Calcium : outside/inside homeostasis and signalling R.J.P. Williams * Inorganic Chemistry Laboratory, Oxford University, South Parks Road, Oxford OX1 3QR, UK Accepted 13 October 1998 Abstract More and more data accumulate concerning calcium dependent effects in all compartments of cells. The higher the organism in evolution the more calcium becomes involved. Inspection of the data while looking for an explanation of the involvement of calcium in metabolism, nuclear functions, control over pumps, external activities, mineralisation and so on leads one to suppose that calcium has an integrating function. The implication is that calcium flow is a large network connecting the environment, the cytoplasm, vesicles, organelles, the nucleus and in higher species, organs. There is the possibility then that calcium ion functions are being analysed, often in vitro, in a bit by bit reductionist manner while in vivo calcium is the equivalent of an electron in complicated electrical circuits. We then should look for its connections to energy, to effects where conformational switching by calcium pulses is equivalent to magnetic triggering by electron flow and where storage in vesicles is equivalent to condenser-like devices and so on. The appearance of pulsing, of time delays in parts of circuits, and other properties of electronic circuits seen in calcium triggering are then explicable as part of calcium circuit design. No other ion can operate in the same way due to the peculiarities of the calcium ion, its size, charge, ionisation potential and its availability which allow it both to flow rapidly yet to bind considerably. ß 1998 Elsevier Science B.V. All rights reserved. Keywords : Homeostasis ; Calcium circuit ; Compartment ; Bio-energetics ; Environment ; Cell control 1. Introduction In my last lecture at the International Meeting on Calcium Proteins, in Lund in 1997, [1] I stressed features of calcium activity which were linked to the activity of a range of other elements including Na ,K , Mg 2 , Mn 2 , Fe 2 and Zn 2 among metal ions and phosphate especially amongst non-metals. I stressed that these connections are part of a general link in all eukaryotes between the activities of the cytoplasm of a cell, of the external environment and of the content of organelles and vesicles, which were all related to calcium levels. The further sugges- tion was that much of homeostasis of a multi-cellular organism is also monitored by calcium controls. In this respect calcium is the dominant messenger for triggering, which crosses membranes of all kinds, while it is also a homeostatic control for both the intracellular and extracellular £uids of higher multi- cellular organisms. Its activity di¡ers greatly from the more primitive messenger uses of hydrogen ions, substrates, phosphate and iron compounds which already in prokaryotes act as the major ho- meostatic controls and triggers of the cytoplasm alone. However, the innovation of the use of calcium in external and vesicle communications with the cy- 0167-4889 / 98 / $ ^ see front matter ß 1998 Elsevier Science B.V. All rights reserved. PII:S0167-4889(98)00139-6 * Corresponding author. Fax: +44 (1865) 272690; E-mail : [email protected] Biochimica et Biophysica Acta 1448 (1998) 153^165

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Calcium: outside/inside homeostasis and signalling
R.J.P. Williams *Inorganic Chemistry Laboratory, Oxford University, South Parks Road, Oxford OX1 3QR, UK
Accepted 13 October 1998
Abstract
More and more data accumulate concerning calcium dependent effects in all compartments of cells. The higher theorganism in evolution the more calcium becomes involved. Inspection of the data while looking for an explanation of theinvolvement of calcium in metabolism, nuclear functions, control over pumps, external activities, mineralisation and so onleads one to suppose that calcium has an integrating function. The implication is that calcium flow is a large networkconnecting the environment, the cytoplasm, vesicles, organelles, the nucleus and in higher species, organs. There is thepossibility then that calcium ion functions are being analysed, often in vitro, in a bit by bit reductionist manner while in vivocalcium is the equivalent of an electron in complicated electrical circuits. We then should look for its connections to energy,to effects where conformational switching by calcium pulses is equivalent to magnetic triggering by electron flow and wherestorage in vesicles is equivalent to condenser-like devices and so on. The appearance of pulsing, of time delays in parts ofcircuits, and other properties of electronic circuits seen in calcium triggering are then explicable as part of calcium circuitdesign. No other ion can operate in the same way due to the peculiarities of the calcium ion, its size, charge, ionisationpotential and its availability which allow it both to flow rapidly yet to bind considerably. ß 1998 Elsevier Science B.V. Allrights reserved.
Keywords: Homeostasis ; Calcium circuit ; Compartment; Bio-energetics ; Environment; Cell control
1. Introduction
In my last lecture at the International Meeting onCalcium Proteins, in Lund in 1997, [1] I stressedfeatures of calcium activity which were linked tothe activity of a range of other elements includingNa�, K�, Mg2�, Mn2�, Fe2� and Zn2� among metalions and phosphate especially amongst non-metals. Istressed that these connections are part of a generallink in all eukaryotes between the activities of thecytoplasm of a cell, of the external environment
and of the content of organelles and vesicles, whichwere all related to calcium levels. The further sugges-tion was that much of homeostasis of a multi-cellularorganism is also monitored by calcium controls. Inthis respect calcium is the dominant messenger fortriggering, which crosses membranes of all kinds,while it is also a homeostatic control for both theintracellular and extracellular £uids of higher multi-cellular organisms. Its activity di¡ers greatly fromthe more primitive messenger uses of hydrogenions, substrates, phosphate and iron compoundswhich already in prokaryotes act as the major ho-meostatic controls and triggers of the cytoplasmalone. However, the innovation of the use of calciumin external and vesicle communications with the cy-
0167-4889 / 98 / $ ^ see front matter ß 1998 Elsevier Science B.V. All rights reserved.PII: S 0 1 6 7 - 4 8 8 9 ( 9 8 ) 0 0 1 3 9 - 6
* Corresponding author. Fax: +44 (1865) 272690;E-mail : [email protected]
BBAMCR 14407 3-12-98
Biochimica et Biophysica Acta 1448 (1998) 153^165

toplasm of eukaryotes had to be married to the useof these internal earlier messenger systems of primi-tive cells since much of eukaryote metabolism is car-ried over from prokaryotes. We therefore found across communication network of messenger systemswhich employ calcium, hydrogen, substrates, phos-phates and iron compounds in the cytoplasm andin the interior of organelles of eukaryote cells. Inthis article I shall continue this theme of calciumfunctions connected to those of many other elementsbut at the same time I shall look at energy use anddistribution often involving ATP and where calciumis involved. I shall be stressing that from soon afterthe time of the appearance of eukaryotes the role ofcalcium was in the management of cell homeostasisthrough continuous energy production and distribu-tion as well as that of material transfer and metabo-lism, while acting as a trigger messenger across allmembranes when required. This £ow of energy is anessential part of homeostasis. There is then an evolu-tionary progression in the use of calcium in a seriesof add-on steps:
1. External use in outer cell walls and membranes ofthe most primitive prokaryotes
2. Messenger for protection and external responsesof single cells of early eukaryotes generating con-traction and similar responses, through binding tointernal matrix ¢bres, e.g. actinomyosin
3. Internal energy homeostasis in single cells of earlyeukaryotes linking organelles, vesicles and the cy-toplasm
4. External structural and messenger use for interac-tions between cell connectivity of multi-cellularorganisms
I dismiss (1) the use of calcium in primitive pro-karyotes, at least where it acts merely as a protectiveexternal cross linking agent, as of little in conse-quence in this article. Undoubtedly today some pro-karyotes can use calcium gradients in message sys-tems too but the constraint in a prokaryote is that itmust multiply rapidly rather than constantly adjust-ing itself to environmental stimuli so that even herethe use of calcium is not very extensive. Growth ismerely a doubling in size before division. It may alsobe that calcium was used early in sporulation againin a protective salt. Prokaryote internal metabolism
is relatively £exible much though it is connected tohydrogen, substrate, phosphate and iron controls.This can be seen by looking at those cytoplasmicmetabolic paths common to all organisms and in-cluding some, as in the following example, whichremain largely independent of calcium to this dayeven in advanced eukaryotes.
Energy directly from sugars without oxidation isgained in the well-known glycolysis pathways by pro-karyotes and eukaryotes alike. Here the products ofglucose internal disproportionation are pyruvate andthen acetate: NAD+(CHOH)nCCH3.CO.CO2H(C3H6O3)+NADHCCH3CO2H (C2H4O2) acetate.The reaction is a special case of energy productionand of the general control of internal metabolic path-ways of all cells by catalysts not linked to external orinternal conditions by calcium messages. In prokary-otes the control by feedback not only of this path-way, but its links to fat production and degradation,to synthesis of amino acids and nucleotide bases andits connection to the citric acid cycle are all managedby the levels of hydrogen, substrate, phosphate andiron compounds without calcium. We need ¢rst thebackground to this control activity linked to energybefore we consider the role of calcium in eukaryotes.
2. Homeostasis
A cell is characterised by activities encoded byDNA which is read by a machinery and then tran-scribed and translated. The activity, including dy-namic structures, is seen in properties of a large num-ber of large and small molecules which can beexamined separately in vitro but in vivo their proper-ties are all related (Fig. 1), so that the activity formsa pattern of £ow of material and energy. The ¢xedpattern in steady state, which is present both in phys-ical form (of all materials) and energy £ows, impliesthat in the steady state all chemicals in the cell are in¢xed ratios. The maintenance of this composition iscalled homeostasis and since the cell is always activein many ways it needs many controls but at the sametime all the activities have to be integrated. Thus itspossible variables need to be controlled by coordinat-ing messengers of all activities of the cell.
Now we need to de¢ne more clearly what are thevariables which have to be controlled. Chemicals are
BBAMCR 14407 3-12-98
R.J.P. Williams / Biochimica et Biophysica Acta 1448 (1998) 153^165154

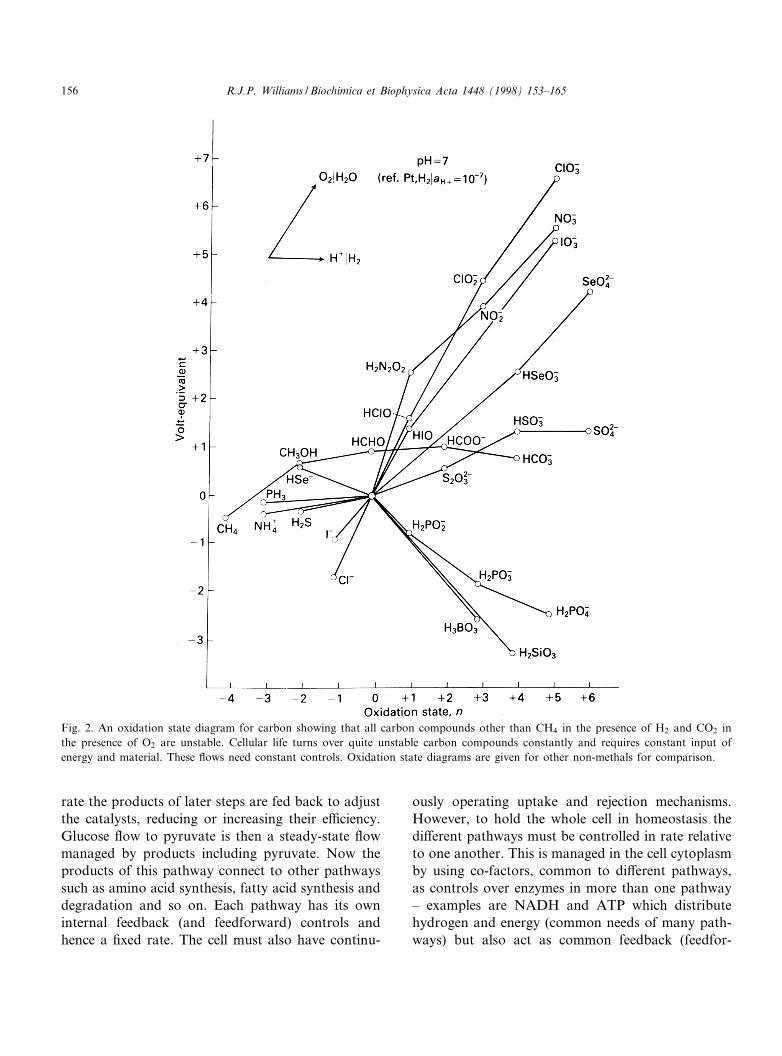
all just combinations of atoms but their combina-tions contain di¡erent amounts of energy when re-ferred to element standard states. For example,CH3COOH, CH4+CO2 and 2C+O2+2H2 all havethe same element composition but di¡er in energycontent (vG) (see Fig. 2). If all three chemicalswere in equilibrium then the ratios of each of thethree compositions would be ¢xed at any given tem-perature and pressure. The variables are then theconcentrations of the elements, C, H, and O, andenergy incorporated and not those of the compoundsCH3COOH, CO2 and CH4 or any others of the samecomposition, per se. Now the cytoplasm of biologicalcells is not in equilibrium but in ¢xed homeostatic£ow. In ¢xed £ow the ratios of say the above threecombinations of C, H and O are maintained both bycontrols on the amount of the elements and by theenergy entering and leaving the cell both of whichleave in di¡erent form from that in which they enter.The £ow has restrictions, catalysts, which decide thecompounds formed in the cell, and cells must alsohave controlled osmotic and electrical solutions.These catalysts and the composition of the aqueousmilieu require many elements other than H, C, N
and O. The variables, not directly the amounts ofindividual combinations of elements such as CO2,CH4 and CH3COOH, in a cell, must therefore beconstrained by the control feedback over uptake(and rejection) of many elements and intake andloss of energy plus controls over their distribution.Thus the entry and exit of metabolic, catalytic andcontrol chemical elements, some twenty, are the var-iables of composition and the actual composition ofchemical compounds in the cell is dependent uponthe energy another variable which is put into theseelements in combination. Consider ¢rst the homeo-stasis in a prokaryote.
The way in which prokaryote homeostasis is man-aged is to have (1) a set of catalysts which see to itthat the reactions of a given substrate, e.g. glucose,connect only to a special pathway and (2) a set ofions to maintain ¢xed solution conditions and rateconstants. The pathway rates, for example glycolysis,are such that the path is decided both in direction asis a walking path, and in rate by a series of selectivereduced energy barriers to chemical change, comparestyles in hedge-rows on walking paths, by catalysts.Now to make sure the £ow is maintained at a steady
Fig. 1. The generalised pattern of activities, £ow of energy and material in a cell.
BBAMCR 14407 3-12-98
R.J.P. Williams / Biochimica et Biophysica Acta 1448 (1998) 153^165 155
rate the products of later steps are fed back to adjustthe catalysts, reducing or increasing their e¤ciency.Glucose £ow to pyruvate is then a steady-state £owmanaged by products including pyruvate. Now theproducts of this pathway connect to other pathwayssuch as amino acid synthesis, fatty acid synthesis anddegradation and so on. Each pathway has its owninternal feedback (and feedforward) controls andhence a ¢xed rate. The cell must also have continu-
ously operating uptake and rejection mechanisms.However, to hold the whole cell in homeostasis thedi¡erent pathways must be controlled in rate relativeto one another. This is managed in the cell cytoplasmby using co-factors, common to di¡erent pathways,as controls over enzymes in more than one pathway^ examples are NADH and ATP which distributehydrogen and energy (common needs of many path-ways) but also act as common feedback (feedfor-
Fig. 2. An oxidation state diagram for carbon showing that all carbon compounds other than CH4 in the presence of H2 and CO2 inthe presence of O2 are unstable. Cellular life turns over quite unstable carbon compounds constantly and requires constant input ofenergy and material. These £ows need constant controls. Oxidation state diagrams are given for other non-methals for comparison.
BBAMCR 14407 3-12-98
R.J.P. Williams / Biochimica et Biophysica Acta 1448 (1998) 153^165156

ward) allosteric controls over enzymes in many path-ways. There are other coenzymes for distributing car-bon and nitrogen. This means that a steady state canbe characterised by a level of such coenzymes asNADH and ATP but also all the intermediates inall paths so the resultant cell is ¢xed. The cofactorshave to act on the catalysts. However, while somepathways generate energy others require it. Now sev-eral co-enzymes connect to energy ^ either H� gra-dients or ATP itself ^ and then are related to path-ways. To keep all the elements in balance input andexit pumps and exchanges of all required elementsfor catalysts and control of ionic solution conditions(Table 1) also must be connected to ATP or H� £ow.The result is a steady £ow of chemicals throughmembranes and the cytoplasm while small amountsof material and energy are syphoned o¡ to makeDNA keeping the protein complement roughly con-stant in composition. The overall result for a pro-karyote is just rapid reproduction against a back-ground of almost ¢xed metabolic activity, i.e. ofhomeostasis ^ a totally dynamic (£ow) conceptinvolving 20 elements in cyclic £ow and an almost¢xed £ow of energy degradation (Fig. 3).
The managed required elements as well as H, C, N
and O of almost all if not all prokaryote cells are S,P, Cl, Na, K, Mg, Mn, Fe, Co, (Ni), (Cu), Zn, Mo(W) [2]. All are held at closely ¢xed concentrationratios within any cell. Since there are always lossesinput must be constantly maintained and monitored.This state of a¡airs can only be procured by the ¢xeddivision of the use of energy seen by the level of ATP.We see that the essence of total homeostasis is thatcertain substances, mainly coenzymes and simple ions,form £ow (currents) connecting the pathways in cir-cuits (Fig. 4). The currents are not shown in thepathways but we give biochemical charts much asis usually the case in electronic circuit diagrams.There are, however, £ows with di¡erent rate con-stants in di¡erent parts of the circuit, e.g. of phos-phorylation and in the feedbacks between large mol-ecules (see Table 3) [3,4].
Fig. 3. A scheme for the maintenance of cellular £ow. Overall acell cycle involves energy build up with an average steady stateof vG. This vG must be constantly renewed from the environ-ment as compounds degrade. To maintain the condition energydegradation, creating entropy, is needed e.g. from the sun toyield eventually heat. Hence the application of energy as wellas basic material to synthesis needs constant control.
Table 1Connections to ATP or gradients in anaerobic prokaryotes
Reaction Connection
Glycolysis ATP production and feedback restrictionFe/S systems Production of H� gradients prior to ATP
formationIon pumps Rejection of Na�, Ca2�, Cl3
Inward movement of K�, HPO234 , Fe2�,
Mg2�, etc.Exchange for H�
or Na�Inward movement of substrates
Table 2Calcium in organelles+Golgi
Organelle Activity of calcium
Chloroplast Associated with dioxygen release (Mn, Cl)(Fe/S)
Mitochondria Associated with dehydrogenases (Fe/S)(Mn?)
Golgi Associated with folding (Cu, Fe, (Zn))(Mn and glycosylation)
BBAMCR 14407 3-12-98
R.J.P. Williams / Biochimica et Biophysica Acta 1448 (1998) 153^165 157

In the prokaryotes the energy distribution, whichwe have linked obviously and directly to ATP, doesnot involve calcium except that the ATP is used topump this element out down to a level of about 1035
M minimally, while pumping in and out various oth-er ions and molecules [1]. As stated before we believethat the role of calcium in early prokaryotes wasrelatively trivial. (Note that prokaryote £ow has acertain £exibility due to the ease with which newbits of circuitry can be made (synthesised) and in-
serted, e.g. when the external food supply is switchedsay to lactose.)
Now in the above we have referred to the controlof the major hydrolytic and synthetic metabolismusing H, C, N and O substrates and linked to chem-ical energy (ATP) and ion gradients. There are fur-ther reaction pathways in the earliest known anaero-bic prokaryotes which utilise redox reactions. Theircontrol involves message systems especially utilisingsulphur and iron compounds in signalling so thatredox homeostasis is managed. Probably all otheressential element uptake and management are linkedto a combination of these major controls. However,once again there does not appear to be a signi¢cantinvolvement of calcium but we have to observe thatdue to leakiness of membranes even at this stage ofcellular life there is a constant calcium current to andfro across the cytoplasmic membrane. It is the evo-lution of this and other calcium circuits and currentswhich we shall follow in this article.
3. The development of eukaryotes
We start discussion of the eukaryotes from theassumption that they developed from anaerobic pro-karyotes. The great advantage of the eukaryote is thedivision of cellular space into compartments whichseparate activities and which are held together by¢laments in tension. The ¢laments and their possiblechanges in tension allow a great increase in size andan ability to change shape of a cell and make possi-ble £exible external membranes used even to con-sume prokaryotes. All these additional activities re-quire energy and it therefore has to be distributedand controlled sometimes in bursts of activity whileit is constantly used at rest. During their longer life-time the eukaryotes must then have careful internal
Fig. 4. A deliberate attempt to liken the metabolic pathways ofa cell to the outputs of a computer circuit : (a) glycolytic path-way, (b) citric acid cycle. All such circuitry which is here of ba-sic H, C, N, O metabolism must have controls (see text). (AfterKau¡man.)
Table 3Timed responses
Time Coupling Response
1033 s ATPases Contraction (tension)1032 s Kinases and phosphases Metabolic pathwaysseconds or longer Transcription factors after phosphorylation:dephosphorylation DNA production of mRNA and proteinsa
aThe activity of Ca2� in the nucleus is closely linked to phosphate mobilisation. It may well be that nuclear systems like those ofmitochondria and chloroplasts are based on di¡erently timed response [3,4].
BBAMCR 14407 3-12-98
R.J.P. Williams / Biochimica et Biophysica Acta 1448 (1998) 153^165158

control of vesicles, ¢laments and cytoplasm whilethey also su¡er and respond to environmentalchanges. The great disadvantage of eukaryotes isthat they live longer and, since they develop, theymust protect themselves before reproduction. At¢rst, they used the £exible adjustment just to assistactions such as endo- and exocytosis to eat and toattack enemies. They had therefore to be aware oftheir environment and to obtain signals from it. Weobserve that the anaerobic eukaryotes developed cal-cium £ow in new circuits simultaneously with thenovel introduction in them of useful ¢laments andvesicles, to aid maintenance and changes of shape.Calcium was chosen as the messenger because it hadlittle previous use and because of its intrinsic proper-ties [2] but also because it had already been rejectedfrom the cytoplasm of prokaryotes to give a consid-erable gradient. In other words calcium managed thenew homeostasis of internal cell tension and the newtriggering of cell shape and both endo- and exocyto-sis using vesicles. Of course this required re¢nementsin the handling of calcium inward and outwardmovements, channels and pumps, which had to beplaced in the plasma membrane. Later we can as-sume that ampli¢cation of action came about usingconcentration of calcium ions in the vesicular sys-tems such as the endoplasmic reticulum needing fur-ther pumps and channels and storage proteins. Asthis reticulum developed it could protect the newnucleus and signal to it also.
In our previous article ([1] and see above), weshowed that this use of calcium as a messenger inthe cytoplasm connecting the outside environmentwith the cytoplasm, the vesicles, the ¢laments andthe nucleus needed also to link to the pre-existing(prokaryotic) anaerobic cytoplasmic controls. Thecytoplasm is controlled as stated above mainly bysubstrates, phosphate and sulphur compounds, ironand hydrogen ions, which were inter-linked. Calciumhad then to be linked to all of these control signals.We know that today it interacts directly with thesignalling phosphate system, the energy system, andthe ATPase pumps even for calcium itself. Phos-phates interact with all the other primitive controls.The signalling network is then complete for the basicanaerobic eukaryote steady state by a calcium circuitbut calcium could also be used to trigger. The calciumcurrent £ow noted in prokaryotes has been made
much more sophisticated even in the resting statesince it has localised features due to the dispositionof pumps, channels and storage devices which £uc-tuate with the shape of the cell. The control overshape and then morphogenesis by local calcium cur-rents is beautifully illustrated in work on acetabu-laria [5,6]. This situation was made more complicatedby the introduction of the two more e¡ective ways ofobtaining energy by aerobic eukaryotes: the incorpo-ration of prokaryotes as organelles in the form ofchloroplasts and mitochondria. Now we have statedthat these prokaryotes themselves did little with cal-cium! How did calcium come to link incorporatedprokaryotes in the network of controls of eukary-otes?
Before we progress further we note that whereasearly signalling in the cytoplasm is based on relativeconcentration of products of reaction, i.e. such ratiosas [ATP]/[ADP][P], [NADH]/[NAD][H�], and Fe2�/Fe3�, the calcium signal is based on the gradientratio [Ca2�]out/[Ca2�]in. We assume that in evolutionthe calcium gradient increased (see [1]) by loweringin-cell calcium to 1037^1038 M in eukaryotes fromsome 1035M in primitive prokaryotes so enablinge¡ective signalling. This use of a strong gradientdoes not appear to have developed for other avail-able ions H�, Mg2�, K� or Cl3, much though small-er gradients are used in later eukaryotes in nerves etc.Calcium in life is unique for good chemical reasons.It is the only available ion which gives fast, relativelystrong, reversible binding to organic materials andfast di¡usion. We note that if this summary is correctthe incorporated organelles always could manage ahigher calcium content than the eukaryote cyto-plasm.
4. Homeostasis and energy of aerobic unicellulareukaryotes
As stated homeostasis in eukaryotes is more com-plicated than in prokaryotes since the new internalcompartments, vesicles or organelles, all have to bein communication with the cytoplasm. Now homeo-stasis is a dynamic condition and costs energy. Inorder to maintain element homeostasis in eukaryotesmuch energy is required therefore in pumping ions,energy itself, and material across many membranes.
BBAMCR 14407 3-12-98
R.J.P. Williams / Biochimica et Biophysica Acta 1448 (1998) 153^165 159

For a steady state this means that cytoplasmic ATPlevels must be kept approximately constant so thatall gradients across membranes are ¢xed. ATP islargely maintained in aerobic eukaryotes by newprocesses: the fundamental production of protongradients in both the mitochondria and chloroplastsgiving the cell ATP. Homeostasis (and triggering)demands signalling to and from these organelles inboth resting and triggered states.
The activity of chloroplasts or mitochondria de-pends on the supply of light and CO2, or dioxygenand reduced carbon metabolites respectively. Let uscombine and describe both organelles as having afundamental exterior energy supply, hX and O2/hy-dride respectively, which is converted to a protongradient or used to generate reduced carbon (plants)or heat (animals). We should like to know the way inwhich light or dioxygen use can be controlled. Onegeneral control apart from the activity of phosphatecompounds appears to rest with the calcium levels inmitochondria or chloroplasts.
Now calcium is high when a eukaryote cytoplasmhas been activated by a calcium input from outsidethe cell or from the reticula. The activity inducedinvariably uses ATP. Hence it makes good sense togear up ATP synthesis while calcium is pumped intomitochondria as a partial recovery from activation.In mitochondria calcium moderates the activity ofthe dehydrogenases which produce ATP. We maytherefore presume that homeostasis of the mitochon-drial activity in relationship to the cytoplasm is re-stored and maintained by calcium input followed bymitochondrial Na�/Ca2� exchange or by ATPasepumps for calcium. (We also know that excess cal-cium can so damage mitochondria that the releasefrom them of cytochrome c causes apoptosis.) Notethat the calcium ion relationship to the mitochondriais very localised [7].
The chloroplast supply of energy from light is alsomodulated by calcium which interacts with and in-creases the dioxygen production of photosystem II.This is due to the level of calcium inside the thyla-koid. It may well be that the lower the use of light togive dioxygen, the greater percentage use to give re-duced carbon by photosystem I. If the electrons aretransferred to photosystem II rather than giving re-duced carbon then more ATP is produced ratherthan stored hydrogen. A balance is needed between
the two. High ATP levels outside the thylakoid willhelp to restore the state of the general homeostasis.Does calcium modulate this redistribution constantlyhence keeping homeostasis between chloroplast andcytoplast in the resting state?
The suggestion then is that across the mitochon-drial or chloroplast membrane there is always a gra-dient of calcium. The calcium is higher in the organ-elle ^ after all it is a prokaryote and can toleratehigher calcium. Modulation of this gradient sensitisesthe supply of ATP due to dioxygen or light supplyrespectively. According to need, say the demands forATP, the calcium level in the organelle rises switch-ing on ATP production. We are not suggesting thatcalcium ions are the only controls since we can besure that for example free manganese and chloridelevels are also important in both organelles (Table 2).In this article, however, the stress is on the networkof calcium £ows which now includes the reactions ofthe organelles and the ATP production for the cyto-plasm (Fig. 5). The £ows, currents, of calcium and
Fig. 5. The involvement of calcium in eukaryotes not only inthe circuitry of metabolites (Fig. 4), but also in the links to ten-sion (¢laments), local signalling (endoplasmic reticulum, ER),energy production (mitochondria and chloroplasts), the nucleusvia phosphorylation, and to exocytosis (vesicles). Note that the£ow is at a constant low level due to channels and pumps butcan be augmented by extracellular events ^ triggering.
BBAMCR 14407 3-12-98
R.J.P. Williams / Biochimica et Biophysica Acta 1448 (1998) 153^165160
energy have to be active at all times to maintain thecell steady state as well as recovery from triggering.
Now we can turn back from the sources of energyto the eukaryote to the energy distribution in thecytoplasm itself after the incorporation of mitochon-dria and chloroplasts, i.e. to advanced aerobic singlecell eukaryotes, and their activities. We need to seethat the eukaryote cell not only has internal balanceas in prokaryotes and a new balanced connection tointernal energy but also has a homeostatic steady-state mechanism relating to the environment as wellas triggered response to change. We believe calciumcurrents play a major role in this management. Thesecurrents have local intensity under di¡erential con-trols in di¡erent parts of cells [7,8] but it must beremembered that of necessity £ow is constant at alow level even in the non-triggered state (see Fig. 3).
5. Cytoplasmic activity of aerobic single-celleukaryotes
The major e¡ort in calcium research has been onstructure/function relationships of the individual re-ceptor proteins in eukaryotes (especially multi-cellu-lar) when calcium enters a cell after a message isreceived. The calcium message is often ampli¢ed byeither IP3- or Ca2�-stimulated release of calciumfrom ER stores. The calcium triggers all kinds ofactivity of kinases, phosphatases, pumps, contractionetc. at 1036 M internal concentration, much of whichis linked to energy, ATP, in a circuit. However, assophistication of compartments grew the responseswere required to have very di¡erent time constants.For example, the contractile response (movementaway from danger or toward a target) must be fasterthan internal metabolic rate change which must befaster than gene (nuclear) response. Thus action en-ergy including distribution has to be directed dispro-portionately in time (Table 3) [3,4]. Here then thereare required to be fast and slow calcium responses,which must depend on the way conformationalchanges of calcium proteins are managed. The ma-nipulation of a calcium receptor protein is dependentupon exchange rates between calcium free and boundstates to any other unit e.g. another protein, a mem-brane or phosphate compounds (see [3]). The re-sponses are localised in space and not general to
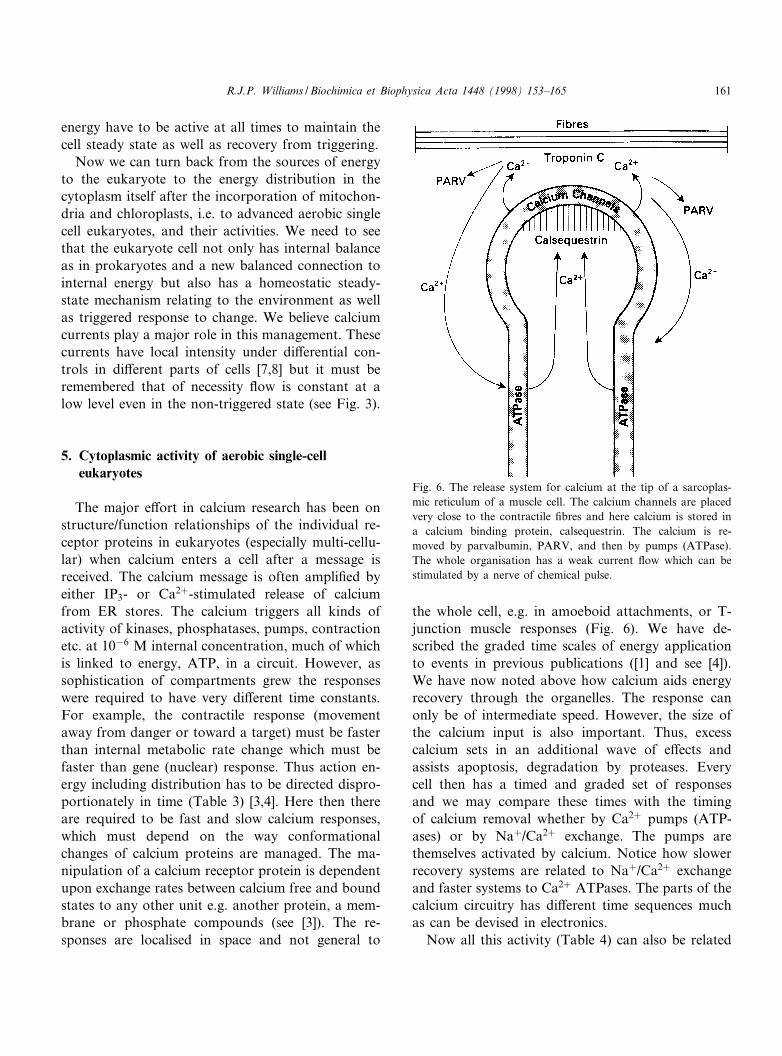
the whole cell, e.g. in amoeboid attachments, or T-junction muscle responses (Fig. 6). We have de-scribed the graded time scales of energy applicationto events in previous publications ([1] and see [4]).We have now noted above how calcium aids energyrecovery through the organelles. The response canonly be of intermediate speed. However, the size ofthe calcium input is also important. Thus, excesscalcium sets in an additional wave of e¡ects andassists apoptosis, degradation by proteases. Everycell then has a timed and graded set of responsesand we may compare these times with the timingof calcium removal whether by Ca2� pumps (ATP-ases) or by Na�/Ca2� exchange. The pumps arethemselves activated by calcium. Notice how slowerrecovery systems are related to Na�/Ca2� exchangeand faster systems to Ca2� ATPases. The parts of thecalcium circuitry has di¡erent time sequences muchas can be devised in electronics.
Now all this activity (Table 4) can also be related
Fig. 6. The release system for calcium at the tip of a sarcoplas-mic reticulum of a muscle cell. The calcium channels are placedvery close to the contractile ¢bres and here calcium is stored ina calcium binding protein, calsequestrin. The calcium is re-moved by parvalbumin, PARV, and then by pumps (ATPase).The whole organisation has a weak current £ow which can bestimulated by a nerve of chemical pulse.
BBAMCR 14407 3-12-98
R.J.P. Williams / Biochimica et Biophysica Acta 1448 (1998) 153^165 161

to triggering and its recovery to steady states but wehave to see that this does not solve a major problemof action which has to be against the background ofhomeostasis. Homeostasis is based upon the restingor developmental level of £ow as well as concentra-tion in a cell which, for a single eukaryote cell, has tobe maintained against a variable environment. It ishere that it appears that calcium is the major com-munication connector. A nice example of the localcircuitry is given next.
Recently the structure of the calcium reservoir inthe endoplasmic and sarcoplasmic reticulum hasbeen uncovered [9]. There is in both vesicular sys-tems a protein, calreticulin or calsequestrin, whichcan hold some 40 Ca2� ions in a cleft of two thio-redoxin-like domains. The proteins are present inalmost crystalline array and are very close to bothchannels for calcium release and pumps for calciumuptake. The positioning of all three proteins (seeFig. 5) allows small circulating calcium currents atall times as well as quick release and recovery of thehomeostatic balance. These local current £ows are ofextreme importance in the management of the cellsince the calcium currents are in communicationwith ¢laments as in a muscle cell and maintain cellmetabolism through kinases, cell shape through ¢l-ament ATPases, and ion levels through plasmamembrane pumps. There are also 2Na�/Ca2� ex-
changers in some of these membranes and perhapsassociated with organelles and energy £ow homeo-stasis. This general local current is reminiscent ofthat of the protons in mitochondria and chloro-plasts. It is also reminiscent of advanced computercircuitry where a low level of £ow is maintained atall times.
Here we draw attention to the new knowledgeconcerning mitochondria, chloroplasts and reticula.In early work electron microscopy apparentlyshowed that these compartments were small, oftendrawn as almost spherical vesicles, with manyvesicles of each kind in a cell, e.g. calsizomes forcalcium storage. In the last few years serial electronmicroscopy has shown that this is a false picture. Infact all three vesicular structures consist of very largeoften single bodies occupying a considerable volumebut composed of waving tubular or laminar mem-branes. The change in the knowledge of these struc-tures requires a re-thinking of the way in which theywork. Instead of treatments which deal with individ-ual small spherical vesicles, with generalised bulkgradients of ions and potentials, we need localisedmodels of activity [10]. Thinking in terms ofsteady-state vesicular thermodynamics, for exampleas characterised by chemi-osmosis, may well missthe sophistication permitted in these large systems,due to localised responses which are so clearly seenby calcium indicators [11], and which will a¡ect lo-calised mitochondrion proton activity.
6. The further needs of a homeostasis in multi-cellularorganism
A multi-cellular organism has additional require-ments to those of a single eukaryote cell. It musthave:
1. An extracellular matrix to hold separate cells inplace in organs yet to allow growth this systemmust be adjusted using energy
2. An extracellular £uid for transfer of materialwhich has to be in a protected container, skin,cuticle or shell, and circulated using energy
3. Messenger systems to give information from re-ceptors in one area (e.g. the head) to all otherareas. Again there is an energy cost.
Table 4Components of the ¢xed intracellular calcium current in eukar-yotes
Component Function
Ca2� ATPase pumps,Na�/Ca2� exchangers
Removing calcium from the cytoplasmto external £uids, vacuoles or organ-elles
Ca2� channels Input of calcium from external £uidsor vesicles to cytoplasm
Actinomyosin ¢laments Maintenance of cell tensionEnzymes Activation by calmodulins, annexins,
S100 and so on, of kinases, phospha-tases, proteases and so on (relayed tonuclear activity)
Proteins in vesicles Calsiquestrin and others as storage oflarge amounts of calcium, internal con-densers
NB. All these systems are constantly active at a low level atrest. Activity general or selective causes changes in the calciumcurrent in the cell stimulated by external events.
BBAMCR 14407 3-12-98
R.J.P. Williams / Biochimica et Biophysica Acta 1448 (1998) 153^165162

Note that as well as being used in responsivechanges the above must act in the maintenance ofthe steady state of cell-cell and organ-organ relation-ships. This implies a constant current £ow of manyions and molecules throughout the organism. Theseneeds are apart from the requirements for materialand energy £ow in single cells which we have justdescribed. No synthesis occurs outside cells and noATP energy is available there so that whereas thereare the sources of energy in the cytoplasm ^ chemicalbond energies (ATP), tension, and gradients ^ thereare really only two sources of energy in the extra-cellular £uids ^ tension and gradients. Changing ten-sion is supplied by contractile devices in muscles,heart and arteries for example ^ all calcium acti-vated. The fact that the cell is a massive store ofmolecules both in the cytoplasm and in vesiclesmeans that relative to the extracellular £uids thecell stores a multitude of energised organic chemicalgradients especially in its vesicles. There are calcium-dependent release mechanisms for these gradients ^exocytosis from vesicles ^ which allow the external£uid to carry messages, e.g. transmitters and hor-mones, or to transform the ejected proteins for useexternally. Some of these proteins are for the syn-thesis of the extracellular matrix and some for pro-tection, digestion, and so on. The message systems tomaintain the workings of the whole are a variety oftransmitters and hormones which are exposed to a¢xed extracellular calcium. The activities of verymany extracellular proteins as well as the exocytosisof hormones etc. depend on calcium as in the follow-ing example. (Note the reverse of cell activationwhere changing calcium adjusts ¢xed levels of en-zymes making them active while in the extracellular£uids changing organic chemical levels are activatedby ¢xed calcium.)
7. Exocytosis and signals
As stated there is no synthesis outside cells. Henceall the organic requirements for maintenance (signal-ling) and growth come from inside cells even if someof the basic units have to be transported into thecells from other cells. The demand is for a controlledsteady level of messenger molecules in the circulating£uids released by cells as well as a controlled release
of structural proteins and protective enzymes for ex-tracellular use. Nothing is permanent in organisms.There has to be both steady-state £ow and growth ofagents which communicate to the inside from out-side. We know that external events cause excitationand calcium entry but internal stress, which could befor example oscillating mechanical stretching usingthe extracellular ¢laments, may also allow calciumto enter the cell. Both may stimulate regular vesicleexocytosis so that calcium is a maintenance andgrowth signal between cells assisted by speci¢c andselective transmitters and hormones and excited pro-teins all of which circulate at certain levels. Do notforget that these hormones and proteins are releasedfrom vesicles but are constantly degraded so must beconstantly distributed. Thus calcium governs growthand maintenance systems in that it governs release ofall these substances by exocytosis. We have to recog-nise that the functioning of a large multi-cellular or-ganism and equally for each single cell requires con-stant £ow even at rest so that homeostasis is basedon current £ow, carefully controlled.
Now such mechanisms as these require very closemanagement at all times of the calcium levels in theextracellular £uids, and simultaneously the control ofthe extracellular matrix itself. These controls operateover time scales of weeks, months and years and arequite unlike those used in the cytoplasm of singlecells. Thus we have to consider new additions to
Table 5Components of the extracellular calcium current in multi-cellu-lar organisms
Component Function
Epithelial cells Calcium input using asymmetrically placedpumps and channels and carrier proteins,calbindin
Kidney cells Excretion of excess calciumBone and bonecells
Maintenance of calcium levels in the extra-cellular £uids
Muscle cells Maintenance of £ow in body £uidsGlands Maintenance of circulating hormones to
relay information about calcium levels toDNA protein synthesis
NB. All these systems are constantly active at a low level atrest. Di¡erent hormone or ion current level (Na�/K�) changescan switch the calcium levels temporally. During developmentthe calcium currents are changing continuously e.g. in fertilisa-tion.
BBAMCR 14407 3-12-98
R.J.P. Williams / Biochimica et Biophysica Acta 1448 (1998) 153^165 163

the calcium network since calcium in the extracellu-lar £uid can only be ¢xed if it is replaced while it isconstantly lost (see Table 5).
8. Extracellular £uid calcium
Calcium is such a major player in the maintenanceof multi-cellular homeostasis that its extracellularconcentration must be very ¢nely managed. Herewe meet an intricate network of interactions associ-ated with mineral deposits especially in higher ani-mals. What is required is regulated uptake and dep-osition of calcium itself at a steady rate so thatresources are constant, keeping calcium £ow in thewhole extracellular system ¢xed. The deposition is ofcalcium in many structures outside of cells includingminerals such as bone. The need is to take in calciumsteadily while rejecting excess. Thus there are epithe-lial cells for uptake with specially placed calcium in-lets and pumps for injection into the organism atdi¡erent sides of these cells and calcium proteinsfor transport across their cytoplasm, for examplecalbindin. Excretion is via the kidney. This activityis regulated by the production of the appropriateproteins which means that there has to be feedbackinstruction to DNA from the circulating levels ofparticular units. One instruction is via the main-tained levels of vitamin D generating a message tozinc-¢nger transcription factors. This sterol com-mands the synthesis of the calbindins and also theprotein production for the control of calcium depo-sition in bone. Thus bone is maintained as a depositof calcium and as far as this article is concerned boneis a condenser of the current carrying ion, calcium(Table 5).
Since the uptake of calcium is necessary to keepbone in a good condition a very small excess concen-tration over the solubility product is desirable since alarge excess will lead to general precipitation. Whatwe observe is that bone is constantly monitored bycells so that the calcium level is very precisely main-tained at a precise e¡ectively bu¡ered level. A majorprotein is osteocalcin. The management of extracel-lular calcium current is emphasised by a second sys-tem.
Recently a second mode of management of extra-cellular calcium has been discovered [12]. The level of
calcium in the extracellular £uid a¡ects via G-pro-teins and IP3 the level of cytoplasmic calcium insidecertain cells. The rise of internal cytoplasmic calciumlevel with fall in external calcium causes release ofparathyroid hormone from vesicles to the extracellu-lar £uid where it acts to increase calcium releasefrom bone. The functions of vitamin D and of theparathyroid hormone are complementary.
We see that the circuitry of extracellular calcium islinked to the circuitry of intracellular calcium inmany ways making the total circuit network verycomplex.
9. Brain growth: a special calcium system
The brain is a peculiar organ in that it develops(grows) to the dictates of the environment as per-ceived by the animal but not under genetic control.Thus it is now experience which makes growth andsynapse connection. The initial event is a Na�/K�
current which releases organic chemicals fromvesicles of nerves to stimulate activate growth con-nections. However, it is again modulation of calcium£ow which controls release and synapse growth[8,13]. There is an extraordinary feature in that theextracellular £uid is now quite di¡erent in calciumconcentration from that of the whole body sincethe brain is immersed in cerebrospinal £uids (CSF).For how long has this been true? Why is it neces-sary? Why is CSF low in calcium? It needs energy tokeep it so. One possibility concerns the sensitivity ofglutamate release to the levels of external calcium[13]. Much as is the case for parathyroid hormonerelease, external calcium concentration a¡ects gluta-mate release. Another possibility is that the low cal-cium is an insurance against precipitation of calciumcarbonate, as in the ear, or of calcium phosphate, oragainst any calcium sensitive reactions such as clot-ting of proteins. However, much of the operation ofthe brain represents another separate calcium-linkedcircuit.
10. Conclusions
This article has as its objectives the demonstrationthat energy dissipation as well as material transport
BBAMCR 14407 3-12-98
R.J.P. Williams / Biochimica et Biophysica Acta 1448 (1998) 153^165164

across membranes is more often that not calciumrelated. The connection is made in several ways:
1. Direct control over ATP production by organelles2. In£ux of Ca2� to cytoplasm as a general signal3. E¥ux of many proteins and other molecules to
the high calcium of extracellular £uids4. Control by Ca2� over contractile poise5. Control by Ca2� over the other gradients of ele-
ments e.g. links to Na�, K� etc.6. Whole body movement of calcium between or-
gans.
Ca2� is certainly a fast trigger through its in£uxbut it can also create a slow sustained response.However, it is linked to so many other cellular fea-tures that its additional role could well be in generalhomeostasis of the resting state. If this is so each celland each organism has to be looked at as if it were
an electrolytic device parallel to an electronic system,e.g. a computer (Fig. 7). If we look at cells as welook at modern computers then the equivalent devi-ces have to be found. The equivalent of the battery isthe storage of calcium in gradients, water replaceswires, switches are current (calcium) induced confor-mation changes, condensers are vesicles (ER etc.) in-side the cell, retaining calcium, or bone. As in acomputer (compare especially the brain) there is con-stant low-level current £ow in the maintained state.The implications is the calcium ions constantly mon-itor and maintain homeostasis as well as acting as atrigger. The parallel is between an electron and thecalcium ion. If this is accepted then we can notunderstand calcium function except in terms of inte-grated circuits.
References
[1] R.J.P. Williams, Cell Calcium 20 (1996) 87^93.[2] R.J.P. Williams, J.J.R. Frausto da Silva, The Natural Selec-
tion of the Chemical Elements, Oxford University Press,Oxford, 1996.
[3] J.W. Putney, Science 279 (1998) 191^193.[4] R.J.P. Williams, Cell Calcium 16 (1994) 339^346.[5] C. Brie©re, B.C. Goodwin, J. Theor. Biol. 131 (1985) 461^475.[6] L.G. Harrison, N.A. Hillier, J. Theor. Biol. 114 (1985) 177^
192.[7] R. Rizzuto, P. Pinton, W. Carrington, F.S. Fay, K.E.
Fogarty, L.M. Lifshitz, R.A. Tuft, T. Pozzan, Science 280(1998) 1763^1765.
[8] R.J.P. Williams, Bioenergetics 1 (1970) 215^225.[9] S. Wang, W.R. Trumble, H. Liao, C.R. Wesson, A.K.
Dunker, C. Kang, Nature (1998) in press.[10] R.J.P. Williams, Biochim. Biophys. Acta 505 (1978) 1^44.[11] L.A. Blatter, E. Niggli, Cell Calcium 23 (1998) 269^279.[12] J.E. Garrett, I.V. Capuano, L.G. Hammerland, B.C.P.
Hung, E.F. Nenneth, F. Fuller, J. Biol. Chem. 270 (1995)12919^12925.
[13] Y. Kubo, T. Miyashita, Y. Murata, Science 279 (1998) 1722^1725.
Fig. 7. A much simpli¢ed relationship between the circuits ofelectrons in man's electronic devices and the electrolytic circuitsof calcium in cells and organisms. The level of comparison ex-tends to the brain and senses in that triggering of both electronand calcium circuits are similar. Energy: Ca2� battery storagecell ; Switches: Ca2� channel openings; Condensers : vesicles,ER etc., (bone); Device: protein receptors; Connectors: Ca2�
transfer through membrane pumps.
BBAMCR 14407 3-12-98
R.J.P. Williams / Biochimica et Biophysica Acta 1448 (1998) 153^165 165
Related Documents











