Calcareous nannofossil biostratigraphy and paleoecology of the Cretaceous^Tertiary transition in the central eastern desert of Egypt Abdel Aziz A.M. Tantawy Department of Geology, Faculty of Science, South Valley University, Aswan 81528, Egypt Received 2 January 2002; received in revised form 26 September 2002; accepted 26 September 2002 Abstract The Gebel Qreiya and nearby Wadi Hamama sections of the central Eastern Desert are among the most complete K/T boundary sequences known from Egypt. The two sections were analyzed spanning an interval from l.83 Myr below to about 3 Myr above the K/T boundary. A 1-cm-thick red clay layer at the K/T boundary at Gebel Qreiya contains an Ir anomaly of 5.4 ppb. The high-resolution study and well-preserved nannoflora provide good age control and the first quantitative records of calcareous nannofossil assemblages for paleoecological interpretations across the K/T transition in Egypt. Four zones (Micula murus, Micula prinsii, NP1, and NP2) were distinguished and correlated with other nannofossil and planktonic foraminiferal zonations that are broadly applicable for the eastern Tethys region. Latest Maastrichtian assemblages are abundant and diverse, though Cretaceous species richness progressively decreased across the K/T boundary. Dominant species include Arkhangelskiella cymbiformis, Micula decussata and Watznaueria barnesae, with high abundance of dissolution-resistant M. decussata reflecting periods of high environmental stress. Thoracosphaera blooms mark the K/T boundary and are followed by an acme of the opportunistic survivor Braarudosphaera bigelowii, the first appearance of the new Tertiary species Cruciplacolithus primus, and an acme of Coccolithus cavus/pelagicus. These successive abundance peaks provide the basis for subdivision of the Early Danian Zones NP1 and NP2 into five subzones. Correlation of selected nannofossil taxa from the Egyptian sections with those from various onshore marine and deep-sea sections provides insights into their paleoenvironmental and paleoecological affinities. ȣ 2002 Elsevier Science B.V. All rights reserved. Keywords: calcareous nannofossils; biostratigraphy; paleoecology; K/T boundary; Eastern Desert; Egypt 1. Introduction In most localities throughout Egypt, Paleocene sediments unconformably overlie Maastrichtian marls. Erosion was at a maximum in the southern Nile valley and southern and central western Desert, where sediment deposition occurred in a shallow marginal sea. In these regions erosion is primarily the result of local tectonic activity and sea-level regression (El-Naggar, 1966; Issawi, 1972; Barthel and Herrmann-Degen, 1981; Her- 0377-8398 / 02 / $ ^ see front matter ȣ 2002 Elsevier Science B.V. All rights reserved. PII:S0377-8398(02)00135-4 * Corresponding author. E-mail address: [email protected] (A.A.A.M. Tantawy). Marine Micropaleontology 47 (2003) 323^356 R Available online at www.sciencedirect.com www.elsevier.com/locate/marmicro

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Calcareous nannofossil biostratigraphy and paleoecology ofthe Cretaceous^Tertiary transition inthe central eastern desert of Egypt
Abdel Aziz A.M. Tantawy �
Department of Geology, Faculty of Science, South Valley University, Aswan 81528, Egypt
Received 2 January 2002; received in revised form 26 September 2002; accepted 26 September 2002
Abstract
The Gebel Qreiya and nearby Wadi Hamama sections of the central Eastern Desert are among the most completeK/T boundary sequences known from Egypt. The two sections were analyzed spanning an interval from l.83 Myrbelow to about 3 Myr above the K/T boundary. A 1-cm-thick red clay layer at the K/T boundary at Gebel Qreiyacontains an Ir anomaly of 5.4 ppb. The high-resolution study and well-preserved nannoflora provide good age controland the first quantitative records of calcareous nannofossil assemblages for paleoecological interpretations across theK/T transition in Egypt. Four zones (Micula murus, Micula prinsii, NP1, and NP2) were distinguished and correlatedwith other nannofossil and planktonic foraminiferal zonations that are broadly applicable for the eastern Tethysregion. Latest Maastrichtian assemblages are abundant and diverse, though Cretaceous species richness progressivelydecreased across the K/T boundary. Dominant species include Arkhangelskiella cymbiformis, Micula decussata andWatznaueria barnesae, with high abundance of dissolution-resistant M. decussata reflecting periods of highenvironmental stress. Thoracosphaera blooms mark the K/T boundary and are followed by an acme of theopportunistic survivor Braarudosphaera bigelowii, the first appearance of the new Tertiary species Cruciplacolithusprimus, and an acme of Coccolithus cavus/pelagicus. These successive abundance peaks provide the basis forsubdivision of the Early Danian Zones NP1 and NP2 into five subzones. Correlation of selected nannofossil taxafrom the Egyptian sections with those from various onshore marine and deep-sea sections provides insights into theirpaleoenvironmental and paleoecological affinities.> 2002 Elsevier Science B.V. All rights reserved.
Keywords: calcareous nannofossils ; biostratigraphy; paleoecology; K/T boundary; Eastern Desert; Egypt
1. Introduction
In most localities throughout Egypt, Paleocene
sediments unconformably overlie Maastrichtianmarls. Erosion was at a maximum in the southernNile valley and southern and central westernDesert, where sediment deposition occurred in ashallow marginal sea. In these regions erosion isprimarily the result of local tectonic activity andsea-level regression (El-Naggar, 1966; Issawi,1972; Barthel and Herrmann-Degen, 1981; Her-
0377-8398 / 02 / $ ^ see front matter > 2002 Elsevier Science B.V. All rights reserved.PII: S 0 3 7 7 - 8 3 9 8 ( 0 2 ) 0 0 1 3 5 - 4
* Corresponding author.E-mail address: [email protected]
(A.A.A.M. Tantawy).
MARMIC 905 27-1-03
Marine Micropaleontology 47 (2003) 323^356
R
Available online at www.sciencedirect.com
www.elsevier.com/locate/marmicro

mina, 1990; Hendriks et al., 1984; Tantawy et al.,2001). The K/T unconformity lies within theDakhla Formation at the base of a thin phos-phatic conglomerate (or sandy limestone) that iswidespread in the Dakhla^Farafra region andmarks the base of the Abu Minqar Horizon (Ab-bas and Habib, 1969; Barthel and Herrmann-De-gen, 1981; Tantawy et al., 2001). In the centraland northern parts of the Eastern Desert and theSinai, variable erosion occurred in middle to outershelf environments (Sestini, 1984; Hendriks andLuger, 1987; Luger, 1988; Jenkens, 1990; Kassaband Keheila, 1994; Luger et al., 1998), althoughhiatuses are relatively short and intra-zonal.
The Gebel Qreiya and nearby Wadi Hamamasections in the central eastern Desert are amongthe most complete Upper Maastrichtian to Dan-ian sequences in Egypt and among the very fewlocalities where the Plummerita hantkeninoides(CF1) and Parvularugogloberina eugubina (P1a)planktonic foraminiferal Zones, as well as the Mi-cula prinsii and Markalius inversus (NP1) calcare-ous nannofossil Zones are present (e.g. East Qenaregion, Luger, 1988; Tantawy, 1998; Luger et al.,1998; Duwi region, Tantawy, 1998; St. Paul,South Galala, Strougo et al., 1992; Faris, 1995a,1997).
During the K/T transition, the Gebel Qreiyaand Wadi Hamama sections were located on thestable shelf of the Asyut Basin (Fig. 1) at middleto upper-shelf depths (Luger, 1988). The regionwas subject to global sea-level £uctuationsthroughout the Late Cretaceous and Early Paleo-cene (Hendriks et al., 1987; Luger and Gro«schke,1989; Klitzsch et al., 1990).
Calcareous nannofossil biostratigraphy andturnover across the K/T boundary have been in-vestigated intensively during the last several yearsin order to understand the nature of the massextinction that a¡ect groups in di¡erent environ-ments and latitudes (Worsley, 1974; Monechi,1977, 1979; Percival and Fischer, 1977; Romein,1977; Perch-Nielsen, 1979a-c; Thierstein andOkada, 1979; Thierstein, 1981; Perch-Nielsen,1981c; Romein and Smit, 1981; Perch-Nielsen etal., 1982; Jiang and Gartner, 1986; Seyve, 1990;Gorostidi and Lamolda, 1991; Eshet et al., 1992;Pospichal, 1991, 1994, 1995; Alcala¤-Herrera et al.,
1992; Ehrendorfer and Aubry, 1992; Lamoldaand Gorostidi, 1992; Pospichal and Bralower,1992; Gartner, 1996; Henriksson, 1996; Gardinand Monechi, 1998; Gardin, 2002). A numberof studies have investigated calcareous nannofos-sil biostratigraphy across the K/T boundary inEgypt (e.g. El-Dawoody, 1973, 1983, 1990a,b;El-Dawoody and Barakat, 1973; El-Dawoodyand Zidan, 1976; Abdelmalik et al., 1978; Faris,1984, 1985, 1988, 1995a, 1997; Schrank andPerch-Nielsen, 1985; Faris and Abd Hamid,1986a,b; Faris et al., 1985; Bassiouni et al.,1991), but no previous studies were quantitativein nature or carried out at high resolution.
This study di¡ers from earlier reports in that anintegrated quantitative database is used for cal-careous nannofossils to provide improved biozo-nation and age control and to infer paleoenviron-mental conditions during the latest Maastrichtianand earliest Danian in the central Eastern Desertof Egypt. The primary objectives are: (1) calcar-eous nannofossil biostratigraphy; (2) analysis ofcalcareous nannofossil assemblages; and (3) dis-tribution and paleoecology of selected species.
2. Location and lithology
The material used in this study was obtainedfrom the Gebel Qreiya and nearby Wadi Hamamasections of the Qena region in the central EasternDesert. Gebel Qreiya is located at the southernmouth of the Wadi Qena (lat. 26‡21PN, long.33‡01PE), about 50 km northeast of Qena Cityand about 18 km north of km 53 of the Qena^Safaga road. The Wadi Hamama section is lo-cated in the southwest of the Wadi Hamama,just north of the extreme eastern end of theWadi Umm Solimat (lat. 26‡18PN, long. 33‡02PE),about 28 km south of Gebel Qreiya and 10km south of km 44 of the Qena^Safaga road(Fig. 1).
Local variations in lithology in the two sectionsare expressed by di¡erent members of the DakhlaFormation. The lithostratigraphic classi¢cationand equivalent ages of this succession in theQena area are discussed by many authors (e.g.Barron and Hume, 1902; Said, 1962; Abdel Ra-
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356324

zik, 1969, 1972; Faris, 1974, Soliman et al., 1986).According to Abdel Razik (1972), the DakhlaFormation of Maastrichtian age comprises yel-lowish marls with numerous casts of Pecten fara-frensis alternating with dark gray calcareousshales. They are equivalent to the upper part ofHamama marl member of Abdel Razik (1972)and are overlain by greenish-gray, laminatedshales and claystones of Paleocene age (equivalent
to the Beida shale member of the Dakhla Forma-tion, Abdel Razik, 1972; Luger, 1988). The thinred K/T boundary clay found in the Qreiya sec-tion contains an Ir anomaly of 5.4 ppb (Keller etal., in press) and represents the ¢rst such claylayer observed in Egypt. Similar thin red claylayers are present in all complete or nearly com-plete K/T boundary sections in low and middlelatitudes (Keller et al., 1995). At the Qreiya
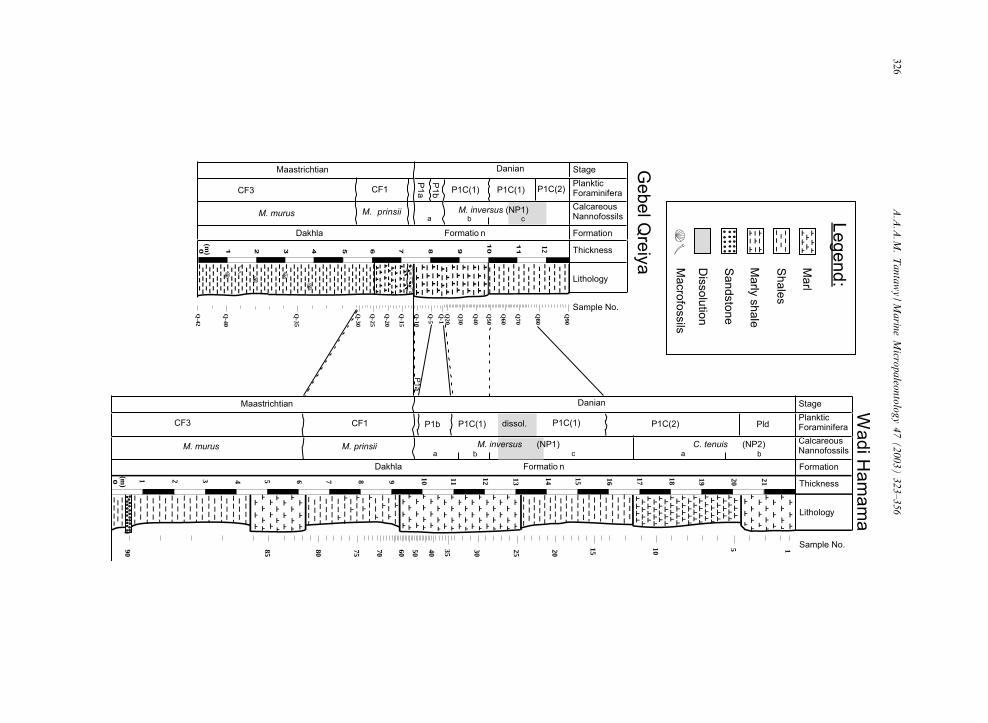
Fig. 1. Location and geological map with the studied sections.
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356 325
12
Q-40
Q-35
Q20
Q30
Q80
Q90
Q40
Q50
Q60
Q70
Q-30
Q-25
Q-20
Q-15
Q-10
Q-5
Q-1
(m)
CalcareousNannofossils
PlankticForaminiferaCF3 CF1
P1
a
P1
b P1C(1) P1C(2)
DanianMaastrichtian Stage
M. prinsii M. murus
Lithology
Thickness
Sample No.
Dakhla Formatio n Formation
10 2 3 4 5 6 7 8 9
10
11
151015202530354050607075808590
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
CalcareousNannofossils
PlankticForaminifera
CF3 CF1
P1
a
P1b P1C(1) P1C(2)
DanianMaastrichtian Stage
M. murus a b c a b
Dakhla Formatio n
Lithology
Thickness
Sample No.
a b c
Ge
be
l Qre
iya
Wa
di H
am
am
a
P1C(1)
dissol. P1C(1) Pld
M. inversus (NP1)
M. inversus (NP1)
M. prinsii
Formation
Marl
Sh
ale
s
Sa
nd
sto
ne
Marly
shale
Macro
fossils
Dis
so
lutio
n
Le
ge
nd
:
Q-42
0
(m)
C. tenuis (NP2)
MARMIC
90527-1-03
A.A.A.M.Tantaw
y/M
arineMicropaleontology
47(2003)
323^356326

section the red clay layer is 1 cm thick with a thingypsum layer (Fig. 2) and is devoid of calcareousmicrofossils.
The K/T red clay layer overlies an undulatingerosional surface which lies 10 cm above a seconderosional surface. These erosion surfaces indicatean incomplete uppermost Maastrichtian sedimentrecord and are characteristic of the widespreaddisconformities observed in many sections in cen-tral and southern Egypt (El-Naggar, 1966; Issawi,1972; Faris, 1982, 1984, 1985; Tantawy, 1998).Nevertheless, erosion at Gebel Qreiya is relativelyminor as indicated by the presence of the latestMaastrichtian Micula prinsii and Plummeritahantkeninoides zones and for this reason the sec-tion is considered one of the most complete K/Ttransitions in central Egypt (Luger, 1988; Lugeret al., 1998; Tantawy, 1998; Tantawy et al.,2000). Similar observations are reported fromthe Galala plateau to the north in the EasternDesert (Strougo et al., 1992; Faris, 1995a, 1997)and the Sinai (Faris, 1988, 1992; Shahin, 1992).
3. Methods
To avoid losing small-sized coccoliths, sampleswere processed by smear slide preparation fromraw sediment samples as described by Perch-Niel-sen (1985). Smear slides were examined using alight photomicroscope with 1000^2000U magni¢-cation. Each slide was observed under cross-po-larized light and with a gypsum plate and phasecontrast was used where necessary. Index speciesare illustrated in Plates 1^3.
Calcareous nannofossils are generally abundantand well-preserved in both sections, which facili-tates detailed quantitative analysis of the £oralassemblages. The counting technique describedby Jiang and Gartner (1986) was used for quanti-tative analysis. All samples were prepared simi-larly to insure uniformity in the distribution ofsample material and to minimize bias. Relative
species abundances were determined by countinga population of about 300 specimens along a ran-dom traverse with a light microscope at a magni-¢cation of about 1250U. Rare species were in twoadditional traverses to assure that no stratigraphicmarker was overlooked. These species are markedby an X in the distribution charts. The relativeabundance of dominant individual species, andthe presence of specimens of all species are plottedand shown in Tables 1 and 2. The number of¢elds of view traversed was also counted.
Total nannofossil abundance was calculated asthe total number of specimens counted per num-ber of ¢elds of view traversed. The species rich-ness (diversity) is given as the total number ofspecies recorded in each sample. The procedurefor counting broken or fragmented specimens(e.g. Thoracosphaera spp. and Braarudosphaera bi-gelowii) is similar to that described by Jiang andGartner (1986), Pospichal (1991, 1995) and Pos-pichal and Bralower (1992). For Thoracosphaeraspp., specimens larger than about one quarter toone third of the whole spherical forms arecounted and added up to one. Every pentalithof B. bigelowii constructed of three or four joinedelements was counted as one specimen and everyfour isolated elements were also counted as onewhole specimen. In order to facilitate counting,the following species were combined: all Thora-cosphaera spp., Arkhangelskiella cymbiformis andArkhangelskiella speciallata ; Cyclagelosphaeraalta and Cyclagelosphaera reinhardtii ; Coccolithuspelagicus, Coccolithus cavus, and Coccolithus ova-lis ; Prinsius tenuiculum and Prinsius dimorphosus.
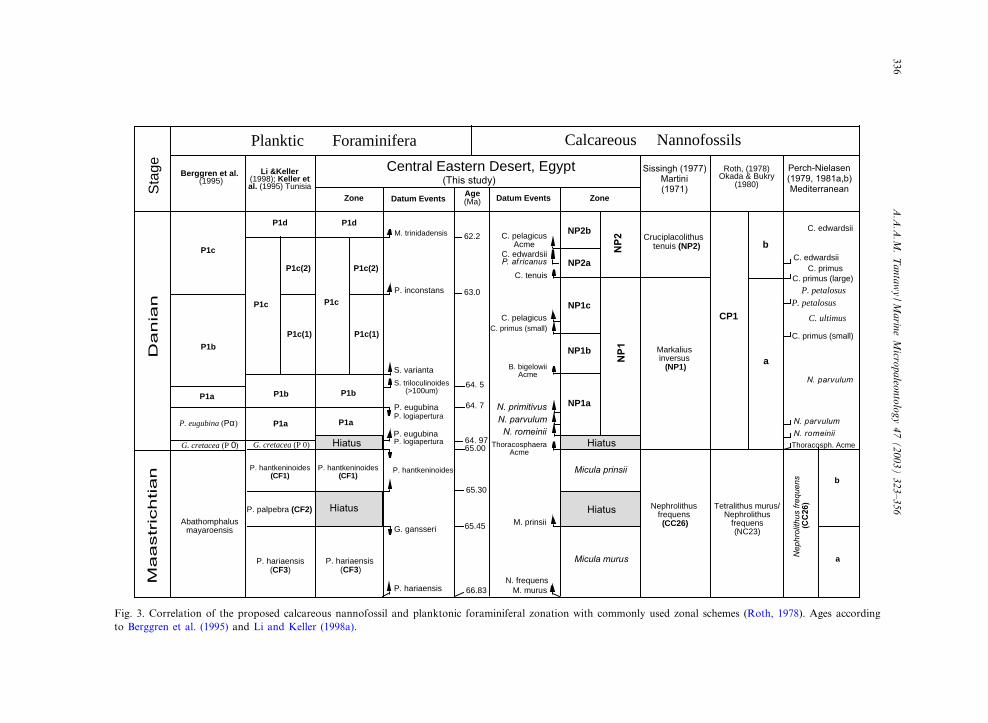
4. Biostratigraphy
The standard calcareous nannofossil zonationsused in this study are those of Sissingh (1977) forthe Maastrichtian and Martini (1971) for theDanian. In addition, the subdivisions of biozonesproposed by Romein (1979) and Perch-Nielsen
Fig. 2. Lithological columns, sample intervals, integrated biostratigraphies of the studied Cretaceous^Tertiary interval in the Ge-bel Qreiya and Wadi Hamama sections.
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356 327
1 2 3 4 5
109876
11 12 13 14 15
2019181716
21 22 23 24 25
3029282726
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356328

(1981a,b) are followed. Quantitative species anal-yses, as well as the ¢rst and last appearance ofspecies, are used in this study to obtain high-res-olution age control. The planktonic foraminiferalzonation for the Qreiya section is from Keller etal. (in press) based on the zonal scheme by Kelleret al. (1995) and Li and Keller (1998a,b).
4.1. Micula murus Zone
The Micula murus Zone (Bukry and Bramlette,1970; emended Perch-Nielsen, 1981c) spans fromthe ¢rst occurrence (FO) of M. murus to the FOof Micula prinsii. This zone is equivalent to thelower part of the Nephrolithus frequens Zone (Ce-pek and Hay, 1969; emended Romein, 1979) orCC26 of Sissingh (1977). The M. murus Zone isprobably restricted to low latitudes (e.g. Worsleyand Martini, 1970; Worsley, 1974; Romein,1979), whereas the N. frequens Zone is mainlyapplicable in higher latitudes (Perch-Nielsen,1985). In this study, N. frequens is rare in bothsections and restricted to short intervals. Miculamurus spans the lower 7.5 m and 6.2 m at theQreiya and Wadi Hamama sections, respectively,and corresponds to the CF3 planktonic foraminif-eral Zone which de¢nes the interval from the FOof Plummerita hariaensis to the LO (last occur-rence) of Gansserina gansseri (66.83^65.45 Ma,see Keller et al. (in press)). The calcareous nanno-fossil assemblages characteristics of this zone con-
tain common to abundant Arkhangelskiella cym-biformis, Micula decussata, Watznaueria barnesae,Cribrosphaerella ehrenbergii, Ahmuellerella octora-diata, Lithraphidites carniolensis, Prediscosphaeracretacea and few to rare Lithraphidites quadratus,Ei¡ellithus turrisei¡elii, Octolithus multiplus, M.murus, Rhagodiscus angustus, Thoracosphaeraspp., Braarudosphaera bigelowii, and Zygodiscusspiralis.
4.2. Micula prinsii Zone
The Micula prinsii Zone (Perch-Nielsen, 1979a;emended Romein and Smit, 1981) spans the inter-val from the FO of M. prinsii to the ¢rst commonoccurrence of Thoracosphaera operculata andThoracosphaera spp. This zone is equivalent tothe upper part of the Nephrolithus frequens Zone(Cepek and Hay, 1969; emended Romein, 1979)or CC26 of Sissingh (1977). The typical £oral as-semblages in this zone is similar to that of theunderlying Micula murus Zone, but is distin-guished by the presence of M. prinsii, the young-est representative of the Micula lineage (Roth andBowdler, 1979). At the Qreiya and Wadi Hama-ma sections, M. prinsii ¢rst occurs at 1.8 m and3.6 m respectively below the K/T boundary (Fig.2). The M. prinsii Zone encompasses the CF1(Plummerita hantkeninoides) and CF2 planktonicforaminiferal Zones and spans the last 500 kyr ofthe Maastrichtian and the interval of Chron 29R
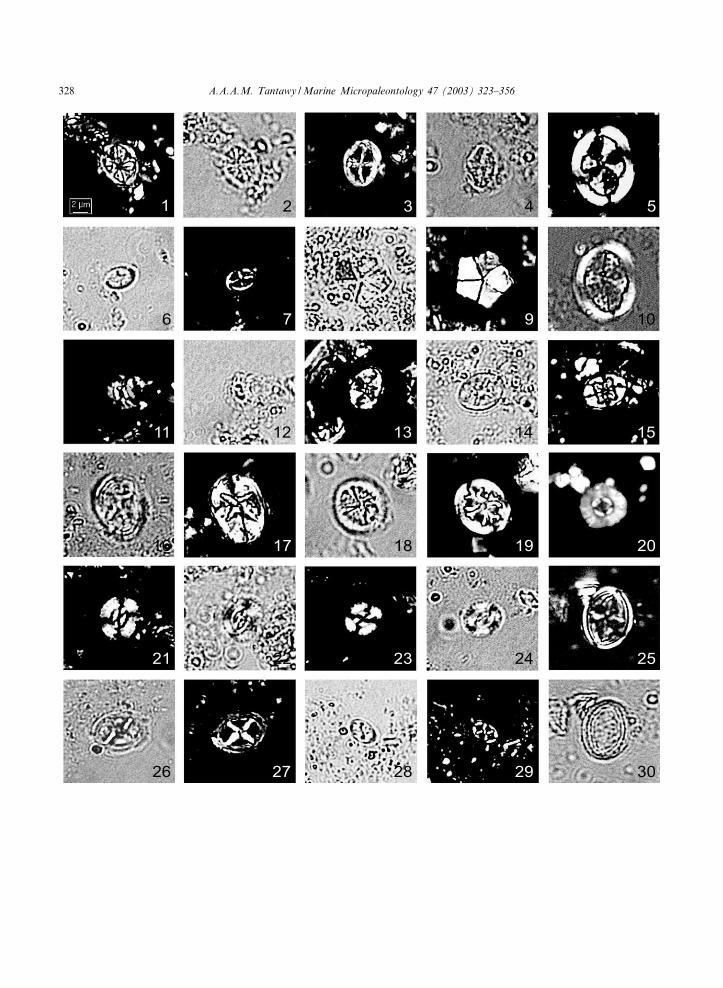
Plate I. (The scale bar for all ¢gures equals approximately 2 Wm. XPL=cross-polarized light; TL= transmitted light)
1, 2. Ahmuellerella octoradiata, sample Q-34, Micula murus Zone, 1: XPL; 2: TL.3, 4. Vekshinella stradneri, Q-20, Micula prinsii Zone, 3: XPL; 4: TL.5, 10. Arkhangelskiella cymbiformis, sample Q-12, M. prinsii Zone, 5: XPL; 10: TL.6, 7. Vekshinella crux, sample Q-14, M. prinsii Zone, 6: TL; 7: XPL.8, 9. Braarudosphaera bigelowii, sample Q44, Zone NP1b, 8: TL; 9: XPL.11, 12. Biscutum constans, sample S62, M. prinsii Zone, 11: XPL; 12: TL.13. Ei¡ellithus gorkae, sample Q-22, M. prinsii Zone, XPL.14, 15. Ei¡ellithus parallelus, sample S58, M. prinsii Zone, 14: TL; 15: XPL.16, 17. Ei¡ellithus turrisei¡elii, sample Q-12, M. prinsii Zone, 16: TL; 17: XPL.18, 19. Cretarhabdus schizobrachiatus, sample S58, M. prinsii Zone, 18: TL; 19: XPL.20. Markalius inversus, sample S14, Zone NP1c, XPL.21^24. Watznaueria barnesae, sample Q-12, M. prinsii Zone, 21 and 23: XPL; 22 and 24: TL.25, 30. Gartnerago obliquum, sample S87, M. murus Zone, 25: XPL; 30: TL.26, 27. Chiastozygus amphipons, sample Q-12, M. prinsii Zone, 26: TL XPL; 27: XPL.28, 29. Corollithion? madagaskarensis, sample Q-20, M. prinsii Zone, 28: TL; 29: XPL.
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356 329
1 2 3 4
5
10
9876
11 13 1412
15
20
19181716
21 22 23 24
25 29282726
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356330

below the K/T boundary (Henriksson, 1993; Par-do et al., 1996; Keller et al., in press). A shorthiatus is present in the lower part of the M. prinsiiZone (Figs. 2 and 3), as indicated by the absenceof planktonic foraminiferal Zone CF2 (Keller etal., in press).
4.3. Markalius inversus Zone (NP1)
The Markalius inversus Zone (Hay and Mohler,1967; emended Martini, 1970), or Zone NP1,spans from the acme of Thoracosphaera spp. tothe FO of Cruciplacolithus tenuis. This zone cor-relates to Subzone P1a of Okada and Bukry(1980), Biantholithus sparsus Zone of Perch-Niel-sen (1971), and the Zygodiscus sigmoides Zone ofPerch-Nielsen (1979a). The lower boundary ofthis zone is characterized by the ¢rst commonThoracosphaera spp. At the Wadi Hamama sec-tion, this horizon is accompanied by the FO ofNeobiscutum romeinii. At the Qreiya section, theThoracosphaera acme starts directly above theK/T boundary clay, whereas N. romeinii appears5 cm above the K/T boundary. The upper limit ofZone NP1 is marked by the FO of large (s 9 Wm)C. tenuis and Cruciplacolithus primus and occursat 6.85 m above the K/T boundary at the Wadi
Hamama section. The upper limit of this zone isnot recovered at the Qreiya section. Planktonicforaminifera indicate the presence of a hiatus inthe earliest Danian (P0/P1a) interval (Figs. 2 and3; Keller et al., in press). The nannofossil assem-blage of Zone NP1 is characterized by Late Maas-trichtian species, which may be redeposited, andby the sequential appearance of many Tertiaryspecies (see Tables 1 and 2). The ¢rst bloom ofthe survivor species Braarudosphaera bigelowii isrecorded within Zone NP1 in both sections. TheM. inversus Zone can be divided into three sub-zones using the FO of the B. bigelowii acme, andthe FO of C. primus as noted below.
4.4. Thoracosphaera imperforata Subzone(NP1a)
The Thoracosphaera imperforata Subzone isproposed here to include the interval from theFO of Thoracosphaera spp. acme to the FO ofthe Braarudosphaera bigelowii bloom. This sub-zone can be recognized in both the Qreiya andWadi Hamama sections where it spans 1.20 mand 1.35 m, respectively. Although Jiang andGartner (1986) originally placed the upper bound-ary of this subzone at the FO of Neobiscutum
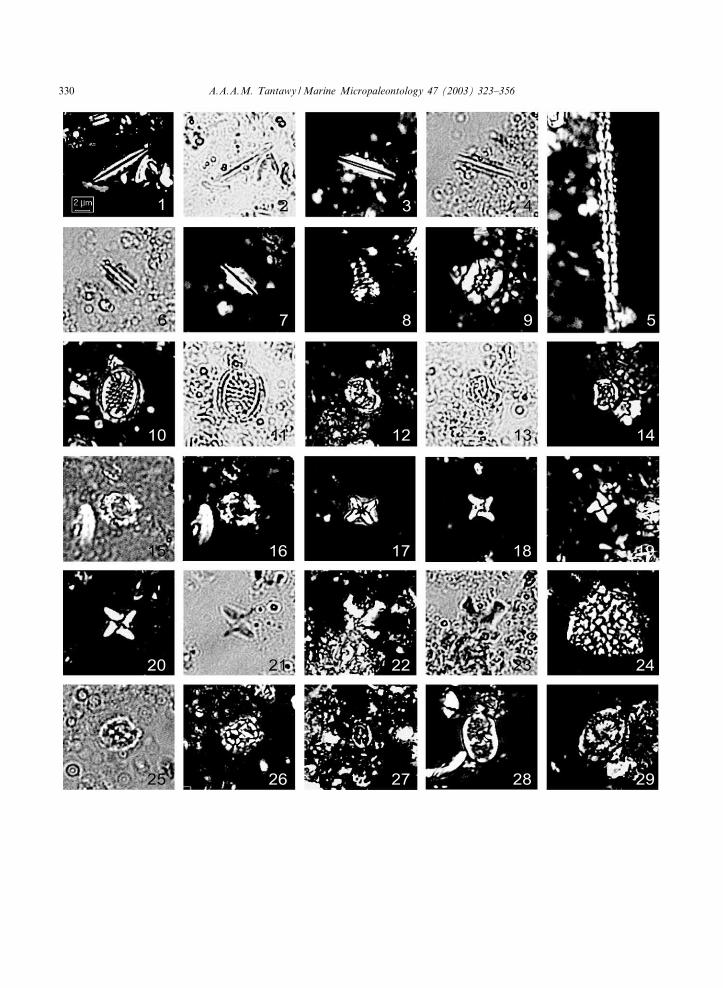
Plate II. (The scale bar for all ¢gures equals approximately 2 Wm. XPL= cross-polarized light; TL= transmitted light)
1, 2. Lithraphidites carniolensis, sample Q-14, Micula prinsii Zone, 1: XPL; 2: TL.3, 4. Lithraphidites praequadratus, sample Q-38, Micula murus Zone, 3: XPL; 4: TL.5. Microrhabdulus attenuatus, sample Q-30, M. prinsii Zone, XPL.6, 7. Lithraphidites quadratus, sample Q-40, M. murus Zone, 6: TL; 7: XPL.8. Podorhabdus decorus, sample Q-12, M. prinsii Zone, XPL.9. Cretarhabdus crenulatus, sample Q-16, M. prinsii Zone, XPL.10, 11. Cribrosphaerella ehrenbergii, sample Q-30, M. prinsii Zone, 10: XPL; 11: TL.12, 13. Nephrolithus frequens, sample Q-30, M. murus Zone, 12: XPL; 13: TL.14. N. frequens, sample Q34, M. prinsii Zone, XPL.15, 16. Cribrocorona gallica, sample S73, M. prinsii Zone, 15: TL; 16: XPL.17. Micula decussata, sample Q-22, M. prinsii Zone, XPL.18. Micula murus, sample Q-40, M. murus Zone, XPL.19. Micula murus, sample Q-12, M. prinsii Zone, XPL.20, 21. Micula murus, sample, S86, M. murus Zone, 20: XPL; 21: TL.22, 23. Micula prinsii, sample Q-16, M. prinsii Zone, 22: XPL; 23: TL.24. Thoracosphaera sp., sample S50, Zone NP1a, XPL.25, 26. Prediscosphaera cretacea, sample Q-12, M. prinsii Zone, 25: TL; 26: XPL.27. Prediscosphaera stoveri sample Q-16, M. prinsii Zone, XPL.28. Rhagodiscus angustus, sample Q-34, M. murus Zone, XPL.29. Rhagodiscus asper, sample S66, M. prinsii Zone, XPL.
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356 331
1 2 3 4 5
109876
11 12 13 14 15
2019181716
21 22 23 24 25
3029282726
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356332

romeinii, this study recommends its placement atthe ¢rst common occurrence of B. bigelowii be-cause N. romeinii is present (rarely) at the baseof this subzone and also extends downward intothe Uppermost Maastrichtian (e.g. Mai et al.,1997; Gardin and Monechi, 1998; Gardin,2002). The T. imperforata Subzone is character-ized by the abundant occurrence of Thoraco-sphaera spp., but only few to rare B. bigelowii.Species which have their ¢rst occurrences in thissubzone include N. romeinii, Neobiscutum parvu-lum, Neochiastozygus primitivus and very rareBiantholithus sparsus. This subzone is equivalentto the planktonic foraminiferal Subzones P1a andP1b of Keller et al. (1995, in press, Fig. 3).
4.5. Braarudosphaera bigelowii Subzone (NP1b)
The Braarudosphaera bigelowii Subzone (NP1b)is proposed here to include the interval from theFO of B. bigelowii acme to the FO of Cruciplaco-lithus primus. The lower boundary of this subzoneoccurs at about 1.3 meters above the K/T bound-ary at both sections and is characterized by the¢rst bloom of B. bigelowii. The nannofossil assem-blage in this subzone contains abundant B. bige-lowii in addition to the same £oral assemblagespresent in the underlying Thoracosphaera imper-forata Subzone. The B. bigelowii Subzone spans1.50 m and 1.25 m in the Qreiya and Wadi Ha-mama sections, respectively, and corresponds to
the lower part of planktonic foraminiferal Sub-zone P1c(1) of Keller et al. (1995) (Fig. 3).
4.6. Cruciplacolithus primus Subzone (NP1c)
The Cruciplacolithus primus Subzone (Romein,1979 emend. Jiang and Gartner, 1986) spans fromthe FO of C. primus to the FO of Cruciplacolithustenuis. Cruciplacolithus primus ¢rst appears at 2.85m and 2.35 m above the K/T boundary in theQreiya (sample 52) and Wadi Hamama sections(sample 28), respectively. At Wadi Hamama theC. primus Subzone spans 6.85 m. The top of thissubzone was not recovered at the Qreiya section.This subzone is equivalent to the upper part ofSubzone P1c(1) and the lower part of SubzoneP1c(2) of Keller et al. (1995) (Fig. 3). The C.primus Subzone is characterized by abundant tocommon Thoracosphaera spp., C. primus, Braaru-dosphaera bigelowii and Zygodiscus sigmoides. The¢rst appearance of Coccolithus cavus/Coccolithuspelagicus is found at the base of this subzone,and the maximum abundance of Neobiscutumparvulum occurs in the middle part of the sub-zone. Carbonate dissolution marks a 1.5-m-thickinterval within the lower part of this subzone inthe studied area.
4.7. Cruciplacolithus tenuis Zone (NP2)
The Cruciplacolithus tenuis Zone or NP2 (Hay
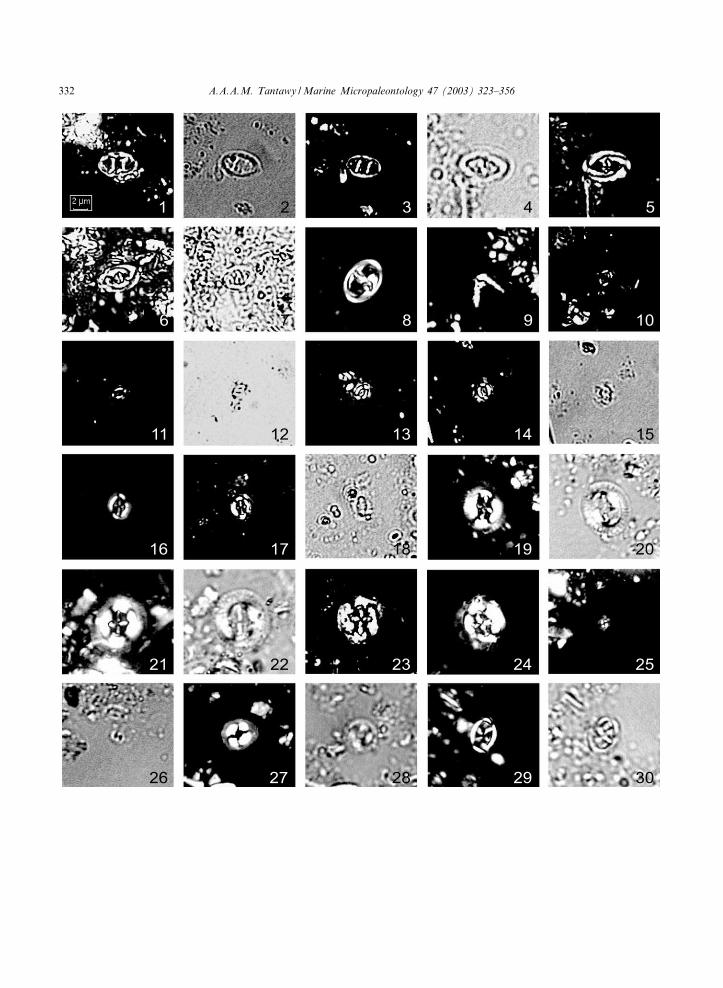
Plate III. (The scale bar for all ¢gures equals approximately 2 Wm. XPL=cross-polarized light; TL= transmitted light)
1^3. Zygodiscus compactus, sample Q-14, Micula prinsii Zone, 1 and 3: XPL, 2: TL.4, 5. Zygodiscus spiralis, sample Q-30, M. prinsii Zone, 4: TL; 5: XPL.6, 7. Z. spiralis, sample Q-20, M. prinsii Zone, 6: XPL; 7: TL.8. Zygodiscus sigmoides, sample S14, Zone Np1c, XPL.9. Ceratolithoides kamptneri, sample Q-13, M. prinsii Zone, XPL.10. Neobiscutum romeinii, sample Q-11, Zone NP1a, XPL.11, 12. N. romeinii, sample Q-10, Zone NP1a, 11: XPL; 12: TL.13^15. Neobiscutum parvulum, samples Q9, Q-11, Zone NP1a, 13 and 14: XPL; 15: TL.16. Cruciplacolithus primus, sample S14, Zone NP1c, XPL.17, 18. C. primus, sample Q90, Zone NP1c. 17: XPL; 18: TL.19, 20. Cruciplacolithus edwardsii, sample S3, Zone NP2b, 19: XPL; 20: TL.21, 22. C. edwardsii, sample S4, Zone NP2b, 21: XPL; 22: TL.23, 24. Cruciplacolithus tenuis, sample S12, S4, Zone NP2, XPL.25, 26. Prinsius africanus, sample S4, Zone NP2b, 25: XPL; 26: TL.27, 28. Coccolithus pelagicus, sample S2, Zone NP2b, 27: XPL; 28: TL.29, 30. Neochiastozygus primitivus, sample S2, Zone NP2b, 29: XPL; 30: TL.
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356 333
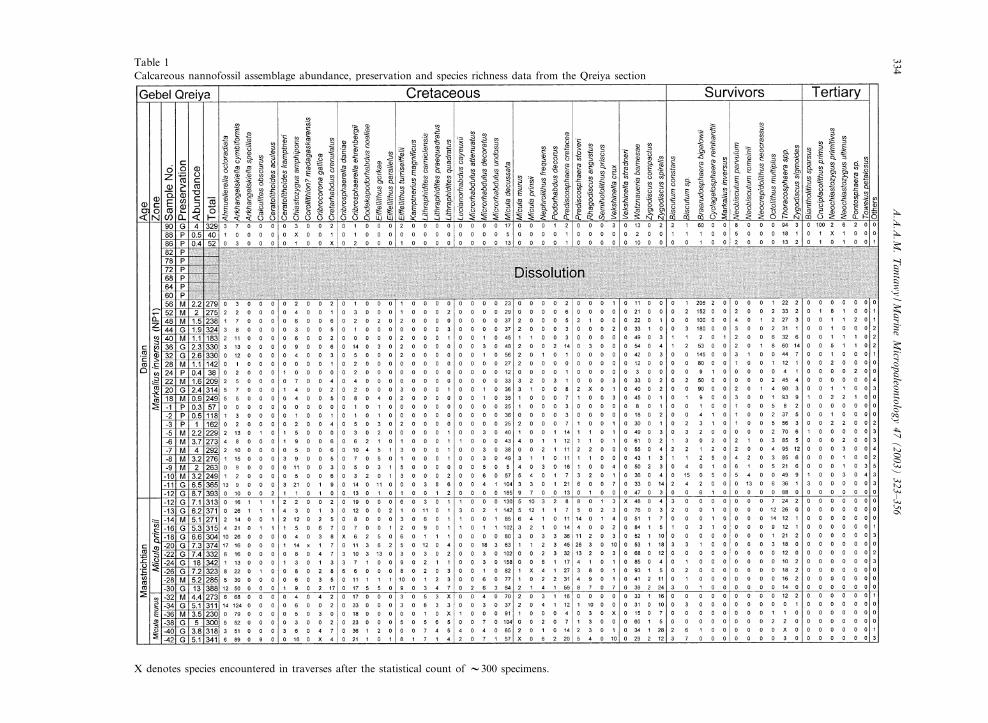
Table 1Calcareous nannofossil assemblage abundance, preservation and species richness data from the Qreiya section
X denotes species encountered in traverses after the statistical count of V300 specimens.
MARMIC
90527-1-03
A.A.A.M.Tantaw
y/M
arineMicropaleontology
47(2003)
323^356334
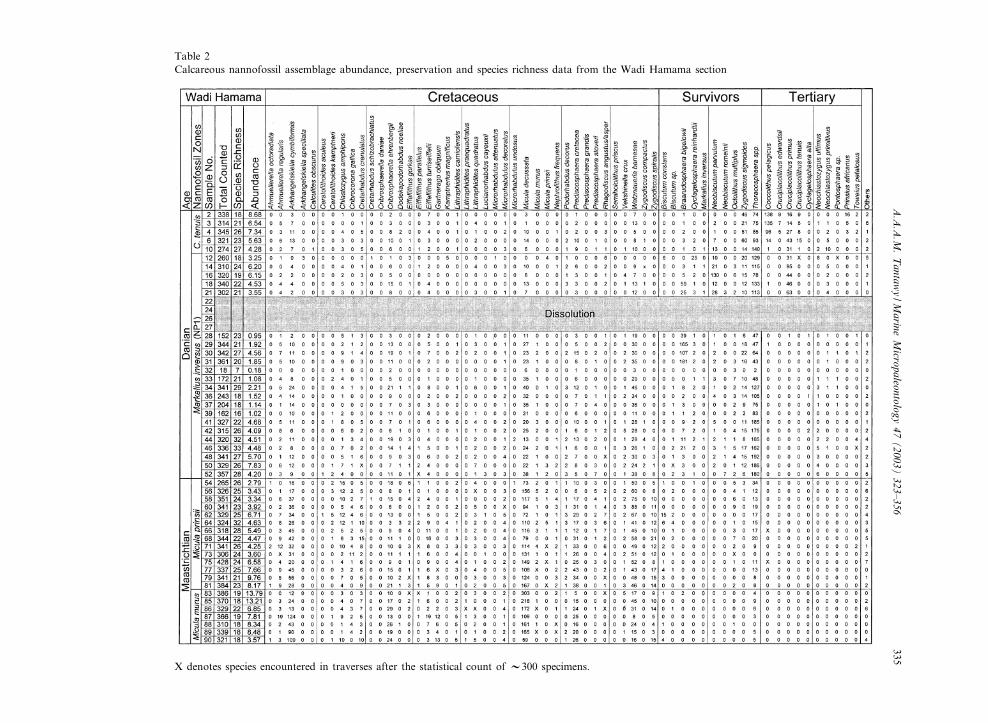
Table 2Calcareous nannofossil assemblage abundance, preservation and species richness data from the Wadi Hamama section
X denotes species encountered in traverses after the statistical count of V300 specimens.
MARMIC
90527-1-03
A.A.A.M.Tantaw
y/M
arineMicropaleontology
47(2003)
323^356335
Datum Events
Li &Keller(1998); Keller etal. (1995) Tunisia
P. hantkeninoides(CF1)
G. gansseri
P. hantkeninoides
Age (Ma)
Abathomphalusmayaroensis 65.45
65.30
65.00
P. hariaensis (CF3)
P. hariaensis 66.83
G. cretacea (P 0)
P. eugubina (Pα)
Berggren et al. (1995)
P1c(1)
P1c(2)
P1d
P1b
P. eugubina 64. 97
S. triloculinoides(>100um)
64. 7
P. inconstans 63.0
Central Eastern Desert, Egypt
P. hariaensis(CF3)
P1d
Markaliusinversus (NP1)
Cruciplacolithus tenuis (NP2)
Tetralithus murus/Nephrolithus
frequens(NC23)
Roth, (1978)Okada & Bukry
(1980)
Nephrolithusfrequens (CC26) M. prinsii
N. frequens
Thoracosph. Acme
C. tenuis
CP1
b
a
Calcareous Nannofossils Planktic Foraminifera S
tage
Ma
astr
ich
tian
P1a
M. trinidadensis
S. varianta
P1cP1c
62.2
Da
nia
n
C. primus (small)
a
b
M. murus
N. primitivus
B. bigelowiiAcme
C. primus (small)
P1b
P. hantkeninoides(CF1)
P. palpebra (CF2)
G. cretacea (P 0)
P1c(1)
P1c(2)
P1b
P1a
Zone Zone
N. parvulum
N. romeinii
C. pelagicus
C. edwardsii
C. pelagicusAcme
NP
2
NP
1
NP2a
NP2b
NP1c
NP1b
NP1a
Ne
ph
rolit
hu
s f
req
ue
ns
(C
C26)
C. primus (large)
P. petalosusP. petalosus
C. ultimus
C. edwardsii
C. edwardsii
C. primus
ThoracosphaeraAcme
Hiatus Hiatus
P1a
P1c
64. 5
P. logiapertura
P. logiapertura
P. eugubina
HiatusHiatus
Micula prinsii
Micula murus
N. romeinii
N. parvulum
N. parvulum
P. af ricanus
Sissingh (1977)Martini(1971)
Perch-Nielasen(1979, 1981a,b)Mediterranean
(This study)
Datum Events
Fig. 3. Correlation of the proposed calcareous nannofossil and planktonic foraminiferal zonation with commonly used zonal schemes (Roth, 1978). Ages accordingto Berggren et al. (1995) and Li and Keller (1998a).
MARMIC
90527-1-03
A.A.A.M.Tantaw
y/M
arineMicropaleontology
47(2003)
323^356336

and Mohler, 1967; emended Martini, 1970) spansthe interval from the FO of C. tenuis to the FO ofChiasmolithus danicus. This zone was only recov-ered at the Wadi Hamama section (samples from9^1), where the lower part of the zone is charac-terized by C. tenuis and common Thoracosphaeraspp., Zygodiscus sigmoides, Cruciplacolithus pri-mus (large, s 9W), few to rare Coccolithus pelagi-cus, Neobiscutum parvulum, Braarudosphaera bige-lowii and rare re-deposited Cretaceous species(Table 2). The upper part of this zone is markedby the ¢rst appearance of Prinsius africanus, Cru-ciplacolithus edwardsii and dominated by C. pela-gicus. The sudden increase of the latter species canbe used to subdivide this zone into SubzonesNP2a and NP2b. Zone NP2 corresponds to theupper two thirds of planktonic foraminiferal Sub-zones P1c(2) and P1d (Fig. 3).
5. Calcareous nannofossil assemblages
Sixty-three calcareous nannofossil species from40 genera were found in the course of this study(Tables 1 and 2). Among these, three distinct spe-cies groups can be recognized: (a) vanishing orCretaceous species, (b) persistent or survivor spe-cies, and (c) incoming or Tertiary species (Figs. 4and 5). Similar species groups have been distin-guished by other workers (e.g. Percival and Fisch-er, 1977; Perch-Nielsen et al., 1982; Jiang andGartner, 1986; Pospichal, 1991, 1995; Pospichaland Bralower, 1992).
5.1. Vanishing species
Vanishing species include taxa that are consid-ered to have become extinct before or at the endof the Maastrichtian. About 40 typical Cretaceousspecies were recorded in both sections (Tables 1,2). The most abundant are Arkhangelskiella cym-biformis, Micula decussata, Watznaueria barnesae,Prediscosphaera cretacea, and Cribrosphaerella eh-renbergii. Species that are common include Ei¡el-lithus turrisei¡elii, Chiastozygus amphipons, Lith-raphidites carniolensis, Ahmuellerella octoradiata,and Zygodiscus spiralis. All other species arequantitatively insigni¢cant. The vanishing Creta-
ceous nannofossil assemblages encompass 93% to95% of the total assemblages in the Micula murusZone and gradually decrease in the Micula prinsiiZone by about 10% (Figs. 4 and 5). Previousstudies reported little or no changes in Cretaceousnannofossil assemblages below the K/T boundary(e.g. Perch-Nielsen et al., 1982; Romein, 1977;Thierstein, 1981; Jiang and Gartner, 1986; Pospi-chal, 1994). Thus the decrease in the Eastern Des-ert of Egypt may be a local phenomenon.
A major change is observed in calcareous nan-nofossil assemblages across the K/T boundary asobserved in other sections worldwide (e.g. Pospi-chal, 1994, 1995). In the Qreiya and Hamamasections, this £oral turnover is marked by a dra-matic decrease in the relative abundance of com-bined Cretaceous species and a corresponding in-crease in the survivor group. At the Hamamasection, Cretaceous nannofossils are reduced by50% at the K/T boundary, average 40% in Sub-zone NP1a and 10^20% in Subzones NP1b and cand Zone NP2 (Fig. 4). But at the Qreiya sectiona more gradual transition is indicated by a de-crease of 25% in the Cretaceous nannofossil as-semblage in the 15 cm overlying the K/T bound-ary, and a gradual decrease to 40% in SubzoneNP1a (Fig. 5).
The di¡erence between the two sections is likelyto be due to erosion and reworking in the EarlyDanian. For example, the basal Danian forami-niferal Zone P0 and most of Subzone P1a (Parvu-larugogloberina eugubina Zone), which span thelower part of Subzone NP1a, are missing at theHamama section, whereas at Qreiya Zone P0 isreduced to the red clay layer and P1a is only 50cm thick (Figs. 4 and 5; Keller et al., in press).Reworked planktonic foraminifera are abundantimmediately above the K/T boundary at theQreiya section, and similar reworking of calcare-ous nannofossils may partly account for the over-all higher percentage of nannofossil assemblagesrelative to Hamama. Reworking in this interval isindicated by abundant reworked foraminifera(Keller et al., in press). Anomalous Lower Danianpeaks in Cretaceous assemblages at both sectionsare also due to reworking and re-deposition par-ticularly in intervals of hiatuses (e.g. at 11.5 m atQreiya and 11 m at Wadi Hamama).
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356 337

The observed pattern of K/T and Lower Dan-ian Cretaceous species assemblage variations isconsistent with previous quantitative studies thatshow a gradual decrease up-section within NP1and NP2. For example, Pospichal and Wise(1990) documented this pattern at the AntarcticOcean ODP Site 690 Perch-Nielsen et al. (1982),and Keller and von Salis-Perch-Nielsen (1995) atEl Kef, and Jiang and Gartner (1986) at BrazosRiver, Texas.
5.2. Survivors
Survivors are de¢ned as Cretaceous species that
survived into the Paleocene or have direct de-scendants surviving into the Paleocene (Perch-Nielsen et al., 1982). Eleven survivor specieswere recorded in Danian sediments at Qreiyaand Hamama, including Biscutum constans, Bis-cutum sp., Braarudosphaera bigelowii, Cyclagelo-sphaera reinhardtii, Markalius inversus, Neobis-cutum parvulum, Neobiscutum romeinii, Neocrepi-dolithus neocrassus, Octolithus multiplus, Thoraco-sphaera spp., and Zygodiscus sigmoides (Tables 1and 2). The rare occurrence of N. romeinii andN. parvulum were previously recorded in theMaastrichtian sediment by several authors, e.g.in the Geulhemmerberg section, the Netherlands
Dakh
la F
orm
atio
n
Ma
ast
rich
tian
Da
nia
nS
tage
Form
atio
n
Th
ickn
ess
Lith
olo
gy
Sa
mp
le N
o.
M. p
rinsi
iM
. m
uru
sN
P1
bc
a
Survivors
Gebel Qreiya
Percent Abundance (%)
K/T
12
Q-40
Q-35
Q20
Q30
Q80
Q90
Q40
Q50
Q60
Q70
Q-30
Q-25
Q-20
Q-15
Q-10
Q-5
Q-1
(m)
1
0
2
3
4
5
6
7
8
9
10
11
Cretaceous species
Tertiary species
Dissolution
Q-42
Pla
nkt
icF
ora
minife
raC
F3
CF
1
P1a
P1b
P1
C(1
)P
1C
(2)
P1
C(1
)
Ca
lca
reo
us
Na
nn
ofo
ssils
60 8020 40
Fig. 4. Percent abundance of Cretaceous, survivor and Tertiary nannofossil assemblages across the K/T boundary in the GebelQreiya section.
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356338

(Mai et al., 1997), and the Elles section, Tunisia(Gardin, 2002). These species are present onlyabove the K/T boundary at the Qreiya and Ha-mama sections. The survivor species are generallyrare (6 3%) in the Micula murus Zone, though amarked increase in the upper Micula prinsii Zoneimmediately below the K/T boundary is observedin both sections (Figs. 4 and 5). Above the bound-ary, the survivor assemblage (here mainly Thora-cosphaera and B. bigelowii) shows a rapid and
progressive increase in the relative abundance toabout 40% near the NP1a/b Subzonal boundaryand a further increase in Subzone NP1c to about70% in both sections (Figs. 4 and 5).
5.3. Tertiary species
Tertiary species are de¢ned as species that ¢rstappear at or above the K/T boundary. A total ofeleven Tertiary species are identi¢ed (Tables 1 and
1
5
10
15
20
25
30
35
40
50
60
70
75
80
85
90
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
Wadi Hamama
Cretaceous
Survivors
K/T
Percent Abundance (%)
Tertiary
Dissolution
60 8020 40
Fig. 5. Percent abundance of Cretaceous, survivors and Tertiary nannofossil assemblages across the K/T boundary in the WadiHamama section.
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356 339

2), including common to abundant Cruciplacoli-thus primus and Coccolithus cavus/pelagicus. Otherspecies are rare or occur sporadically, includingCruciplacolithus tenuis, Cruciplacolithus edwardsii,Cyclagelosphaera alta, Neochiastozygus ultimus,Neochiastozygus primitivus, Prinsius dimorphosus/tenuiculum, Prinsius africanus, and Toweius petal-osus. The Tertiary assemblage is a minor compo-nent (V2^3%) of the Danian nannofossil assem-blages in Subzones NP1a and NP1b, increases toabout 20% in the upper part of Subzones NP1cand NP2a, and a rapid increase to 56% in theupper part of Zone NP2 (Figs. 4 and 5).
6. Nannofossil abundance and species richness
6.1. Species richness (number of species persample)
6.1.1. Cretaceous speciesSpecies richness in the Cretaceous assemblage
varies considerably below the K/T boundary. Atthe Qreiya section, the species richness of the Cre-taceous assemblage decreases gradually from 27at the base of the section to 14 species withinthe middle part of Micula murus Zone, then re-turns gradually to 27 species at the top of this
Q90
Q70
Q80
Q60
Q50
Q40
Q20
Q30
Q-10
Q-1
Q-5
Q-15
Q-25
Q-30
Q-35
Q-40
Q-20
Q-42
1
0
2
3
4
5
6
7
8
9
10
11
12
(m)
Nannofossil Abundance(Specimens/Field of view)
Gebel Qreiya section
Cretaceousspecies
Survivor +Tertiary species
Species Richness (Species/Sample)
Q-1.5
Dissolution
10 20 5 10 15
Fig. 6. Calcareous nannofossil abundance and species richness across the K/T boundary in the Gebel Qreiya section.
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356340

zone. Within the Micula prinsii Zone, the speciesrichness shows a gradual decrease in the lowerpart, an abrupt increase at 85 cm below the K/Tboundary, and £uctuates around 25 species up tojust below the boundary.
At the Wadi Hamama section, the species rich-ness increases slightly from the base upward andaverages 18 species within the Micula murus Zoneand 21 species within the Micula prinsii Zone witha peak of 27 species at 45 cm below the K/Tboundary. The di¡erences in species richness re-corded at the Gebel Qreiya and Wadi Hamama
sections may be due to the lithologic changes,preservation and more shallow-water conditionsin the Wadi Hamama area during the Late Maas-trichtian M. murus and M. prinsii Zones.
Species richness at Qreiya shows a progressivedecrease from 27 to 17 species within the upper-most 20 cm of the Maastrichtian, then increasesagain to 22 species at 5 cm above the K/T bound-ary (Figs. 6 and 7). This trend is not recorded atWadi Hamama, where the species richness re-mained similar below and above the boundary.This di¡erence may be due to the hiatus in the
1
5
10
15
20
25
30
35
40
50
60
70
75
80
85
90
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
Wadi Hamama section
Nannofossil Abundance(Specimens/Field of view)
Cretaceousspecies
Survivor +Tertiary species
Species Richness (Species/Sample)
Dissolution
1020 510
Fig. 7. Calcareous nannofossil abundance and species richness across the K/T boundary in the Wadi Hamama section.
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356 341

basal Danian, where most of Subzone P1a andlowermost part of Subzone NP1a are missing(see Figs. 2 and 7).
In both sections, the species richness of the Cre-taceous assemblage shows a general decreasingtrend with a strong oscillation in the Lower Dan-ian and the presence of nine species at the top ofboth sections (Figs. 8 and 9). The species abun-dance changes possibly as a result of dissolution,and intermittent reworking and re-depositionwithin the lower part of Zone NP1.
6.1.2. Survivors and Tertiary speciesThe species richness of the survivor assemblage
averages about 2.5 species in the Maastrichtian atboth sections and increases slightly just below theK/T boundary (Figs. 6 and 7). Survivor and Ter-tiary species combined at the Qreiya section showan abrupt increase in species richness across theboundary and £uctuate between 5 and 10 speciesin the Danian. However, at Wadi Hamama, thesurvivors and Tertiary species show a more grad-ual increase in the species richness across theboundary and an increase from two species at45 cm below the K/T boundary to 13 species at45 cm above the boundary (Fig. 7). Fluctuationsin the combined survivor and Tertiary speciesrichness parallel to those of Cretaceous speciesare also observed in the upper part of SubzoneNP1a and lower part of Subzone NP1b. Carbon-ate dissolution and poor preservation appear tobe responsible for these £uctuations. In the upperpart of Subzone NP1b and Zone NP2, speciesrichness is constant, averaging nine species persample (Fig. 7).
6.2. Nannofossil abundance (specimens per ¢eld ofview)
The relative abundance of nannofossils, calcu-lated by counting the number of specimens per¢eld of view (s/fv), provides a measure of environ-mental change. At Qreiya the relative abundanceof the Cretaceous assemblages in the UpperMaastrichtian Micula murus Zone £uctuates be-tween four and ¢ve specimens per ¢eld of view.In two intervals in the Micula prinsii Zone at1.25 m and 1.85 m, the relative abundance reaches
17 and 12 s/fv, respectively (Fig. 6). These peakvalues were not observed at the Wadi Hamamasection, in which the Cretaceous assemblagesshow variable abundances below the K/T bound-ary parallel to the lithologic changes. In the lattersection, the minimum abundance is recordedwithin a sandstone marker bed at the base ofthe section at the base of M. murus Zone, sample90, 3.4 s/fv, and appears to be due to poor pres-ervation. Maximum abundance of 13.5 s/fv isreached in the marls within the uppermost twometers of the M. murus Zone (Fig. 7).
An abrupt decrease in the Cretaceous assem-blage abundance from seven to one s/fv occursin the 25-cm interval above the K/T boundaryat the Qreiya section (Fig. 6). On the otherhand, a gradual decline in nannofossil abundancefrom six s/fv at 45 cm below the K/T boundary(sample 62) to 0.5 s/fv at 60 cm above the bound-ary (sample 39) is observed at the Wadi Hamama(uppermost Micula prinsii Zone and SubzoneNP1a, Fig. 7).
Up-section in the Danian, the abundance of theCretaceous assemblage is very low and £uctuatesbetween 0 and 1.5 s/fv. The combined relativeabundance of survivor and Tertiary assemblagesis very low (6 1 s/fv) below the K/T boundary,and peaks just above the boundary (2 s/fv) atQreiya and (5 s/fv) at Wadi Hamama. This di¡er-ence may be due to the shallower depositionalenvironment and possibly higher salinity in thesouthern Wadi Hamama area, which resulted inthe bloom of survivor and opportunistic Thora-cosphaera spp. during the earliest Danian. Therelative abundance of survivors and Tertiaryspecies combined at Wadi Hamama increasesgradually upward to reach eight s/fv with minor£uctuations at the top of the section (upper partof Zone NP2, Fig. 7).
7. Paleoecology of selected species
7.1. Late Maastrichtian
During the Late Maastrichtian, calcareous nan-nofossil assemblages in central Egypt are similarto assemblages reported from low to mid-latitudes
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356342

Fig. 8. Relative abundances of selected calcareous nannofossil species across the K/T boundary in the Gebel Qreiya section.
MARMIC
90527-1-03
A.A.A.M.Tantaw
y/M
arineMicropaleontology
47(2003)
323^356343

Fig. 9. Relative abundances of selected calcareous nannofossil species across the K/T boundary in the Wadi Hamama section.
MARMIC
90527-1-03
A.A.A.M.Tantaw
y/M
arineMicropaleontology
47(2003)
323^356344

(Thierstein, 1981) in that they contain a mixtureof low and high-latitude species.Micula decussata,the dominant species, averages between 20% and30% of the total nannofossil assemblage (Figs. 8and 9). Four well-developed peaks of M. decus-sata are observed at Qreiya and Hamama. Thelower two peaks occur in the middle and upperMicula murus Zone, and the upper two peakswithin the middle and upper Micula prinsii Zone(Figs. 8 and 9). The M. decussata blooms belowthe K/T boundary have also been recognized inother Upper Maastrichtian sections (e.g. Braggs,Alabama (Worsley, 1974); San Tolmo, Spain(Percival and Fischer, 1977); Brazos River andLittig Quarry, Texas (Jiang and Gartner, 1986);Hor Hahar, Israel (Eshet et al., 1992; Eshet andAlmogi Labin, 1996); Gubbio, Italy, and DSDPsite 577A (Gardin and Monechi, 1998); Elles, Tu-nisia (Gardin, 2002)).Micula decussata has been reported as a major
constituent of Late Cretaceous tropical and sub-tropical assemblages (Thierstein and Haq, 1977;Wind, 1979). Its distribution may in fact bethe result of latitudinal preservation patterns,possibly related to fertility. According to Thier-stein (1980, 1981), M. decussata is a highly dis-solution-resistant form and is considered a goodindicator of poor nannofossil preservation anddiagenetic enhancement (see also Roth, 1983;Pospichal, 1991; Eshet and Almogi Labin,1996). However, in the two sections examinedfrom central Egypt, the high abundance ofM. decussata does not appear to be an artifactof dissolution because most other species, in-cluding dissolution-prone forms (e.g. Biscutumconstans, Prediscosphaera cretacea and Cribro-sphaerella ehrenbergii), are well-preserved. Simi-larly, abundant M. decussata was reported byMoshkovitz and Eshet (1989) and Eshet et al.(1992) from Israel in samples with good preser-vation and no evidence of strong dissolutionor overgrowth. Hence, the high abundances ofM. decussata in well-preserved assemblages inUppermost Maastrichtian sediments in sectionsin the Negev, Egypt and elsewhere could re£ecta natural increase in abundance (Pospichal, 1991)due to high-stress marine environments (Eshet etal., 1992). There is other support for this interpre-
tation. Stable isotope data from the Qreiya sec-tion indicate very low surface productivity formost of the Micula murus and the lower anduppermost parts of the Micula prinsii Zones(Fig. 10; Keller et al., in press), and planktonicforaminiferal assemblages are dominated by thedisaster species Guembelitria cretacea.
Following Thierstein (1980), the studied sec-tions show the presence of some warm-water in-dicators such as Micula murus, Micula prinsii,Lithraphidites quadratus, and Watznaueria barne-sae.Watznaueria barnesae comprises up to about
29% of the nannofossil assemblage in the Upper-most Maastrichtian Micula prinsii Zone at Qreiya(Figs. 8 and 9), and re£ects a global warmingtrend identi¢ed in planktonic foraminiferal ZoneCF1 based on oxygen isotopes (Li and Keller,1998c). In the Micula murus Zone, W. barnesaedoes not exceed more than 12% of the total nan-nofossil assemblage, except for sample Q-38,where this species reaches 20% in an intervalmarked by poor preservation, which could explainthe high abundance of this very dissolution-resis-tant species.
Several species are commonly recognized ascool-water indicators. The cool-water species Ar-khangelskiella cymbiformis is relatively abundantin the Micula murus Zone in both sections (Figs. 8and 9) and re£ects a major Late Maastrichtianglobal cooling event (Barrera, 1994; Li and Kel-ler, 1998c). There is a signi¢cant decrease in thesize of A. cymbiformis specimens towards the topof the Maastrichtian in the Micula prinsii Zone, inagreement with previous ¢ndings of Girgis (1989)and Faris (1995b) in Egypt and of Gardin (2002)in Tunisia. The decrease in size of this speciescoincides with warming as indicated by stable iso-topes data (Barrera, 1994; Li and Keller, 1998c).Another cool-water taxon, Nephrolithus frequens,is sporadically present in low abundance (6 1%).This species was previously recorded from otherLate Maastrichtian sections in Egypt by Sha¢kand Stradner (1971), Perch-Nielsen et al. (1978)and Faris (1997). Prediscosphaera stoveri, anothercool-water species, increases in abundance in highlatitudes (ODP Sites 690, 738, and 752) during theLate Maastrichtian (e.g. Pospichal and Wise,
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356 345
CF3
CF1
P1a
P1
c(1
)
2
3
4
5
6
7
8
9
10
11
La
te M
aastr
ich
tia
nE
arl
y D
an
ian
m
Qreiya, Egypt
P1
c(1
)
M.
mu
rus
M.
pri
nsii
NP
1a
NP
1b
NP
1c
P1b
68
6460565248444036
32282420-1-3-5-7-9-11
-14-16-18-20-22-24-26
-28-30
-33
-34
-35
-36
-37
-38
-39
-40
-41
-42
δ13C
-0 .5 0
Permil (PDB)-1 -5 -4 -3 -2
Permil (PDB)
-8 -7 -6
δ18O
0 0 50
PercentPercent
30 300 400 30
A.
cy
mb
ifo
rmis
M.
de
cu
ssata
W.
barn
esae
Th
ora
co
sp
hae
ra s
pp
.
Bra
aru
do
sp
hae
ra b
ige
low
ii
Calcareous Nannofossils
selected species
Calcareous Nannofossils
selected species
Cretaceous
Terttiary +Survivor
5 10 15 20 25
Number of Species
Species
Richness
1
K/ T
Rugoglobigerina rugosa (planktic)
Cibicidoides pseudoacutus
Hiat us
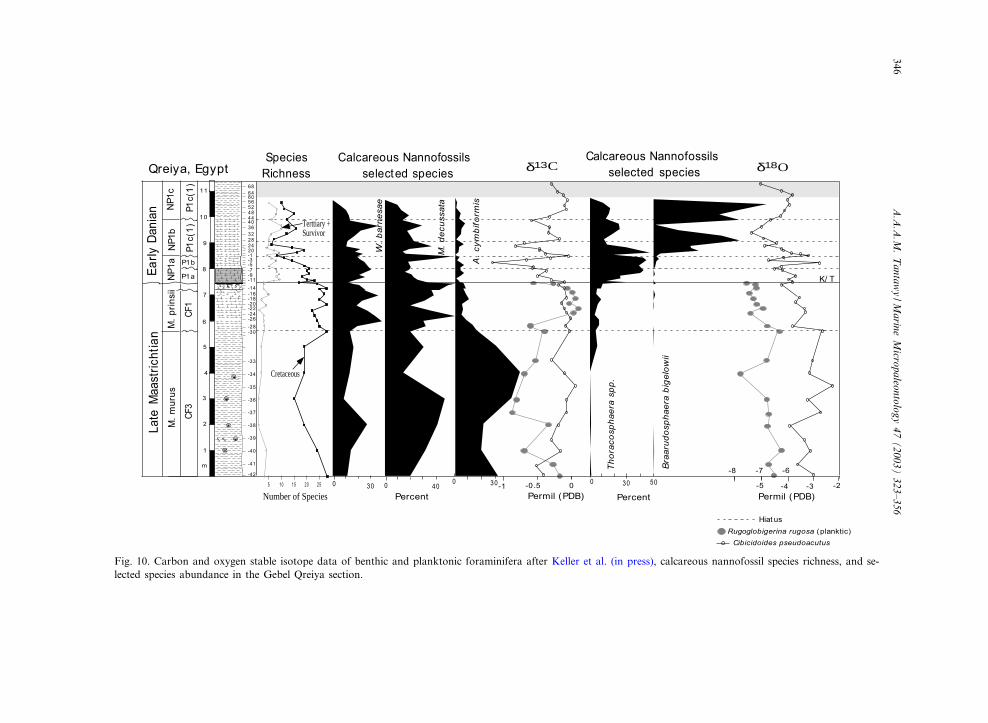
Fig. 10. Carbon and oxygen stable isotope data of benthic and planktonic foraminifera after Keller et al. (in press), calcareous nannofossil species richness, and se-lected species abundance in the Gebel Qreiya section.
MARMIC
90527-1-03
A.A.A.M.Tantaw
y/M
arineMicropaleontology
47(2003)
323^356346

1990; Pospichal, 1991; Wei and Pospichal, 1991;Pospichal and Bralower, 1992) and reaches nearly70% below the K/T boundary at ODP Sites 690and 738. In low-latitude continental shelf sections,this species is common (V15%) near the top ofthe Maastrichtian as observed at Brazos River,Texas, and El Kef, Tunisia (Jiang and Gartner,1986). Peak abundance of P. stoveri in the Qreiyasection is only about 7% in sample Q20, about 85cm below the K/T boundary (Fig. 8).
Other cold-water species present in centralEgypt include Ahmuellerella octoradiata, Biscutumconstans, Ei¡ellithus turrisei¡elii, and Kamptneriusmagni¢cus. The presence, though low abundance,of this cool-water or high-latitude assemblage(Thierstein, 1981; Pospichal, 1991) in the Miculaprinsii Zone suggests cooler conditions near theend of the Maastrichtian.
Thus the Upper Maastrichtian relative abun-dances of preserved cool and warm-water speciessuggest alternating cool/warm £uctuations duringthe latest Maastrichtian (Fig. 10).
7.2. Cretaceous/Tertiary and Early Danian
The K^P and Lower Danian intervals at Qreiyaand Wadi Hamama show a succession of bloomsof Thoracosphaera spp., Braarudosphaera bigelo-wii, Neobiscutum parvulum, Cruciplacolithus pri-mus, Zygodiscus sigmoides, and Coccolithus pela-gicus. Blooms of Thoracosphaera are known fromseveral middle to lower-latitude sites (Percival andFischer, 1977; Perch-Nielsen et al., 1982; Jiangand Gartner, 1986; Gardin and Monechi, 1998).In the Eastern Desert of Egypt, this species showsa sudden increase in relative abundance to 57%that parallels a decrease in Cretaceous speciesrichness in the 120 cm above the K/T boundary(Tables 1 and 2; Figs. 8 and 9). Thoracospahera isconsidered an opportunistic species that toleratesunusual marine conditions, including majorchanges in primary productivity (Eshet et al.,1992). Recent representatives of Thoracosphaerain the Gulf of Mexico were commonly observedin surface waters with high salinity values (Gaard-er and Hasle (1971).
During Danian Zone NP1/NP2 transition,Thoracosphaera also dominates, reaching about
V51% of the total assemblage in the Wadi Ha-mama section (samples 10^12) and 28% in theQreiya section (sample Q90) (see Tables 1 and 2,and Figs. 8 and 9). A similar Thoracosphaerabloom was observed by Eshet et al. (1992) atthe same stratigraphic level. Eshet et al. (1992)assumed that the presence of both Thoracosphaeraand re-deposited material indicates a second ma-rine regression in the Early Tertiary.
The taxon Braarudosphaera bigelowii reaches apeak abundance of about 25% at 120 cm and 180cm above the boundary at Qreiya and Wadi Ha-mama, respectively (Figs. 8 and 9).
Although Braarudosphaera bigelowii bloomsjust after the bloom of Thoracosphaera in severalmid^low-latitude sections (e.g. Brazos, Texas,Jiang and Gartner, 1986; Agost, Pospichal,1995; Zumaya, Spain, Lamolda and Gorostidi,1992; Biarritz, France, Perch-Nielsen, 1979c; ElKef, Tunisia, Gardin and Monechi, 1998), thistaxon is absent from other low-latitude K/T sec-tions, including all sections in Israel (Magaritz etal., 1985; Eshet et al., 1992), DSDP Site 577A(Gardin and Monechi, 1998), and Elles, Tunisia(Gardin, 2002). Braarudosphaera bigelowii thusseems to respond to di¡erent and, as yet un-known, paleoenvironmental conditions in middleand low-latitude locations. It has been variouslysuggested that B. bigelowii blooms indicate near-shore depositional environments characterized byrelatively shallow water (Roth and Thierstein,1972; Mu«ller, 1985), low salinity (Bukry, 1974),increased fresh-water in£ux and lowered salin-ities after the K/T boundary (Thierstein and Ber-ger, 1979; Askin, 1992; Barrera and Keller, 1994),or eutrophication (Cunha and Shimabukuro,1996).Cyclagelosphaera reinhardtii is very rare in the
Qreiya and Hamama sections, as was also ob-served at El Kef, Tunisia, and Agost, Spain, byPospichal (1995). In sections of northern Spainand southern France (Seyve, 1990; Lamolda andGorostidi, 1992), where blooms of C. reinhardtiiare observed, this species is restricted to a veryshort interval between the initial peaks of Thora-cosphaera and Braarudosphaera.
The Zygodiscus sigmoides bloom or acme ismost pronounced in high-latitude sites (e g. Den-
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356 347

mark, Perch-Nielsen, 1979b; ODP Site 690,Pospichal and Wise, 1990; Site 752, Pospichal,1991). This species, however, is rare in Lower Pa-leocene assemblages from middle to low-latituderegions (e.g. Biarritz, France, Perch-Nielsen,1979c; Gubbio, Monechi, 1979; Zumaya, Percivaland Fischer, 1977). In the Qreiya and Hamamasections, an increase in the abundance of Z. sig-moides is observed in Zone NP2, where it rangesfrom 4% to 13% of the total nannofossil assem-blage.Neobiscutum romeinii and Neobiscutum parvu-
lum, two very small forms, are usually present invery low abundances in Subzones NP1a andNP1b. The latter species is more dominant inthe middle part of Subzone NP1c, where itreaches 40% of the assemblage at Wadi Hamama(sample 16) about 2 m above the K/T boundary(Fig. 9). Perch-Nielsen (1985) observed that N.romeinii and N. parvulum are present in the Te-thys region but not in higher northern or southernlatitudes.Cruciplacolithus primus (6 5 Wm) is very rare in
the basal part of Subzone NP1c of the Qreiya andHamama sections and very abundant just abovethe dissolution interval, reaching a maximum of30% of the total nannofossil assemblage at 16 mabove the base of both sections. At Wadi Hama-ma, both small (6 5 Wm) and larger (6 9 Wm)morphotypes are present in Zone NP2 (Fig. 9),where they reach a combined abundance of 5%to 13%. The C. primus bloom has been recordedin many low and middle-latitude sections by var-ious workers (e.g. Pospichal, 1991, 1995; Gardinand Monechi, 1998). Gardin and Monechi (1998)and Gardin (2002) suggested that the appearanceof C. primus marks the onset of the return tomore stable environmental conditions after theperturbations at the K/T boundary.Coccolithus pelagicus is relatively abundant at
the top of the Wadi Hamama section, where itreaches more than 40% of the total nannofossilassemblage (Fig. 9). Wei and Pospichal (1991)observed that C. pelagicus decreased in abundancetowards higher latitudes. In the modern ocean, C.pelagicus is most abundant in the cool waters ofthe northern high latitudes (McIntyre and Be¤,1967). This was interpreted by Haq and Lohmann
(1976) as indicating that C. pelagicus has under-gone an evolutionary shift in its ecological re-sponse.
8. Discussion
8.1. Productivity
Eshet et al. (1992) and Eshet and Almogi Labin(1996) used calcareous nannofossil species abun-dances as indicators of productivity based on thecomparison of their distribution with independentdata from other sources, such as planktonic fora-minifera (Almogi Labin et al., 1993) and organic-walled dino£agellate cysts (Eshet et al., 1994).They concluded that nannoplankton reach theirhighest species diversity and abundance in inter-vals of lowered productivity and that relativespecies abundances are suppressed by highproductivity conditions. Similarly, Thunell andSautter (1992) suggested an inverse relationshipbetween nannofossils and foraminiferal popula-tions with planktonic foraminiferal populationsincreasing and nannofossils declining duringpeaks of productivity. On the other hand, Young(1994) suggested that the number of nannofossilsincreased with productivity, but is suppressed inextreme eutrophic environments.
Further clues to the environmental signi¢canceof calcareous nannofossil abundance and speciesdiversity can be obtained from the Qreiya sec-tion of Egypt (Fig. 11), where planktonic forami-nifera have been analyzed quantitatively alongwith carbon isotope studies of the benthic spe-cies Cibicidoides pseudoacutus and the planktonicforaminifera Rugoglobigerina rugosa (Keller etal., in press). These studies show a positive cor-relation between calcareous nannofossil speciesrichness and planktonic foraminifera, and a pos-itive correlation of both with primary productiv-ity as indicated by carbon isotopes (Fig. 10). Inthe Upper Maastrichtian Micula murus Zone, thecalcareous nannofossil species richness increasedin the basal part of the section, correspondingto a signi¢cant increase in surface productivityas might be indicated by higher N
13C values(Fig. 10; Keller et al., in press). However, in the
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356348
P1a
Gebel Qreiya Wadi Hamama
A. cymbiformisbloom
Wa
rm-w
ate
r t
axa
in
flu
x
Maximum abundance
Max. species richness
Species richness &Abundance decrease
1st Thoracosphaera bloom
Braarudosphaera bigelowii bloom
C. pelagicus bloom
2nd Thoracosphaera bloom,re-entry of some Cretaceous taxa
Appearance of . C. primus
N. parvulum bloom
Increased abundance of Z. sigmiodes
Incre
ase
d a
bu
nd
an
ce
of
Mic
ula
de
cu
ssa
ta
Co
ol-
wa
ter
taxa
in
flu
x
Paleoenvironmental Interpretation
Nannofossil EventsSea Level
(Li et al., 2000: Keller &Stinnesbeck, 1996; this study)
1 2
Q - 4 0
Q - 3 5
Q 2 0
Q 3 0
Q 8 0
Q 9 0
Q 4 0
Q 5 0
Q 6 0
Q 7 0
Q -3 0
Q - 2 5
Q - 2 0
Q - 1 5
Q - 1 0
Q - 5
Q - 1
( m )
Ca
lca
reo
us
Na
nn
ofo
ssils
Pla
nktic
Fo
ram
inife
raC
F3
CF
1
P1a
P1b
P1
C(1
)P
1C
(2)
Da
nia
nM
aa
str
ich
tian
Sta
ge
M. p
rinsii
M. m
uru
s
Lith
olo
gy
Th
ickn
ess
Sa
mp
le N
o.
Dakhla
F
orm
atio
nF
orm
atio
n
1
0
2
3
4
5
6
7
8
9
10
11
ab
c
P1
C(1
)
M. in
vers
us (
NP
1)
Q-42
1
5
1 0
1 5
2 0
2 5
3 0
3 5
4 0
5 0
6 0
7 0
7 5
8 0
8 5
9 0
1
2
3
4
5
6
7
8
9
1 0
1 1
1 2
1 3
1 4
1 5
1 6
1 7
1 8
1 9
2 0
2 1
0
( m )
CF
3C
F1
P1
bP
1C
(1)
P1
C(2
)
M. m
uru
s
ab
ca
b
dis
so
l.P
1C
(1)
Pld
M. invers
us (N
P1
)M
. prinsii
Cool climate,? Low productivity,Highly stressed environment
Warm climate,? High productivity
Productivity minimum,High salinity, low sea level
Nearshore, shallow water,increased fresh water influx,lower salinity
Return of normal marine conditions.
Major sea-level lowstand.
Warm climate,High productivty.
Relatively cool climate.
High productivity,poor water mass stratification.
low high
C.
ten
uis
(N
P2
)Hiatus
Hiatus
Hiatus
Hiatus
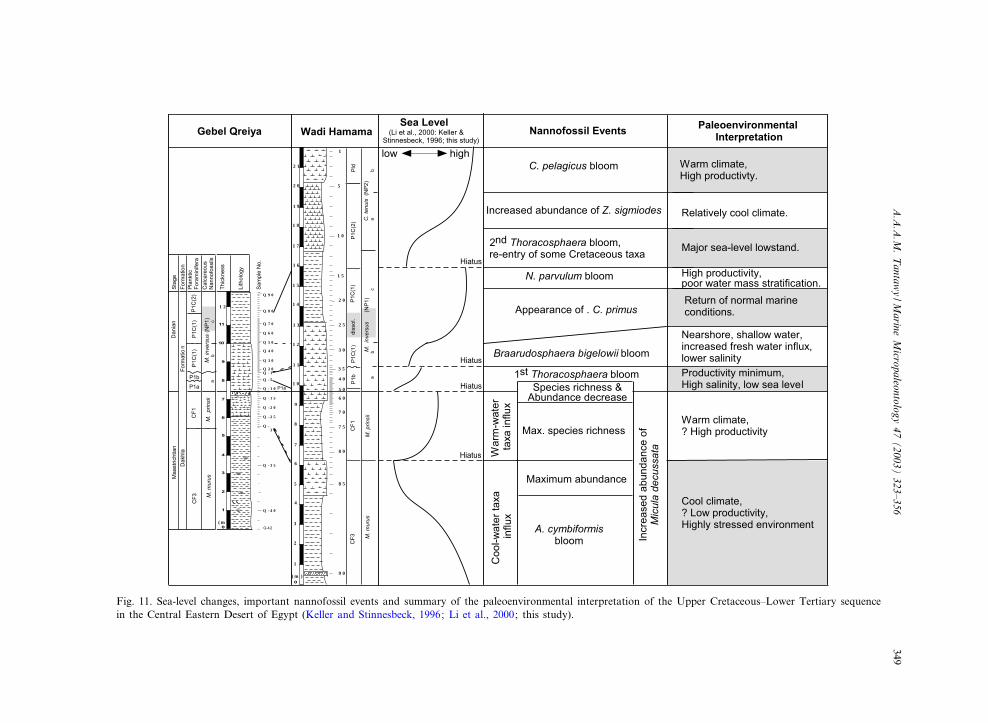
Fig. 11. Sea-level changes, important nannofossil events and summary of the paleoenvironmental interpretation of the Upper Cretaceous^Lower Tertiary sequencein the Central Eastern Desert of Egypt (Keller and Stinnesbeck, 1996; Li et al., 2000; this study).
MARMIC
90527-1-03
A.A.A.M.Tantaw
y/M
arineMicropaleontology
47(2003)
323^356349

middle part of the M. murus Zone, calcareousnannofossils exhibit lower species richness associ-ated with the breakdown in ocean primary pro-ductivity, as might be suggested by lower N
13Cvalues. In the Micula prinsii Zone, primary pro-ductivity increased signi¢cantly and calcareousnannofossils attain highest species richness values(Fig. 10; Keller et al., in press). A decrease insurface productivity near the top of the Maas-trichtian is associated with the apparent decreasein nannofossil species richness and abundance(Figs. 6 and 10). The decrease in surface pro-ductivity at the top of the Maastrichtian is con-sistent with this pattern elsewhere in the TethysOcean (e.g. Keller and Lindinger, 1989; Zachoset al., 1989; Barrera, 1994; Barrera and Keller,1994; Li and Keller, 1998a; Keller et al., inpress).
At Wadi Hamama, the Cretaceous calcareousnannofossil species richness shows trends similarto the nearby Qreiya section, except for the lowerpart of the section in the lower Micula murusZone, where species richness decreased (Fig. 7).This decrease may be due to the poor preserva-tion at Wadi Hamama and the shallower deposi-tional environment.
At the K/T boundary, species richness and rel-ative abundances of Cretaceous species decrease,whereas Cretaceous survivor species and Tertiaryspecies increase. In the Lower Danian (SubzonesNP1a and NP1b), both calcareous nannofossilgroups show low abundances and marked £uctu-ations in species richness. Carbon isotope dataindicate low, but £uctuating productivity duringthis interval. In Subzone NP1c and Zone NP2, thespecies richness and relative abundances increasealong with surface productivity, as indicated byincreased N
13C values (Fig. 10) and the returnto normal marine condition in the upper partof the Danian. These observations parallel thepost-K/T recovery of planktonic foraminifera,the return to normal marine productivity, andre-establishment of the normal surface-to-deepgradient in low to middle latitudes in the upperpart of Subzone P1c(1), nearly 2 Myr after theK/T boundary event (Keller, 1988; 1996; Zachoset al., 1989; Barrera and Keller, 1994; Keller etal., in press).
8.2. Climate
The Upper Maastrichtian Micula murus Zone ischaracterized by global cooling, culminating in asea-level lowstand at 65.5 Ma that is generallyassociated with a hiatus at the top of planktonicforaminiferal Zone CF3 (Barrera et al., 1997; Liand Keller, 1998c). In the present study, the M.murus Zone at Gebel Qreiya exhibits a similarcooling trend, as might be indicated by more pos-itive N
18O values (Fig. 10; Keller et al., in press).This cooling event is marked by a decrease in thewarm-water species Watznaueria barnesae and theincreased abundance of the cool-water species(e.g. Arkhangelskiella cymbiformis, Gartnerago ob-liquum and Nephrolithus frequens Figs. 8 and 9) atboth Qreiya and Wadi Hamama. In the Miculaprinsii Zone, a warming trend is indicated by theincrease in the warm-water species W. barnesaeand by the lower benthic and planktonic N
18Ovalues (Fig. 10). This warming is consistent withthe middle to high-latitude climate trends, wherecooling in Zone CF3 is followed by 3^4‡C warm-ing in Zones CF1^2 interval (Li and Keller,1998c).
A warm climate prevailed during the depositionof the Lower Danian Subzones NP1a^b, as indi-cated by benthic N
18O values, whereas cooler cli-matic conditions were re-established during thesedimentation of the upper part of SubzoneP1c(1) and Subzone NP1c, as indicated by bothbenthic and planktonic N
18O values (Fig. 10; Kel-ler et al., in press). At Wadi Hamama, coolerclimatic conditions prevailed into planktonic fora-miniferal Subzone P1c(2) and into the lower partof calcareous nannofossil Zone NP2, as indicatedby a marked increase in the abundance of Zygo-discus sigmoides. This cooling trend is accompa-nied by the recovery of surface productivity andre-establishment of normal marine conditions.The top of the Wadi Hamama section is markedby increased abundance of Coccolithus pelagicus,which may indicate a warmer climate during theupper part of Zone NP2.
8.3. Paleoenvironment of Central Egypt
The paleoenvironmental signi¢cance of various
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356350

calcareous nannoplankton species in the Late Cre-taceous and Early Tertiary is not yet well under-stood. The present study of the Central EasternDesert of Egypt yields further clues to environ-mental a⁄nities, as summarized in Fig. 11.
During the Late Maastrichtian, the Micula mu-rus Zone is characterized by a cooler and rela-tively low-productivity, high-stress environment,as indicated by an in£ux of cool-water taxa, abloom of Arkhangelskiella cymbiformis, abun-dance of Micula decussata and low carbon isotopevalue. The subsequent Micula prinsii Zone is char-acterized by a warmer climate and increased pro-ductivity, as indicated by an in£ux of warm-watertaxa, higher nannofossil species richness and in-creased carbon isotope values. At the K/T bound-ary and in the Early Danian Subzone NP1a, theenvironment was characterized by very low pro-ductivity, high salinity and low sea level, as indi-cated by decreased nannofossil species richnessand low carbon isotope values, and high-stressconditions, as indicated by a bloom of the oppor-tunistic Thoracosphaera spp.
The Early Danian Subzone NP1b is character-ized by increased fresh-water in£ux and lower sa-linity, as indicated by the bloom of the disasterspecies Braarudosphaera bigelowii. The appear-ance of Cruciplacolithus primus in Subzone NP1cmarks the gradual return to more stable environ-mental conditions, characterized by increased pro-ductivity and poor water mass strati¢cation, asindicated by the dwarfed Neobiscutum parvulumbloom. A major sea-level regression occurred atthe top of Subzone NP1c and the base of SubzoneNP2a, as inferred from the second bloom of Thor-acosphaera spp. and the re-entry of some Creta-ceous taxa. A relatively cool climate prevailedduring the upper part of Subzone NP2a, as sug-gested by the increased abundance of Zygodiscussigmoides, followed by a warmer climate, highproductivity, and high sea level, as indicated bya Coccolithus pelagicus bloom and increased N
13Cvalues within Subzone NP2b (Fig. 11).
9. Conclusions
1. The Gebel Qreiya and nearby Wadi Hama-
ma sections of the Eastern Desert of Egypt areamong the very few localities where the plankton-ic foraminifera Plummerita hantkeninoides (ZoneCF1) and Parvularugogloberina eugubina (P1a)Subzone and calcareous nannofossilMicula prinsiiand Markalius inversus (NP1) Zones are present,and where the K/T boundary is marked by a redlayer and Ir anomaly.
2. This is the ¢rst study that quantitatively an-alyzes calcareous nannofossil assemblages acrossthe K/T boundary in Egypt, based on closelyspaced samples and high-resolution age control,that provides a more accurate assessment of the£oral assemblage turnovers than is possible withstandard species ¢rst and last appearance studies.
3. The Late Maastrichtian assemblages in cen-tral Egypt are typical of low to mid-latitudes withthe vanishing Cretaceous nannofossil assemblageabundant and diverse (99.5% to 93.3%) in theMicula murus Zone and gradually decreasing inthe Micula prinsii Zone correlative with an in-crease in the survivor assemblage (mainly Thora-cosphaera and Braarudosphaera bigelowii). TheTertiary assemblage is a minor component(V2.5%) of the Early Danian nannofossil assem-blages, but increases rapidly to 56% at the base ofNP2.
4. Calcareous nannofossil abundance and spe-cies richness indicate that the Late MaastrichtianMicula murus Zone is characterized by a coolerclimate, a high-stress environment and low pro-ductivity, whereas a warmer climate and higherproductivity characterize the Micula prinsii Zone.
5. Minimum productivity, high salinity and alow sea-level mark the K/T boundary and Sub-zone NP1a. Subzone NP1b is characterized byincreased fresh-water in£ux and lower salinity.
6. Normal marine environmental conditions re-turned in Subzone NP1c, marked by increasedproductivity and possibly poor water mass strat-i¢cation. A major sea-level regression occurred atthe NP1c/NP2a transition.
Acknowledgements
I am grateful to Prof. Gerta Keller, Departmentof Geosciences, Princeton Uinversity, USA for
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356 351

discussions, comments, critical review of thismanuscript, and for making available her lab fa-cilities. I also thank the reviewers, Prof. Kathari-na von Salis Perch-Nielsen and Prof. T. Bralower,for critical comments and helpful suggestions forimprovement of this manuscript, Prof. ThierryAdatte for ¢eldwork collaboration, and Prof. W.Berggren and Prof. M.-P. Aubry for their helpfulcomments and suggestions. This publication wassponsored by the US^Egypt Science and Technol-ogy Joint Fund in cooperation with NSF andNRC under Project OTH2-008-001-98.
References
Abbas, H.L., Habib, M.M., 1969. Stratigraphy of west Ma-whoob area, Southwestern Desert. Egypt. Bull. Desert In-stitute, Egypt 19, 48^108.
Abdelmalik, W.M., Bassiouni, M.A., Kerdany, M.T., Obeid,F.L., 1978. Biostratigraphy of Upper Cretaceous^LowerTertiary rocks from West central Sinai. 2. Calcareous nan-noplankton. Ann. Mines Geol. 28, 217^241.
Abdel Razik, T.M., 1969. Stratigraphical studies on the phos-phate deposits between River Nile and Red Sea (south oflatitude 27‡N). Fac. Sci. Bull. Cairo Univ. 42, 299^324.
Abdel Razik, T.M., 1972. Comparative studies on the UpperCretaceous^Early Paleogene sediments on the Red Seacoast, Nile Valley and Western Desert, Egypt. 6th Arab Pe-troleum Congr., Algeria, 71: 1^23.
Alcala¤-Herrera, J.A., Grossman, E.L., Gartner, S., 1992. Nan-nofossil diversity and equitability and ¢ne-fraction D
13Cacross the Cretaceous/Tertiary at Walvis Ridge Leg 74,South Atlantic. Mar. Micropaleontol. 20, 77^88.
Almogi Labin, A., Bein, A., Sass, E., 1993. Late Cretaceousupwelling system along the southern Tethys margins (Isra-el): interrelationship between productivity, bottom-water en-vironments and organic matter preservation. Paleoceanogra-phy 8, 671^690.
Askin, R.A., 1992. Preliminary palynology and stratigraphicinterpretation from a new Cretaceous^Tertiary boundarysection from Seymour Island. Antarct. J. US 25, 42^44.
Barrera, E., 1994. Global environmental changes preceding theCretaceous^Tertiary boundary: Early^Late Maastrichtiantransition. Geology 22, 877^880.
Barrera, E., Keller, G., 1994. Productivity across the K/Tboundary in high latitudes. Geol. Soc. Am. Bull. 106,1254^1266.
Barrera, E., Savin, S.M., Thomas, E., Jones, C.E., 1997. Evi-dence for thermohaline circulation reversals controlled bysea-level change in the latest Cretaceous. Geology 25, 715^718.
Barron, T., Hume, W.F., 1902. Topography and geology ofthe Eastern Desert of Egypt (Central Portion), Survey Dept.Cairo.
Barthel, W.K., Herrmann-Degen, W., 1981. Late Cretaceousand Early Tertiary stratigraphy in the Great Sand Sea andits SE Margins (Farafra and Dakhla Oases, SW DesertEgypt. Mitt. Bayer. Staatssamml. Palaeontol. Hist. Geol.21, 141^182.
Bassiouni, M.A., Faris, M., Sharaby, S., 1991. Late Maas-trichtian and Paleocene calcareous nannofossils from AinDabadib section: NW Kharga Oasis. Qatar Univ. Sci.Bull. J. 11, 357^375.
Berggren, W.A., Kent, D.V., Swisher, C.C. III, Aubry, M.-P.,1995. A revised Cenozoic geochronology and chronostratig-raphy. In: Berggren, W., Kent, D.V., Aubry, M.-P., Hard-enbol, J. (Eds.), Geochronology, Time Scales and GlobalStratigraphic Correlation. Soc. Sed. Geol. Spec. Publ. 54:129^212.
Bukry, D., 1974. Coccoliths as paleosalinity indicators: evi-dence from Black Sea. AAPG Mem. 20, 353^363.
Bukry, D., Bramlette, M.N., 1970. Coccolith age determina-tions, Leg 3, Deep Sea Drilling Project. In: Maxwell, A.E.,Von Herzen, R.P., et al. (Eds.), Init. Rep. DSDP 3, 589^611.
Cepek, P., Hay, W.W., 1969. Calcareous nannoplankton andstratigraphic subdivision of the Upper Cretaceous. Trans.Gulf Coast Assoc. Geol. Soc. 19, 323^336.
Cunha, A.S., Shimabukuro, S., 1996. Evidence of Braarudo-sphaera blooms at the Cenomanian-Turonian boundary ofthe Santos basin, Brazil. IGCP 381 News, South AtlanticMesozoic.
Ehrendorfer, T., Aubry, M-P., 1992. Calcareous nannoplank-ton changes across the Cretaceous/Paleocene boundary inthe southern Indian Ocean (Site 750). Proc. ODP Sci. Re-sults 120, 451^459.
El-Dawoody, A.S., 1973. Microbiostratigraphy of some UpperCretaceous and Lower Tertiary sediments in Egypt. Osleny-tani vitak (Discussions Paleontologicae), Budapest 21, 117^118.
El-Dawoody, A.S., 1983. Nannobiostratigraphy of the LateCretaceous^Early Eocene sediments in the Dakhla/Khargaregion, Western Desert, Egypt. 12th Colloq. Afr. Geol., Ter-vuren, Brussels, 27 pp.
El-Dawoody, A.S., 1990a. Nannobiostratigraphy of the LateCretaceous^Paleocene succession in Esh El-Mellaha Range,Eastern Desert, Egypt. Qatar Univ. Sci. Bull. J. 10, 315^557.
El-Dawoody, A.S., 1990b. Nannobiostratigraphy of the LateCretaceous^Early Tertiary succession in the Nile Valley andEastern Desert, Egypt. Proc. Zool. Soc. Egypt, Cairo 18,465^486.
El-Dawoody, A.S., Barakat, M.G., 1973. Nannobiostratigra-phy of the Upper Cretaceous^Paleocene contact in DuwiRange, Quseir District, Egypt. Riv. Ital. Paleont. 79, 103^124.
El-Dawoody, A.S., Zidan, M.A., 1976. Micro and nannopa-leontology of the Upper Cretaceous^Paleocene succession inWest Mawhoob area, Dakhla Oasis, Egypt. Rev. Espan.Micropaleont. 8, 401^428.
El-Naggar, Z.R., 1966. Stratigraphy and planktonic foraminif-
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356352

era of the Upper Cretaceous^Lower Tertiary succession inthe Esna^Idfu Region, Nile Valley, Egypt. Br. Mus. Nat.Hist. Bull. Suppl. 2, London, 279 pp..
Eshet, Y., Almogi Labin, A., 1996. Calcareous nannofossils aspaleoproductivity indicators in Upper Cretaceous organic-rich sequences in Israel. Mar. Micropaleontol. 29, 37^61.
Eshet, Y., Almogi Labin, A., Bein, A., 1994. Dino£agellatecysts, paleoproductivity and upwelling systems: a Late Cre-taceous example from Israel. Mar. Micropaleontol. 18, 199^228.
Eshet, Y., Moshkovitz, S., Habib, D., Benjamini, C., Magar-itz, M., 1992. Calcareous nannofossil and dino£agellate stra-tigraphy across the Cretaceous/Tertiary boundary at HorHahar, Israel. Mar. Micropaleontol. 18, 199^228.
Faris, M., 1974. Geological and Paleontological Studies on theLate Cretaceous^Early Tertiary Succession in Qena Regionand Kharga Oasis. M.Sc. Thesis, Assiut Univ., Egypt, 189pp.
Faris, M., 1982. Micropale¤ontologie et biostratigraphie duCre¤tace¤ Supe¤rieur a' l’Eocene infe¤rieur de l’Egypte centrale(Region de Duwi Valle¤e du Nil, Oasis de Kharga et deDakhla). Doctorat d’Etat, Univ. de Paris Vl, France, 438pp., 50 pls.
Faris, M., 1984. The Cretaceous^Tertiary boundary in CentralEgypt (Duwi region, Nile Valley, Kharga and Dakhlaoases). Neues Jahrb. Geol. Palaeontol. Abh. 7, 385^392.
Faris, M., 1985. Stratigraphy of the Late Cretaceous/EarlyTertiary sediments in Ghanima and Ain Amur sections,Kharga area, Egypt. Newsl. Stratigr. 14, 36^47.
Faris, M., 1988. Late Cretaceous/Early Tertiary calcareousnannofossils from El Qusaima area, NE Sinai, Egypt. Bull.Fac. Sci. Qena (Egypt) 2, 263^275.
Faris, M., 1992. Calcareous nannoplankton from the Turo-nian^Maastrichtian sequence, East of El Qusaima, NE Si-nai, Egypt. Qatar Univ. Sci. 12, 166^175.
Faris, M., 1995a. Late Cretaceous and Paleocene calcareousnannofossil biostratigraphy at St. Paul section, SouthernGalala Plateau, Egypt. Ann. Geol. Surv. Egypt 20, 527^538.
Faris, M., 1995b. Morphometric analysis of Arkhangelskiellacymbiformis Vekshina, 1959, in the Upper Cretaceous Rocksof Egypt and its stratigraphic importance. Ann. Geol. Surv.Egypt 20, 585^601.
Faris, M., 1997. Biostratigraphy of calcareous nannofossilsacross the K/T boundary in Egypt. Neues Jahrb. Geol. Pa-laeont. Monatsh. 8, 447^464.
Faris, M., Abd El-Hamid, A.T., 1986a. Contribution to theLate Maastrichtian^Paleocene Chalk at Jiran El Ful area,Abu Roash, West of Cairo, Egypt. Newsl. Stratigr. 15, 139^149.
Faris, M., Abd El-Hamid, A.T., 1986b. The Cretaceous/Ter-tiary boundary in Tramsa section, West of Qena, Nile Val-ley, Egypt. Newsl. Stratigr. 16, 85^97.
Faris, M., Allam, A., Marzouk, A.M., 1985. Biostratigraphyof the Late Cretaceous/Early Tertiary rocks in the Nile Val-ley (Qena Region), Egypt. Ann. Geol. Surv. Egypt 15, 287^300.
Gaarder, K.R., Hasle, G.R., 1971. Coccolithophorids of theGulf of Mexico. Bull. Mar. Sci. Gulf Carrib. 21, 519^544.
Gardin, S., 2002. Late Maastrichtian to Early Danian calcar-eous nannofossils at Elles (northwest Tunisia) : a tale of onemillion years across the K/T boundary. Paleogeogr. Paleo-climatol. Paleoecol. 2756, 1^21.
Gardin, S., Monechi, S., 1998. Paleoecological change in mid-dle to low-latitude calcareous nannoplankton at the Creta-ceous/Tertiary boundary. Bull. Soc. Geol. Fr. 169, 709^723.
Gartner, S., 1996. Calcareous nannofossil at the Cretaceous/Tertiary boundary. In: Keller, G., MacLeod, N. (Eds.), Cre-taceous Mass Extinction. Norton and Company, New York,pp. 27^47.
Girgis, M.H., 1989. A morphometric analysis of the Arkhan-gelskiella group and its stratigraphical and paleoenviron-mental imortance. In: Crux, J., van Heck, S. (Eds.), Nanno-fossils and their Applications. Ellis Horwood, Chichester,pp. 327^339.
Gorostidi, A., Lamolda, M.A., 1991. Variaciones de la nano-£ora calcarea en el paso Cretacico^Terciario de Monte Urko(N de Espan‹a). Rev. Esp. Paleontol. 6, 89^97.
Haq, B.U., Lohmann, G.P., 1976. Early Cenozoic calcareousnannoplankton biogeography of the Atlantic Ocean. Mar.Micropaleontol. 1, 119^194.
Hay, W.W., Mohler, H.P., 1967. Calcareous nannoplanktonfrom Early Tertiary rocks at Pont Labau, France, and Pa-leocene^Eocene correlation. J. Paleontol. 41, 1505^1541.
Hendriks, F., Luger, P., Kallenbach, H., Schroeder, J.H., 1984.Stratigraphical and sedimentological framework of theKharga-Sin El Kaddab stretch (western and southern partof the upper Nile basin), Western Desert, Egypt. Berl. Geo-wiss. Abh. 50(A), 117^151.
Hendriks, F., Luger, P., 1987. The Rakhiyat Formation of theGebel Qreiya area: evidence of middle Campanian to EarlyMaastrichtian synsedimentary tectonism. Berl. Geowiss.Abh. 75(A), 83^96.
Hendriks, F., Luger, P., Biwitz, J., Kallenbach, H., 1987. Evo-lution of the depositional environments of SE Egypt duringthe Cretaceous and Lower Tertiary. Berl. Geowiss. Abh.75(A), 49^82.
Henriksson, A.S., 1993. Biochronology of the terminal Creta-ceous calcareous nannofossil Zone of Micula prinsii. Creta-ceous Res. 14, 59^68.
Henriksson, A.S., 1996. Calcareous nannoplankton productiv-ity and succession across the Cretaceous^Tertiary boundaryin the Paci¢c (DSDP Site 465) and Atlantic (DSDP Site 527)Oceans. Cretaceous Res. 17, 451^477.
Hermina, M., 1990. The surroundings of Kharga, Dakhla andFarafra Oases. In: Said, R. (Ed.), The Geology of Egypt.Balkema, Rotterdam, pp. 259^292.
Issawi, B., 1972. Review of Upper Cretaceous^Lower Tertiarystratigraphy in central and southern Egypt. Bull. Am. Assoc.Petrol. Geol. 56, 1448^1463.
Jenkens, D., 1990. North and Central Sinai. In: Said, R. (Ed.),The Geology of Egypt. Balkema, Rotterdam, pp. 361^380.
Jiang, M.J., Gartner, S., 1986. Calcareous nannofossil succes-
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356 353

sion across the Cretaceous^Tertiary boundary in east-centralTexas. Micropaleontology 32, 232^255.
Kassab, A., Keheila, E.A., 1994. Paleocene biostratigraphyand sea level changes of the northern Eastern Desert ofEgypt. Newsl. Stratigr. 31, 85^100.
Keller, G., 1988. Extinction, survivorship and evolution ofplanktic foraminifera across the Cretaceous/Tertiary bound-ary at El Kef, Tunisia. Mar. Micropaleontol. 13, 239^263.
Keller, G., 1996. Cretaceous/Tertiary mass extinction in plank-tonic foraminifera: biotic constraints for catastrophe theo-ries. In: MacLeod, N., Keller, G. (Eds.), The Cretaceous/Tertiary boundary mass extinction: biotic and environmen-tal changes. W.W. Norton p Co., New York, pp. 49^84.
Keller, G., Lindinger, M., 1989. Stable isotope, TOC andCaCO3 record across the Cretaceous^Tertiary boundary atEl Kef, Tunisia. Palaeogeogr. Palaeoclimatol. Palaeoecol.73, 243^265.
Keller, G., von Salis-Perch-Nielsen, K., 1995. Cretaceous^Ter-tiary (K/T) mass extinction: e¡ect of global change on cal-careous microplankton. In: Stanley, S. (Ed.), The E¡ects ofPost-Global Change on Life. Nat. Acad. Sci./Nat. Res.Council, Washington DC, pp. 72^93.
Keller, G., Stinnesbeck, W., 1996. Sea level changes, clasticdeposits, and megatsunamis across the Cretaceous^Tertiaryboundary. In: MacLeod, N., Keller, G. (Eds.), The Creta-ceous/Tertiary Boundary Mass Extinction: Biotic and Envi-ronmental Changes. Norton and Company, New York,pp. 415^450.
Keller, G., Adatte, T., Burns, S., Tantawy, A., in press. High-stress paleoenvironment during the Late Maastrichtian toEarly Paleocene in Central Egypt. Paleogeogr. Paleoclima-tol. Paleoecol.
Keller, G., Li, L., MacLeod, N., 1995. The Cretaceous/Terti-ary boundary stratotype section at El Kef, Tunisia: howcatastrophic was the mass extinction? Paleogeogr. Paleocli-matol. Paleoecol. 119, 221^254.
Klitzsch, E., Groeschke, M., Herrmann-Degen, W., 1990.Wadi Qena: Paleozoic and pre-Campanian Cretaceous stra-ta. In: Said, R. (Ed.), The Geology of Egypt. Balkema,Rotterdam, pp. 321^328.
Lamolda, M.A., Gorostidi, A., 1992. Nannofossil stratigraphicrecord in Upper Maastrichtian^lowermost Danian at Zu-maya. Mem. Sci. Geol. 43, 149^161.
Li, L., Keller, G., 1998a. Maastrichtian climate, productivityand faunal turnovers in planktic foraminifera in South At-lantic DSDP Sites 525 and 21. Mar. Micropaleontol. 33, 55^86.
Li, L., Keller, G., 1998b. Maastrichtian diversi¢cation ofplanktic foraminifera at El Kef and Elles, Tunisia. EclogaeGeol. Helv. 91, 75^102.
Li, L., Keller, G., 1998c. Abrupt deep-sea warming at the endof the Cretaceous. Geology 26, 995^999.
Li, L., Keller, G., Adatte, T., Stinnesbeck, W., 2000. LateCretaceous sea level changes in Tunisia: a multi-disciplinaryapproach. J. Geol. Soc. London 157, 447^458.
Luger, P., 1988. Maastrichtian to Paleocene facies evolutionand Cretaceous/Tertiary boundary in middle and southern
Egypt. Rev. Espan. Micropaleont. Numero Extraordinario,pp. 83^90.
Luger, P., Gro«schke, M., 1989. Late Cretaceous ammonitesfrom the Wadi Qena area in the Egyptian Eastern Desert.Palaeontology 32, 355^407.
Luger, P., El-Beshtawy, M., Mai, H., 1998. Maaastrichtianplanktonic foraminiferal biozonation and K/T boundary incentral Egypt. Rev. Espan. Micropaleont. 30, 37^49.
Magaritz, M., Moshkovitz, S., Benjamini, Hansen H.J., Ha-kansson, E., Rasmussen, K.L., 1985. Carbon isotope-, bio-and magnetostratigraphy across the Cretaceous^Tertiaryboundary in the Zin Valley Negev, Israel. Newsl. Stratigr.15, 100^113.
Mai, H., v. Salis Perch-Nielsen, K., Willems, H., Romein, T.,1997. Fossil coccospheres from the K/T boundary sectionfrom Geulhemmerberg, the Netherlands. Micropaleontology43, 281^302.
Martini, E., 1970. Standard Paleogene calcareous nannoplank-ton zonation. Nature 226, 560^561.
Martini, E., 1971. Standard Tertiary and Quaternary calcare-ous nannoplankton zonation. In: Farinacci, A. (Ed.), Proc.2nd Planktonic Conf. Rome 1970, Ed. Technoscienza 2:739^785.
McIntyre, A., Be¤, A., 1967. Modern Coccolithophoridae of theAtlantic Ocean. I. Pacoliths and cyrtoliths. Deep-Sea Res.Oceanogr. 14, 561^597.
Monechi, S., 1977. Upper Cretaceous and Early Tertiary nan-noplankton from the Scaglia Umbra Formation (Gubbio,Italy). Rev. Ital. Paleont. 83, 759^802.
Monechi, S., 1979. Variations in nannofossils assemblage atthe Cretaceous^Tertiary boundary in the Bottaccione section(Gubbio, Italy). Cretaceous^Tertiary Events Symp., Copen-hagen, 2: 164^167, Copenhagen, Denmark.
Moshkovitz, S., Eshet, Y., 1989. Inter-relative aspects of nan-nofossil and palynomorph biostratigraphy and paleoecologyat the K/T boundary, Hor Hahar section, southern Israel.Abstract. INA Newsl. 11, 80^81.
Mu«ller, C., 1985. Biostratigraphic and paleoenvironmental in-terpretation of the Goban Spur region based on a study ofcalcareous nannoplankton. In: de Gracinsky, P.C., Poag,C.W., et al. (Eds.), Init. Rept. DSDP 80, 573^599.
Okada, H., Bukry, D., 1980. Supplementary modi¢cation andintroduction of code numbers to the low-latitude coccolithbiostratigraphic zonation (Bukry, 1973, 1975). Mar. Micro-paleontol. 5, 321^325.
Pardo, A., Ortiz, N., Keller, G., 1996, Latest Maastrichtianand K/T boundary foraminiferal turnover and environmen-tal changes at Agost, Spain. In: MacLeod, N., Keller, G.(Eds.), The Cretaceous^Tertiary Boundary Mass Extinction:Biotic and Environmental Events Across the Cretaceous/Tertiary Boundary. Norton and Company, New York, pp.139^172.
Perch-Nielsen, K., 1971. Neue Coccolithen aus dem Pala«oza«nvon Da«nemark, der Bucht von Biskaya und dem Eoza«n derLabrador See. Bull. Geol. Soc. Denmark 21, 51^66.
Perch-Nielsen, K., 1979a. Calcareous nannofossil zonation atthe Cretaceous^Tertiary boundary in Denmark. Proc. Creta-
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356354

ceous^Tertiary Boundary Events Symp., Copenhagen, 1,115^135.
Perch-Nielsen, K., 1979b. Calcareous nannofossils in the Cre-taceous^Tertiary boundary sections in Denmark. Proc. Cre-taceous^Tertiary Boundary Events Symp., Copenhagen, 2,120^126.
Perch-Nielsen, K., 1979c. Calcareous nannofossils at the Cre-taceous^Tertiary boundary near Biarritz, France. Proc. Cre-taceous^Tertiary Boundary Events Symp., Copenhagen, 2,151^155.
Perch-Nielsen, K., 1981a. New Maastrichtian and Paleocenecalcareous nannofossils from Africa, Demnark, the USAand the Atlantic, and some Paleocene lineages. EclogaeGeol. Helv. 73, 831^863.
Perch-Nielsen, K., 1981b. Les coccolithes du Pale¤oce'ne pre's deEl kef, Tunisie et leurs ance“tres. Cah. Micropaleontol. 3, 7^25.
Perch-Nielsen, K., 1981c. Les nannofossiles calcaires a' la limiteCre¤tace¤^Tertiaire pre's de El Kef, Tunisie. Cah. Micropa-leontol. 3, 25^37.
Perch-Nielsen, K., 1985. Cennozoic calcareous nannofossils.In: Bolli, H.M., Saunders, J.B., Perch-Nielsen, K. (Eds.),Plankton Stratigraphy. Cambridge University Press, Cam-bridge, pp. 422^454.
Perch-Nielsen, K., McKenzie, J., He¤, Q., 1982. Biostratigraphyand isotope stratigraphy and the ‘catastrophic’ extinction ofcalcareous nannoplankton at the Cretaceous/Tertiaryboundary. Geol. Soc. Am. Spec. Paper 190, 353^371.
Perch-Nielsen, K., Sadek, A., Barakat, M.G., Teleb, F., 1978.Late Cretaceous and Early Tertiary calcareous nannofossiland planktonic foraminifera zones from Egypt. Sixth Afri-can Micropaleont., Coll. Tunis (1974) 28 (II), 337^403.
Percival, S.F., Fischer, A.G., 1977. Changes in calcareous nan-noplankton in the Cretaceous^Tertiary biotic crisis at Zu-maya, Spain. Evolutionary Theory 2, 1^35.
Pospichal, J.J., 1991. Calcareous nannofossils across the Creta-ceous/Tertiary boundary at Site 752, eastern Indian Ocean,In: Weissel, J., Peirce, J., et al. (Eds.), Proc. ODP Sci. Re-sults 121, 395^414.
Pospichal, J.J., 1994. Calcareous nannofossils at the K/Tboundary, El Kef: no evidence for stepwise, gradual or se-quential extinctions. Geology 22, 99^102.
Pospichal, J.J., 1995. Cretaceous Tertiary boundary calcareousnannofossils from Agost, Spain. In: Flores, J.A., Sierro,F.J. (Eds.), 5th INA Conf. in Salamanca, Proc., pp. 185^217.
Pospichal, J.J., Bralower, T.J., 1992. Calcareous nannofossilsacross the Cretaceous^Tertiary boundary, Site 761, North-west Australian margin. Proc. ODP Sci. Results 122, 515^532.
Pospichal, J.J., Wise, S.W., Jr., 1990. Calcareous nannofossilsacross the K/T boundary, ODP Hole 690C, Maud Rise,Weddell Sea. Proc. ODP Sci. Results 113, 515^532.
Romein, A.J.T., 1977. Calcareous nannofossils from the Creta-ceous^Tertiary boundary interval in the Barranco del Gre-dero (Caravaca Province, Murcia, S.E. Spain), Parts I andII. Proc. K. Ned. Akad. Wet. Ser. B, 256^279.
Romein, A.J.T., 1979. Lineages in Early Paleocene nanno-plankton. Utrecht Micropaleontol. Bull. 22, 18^22.
Romein, A.J.T., Smit, J., 1981. The Cretaceous/Tertiaryboundary: calcareous nannofossils and stable isotopes. Pa-leontol. Proc. B 84, 295^314.
Roth, P.H., 1978. Calcareous nannoplankton biostratigraphyand oceanography of the Northwestern Atlantic Ocean. Init.Rep. DSDP 44, 731^759.
Roth, P.H., 1983. Calcareous nannofossils in mid-Cretaceousblack shale cycles from the Atlantic and Paci¢c: e¡ects ofdiagenesis. EOS 64, 733^734.
Roth, P.H., Bowdler, J.L., 1979. Evolution of the calcareousnannofossil genus Micula in the Late Cretaceous. Micropa-leontology 25, 272^280.
Roth, P.H., Thierstein, H.R., 1972. Calcareous nannoplank-ton: Leg 14 of the Deep Sea Drilling Project. In: Hayes,D.E., Pimm, A.C., et al. (Eds.), Init. Rep. DSDP 14, 421^485.
Said, R., 1962. The Geology of Egypt. Elsevier, Amsterdam,377 pp.
Schrank, E., Perch-Nielsen, K., 1985. Late Cretaceous paly-nostratigraphy in Egypt with comments on Maastrichtianand Early Tertiary calcareous nannofossils. Newsl. Stratigr.15, 81^99.
Sestini, G., 1984. Tectonic and sedimentary history of NEAfrican margin (Egypt/Lybia). In: Dikson, J.E., Robertson,A.H.F. (Eds.), The geological evolution of the eastern Med-iterranean, Blackwell Scienti¢c Publisher, Oxford, pp. 161^175.
Seyve, C., 1990. Nannofossil biostratigraphy of the Creta-ceous^Tertiary boundary in the French Basque Country.Bull. Centr. Rech. Explor. Prod. Elf Aquitaine 14, 553^572.
Sha¢k, S., Stradner, H., 1971. Nannofossils from the EasternDesert, Egypt, with reference to Maastrichtiaan nannofossilsfrom the USSR, Wien. Jahrb. Geol. Bundesanst. Sonderb.17, 69^104.
Shahin, A., 1992. Contribution to the foraminiferal biostratig-raphy and paleobathymetry of the Late Cretaceous andEarly Tertiary in the western central Sinai, Egypt. Rev. Mi-cropaleontol. 35, 157^175.
Sissingh, W., 1977. Biostratigraphy of Cretaceous calcareousnannoplankton. Geol. Mijnbouw 56, 37^65.
Soliman, M., Habib, M., Ahmed, E., 1986. Sedimentologicand tectonic evolution of the Upper Cretaceous^Lower Ter-tiary succession at Wadi Qena, Egypt. Sediment. Geol. 46,11^133.
Strougo, A,Haggag,M.A., Luterbacher, H.P., 1992. The basalPaleocene ‘GIobigerina’ eugubina Zone in the Eastern Des-ert (St. Paul’s Monastery, South Galala), Egypt. NeuesJahrb. Geol. Palaeontol. Monatsh. 2, 97^102.
Tantawy, A.A., 1998. Stratigraphical and paleontological stud-ies on some Paleocene-Eocene successions in Egypt. Ph.D.Thesis, Aswan Fac. Sci., South Valley Univ., Egypt, 273 pp.
Tantawy, A.A., Keller, G., Adatte, T., Burns, S., 2000. TheCretaceous^Tertiary transition in central Egypt: biostratig-raphy, mineralogy and stable isotopes. 6th InternationalCretaceous Symposium (Abstract), Vienna, Austria, p. 170.
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356 355

Tantawy, A.A., Keller, G., Adatte, T., Stinnesbeck, W., Kas-sab, A., Schulte, P., 2001. Maastrichtian to Paleocene(Dakhla Formation) depositional environment of the West-ern Desert in Egypt: sedimentology, mineralogy and inte-grated micro- and macrofossil biostratigraphies. CretaceousRes. 22, 795^827.
Thierstein, H.R., 1980. Selective dissolution of Late Creta-ceous and Earliest Tertiary calcareous nannofossils: exper-imental evidence. Cretaceous Res. 2, 165^176.
Thierstein, H.R., 1981. Late Cretaceous nannoplankton andthe change at the Cretaceous^Tertiary boundary. SEPMSpec. Publ. 32, 355^394.
Thierstein, H.R., Berger, W., 1979. On phanerozoic mass ex-tinctions. Naturwissenschaften 66, S46.
Thierstein, H.R., Haq, B.U., 1977. Maestrichtian/Danian bio-geographic variations in calcareous nannoplankton (ab-stract). J. Paleontol. 51, 28.
Thierstein, H.R., Okada, H., 1979. The Cretaceous/Tertiaryboundary event in the North Atlantic. In: Tucholke, B.E.,Vogt, P.R., et al. (Eds.), Init. Rep. DSDP 43, 601^616.
Thunell, R.C., Sautter, L.R., 1992. Planktonic foraminiferal,fauna and stable isotope indices of upwelling: a sediment-trap study in the San Pedro Basin, Southern California
Bight. In: Summerhayes, C.P., Prell, W.L., Emeis, K.C.(Eds.), Upwelling system: evolution since the Early Mio-cene. Geol. Soc. Spec. Publ. 64, 77^91.
Wei, W., Pospichal, J., 1991. Danian calcareous nannofossilsuccession at ODP Site 738 in the southern Indian Ocean.In: Barron, J., Larsen, B., et al. (Eds.), Proc. ODP Sci.Results 119, 495^512.
Wind, F.H., 1979. Maestrichtian^Campanian nanno£oralprovinces of the southern Atlantic and Indian Oceans. In:Talwani, M., et al. (Eds.), Deep Drilling Results in the At-lantic Ocean: Continental Margins and Paleoenvironment.Washington, Am. Geophys. Union, pp. 123^137.
Worsley, T.R., 1974. The Cretaceous^Tertiary boundary event.In: Hay, W.W. (Ed.), Studies in Paleoceanography. SEPMSpec. Publ. 20, 94^125.
Worsley, T., Martini, E., 1970. Late Maastrichtian Nanno-plankton provinces. Nature 225, 1242^1243.
Young, J.R., 1994. Functions of coccoliths. In: Winter, A.,Siesser, W.G. (Eds.), Coccolithophores. Cambridge Univer-sity Press, Cambridge, pp. 63^82.
Zachos, J.C., Arthur, M.A., Dean, W.E., 1989. Geochemicalevidence for suppression of pelagic marine productivity atthe Cretaceous^Tertiary boundary. Nature 337, 61^64.
MARMIC 905 27-1-03
A.A.A.M. Tantawy /Marine Micropaleontology 47 (2003) 323^356356
Related Documents











