European Journal of Pharmacology, 210 (1992) 137-147 137 ~ 1992 Elsevier Science Publishers B.V. All rights reserved 0014-2999/92/$05.00 EJP 52218 Cardiovascular responses elicited by intrathecal kinins in the conscious rat Paulo Lopes and R~jean Couture Groupe de Recherche sur le Systbrne Nert~eux Autonorne, D~Xpartement de Physiologie, Facultd de Mddecine, Unit'ersitd de MontrtXal, C.P. 6128, Succursale A, Montr¢;al, Qudbec, Canada H3C 3.17 Received 6 June 1991, revised MS received 3 September 1991, accepted 15 October 1991 In the conscious, unrestrained rat, intrathccal (i.t.) injection of 0.81 pmol-81 nmol bradykinin (BK), kallidin (KD) and T-kinin at the T-9 spinal cord level produced transient (< 10 mini increases in mean arterial pressure (MAP) and longer lasting decreases in heart rate (HR). These effects were dose-dependent and similar with respect to intensity and time course for the three kinins. The des-Arg'-BK fragment, a selective agonist for Bj receptors, was active only at 81 nmol. The pressor response induced by BK was enhanced by propranolol and by transection of the cervical spinal cord but was converted to a vasodcpressor effect by prazosin. The bradycardia was converted to tachycardia by prazosin, atropine, pentolinium, capsaicin and in spinal transccted rats. However, the cardiovascular responses to BK remained unaffected by diphenhydraminc plus cimetidine, morphine, indomethacin, adrenal medullectomy, i.t. idazoxan and after bulbospinal noradrenalinc deafferentation with 6-hy- droxydopaminc. These results suggest that the increase in MAP induced by i.t. BK is mediated by the sympathoadrcnal system while the decrease in HR is ascribable to a vagal reflex involving sensory C-fibers and a spinobulbar pathway. This pharmacological evidence therefore supports a role for kinins in cardiovascular regulation in the spinal cord. Kinins; Spinal cord; Cardiovascular system; Catecholamines; Vagal reflex; C-fibers 1. Introduction Kinins are biologically active peptides generated in plasma (bradykinin, BK) and peripheral tissues (kalli- din, KD) after cleavage of large protein precursors (kininogens) under the action of specific enzymes termed kallikreins. BK and KD are produced during tissue dammage and are involved in inflammatory reac- tions, causing pain by activating peripheral sensory endings, vasodilation and increased vascular permeabil- ity (Regoli and Barab6, 1980; Regoli, 1987). In addi- tion, rat plasma contains another kinin named T-kinin (Ile-Ser-BK) (Okamoto and Greenbaum, 1983a, b) which is apparently a mediator of the inflammatory response in the rat (Barlas et al., 1985; Adam et al., 1989). Several reports suggests that kinins may also be involved in central cardiovascular regulation (Diz, 1985). The presence of kinins, kininogen, kallikreins and kininase activity has been shown in the brain, the spinal cord and the cerebrospinal fluid of humans Correspondence to: R. Couture, Groupe de Recherche sur le Systbme Nerveux Autonome, D~partement de Physiologie, Facult~ de M6de- cine, Universit6 de Montreal, C.P. 6128, Succursale A, Montreal, Quebec, Canada H3C 3J7. Tel. 1.514.343 7060, fax 1.514.343 2111. (Scicli et al., 1984) and various other species (Perry and Snyder, 1984; Thomas et al., 1984; Kariya et al., 1985; Hermann et al., 1986; Madeddu et al., 1990; Yoshida and Nosaka, 1990). Autoradiographic studies (Steranka et al., 1988) and binding studies (Fujiwara et al., 1989; Sharif and Whiting, 1991) have revealed the presence of [3H]BK receptor binding sites (identified as B z re- ceptors) in the guinea pig spinal cord and other central areas of cardiovascular regulation. The intracere- broventricular (i.c.v.) injection of BK and/or KD pro- duces changes (decreases and/or increases) of blood pressure in cats, dogs, rabbits and rats (see Diz, 1985 for a review). The pressor effect is prevalent in the awake rat, indicating that anesthesia affects the blood pressure response to centrally administered BK (Lam- bert and Lang, 1970; Corr~a and Graeff, 1974; Hoff- man and Schmid, 1978; Buccafusco and Serra, 1985; Lindsey et al., 1989). Concomitant increases of blood pressure and levels of kinins in the cerebrospinal fluid have been reported in dogs (Thomas et al., 1984, 1987) and rats (Thomas and Hiley, 1988). In spite of these observations, no studies have attempted to determine whether the kinins can affect the cardiovascular system through an action in the spinal cord. The present study was therefore undertaken to de- termine the intrathecal (i.t.) action of BK, KD and T-kinin on mean arterial pressure (MAP) and heart

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

European Journal of Pharmacology, 210 (1992) 137-147 137 ~ 1992 Elsevier Science Publishers B.V. All rights reserved 0014-2999/92/$05.00
EJP 52218
Cardiovascular responses elicited by intrathecal kinins in the conscious rat
Pau lo L o p e s and R~ jean C o u t u r e
Groupe de Recherche sur le Systbrne Nert~eux Autonorne, D~Xpartement de Physiologie, Facultd de Mddecine, Unit'ersitd de MontrtXal, C.P. 6128, Succursale A, Montr¢;al, Qudbec, Canada H3C 3.17
Received 6 June 1991, revised MS received 3 September 1991, accepted 15 October 1991
In the conscious, unrestrained rat, intrathccal (i.t.) injection of 0.81 pmol-81 nmol bradykinin (BK), kallidin (KD) and T-kinin at the T-9 spinal cord level produced transient (< 10 mini increases in mean arterial pressure (MAP) and longer lasting decreases in heart rate (HR). These effects were dose-dependent and similar with respect to intensity and time course for the three kinins. The des-Arg'-BK fragment, a selective agonist for Bj receptors, was active only at 81 nmol. The pressor response induced by BK was enhanced by propranolol and by transection of the cervical spinal cord but was converted to a vasodcpressor effect by prazosin. The bradycardia was converted to tachycardia by prazosin, atropine, pentolinium, capsaicin and in spinal transccted rats. However, the cardiovascular responses to BK remained unaffected by diphenhydraminc plus cimetidine, morphine, indomethacin, adrenal medullectomy, i.t. idazoxan and after bulbospinal noradrenalinc deafferentation with 6-hy- droxydopaminc. These results suggest that the increase in MAP induced by i.t. BK is mediated by the sympathoadrcnal system while the decrease in HR is ascribable to a vagal reflex involving sensory C-fibers and a spinobulbar pathway. This pharmacological evidence therefore supports a role for kinins in cardiovascular regulation in the spinal cord.
Kinins; Spinal cord; Cardiovascular system; Catecholamines; Vagal reflex; C-fibers
1. Introduction
Kinins are biologically active peptides generated in plasma (bradykinin, BK) and peripheral tissues (kalli- din, KD) after cleavage of large protein precursors (kininogens) under the action of specific enzymes termed kallikreins. BK and KD are produced during tissue dammage and are involved in inflammatory reac- tions, causing pain by activating peripheral sensory endings, vasodilation and increased vascular permeabil- ity (Regoli and Barab6, 1980; Regoli, 1987). In addi- tion, rat plasma contains another kinin named T-kinin (Ile-Ser-BK) (Okamoto and Greenbaum, 1983a, b) which is apparently a mediator of the inflammatory response in the rat (Barlas et al., 1985; Adam et al., 1989).
Several reports suggests that kinins may also be involved in central cardiovascular regulation (Diz, 1985). The presence of kinins, kininogen, kallikreins and kininase activity has been shown in the brain, the spinal cord and the cerebrospinal fluid of humans
Correspondence to: R. Couture, Groupe de Recherche sur le Systbme Nerveux Autonome, D~partement de Physiologie, Facult~ de M6de- cine, Universit6 de Montreal, C.P. 6128, Succursale A, Montreal, Quebec, Canada H3C 3J7. Tel. 1.514.343 7060, fax 1.514.343 2111.
(Scicli et al., 1984) and various other species (Perry and Snyder, 1984; Thomas et al., 1984; Kariya et al., 1985; Hermann et al., 1986; Madeddu et al., 1990; Yoshida and Nosaka, 1990). Autoradiographic studies (Steranka et al., 1988) and binding studies (Fujiwara et al., 1989; Sharif and Whiting, 1991) have revealed the presence of [3H]BK receptor binding sites (identified as B z re- ceptors) in the guinea pig spinal cord and other central areas of cardiovascular regulation. The intracere- broventricular (i.c.v.) injection of BK a n d / o r KD pro- duces changes (decreases a n d / o r increases) of blood pressure in cats, dogs, rabbits and rats (see Diz, 1985 for a review). The pressor effect is prevalent in the awake rat, indicating that anesthesia affects the blood pressure response to centrally administered BK (Lam- bert and Lang, 1970; Corr~a and Graeff, 1974; Hoff- man and Schmid, 1978; Buccafusco and Serra, 1985; Lindsey et al., 1989). Concomitant increases of blood pressure and levels of kinins in the cerebrospinal fluid have been reported in dogs (Thomas et al., 1984, 1987) and rats (Thomas and Hiley, 1988). In spite of these observations, no studies have a t tempted to determine whether the kinins can affect the cardiovascular system through an action in the spinal cord.
The present study was therefore undertaken to de- termine the intrathecal (i.t.) action of BK, KD and T-kinin on mean arterial pressure (MAP) and heart

138
rate (HR). The cardiovascular response to des-Arg '~- BK, a metabolite of BK and a selective stimulant of the BI receptor, was also measured. The second objective was to investigate the spinal and peripheral mecha- nisms underlying the cardiovascular responses to BK. Intrathecal BK may affect the cardiovascular system through three possible mechanisms: (1) by direct acti- vation of preganglionic sympathetic fibres in the inter- mediolateral cell column, (2) indirectly, by facilitating the release of substance P or other transmitters from central sensory nerve terminals, as suggested by the presence of BK binding sites in the substantia gelati- nosa (Steranka et al., 1988) and by the behavioral nociceptives responses produced by i.t. BK in the rat (Laneuville and Couture, 1987; Laneuville et al., 1989), (3) indirectly, by activating bulbospinal catecholaminer- gic fibers which appear to play a role in central cardio- vascular regulation (Coote, 1988). A previous study from our laboratory had provided evidence that BK may inhibit spinal nociceptive sensory transmission and produce analgesia by activating bulbospinal noradren- ergic fibers (LaneuviIle et al., 1989). Thus, a possible interaction with sensory C-fibers and bulbospinal cate- cholaminergic fibers was examined. We also sought to examine the role played by the sympathoadrenal sys- tem, the vagus nerve, prostaglandins and other endoge- nous mediators in the spinal action of BK on the rat cardiovascular system. In order to exclude any influ- ence of anesthesia, all experiments were performed in conscious unrestrained rats. Some of these results have been presented as a preliminary communication (Lopes and Couture, 1991).
2. Materials and methods
2.1. Implantation of catheters and measurement of car- diot,ascular parameters
Male Wistar rats (Charles River, St-Constant, Qu6bec, Canada) weighing 200-250 g were implanted with three catheters under sodium pentobarbital anes- thesia (65 m g / k g intraperitoneally (i.p.)), as previously described (Hass6ssian and Couture, 1989; Hassdssian et al., 1990). Briefly, intraarterial (i.a.) and intravenous (i.v.) polyethylene tubes (Intramedic, Clay Adams, N J, U.S.A.; PE-50) were inserted into the medial tail artery and the right jugular vein while an i.t. catheter (PE-10) reached the 9th thoracic (T-9) segment. The experi- ments were performed in awake unrestrained rats, at least 24 h after catheter implantations, in a quiet room, with the cage covered by an opaque cloth to avoid visual cues to the rats. Only rats with no apparent motor deficit and showing normal behavior were used. The correct positioning of the i.t. catheter was verified by post-mortem examination at the end of each experi-
ment. The arterial blood pressure was monitored via the i.a. catheter using a Statham pressure transducer (P231D) and the heart rate (HR) was measured with a cardiac tachometer (model 7P4) triggered by the arte- rial pressure pulses. The arterial blood pressure and the H R were recorded with a Grass T M polygraph model 79 (Quincy, MA, U.S.A.). Each experiment was started with an i.t. injection of 30 ~1 of cerebrospinal fluid (CSF; for composition see Hassdsian and Couture, 1989, void volume of the i.t. catheter was 20 ~1) as a vehicle control. The arterial blood pressure and H R were then monitored for 30 min, after which the first test drug was injected i.t. in a volume of 10 #1. The drug and the CSF (20 ~zl) flush were given within a total period of 60 s. Changes in the MAP and H R represent the difference between recordings obtained 30 s before the start of injection and the value at the designed time after injection.
2.2. Adrenal medtdlectomy and spinal chemical sympa- thectomy
Adrenal medullectomy was performed 72 h prior to experiments in rats under pentobarbital anesthesia. The two adrenal glands were exposed through dorsal incisions and the medullas were extruded with gentle forceps pressure on the adrenal after slitting the cap- sule. The skin was sutured and rats were allowed to recover from anesthesia. Two days later, the rats were anesthetized again for catheter implantation as de- scribed above. The neurotoxin, 6-hydroxydopamine hy- drochloride (6-OHDA) (prepared in CSF containing 1% ascorbic acid), was administered in 50-~g (free base) doses at the T-2 and T-9 spinal cord levels through an i.t. catheter in pentobarbital-anesthetized rats. The i.t. catheter was removed thereafter, the surgical wound was sutured and the rats were returned to their cages for one week after which the chronic i.t. (T-9 level) and intravascular catheters were implanted. The noradrenaline content was reduced by around 60% in the spinal cord while serotonin, dopamine, adrenaline and their main metabolites were unaffected in rats that had received a single i.t. injection of 20/_Lg 6 -OHDA one week earlier (Laneuville ct al., 1989).
2.3. capsaicin treatment
The rats were treated with capsaicin according to the following protocol: 125 m g / k g given subcuta- neously (s.c.) over 3 days, with 25, 40 and 60 m g / k g on the first, second and third day, respectively. Capsaicin was made up in 10% ethanol and 10% Tween 80 in saline. Experiments were conducted four days after the last injection of capsaicin. When administered in adult rats, capsaicin produces a prolonged inactivation of

139
primary sensory C-fibers (Maggi and Meli, 1988). Rats pretreated with capsaicin failed to react to the applica- tion of a drop of solution containing 25 mg/ml cap- saicin in one eye. Similar treatment of control animals caused discomfort reactions.
2.4. Spinal cord transection
Rats assigned to the spinal group had surgically implanted chronic i.t. and intravascular catheters. The spinal cord was transected at the C-6-C-7 vertebral level under halothane anesthesia (3-5% volume in a gas mixture of 95% O : - 5 % CO2). The dorsal cervical muscles were exposed; the intravertebral disc was cut with micro-scissors, and the spinal cord was transected while the i.t. cannula remained intact. Bleeding was minimal and usually stopped within 30 s with a ball of cotton wool inserted between the rostral and caudal segments of the transected spinal cord. The wound was closed and the animals recovered from anesthesia on inhaling a gas mixture containing 95% 0 2 - 5 % CO 2. The experiments were started 45 min after surgery in awake animals.
2.5. Experimental protocols
In the first series of experiments, the time course effects of several doses of BK, KD, T-kinin and des- ArgLBK on MAP and HR was measured after i.t. injection at the T-9 spinal cord level. The experiments were started by the injection of CSF, followed 20 min later by increasing doses of peptide. A period of 30-45 min was allowed between doses so that the MAP and HR could return to pre-injection values. Only one peptide was given to each rat.
The second series of experiments was designed to study the mechanism underlying the cardiovascular ef- fect of BK. The roles played by the sympathetic and parasympathetic components of the autonomic nervous system were examined using antagonists of al-adreno- ceptors (prazosin) and peripheral muscarinic acety[- choline receptors (methylnitrate atropine). The re- sponses to BK were also evaluated in rats following bilateral adrenal medullectomy. The participation of prostaglandins and histamine was determined with a cyclooxygenase inhibitor (indomethacin) and antihis- taminics for H~ (diphenhydramine) and H 2 (cimeti- dine) receptors. A ganglionic nicotinic receptor blocker (pentolinium) was administered to rats to determine whether the cardiovascular responses to BK were me- diated through a reflex mechanism. The rats received systemic capsaicin and i.t. 6-OHDA to assess the con- tribution of primary sensory C-fibers and bulbospinal catecholaminergic fibers on the effect of BK. The central contribution of catecholamines was also as- sessed with i.t. idazoxan, an a2-adrenoceptor antago-
nist known to Mock the antinociceptive response in- duced by i.t. BK (Laneuville et al., 1989). A group of rats was treated with morphine (opioid agonist) to test for a nociceptive component of the cardiovascular re- sponse. The potential contribution of supraspinal cen- ters was studied in spinal rats.
2.6. Peptides and other compounds
The peptides BK, KD, T-kinin and des-ArgKBK were purchased from Hfikabel Scientific Ltd. (Longueuil, Canada). Indomethacin, methylnitrate at- ropine, cimetidine, nicotine, pentolinium tartratc salt, diphenhydramine hydrochloride, propranolol hydro- chloride, capsaicin (8-methyl-N-vanillyl-6-nonenamide), heparin sodium salt (porcine, grade II) were all pur- chased from Sigma (St. I~uis, MO, U.S.A.), idazoxan was obtained from Reckitt and Colman (UK), 6-OHDA hydrochloride from Labkemi AB (Stockholm, Sweden), prazosin hydrochloride from Pfizer (Kirkland, Canada), L-ascorbic acid from Baker (Phillipsburg, N J, USA), morphine sulphate from BDH Pharmaceuticals (Toronto, Canada), sodium pentobarbital (somnotol) from MTC Pharmaceuticals (Missisauga, Canada), halothane (fluothane) from Ayerst (Montrdal, Canada) and Tween 80 from Fisher (Montrdal, Canada). The peptides were prepared in CSF as 1 m g /m l stock solutions, divided into aliquots of 100 /xl each, and stored at - 2 0 ° C until used. Indomethacin was pre- pared in trizma base (0.2 M, Sigma)just before utiliza- tion, and the other drugs were prepared in saline. Doses of peptides are given in mo[ of the salt.
2. 7. Statistical analysis of data
The results are expressed as means _+ S.E.M. Statis- tical differences were evaluated with Student's t-test for paired or unpaired samples as required. A one-way analysis of variance (ANOVA) with a post-hoc Dun- nctt test was used for multiple comparisons with one control group. The time course of the effects was analysed with a two-way ANOVA in conjunction with Bonferroni confidence intervals. Only probability val- ues (P) smaller than 0.05 were considered to be statisti- cally significant.
3. Results
3. l. Effects of i.t. kinins on MAP and HR
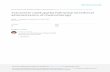
An original tracing showing the cardiovascular effect of 81 pmol BK given i.t. at T-9 to a freely moving rat is presented in fig. 1. A transient increase of blood pres- sure and decrease of HR were seen immediately after the peptide injection.
140
LU 200 l C jS :~ oDI 150
=~g 100 a_ 50
~_~ 500 ( <_c E 400
< ~ 300 w ~ I 200
CSF BK (81 pmol) 1 mm
Fig. 1. Typical tracings of blood pressure and HR responses to i.t. injection of 81 pmol BK at the T-9 spinal thoracic level in a
conscious rat. Note the lack of response to i.t. injection of CSF.
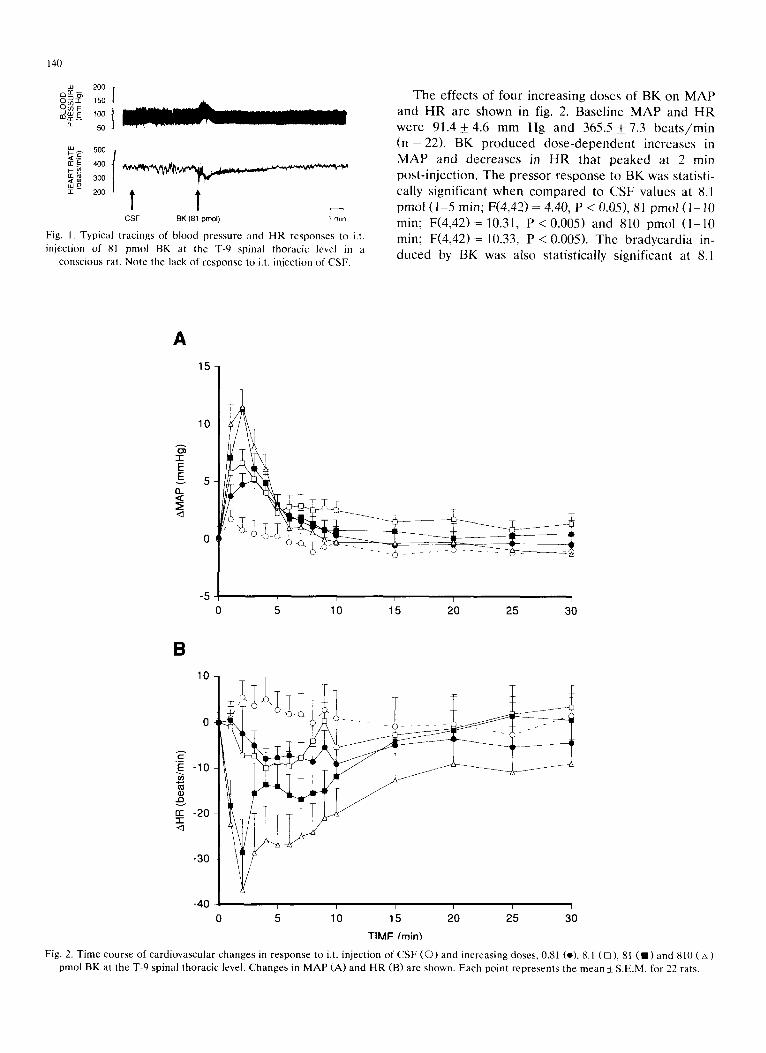
The effects of four increasing doses of BK on MAP and HR are shown in fig. 2. Baseline MA P and H R were 91.4_+4.6 mm Hg and 365.5_+ 7.3 b e a t s / m i n (n = 22). BK produced dose -dependen t increases in M A P and decreases in HR that peaked at 2 rain
post- injection, The pressor response to BK was statisti- cally significant when compared to CSF values at 8.1
pmol (1 -5 rain; F(4,42) - 4.40, P < 0.05), 81 pmol (1 -10 rain; F (4 ,42)= 10.31, P < 0.005) and 810 pmol (1 -10 rain; F (4 ,42)= 10.33, P < 0.005). The bradycardia in- duced by BK was also statistically significant at 8.1
A 15
10
0
-5 0
"I- E E 5 0_ <
<~
t i i I i i
5 10 15 20 25 30
B 10
E -10
n- -20 -'r-
-30
-40
0
Fig. 2. Time course of cardiovascular changes pmol BK at the T-9 spinal thoracic level. Changes in MAP (A) and HR (B) are shown. Each point represents the mean + S.E.M. for 22 rats.
i i I i i 5 10 15 20 25 30
TIMI= ~'min~
m response to i.t. injection of CSF (O) and increasing doses, 0.81 Co), 8.1 ( [] ), 81 ( • ) and 810 ( ~ )
pmol ( 1 - 5 min; F(4,42) = 4.40, P < 0.05), 81 pmol ( 1 - 1 0 min; F ( 4 , 4 2 ) = 10.8, P < 0.005) and 810 pmol ( 1 - 1 0 rain; F(4,42) = 14.61, p < 0.005). The p resso r r e sponse r e t u r n e d to base l ine within 10 min while the b radyca r - dia las ted longer and had not comple t e ly r e t u r n e d to p re - in jec t ion values 30 min af te r inject ion of 810 pmol BK. The ca rd iovascu la r r e sponses e l ic i ted by the i.t. in ject ion of 81 pmol BK dur ing and 30 min af te r the assessment of the d o s e - r e s p o n s e curve were not statis- t ically d i f fe ren t ( A M A P : F(1,28) = 2.74, P > 0.05; A H R : F(1,28) = 0.48, P > 0.05). Hence , no tachyphy- laxis was obse rved with BK u n d e r these expe r ime n t a l
141
condi t ions . T h e i.t. in ject ion of C S F was wi thout app re - c iable effect on M A P and HR.
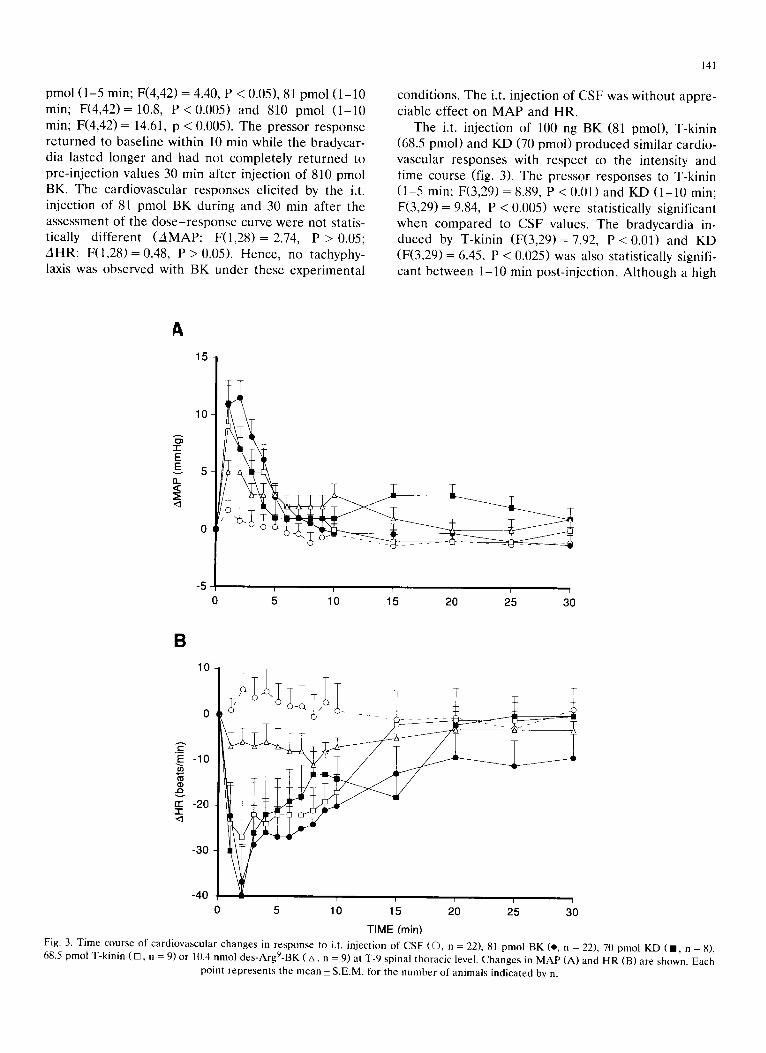
The i.t. in ject ion of 100 ng BK (81 pmol) , T-kinin (68.5 pmol ) and K D (70 pmol ) p r o d u c e d s imilar card io- vascu la r r e sponses with r e spec t co the in tensi ty and t ime course (fig. 3). The p resso r r esponses to T-kinin ( 1 - 5 min; F(3,29) = 8.89, P < 0.01) and KD ( 1 - 1 0 min; F(3,29) = 9.84, P < 0.005) were s ta t is t ical ly s ignif icant when c o m p a r e d to CSF values. The b r adyca rd i a in- duced by T-kinin ( F ( 3 , 2 9 ) = 7.92, P < 0 . 0 1 ) and KD (F(3,29) = 6.45, P < 0.025) was also s tat is t ical ly signifi- cant be tween 1-10 min pos t - in jec t ion . A l though a high
A 15
10
-1- E
g 5 EL
- 5 i i T i i i
0 5 10 15 20 25 30
B 10
E -10
n- -20 "1- <1
-30
-40 - - p
0 5 i p i i i
10 15 20 25 30
TIME (mini
Fig. 3. Time course of cardiovascular changes in response to i.t. injection of CSF (©, n = 22), 81 pmol BK (e, n = 22), 70 pmol KD (11, n = 8), 68.5 pmol T-kinin ( D, n = 9) or 10.4 nmol des-Argg-BK ( A, n = 9) at T-9 spinal thoracic level. Changes in MAP (A) and HR (B) are shown. Each
point represents the mean + S.E.M. for the number of animals indicated by n.
142
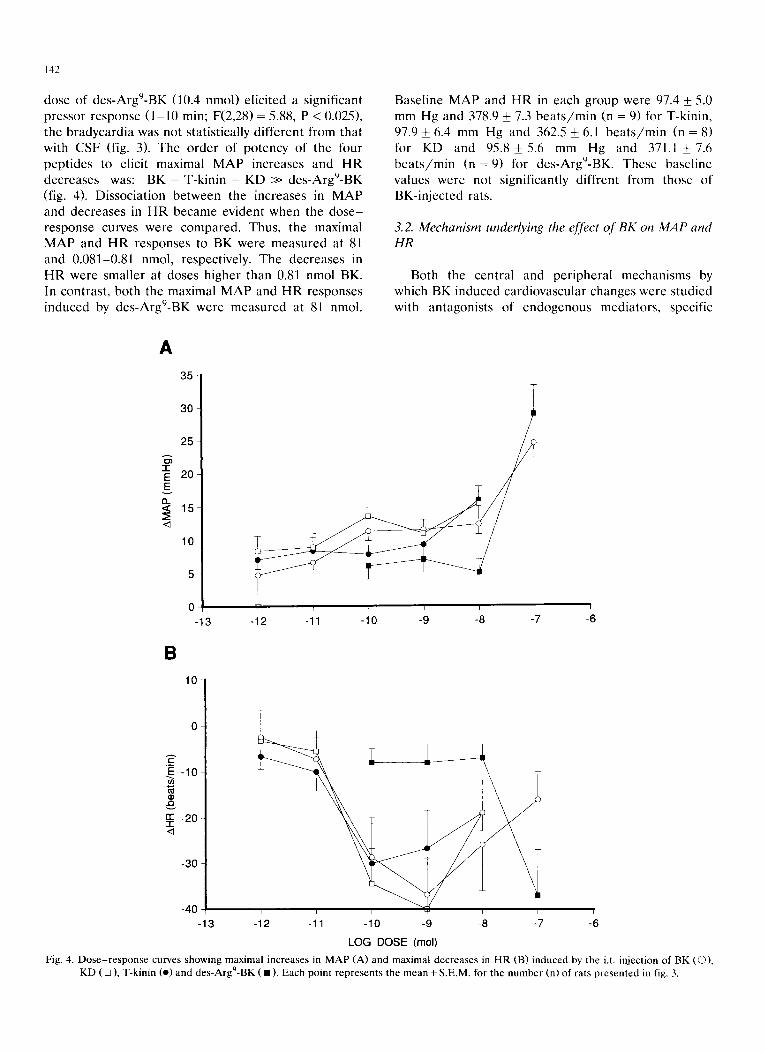
dose of des -Argg-BK (10.4 nmol) e l ic i ted a s ignif icant p ressor response ( 1 - 1 0 min; F(2,28) = 5.88, P < 0.025), the b r a d y c a r d i a was not s tat is t ical ly d i f fe ren t from that with C S F (fig. 3). The o r d e r of po tency of the four pep t ide s to elicit maximal M A P increases and H R dec reases was: BK = T-kin in = K D >> des -Argg-BK (fig. 4). Dissoc ia t ion be tween the increases in M A P and dec reases in H R b e c a m e evident when the d o s e - r e sponse curves were c o m p a r e d . Thus, the maximal M A P and H R responses to BK were m e a s u r e d at 81 and 0.081-0.81 nmol, respect ively. The dec reases in H R were smal le r at doses h igher than 0.81 nmol BK. In contras t , bo th the maximal M A P and H R responses induced by des -Argg-BK were m e a s u r e d at 81 nmol.
Base l ine M A P and H R in each group were 97.4 +- 5.0 mm Hg and 378.9 _+ 7.3 b e a t s / m i n (n = 9) for T-kinin , 97.9 _+ 6.4 mm Hg and 362.5 +- 6.1 b e a t s / r a i n (n = 8) for KD and 95.8_+5.6 mm Hg and 371.1 +_7.6 b e a t s / r a i n (n = 9) for d e s - A r g < B K . These base l ine values were not s ignif icantly di f f rent from those of BK- in jec ted rats.
3.2. Mechanism underlying the effect of BK on MAP and HR
Both the centra l and pe r iphe ra l mechan i sms by which BK induced ca rd iovascu la r changes were s tud ied with an tagonis t s of e n d o g e n o u s media tors , specific
A
1"
E E
r l ,<
<1
35
30
25
20
15
10
5
0 -13
i
I r i i i i I
-12 -11 -10 -9 -8 -7 -6
B 10
0
. ~ - 1 0 -
-20 - I
-30 -
-40
-13 -12 -11 -10 -9 -8 -7 -6
LOG DOSE (mol)
Fig. 4. Dose-response curves showing maximal increases in MAP (A) and maximal decreases in HR (B) induced by the i.t. injection of BK (O), KD ( [] ), T-kinin 9 (o) and des-Arg -BK ( • ) . Each point represents the mean + S.E.M. for the number (n) of rats presented in fig. 3.
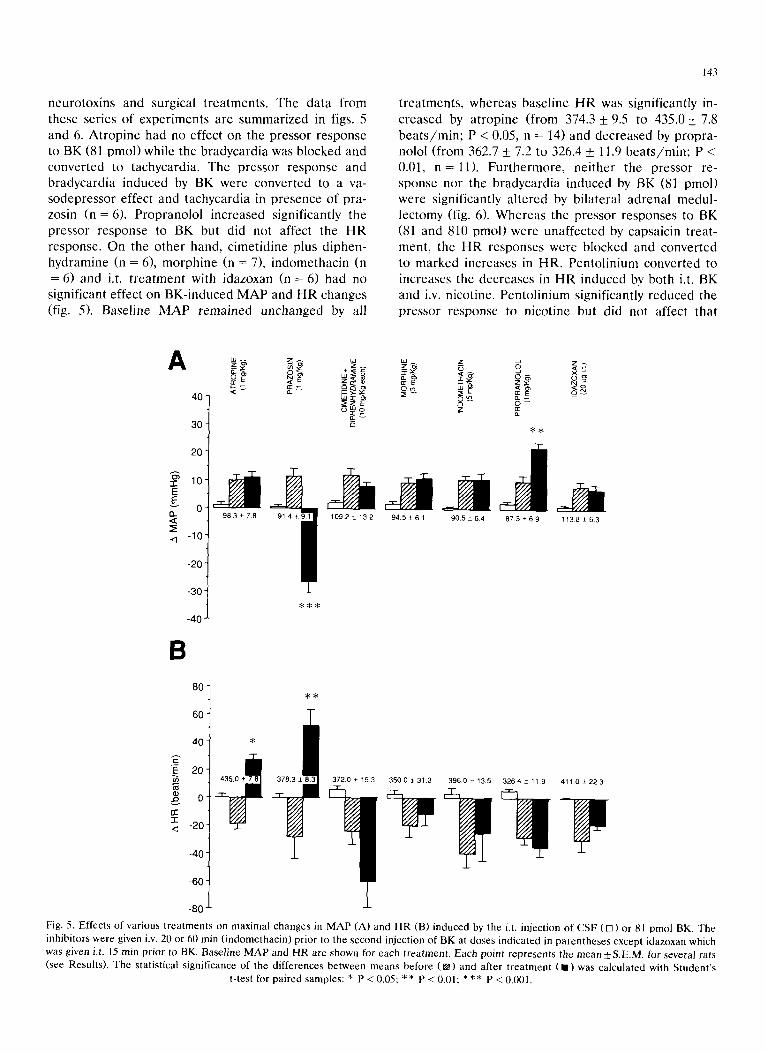
neuro tox ins and surgical t r ea tmen t s . The da ta f rom these ser ies of expe r imen t s a re s u m m a r i z e d in figs. 5 and 6. A t r o p i n e had no effect on the p re s so r response to BK (81 pmol ) while the b r adyca rd i a was b locked and conver t ed to tachycard ia . The p resso r r e sponse and b radyca rd i a induced by BK were conver t ed to a va- sodep res so r effect and t achycard ia in p r e sence of pra- zosin (n = 6). P rop rano lo l inc reased signif icant ly the p ressor r e sponse to BK but did not affect the H R response . On the o the r hand, c imet id ine plus d iphen- hydramine (n = 6), m o r p h i n e (n = 7), i ndome thac in (n = 6) and i.t. t r e a t m e n t with idazoxan (n = 6) had no signif icant effect on BK- induced M A P and H R changes (fig. 5). Base l ine M A P r e m a i n e d unchanged by all
143
t r ea tmen t s , whe reas base l ine H R was signif icant ly in- c r ea sed by a t rop ine (from 374.3 + 9.5 to 435.0 _+ 7.8 b e a t s / m i n ; P < 0.05, n = 14) and dec rea sed by p rop ra - nolol ( f rom 362.7 + 7.2 to 326.4 _+ 11.9 b e a t s / r a i n ; P < 0.01, n = l l ) . F u r t h e r m o r e , ne i the r the p ressor re- sponse nor the b r adyca rd i a induced by BK (81 pmol ) were s ignif icant ly a l t e red by b i la te ra l ad rena l medu l - lec tomy (fig. 6). W h e r e a s the p ressor responses to BK (81 and 810 pmol ) were unaf fec ted by capsaic in t rea t - ment , the H R responses were b locked and conver ted to m a r k e d increases in HR. Pen to l in ium conver t ed to increases the dec reases in H R induced by both i.t. BK and i.v. nicot ine. Pen to l in ium signif icant ly r e d u c e d the p re s so r r e sponse to n icot ine but did not affect tha t
A 4 0
30-
2 0
10 !o ,,~ -10
-20
-30
-40
o _ - -
98.3 7.8 . - • 109.2±132 94.5_61 90.5 6.4 673+6.9 1138+63
8 80
60
4 0
20 ¸
.~, 0 ̧ < - 2 0
- 4 0
- 6 0
-80
~ ! 3 5 0 0 :t 31.3 ~ 326.4 -+ 11.9 411.0 ± 22~
Fig. 5. Effects of various treatments on maximal changes in MAP (A) and HR (B) induced by the i.t. injection of CSF ([]) or 81 pmol BK. The inhibitors were given i.v. 20 or 60 rain (indomethacin) prior to the second injection of BK at doses indicated in parentheses except idazoxan which was given i.t. 15 rain prior to BK. Baseline MAP and HR are shown for each treatment. Each point represents the mean+S.E.M, for several rats (see Results). The statistical significance of the differences between means before ([]) and after treatment (11)was calculated with Student's
t-test for paired samples: * P < 0.05; * * P < 0.01; * ** P < 0.001.
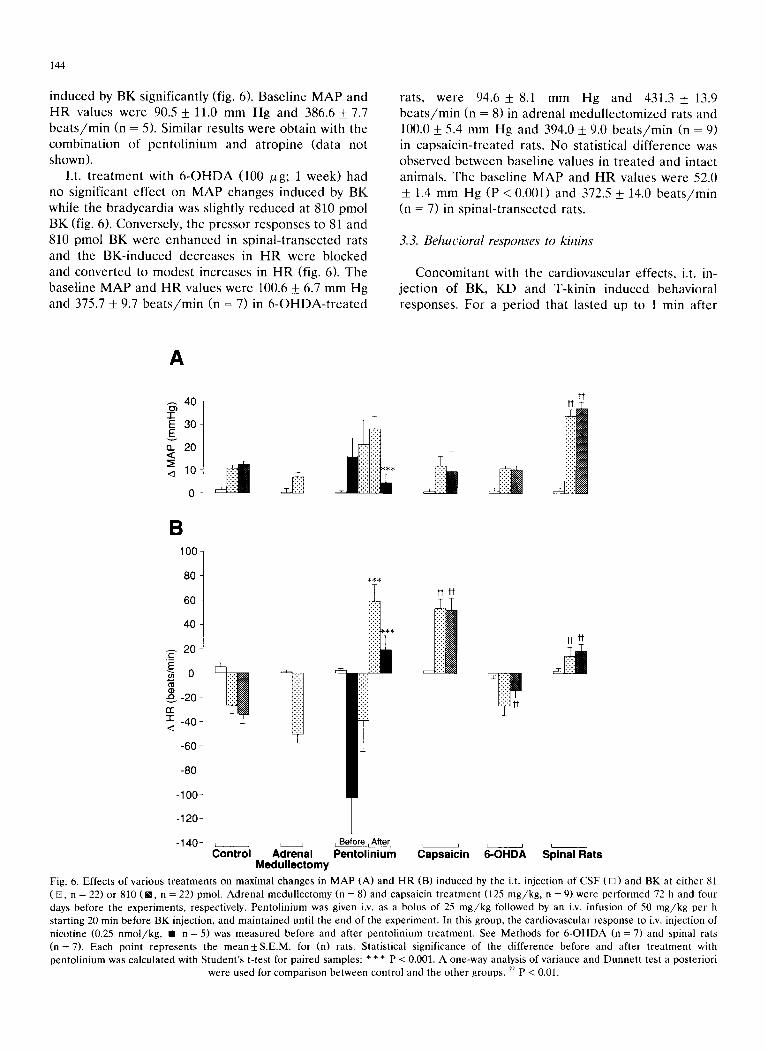
144
i n d u c e d by B K s ign i f i can t ly (fig. 6). B a s e l i n e M A P a n d
H R v a l u e s w e r e 90.5 _+ 11.0 m m H g and 386.6 _+ 7.7
b e a t s / m i n (n = 5). S imi l a r resu l t s w e r e o b t a i n wi th t he
c o m b i n a t i o n o f p e n t o l i n i u m a n d a t r o p i n e (da t a no t
shown) .
I.t. t r e a t m e n t wi th 6 - O H D A (100 Ixg; 1 w e e k ) had
no s ign i f i can t e f f ec t on M A P c h a n g e s i n d u c e d by B K
wh i l e t he b r a d y c a r d i a was s l ight ly r e d u c e d at 810 p m o l
B K (fig. 6). C o n v e r s e l y , t h e p r e s s o r r e s p o n s e s to 81 and
810 p m o l B K w e r e e n h a n c e d in s p i n a l - t r a n s e c t e d rats
and the B K - i n d u c e d d e c r e a s e s in H R w e r e b l o c k e d
and c o n v e r t e d to m o d e s t i n c r e a s e s in H R (fig. 6). T h e
b a s e l i n e M A P a n d H R v a l u e s w e r e 100.6 _+ 6.7 m m H g
and 375.7 _+ 9.7 b e a t s / r a i n (n = 7) in 6 - O H D A - t r e a t e d
rats , w e r e 94.6 _+ 8.1 m m H g and 431.3 _+ 13.9
b e a t s / m i n (n = 8) in a d r e n a l m e d u l l e c t o m i z e d rats and
100.0 _+ 5.4 m m H g and 394.0 +_ 9.0 b e a t s / m i n (n = 9)
in c a p s a i c i n - t r e a t e d rats. N o s ta t i s t ica l d i f f e r e n c e was
o b s e r v e d b e t w e e n b a s e l i n e va lues in t r e a t e d and in tac t
an imals . T h e ba se l i ne M A P and H R va lues w e r e 52.0
_+ 1.4 m m H g (P < 0.001) and 372.5 _+ 14.0 b e a t s / m i n
(n = 7) in s p i n a l - t r a n s e c t e d rats.
3.3. Behadora l responses to kinins
C o n c o m i t a n t wi th t he c a r d i o v a s c u l a r e f fec ts , i.t. in-
j e c t i on o f BK, K D and T -k in in i n d u c e d b e h a v i o r a l
r e sponse s . F o r a p e r i o d tha t l a s ted up to 1 min a f t e r
A
4O t
E 30 E n 20 < Z; .,~ 10
11
B 100-
8O
60
40
20
) o
-20 n-"
-1- -40 , d
-60
-80
-100
-120
-140 L J Control
# t t ] #11
J
L i i B e f o r e i A f t e q t J t J i i
Adrenal Pentolinium Capsaiein 6-OHDA Spinal Rats Medullectomy
Fig. 6. Effects of various treatments on maximal changes in MAP (A) and HR (B) induced by the i.t. injection of CSF ([]) and BK at either 81 (m, n = 22) or 810 (m, n = 22) pmol. Adrenal medullectomy (n = 8) and capsaicin treatment (125 mg/kg, n = 9) were performed 72 h and four days before the experiments, respectively. Pentolinium was given i.v. as a bolus of 25 mg/kg followed by an i.v. infusion of 50 mg/kg per h starting 20 min before BK injection, and maintained until the end of the experiment. In this group, the cardiovascular response to i.v. injection of nicotine (0.25 nmol/kg, • n = 5) was measured before and after pentolinium treatment. See Methods for 6-OHDA (n = 7) and spinal rats (n = 7). Each point represents the mean_+S.E.M, for (n) rats. Statistical significance of the difference before and after treatment with pentolinium was calculated with Student's t-test for paired samples: * * * P < 0.001. A one-way analysis of variance and Dunnen test a posteriori
were used for comparison between control and the other groups. ~ P < 0.01.

145
injection of kinins, the rat became restless and showed exploratory activity at all doses tested. A lateral rock- ing motion on the posterior limbs was also observed in most of the rats. At high doses of BK (0.81 to 8.1 nmol), almost all animals vocalized while at smaller doses, vocalization was less frequent. None of the animals in this study showed grooming or scratching. The initial behavioral excitation was followed by seda- tion which lasted for more than 15 min and was more apparent after repeated administration of BK. How- ever, at 8.1 nmol and higher doses, 50% of the animals collapsed and died within 5 min. Repeated administra- tion of BK (8.1 nmol) also produced hindlimb paralysis in several animals. The behavioral excitation induced by 81 pmol BK was absent in capsaicin-treated and morphine-treated rats, but its intensity and duration were potentiated in 6-OHDA-treated animals. In all cases, vehicle injection had no behavioral effects but [des-Argg]BK caused minor behavioral responses with- out vocalization, paralysis or death.
4. Discussion
The i.t. injection of BK, KD and T-kinin was found to increase blood pressure and decrease HR in con- scious rats. These cardiovascular responses had a rapid onset, suggesting an action directly at the segment of injection. Indeed, the biological half-life of BK after i.t. injection is thought to be as short as that reported (< 30 s) after i.c.v, administration in conscious rats (Kariya et al., 1982). Peripheral leakage of the injected peptide into systemic circulation cannot account for these responses as BK and the other kinins produced vasodepressor responses after peripheral (i.v. and i.a.) injections in rats (Kondo et al., 1979; Regoli and Barab6, 1980). Previous studies using dyes and radio- tracers have documented that drugs injected to rats by an i.t. catheter do not reach supraspinal structures within the time of the experiments (Yaksh and Rudy, 1976; Cridland et al., 1987). Furthermore, in the pre- sent study doses as small as 0.81-8.1 pmol BK pro- duced cardiovascular changes while doses a 100-fold higher were required to evoke a pressor response after i.c.v, injection in consicious rats (Lambert and Lang, 1970; Corr~a and Graeff, 1974; Kondo et al., 1979; Lewis and Phillips, 1984; Buccafusco and Serra, 1985; Lindsey et al., 1989; Madeddu et al., 1990). Contrary to its i.t. injection, i.c.v. BK increased heart rate in the conscious rat (Buccafusco and Serra, 1985). Addition- ally, the pressor response to i.t. BK was markedly enhanced rather than blocked after transection of the cervical spinal cord. Together, these results rule out peripheral and supraspinal sites of action for i.t. BK and related peptides.
To address the possibility that the cardiovascular
response observed in conscious rat is a consequence of perception of a noxious stimulus induced by i.t. injec- tion of kinins, experiments were conducted in mor- phine-treated and capsaicin-treated animals. The sys- temic dose of morphine that was used is sufficient to prevent a HR response to noxious stimulation (Kissin et al., 1984). In the present study, morphine abolished all the behavioral responses while the lateral rocking movement and cardiovascular responses induced by BK remained unaffected. On the other hand, behav- ioral responses induced by BK were enhanced in 6- OHDA-trea ted rats although the cardiovascular effects of BK remained unaltered. BK seems to affect noci- ception through two mechanisms in the spinal cord. BK causes both nociceptive and antinociceptive re- sponses (hypoalgesia) after i.t. injection in the awake rat (Laneuville et al., 1989). The former response may be related to a direct action of BK on primary sensory terminals, as an autoradiographic study had shown the presence of B e receptor binding sites in the substantia gelatinosa of the guinea pig spinal cord (Steranka et al., 1988). The latter response was ascribed to an action of BK on bulbospinal noradrenergic inhibitory fibers (Laneuville et al., 1989). The specific noradrenergic deafferentation of the spinal cord with 6-OHDA elimi- nated the antinociceptive component and left intact the nociceptive counterpart. Results from capsaicin- treated animals eliminate a potential sensory compo- nent in the BK-induced cardiovascular responses. Cap- saicin treatment of the adult rat is known to impair primary sensory C-fibers (Maggi and Meli, 1988). This treatment abolished the nociceptive responses induced by i.t. BK without changing the pressor response. Moreover, the pressor response to i.t. BK was en- hanced rather than inhibited in spinal-transected rats. This latter potentiation may be related to the blockade of vagal influence as well as to the decrease in baseline MAP, leading to greater changes. Thus, cardiovascular responses to kinins do not appear to derive from the initial nociceptive-related behaviors elicited by i.t. in- jection of BK.
The i.t. effect of kinins appears to be mediated by a B 2 kinin receptor in the spinal cord. The des-Argg-BK fragment, a potent stimulant of the B~ kinin receptor subtype which has less than 1% the relative affinity of BK on the B 2 receptor in peripheral bioassays (Regoli and Barab& 1980), was two log units less potent than BK on HR and increased MAP only at a very high dose (81 nmol) although, at this dose, the pressor response was similar for BK and the B~ kinin agonist. Interest- ingly, it has been reported that the pressor responses to i.c.v. BK and other kinins are mediated by receptors of the B 2 subtype as demonstrated with selective B~ and B 2 receptor-selective antagonists (Lindsey et al., 1989). These findings are consistent with the absence of kininase I in the rat CSF (Yoshida and Nosaka,

146
1990). Thus, the formation of the highly selective B, receptor agonist des-Argg-BK which is a metabolite of BK produced by kininase I is thought not to take place, at least in the CSF.
4.1. Central mechanism underlying the spinal action of BK on the cardiot,ascular system
The cardiovascular (present study) and antinocicep- tive (Laneuville et al., 1989) responses elicited by i.t. BK seem to be mediated by separate mechanisms in the rat. Whereas the antinociceptive response to BK (81 pmol) was blocked by prior i.t. administration of idazoxan (15 rain earlier) or 6 -OHDA (1 week earlier), the latter t reatments preserved both the pressor effect and the bradycardia induced by BK (81 pmol). Hence, contrary to the BK-induced antinociception, the effect of i.t. BK on the rat cardiovascular system is unlikely to be attributable to a spinal noradrenergic mechanism. Furthermore, as discussed above, an action of BK on spinal sensory terminals cannot explain the cardiovas- cular response to BK. Hence, the possibility must be considered that BK acts directly on preganglionic sym- pathetic fibers in the intermediolateral cell column of the spinal cord.
4.2. Peripheral mechanism underlying the spinal action of BK on the cardiovascular system
The spinal action of BK on blood pressure appears to be attributable to the release of catecholamines from sympathetic fibers and the adrenal medulla as suggested by the inhibitory action of the a , -adrenocep- tor blocker prazosin on this vascular response. The release of catecholamines from the adrenal medulla does not appear to be essential as the pressor response to BK remained unaffected after bilateral adrenal medullectomy. Nevertheless the vasodepressor re- sponse induced by BK in prazosin-treated rats was prevented by propranolol (results not shown), which indicates that a vascular /3-adrenergic component is unmasked after a-adrenergic inhibition. Thus, in intact animals, the adrenal medulla seems to be activated by i.t. BK as circulating adrenaline is the natural agonist for both a- and /3-adrenoceptors while noradrenaline has greater affinity for a-adrenoceptors . In agreement with this proposal, propranolol enhanced the pressor response to BK while prazosin converted the pressor response of BK to a vasodepressor response in intact (fig. 5) but not in adrenal medullectomized rats (un- published observations). Therefore, the two vascular adrenoceptors (a and/3) appear to be activated follow- ing the release of peripheral catecholamines induced by i.t. BK. These two opposite effects on vascular tone are thought to modulate the vascular response to BK
by reducing significantly the magnitude of the pressor r e s p o n s e .
A finding which remains to be explained is the inability of pentolinium (alone or in combination with atropine) to prevent the pressor response to BK. Nev- ertheless, this t reatment was able to reduce signifi- cantly the pressor response induced by systemic admin- istration of nicotine and to block the bradycardia in- duced by both nicotine and BK. The participation of other mediators such as histamine, acetylcholine and prostaglandins in the pressor response to BK can be excluded.
That prostaglandins and histamine are not involved in the bradycardia induced by BK is evidenced by experiments with indomethacin and antihistaminics. The bradycardia induced by BK was abolished and converted to tachycardia by methylnitrate atropine, pentolinium and capsaicin, suggesting that the brady- cardia is due to the reflex activation of the vagus nerve. Sensory C-fibers are also involved in this reflex re- sponse. Although the MAP and H R responses to BK appeared to change in opposite directions (see results with prazosin), three reasons make it difficult to con- clude that there is a baroreflex mechanism. First, large decreases of H R were measured, with small increases ( ~ 10 mm Hg) of MAP; second, there was no correla- tion between MAP and H R changes when dose- re - sponse curves to kinins were compared and third, capsaicin sensitive sensory fibers do not appear to play a role in baroreflex activity in the rat (Takano et al., 1988). The possibility that the vagus nerve is activated secondary to the spinal activation of ascending fibers to supraspinal centers by BK is supported by the absence of bradycardia in spinal-transected rats. Such a possi- bility was suggested in a previous study demonstrating that transection of the cervical spinal cord prevented the increase of cutaneous vascular permeability pro- duced by i.t. BK (Jacques and Couture, 1990). In the latter study, the vagus nerve and capsaicin-sensitive sensory fibers were also required.
In summary, this is the first report demonstrating that kinins can affect the cardiovascular system through an action directly in the spinal cord of the conscious rat. The effects of BK on MAP and H R appear to be mediated by two separate mechanisms. Thus, spinal activation of the sympathoadrenal system is the mecha- nism likely to explain the pressor response while a reflex mechanism involving sensory C-fibers and the vagus nerve can account for the bradycardia. This latter response is likely to be triggered by activation of a spinobulbar pathway. This spinal action of BK on the cardiovascular system is of interest and deserves fur- ther investigations as kinins are present in the CSF and the spinal cord (see Introduction) and may reach criti- cal levels during spinal injury, surgery or noxious ma- nipulations.

Acknowledgements
This work was supported by a Grant-in-Aid from the Medical Research Council of Canada. R.C. is a senior Scholar of the Fonds de la Recherche en Sant~ du Qudbec. The secretarial assistance of Christiane Laurier and the graphic work of Claude Gauthier are gratefully acknowledged.
References
Adam, A., J. Damas, G. Calay, C. Renard, G. Remacle-Volon and V. Bourdon, 1989, Quantification of rat T-kininogen using im- munological methods, Biochem. Pharmacol. 38, 1569.
Barlas, A,, H. Okamoto and L.M. Greenbaum. 1985, T-kininogen - the major plasma kininogen in rat adjuvant arthritis, Biochem. Biophys. Res. Commun. 129, 280.
Buccafusco, J.J. and M. Serra, 1985, Role of cholinergic neurons in the cardiovascular responses evoked by central injection of bradykinin or angiotensin II in conscious rats, Eur. J. Pharmacol. 113, 43.
Coote, J.H., 1988, The organisation of cardiovascular neurons in the spinal cord, Rev. Physiol. Biochem. Pharmacol. 110, 147.
Corrgaa, F.M.A. and F.G. Graeff, 1974, Central mechanisms of the hypertensive action of intraventricular bradykinin in the unanaes- thetized rat, Neuropharmacology 13, 65.
Cridland, R.A., K. Yashpal, V.V. Romita, S. Gauthier and J.L. Henry, 1987, Distribution of label after intrathecal administration of t>l-substance P in the rat, Peptides 8, 213.
Diz, D.I., 1985, Bradykinin and related peptides in central control of the cardiovascular system, Peptides 6, 57.
Fujiwara, Y., C.R. Mantione, R.J. Vavrek, J.M. Stewart and H.I. Yamamura, 1989, Characterization of [ -~ H]bradykinin binding sites in guinea-pig central nervous system: possible existence of B e subtypes, Life Sci. 44, 1645.
Hassdssian, H. and R. Couture, 1989, Cardiovascular responses induced by intrathecal substance P in the conscious freely moving rat, J. Cardiovasc. Pharmacol. 13, 594.
Hass&sian, H., R. Couture and J. De Champlain, 1990, Sympathoad- renal mechanisms underlying cardiovascular responses to in- trathecal substance P in conscious rats, J. Cardiovasc. Pharmacol. 15, 736.
Hermann, K., G. Schaechtelin and M. Marin-Grez, 1986, Kinins in cerebrospinal fluid: reduced concentration in spontaneously hy- pertensive rats, Experientia 42, 1238.
Hoffman, W.E. and P.G. Schmid, 1978, Separation of pressor and antidiuretic effects of intraventricular bradykinin, Neuropharma- cology 17, 999.
Jacques, L. and R. Couture, 1990, Studies on the vascular permeabil- ity induced by intrathecal substance P and bradykinin in the rat, Eur. J. Pharmacol. 184, 9.
Kariya, K., A. Yamauchi, S. Hattori, Y. Tsuda and Y. Okada, 1982, The disappearance rate of intraventricular bradykinin in the brain of the conscious rat, Biochem. Biophys. Res. Commun. 107, 1461.
Kariya, K., A. Yamauchi and T. Sasaki, 1985, Regional distribution and characterization of kinin in the CNS of the rat, J. Neu- rochem. 44, 1892.
Kissin, l., C.R. Kerr and L.R. Smith, 1984, Effect of morphine on the heart rate response to noxious stimulation: interaction with halothane and naloxone, Pain 18, 351.
Kondo, K.+ T. Okuno, K. Konishi, T. Saruta and E. Kato, 1979, Central and peripheral effects of bradykinin and prostaglandin E2 on blood pressure in concious rats, Naunyn-Schmiedeb. Arch. Pharmacol. 308, 111.
Lambert, G.A. and W.J. Lang, 1970, The effects of bradykinin and
147
eledoisin injected into the cerebral ventricles of conscious rats, Eur. J. Pharmacol. 9, 383.
Laneuville, O. and R. Couture, 1987, Bradykiniu analogue blocks bradykinin-induced inhibition of a spinal nociceptive reflex in the rat, Eur. J. Pharmacol. 137. 281.
Laneuville, O., T.A. Reader and R. Couture, 1989, Intrathecal bradykinin acts presynaptically on spinal noradrenergic terminals to produce antinociception in the rat, Eur. J. Pharmacol. 159, 273.
Lewis, R.E. and M.I. Phillips, 1984, Localization of the central pressor action of bradykinin to the cerebral third ventricle, Am. J. Physiol. 247, R63.
Lindsey, C.J., C.R. Nakaie and D.T.O. Martins, 1989, Central ner- vous system kinin receptors and the hypertensive response medi- ated by bradykinin, Br. J. Pharmacol. 97, 763.
Lopes, P. and R. Couture, 1991, Cardiovascular responses induced by the intrathecal injection of kinins in the freely moving rat, Can. J. Physiol. Pharmacol. 69, Abstract AXVii.
Madeddu, P., N. Glorioso, A. Soro, G. Tonolo, P. Manunta, C. Troffa, M.P. Demontis+ M.V. Varoni and V. Anania, 1990, Brain kinins are responsible for the pressor effect of intracerebroven- tricular captopril in spontaneously hypertensive rats, Hyperten- sion 15, 407.
Maggi, C.A. and A. Meli, 1988, The sensory-efferent function of capsaicin-sensitive sensory neurons, Gen. Pharmacol. 19, 1.
Okamoto, H. and L.M. Greenbaum, 1983a, Isolation and structure of T-kinin, Biochem. Biophys. Res. Commun. 112, 701.
Okamoto, H. and L.M. Greenbaum, 1983b, Pharmacological proper- ties of T-kinin (isoleucyl-seryl-bradykinin) from rat plasma, Biochem. Pharmacol. 32, 2637.
Perry,, D.C. and S.H. Snyder, 1984, Identification of bradykinin in mammalian brain, J. Neurochem. 43, 1072.
Regoli, D., 1987, Kinins, Br. Med. Bull. 43, 270. Regoli, D. and J. Barab~, 1980, Pharmacology of bradykinin and
related kinins, Pharmacol. Rev. 32, 1. Scicli, A.G., G. Forbes, H. Nolly, M. Dujovny and O.A. Carretero,
1984, Kallikrein-kinins in the central nervous system, Clin. Exp. Hypert, A6, 1731.
Sharif, N.A. and R.L+ Whiling, 1991, Identification of Be-bradykinin receptors in guinea pig brain regions, spinal cord and peripheral tissues, Neurochem. Int. 18, 89.
Steranka, L.R., D.C. Manning, C.J. DeHaas, J.W. Ferkany, S.A. Borosky, J.R. Connor, R.J. Vavrek, J.M. Stewart and S.H. Sny- der, 1988, Bradykinin as a pain mediator: receptors are localized to sensory neurons, and antagonists have analgesic actions, Proc. Natl. Acad. Sci. 85, 3245.
Takano, Y., A. Nagashima, H. Kamiya, M. Kurosawa and A. Sato, 1988, Well-maintained reflex responses of sympathetic nerve ac- tivity to stimulation of baroreceptor, chemoreceptor and cuta- neous mechanoreceptors in neonatal capsaicin-treated rats, Brain Res. 445, 188.
Thomas, G.R. and C.R. Hiley, 1988, Cardiovascular effects of intra- cerebro-ventricular bradykinin and melittin in the rat, J. Pharm. Pharmacol. 40, 721.
Thomas G.R., H. Thibodeaux, H.S. Margolius and P.J. Provitera, 1984, Cerebrospinal fluid kinins and cardiovascular function ef- fects of cerebroventricular melittin, Hypertension 6 (Suppl. 1), 1-46.
Thomas, G.R., H. Thibodeaux, H.S. Margolius, J.G. Webb and P.J. Privitera, 1987, Afferent vagal stimulation, vasopressin, and nitro- prusside alter cerebrospinal fluid kinin, Am. J. Physiol. 253, R136.
Yaksh, T.L. and T.A. Rudy, 1976, Chronic catheterization of the spinal subarachnoid space, Physiol. Behav. 17, 1031.
Yoshida, T, and S. Nosaka, 1990, Some characteristics of a peptidyl dipeptidase (kininase I1) from rat CSF: differential effects of NaCI on the sequential degradation steps of bradykinin, J. Neu- rochem. 55, 1861.
Related Documents