Carnivorous leaves from Baltic amber Eva-Maria Sadowski a , Leyla J. Seyfullah a , Friederike Sadowski b , Andreas Fleischmann c , Hermann Behling d , and Alexander R. Schmidt a,1 a Department of Geobiology, University of Göttingen, 37077 Göttingen, Germany; b Institute for Interdisciplinary Research on Conflict and Violence, Bielefeld University, 33615 Bielefeld, Germany; c Botanische Staatssammlung München, 80638 Munich, Germany; and d Department of Palynology and Climate Dynamics, Albrecht von Haller Institute of Plant Sciences, University of Göttingen, 37073 Göttingen, Germany Edited by Peter R. Crane, Yale School of Forestry and Environmental Studies, New Haven, CT, and approved November 3, 2014 (received for review August 1, 2014) The fossil record of carnivorous plants is very scarce and macro- fossil evidence has been restricted to seeds of the extant aquatic genus Aldrovanda of the Droseraceae family. No case of carnivo- rous plant traps has so far been reported from the fossil record. Here, we present two angiosperm leaves enclosed in a piece of Eocene Baltic amber that share relevant morphological features with extant Roridulaceae, a carnivorous plant family that is today endemic to the Cape flora of South Africa. Modern Roridula species are unique among carnivorous plants as they digest prey in a complex mutualistic association in which the prey-derived nutrient uptake depends on heteropteran insects. As in extant Roridula, the fossil leaves possess two types of plant trichomes, in- cluding unicellular hairs and five size classes of multicellular stalked glands (or tentacles) with an apical pore. The apices of the narrow and perfectly tapered fossil leaves end in a single tentacle, as in both modern Roridula species. The glandular hairs of the fossils are re- stricted to the leaf margins and to the abaxial lamina, as in extant Roridula gorgonias. Our discovery supports current molecular age estimates for Roridulaceae and suggests a wide Eocene distribution of roridulid plants. plant carnivory | Roridulaceae | Eocene | Ericales P lant carnivory is traditionally defined as the attraction, cap- ture, and digestion of prey by vegetative traps, with the subsequent uptake of nutrients (1, 2). Some carnivorous plants, however, challenge the boundary of the botanical carnivory con- cept because they depend on commensal organisms for the di- gestion of their prey (2, 3). The most famous representative of those plants is Roridula, placed in the monogeneric family Roridulaceae that is endemic to a few localities in the south- western Cape of South Africa (4, 5). The resinous glandular leaves of both extant species, Roridula dentata and Roridula gorgonias, capture plenty of arthropods. The sticky trapping glue of Roridula is a viscous lipophilic resin containing triterpenoids as major component, which does not allow dissolution of digestive enzymes (6). Consequently, the secretory glands of Roridulaceae lack enzymatic activity (7, 8). For prey-derived nutrient uptake, Roridula depends on two ob- ligately associated heteropteran Pameridea species (family Miridae, “capsid bugs”), which feed on the trapped animals (5, 9). In this “digestive mutualism” (10), the nutrient-rich fecal compounds of these “Roridula bugs” are incorporated by Roridula through nanometer-sized cuticular gaps and serve for a better alimenta- tion in a nutrient-poor habitat (7, 8, 10, 11). The benefit of nu- trient uptake from captured prey is the essential criterion for the concept of botanical carnivory (1, 2) and thus includes Roridulaceae (11, 12). Here, we report two leaf fossils from Eocene Baltic amber possessing the relevant morphological features of an adhesive flypaper trap plant that we assign to the Roridulaceae lineage (Figs. 1–3). Both specimens originate from the Jantarny amber mine near Kaliningrad (Russia). The amber-bearing sediments of this fossil site date to 35–47 million years ago (13, 14). Results The linear-lanceolate leaves are 5 and 4.5 mm long and 0.2 mm wide at the base, and they narrow gradually toward the leaf tip, which terminates in a stalked gland (tentacle; Fig. 1). The leaves possess two trichome types: tentacles and nonglandular hyaline hairs (Figs. 1 and 2). The hyaline trichomes are located on both sides and the margins of the lamina, whereas the tentacles are exclusively found along the margins and on the abaxial side without a definite arrangement (Fig. 1). The tentacles are multi- cellular, consisting of a tapering stalk and a clavate to ovoid glandular head, which shows a small pore at the center of its distal side (Fig. 1 E and F). The stalks of the glands measure between 20 and 350 μm in length (Figs. 1 and 2 A and C), whereas an ex- ceptional stalk exceeds this size, reaching 1.4 mm (Fig. 3A). As with the stalks, the glandular heads vary in size (20–120 μm long, 10–40 μm wide). Adhered organic remains as well as trichomes of other plants attached to the glandular tentacle heads (Figs. 1 A and D and 3A) indicate that they excreted a sticky secretion, as known from adhesive traps of extant carnivorous plants. The non- glandular trichomes are hyaline, unicellular, and arcuate to straight, tapering toward an acute apex (Figs. 1C and 2E). Their length ranges from 10 to 80 μm, and their width reaches up to 12 μm. Both leaves exhibit a well-preserved epidermis with small tetrag- onal cells at the leaf base and elongated larger cells from the middle part toward the leaf tip. These cells measure 3–54 × 6–18 μm. Stomata of 20–38 × 15–25 μm are present on the abaxial leaf side (Figs. 1C and 2G). Our statistical cluster analyses (SI Text) revealed that the fossil specimens and Roridula gorgonias show the same mor- phological pattern among the tentacles. R. gorgonias exhibits five Significance Amber, fossil tree resin, preserves organisms in microscopic fidelity, and frequently fossils preserved in amber are other- wise absent in the entire fossil record. Plant remains, however, are rarely entrapped in amber, compared with the vast amount of insects and other animals. Our newly discovered fossils from Eocene Baltic amber are the only documented case of fossilized carnivorous plant traps and represent the first fossil evidence of the carnivorous plant family Roridulaceae, which is today a narrow endemic of South Africa. Hence, our results shed light onto the paleobiogeography of the Roridulaceae, indicating a wide Eocene distribution of the roridulid ancestors and chal- lenging previous notions about a Gondwanan origin of this plant family. Author contributions: E.-M.S., L.J.S., H.B., and A.R.S. designed research; E.-M.S., L.J.S., F.S., A.F., H.B., and A.R.S. performed research; E.-M.S., L.J.S., F.S., A.F., and A.R.S. analyzed data; and E.-M.S., L.J.S., A.F., and A.R.S. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. 1 To whom correspondence should be addressed. Email: [email protected] goettingen.de. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1414777111/-/DCSupplemental. www.pnas.org/cgi/doi/10.1073/pnas.1414777111 PNAS Early Edition | 1 of 6 EVOLUTION

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Carnivorous leaves from Baltic amberEva-Maria Sadowskia, Leyla J. Seyfullaha, Friederike Sadowskib, Andreas Fleischmannc, Hermann Behlingd,and Alexander R. Schmidta,1
aDepartment of Geobiology, University of Göttingen, 37077 Göttingen, Germany; bInstitute for Interdisciplinary Research on Conflict and Violence, BielefeldUniversity, 33615 Bielefeld, Germany; cBotanische Staatssammlung München, 80638 Munich, Germany; and dDepartment of Palynology and ClimateDynamics, Albrecht von Haller Institute of Plant Sciences, University of Göttingen, 37073 Göttingen, Germany
Edited by Peter R. Crane, Yale School of Forestry and Environmental Studies, New Haven, CT, and approved November 3, 2014 (received for review August1, 2014)
The fossil record of carnivorous plants is very scarce and macro-fossil evidence has been restricted to seeds of the extant aquaticgenus Aldrovanda of the Droseraceae family. No case of carnivo-rous plant traps has so far been reported from the fossil record.Here, we present two angiosperm leaves enclosed in a piece ofEocene Baltic amber that share relevant morphological featureswith extant Roridulaceae, a carnivorous plant family that is todayendemic to the Cape flora of South Africa. Modern Roridulaspecies are unique among carnivorous plants as they digest preyin a complex mutualistic association in which the prey-derivednutrient uptake depends on heteropteran insects. As in extantRoridula, the fossil leaves possess two types of plant trichomes, in-cluding unicellular hairs and five size classes of multicellular stalkedglands (or tentacles) with an apical pore. The apices of the narrowand perfectly tapered fossil leaves end in a single tentacle, as in bothmodern Roridula species. The glandular hairs of the fossils are re-stricted to the leaf margins and to the abaxial lamina, as in extantRoridula gorgonias. Our discovery supports current molecular ageestimates for Roridulaceae and suggests a wide Eocene distributionof roridulid plants.
plant carnivory | Roridulaceae | Eocene | Ericales
Plant carnivory is traditionally defined as the attraction, cap-ture, and digestion of prey by vegetative traps, with the
subsequent uptake of nutrients (1, 2). Some carnivorous plants,however, challenge the boundary of the botanical carnivory con-cept because they depend on commensal organisms for the di-gestion of their prey (2, 3). The most famous representativeof those plants is Roridula, placed in the monogeneric familyRoridulaceae that is endemic to a few localities in the south-western Cape of South Africa (4, 5).The resinous glandular leaves of both extant species, Roridula
dentata and Roridula gorgonias, capture plenty of arthropods.The sticky trapping glue of Roridula is a viscous lipophilic resincontaining triterpenoids as major component, which does notallow dissolution of digestive enzymes (6). Consequently, thesecretory glands of Roridulaceae lack enzymatic activity (7, 8).For prey-derived nutrient uptake, Roridula depends on two ob-ligately associated heteropteran Pameridea species (family Miridae,“capsid bugs”), which feed on the trapped animals (5, 9). In this“digestive mutualism” (10), the nutrient-rich fecal compounds ofthese “Roridula bugs” are incorporated by Roridula throughnanometer-sized cuticular gaps and serve for a better alimenta-tion in a nutrient-poor habitat (7, 8, 10, 11). The benefit of nu-trient uptake from captured prey is the essential criterionfor the concept of botanical carnivory (1, 2) and thus includesRoridulaceae (11, 12).Here, we report two leaf fossils from Eocene Baltic amber
possessing the relevant morphological features of an adhesiveflypaper trap plant that we assign to the Roridulaceae lineage(Figs. 1–3). Both specimens originate from the Jantarny ambermine near Kaliningrad (Russia). The amber-bearing sedimentsof this fossil site date to 35–47 million years ago (13, 14).
ResultsThe linear-lanceolate leaves are 5 and 4.5 mm long and 0.2 mmwide at the base, and they narrow gradually toward the leaf tip,which terminates in a stalked gland (tentacle; Fig. 1). The leavespossess two trichome types: tentacles and nonglandular hyalinehairs (Figs. 1 and 2). The hyaline trichomes are located on bothsides and the margins of the lamina, whereas the tentacles areexclusively found along the margins and on the abaxial sidewithout a definite arrangement (Fig. 1). The tentacles are multi-cellular, consisting of a tapering stalk and a clavate to ovoidglandular head, which shows a small pore at the center of its distalside (Fig. 1 E and F). The stalks of the glands measure between 20and 350 μm in length (Figs. 1 and 2 A and C), whereas an ex-ceptional stalk exceeds this size, reaching 1.4 mm (Fig. 3A). Aswith the stalks, the glandular heads vary in size (20–120 μm long,10–40 μm wide). Adhered organic remains as well as trichomes ofother plants attached to the glandular tentacle heads (Figs. 1A and D and 3A) indicate that they excreted a sticky secretion, asknown from adhesive traps of extant carnivorous plants. The non-glandular trichomes are hyaline, unicellular, and arcuate to straight,tapering toward an acute apex (Figs. 1C and 2E). Their lengthranges from 10 to 80 μm, and their width reaches up to 12 μm.Both leaves exhibit a well-preserved epidermis with small tetrag-
onal cells at the leaf base and elongated larger cells from the middlepart toward the leaf tip. These cells measure 3–54 × 6–18 μm.Stomata of 20–38 × 15–25 μm are present on the abaxial leaf side(Figs. 1C and 2G).Our statistical cluster analyses (SI Text) revealed that the
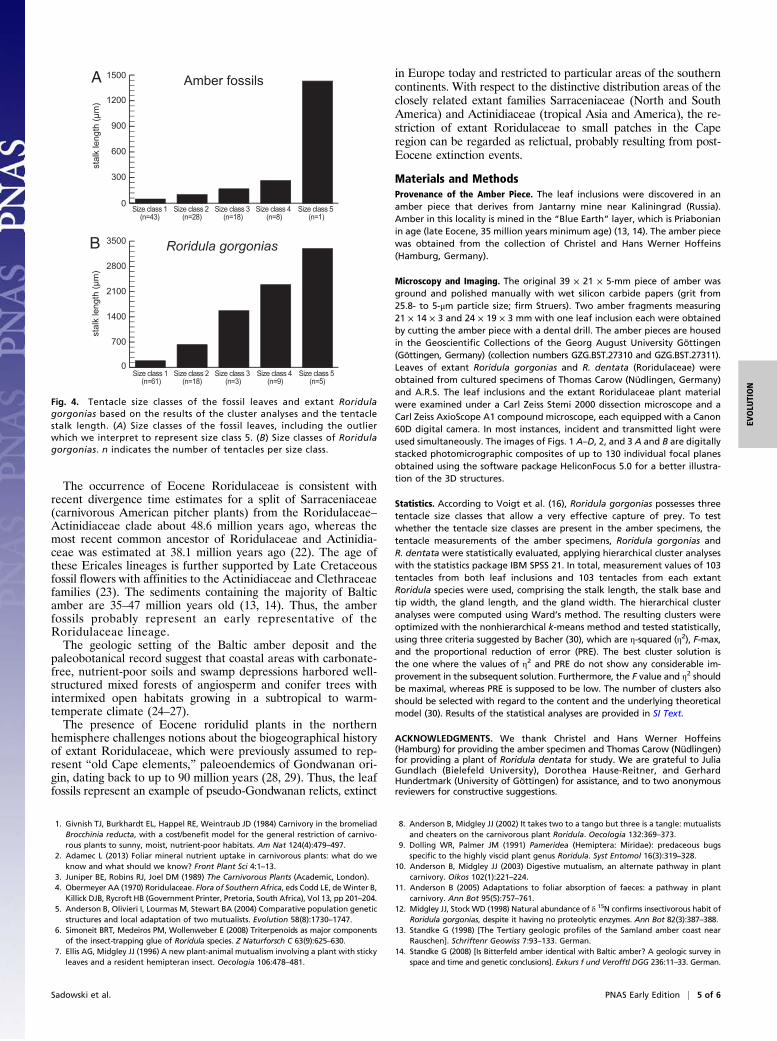
fossil specimens and Roridula gorgonias show the same mor-phological pattern among the tentacles. R. gorgonias exhibits five
Significance
Amber, fossil tree resin, preserves organisms in microscopicfidelity, and frequently fossils preserved in amber are other-wise absent in the entire fossil record. Plant remains, however,are rarely entrapped in amber, compared with the vast amountof insects and other animals. Our newly discovered fossils fromEocene Baltic amber are the only documented case of fossilizedcarnivorous plant traps and represent the first fossil evidenceof the carnivorous plant family Roridulaceae, which is today anarrow endemic of South Africa. Hence, our results shed lightonto the paleobiogeography of the Roridulaceae, indicating awide Eocene distribution of the roridulid ancestors and chal-lenging previous notions about a Gondwanan origin of thisplant family.
Author contributions: E.-M.S., L.J.S., H.B., and A.R.S. designed research; E.-M.S., L.J.S., F.S.,A.F., H.B., and A.R.S. performed research; E.-M.S., L.J.S., F.S., A.F., and A.R.S. analyzeddata; and E.-M.S., L.J.S., A.F., and A.R.S. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1414777111/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1414777111 PNAS Early Edition | 1 of 6
EVOLU
TION
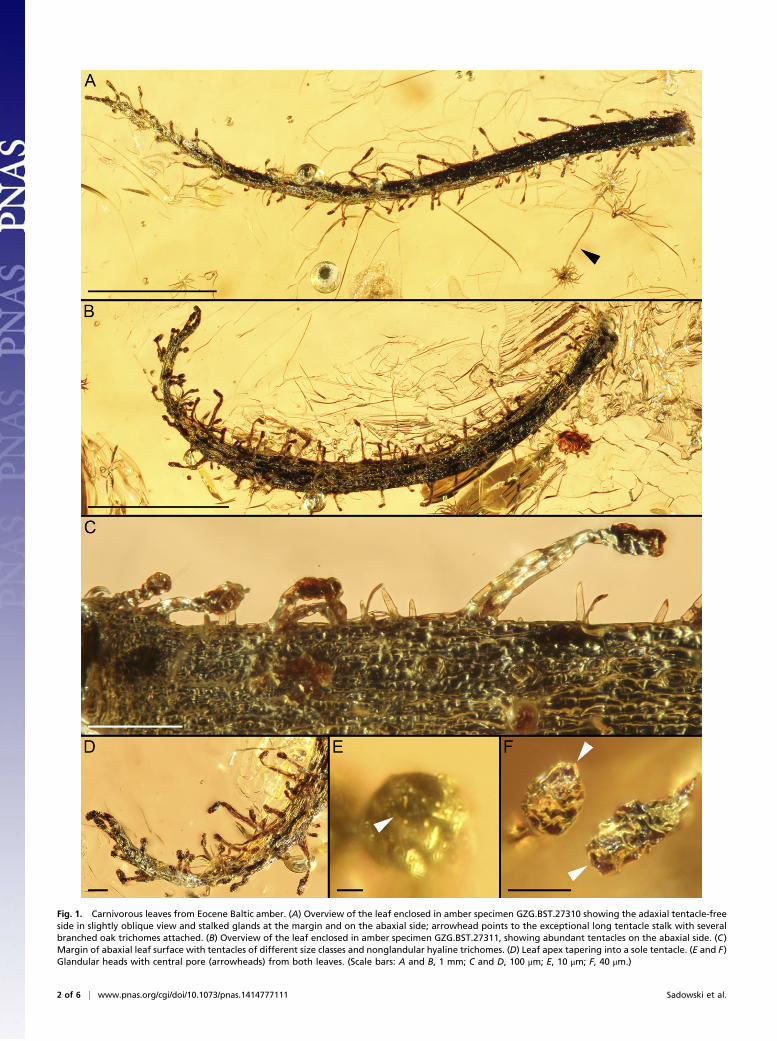
Fig. 1. Carnivorous leaves from Eocene Baltic amber. (A) Overview of the leaf enclosed in amber specimen GZG.BST.27310 showing the adaxial tentacle-freeside in slightly oblique view and stalked glands at the margin and on the abaxial side; arrowhead points to the exceptional long tentacle stalk with severalbranched oak trichomes attached. (B) Overview of the leaf enclosed in amber specimen GZG.BST.27311, showing abundant tentacles on the abaxial side. (C)Margin of abaxial leaf surface with tentacles of different size classes and nonglandular hyaline trichomes. (D) Leaf apex tapering into a sole tentacle. (E and F)Glandular heads with central pore (arrowheads) from both leaves. (Scale bars: A and B, 1 mm; C and D, 100 μm; E, 10 μm; F, 40 μm.)
2 of 6 | www.pnas.org/cgi/doi/10.1073/pnas.1414777111 Sadowski et al.

Fig. 2. Morphological comparison of the carnivorous leaf fossils from Baltic amber (Left) and extant Roridula species (Right). (A and B) Leaf tip ending in a soletentacle. (C and D) Stalked glands of different size classes. (E and F) Hyaline unicellular nonglandular trichomes. (G and H) Epidermal cells and stomata. (I–L) Mul-ticellular tentacles. (A, C, E, and G) GZG.BST.27310. (I and J) GZG.BST.27311. (B, D, K, and L) R. gorgonias. (F and H) R. dentata. (Scale bars: A–D, 100 μm; E–L, 50 μm.)
Sadowski et al. PNAS Early Edition | 3 of 6
EVOLU
TION

size clusters of tentacle stalk lengths whereas four clusters weredetected for the amber inclusions. As an outlier, the longest ten-tacle was excluded from the cluster analyses of the fossils, but itspresence indicates that a fifth size class was present in the Eoceneleaves, as in extant Roridulaceae (Fig. 4).
DiscussionAlthough glandular secreting trichomes appear in about 30% ofvascular plants (15), the unique character combination in thefossils bears most similarities to extant representatives of theRoridulaceae. They share the long, narrow, and perfectly ta-pered leaf lamina ending in a single tentacle, the presence andmorphology of two trichome types (tentacles and nonglandularhairs), the possession of glandular hairs along the leaf marginsand on the abaxial lamina, the tentacle head with a central pore,and the size and shape of the epidermal cells and stomata. Theglabrous adaxial side of the amber inclusions and the hyalinetrichomes being located on both leaf surfaces only appear in thesepals of extant Roridula gorgonias. Besides smaller tentaclesizes, the fossils are distinguished from leaves of extant Roridulaspecies by the absence of a prominent midrib on the abaxial leafside, and are thus most similar to sepals of Roridula gorgonias.
Extant Roridula plants are very effective traps for all kinds ofarthropods due to the sticky resinous trapping glue and the hier-archical organization of the tentacles into functional units for ef-fective prey capture (16, 17). The longest tentacles make the firstcontact with the prey. Due to the high flexibility of these prom-inent tentacles, the moving prey then gets stuck to the medium-sized tentacles, which slow down the caught animal. Finally, thesmallest and stiffest tentacles immobilize the prey (16). As inmodern Roridulaceae, the leaf fossils have different size classes oftentacles that fulfill the functional roles for prey capture (entan-glement, slow-down, and immobilization) and comply with therequirements for a carnivorous nature. In addition, the pore of thetentacle heads distinguishes the fossils from any other extantcarnivorous plants with glandular adhesive traps such as sundews(Drosera) (3, 18, 19).In the fossil record, evidence of carnivorous plants is exceed-
ingly rare and macrofossils are restricted to seeds of the aquaticcarnivore Aldrovanda (Droseraceae), which are recorded sincethe Eocene (20, 21). Hence, the fossil leaves from Baltic amberare (to our knowledge) the first documented case of carnivorousplant traps being fossilized.
Fig. 3. Carnivorous leaf from Eocene Baltic amber (A and B; GZG.BST.27310) and leaves of extant Roridula gorgonias (C and D). (A) Exceptionallylong tentacle stalk (with several branched oak trichomes attached) of the fossil leaf representing the fifth size class of stalked glands. (B and C ) Overviewsshowing the tentacle-free adaxial surface and tentacles along the leaf margins. (D) Partial leaf tip showing different size classes of stalked glands. (Scalebars: A, 100 μm; B, 500 μm; C and D, 1 mm.)
4 of 6 | www.pnas.org/cgi/doi/10.1073/pnas.1414777111 Sadowski et al.
The occurrence of Eocene Roridulaceae is consistent withrecent divergence time estimates for a split of Sarraceniaceae(carnivorous American pitcher plants) from the Roridulaceae–Actinidiaceae clade about 48.6 million years ago, whereas themost recent common ancestor of Roridulaceae and Actinidia-ceae was estimated at 38.1 million years ago (22). The age ofthese Ericales lineages is further supported by Late Cretaceousfossil flowers with affinities to the Actinidiaceae and Clethraceaefamilies (23). The sediments containing the majority of Balticamber are 35–47 million years old (13, 14). Thus, the amberfossils probably represent an early representative of theRoridulaceae lineage.The geologic setting of the Baltic amber deposit and the
paleobotanical record suggest that coastal areas with carbonate-free, nutrient-poor soils and swamp depressions harbored well-structured mixed forests of angiosperm and conifer trees withintermixed open habitats growing in a subtropical to warm-temperate climate (24–27).The presence of Eocene roridulid plants in the northern
hemisphere challenges notions about the biogeographical historyof extant Roridulaceae, which were previously assumed to rep-resent “old Cape elements,” paleoendemics of Gondwanan ori-gin, dating back to up to 90 million years (28, 29). Thus, the leaffossils represent an example of pseudo-Gondwanan relicts, extinct
in Europe today and restricted to particular areas of the southerncontinents. With respect to the distinctive distribution areas of theclosely related extant families Sarraceniaceae (North and SouthAmerica) and Actinidiaceae (tropical Asia and America), the re-striction of extant Roridulaceae to small patches in the Caperegion can be regarded as relictual, probably resulting from post-Eocene extinction events.
Materials and MethodsProvenance of the Amber Piece. The leaf inclusions were discovered in anamber piece that derives from Jantarny mine near Kaliningrad (Russia).Amber in this locality is mined in the “Blue Earth” layer, which is Priabonianin age (late Eocene, 35 million years minimum age) (13, 14). The amber piecewas obtained from the collection of Christel and Hans Werner Hoffeins(Hamburg, Germany).
Microscopy and Imaging. The original 39 × 21 × 5-mm piece of amber wasground and polished manually with wet silicon carbide papers (grit from25.8- to 5-μm particle size; firm Struers). Two amber fragments measuring21 × 14 × 3 and 24 × 19 × 3 mm with one leaf inclusion each were obtainedby cutting the amber piece with a dental drill. The amber pieces are housedin the Geoscientific Collections of the Georg August University Göttingen(Göttingen, Germany) (collection numbers GZG.BST.27310 and GZG.BST.27311).Leaves of extant Roridula gorgonias and R. dentata (Roridulaceae) wereobtained from cultured specimens of Thomas Carow (Nüdlingen, Germany)and A.R.S. The leaf inclusions and the extant Roridulaceae plant materialwere examined under a Carl Zeiss Stemi 2000 dissection microscope and aCarl Zeiss AxioScope A1 compound microscope, each equipped with a Canon60D digital camera. In most instances, incident and transmitted light wereused simultaneously. The images of Figs. 1 A–D, 2, and 3 A and B are digitallystacked photomicrographic composites of up to 130 individual focal planesobtained using the software package HeliconFocus 5.0 for a better illustra-tion of the 3D structures.
Statistics. According to Voigt et al. (16), Roridula gorgonias possesses threetentacle size classes that allow a very effective capture of prey. To testwhether the tentacle size classes are present in the amber specimens, thetentacle measurements of the amber specimens, Roridula gorgonias andR. dentata were statistically evaluated, applying hierarchical cluster analyseswith the statistics package IBM SPSS 21. In total, measurement values of 103tentacles from both leaf inclusions and 103 tentacles from each extantRoridula species were used, comprising the stalk length, the stalk base andtip width, the gland length, and the gland width. The hierarchical clusteranalyses were computed using Ward’s method. The resulting clusters wereoptimized with the nonhierarchical k-means method and tested statistically,using three criteria suggested by Bacher (30), which are η-squared (η2), F-max,and the proportional reduction of error (PRE). The best cluster solution isthe one where the values of η2 and PRE do not show any considerable im-provement in the subsequent solution. Furthermore, the F value and η2 shouldbe maximal, whereas PRE is supposed to be low. The number of clusters alsoshould be selected with regard to the content and the underlying theoreticalmodel (30). Results of the statistical analyses are provided in SI Text.
ACKNOWLEDGMENTS. We thank Christel and Hans Werner Hoffeins(Hamburg) for providing the amber specimen and Thomas Carow (Nüdlingen)for providing a plant of Roridula dentata for study. We are grateful to JuliaGundlach (Bielefeld University), Dorothea Hause-Reitner, and GerhardHundertmark (University of Göttingen) for assistance, and to two anonymousreviewers for constructive suggestions.
1. Givnish TJ, Burkhardt EL, Happel RE, Weintraub JD (1984) Carnivory in the bromeliadBrocchinia reducta, with a cost/benefit model for the general restriction of carnivo-rous plants to sunny, moist, nutrient-poor habitats. Am Nat 124(4):479–497.
2. Adamec L (2013) Foliar mineral nutrient uptake in carnivorous plants: what do weknow and what should we know? Front Plant Sci 4:1–13.
3. Juniper BE, Robins RJ, Joel DM (1989) The Carnivorous Plants (Academic, London).4. Obermeyer AA (1970) Roridulaceae. Flora of Southern Africa, eds Codd LE, deWinter B,
Killick DJB, Rycroft HB (Government Printer, Pretoria, South Africa), Vol 13, pp 201–204.5. Anderson B, Olivieri I, Lourmas M, Stewart BA (2004) Comparative population genetic
structures and local adaptation of two mutualists. Evolution 58(8):1730–1747.6. Simoneit BRT, Medeiros PM, Wollenweber E (2008) Triterpenoids as major components
of the insect-trapping glue of Roridula species. Z Naturforsch C 63(9):625–630.7. Ellis AG, Midgley JJ (1996) A new plant-animal mutualism involving a plant with sticky
leaves and a resident hemipteran insect. Oecologia 106:478–481.
8. Anderson B, Midgley JJ (2002) It takes two to a tango but three is a tangle: mutualistsand cheaters on the carnivorous plant Roridula. Oecologia 132:369–373.
9. Dolling WR, Palmer JM (1991) Pameridea (Hemiptera: Miridae): predaceous bugsspecific to the highly viscid plant genus Roridula. Syst Entomol 16(3):319–328.
10. Anderson B, Midgley JJ (2003) Digestive mutualism, an alternate pathway in plantcarnivory. Oikos 102(1):221–224.
11. Anderson B (2005) Adaptations to foliar absorption of faeces: a pathway in plantcarnivory. Ann Bot 95(5):757–761.
12. Midgley JJ, Stock WD (1998) Natural abundance of δ 15N confirms insectivorous habit ofRoridula gorgonias, despite it having no proteolytic enzymes. Ann Bot 82(3):387–388.
13. Standke G (1998) [The Tertiary geologic profiles of the Samland amber coast nearRauschen]. Schriftenr Geowiss 7:93–133. German.
14. Standke G (2008) [Is Bitterfeld amber identical with Baltic amber? A geologic survey inspace and time and genetic conclusions]. Exkurs f und Verofftl DGG 236:11–33. German.
Amber fossils
Size class 5 (n=5)
Size class 4 (n=9)
Size class 3 (n=3)
Size class 2 (n=18)
Size class 1 (n=61)
B
0
700
1400
2100
2800
3500 Roridula gorgonias
stal
k le
ngth
(µm
)st
alk
leng
th (µ
m)
0
300
600
900
1200
1500
Size class 5 (n=1)
Size class 4 (n=8)
Size class 3 (n=18)
Size class 2 (n=28)
Size class 1(n=43)
A
Fig. 4. Tentacle size classes of the fossil leaves and extant Roridulagorgonias based on the results of the cluster analyses and the tentaclestalk length. (A) Size classes of the fossil leaves, including the outlierwhich we interpret to represent size class 5. (B) Size classes of Roridulagorgonias. n indicates the number of tentacles per size class.
Sadowski et al. PNAS Early Edition | 5 of 6
EVOLU
TION

15. Wagner GJ, Wang E, Shepherd RW (2004) New approaches for studying and ex-ploiting an old protuberance, the plant trichome. Ann Bot 93(1):3–11.
16. Voigt D, Gorb E, Gorb S (2009) Hierarchical organisation of the trap in the proto-carnivorous plant Roridula gorgonias (Roridulaceae). J Exp Biol 212(19):3184–3191.
17. Voigt D, Gorb S (2010) Desiccation resistance of adhesive secretion in the proto-carnivorous plant Roridula gorgonias as an adaptation to periodically dry environ-ment. Planta 232(6):1511–1515.
18. Lloyd FE (1934) Is Roridula a carnivorous plant? Can J Res 10:780–786.19. Lloyd FE (1942) The Carnivorous Plants, ed Verdoorn F (Chronica Botanica Company,
Waltham, MA).20. Degreef JD (1997) Fossil Aldrovanda. Int Carn Pl Newsletter 26:93–97.21. He�rmanová Z, Kvacek J (2010) Late Cretaceous Palaeoaldrovanda, not seeds of a
carnivorous plant, but eggs of an insect. Journal of the National Museum (Prague)179(9):105–118.
22. Ellison AM, et al. (2012) Phylogeny and biogeography of the carnivorous plant familySarraceniaceae. PLoS One 7(6):e39291.
23. Schönenberger J, et al. (2012) Glandulocalyx upatoiensis, a fossil flower of Ericales(Actinidiaceae/Clethraceae) from the Late Cretaceous (Santonian) of Georgia, USA.Ann Bot (Lond) 109(5):921–936.
24. Kohlman-Adamska A (2001) A graphic reconstruction of an “amber” forest. The
Amber Treasure Trove. Part 1. The Tadeusz Giecewicz’s Collection at the Museum of
the Earth, Polish Academy of Sciences, Warsaw, ed Kosmowska-Ceranowicz B
(Oficyna Wydawnicza Sadyba, Warsaw), pp 15–18.25. Mosbrugger V, Utescher T, Dilcher DL (2005) Cenozoic continental climatic evolution
of Central Europe. Proc Natl Acad Sci USA 102(42):14964–14969.26. Collinson ME (1992) Vegetational and floristic changes around the Eocene/Oli-
gocene boundary in western and central Europe. Eocene-Oligocene Climatic and
Biotic Evolution, eds Prothero DR, Berggren WA (Princeton Univ Press, Princeton),
pp 437–450.27. Collinson ME, Hooker JJ (2003) Paleogene vegetation of Eurasia: framework for
mammalian faunas. Deinsea 10:41–83.28. Warren BH, Hawkins JA (2006) The distribution of species diversity across a flora’s
component lineages: dating the Cape’s “relicts.” Proc Biol Sci 273:2149–2158.29. Goldblatt P, Manning JC (2002) Plant diversity of the Cape Region of southern Africa.
Ann Mo Bot Gard 89(1):281–302.30. Bacher J (2001) [Test statistics to determine the number of clusters in Quick Cluster].
ZA-Information/Zentralarchiv für Empirische Sozialforschung 48:71–97. German.
6 of 6 | www.pnas.org/cgi/doi/10.1073/pnas.1414777111 Sadowski et al.

Supporting InformationSadowski et al. 10.1073/pnas.1414777111SI Text
Results of the Cluster AnalysesCluster Analyses of the Amber Inclusions. Before cluster analyseswere performed, it was tested whether the assumptions for clusteranalyses, ametric level ofmeasurement and the absence of outliers,weremet. The accuracy of data entry, missing values, skewness, andkurtosis were also examined. Five outliers were excluded (TableS1, numbers 9, 28, 62, 73, and 99).Due to high correlations between the variables stalk length and
gland length (r = 0.6), it was decided to run the cluster analysistwice, excluding the length of the glandular head in the first runand without the stalk length in the second run.The four-cluster solution is supported by the increase of η2
from 0.84 to 0.91 (Table S2). Moreover, the values of η2 do notincrease considerably in the subsequent solution. Hence, the ad-dition of more clusters would not improve the results. Regardingthe proportional reduction of error (PRE) coefficent, the value forfour clusters is low (0.46) and decreases to 0.28 if another clusteris added, indicating minor improvements for the five-cluster so-lution. However, the value of F-max is not maximal. The six-cluster solution is supported by the maximal F value and the lowPRE (0.26), but the η2 value does not increase substantially for thesolution of six clusters (Table S2).All in all, the first analysis revealed a clear size pattern for the
tentacle morphology: with increasing stalk length the width ofthe glands, the stalk base width and the tip width rise as well(Table S3).In the second cluster analysis excluding the stalk length, three-
or four-cluster solutions are reasonable (Table S4). The three-cluster solution is supported by the maximal F value as well as bythe strong decrease of PRE from 0.55 to 0.38. The increase ofη2 supports the solution of three clusters as well. However, fourclusters would be possible, too. The PRE is still very low and η2relatively high. Moreover, the four-cluster solution also containsthe second largest F value.As in the first run of the analysis, a size pattern became visible
(Table S5), indicating that with longer glands the width of theglands as well as the width of the stalk increased.
Cluster Analyses of Roridula gorgonias.The cluster analysis was onlyapplied for Roridula gorgonias, because the variables of R. dentatawere correlated too highly (up to r = 1). As in the amber in-clusions, the assumptions for cluster analyses (metric level ofmeasurement and the absence of outliers) were tested, as well asthe accuracy of data entry, missing values, skewness, and kur-tosis. Seven outliers were excluded from the analysis (Table S1,numbers 167, 168, 170, 190, 197, 202, and 206).Similar to the prior analyses of the amber inclusions, corre-
lations between stalk length and width of the stalk base (r = 0.85),stalk length and gland width (r = 0.80), as well as gland lengthand gland width (r = 0.88) were again quite high. However, theconduction of the cluster analysis was still reasonable, becausethese correlations did not exceed r = 0.9 and only appeared inthree cases. To avoid high correlations and to maintain a goodcomparability to the cluster analyses of the amber inclusions, thevariable gland length was excluded from the first run. As in theamber inclusion, a second cluster analysis was computed, leavingout the stalk length.
The results of the first cluster analysis excluding the glandlength (Table S6) revealed two possible solutions: two and fiveclusters. The solution for two clusters is supported by the PREvalue, because it drops strongly from 0.86 to 0.46 and thusindicates that the following cluster solution does not show con-siderable improvements. However, F-max is not maximal and thechange in η2 from 0.86 to 0.93 is rather small. The maximal F valueis the highest for the 10-cluster solution, but with regards to thecontent not very feasible.Another possible solution would be five clusters, because the
PRE value is low, followed by another decrease, revealing minorimprovements if more clusters are added. In this scenario, F-maxis not maximal but higher than in the three-cluster solution.However, η2 does not change substantially (Table S6).As for the amber inclusions, the mean values of the size classes
show that, with increasing stalk length, the width of the stalk andthe glands increase, too (Table S7).In the second run of the cluster analyses (without the variable
stalk length), several solutions were indicated (Table S8). Themaximal F value speaks for two clusters, but in this case, therelatively high PRE and the very low η2 do not support thisscenario. The three-cluster solution is indicated by the low PRE,which drops considerably from 0.70 to 0.32. However, η2 doesnot change substantially and F-max is not maximal. Anotherpossible solution is a four-cluster scenario, because PRE is lowand decreases from 0.31 to 0.19. The following PRE values donot change strongly. The F value is not maximal but the secondlargest. The four-cluster solution is also supported by η2, whichincreases from 0.79 to 0.86 and does not show any considerablechanges afterward.The mean values of the variables (Table S9) indicate a similar
morphological pattern as in the amber inclusion: longer glandsare broader, and with increasing gland size, the stalk width at thebase and the tip increase as well.
Statistical Evaluation of the Fossil and Roridula gorgonias. In the firstcluster analysis of the amber fossil, the most reasonable solution isthe four-cluster result, which is supported by the clear pattern ofincreasing values in each cluster (Table S3) and by the second runof the analysis without the stalk length, also resulting in fourclusters (Tables S4 and S5).In contrast to the amber inclusions, the most feasible result in
the cluster analyses of Roridula gorgonias is the five-cluster solu-tion. The two-cluster solution does not appear very likely, becauseit does not comprise the wide range of the tentacle length from60 to 3,950 μm.Although the number of size classes was not the same in the leaf
inclusions and R. gorgonias, both showed the same pattern oflonger tentacles possessing broader stalks and glandular heads.Moreover, it is likely that more size classes were present in theEocene roridulid plant but are underrepresented in the fossils. Inparticular, one fossil leaf has a very long tentacle with a rupturedglandular head (Fig. 3A), which was an outlier and hence ex-cluded from the analyses. We see this excluded tentacle as evi-dence for the additional size class 5.If the stalk length was excluded from the cluster analyses, the
results of the amber fossils and R. gorgonias indicated four clusterswith the same size pattern: increasing length of the glandularheads is accompanied by rising width of the glandular head and ofthe tentacle stalk.
Sadowski et al. www.pnas.org/cgi/content/short/1414777111 1 of 7

Table S1. Measured data (values in μm) from the amber inclusions (group 1) and the leaves ofRoridula gorgonias (group 2)
No. Group Stalk length Stalk base width Stalk tip width Gland length Gland width
1 1 30 20 10 20 202 1 60 20 10 50 303 1 90 20 10 60 304 1 40 20 10 30 205 1 50 20 10 30 206 1 40 20 10 40 207 1 60 20 10 40 208 1 180 40 10 60 309 1 1,430 40 10 — —
10 1 80 20 10 50 2011 1 80 30 — 50 3012 1 80 20 10 60 3013 1 80 30 20 70 3014 1 60 20 20 50 3015 1 60 20 10 40 3016 1 220 40 20 80 4017 1 70 20 20 50 3018 1 200 30 10 120 4019 1 70 20 20 60 3020 1 40 20 10 30 2021 1 150 50 20 80 4022 1 90 20 10 60 2023 1 60 20 20 50 3024 1 170 40 20 100 3025 1 60 20 10 40 2026 1 70 20 20 50 2027 1 110 30 20 70 3028 1 350 20 20 110 3029 1 70 20 20 50 2030 1 70 30 10 60 3031 1 200 40 20 80 3032 1 90 20 10 50 3033 1 40 30 20 40 3034 1 90 30 10 50 3035 1 130 20 10 50 3036 1 150 30 20 70 3037 1 300 50 20 120 3038 1 60 20 20 50 3039 1 250 40 20 100 3040 1 40 30 20 40 3041 1 60 30 20 60 3042 1 50 20 10 50 2043 1 130 30 20 110 3044 1 200 40 20 70 3045 1 80 30 20 60 3046 1 100 20 10 60 3047 1 250 20 20 110 3048 1 120 30 20 90 2049 1 140 30 10 90 2050 1 60 30 10 80 3051 1 140 40 30 60 4052 1 90 30 30 70 3053 1 110 20 10 50 3054 1 40 20 20 40 2055 1 120 20 20 60 2056 1 70 20 20 60 3057 1 200 60 20 100 4058 1 30 20 20 40 2059 1 100 30 20 110 3060 1 120 30 30 50 3061 1 50 20 10 30 2062 1 300 50 20 60 40
Sadowski et al. www.pnas.org/cgi/content/short/1414777111 2 of 7

Table S1. Cont.
No. Group Stalk length Stalk base width Stalk tip width Gland length Gland width
63 1 300 50 20 60 3064 1 50 20 10 40 2065 1 120 40 20 50 3066 1 100 30 10 60 3067 1 190 30 20 80 3068 1 40 20 10 30 2069 1 100 30 20 50 2070 1 140 40 10 70 3071 1 110 30 10 30 3072 1 250 40 10 60 3073 1 110 30 10 120 3074 1 110 30 10 70 3075 1 90 30 20 30 2076 1 40 20 10 30 2077 1 50 20 10 30 2078 1 30 20 20 30 2079 1 200 30 10 70 3080 1 170 40 20 80 3081 1 50 20 10 30 2082 1 40 20 10 30 2083 1 40 20 20 20 2084 1 40 10 10 20 1085 1 40 10 10 30 2086 1 150 40 20 80 3087 1 50 30 10 30 3088 1 100 40 20 50 2089 1 280 30 10 60 3090 1 270 40 10 50 3091 1 20 20 20 20 2092 1 50 20 10 40 2093 1 30 20 20 30 3094 1 80 20 10 60 2095 1 60 20 10 40 3096 1 40 20 10 30 2097 1 140 30 20 50 3098 1 150 10 10 50 2099 1 50 50 10 50 20100 1 160 20 20 60 30101 1 120 40 10 50 20102 1 30 10 10 40 20103 1 110 20 20 50 20104 2 2,200 120 50 260 120105 2 1,600 100 40 200 120106 2 3,000 200 40 220 120107 2 2,400 120 60 260 120108 2 3,500 160 60 260 120109 2 1,400 100 40 200 120110 2 2,000 100 60 240 120111 2 2,540 120 40 260 140112 2 2,500 140 60 240 140113 2 2,440 140 60 220 120114 2 3,360 200 60 260 120115 2 2,000 140 60 240 120116 2 3,640 180 60 260 120117 2 2,240 160 60 260 120118 2 2,300 180 60 240 140119 2 1,720 120 60 220 120120 2 2,980 140 60 240 120121 2 120 50 30 220 140122 2 200 30 30 100 50123 2 150 30 30 70 50124 2 230 50 30 80 50
Sadowski et al. www.pnas.org/cgi/content/short/1414777111 3 of 7

Table S1. Cont.
No. Group Stalk length Stalk base width Stalk tip width Gland length Gland width
125 2 200 40 30 100 60126 2 230 40 30 80 50127 2 450 60 30 90 60128 2 140 40 30 110 70129 2 220 50 40 90 40130 2 170 40 30 90 60131 2 60 20 15 80 50132 2 150 40 30 40 20133 2 310 70 40 70 50134 2 150 30 40 100 60135 2 180 50 30 70 50136 2 150 30 30 80 50137 2 220 40 30 70 50138 2 180 40 30 80 50139 2 110 20 20 80 50140 2 180 30 30 60 30141 2 200 40 30 70 50142 2 100 30 20 70 40143 2 200 30 30 60 30144 2 370 50 30 90 50145 2 120 30 20 120 70146 2 140 30 30 90 40147 2 180 40 30 90 50148 2 700 90 40 90 60149 2 150 30 20 110 110150 2 190 40 30 60 50151 2 200 40 30 90 50152 2 120 30 30 90 50153 2 700 70 50 60 40154 2 300 50 40 130 100155 2 180 30 20 100 60156 2 170 40 30 80 40157 2 1,050 90 50 70 50158 2 440 70 30 120 120159 2 230 40 30 110 70160 2 570 100 40 80 60161 2 200 30 30 120 70162 2 250 50 30 70 50163 2 170 50 50 100 60164 2 180 50 40 100 70165 2 300 70 60 90 80166 2 550 110 80 150 110167 2 160 50 40 220 190168 2 490 120 100 80 60169 2 120 40 40 200 160170 2 600 150 100 100 70171 2 240 70 60 200 170172 2 550 130 90 120 90173 2 280 80 80 230 180174 2 550 140 90 130 110175 2 150 50 40 210 150176 2 450 100 80 90 60177 2 150 50 40 180 140178 2 150 50 40 80 60179 2 600 140 80 100 80180 2 250 70 50 200 150181 2 200 60 50 120 90182 2 140 40 30 100 90183 2 130 50 30 70 50184 2 100 40 30 60 50185 2 120 30 20 80 60186 2 150 50 40 70 40
Sadowski et al. www.pnas.org/cgi/content/short/1414777111 4 of 7
Table S1. Cont.
No. Group Stalk length Stalk base width Stalk tip width Gland length Gland width
187 2 440 100 30 100 60188 2 170 50 40 90 60189 2 230 80 40 70 60190 2 2,300 240 50 200 90191 2 120 30 20 40 30192 2 630 120 30 110 70193 2 180 50 30 70 50194 2 820 150 40 130 90195 2 200 50 40 70 50196 2 120 30 20 50 40197 2 3,950 250 70 250 130198 2 220 50 40 70 50199 2 800 140 50 130 70200 2 230 60 40 80 60201 2 420 90 40 100 60202 2 2,650 250 30 200 90203 2 500 90 50 120 80204 2 1,090 150 50 150 80205 2 100 40 20 50 30206 2 3,050 270 60 290 120
Table S2. Results of the first cluster analysis of the leafinclusions, excluding the gland length, with the three statisticaltest criteria, η2, F-max, and PRE
Cluster η2 F-max PRE
1 0.00 −99 −992 0.69 215.20 0.693 0.84 243.19 0.474 0.91 321.93 0.465 0.94 342.95 0.286 0.95 372.49 0.267 0.96 351.62 0.128 0.96 350.62 0.149 0.97 340.66 0.1110 0.97 326.11 0.08
Indicative values are highlighted in bold.
Table S3. Mean values (in μm) of the four-cluster solution forthe leaf inclusions
Variables Size class 1 Size class 2 Size class 3 Size class 4
Stalk length 49.07 101.79 168.33 265.00Stalk base width 20.70 27.14 35.56 38.75Stalk tip width 13.95 15.71 17.22 16.25Gland width 23.49 26.43 31.11 31.25No. of tentacles 43 28 18 8
Length of the glandular head is excluded.
Sadowski et al. www.pnas.org/cgi/content/short/1414777111 5 of 7
Table S4. Results of the second cluster analysis of the leafinclusions, excluding the stalk length, with the three statisticaltest criteria, η2, F-max, and PRE
Cluster η2 F-max PRE
1 0.00 −99 −992 0.55 116.81 0.553 0.72 121.61 0.384 0.79 117.90 0.255 0.82 102.09 0.126 0.83 90.92 0.097 0.85 84.27 0.098 0.86 79.12 0.089 0.87 76.20 0.0910 0.88 73.81 0.08
Indicative values are highlighted in bold.
Table S5. Mean values (in μm) of the four-cluster solution forthe leaf inclusions
Variables Size class 1 Size class 2 Size class 3 Size class 4
Gland length 32.00 53.16 71.58 105.00Gland width 21.33 26.84 31.58 30.00Stalk base width 20.00 24.72 26.84 36.00Stalk tip width 12.33 15.53 17.89 18.00No. of tentacles 30 38 19 10
Length of the stalk is excluded.
Table S6. Results of the first cluster analysis of Roridulagorgonias, excluding the gland length, with the three statisticaltest criteria, η2, F-max, and PRE
Cluster η2 F-max PRE
1 0.00 −99 −992 0.86 582.96 0.863 0.93 576.90 0.464 0.96 781.77 0.495 0.98 979.70 0.406 0.99 1,090.73 0.287 0.99 1,202.36 0.258 0.99 1,354.45 0.259 0.99 1,458.88 0.2010 0.99 1,604.54 0.20
Indicative values are highlighted in bold.
Table S7. Mean values (in μm) of the five-cluster solution for Roridula gorgonias
Variables Size class 1 Size class 2 Size class 3 Size class 4 Size class 5
Stalk length 181.15 628.33 1,573.33 2,291.11 3,296Stalk base width 43.61 107.78 106.67 135.56 176Stalk tip width 33.52 52.78 46.67 56.67 56Gland width 66.23 75 120 126.67 120No. of tentacles 61 18 3 9 5
Length of the glandular head is excluded.
Sadowski et al. www.pnas.org/cgi/content/short/1414777111 6 of 7

Table S8. Results of the second cluster analysis of Roridulagorgonias, excluding the stalk length, with the three statisticaltest criteria, η2, F-max, and PRE
Cluster η2 F-max PRE
1 0.00 −99 −992 0.70 214.85 0.703 0.79 178.59 0.324 0.86 184.43 0.315 0.88 174.15 0.196 0.91 171.12 0.187 0.92 168.48 0.158 0.93 165.33 0.139 0.94 157.11 0.0810 0.94 154.96 0.10
Indicative values are highlighted in bold.
Table S9. Mean values (in μm) of the four-cluster solution forRoridula gorgonias, excluding the stalk length
Variables Size class 1 Size class 2 Size class 3 Size class 4
Gland length 80.37 114.44 204.44 245.33Gland width 52.41 81.67 147.78 124.00Stalk base width 42.41 105.56 67.78 148.00Stalk tip width 31.20 53.89 46.67 56.67No. of tentacles 54 18 9 15
Sadowski et al. www.pnas.org/cgi/content/short/1414777111 7 of 7
Related Documents











