Capsicum annuum S (CaS) promotes reproductive transition and is required for flower formation in pepper (Capsicum annuum) Oded Cohen, Yelena Borovsky, Rakefet David-Schwartz and Ilan Paran Institute of Plant Science, Agricultural Research Organization, The Volcani Center, PO Box 6, Bet Dagan 50250, Israel Author for correspondence: Ilan Paran Tel: +972 3 9683943 Email: [email protected] Received: 2 December 2013 Accepted: 7 January 2014 New Phytologist (2014) doi: 10.1111/nph.12711 Key words: flower formation, pepper (Capsicum annuum), petunia (Petunia hybrida), sympodial growth, tomato (Sola- num lycopersicum), transition to flowering. Summary The genetic control of the transition to flowering has mainly been studied in model species, while few data are available in crop species such as pepper (Capsicum spp.). To elucidate the genetic control of the transition to flowering in pepper, mutants that lack flowers were iso- lated and characterized. Genetic mapping and sequencing allowed the identification of the gene disrupted in the mutants. Double mutants and expression analyses were used to characterize the relationships between the mutated gene and other genes controlling the transition to flowering and flower differentiation. The mutants were characterized by a delay in the initiation of sympodial growth, a delay in the termination of sympodial meristems and complete inhibition of flower formation. Capsicum annuum S (CaS), the pepper (Capsicum annuum) ortholog of tomato (Solanum lycopersicum) COMPOUND INFLORESCENCE and petunia (Petunia hybrida) EVERGREEN, was found to govern the mutant phenotype. CaS is required for the activity of the flower meristem identity gene Ca-ANANTHA and does not affect the expression of CaLEAFY. CaS is epistatic over other genes controlling the transition to flowering with respect to flower formation. Comparative homologous mutants in the Solanaceae indicate that CaS has uniquely evolved to have a critical role in flower formation, while its role in meristem maturation is con- served in pepper, tomato and petunia. Introduction Flowering plants undergo major transitions in their life cycles: first a transition from the vegetative juvenile phase to the vegeta- tive adult phase and then a transition to the reproductive phase (Huijser & Schmid, 2011). The transition from the vegetative phase to the flowering phase has been most extensively character- ized in Arabidopsis (recently reviewed by Srikanth & Schmid, 2011). The shoot apical meristem (SAM) controls the aerial part of the plant, first producing the stem and leaves and then, upon the transition to flowering, producing the inflorescence meristem (IM). Plants such as Arabidopsis and Antirrhinum majus are char- acterized by monopodial shoot architecture in which the SAM is active throughout the plant’s life (indeterminate); that is, after the vegetative phase, flowers develop continuously on the flank of the SAM. By contrast, plants in the family Solanaceae, such as petunia (Petunia hybrida), tomato (Solanum lycopersicum) and pepper (Capsicum annuum), are characterized by sympodial shoot architecture: the SAM terminates in an IM, and plant growth continues from lateral meristems, termed sympodial meristems (SYMs), which develop in the axil of the youngest leaf below the apical inflorescence (Schmitz & Theres, 1999). The SYM devel- ops into a shoot segment termed the sympodial unit (SU), consisting of vegetative and reproductive organs in a species- specific pattern, and new SYMs are repeatedly formed from the axils of the uppermost leaves of the preceding SU. Two of the most important genes determining inflorescence architecture and floral meristem (FM) identity in Arabidopsis are LEAFY (LFY) and TERMINAL FLOWER1 (TFL1). LFY is a transcription factor that responds to external stimuli, and directly and indirectly activates or represses a large network of genes that determine the identity of floral organs (Winter et al., 2011; Grandi et al., 2012). LFY has a role in regulating flowering in all flowering plants, and it also exists in nonflowering plants; how- ever, its role in these latter plants is not well understood (Moy- roud et al., 2010). TFL1 is a flowering repressor required to maintain the indeterminate state of the inflorescence (Bradley et al., 1997). Theoretical modeling suggests that variation in the architecture of the inflorescence may be explained by alterations in the spatio-temporal expression patterns of LFY and TFL1 (Prusinkiewicz et al., 2007). While research on the regulation of flowering and shoot archi- tecture has focused mainly on monopodial species, in particular Arabidopsis (Srikanth & Schmid, 2011), knowledge of the genes, and their interactions, regulating these processes in sympodial plants has lagged behind. The most studied sympodial model Ó 2014 Agricultural Research Organization New Phytologist Ó 2014 New Phytologist Trust New Phytologist (2014) 1 www.newphytologist.com Research

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Capsicum annuum S (CaS) promotes reproductive transition andis required for flower formation in pepper (Capsicum annuum)
Oded Cohen, Yelena Borovsky, Rakefet David-Schwartz and Ilan Paran
Institute of Plant Science, Agricultural Research Organization, The Volcani Center, PO Box 6, Bet Dagan 50250, Israel
Author for correspondence:Ilan ParanTel: +972 3 9683943
Email: [email protected]
Received: 2 December 2013
Accepted: 7 January 2014
New Phytologist (2014)doi: 10.1111/nph.12711
Key words: flower formation, pepper(Capsicum annuum), petunia (Petuniahybrida), sympodial growth, tomato (Sola-num lycopersicum), transition to flowering.
Summary
� The genetic control of the transition to flowering has mainly been studied in model species,
while few data are available in crop species such as pepper (Capsicum spp.). To elucidate the
genetic control of the transition to flowering in pepper, mutants that lack flowers were iso-
lated and characterized.� Genetic mapping and sequencing allowed the identification of the gene disrupted in the
mutants. Double mutants and expression analyses were used to characterize the relationships
between the mutated gene and other genes controlling the transition to flowering and flower
differentiation.� The mutants were characterized by a delay in the initiation of sympodial growth, a delay in
the termination of sympodial meristems and complete inhibition of flower formation.
Capsicum annuum S (CaS), the pepper (Capsicum annuum) ortholog of tomato (Solanum
lycopersicum) COMPOUND INFLORESCENCE and petunia (Petunia hybrida) EVERGREEN,
was found to govern the mutant phenotype. CaS is required for the activity of the flower
meristem identity gene Ca-ANANTHA and does not affect the expression of CaLEAFY. CaS is
epistatic over other genes controlling the transition to flowering with respect to flower
formation.� Comparative homologous mutants in the Solanaceae indicate that CaS has uniquely
evolved to have a critical role in flower formation, while its role in meristem maturation is con-
served in pepper, tomato and petunia.
Introduction
Flowering plants undergo major transitions in their life cycles:first a transition from the vegetative juvenile phase to the vegeta-tive adult phase and then a transition to the reproductive phase(Huijser & Schmid, 2011). The transition from the vegetativephase to the flowering phase has been most extensively character-ized in Arabidopsis (recently reviewed by Srikanth & Schmid,2011). The shoot apical meristem (SAM) controls the aerial partof the plant, first producing the stem and leaves and then, uponthe transition to flowering, producing the inflorescence meristem(IM). Plants such as Arabidopsis and Antirrhinum majus are char-acterized by monopodial shoot architecture in which the SAM isactive throughout the plant’s life (indeterminate); that is, afterthe vegetative phase, flowers develop continuously on the flank ofthe SAM. By contrast, plants in the family Solanaceae, such aspetunia (Petunia hybrida), tomato (Solanum lycopersicum) andpepper (Capsicum annuum), are characterized by sympodial shootarchitecture: the SAM terminates in an IM, and plant growthcontinues from lateral meristems, termed sympodial meristems(SYMs), which develop in the axil of the youngest leaf below theapical inflorescence (Schmitz & Theres, 1999). The SYM devel-ops into a shoot segment termed the sympodial unit (SU),
consisting of vegetative and reproductive organs in a species-specific pattern, and new SYMs are repeatedly formed from theaxils of the uppermost leaves of the preceding SU.
Two of the most important genes determining inflorescencearchitecture and floral meristem (FM) identity in Arabidopsis areLEAFY (LFY) and TERMINAL FLOWER1 (TFL1). LFY is atranscription factor that responds to external stimuli, and directlyand indirectly activates or represses a large network of genes thatdetermine the identity of floral organs (Winter et al., 2011;Grandi et al., 2012). LFY has a role in regulating flowering in allflowering plants, and it also exists in nonflowering plants; how-ever, its role in these latter plants is not well understood (Moy-roud et al., 2010). TFL1 is a flowering repressor required tomaintain the indeterminate state of the inflorescence (Bradleyet al., 1997). Theoretical modeling suggests that variation in thearchitecture of the inflorescence may be explained by alterationsin the spatio-temporal expression patterns of LFY and TFL1(Prusinkiewicz et al., 2007).
While research on the regulation of flowering and shoot archi-tecture has focused mainly on monopodial species, in particularArabidopsis (Srikanth & Schmid, 2011), knowledge of the genes,and their interactions, regulating these processes in sympodialplants has lagged behind. The most studied sympodial model
� 2014 Agricultural Research Organization
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 1
www.newphytologist.com
Research

species are tomato and petunia. In both, generation and charac-terization of developmental mutants have enabled the identifica-tion of key genes controlling flowering time, inflorescencearchitecture, axillary branching and sympodial shoot develop-ment (reviewed by Angenent et al., 2005; Lozano et al., 2009;Castel et al., 2010). Those studies have revealed that, in mostcases, genes that function in regulating these processes in sympo-dial plants have a related function in monopodial plants.
The tomato reproductive shoot is composed of reiterated SUseach consisting of three leaves and an inflorescence. The inflores-cence of tomato has a reiterated sympodial shoot structure com-posed of one-nodal SUs each terminated by a single flower(Lippman et al., 2008). The tomato inflorescence architectureexhibits large phenotypic variation in the degree of branching.The inflorescences of mutants such as single flower truss (sft) anduniflora (uf) consist of single flowers (Dielen et al., 2004;Lifschitz et al., 2006), while those of anantha (an), falsiflora (fa)and compound inflorescence (s) are highly branched (Lippmanet al., 2008). SINGLE FLOWER TRUSS (SFT ) is the ortholog ofthe florigen-encoding gene FLOWERING LOCUS T (FT ) inArabidopsis. ANANTHA (AN ) and FALSIFLORA (FA ) are theorthologs of the FM identity genes UNUSUAL FLORALORGANS (UFO) and LFY in Arabidopsis, respectively, andCOMPOUND INFLORESCENCE (S ) encodes a Wuschel-homeobox (WOX) transcription factor that functions in themaintenance of stem cells and is not known to have a role ininflorescence architecture in Arabidopsis.
After termination of the primary stem by a single flower, thepetunia reproductive shoot is composed of reiterated SUs eachconsisting of two leaf-like organs (bracts) and a single flower(Castel, 2009). Unlike UFO in Arabidopsis, which has a limitedrole in regulating inflorescence architecture and IM formation,the role of the petunia ortholog DOUBLE TOP (DOT) in deter-mining inflorescence architecture is more critical, and its expres-sion is necessary and sufficient to induce flowering (Souer et al.,2008). In dot mutants, the inflorescence has a leafy appearanceand it lacks flowers, resembling the phenotype of the tomato or-tholog an. Expression of DOT is dependent on prior expressionof EVERGREEN (EVG), the homolog of tomato S (Rebochoet al., 2008). DOT further activates ABERRANT LEAF ANDFLOWER (ALF ), the petunia ortholog of LFY. In both tomato sand petunia evg mutants, expression of AN and DOT is delayedor reduced (Lippman et al., 2008; Rebocho et al., 2008). Thedelay of AN expression in s mutants is postulated to extend theperiod of IM indeterminacy, leading to the mutant’s highlybranched inflorescence phenotype (Lippman et al., 2008). Bycontrast, lack of DOT activation in evg results in loss of flowering(Rebocho et al., 2008).
Pepper (Capsicum spp.) is a member of the Solanaceae, havinga sympodial shoot structure with solitary flowers. The distinctshoot architecture of pepper compared with other studied Solan-aceae such as tomato and petunia suggests possible diversificationof genes and pathways involved in controlling this trait. There-fore, to extend our understanding of the genetic control of shootarchitecture in the Solanaceae, natural and induced variation inpepper mutants altered in their shoot architecture was isolated
and characterized. Those studies enabled the identification of keygenes controlling sympodial shoot development (FASCICULATEand CaJOINTLESS), flower formation (Ca-ANANTHA) andaxillary branching (CaBLIND and CaHAM) in pepper (Lippmanet al., 2008; Elitzur et al., 2009; Jeifetz et al., 2011; Cohen et al.,2012; David-Schwartz et al., 2013). In the course of screening anethyl methanesulfonate (EMS)-mutagenized population (Paranet al., 2007), allelic mutants that lack flowers were observed. Thecomplete lack of flowers and additional associated phenotypesindicated that the disrupted gene in those mutants has a centralrole in regulation of shoot architecture and flower formation inpepper. The goals of the present work were to comprehensivelycharacterize the unique nonflowering mutation, to isolate thegene disrupted by the mutation, and to compare gene function inthe Solanaceae by utilizing homologous mutants in other species.We report the identification of CaS, the pepper homolog of Sand EVG in tomato and petunia, respectively, as controlling themutant phenotype. The role of CaS as a flowering promoter andits interaction with other genes controlling reproductive differen-tiation and shoot architecture in pepper are further described.
Materials and Methods
Plant material
The mutants Cas (E-327 and E-648), Cajointless (Caj),Ca-anantha (Ca-an) and E-62 were isolated from an EMS-mutagenized population with Capsicum annuum cv Maor as thewild-type parent (Paran et al., 2007; Lippman et al., 2008; Cohenet al., 2012). Fasciculate (fa) is a mutant with a spontaneous natu-ral mutation described by Elitzur et al. (2009) that was back-crossed to the ‘Maor’ background. An F2 segregating populationwas generated by crossing heterozygotes for the E-327 mutationto Capsicum frutescens BG 2816. Double mutants with Cas werederived from F2 populations obtained by crossing the corre-sponding mutants with individuals heterozygous for the Cas alleleE-327, followed by self-pollination and marker-assisted selection.
Mapping and isolation of CaS
To map the gene governing the E-327 phenotype, the bulkedsegregant analysis (BSA) approach was used (Michelmore et al.,1991). Two bulks of DNA composed of 15 individuals eachfrom mutant and wild-type plants of the F2 segregating popula-tion were constructed and screened with 400 random amplifiedpolymorphic DNA (RAPD) primers. Polymorphic bands wererecovered from an agarose gel, cloned into the pGEM-T EasyVector system (Promega) and used as restriction fragment lengthpolymorphism (RFLP) probes to genotype the F2 population andmap the pepper genome (Rao et al., 2003).
To clone CaS, tomato primers Sl-WoxF1 and Sl-WoxR1 (kindlyprovided by Z. Lippman; Supporting Information Table S1) wereused to amplify an 834-bp fragment of CaS with pepper genomicDNA as the template. To extend the sequence of CaS toward the5′ end of the open reading frame (ORF), inverse PCR (Ochmanet al., 1988) was performed with the primers CaS-IPCRF1,
New Phytologist (2014) � 2014 Agricultural Research Organization
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist2

CaS-IPCRF2, CaS-IPCRR1 and CaS-IPCRR2 (Table S1) andgenomic DNA as the template. To extend the CaS sequencetoward the 3′ end of the ORF, 3′ rapid amplification of cDNAends (RACE) was performed with a Smart RACE kit (Clontech,Palo Alto, CA, USA) and total RNA from the shoot apices. TheORF of CaS from ‘Maor’ was deposited in GenBank (accessionnumber KC414761). Screening for Cas mutants in subsequentexperiments was performed by developing a cleaved amplifiedpolymorphic sequence (CAPS) marker using the primersCaS-DdeF and CaS-DdeR (Table S1) followed by restrictiondigestion with DdeI.
Gene expression analysis
Total RNA was extracted from shoot apices using the GeneEluteMammalian Total RNA Extraction Miniprep kit (Sigma) fol-lowed by DNaseI treatment (Sigma). Total RNA (400 ng) wasused for first-strand cDNA synthesis by reverse transcriptionPCR (RT-PCR) using a PrimeScript RT Reagent kit (Takara BioInc., Otsu, Japan). For real-time quantitative PCR (qRT-PCR),three biological and two technical repeats were used for each sam-ple. For the qRT-PCR experiments, plants were grown in a glass-house under natural daylight during the winter season in Israel.PCR amplification was performed using the primers CaS-qRTFand CaS-qRTR for CaS, CaLFY-qRTF and CaLFY-qRTR forCaLFY and Ca-AN-qRTF and Ca-AN-qRTR for Ca-AN (TableS1). Amplified products were detected using SYBR Premix ExTaq II (Takara) in a Rotor-Gene 6000 thermal cycler (CorbettResearch, Mortlake, Australia). Results were analyzed usingROTOR-GENE 6000 SERIES SOFTWARE 1.7 (Corbett). The relativeexpression levels of the genes were normalized againstCaUBIQUITIN (DQ975458.1) using the primers UBQ-qRTFand UBQ-qRTR.
In situ hybridization
In situ analysis was performed with digoxigenin (DIG)-labeledprobes as previously described (Neta et al., 2011). Meristemswere fixed in FAA (formaldehyde : acetic acid : 70% ethanol,10 : 5 : 85, v/v), then dehydrated and embedded in ParaPlast(McCormick Scientific, St Louis, MO, USA). The tissue wasthen cut (10 lm) on a Leica RM2245 microtome (VectaMount;Vector Laboratories, Peterborough, UK) and sections were placedon SuperFrost Plus slides (Menzel-Glaser, Braunschweig,Germany) for 2 d on a 42°C hot plate. An antisense DIG-labeledRNA probe was synthesized from the 3′ end of CaS cDNA,excluding the conserved homeodomain, using the MEGAscript kit
(Ambion, Austin, TX, USA) and DIG RNA labeling mix (RocheApplied Science, Mannheim, Germany).
Scanning electron microscopy (SEM)
Samples for SEM were fixed directly in 70% ethanol, and criticalpoint dried as described by Alvarez et al. (1992). SEM wasperformed in a Hitachi S-3500N instrument (Hitachi, Tokyo,Japan).
Phylogenetic analysis
Multiple sequence alignments were performed with a web-basedversion of CLUSTALW ( http://www.ebi.ac.uk/Tools/msa/) usingthe default settings. The phylogenetic tree was calculated by theneighbor-joining method and bootstrap analysis with 1000 repli-cates using the MEGA4 software (http://www.megasoftware.net/mega4/mega.html). The tree was calculated from alignments ofthe WOX homeodomain consisting of c. 64 of the proteins’amino acids. Accession numbers were as follows: petunia: Ph-EVG (EF187281) and SISTER OF EVERGREEN (Ph-SOE)(EF187282); Arabidopsis: WUSCHEL (WUS) (CAA09986),WOX1 (AAP37133), WOX2 (AAP37131), WOX3(AAP37135), WOX4 (AAP37134), WOX5 (AAP37136),WOX6 (AAP37137), WOX8 (AAP37138), WOX9(AAP37139), WOX11 (AAP37140), WOX12 (AAP37141) andWOX13 (AAP37142); Antirrhinum majus: Am-WUS(AAO23113); tomato: Sl-S (NP_001234072); Phaseoluscoccineus: Pc-WOX9-like (ACL11801); Populus trichocarpa:Pt-WOX9 (CAJ84153); rice (Oryza sativa): WOX-like(Os07g34880) and WUS-like (Os05g48990); pepper (C.annuum): CaS (KC414761).
Results
Wild-type and mutant phenotypes
After germination and during vegetative growth, the pepperSAM gives rise to a stem and leaves arranged in an alternate spiralpattern. After the development of 10.3� 1 leaves on the primarystem of ‘Maor’ (Table 1), the SAM undergoes a transition toflowering by converting to an FM that subsequently developsinto a flower. Further shoot growth continues from the lateralmeristems which develop in the axils of the two uppermost leavesbelow the apical flower. The subtending leaves from the primaryshoot are carried up and also carry SYMs in their axils to allowcontinuous growth of the shoot. The development of two
Table 1 Transition to flowering of pepper mutants
Maor Cas Caj fa E-62 Cas Caj Cas fa Cas E-62
No. of leaves on primary stem until first flower 10.3� 1.0 21.3� 1.1 17� 0.6 8.6� 0.5 4 61.5� 1.5 20� 1.5 17.7� 1.5
For Casmutants, the number of leaves on the primary stem was measured until initiation of sympodial growth, as reflected by formation of a dichasiallyforked shoot.Caj, Cajointless; fa, fasciculate.
� 2014 Agricultural Research Organization
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014)
www.newphytologist.com
NewPhytologist Research 3

meristems results in the typical dichasially forked shoot of wild-type pepper (Child, 1979; Fig. 1a,b). During early sympodiumdevelopment, the SYM generates primordia of two leaves andterminates by forming a single flower, resulting in an SU thatconsists of two leaves and a flower (Fig. 1a,b). Later in develop-ment, the two leaves of the SYM grow upward and away fromthe flower. The next two SUs emerge at the axils of the leaves ofthe preceding SU and are separated by its flower. This processrepeats itself and results in a shoot architecture that consists ofmultiple SUs arranged in a perpendicular pattern.
An EMS-mutagenized population was generated using theblocky cv Maor as the wild-type parent (Paran et al., 2007) andscreened for alterations in shoot architecture. Two M2 families(10 plants per family), E-327 and E-648, were identified, whichsegregated for a similar lack of flower phenotype in a monogenicrecessive manner. No differences in the severity of the phenotypewere observed between the two mutants. The two mutants werecrossed (in a heterozygous state as revealed by a progeny test) andthe mutant phenotype that segregated in the F1 progeny indi-cated that E-327 and E-648 are allelic. Further work focused onlyon E-327. The mutants were indistinguishable from wild-typeplants during the vegetative growth phase. However, whereas
wild-type plants initiate sympodial growth and floral formationafter the development of 10 leaves on the primary stem, initiationof sympodial growth in the mutant as manifested by forming thedichasially forked shoot occurs later after the development of21.3� 1.1 leaves, and the sympodial branches remain fused toeach other, forming a fasciated shoot (Fig. 1c). The fasciation isreleased after formation of a few additional leaves, allowing sepa-ration of the shoots (Fig. 1d). Subsequent sympodial growth con-tinues in cycles of SU formation in which the SU consists of amultiple, unfixed number of leaves and occasional fasciation ofbranches without formation of flowers, giving rise to the leafyphenotype of the mutant (Fig. 1e).
To determine the ontogeny of the aberrations in E-327, aseries of dissected apices were examined by SEM. In the wild-type, at the stage of two expanding leaves (defined as the numberof leaves on the primary stem of at least 3 cm in length), the api-cal meristem remains vegetative (Fig. 2a). A primordial flower isobserved at the six-expanding-leaf stage (Fig. 2b). At the12-expanding-leaf stage, the terminal flower is flanked by twoleaves and the subsequent SUs, each consisting of two leaves anda flower, develop in the axils of the two preceding leaves (Fig. 2c).In E-327, the SAM remains vegetative for a longer time than in
(a)
(c) (d) (e)
(b)
Fig. 1 Characterization of wild-type andE-327 pepper (Capsicum annuum) plants. (a)Schematic diagram of wild-type parent‘Maor’. At the transition to flowering, theapical flower (flower of the primary shoot,Fp) and the first two leaves (leaves of theprimary shoot, Lp) originate from the primaryshoot. The next leaves belong to the first twosympodial units (SUs), each consisting of twoleaves (leaves of the sympodial shoot, Ls)and a flower (flower of the sympodial shoot,Fs). (b) Wild-type SU consisting of a flower(Fs) and two leaves (Ls). (c) Schematicdiagram of the E-327mutant showing stemfasciation, lack of flowers and leafyappearance. (d) Initiation of sympodialgrowth, primary stem fasciation and itsrelease after formation of a few additionalleaves. (e) The E-327 sympodial shootshowing stem fasciation and multiple leaves.
New Phytologist (2014) � 2014 Agricultural Research Organization
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist4
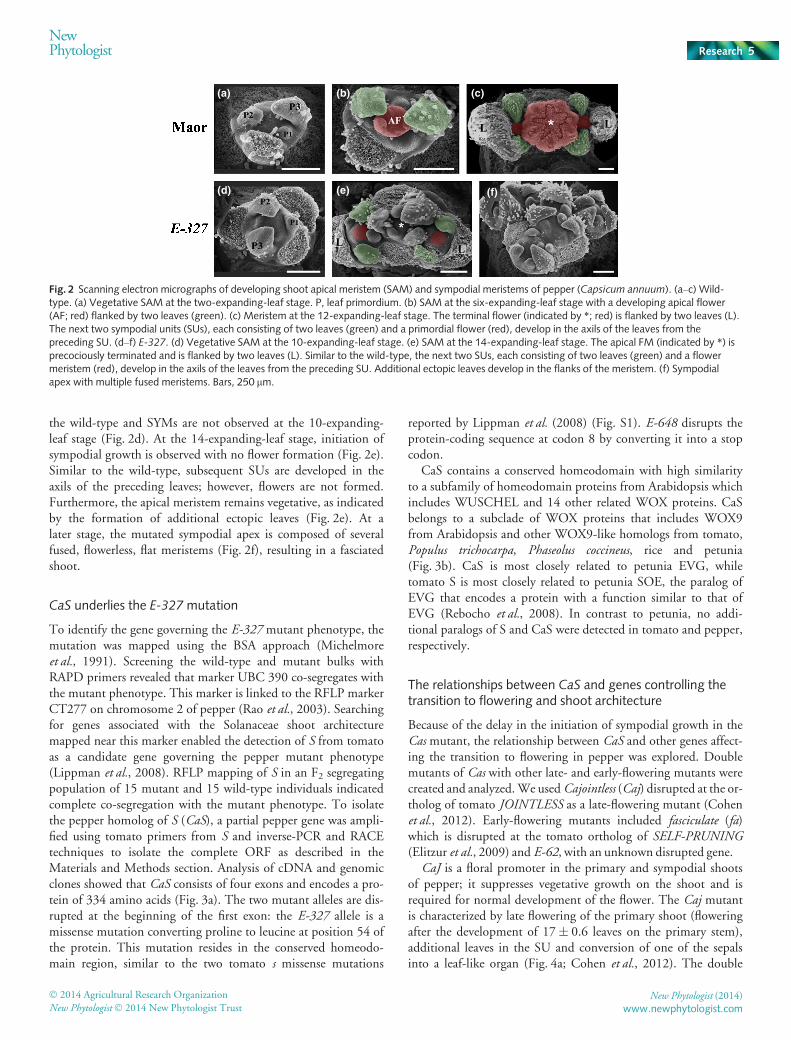
the wild-type and SYMs are not observed at the 10-expanding-leaf stage (Fig. 2d). At the 14-expanding-leaf stage, initiation ofsympodial growth is observed with no flower formation (Fig. 2e).Similar to the wild-type, subsequent SUs are developed in theaxils of the preceding leaves; however, flowers are not formed.Furthermore, the apical meristem remains vegetative, as indicatedby the formation of additional ectopic leaves (Fig. 2e). At alater stage, the mutated sympodial apex is composed of severalfused, flowerless, flat meristems (Fig. 2f), resulting in a fasciatedshoot.
CaS underlies the E-327mutation
To identify the gene governing the E-327 mutant phenotype, themutation was mapped using the BSA approach (Michelmoreet al., 1991). Screening the wild-type and mutant bulks withRAPD primers revealed that marker UBC 390 co-segregates withthe mutant phenotype. This marker is linked to the RFLP markerCT277 on chromosome 2 of pepper (Rao et al., 2003). Searchingfor genes associated with the Solanaceae shoot architecturemapped near this marker enabled the detection of S from tomatoas a candidate gene governing the pepper mutant phenotype(Lippman et al., 2008). RFLP mapping of S in an F2 segregatingpopulation of 15 mutant and 15 wild-type individuals indicatedcomplete co-segregation with the mutant phenotype. To isolatethe pepper homolog of S (CaS), a partial pepper gene was ampli-fied using tomato primers from S and inverse-PCR and RACEtechniques to isolate the complete ORF as described in theMaterials and Methods section. Analysis of cDNA and genomicclones showed that CaS consists of four exons and encodes a pro-tein of 334 amino acids (Fig. 3a). The two mutant alleles are dis-rupted at the beginning of the first exon: the E-327 allele is amissense mutation converting proline to leucine at position 54 ofthe protein. This mutation resides in the conserved homeodo-main region, similar to the two tomato s missense mutations
reported by Lippman et al. (2008) (Fig. S1). E-648 disrupts theprotein-coding sequence at codon 8 by converting it into a stopcodon.
CaS contains a conserved homeodomain with high similarityto a subfamily of homeodomain proteins from Arabidopsis whichincludes WUSCHEL and 14 other related WOX proteins. CaSbelongs to a subclade of WOX proteins that includes WOX9from Arabidopsis and other WOX9-like homologs from tomato,Populus trichocarpa, Phaseolus coccineus, rice and petunia(Fig. 3b). CaS is most closely related to petunia EVG, whiletomato S is most closely related to petunia SOE, the paralog ofEVG that encodes a protein with a function similar to that ofEVG (Rebocho et al., 2008). In contrast to petunia, no addi-tional paralogs of S and CaS were detected in tomato and pepper,respectively.
The relationships between CaS and genes controlling thetransition to flowering and shoot architecture
Because of the delay in the initiation of sympodial growth in theCas mutant, the relationship between CaS and other genes affect-ing the transition to flowering in pepper was explored. Doublemutants of Cas with other late- and early-flowering mutants werecreated and analyzed.We usedCajointless (Caj) disrupted at the or-tholog of tomato JOINTLESS as a late-flowering mutant (Cohenet al., 2012). Early-flowering mutants included fasciculate (fa)which is disrupted at the tomato ortholog of SELF-PRUNING(Elitzur et al., 2009) and E-62, with an unknown disrupted gene.
CaJ is a floral promoter in the primary and sympodial shootsof pepper; it suppresses vegetative growth on the shoot and isrequired for normal development of the flower. The Caj mutantis characterized by late flowering of the primary shoot (floweringafter the development of 17� 0.6 leaves on the primary stem),additional leaves in the SU and conversion of one of the sepalsinto a leaf-like organ (Fig. 4a; Cohen et al., 2012). The double
(a) (b) (c)
(d) (e) (f)
Fig. 2 Scanning electron micrographs of developing shoot apical meristem (SAM) and sympodial meristems of pepper (Capsicum annuum). (a–c) Wild-type. (a) Vegetative SAM at the two-expanding-leaf stage. P, leaf primordium. (b) SAM at the six-expanding-leaf stage with a developing apical flower(AF; red) flanked by two leaves (green). (c) Meristem at the 12-expanding-leaf stage. The terminal flower (indicated by *; red) is flanked by two leaves (L).The next two sympodial units (SUs), each consisting of two leaves (green) and a primordial flower (red), develop in the axils of the leaves from thepreceding SU. (d–f) E-327. (d) Vegetative SAM at the 10-expanding-leaf stage. (e) SAM at the 14-expanding-leaf stage. The apical FM (indicated by *) isprecociously terminated and is flanked by two leaves (L). Similar to the wild-type, the next two SUs, each consisting of two leaves (green) and a flowermeristem (red), develop in the axils of the leaves from the preceding SU. Additional ectopic leaves develop in the flanks of the meristem. (f) Sympodialapex with multiple fused meristems. Bars, 250 lm.
� 2014 Agricultural Research Organization
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014)
www.newphytologist.com
NewPhytologist Research 5

mutant Cas Caj (Fig. 4b) has an overall phenotype that resemblesthat of Cas (Fig. 4c), but initiation of sympodial growth occursvery late, after the development of 61.5� 1.5 leaves on theprimary stem, and it lacks flowers. These phenotypes suggest thatCas is epistatic over Caj with respect to flower formation, whileCas and Caj act in a positive synergistic manner with respect toinitiation of sympodial growth (i.e. the effect of the doublemutant, measured as the difference between the double mutantand the wild-type, is larger than the combined effects of the indi-vidual mutants). Interestingly, heterozygous plants at the CaSlocus in a Caj background initiate transition to sympodial growthvery late, after the development of 47.5� 2.5 leaves on theprimary stem, but, unlike Cas, they do form intact flowers, fur-ther indicating an interaction between the two genes.
FA is a floral repressor in the primary and sympodial shoots ofpepper. The fa mutant is characterized by a reduction of flower-ing time (8.6� 0.5 leaves developing before the first flower), and
shortening of the SUs (Elitzur et al., 2009). These phenotypesresult in a compact ‘determinate’ plant carrying condensed clus-ters of flowers. Double-mutant Cas fa plants are indistinguishablefrom Cas plants; that is, they initiate late transition to sympodialgrowth after the development of 20� 1.5 leaves and completelylack flowers, indicating epistasis of Cas over fa.
E-62 is a new monogenic recessive early-flowering mutant thatwas identified in the EMS-mutagenized pepper population(Paran et al., 2007). E-62 flowers after four leaves have developedon the primary stem, while no change in flowering pattern andflower structure is observed in the sympodial shoot (Fig. 4d). Thedouble mutant Cas E-62 has the overall phenotype of Cas but itstransition to sympodial growth occurs after the development of17.7� 1.5 leaves on the primary stem, an intermediate pheno-type between those of the two single mutants, and it completelylacks flowers (Fig. 4e). These phenotypic characteristics suggestthat, similar to the relationship with other mutants, Cas is epi-static over E-62 with respect to flower formation. However, theintermediate phenotype of transition to sympodial growth of CasE-62 suggests an additive effect of the two genes with respect toinitiation of sympodial growth (i.e. the effect of the doublemutant, measured as the difference between the double mutantand the wild-type, is equal to the combined effects of the individ-ual mutants).
The relationships between CaS and FM identity genes
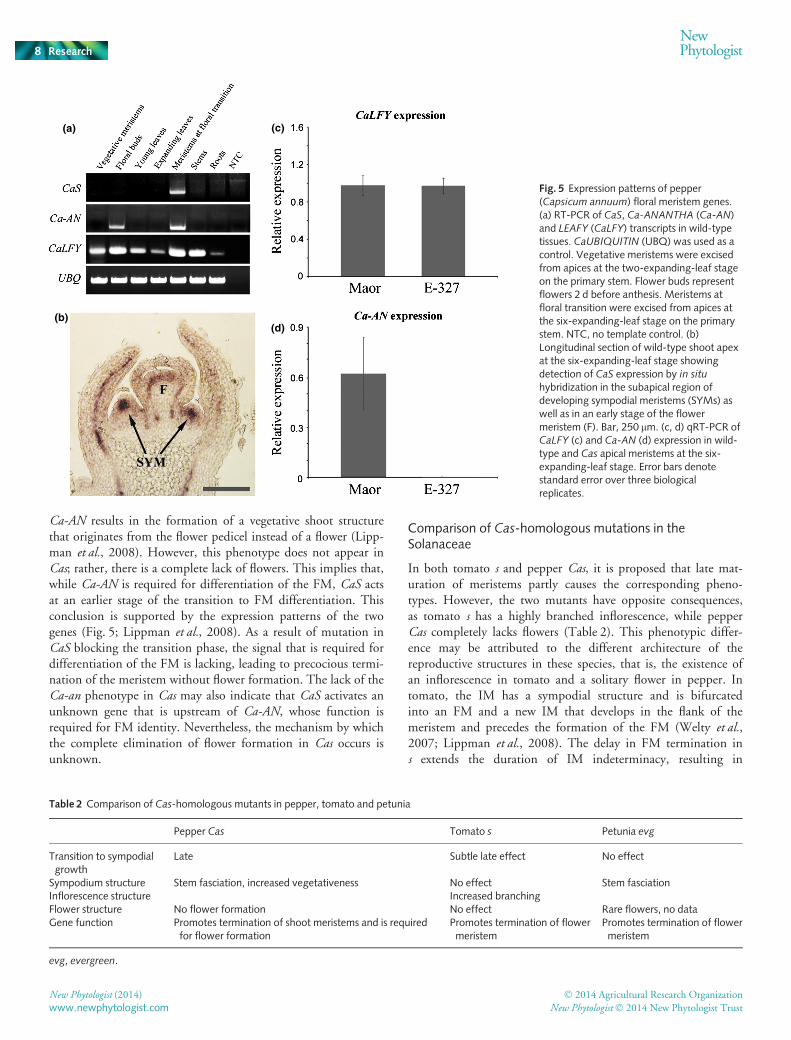
To better understand the genetic hierarchy that governs flowerformation in pepper, the expression patterns of CaS and the FMidentity genes CaLFY and Ca-AN were analyzed in the wild-type.Expression of CaS was detected at apical meristems after floraltransition but not in vegetative meristems or in more developedflower buds (Fig. 5a). CaS expression was further localized byin situ hybridization to the inner, central cells of the SYM beforedifferentiation into a flower primordium (Fig. 5b). Lowexpression was also observed in an early stage of the FM (Fig. 5b).Ca-AN expression was detected in both apical meristems at floraltransition and at flower buds of wild-type plants, indicating par-tial overlap in the timing of expression of the two genes (Fig. 5a).CaLFY transcripts were detected at various levels in all examinedtissues (Fig. 5a). The expression levels of CaLFY and CaJ werenot altered in the apical and sympodial meristems of the Casmutant compared with the wild-type (Figs 5c, S2), whereasexpression of Ca-AN was not detected in the Cas mutant in thesemeristems (Figs 5d, S2). These results suggest that CaS does notaffect CaLFY expression and that the two genes probably actindependently with respect to FM differentiation, while CaS isrequired for the expression of Ca-AN and is located upstream ofit in the pepper flower formation pathway.
Discussion
In the present study, the identification of CaS which controlsmeristem termination in pepper is reported. CaS functions inpromoting the reproductive shift of the apical and sympodialshoots and is required for flower formation.
(a)
(b)
Fig. 3 Gene structure and phylogeny of pepper (Capsicum annuum) CaS.(a) Schematic representation of CaS structure. Introns and exons areindicated by solid lines and white boxes, respectively. Regions encodingthe conserved Wuschel-homeobox (WOX)-homeodomain in exons 1 and2 are indicated by black boxes and numbers indicate the homeodomainposition on the gene sequence. Asterisks indicate the position of themutations in E-327 and E-648. (b) Phylogenetic tree of CaS and itshomologs from tomato, petunia, rice, Phaseolus coccineus, Populustrichocarpa, Antirrhinum majus and Arabidopsis. Numbers indicatepercentage bootstrap support for each branch (1000 replicates).
New Phytologist (2014) � 2014 Agricultural Research Organization
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist6

Role of CaS in controlling the transition to sympodialgrowth
While initiation of sympodial growth marks the transition toflowering and flower formation in wild-type pepper, initiation ofsympodial growth in Cas is associated with formation of a dicha-sially forked shoot, stem fasciation and lack of flowers. Casmutants display four major pleiotropic effects: delay in termina-tion of the apical meristem; delay in termination of sympodialmeristems; fasciated sympodial shoot; and loss of flower forma-tion. These phenotypes, as well as the expression of CaS in theapical meristem at the transition to flowering and in the SYMs(Fig. 5a,b), suggest that its primary role is to promote termina-tion of apical and sympodial meristems. According to Park et al.(2012), the rate of meristem maturation determines inflorescencearchitecture in tomato and in other Solanaceae. The delay in thetermination of apical and sympodial meristems in Cas results indelayed transition of the primary shoot to sympodial growth andincreased vegetativeness of the meristems to a greater degree thanin other known late-flowering pepper mutants (Jeifetz et al.,2011; Cohen et al., 2012). The fasciated shoot of Cas is a conse-quence of fusion of the SYMs, which may result because theflower that provides a physical barrier between adjacent SUs islacking. Release of stem fasciation occurs after formation of a fewsubsequent SYMs, possibly because of physical constraintsimposed on the apex organization.
Double-mutant analyses of CaS with other genes controllingthe transition to flowering showed complex patterns of relation-ships. CaS interacts in a positive synergistic manner with theflowering promoter CaJ, while it is epistatic to the flowering
repressor FA and acts additively to the flowering repressor E-62.These relationships imply that the activities of CaS, CaJ and FAconverge at some point in the sympodial growth transition path-way, while the gene disrupted in E-62 acts independently to CaS.Because the expression of CaJ is not affected by the Cas mutation(Fig. S2), CaJ is probably upstream of CaS.
Role of CaS in controlling flower formation
In wild-type pepper, termination of the apical and sympodialmeristems is associated with the transition to flowering of theprimary and sympodial shoot, and, for both meristems, this tran-sition is associated with flower formation. In Cas, termination ofboth apical and sympodial meristems is delayed, as manifested bya delay in the initiation of sympodial growth and increased vege-tativeness of the SU. The failure to produce flowers in Cas is nota consequence of the delay in meristem termination, as, in theCas E-62 double mutant, transition to sympodial growth isenhanced relative to Cas as a result of a mutation in the floweringrepressor gene of E-62; however, flowers are not formed.
In petunia, expression of DOT is downstream of EVG (Rebo-cho et al., 2008) and is required for the formation of flowers(Souer et al., 2008). Ectopic expression of DOT in evg restoresflower formation. Therefore, inhibition of flower formation inevg is interpreted as resulting from loss of DOT expression. Intomato, expression of S precedes that of AN, and the two genespromote successive stages in inflorescence and flower develop-ment (Lippman et al., 2008). Similar to petunia and tomato,expression of Ca-AN, the pepper ortholog of DOT and AN,is dependent on the activity of CaS (Figs 5d, S2). Mutation in
(a) (b) (c)
(d) (e)
Fig. 4 Pepper (Capsicum annuum) mutantsaffected in flowering time and shootarchitecture and their interaction with Cas.(a) Cajointless (Caj) exhibits late floweringcompared with ‘Maor’. (b) The Cas Cajdouble mutant exhibits extreme inhibition ofinitiation of sympodial growth. (c) E-327exhibits late transition to sympodial growthand lack of flowers. (d) E-62 exhibits earlytransition to flowering compared with‘Maor’. (e) Cas E-62 double mutant showingintermediate transition to sympodial growthand lack of flowers.
� 2014 Agricultural Research Organization
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014)
www.newphytologist.com
NewPhytologist Research 7
Ca-AN results in the formation of a vegetative shoot structurethat originates from the flower pedicel instead of a flower (Lipp-man et al., 2008). However, this phenotype does not appear inCas; rather, there is a complete lack of flowers. This implies that,while Ca-AN is required for differentiation of the FM, CaS actsat an earlier stage of the transition to FM differentiation. Thisconclusion is supported by the expression patterns of the twogenes (Fig. 5; Lippman et al., 2008). As a result of mutation inCaS blocking the transition phase, the signal that is required fordifferentiation of the FM is lacking, leading to precocious termi-nation of the meristem without flower formation. The lack of theCa-an phenotype in Cas may also indicate that CaS activates anunknown gene that is upstream of Ca-AN, whose function isrequired for FM identity. Nevertheless, the mechanism by whichthe complete elimination of flower formation in Cas occurs isunknown.
Comparison of Cas-homologous mutations in theSolanaceae
In both tomato s and pepper Cas, it is proposed that late mat-uration of meristems partly causes the corresponding pheno-types. However, the two mutants have opposite consequences,as tomato s has a highly branched inflorescence, while pepperCas completely lacks flowers (Table 2). This phenotypic differ-ence may be attributed to the different architecture of thereproductive structures in these species, that is, the existence ofan inflorescence in tomato and a solitary flower in pepper. Intomato, the IM has a sympodial structure and is bifurcatedinto an FM and a new IM that develops in the flank of themeristem and precedes the formation of the FM (Welty et al.,2007; Lippman et al., 2008). The delay in FM termination ins extends the duration of IM indeterminacy, resulting in
(a) (c)
(b)(d)
Fig. 5 Expression patterns of pepper(Capsicum annuum) floral meristem genes.(a) RT-PCR of CaS, Ca-ANANTHA (Ca-AN)and LEAFY (CaLFY) transcripts in wild-typetissues. CaUBIQUITIN (UBQ) was used as acontrol. Vegetative meristems were excisedfrom apices at the two-expanding-leaf stageon the primary stem. Flower buds representflowers 2 d before anthesis. Meristems atfloral transition were excised from apices atthe six-expanding-leaf stage on the primarystem. NTC, no template control. (b)Longitudinal section of wild-type shoot apexat the six-expanding-leaf stage showingdetection of CaS expression by in situhybridization in the subapical region ofdeveloping sympodial meristems (SYMs) aswell as in an early stage of the flowermeristem (F). Bar, 250 lm. (c, d) qRT-PCR ofCaLFY (c) and Ca-AN (d) expression in wild-type and Cas apical meristems at the six-expanding-leaf stage. Error bars denotestandard error over three biologicalreplicates.
Table 2 Comparison of Cas-homologous mutants in pepper, tomato and petunia
Pepper Cas Tomato s Petunia evg
Transition to sympodialgrowth
Late Subtle late effect No effect
Sympodium structure Stem fasciation, increased vegetativeness No effect Stem fasciationInflorescence structure Increased branchingFlower structure No flower formation No effect Rare flowers, no dataGene function Promotes termination of shoot meristems and is required
for flower formationPromotes termination of flowermeristem
Promotes termination of flowermeristem
evg, evergreen.
New Phytologist (2014) � 2014 Agricultural Research Organization
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist8

increased branching. In pepper, the mutation in Cas results indelayed termination of the shoot meristems. However, the lackof IM does not provide a developmental window for branch-ing. Interestingly, while in pepper Cas, termination of all shootmeristems is coordinately delayed or blocked, in tomato s,delay in meristem termination is mostly restricted to the FMas only a subtle change is observed in the timing of the transi-tion to flowering (Quinet et al., 2006a) and no change isobserved in the sympodial shoot structure.
The Cas phenotype more closely resembles that of petunia evg(Rebocho et al., 2008), which is characterized by a bushy vegeta-tive inflorescence that lacks flowers. However, compared withpepper Cas, which exhibits a delayed transition to sympodialgrowth and prevention of flower formation, evg switches to theflowering phase (as indicated by the appearance of bracts in theinflorescence nodes) at the same time as the wild-type and occa-sionally produces flowers (Table 2). Furthermore, similar to thefasciated shoot of Cas, the inflorescence stems in evg often fail tobifurcate into flower and inflorescence meristems as occurs in thewild-type, resulting in a fasciated shoot. Therefore, shoot fascia-tion and lack of flowers are common to both Cas and evg,although these characteristics are more strongly expressed inpepper. Furthermore, while S and EVG act predominantly in theIM and FM, CaS has a wider function in shoot meristems. Thesmaller phylogenetic distance between CaS and EVG thanbetween CaS and S further supports the greater phenotypicresemblance between the pepper and petunia mutants thanbetween the pepper and tomato ones.
An additional factor that may affect diversification of genefunction in different species is differential patterns of geneinteractions. CaS homologs exhibit different patterns of inter-actions in pepper compared with tomato and petunia. Inpepper, all tested double mutants involving Cas fail toundergo floral formation; that is, Cas is epistatic to othermutations in genes controlling sympodial development inaffecting flower formation. By contrast, in tomato, the solitaryflower mutation uniflora, as well as jointless, are epistatic to s(Quinet et al., 2006b; Thouet et al., 2012). Similarly, in petu-nia, the mutations extrapetals (exp) and hermit (her), having asingle flower inflorescence and arrest of sympodial growth, areepistatic to evg, as double mutants with evg show the singleflower phenotype (Rebocho et al., 2008). EXP is the homologof CaJ (Table S2; Castel, 2009), while mutations in pepperhomologous to uniflora and her have not been reported.Therefore, in contrast to petunia, in which the requirementof EVG for flower formation can be bypassed by altering theinflorescence architecture, this requirement of CaS is manda-tory. This indicates that, while S and EVG have a primaryrole in specifying FM maturation (Park et al., 2014), CaS hasevolved to have a critical role in flower formation in additionto its role in controlling meristem maturation. Furthermore,the opposite epistatic effects of Cas Caj homologous mutantsin pepper compared with tomato and petunia indicatethat different modes of gene interaction exist that may con-tribute to diversification of sympodial development in theSolanaceae.
Acknowledgements
We thank Saadia Nahon for technical support, Arnon Brand forgraphic design and Hanita Zemach for assistance with micro-scopic analyses. We thank Zach Lippman (Cold Spring HarborLaboratory) for fruitful discussions and critical reading of themanuscript, and Prof. Dani Zamir (Hebrew University) for PhDguidance of O.C. This research was supported by The IsraelScience Foundation (grant no. 1349/10).
References
Alvarez J, Guli CL, Yu XH, Smyth DR. 1992. TERMINAL FLOWER: a geneaffecting inflorescence development in Arabidopsis thaliana. Plant Journal 2:103–116.
Angenent GC, Stuurman J, Snowden KC, Koes R. 2005. Use of Petunia to
unravel plant meristem functioning. Trends in Plant Science 10: 243–250.Bradley D, Ratcliffe O, Vincent C, Carpenter R, Coen E. 1997. Inflorescence
commitment and architecture in Arabidopsis. Science 275: 80–83.Castel R. 2009.Molecular analysis of the developmental mechanisms that establishthe body plan of petunia. PhD thesis, Vrije Universiteit, Amsterdam, the
Netherlands.
Castel R, Kusters E, Koes R. 2010. Inflorescence development in petunia:
through the maze of botanical terminology. Journal of Experimental Botany 61:2235–2246.
Child A. 1979. A review of branching patterns in the Solanaceae. In: Hawkes JG,
Lester RN, Skelding AD, eds. The biology and taxonomy of the Solanaceae.London, UK: Academic Press, 345–356.
Cohen O, Borovsky Y, David-Schwartz R, Paran I. 2012. CaJOINTLESS is aMADS-box gene involved in suppression of vegetative growth in all shoot
meristems in pepper. Journal of Experimental Botany 63: 4947–4957.David-Schwartz R, Borovsky Y, Zemach H, Paran I. 2013. CaHAM is
autoregulated and regulates CaSTM expression and is required for shoot apical
meristem organization in pepper. Plant Science 203–204: 8–16.Dielen V, Quinet M, Chao J, Batoko H, Havelange A, Kinet J-M. 2004.
UNIFLORA, a pivotal gene that regulates floral transition and meristem
identity in tomato (Lycopersicon esculentumMill.). New Phytologist 161: 393–400.
Elitzur T, Nahum H, Borovsky Y, Pekker I, Eshed Y, Paran I. 2009.
Co-ordinated regulation of flowering time, plant architecture and growth by
FASCICULATE: the pepper orthologue of SELF PRUNING. Journal ofExperimental Botany 60: 869–880.
Grandi V, Gregis V, Kater MM. 2012. Uncovering genetic and molecular
interactions among floral meristem identity genes in Arabidopsis thaliana. PlantJournal 69: 881–893.
Huijser P, Schmid M. 2011. The control of developmental phase transitions in
plants. Development 138: 4117–4129.Jeifetz D, David-Schwartz R, Borovsky Y, Paran I. 2011. CaBLIND regulates
axillary meristem initiation and transition to flowering in pepper. Planta 234:1227–1236.
Lifschitz E, Eviatar T, Rozman A, Shalit A, Goldshmidt A, Amsellem Z, Alvarez
JP, Eshed Y. 2006. The tomato FT ortholog triggers systemic signals that
regulate growth and flowering and substitute for diverse environmental stimuli.
Proceedings of the National Academy of Sciences, USA 103: 6398–6403.Lippman ZB, Cohen O, Alvarez JP, Abu-Abied M, Pekker I, Paran I, Eshed Y,
Zamir D. 2008. The making of a compound inflorescence in tomato and
related nightshades. PLoS Biology 6: e288.Lozano R, Gim�enez E, Cara B, Capel J, Angosto T. 2009. Genetic analysis of
reproductive development in tomato. International Journal of DevelopmentalBiology 53: 1635–1648.
Michelmore RW, Paran I, Kesseli RV. 1991. Identification of markers linked to
disease-resistance genes by bulked segregant analysis: a rapid method to detect
markers in specific genomic regions by using segregating populations.
Proceedings of the National Academy of Sciences, USA 88: 9828–9832.
� 2014 Agricultural Research Organization
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014)
www.newphytologist.com
NewPhytologist Research 9

Moyroud E, Kusters E, Monniaux M, Koes R, Parcy F. 2010. LEAFY blossoms.
Trends in Plant Science 15: 346–352.Neta R, David-Schwartz R, Peretz Y, Sela I, Rabinowitch HD, Flaishman M,
Kamenetsky R. 2011. Flower development in garlic: the ups and downs of
gaLFY expression. Planta 233: 1063–1072.Ochman H, Gerber AS, Hartl DL. 1988. Genetic applications of an inverse
polymerase chain reaction. Genetics 120: 621–623.Paran I, Borovsky Y,Nahon S, CohenO. 2007.The use of inducedmutations to
study shoot architecture inCapsicum. Israel Journal of Plant Sciences 55: 125–131.Park SJ, Eshed Y, Lippman ZB. 2014.Meristem maturation and inflorescence
architecture – lessons from the Solanaceae. Current Opinion in Plant Biology 17:70–77.
Park SJ, Jiang K, Schatz MC, Lippman ZB. 2012. Rate of meristem maturation
determines inflorescence architecture in tomato. Proceedings of the NationalAcademy of Sciences, USA 109: 639–644.
Prusinkiewicz P, Erasmus Y, Lane B, Harder LD, Coen E. 2007. Evolution and
development of inflorescence architectures. Science 316: 1452–1456.Quinet M, Dielen V, Batoko H, Boutry M, Havelange A, Kinet J-M. 2006b.
Genetic interactions in the control of flowering time and reproductive
structure development in tomato (Solanum lycopersicum). New Phytologist 170:701–710.
Quinet M, Dubois C, Goffin M-C, Chao J, Dielen V, Batoko H, Boutry M,
Kinet J-M. 2006a. Characterization of tomato (Solanum lycopersicum L.)
mutants affected in their flowering time and in the morphogenesis of their
reproductive structure. Journal of Experimental Botany 57: 1381–1390.Rao GU, Ben-Chaim A, Borovsky Y, Paran I. 2003.Mapping of yield-related
QTLs in pepper in an interspecific cross of Capsicum annuum and C. frutescens.Theoretical and Applied Genetics 106: 1457–1466.
Rebocho AB, Bliek M, Kusters E, Castel R, Procissi A, Roobeek I, Souer E,
Koes R. 2008. Role of EVERGREEN in the development of the cymose
petunia inflorescence. Developmental Cell 15: 437–447.Schmitz G, Theres K. 1999. Genetic control of branching in Arabidopsis andtomato. Current Opinion in Plant Biology 2: 51–55.
Souer E, Rebocho AB, Bliek M, Kusters E, De Bruin RAM, Koes R. 2008.
Patterning of inflorescences and flowers by the F-Box protein DOUBLE TOP
and the LEAFY homolog ABERRANT LEAF AND FLOWER of petunia. ThePlant Cell 20: 2033–2048.
Srikanth A, Schmid M. 2011. Regulation of flowering time: all roads lead to
Rome. Cellular and Molecular Life Sciences 68: 2013–2037.Thouet J, Quinet M, Lutts S, Kinet JM, Perilleux C. 2012. Repression of floral
meristem fate is crucial in shaping tomato inflorescence. PLoS ONE 7: e31096.
Welty N, Radovich C, Meulia T, van der Knaap E. 2007. Inflorescence
development in two tomato species. Canadian Journal of Botany 85: 111–118.Winter CM, Austin RS, Blanvillain-Baufum�e S, Reback MA, Monniaux M,
Wu M-F, Sang Y, Yamaguchi A, Yamaguchi N, Parker JE et al. 2011. LEAFYtarget genes reveal floral regulatory logic, cis motifs, and a link to biotic
stimulus response. Developmental Cell 20: 430–443.
Supporting Information
Additional supporting information may be found in the onlineversion of this article.
Fig. S1Multiple sequence alignment of CaS homologs in pepper,tomato and petunia.
Fig. S2 Expression of pepper flowering-related genes in SYMs of‘Maor’ and E-327.
Table S1 Pepper primers used in this study
Table S2 Comparison of Caj homologous mutants in pepper,tomato and petunia
Please note: Wiley Blackwell are not responsible for the contentor functionality of any supporting information supplied by theauthors. Any queries (other than missing material) should bedirected to the New Phytologist Central Office.
New Phytologist (2014) � 2014 Agricultural Research Organization
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist10
Related Documents











