminescence spectral analysis of synthetic doped minerals:anhydrite, apatite, calcite, fluorite, scheelite and zircon. In: Pagel, M., Barbin, V.,Blanc, Ph., Ohnenstetter, D. (Eds.), Cathodoluminescence in Geosciences. Springer-Verlag, Berlin, pp. 127–160.
Bryant, J.D., Luz, B., Froelich, P.N.,1994. Oxygen isotopic composition of fossil horse toothphosphate as a record of continental paleoclimate. Palaeogeogr. Palaeoclimatol.Palaeoecol. 107, 303–316.
Burton, J.H., Wright, L.E., 1995. Nonlinearity in the relationship between bone Sr/Ca anddiet: palaeodietary implications. Am. J. Phys. Anthropol. 96, 273–282.
Darling, W.G., Gizaw, B., Arusei, M.K., 1996. Lake-groundwater relationships and fluid–rock interaction in the East African Rift Valley: isotopic evidence. J. Afr. Earth Sci. 22,423–431.
de Rafélis, M., Renard, M., Emmanuel, L., Durlet, C., 2000. Apports de la cathodolumi-nescence du Mn dans les carbonates pélagiques. C. R. Acad. Sci. Paris 330, 391–398.
Elliott, J.C., 2002. Calcium phosphate biominerals. Rev. Mineral. Geochem. 48, 427–453.Farlow, J.O., Argast, A., 2006. Preservation of fossil bone from the pipe creek sinkhole
(late Miocene, Grant County, Indiana, U.S.A.). J. Paleontol. Soc. Korea 22, 51–75.Gaft, M., Shoval, S., Panczer, G., Nathan, Y., Champagon, B., Garapon, C., 1996. Lumi-
nescence of uranium and rare-earth elements in apatite of fossil fish teeth.Palaeogeogr. Palaeoclimatol. Palaeoecol. 126, 187–193.
Gilbert, C., Sealy, J., Sillen, A., 1994. An investigation of barium, calcium and strontium aspalaeodietary indicators in the Southwestern Cape, South Africa. J. Archaeol. Sci. 21,173–184.
Götze, J., Heimann, R.B., Hildebrandt, H., Gburek, U., 2001. Microstructural investigationinto calcium phosphate biomaterials by spatially resolved cathodoluminescence.Mat.-wiss. U. Werkstofftech. 32, 130–136.
Habermann, D., Götte, T., Meijer, J., Stephan, A., Richter, D.K., Niklas, J.R., 2000. Highresolution rare-earth element analyses of natural apatite and its application in geo-sciences: combined micro-PIXE, quantitative CL spectrometry and electron spinresonance analyses. Nucl. Instrum. Methods Phys. Res., B 161–163, 846–851.
Hendey, Q.B., 1978. Preliminary report on the Miocene vertebrates from Arrisdrift,South West Africa. Ann. S. Afr. Mus. 76, 1–41.
Kohn, M.J., Schoeninger, M.J., Barker, W.W., 1999. Altered states: effects of diagenesis onfossil tooth chemistry. Geochim. Cosmochim. Acta 63, 2737–2747.
Lee-Thorp, J.A., Sponheimer, M., 2003. Three case studies used to reassess the reliabilityof fossil bone and enamel isotope signals for paleodietary studies. J. Anthropol.Archaeol. 22, 208–216.
LeGeros, R.Z., 1981. Apatites in biological systems. Prog. Cryst. Growth Charact. 4, 1–45.LeGeros, R.Z., 1991. Calcium Phosphates in Oral Biology andMedicine. Karger, New York,
pp. 1–201.Machel, H.G., Burton, E.A., 1991. Factors governing cathodoluminescence in calcite and
dolomite, and their implications for studies of carbonate diagenesis. In: Barker, C.E.,Kopp, O.C. (Eds.), Luminescence Microscopy: Quantitative and Qualitative Aspects.SEPM short course, vol. 25, pp. 37–57.
Machel, H.G., Mason, R.A., Mariano, A.N., Mucci, A., 1991. Causes and emission ofluminescence in calcite and dolomite. In: Barker, C.E., Kopp, O.C. (Eds.),Luminescence Microscopy: Quantitative and Qualitative Aspects. SEMP, pp. 9–25.
Person, A., Bocherens, H., Saliège, J.F., Paris, F., Zeitoun, V., Gerard, M., 1995. Earlydiagenetic evolution of bone phosphate: an X-ray diffractometry analysis.J. Archaeol. Sci. 22, 211–221.
Person, A., Bocherens, H., Mariotti, A., Renard, M., 1996. Diagenetic evolution andexperimental heating of bone phosphate. Palaeogeogr. Palaeoclimatol. Palaeoecol.126, 135–149.
Pickford, M., Senut, B., 2000. Geology and Palaeobiology of the Namib Desert, South-western Africa. . Memoir, vol. 18. Geological Survey of Namibia. 155 pp.
Pickford, M., Senut, B., 2001. The geological and faunal context of Late Miocene hominidremains from Lukeino, Kenya. C. R. Acad. Sci. II 332, 145–152.
Pickford, M., Senut, B., 2003. Geology and Palaeobiology of the Central and SouthernNamib: Vol. 2 Palaeontology of the Orange River Valley, Namibia. . Memoir, vol. 19.Geological Survey of Namibia. 398 pp.
Pickford, M., Senut, B., Mein, P., Morales, J., Soria, D., Nieto, M., Ward, J., Bamford, M.,1995. The discovery of lower and middle Miocene vertebrates at Auchas, southernNamiba. C. R. Acad. Sci. Paris, Série IIa 322, 901–906.
Pickford, M., Senut, B., Mein, P., Gommery, D., Morales, J., Soria, D., Nieto, M., Ward, J.,1996. Preliminary results of new excavations at Arrisdrift, middle Miocene ofsouthern Namibia. C. R. Acad. Sci. Paris, Série IIa 322, 991–996.
Renaut, R.W., Jones, B., Tiercelin, J.-J., Tarits, C., 2002. Sublacustrine precipitation ofhydrothermal silica in rift lakes: evidence from Lake Baringo, central Kenya RiftValley. Sediment. Geol. 148, 235–257.
Schoeninger, M.J., Hallin, K., Reeser, H., Valley, J.W., Fournelle, J., 2003. Isotopic alterationof mammalian tooth enamel. Int. J. Osteoarchaeol. 13, 11–19.
Ségalen, L., Rognon, P., Pickford, M., Senut, B., Emmanuel, L., Renard, M., Ward, J., 2004.Reconstitution des morphologies dunaires et du régime des paléovents dans leProto-Namib au cours du Miocène. Bull. Soc. Geol. Fr. 175, 537–546.
Senut, B., Pickford, M., Gommery, D., Mein, P., Cheboi, K., Coppens, Y., 2001. Firsthominid from the Miocene (Lukeino Formation, Kenya). C. R. Acad. Sci II 332,137–144.
Sillen, A., 1986. Biogenic and diagenetic Sr/Ca in Plio-Pleistocene fossils in the OmoShungura Formation. Paleobiology 12, 311–323.
Sillen, A., 1992. Strontium-calcium ratios (Sr/Ca) of Australopithecus robustus andassociated fauna from Swartkrans. J. Hum. Evol. 23, 495–516.
Sillen, A., Lee-Thorp, J.A., 1994. Trace elements and isotopic aspects of predator–preyrelationships in terrestrial foodwebs. Palaeogeogr. Palaeoclimatol. Palaeoecol. 107,243–255.
Sillen, A., Parkington, J.E., 1996. Diagenesis of bones from Eland's Bay Cave. J. Archaeol.Sci. 23, 535–542.
Sponheimer, M., Lee-Thorp, J.A., 1999. Alteration of enamel carbonate environmentsduring fossilization. J. Archaeol. Sci. 26, 143–150.
Sponheimer, M., Lee-Thorp, J.A., 2006. Enamel diagenesis at South African Australopithsites: implications for paleoecological reconstruction with trace elements.Geochim. Cosmochim. Acta 70, 1644–1654.
Trueman, C.N., Tuross, N., 2002. Trace elements in recent and fossil bone apatite. Rev.Mineral. Geochem. 48, 489–521.
Trueman, C.N., Behrensmeyer, A.K., Tuross, N., Weiner, S., 2004. Mineralogical andcompositional changes in bones exposed on soil surfaces in Amboseli National Park,Kenya: diagenetic mechanisms and the role of sediment pore fluids. J. Archaeol. Sci.31, 721–739.
Tuross, N., Behrensmeyer, A.K., Eanes, E.D., Fisher, L.W., Hare, P.E., 1989. Molecularpreservation and crystallographic alterations in a weathering sequence of wild-ebeest bones. Appl. Geochem. 4, 261–270.
Wang, Y., Cerling, T.E., 1994. A model of fossil tooth and bone diagenesis: implicationsfor paleodiet reconstruction from stable isotopes. Palaeogeogr. Palaeoclimatol.Palaeoecol. 107, 281–289.
Williams, C.T., Henderson, P., 1997. The environment of deposition indicated by thedistribution of rare earth elements in fossil bone from Olduvai Gorge, Tanzania.Appl. Geochem. 12, 537–547.
Related Documents











