REGULAR ARTICLE Catalase, Bax and p53 expression in the visual system of the crab Ucides cordatus following exposure to ultraviolet radiation Nadia Campos de Oliveira Miguel & Inês Júlia R. Wajsenzon & Christina Maeda Takiya & Leonardo Rodrigues de Andrade & Giovane G. Tortelote & Marcelo Einicker-Lamas & Silvana Allodi Received: 8 December 2006 / Accepted: 2 March 2007 / Published online: 4 April 2007 # Springer-Verlag 2007 Abstract In invertebrates, a few studies have suggested apoptosis as the mechanism of choice to protect the retina after exposure to ultraviolet (UV) radiation. We demon- strated previously, by electron microscopy, that the retina and lamina ganglionaris (or lamina) cells of the crab Ucides cordatus displayed subcellular signs of apoptosis after exposure to UVB and UVC. Here, we first ascertained, by the TdT-mediated dUTP-biotin nick end-labeling (TUNEL) technique, that UV irradiation indeed produced the previ- ously reported results. We next tested, in the visual system of U. cordatus, whether the expression (as analyzed by immunohistochemistry and observed with laser scanning microscopy) and levels (as examined by Western blotting) of catalase, Bax, and p53 were affected by the same dose of UV radiation as that used previously. Our data revealed that the intensity of catalase, Bax, and p53 labeling was stronger in irradiated retina and lamina cells than in non-irradiated retina and lamina. However, no significant difference was observed in the concentrations of these proteins isolated from the whole optic lobe. The results thus suggest that UVB and UVC induce apoptosis in the crustacean retina and lamina by increasing catalase expression and activating the Bax- and p53-mediated apoptosis pathways. Keywords Cell death . Visual system . Immunohistochemistry . Western blotting . Mangrove crab, Ucides cordatus (Crustacea) Introduction Human interference in the environment in recent decades has been so immense that studies conducted to understand the mechanisms underlying the resulting stress are essential. In this context, the study of enzymes (and other molecules) involved in responses to stress can lead to important insights. Previous investigations have demonstrated that the role of antioxidant enzymes can be interpreted in terms of the concentration of reactive oxygen species available when cells are submitted to damage (Garcia et al. 2000; Orbea et al. 2002). Cell Tissue Res (2007) 329:159–168 DOI 10.1007/s00441-007-0410-x Nadia Campos de Oliveira Miguel and Inês Júlia R. Wajsenzon contributed equally to this work. This study was supported by the Brazilian Research Council for Science and Technology (CNPq), Rio de Janeiro State Foundation for the Advancement of Science (FAPERJ), José Bonifácio Foundation (FUJB), and Federal University of Rio de Janeiro Research Chamber (SR2/UFRJ). N. C. Miguel : I. J. R. Wajsenzon : C. M. Takiya : L. R. de Andrade : S. Allodi (*) Departamento de Histologia e Embriologia, Instituto de Ciências Biomédicas, Universidade Federal do Rio de Janeiro, Ilha do Fundão, 21941-590 Rio de Janeiro, RJ, Brazil e-mail: [email protected] N. C. Miguel : I. J. R. Wajsenzon : C. M. Takiya : L. R. de Andrade : S. Allodi Programa de Pós-Graduação em Ciências Morfológicas, Instituto de Ciências Biomédicas, Universidade Federal do Rio de Janeiro, 21941-590 Rio de Janeiro, RJ, Brazil G. G. Tortelote : M. Einicker-Lamas Instituto de Biofísica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro, 21941-590 Rio de Janeiro, RJ, Brazil

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

REGULAR ARTICLE
Catalase, Bax and p53 expression in the visual systemof the crab Ucides cordatus following exposureto ultraviolet radiation
Nadia Campos de Oliveira Miguel &Inês Júlia R. Wajsenzon & Christina Maeda Takiya &
Leonardo Rodrigues de Andrade &
Giovane G. Tortelote & Marcelo Einicker-Lamas &
Silvana Allodi
Received: 8 December 2006 /Accepted: 2 March 2007 / Published online: 4 April 2007# Springer-Verlag 2007
Abstract In invertebrates, a few studies have suggestedapoptosis as the mechanism of choice to protect the retinaafter exposure to ultraviolet (UV) radiation. We demon-strated previously, by electron microscopy, that the retinaand lamina ganglionaris (or lamina) cells of the crab Ucidescordatus displayed subcellular signs of apoptosis afterexposure to UVB and UVC. Here, we first ascertained, bythe TdT-mediated dUTP-biotin nick end-labeling (TUNEL)technique, that UV irradiation indeed produced the previ-
ously reported results. We next tested, in the visual systemof U. cordatus, whether the expression (as analyzed byimmunohistochemistry and observed with laser scanningmicroscopy) and levels (as examined by Western blotting)of catalase, Bax, and p53 were affected by the same dose ofUV radiation as that used previously. Our data revealed thatthe intensity of catalase, Bax, and p53 labeling was strongerin irradiated retina and lamina cells than in non-irradiatedretina and lamina. However, no significant difference wasobserved in the concentrations of these proteins isolatedfrom the whole optic lobe. The results thus suggest thatUVB and UVC induce apoptosis in the crustacean retinaand lamina by increasing catalase expression and activatingthe Bax- and p53-mediated apoptosis pathways.
Keywords Cell death . Visual system .
Immunohistochemistry .Western blotting .Mangrove crab,Ucides cordatus (Crustacea)
Introduction
Human interference in the environment in recent decadeshas been so immense that studies conducted to understandthe mechanisms underlying the resulting stress are essential.In this context, the study of enzymes (and other molecules)involved in responses to stress can lead to importantinsights. Previous investigations have demonstrated thatthe role of antioxidant enzymes can be interpreted in termsof the concentration of reactive oxygen species availablewhen cells are submitted to damage (Garcia et al. 2000;Orbea et al. 2002).
Cell Tissue Res (2007) 329:159–168DOI 10.1007/s00441-007-0410-x
Nadia Campos de Oliveira Miguel and Inês Júlia R. Wajsenzoncontributed equally to this work.
This study was supported by the Brazilian Research Council forScience and Technology (CNPq), Rio de Janeiro State Foundation forthe Advancement of Science (FAPERJ), José Bonifácio Foundation(FUJB), and Federal University of Rio de Janeiro Research Chamber(SR2/UFRJ).
N. C. Miguel : I. J. R. Wajsenzon :C. M. Takiya :L. R. de Andrade : S. Allodi (*)Departamento de Histologia e Embriologia,Instituto de Ciências Biomédicas,Universidade Federal do Rio de Janeiro,Ilha do Fundão,21941-590 Rio de Janeiro, RJ, Brazile-mail: [email protected]
N. C. Miguel : I. J. R. Wajsenzon :C. M. Takiya :L. R. de Andrade : S. AllodiPrograma de Pós-Graduação em Ciências Morfológicas,Instituto de Ciências Biomédicas,Universidade Federal do Rio de Janeiro,21941-590 Rio de Janeiro, RJ, Brazil
G. G. Tortelote :M. Einicker-LamasInstituto de Biofísica Carlos Chagas Filho,Universidade Federal do Rio de Janeiro,21941-590 Rio de Janeiro, RJ, Brazil

Catalase serves to remove hydrogen peroxide from cells,splitting it into water and oxygen, thus avoiding thegeneration of hydroxyl radicals (Halliwell and Gutteridge1989). In invertebrates, catalase has been detected in theperoxisomes of the digestive gland (Cajaraville et al. 1992),the blood cells of the mussel Mytilus sp. (Pipe andLivingstone 1993), and in the eyestalk of the crab Ucidescordatus (Miguel et al. 2005). Results regarding catalaseactivity in apoptosis are inconsistent, but evidence for bothanti-apoptotic and pro-apoptotic roles of catalase has beenreported (Reddy et al. 2004). Apoptosis is a criticaldevelopmental and homeostatic process, and its mecha-nisms are involved in numerous human diseases, includingcancers and neurodegenerative disorders (Kerr et al. 1972;Wyllie 1985; for a review, see Guimarães and Linden2004).
Ultraviolet (UV) radiation and other DNA-damage-inducing agents have been shown to trigger apoptosis, firstactivating caspase-2 and then regulating the mitochondrialand post-mitochondrial events. Active caspase-2 engagesmitochondria by promoting B-cell leukemia/lymphoma-2(Bcl-2)-associated protein X (Bax) translocation to themitochondria. Consequently, Bax has been proposed toplay a central role in bridging the active caspase-2 withmitochondria, by affecting mitochondrial permeability,cytochrome c release into the cytosol, and caspase-9activation. In addition, UV-induced apoptosis and caspase-2 activation have been shown to be diminished in Bax-deficient cells, suggesting that Bax plays an important rolein UV-induced apoptosis (He et al. 2004).
DNA breakage can be induced by ionizing radiationthrough the activation of cell cycle checkpoints. Once DNAis damaged, the tumor suppressor p53 protein mediates acell response by regulating the expression of a number ofcell targets (Vousden and Lu 2002). A master regulator ofcellular responses to ionizing radiation is ATM (ataxia-telangiectasia-mutated) kinase, which phosphorylates manydownstream effectors, including p53, in order to inhibit cellcycle progression or to induce apoptosis (Canman et al.1998). Even though UV rays cannot be considered asionizing radiation, UV irradiation leads to the stabilizationof p53 protein, in part attributable to its phosphorylation byextracellular signal-regulated kinase and p38 kinases (Sheet al. 2000; Chouinard et al. 2002).
In a previous paper (Miguel et al. 2002), we havedemonstrated, by electron microscopy, that retina and thelamina ganglionaris cells of the crab U. cordatus exposed toultraviolet B (UVB) and C (UVC) radiation displaysubcellular signs of apoptosis, such as an increase inheterochromatin, karyolyses, and karyorrhexes. Therefore,the purpose of this study has been, first, to confirmapoptosis under the same conditions used previously withthe TdT-mediated dUTP-biotin nick end-labeling (TUNEL)
technique, and then to test, in the visual system of this samespecies, whether catalase, Bax, and p53 expression (byusing immunohistochemistry) and levels (through immuno-blotting) are affected by the same high dose of UVB andUVC radiation used previously. A remark should be addedhere regarding the focus of this study: even though theeffects caused by UVA and UVB have received moreattention than those produced by UVC (Sanford et al. 1996;Duncan et al. 1997; Gouveia et al. 2005), the latter shouldnot be neglected, because, among other reasons, the studyof UVC radiation may help us to understand some of theunderlying mechanisms of UV-induced cell damage.
Materials and methods
Animals
Eighty-one mangrove crabs, U. cordatus (Linnaeus 1763),grouped according to the technique employed, were used:one for routine light microscopy (histological sectionsstained with hematoxylin and eosin), three for TUNEL,12 for immunohistochemistry, and 65 for immunoblotting.Adult male specimens with carapace widths ranging from6.1 cm to 7.6 cm were obtained from Itambi, Niterói, Stateof Rio de Janeiro, Brazil (IBAMA document number02022.003021/06-69). The crabs were maintained undercontrolled laboratory conditions at a temperature of 25±3°C and 12-h/12-h light/dark cycle (natural light) and wereregularly fed with green leaves.
All the procedures adopted for the treatment and use ofthe animals were according to the “NIH Guide and Care ofLaboratory Animals” and were approved by the UseEvaluation Committee of Animals in Research of theCentro de Ciências da Saúde, URFJ.
Radiation factors
The animals were anesthetized by chilling. Prior to the 30-min exposure to UV light, one eye of each crab wascompletely covered with a bandage painted with non-toxicblack enamel. After exposure, the covered eye of each animalhad the bandage removed and was used as a control. Thelight emanated from an Osram 15 W mercury vapor lamp(95% of the emission at 254 nm and 5% above 254 nm).The received UVC dose measured was 7,200 J/cm2, and thereceived UV-B dose was 590 J/cm2. During exposure, thecrab eyes remained 12 cm from the UV-light source, at atemperature of 25±3°C. One week after exposure, theanimals that survived were anesthetized again by chillingand, always in the morning, had their eyestalks ablated andthe optic lobes dissected.
160 Cell Tissue Res (2007) 329:159–168

Routine histology
Optic lobes were fixed in Bouin’s solution, dehydrated inethanol, cleared in xylene, and then infiltrated with andembedded in paraffin. They were sectioned at 6 μmlongitudinal to the long axis of the stalk and stained withhematoxylin and eosin.
TUNEL method
Optic lobes were fixed in Bouin’s solution, dehydrated in agraded ethanol series, cleared in xylene, and embedded inparaffin. They were sectioned at a thickness of 6 μmlongitudinal to the long axis of the stalk. The sections weremounted on gelatin-coated slides.
The ApopTag Peroxidase In situ Apoptosis Detection kit(Chemicon International, Temecula, Calif., USA) was usedfor DNA fragmentation analysis according to the manufac-turer’s instructions, with a few modified procedures.Briefly, the tissue sections were dewaxed, rehydrated with0.1 M phosphate-buffered saline (PBS), and boiled in0.1 M sodium citrate buffer (pH 6.0), as used for antigenretrieval in immunohistochemical procedures (30 min).After being cooled, endogenous peroxidase activity in thesections was quenched by incubation with 3% hydrogenperoxide in PBS, and then the equilibration buffer wasapplied to the tissue samples, followed by the application ofthe working-strength TdT enzyme. The working-strengthstop/wash buffer was applied to the slides, followed by theanti-digoxigenin conjugate. The sections were washed inPBS, and the characteristic reaction color was developed inperoxidase substrate. Finally, the sections were counter-stained with hematoxylin and examined under a lightmicroscope (Zeiss Axioskop). Nuclei with brown stainingwere indicative of DNA fragmentation, suggesting apopto-sis. We counted the nuclei positive for TUNEL, and theresults matched those obtained in our previous paper(Miguel et al. 2002) by toluidine-blue staining. Eyestalksections treated with a TdT-substrate-free solution wereused as a negative control, and sections of rat testes wereused as a positive control.
Immunohistochemistry
Optic lobes were fixed and prepared for sectioning asdescribed above. Sections mounted on gelatin-coated slideswere treated with 10% bovine serum albumin (BSA) toblock nonspecific binding of immunoglobulins to tissuesand then incubated overnight at 4°C in a humid chamberwith one of the primary antibodies: rabbit polyclonal anti-catalase (Chemicon International, Temecula, Calif., USA);rabbit polyclonal anti-Bax (Santa Cruz Biotechnology,Santa Cruz, Calif., USA); mouse monoclonal anti-p53
(clone DO-1; Santa Cruz Biotechnology); mouse monoclo-nal anti-p53 (clone DO-7; DakoCytomation, Carpinteria,Calif., USA). The fluorescent secondary antibodies (anti-rabbit IgG CY-3 conjugate and anti-mouse IgG CY-3conjugate; Sigma, St. Louis, Mo., USA) were then appliedto the sections, which were observed, as a first step, under aconventional fluorescence microscope (Zeiss Axioskop 2standard fluorescence microscope). The sections were alsoobserved under a laser scanning microscope (LSM) Zeiss510 META, at 543 nm wavelength of excitation for theCY-3 fluorochrome. All images were acquired at 512×512pixels of resolution, 75% laser transmittance, optical slices0.7 μm deep, and 1 min scanning-time in the same value ofdetector gain. Negative controls for the reaction wereprepared by omitting the primary antibody, and positivecontrols (data not shown) were prepared by using rat liversections (catalase) and human basal cell carcinoma (Baxand p53).
Materials for SDS-polyacrylamide gel electrophoresisand immunoblotting
All the reagents used were of the highest purity grade. Forprotein separation and Western blotting, Hybond nitrocel-lulose membrane (Amersham Pharmacia Biotech, Ger-many), the chemiluminescence ECL system (Amersham,Buckinghamshire, UK), and a Hyperfilm (AmershamPharmacia Biotech, UK) diagnostic film were used.Solvents and solutions used to develop the radiogramswere purchased from Kodak (Rezende, Brazil). Otherreagents used for protein determination and other biochem-ical procedures were from Sigma-Aldrich (St. Louis, Mo.,USA).
Protein determination assay
The dissected and isolated whole optic lobes (both irradiatedand controls) were kept in a glass tube containing a smallvolume of PBS (0.5 ml) and, in order to maintain proteinintegrity, were stored at the most for 3 days at −20°C. Theseisolated optic lobes were homogenized in a Potter-Elvehjemhomogenizer, and aliquots were then used to determinethe protein concentration according to the Folin-phenolmethod described by Lowry et al. (1951), with BSA as thestandard.
Total proteins from the optic lobes were separated andidentified by 12.5% sodium dodecylsulfate-polyacrylamidegel electrophoresis (SDS-PAGE) on a Mini PROTEAN3System (Bio-Rad Laboratories, Hercules, Calif., USA) at60 mA/gel. The proteins were transferred at 350 mA to anitrocellulose membrane by using the same Bio-Rad systemdescribed above for approximately 90 min. The nitrocellu-lose membrane containing the immobilized proteins was
Cell Tissue Res (2007) 329:159–168 161
first blocked with non-fat dry milk (5%) plus BSA (1%) in0.1 M TRIS-buffered saline containing 0.001% Tween(TBSt), for 90 min. This procedure had previously beenused successfully in blotting proteins obtained from theeyestalks of crabs of the same species (Corrêa et al. 2004;da Silva et al. 2004). After blockage, the membrane waswashed twice in TBSt under constant stirring for 3 min.Next, the membrane was incubated with the same anti-bodies used for immunodetection: anti-catalase polyclonalantibody, anti-Bax polyclonal antibody, or anti-p53 mono-clonal antibody, under constant gentle stirring for 2 h atroom temperature. The membrane was washed again (5×,3 min each) with TBSt. The secondary antibodies used wereanti-mouse or anti-rabbit IgGs conjugated to peroxidase(diluted at 1:500) and were incubated with the membranefor 90 min at room temperature. Membranes were washedas described above. Catalase, Bax, and p53 were detectedby using the chemiluminescence ECL system and aHyperfilm diagnostic film. The molecular weights wereeasily determined by using pre-stained molecular weightstandards from Sigma-Aldrich (St. Louis, Mo., USA).
Results
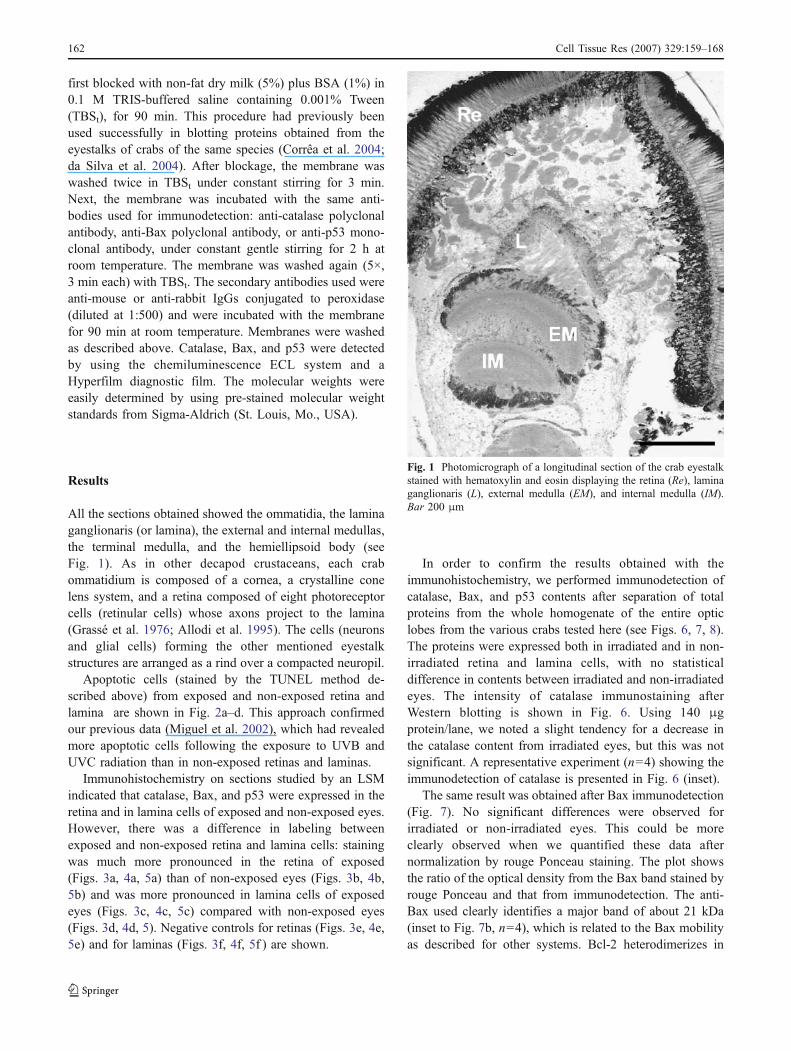
All the sections obtained showed the ommatidia, the laminaganglionaris (or lamina), the external and internal medullas,the terminal medulla, and the hemiellipsoid body (seeFig. 1). As in other decapod crustaceans, each crabommatidium is composed of a cornea, a crystalline conelens system, and a retina composed of eight photoreceptorcells (retinular cells) whose axons project to the lamina(Grassé et al. 1976; Allodi et al. 1995). The cells (neuronsand glial cells) forming the other mentioned eyestalkstructures are arranged as a rind over a compacted neuropil.
Apoptotic cells (stained by the TUNEL method de-scribed above) from exposed and non-exposed retina andlamina are shown in Fig. 2a–d. This approach confirmedour previous data (Miguel et al. 2002), which had revealedmore apoptotic cells following the exposure to UVB andUVC radiation than in non-exposed retinas and laminas.
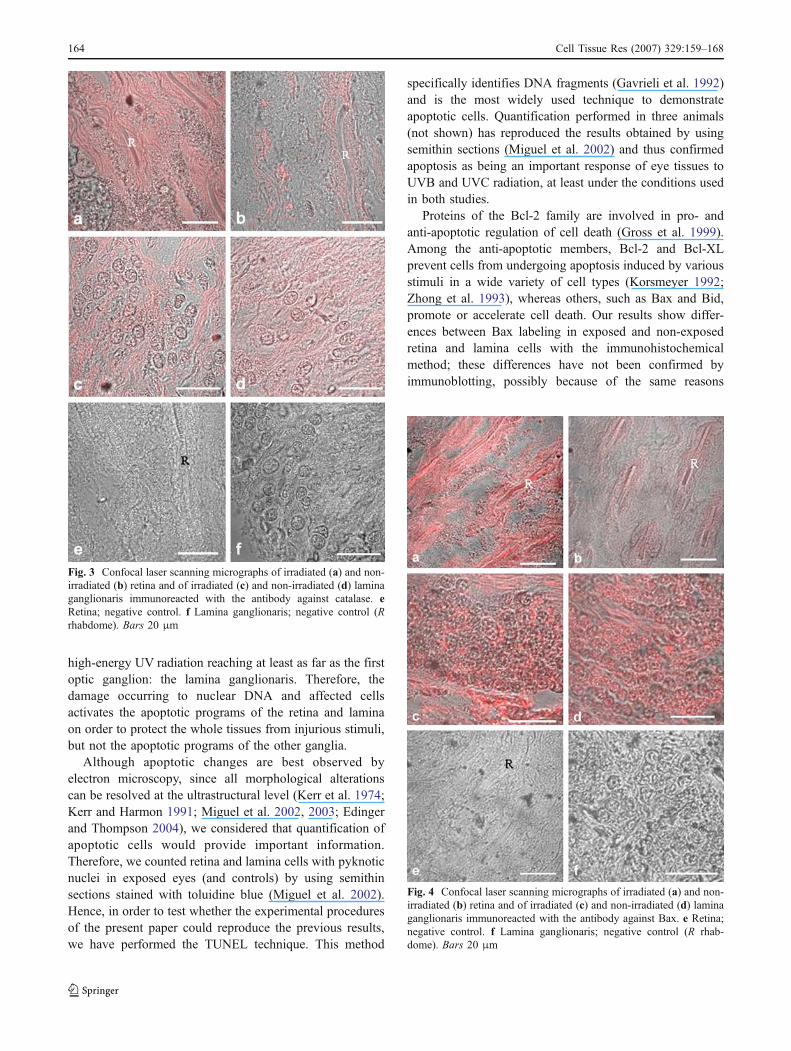
Immunohistochemistry on sections studied by an LSMindicated that catalase, Bax, and p53 were expressed in theretina and in lamina cells of exposed and non-exposed eyes.However, there was a difference in labeling betweenexposed and non-exposed retina and lamina cells: stainingwas much more pronounced in the retina of exposed(Figs. 3a, 4a, 5a) than of non-exposed eyes (Figs. 3b, 4b,5b) and was more pronounced in lamina cells of exposedeyes (Figs. 3c, 4c, 5c) compared with non-exposed eyes(Figs. 3d, 4d, 5). Negative controls for retinas (Figs. 3e, 4e,5e) and for laminas (Figs. 3f, 4f, 5f ) are shown.
In order to confirm the results obtained with theimmunohistochemistry, we performed immunodetection ofcatalase, Bax, and p53 contents after separation of totalproteins from the whole homogenate of the entire opticlobes from the various crabs tested here (see Figs. 6, 7, 8).The proteins were expressed both in irradiated and in non-irradiated retina and lamina cells, with no statisticaldifference in contents between irradiated and non-irradiatedeyes. The intensity of catalase immunostaining afterWestern blotting is shown in Fig. 6. Using 140 μgprotein/lane, we noted a slight tendency for a decrease inthe catalase content from irradiated eyes, but this was notsignificant. A representative experiment (n=4) showing theimmunodetection of catalase is presented in Fig. 6 (inset).
The same result was obtained after Bax immunodetection(Fig. 7). No significant differences were observed forirradiated or non-irradiated eyes. This could be moreclearly observed when we quantified these data afternormalization by rouge Ponceau staining. The plot showsthe ratio of the optical density from the Bax band stained byrouge Ponceau and that from immunodetection. The anti-Bax used clearly identifies a major band of about 21 kDa(inset to Fig. 7b, n=4), which is related to the Bax mobilityas described for other systems. Bcl-2 heterodimerizes in
Fig. 1 Photomicrograph of a longitudinal section of the crab eyestalkstained with hematoxylin and eosin displaying the retina (Re), laminaganglionaris (L), external medulla (EM), and internal medulla (IM).Bar 200 μm
162 Cell Tissue Res (2007) 329:159–168

vivo with a conserved homolog, Bax, that acceleratesprogrammed cell death (Oltval et al. 1993).
Regarding p53, immunodetection (Fig. 8) showed thatits expression was not altered after irradiation, as occurredwith catalase and Bax. p53 detection could be more clearlyobserved when we quantified the data after normalizationby rouge Ponceau staining (n=6).
Discussion
Peroxisomes constitute the primary line of antioxidativedefense, and catalase is the enzyme most used as a markerfor peroxisomes (Cajaraville et al. 1992). Based on the resultsobtained by Cajaraville et al. (2003) and Orbea et al. (2002),increase in the peroxisomal enzymes catalase and acyl-CoAoxidase levels and in peroxisome proliferation have beenproposed as useful biomarkers for exposure to a variety ofpollutants. One of our interests is the effects of UVB andUVC on the visual system of living organisms such as thecrab (Miguel et al. 2002) and rats (Miguel et al. 2003), andcatalase has been detected in organelles in the visual systemcells of U. cordatus by using cytochemistry, immunohisto-chemistry, and immunoelectron microscopy (Miguel et al.2005). In the present work, one of our aims has been to lookfor any alteration in catalase expression in the retina andlamina after the exposure of crabs to UVB and UVCradiation. Our results show a difference in catalase expres-sion as observed by immunohistochemistry, both in theretina and lamina, although this has not been detected by
Western blotting performed on whole optic lobe. Theexposure of tissues to UV is known to produce highly activeintracellular oxygen species (Reddy and Bhat 1999; Schraderet al. 2000); therefore, even though the levels do not changeafter exposure as detected by Western blotting with wholeeyestalk, we suggest that, under the conditions used here,catalase does indeed play a role, as demonstrated by theimmunohistochemistry of retina and lamina cells.
In the case of exposed crabs, the level of catalase afterirradiation may not have been sufficient to protect com-pletely against apoptosis. Based on the available literature,Kahl et al. (2004) have assumed the following with regardto the facilitation or inhibition of apoptosis: (1) theoverexpression or upregulation of catalase leads to aprotection against apoptosis (Tome et al. 2001; Song andLee 2003; Lin et al. 2004); (2) the overexpression orupregulation of catalase facilitates apoptosis, whereas lowcatalase levels confer resistance to apoptotic stimuli(Oberley and Oberley 1997). Our immunohistochemicalresults favor, in principle, the second assumption. However,this difference between the results obtained by immunohis-tochemistry and by immunoblotting could be explained byour having to use most of the eyestalk for the latter, sincethe anatomical organization of the eyestalk (see Fig. 1)makes it unfeasible to dissect the retina and laminaseparately; for the immunohistochemistry, we could directlyobserve the structures themselves under the microscope.Indeed, the apoptotic changes in the retina and lamina cellsof irradiated U. cordatus eyes described in our previouspaper (Miguel et al. 2002) appear to be the direct result of
Fig. 2 TUNEL staining of irra-diated (a) and non-irradiated (b)retina and of irradiated (c) andnon-irradiated (d) lamina gan-glionaris. Note the apoptoticcells (brown, arrows). Counter-staining: hematoxylin. Bars13 μm (a, b),17 μm (c, d)
Cell Tissue Res (2007) 329:159–168 163
high-energy UV radiation reaching at least as far as the firstoptic ganglion: the lamina ganglionaris. Therefore, thedamage occurring to nuclear DNA and affected cellsactivates the apoptotic programs of the retina and laminaon order to protect the whole tissues from injurious stimuli,but not the apoptotic programs of the other ganglia.
Although apoptotic changes are best observed byelectron microscopy, since all morphological alterationscan be resolved at the ultrastructural level (Kerr et al. 1974;Kerr and Harmon 1991; Miguel et al. 2002, 2003; Edingerand Thompson 2004), we considered that quantification ofapoptotic cells would provide important information.Therefore, we counted retina and lamina cells with pyknoticnuclei in exposed eyes (and controls) by using semithinsections stained with toluidine blue (Miguel et al. 2002).Hence, in order to test whether the experimental proceduresof the present paper could reproduce the previous results,we have performed the TUNEL technique. This method
specifically identifies DNA fragments (Gavrieli et al. 1992)and is the most widely used technique to demonstrateapoptotic cells. Quantification performed in three animals(not shown) has reproduced the results obtained by usingsemithin sections (Miguel et al. 2002) and thus confirmedapoptosis as being an important response of eye tissues toUVB and UVC radiation, at least under the conditions usedin both studies.
Proteins of the Bcl-2 family are involved in pro- andanti-apoptotic regulation of cell death (Gross et al. 1999).Among the anti-apoptotic members, Bcl-2 and Bcl-XLprevent cells from undergoing apoptosis induced by variousstimuli in a wide variety of cell types (Korsmeyer 1992;Zhong et al. 1993), whereas others, such as Bax and Bid,promote or accelerate cell death. Our results show differ-ences between Bax labeling in exposed and non-exposedretina and lamina cells with the immunohistochemicalmethod; these differences have not been confirmed byimmunoblotting, possibly because of the same reasons
Fig. 4 Confocal laser scanning micrographs of irradiated (a) and non-irradiated (b) retina and of irradiated (c) and non-irradiated (d) laminaganglionaris immunoreacted with the antibody against Bax. e Retina;negative control. f Lamina ganglionaris; negative control (R rhab-dome). Bars 20 μm
Fig. 3 Confocal laser scanning micrographs of irradiated (a) and non-irradiated (b) retina and of irradiated (c) and non-irradiated (d) laminaganglionaris immunoreacted with the antibody against catalase. eRetina; negative control. f Lamina ganglionaris; negative control (Rrhabdome). Bars 20 μm
164 Cell Tissue Res (2007) 329:159–168

proposed above for catalase (and below for p53), viz.difference in the results between both methods. Thus, eventhough there is no massive production of Bax, perhaps thelevel of activated Bax is enough to trigger cell death inorder to allow the survival of the whole tissue afterirradiation. It follows, then, that tissues are sustained tosome extent by the activation of Bax under conditions oflimiting survival signals. Deprivation of survival factorsactively triggers the activation of Bax (and other mole-cules), rather than simply removing the anti-apoptoticfunctions of certain proteins (Green 2005).
Fig. 8 Total proteins from control (c) and treated (i) animals wereseparated by SDS-PAGE and transferred to nitrocellulose forimmunoblotting analysis. Plot of the ratio obtained by using opticaldensity analysis (O.D.) of the rouge Ponceau staining from the 53-kDaband and the optical density analysis of the 53-kDa band recognizedby the monoclonal anti-p53 antibody. Inset: Immunoblotting detectionof p53
Fig. 7 Total proteins from control (c) and treated (i) animals wereseparated by SDS-PAGE and transferred to nitrocellulose forimmunoblotting analysis. Plot of the ratio obtained by using opticaldensity analysis (O.D.) of the rouge Ponceau staining from the 21-kDaband and the optical density analysis of the 21-kDa band recognizedby the monoclonal anti-Bax antibody. Inset: Immunoblotting detectionof Bax
Fig. 6 Total proteins from control (c) and treated (i) animals wereseparated by SDS-polyacrylamide gel electrophoresis (SDS-PAGE)and transferred to nitrocellulose for immunoblotting analysis. Plot ofthe ratio obtained for the optical density (O.D.) after rouge Ponceaustaining to the optical density of the immunoblot. Inset: Immunoblot-ting detection of catalase by using the polyclonal antibody anti-catalase. A band is clearly immunodetected at about 220 kDa,agreeing with the molecular weight of catalase
Fig. 5 Confocal laser scanning micrographs of irradiated (a) and non-irradiated (b) retina and of irradiated (c) and non-irradiated (d) laminaganglionaris immunoreacted with the antibody against p53. e Retina;negative control. f Lamina ganglionaris; negative control (R rhab-dome). Bars 20 μm
Cell Tissue Res (2007) 329:159–168 165

Awealth of evidence suggests that the damage caused byUV is mediated by more than its ability to affect DNA (Kulmset al. 1999; Kulms and Schwarz 2002). Once DNA isdamaged, p53 is activated, resulting in cell cycle arrest,allowing time for damage repair or promoting cell death(Vousden 2000; Vousden and Lu 2002). In the crab U.cordatus, p53 binding revealed a heavier immunostaining inexposed than in non-exposed retinas and laminas. This resultalone is indicative of cell stress (Giaccia and Kastan 1998)and suggests that non-ionizing radiations such as UVB andUVC lead to the activation of p53, as do ionizing radiations(Brodsky et al. 2000). However, as was the result withcatalase and Bax, p53 expression was not followed by asignificant difference between the immunoblotting of ex-posed and non-exposed eyestalks. Therefore, even thoughthe immunohistochemistry suggests a bias toward anincreased expression of p53, again this difference betweenthe two results could be explained by the finding that, for theimmunoblotting, we had to use most of the eyestalk, whereasfor the immunohistochemistry, the retina and the laminacould be observed separately. A complementary explanationfor this difference could concern the activation of p53.Indeed, p53 levels in control and exposed cells are notextremely different, because irradiation can activate p53 butnot lead to its over-expression. In certain cell types, p53stimulates DNA repair functions, and in others, it inducesapoptosis (Roos et al. 2004; Tomicic et al. 2005).
Several groups have suggested that p53 is involved inthe positive regulation of cell growth (Reich and Levine1984) and in cell transformation (Eliyahu et al. 1984;Parada et al. 1984). In 1989, Finlay et al. established thereal physiological function of p53 in rats: a tumorsuppressor action. In Drosophila, the p53 ortholog Dmp53also acts in the cell response to DNA damage, and over-expression of Dmp53 in the retina can lead to extensive celldeath (Jin et al. 2000; Ollmann et al. 2000). Indeed, UVCradiation exposure at nonlethal doses results in retinaldefects in Drosophila during development. However,whether the effect of the over-expression of p53 on UV-mediated damage alone leads to extensive cell death in theretina has not been assessed (Jassim et al. 2003). Therefore,the determination of whether p53 is also expressed in otheranimal groups such as crustaceans, after exposure to intenseUVB and UVC radiation is of interest. This knowledgemay help confirm the importance of p53 in apoptosisfollowing stress, in addition to contributing to its correla-tion with evolutionary aspects.
When a cell is unable to repair the DNA damage, p53 canbe postulated to induce the activation of the apoptoticsignaling pathways. Among the apoptotic genes controlledby p53 are members of the Bcl-2 family (including Bax), thep53-induced gene family, and death receptor gene expression(CD95, DR5, or PIDD; Ashkenazi and Dixit 1998; Lyn et al.
2000). After p53 stabilization, p53 accumulates in thenucleus to regulate directly the expression of proapoptoticgenes, such as Bax and p53-up-regulated modifier ofapoptosis. Protein p53 also accumulates in the cytoplasmand awaits a secondary death signal allowing mitochondrialpermeabilization (Chipuk et al. 2005).
In conclusion, our experiments have shown that sincecatalase, Bax, and p53, were partially activated byirradiation, UVB and UVC can affect retina and laminacells via a phototransduction cascade. In our experiments,the dose of UVC used was much higher than the dose ofUVB; however, we cannot attribute most of the resultsobtained to UVC. It is possible that both wavelengthscontributed to the activation of the proteins studied.Therefore, knowledge of whether UVB or UVC separatelyproduces the same results in this model would be ofinterest, as would whether UV activates other protectivemechanisms such as that induced by mediators of p53(Sohn et al. 2006).
Acknowledgements We are grateful to Jorge Luís da Silva fortechnical support.
References
Allodi S, Santos LMS, da Silva SF (1995) Histological study of thevisual system in the decapod crustacean Macrobrachium rosen-bergii. Braz J Morphol Sci 12:14–22
Ashkenazi A, Dixit VM (1998) Death receptors: signaling andmodulation. Science 281:1305–1308
Brodsky MH, Nordstrom W, Tsang G, Kwan E, Rubin GM, AbramsJM (2000) Drosophila p53 binds a damage response element atthe reaper locus. Cell 101:103–113
Cajaraville MP, Völkl A, Fahimi HD (1992) Peroxisomes in thedigestive gland cells of the mussel Mytilus galloprovincialisLmk. Biochemical, ultrastructural and immunocytochemicalcharacterization. Eur J Cell Biol 59:255–264
Cajaraville MP, Cancio I, Ibabe A, Orbea A (2003) Peroxisomeproliferation as a biomarker in environmental pollution assess-ment. Microsc Res Tech 61:191–202
Canman CE, Lim DS, Cimprich KA, Taya Y, Tamai K, Sakaguchi K,Appella E, Kastan MB, Siliciano JD (1998) Activation of theATM kinase by ionizing radiation and phosphorylation of p53.Science 281:1677–1679
Chipuk JE, Bouchier-Hayes L, Kuwana T, Newmeyer DD, Green DR(2005) PUMA couples the nuclear and cytoplasmic proapoptoticfunction of p53. Science 309:1732–1735
Chouinard N, Valerie K, Rouabhia M, Huot J (2002) UVB-mediatedactivation of p38 mitogen-activated protein kinase enhancesresistance of normal human keratinocytes to apoptosis bystabilizing cytoplasmic p53. Biochem J 365:133–145
Corrêa CL, Silva SF da, Lowe J, Tortelote GG, Einicker-Lamas M,Martinez AMB, Allodi A (2004) Identification of a neurofila-ment-like protein in the protocerebral tract of the crab Ucidescordatus. Cell Tissue Res 318:609–615
da Silva SF, Corrêa CL, Tortelote GG, Einicker-Lamas M, MartinezAMB, Allodi S (2004) Glial fibrillary acidic protein (GFAP)-likeimmunoreactivity in the visual system of the crab Ucidescordatus (Crustacea, Decapoda). Biol Cell 96:727–734
166 Cell Tissue Res (2007) 329:159–168

Duncan DD, Munoz B, Bandeen-Roche K, West SK (1997)Assessment of ocular exposure to ultraviolet-B for populationstudies. Salisbury eye evaluation project team. PhotochemPhotobiol 66:701–709
Edinger AL, Thompson CB (2004) Death by design: apoptosis,necrosis and autophagy. Curr Opin Cell Biol 16:663–669
Eliyahu D, Raz A, Gruss P, Givol D, Oren M (1984) Participation ofp53 cellular tumour antigen in transformation of normalembryonic cells. Nature 312:646–649
Finlay CA, Hinds PW, Levine AJ (1989) The p53 proto-oncogene canact as a suppressor of transformation. Cell 57:1083–1093
Garcia MX, Foote C, van Es S, Devreotes PN, Alexander S,Alexander H (2000) Differential developmental expression andcell type specificity of Dictyostelium catalases and their responseto oxidative stress and UV-light. Biochim Biophys Acta1492:295–310
Gavrieli Y, Sherman Y, Ben-Sasson SA (1992) Identification ofprogrammed cell death in situ via specific labeling of nuclearDNA fragmentation. J Cell Biol 119:493–501
Giaccia AJ, Kastan MB (1998) The complexity of p53 modulation:emerging patterns from divergent signals. Genes Dev 12:2973–2983
Grassé PP, Poisson RA, Tuzet O (1976) Zoología. 1. Invertebrados.Toray-Masson, Barcelona
Green DR (2005) Apoptotic pathways: ten minutes to dead. Cell121:671–674
Gouveia GR,Marques DS, Cruz BP, Geracitano LA, Nery LE, TrindadeGS (2005) Antioxidant defenses and DNA damage induced byUV-A and UV-B radiation in the crab Chasmagnathus granulata(Decapoda, Brachyura). Photochem Photobiol 81:398–403
Gross A, McDonnell JM, Korsmeyer SJ (1999) BCL-2 familymembers and the mitochondria in apoptosis. Genes Dev13:1899–1911
Guimarães CA, Linden R (2004) Programmed cell death. Apoptosisand alternative deathstyles. Eur J Biochem 271:1638–1650
Halliwell B, Gutteridge JMC (1989) Free radicals in biology andmedicine. Oxford University Press, Oxford
He Q, Huang Y, Sheikh MS, Murphy M, Mabruk MJ, Lenane P, LiewA, McCann P, Buckley A, Billet P, Leader M, Kay E, MurphyGM (2004) Bax deficiency affects caspase-2 activation duringultraviolet radiation-induced apoptosis. Oncogene 23:1321–1325
Jassim OW, Fink JL, Cagan RL (2003) Dmp53 protects theDrosophila retina during a developmentally regulated DNAdamage response. EMBO J 22:5622–5632
Jin S, Martinek S, Joo WS, Wortman JR, Mirkovic N, Sali A, YandellMD, Pavletich NP, Young MW, Levine AJ (2000) Identificationand characterization of a p53 homologue in Drosophila mela-nogaster. Proc Natl Acad Sci USA 97:7301–7306
Kahl R, Kampkotter A, Watjen W, Chovolou Y (2004) Antioxidantand apoptosis. Drug Metab Rev 36:747–762
Kerr JFR, Harmon BV (1991) Definition and incidence of apoptosis:an historical perspective. In: Tornei LD, Cope FO (eds) Themolecular basis of cell death. Cold Spring Harbor Laboratory,Cold Spring Harbor, NY, pp 5–29
Kerr JF, Wyllie AH, Currie AR (1972) Apoptosis: a basic biologicalphenomenon with wide-ranging implications in tissue kinetics.Br J Cancer 26:239–257
Kerr JF, Harmon B, Searle J (1974) An electron-microscope study ofcell deletion in the anuran tadpole tail during spontaneousmetamorphosis with special reference to apoptosis of striatedmuscle fibers. J Cell Sci 14:571–585
Korsmeyer SJ (1992) Bcl-2 initiates a new category of oncogenes:regulators of cell death. Blood 80:879–886
Kulms D, Schwarz T (2002) Independent contribution of threedifferent pathways to ultraviolet-B-induced apoptosis. BiochemPharmacol 64:837–841
Kulms D, Poppelmann B, Yarosh D, Luger TA, Krutman J, Schwarz T(1999) Nuclear and cell membrane effects contribute indepen-dently to the induction of apoptosis in human cells exposed toUVB radiation. Proc Natl Acad Sci USA 96:7974–7979
Lin SJ, Shyue SK, Liu PL, Chen YH, Ku HH, Chen JW, Tam KB,Chen YL (2004) Adenovirus-mediated overexpression of catalaseattenuates oxLDL-induced apoptosis in human aortic endothelialcells via AP-1 and C-Jun N-terminal kinase/extracellular signal-regulated kinase mitogen-activated protein kinase pathways. JMol Cell Cardiol 36:129–139
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Proteinmeasurement with the Folin-phenol reagent. J Biol Chem193:265–275
Lyn D, Liu X, Bennett NA, Emmett NL (2000) Gene expressionprofile in mouse myocardium after ischemia. Physiol Genomics2:93–100
Miguel NCO, Meyer-Rochow VB, Allodi S (2002) Ultrastructuralstudy of first and second order neurons in the visual system of thecrab Ucides cordatus following exposure to ultraviolet radiation.Micron 33:627–637
Miguel NCO, Meyer-Rochow VB, Allodi S (2003) A structural studyof the retinal photoreceptor, plexiform and ganglion cell layersfollowing exposure to UV-B and UV-C radiation in the albino rat.Micron 34:395–404
Miguel NCO, Wajsenzon IJR, Allodi S (2005) The expression ofcatalase in the visual system of the crab Ucides cordatus.Nauplius 13:159–166
Oberley TD, Oberley LW (1997) Antioxidant enzyme levels in cancer.Histol Histopathol 12:525–535
Ollmann M, Young LM, Di Como CJ, Karim F, Belvin M, RobertsonS, Whittaker K, Demsky M, Fisher WW, Buchman A, Duyk G,Friedman L, Prives C, Kopczynski C (2000) Drosophila p53 is astructural and functional homolog of the tumor suppressor p53.Cell 101:91–101
Oltval ZN, Milliman CL, Korsmeyer SJ (1993) Bcl-2 heterodimerizesin vivo with a conserved homolog, Bax, that acceleratesprogrammed cell death. Cell 74:609–619
Orbea A, Ortiz-Zarragoitia M, Solé M, Porte C, Cajaraville MP (2002)Antioxidant enzymes and peroxisome proliferation in relation tocontaminant body burdens of PAHs and PCBs in bivalvemollusks, crabs and fish from the Urdaibai and Plentzia estuaries(Bay of Biscay). Aquat Toxicol 58:75–98
Parada LF, Land H, Weinberg RA, Wolf D, Rotter V (1984)Cooperation between gene encoding P53 tumour antigen andras in cellular transformation. Nature 312:649–651
Pipe RK, Livingstone DR (1993) Antioxidant enzymes associatedwith the blood cell and haemolymph of the mussel Mytilusgalloprovincialis. Fish Shellfish Immunol 3:221–233
Reddy GB, Bhat KS (1999) Protection against UV-B inactivation (invitro) of rat lens enzymes by natural antioxidants. Mol CellBiochem 194:41–45
Reddy VN, Kasahara E, Hiraoka M, Lin LR, Ho YS (2004) Effects ofvariation in superoxide dismutases (SOD) on oxidative stress andapoptosis in lens epithelium. Exp Eye Res 79:859–868
Reich NC, Levine AJ (1984) Growth regulation of a cellular tumourantigen, p53, in nontransformed cells. Nature 308:199–201
Roos W, Baumgartner M, Kaina B (2004) Apoptosis triggered byDNA damage O6-methylguanine in human lymphocytes requiresDNA replication and is mediated by p53 and Fas/CD95/Apo-1.Oncogene 23:359–367
Sanford BE, Beacham S, Hanifin JP, Hannon P, Streletz L, Sliney D,Brainard GC (1996) The effects of ultraviolet-A radiation onvisual evoked potentials in the young human eye. ActaOphthalmol Scand 74:553–557
Schrader M, King SJ, Stroh TA (2000) Real time imaging reveals aperoxisomal reticulum in living cells. J Cell Sci 113:3663–3671
Cell Tissue Res (2007) 329:159–168 167

She QB, Chen N, Dong Z (2000) ERKs and p38 kinase phosphorylatep53 protein at serine 15 in response to UV radiation. J Biol Chem275:20444–20449
Sohn D, Essmann F, Schulze-Osthoff K, Janicke RU (2006) P21blocks irradiation-induced apoptosis downstream of mitochon-dria by inhibition of cyclin-dependent kinase-mediated caspase-9activation. Cancer Res 66:11254–11262
Song JJ, Lee YJ (2003) Catalase, but not MnSOD, inhibits glucosedeprivation-activated ASK1-MEK-MAPK signal transductionpathway and prevents relocalization of daxx: hydrogen peroxideas a major second messenger of metabolic oxidative stress. J CellBiochem 90:304–314
Tome ME, Baker AF, Powis G, Payne CM, Briehl MM (2001)Catalase-overexpressing thymocytes are resistant to glucocorti-
coid-induced apoptosis and exhibit increased net tumor growth.Cancer Res 61:2766–2773
Tomicic MT, Christmann M, Kaina B (2005) Apoptosis in UV-C lightirradiated p53 wild-type, apaf-1 and p53 knockout mouseembryonic fibroblasts: interplay of receptor and mitochondrialpathway. Apoptosis 10:1295–1304
Vousden KH (2000) P53: death star. Cell 101:691–694Vousden KH, Lu X (2002) Live or let die: the cell´s response to p53.
Natl Rev Cancer 2:594–604Wyllie AH (1985) The biology of cell death in tumours. Anticancer
Res 5:131–136Zhong LT, Sarafian T, Kane DJ, Charles AC, Mah SP, Edwards RH,
Bredesen DEZ (1993) Bcl-2 inhibits death of central neural cellsinduced bymultiple agents. Proc Natl Acad Sci USA 90:4533–4537
168 Cell Tissue Res (2007) 329:159–168
Related Documents











