Caffeic Acid 3,4-Dihydroxy-Phenethyl Ester Suppresses Receptor Activator of NF-kB Ligand–Induced Osteoclastogenesis and Prevents Ovariectomy-Induced Bone Loss Through Inhibition of Mitogen-Activated Protein Kinase/Activator Protein 1 and Ca 2þ –Nuclear Factor of Activated T-cells Cytoplasmic 1 Signaling Pathways Xian Wu, 1 * Zhenxi Li, 1,2 * Zhengfeng Yang , 1 Chunbing Zheng , 1 Ji Jing, 1 Yihua Chen , 1 Xiyun Ye , 1 Xiaoyuan Lian, 1 Wenwei Qiu, 3 Fan Yang, 3 Jie Tang, 3 Jianru Xiao, 2 * Mingyao Liu, 1,2,4 * and Jian Luo 1,2 * 1 Shanghai Key Laboratory of Regulatory Biology, Institute of Biomedical Sciences and School of Life Sciences, East China Normal University, Shanghai, China 2 East China Normal University and Shanghai Changzheng Hospital Joint Research Center for Orthopedic Oncology, Shanghai, China 3 Institute of Medicinal Chemistry and Department of Chemistry, East China Normal University, Shanghai, China 4 Alkek Institute of Biosciences and Technology, Texas A&M University Health Science Center, Houston, TX, USA ABSTRACT Receptor activator of NF-kB ligand (RANKL) stimulation leads to the activation of mitogen-activated protein kinase (MAPK)/AP-1 and Ca 2þ –nuclear factor of activated T-cells cytoplasmic 1 (NFATc1) signaling pathways in osteoclastogenesis. Targeting these pathways has been an encouraging strategy for bone-related diseases, such as postmenopausal osteoporosis. In this study, we examined the effects of caffeic acid 3,4-dihydroxy-phenethyl ester (CADPE) on osteoclastogenesis. In mouse bone marrow monocytes (BMMs) and RAW264.7 cells, CADPE suppressed RANKL-induced osteoclast differentiation and actin-ring formation in a dose-dependent manner within non–growth inhibitory concentrations at the early stage, while CADPE had no effect on macrophage colony-stimulating factor (M-CSF)-induced proliferation and differentiation. At the molecular level, CADPE inhibited RANKL-induced phosphorylation of MAPKs, including extracellular signal-regulated kinases 1/2 (ERK1/2), p38, and c-Jun N-terminal kinase (JNK), without significantly affecting the NF-kB signaling pathway. CADPE abrogated RANKL-induced activator protein 1 (AP-1)/FBJ murine osteosarcoma viral oncogene homolog (c-Fos) nuclear translocation and activation. Overexpression of c-Fos prevented the inhibition by CADPE of osteoclast differentiation. Furthermore, CADPE suppressed RANKL-induced the tumor necrosis factor receptor associated factor 6 (TRAF6) interaction with c-src tyrosine kinase (c-Src), blocked RANKL-induced the phosphorylation of protein kinase B (AKT), and inhibited RANKL-induced Ca 2þ oscillation. As a result, CADPE decreased osteoclastogenesis-related marker gene expression, including NFATc1, TRAP, cathepsin K, and c-Src. To test the effects of CADPE on osteoclast activity in vivo, we showed that CADPE prevented ovariectomy- induced bone loss by inhibiting osteoclast activity. Together, our data demonstrate that CADPE suppresses osteoclastogenesis and bone loss through inhibiting RANKL-induced MAPKs and Ca 2þ -NFATc1 signaling pathways. CADPE is a novel agent in the treatment of osteoclast-related diseases, such as osteoporosis. ß 2012 American Society for Bone and Mineral Research. KEY WORDS: CAFFEIC ACID; MAPK/AP-1; Ca 2þ -NFATc1; MENOPAUSE; OSTEOPOROSIS Introduction B one homeostasis is balanced between bone formation by osteoblasts and bone resorption by osteoclasts, which occurs throughout our lives. (1) Deficiency in bone formation by osteoblasts or excessive bone resorption by osteoclasts can cause a number of osteopenic and pathologic diseases, especially osteoporosis. (2–4) Therefore, identification of agents that either increase bone formation or block osteoclast resorption are essential for the development of therapeutic ORIGINAL ARTICLE J JBMR Received in original form March 8, 2011; revised form January 30, 2012; accepted February 6, 2012. Published online February 15, 2012. Address correspondence to: Jian Luo, PhD, East China Normal University, 500 Dongchuan Road, Shanghai 200241, China. E-mail: [email protected] Additional Supporting Information may be found in the online version of this article. *XW, ZL, and JX contributed equally to this work. ML and JL are co-senior authors. Journal of Bone and Mineral Research, Vol. 27, No. 6, June 2012, pp 1298–1308 DOI: 10.1002/jbmr.1576 ß 2012 American Society for Bone and Mineral Research 1298

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Caffeic Acid 3,4-Dihydroxy-Phenethyl Ester SuppressesReceptor Activator of NF-kB Ligand–InducedOsteoclastogenesis and Prevents Ovariectomy-InducedBone Loss Through Inhibition of Mitogen-ActivatedProtein Kinase/Activator Protein 1 and Ca2þ–NuclearFactor of Activated T-cells Cytoplasmic 1 SignalingPathways
Xian Wu,1* Zhenxi Li ,1,2* Zhengfeng Yang,1 Chunbing Zheng ,1 Ji Jing ,1 Yihua Chen ,1 Xiyun Ye ,1
Xiaoyuan Lian,1 Wenwei Qiu,3 Fan Yang,3 Jie Tang,3 Jianru Xiao,2* Mingyao Liu,1,2,4* and Jian Luo1,2*1Shanghai Key Laboratory of Regulatory Biology, Institute of Biomedical Sciences and School of Life Sciences,East China Normal University, Shanghai, China
2East China Normal University and Shanghai Changzheng Hospital Joint Research Center for Orthopedic Oncology, Shanghai, China3Institute of Medicinal Chemistry and Department of Chemistry, East China Normal University, Shanghai, China4Alkek Institute of Biosciences and Technology, Texas A&M University Health Science Center, Houston, TX, USA
ABSTRACTReceptor activator of NF-kB ligand (RANKL) stimulation leads to the activation of mitogen-activated protein kinase (MAPK)/AP-1
and Ca2þ–nuclear factor of activated T-cells cytoplasmic 1 (NFATc1) signaling pathways in osteoclastogenesis. Targeting these pathways
has been an encouraging strategy for bone-related diseases, such as postmenopausal osteoporosis. In this study, we examined the
effects of caffeic acid 3,4-dihydroxy-phenethyl ester (CADPE) on osteoclastogenesis. In mouse bone marrow monocytes (BMMs) and
RAW264.7 cells, CADPE suppressed RANKL-induced osteoclast differentiation and actin-ring formation in a dose-dependent manner
within non–growth inhibitory concentrations at the early stage, while CADPE had no effect on macrophage colony-stimulating factor
(M-CSF)-induced proliferation and differentiation. At the molecular level, CADPE inhibited RANKL-induced phosphorylation of MAPKs,
including extracellular signal-regulated kinases 1/2 (ERK1/2), p38, and c-Jun N-terminal kinase (JNK), without significantly affecting the
NF-kB signaling pathway. CADPE abrogated RANKL-induced activator protein 1 (AP-1)/FBJ murine osteosarcoma viral oncogene
homolog (c-Fos) nuclear translocation and activation. Overexpression of c-Fos prevented the inhibition by CADPE of osteoclast
differentiation. Furthermore, CADPE suppressed RANKL-induced the tumor necrosis factor receptor associated factor 6 (TRAF6)
interaction with c-src tyrosine kinase (c-Src), blocked RANKL-induced the phosphorylation of protein kinase B (AKT), and inhibited
RANKL-induced Ca2þ oscillation. As a result, CADPE decreased osteoclastogenesis-related marker gene expression, including NFATc1,
TRAP, cathepsin K, and c-Src. To test the effects of CADPE on osteoclast activity in vivo, we showed that CADPE prevented ovariectomy-
induced bone loss by inhibiting osteoclast activity. Together, our data demonstrate that CADPE suppresses osteoclastogenesis and bone
loss through inhibiting RANKL-induced MAPKs and Ca2þ-NFATc1 signaling pathways. CADPE is a novel agent in the treatment of
osteoclast-related diseases, such as osteoporosis. � 2012 American Society for Bone and Mineral Research.
KEY WORDS: CAFFEIC ACID; MAPK/AP-1; Ca2þ-NFATc1; MENOPAUSE; OSTEOPOROSIS
Introduction
Bone homeostasis is balanced between bone formation by
osteoblasts and bone resorption by osteoclasts, which
occurs throughout our lives.(1) Deficiency in bone formation by
osteoblasts or excessive bone resorption by osteoclasts can
cause a number of osteopenic and pathologic diseases,
especially osteoporosis.(2–4) Therefore, identification of agents
that either increase bone formation or block osteoclast
resorption are essential for the development of therapeutic
ORIGINAL ARTICLE JJBMR
Received in original form March 8, 2011; revised form January 30, 2012; accepted February 6, 2012. Published online February 15, 2012.
Address correspondence to: Jian Luo, PhD, East China Normal University, 500 Dongchuan Road, Shanghai 200241, China. E-mail: [email protected]
Additional Supporting Information may be found in the online version of this article.
*XW, ZL, and JX contributed equally to this work. ML and JL are co-senior authors.
Journal of Bone and Mineral Research, Vol. 27, No. 6, June 2012, pp 1298–1308
DOI: 10.1002/jbmr.1576
� 2012 American Society for Bone and Mineral Research
1298

drugs for osteoporosis. However, the drugs for the disease are far
from ideal.(5) For example, the therapeutic peptide parathyroid
hormone (PTH), which can induce bone formation, must be
given as an injection, and has a 2-year maximal treatment course
in clinic due to bone cancer concerns.(6) Bisphosphonates, which
can inhibit osteoclast resorption, however, are poorly absorbed
and can cause damage to the gastrointestinal tract.(7) In addition,
estrogen-replacement therapy, often prescribed for postmeno-
pausal women, has been proven to increase the risk for uterine
and breast cancer, heart attack, and blood clots, and its use
has been discouraged for long-term treatment.(8) Therefore,
there is considerable scientific and lay public interest in finding
alternative agents and treatments for osteoporosis.
Previous research in postmenopausal animals shows that the
main reason for postmenopausal osteoporosis is estrogen
deficiency accompanied by production of pro-osteoclastogenic
cytokines, such as tumor necrosis factor a (TNFa) and receptor
activator of NF-kB ligand (RANKL),(9) which strongly increases
osteoclast lifespan and activity.(4,10) Therefore, inhibition of
osteoclastogenesis is a promising strategy for postmenopausal
osteoporosis treatment. Osteoclasts are multinucleated cells
formed by the differentiation and fusion of bone marrow
monocytes (BMMs) and are the unique cells involved in bone
resorption.(11)
Previous studies indicated that two cytokines, macrophage
colony-stimulating factor (M-CSF) and RANKL are essential and
sufficient to promote osteoclastogenesis. M-CSF can induce the
proliferation of BMMs and their differentiation into osteoclast
precursors, and RANKL can subsequently induce the differentia-
tion of osteoclast precursors intomature osteoclasts.(2,12) Binding
of RANKL to its receptor RANK results in the recruitment of the
adaptor molecules TNF receptor-associated factors (TRAFs).(11)
The formation of the RANK-TRAFs complex mediates mitogen-
activated protein kinases (MAPKs) signaling pathways in
osteoclast formation.(13,14) In particular, RANK-TRAF6 complex
can lead to the activation of phosphatidylinositol 3-kinase (PI3K)/
protein kinase B (AKT) signaling pathway by recruiting Src
family kinases.(2,11,15,16) Activation of PI3K finally releases
intracellular Ca2þ,(17–19) which can activate the central transcrip-
tional factor in osteoclastogenesis, nuclear factor of activated
T-cells cytoplasmic 1 (NFATc1).(20) NFATc1 then translocates to
the nucleus and activates the expression of multiple osteoclas-
togenesis-related genes, such as tartrate-resistant acid phospha-
tase (TRAP), latent transforming growth factor beta binding
protein3 (LTBP3), chloride channel (ClC7), MMP9, calcitonin
receptor (CTR), cathepsin K, and c-Src.(21)
Caffeic acid 3,4-dihydroxy-phenethyl ester (CADPE), a com-
pound originally isolated frommedicinal plants Sarcandra glabra
and Teucrium pilosum,(22) has attracted much interest due to its
proven pharmacologic safety and its many biologic activities,
such as induction of cancer senescence,(23) inhibition of
tumor angiogenesis,(24) suppression of hepatocellular carcinoma
growth,(25) and gastric carcinoma cell migration.(26) In the
present study, we investigated the effects of CADPE on
osteoclastogenesis both in vitro and in vivo, and elucidated
the underlying molecular mechanisms. We found that CADPE
suppressed RANKL-induced osteoclast differentiation within
non–growth inhibitory concentrations at an early stage of
osteoclastogenesis. Addition of CADPE disrupted the formation
of the actin-ring during osteoclastogenesis in vitro, and
prevented ovariectomy-induced bone loss in vivo. Furthermore,
we found that CADPE inhibited NFATc1 expression by blocking
the MAPK/AP-1 signaling pathway and Ca2þ oscillation, but
not the NF-kB signaling pathway in RANKL-induced osteoclast
differentiation. Finally, CADPE decreased the expression levels
of osteoclast-related marker genes, including TRAP, cathepsin K,
and c-Src. Therefore, our data show that CADPE suppresses
osteoclastogenesis and osteoclast activity through MAPK/AP-1
and Ca2þ/NFAT signaling pathways.
Materials and Methods
Reagents and antibodies
CADPE was synthesized by the Department of Chemistry at
East China Normal University as described.(5) RAW264.7 cells
were a kind gift from Dr. B.G. Darnay (the University of Texas M.D.
Anderson Cancer Center, Houston, TX, USA). Penicillin, strepto-
mycin, and amodified essential medium (a-MEM) were obtained
from Invitrogen (Carlsbad, CA, USA), and fetal bovine serum (FBS)
was obtained from the FRONT Company. All antibodies were
purchased from Cell Signaling Technology (Beverly, MA, USA)
except the NFATc1 antibody, which was from Santa Cruz
Biotechnology (Santa Cruz, CA, USA). Fugene HD transfection
reagent (Roche, Shanghai, China) was used to transfect
RAW264.7 cells. TRIZOL reagent was purchased from Invitrogen
(Carlsbad, CA, USA), and the PrimeScript RT reagent Kit
was obtained from TaKaRa Biotechnology. Bacteria-derived
recombinant mouse RANKL and M-CSF were from R&D Systems
(Minneapolis, MN, USA). Tris, glycine, NaCl, SDS, and other
regents were obtained from Sigma (St. Louis, MO, USA).
Cytotoxicity assay
We detected the cytotoxic effect of CADPE by the sulforhoda-
mine B (SRB) assay method(27) using a VERSA max microplate
reader (Molecular Devices, Sunnyvale, CA, USA).
In vitro osteoclastogenesis assay
For primary cell cultures, we cultured bone marrow cells isolated
from C57/BL6 mice as described.(28) Briefly, BMMs were isolated
from femurs and tibias of 4-week-old C57BL/6 mice and cultured
in the presence of M-CSF (20 ng/mL) for 3 days. For differentia-
tion into mature osteoclasts, osteoclast precursors were cultured
with M-CSF (20 ng/mL) and RANKL (30 ng/mL) in 48-well culture
plates for 7 days with media changed every 2 days. For TRAP
staining, the cells were fixed and stained for TRAP activity
(Sigma). TRAP-positive multinucleated cells with >5 nuclei
were counted as osteoclasts. For RAW264.7 cells, culture was
performed as described.(28)
Actin-ring formation and pit assay
After stimulation with M-CSF (20 ng/mL) and RANKL (30 ng/mL),
we fixed the cells with 4% paraformaldehyde for 10 minutes
and washed them three times with PBS. Then the cells were
incubated with fluorescein isothiocyanate (FITC)-phalloidin for
Journal of Bone and Mineral Research CADPE INHIBITS RANKL-INDUCED OSTEOCLASTOGENESIS 1299

1 hour followed with 4,6-diamidino-2-phenylindole (DAPI)
staining for 10 minutes. For pit assay, osteoclasts were cultured
on FBS-coated dentin slices as described.(29)
Ovariectomized mouse model and bonehistomorphometric analysis
We created an ovariectomized mouse model as described.(30)
Maintenance, use, and treatment of all animals in this study were
in accordance with accepted standards of the Ethics Committee
at East China Normal University. Briefly, 7 days after ovariectomy,
mice were divided into three groups of six mice each: sham-
operatedmice (SHAM), ovariectomizedmice treated with vehicle
(OVX) and OVX mice treated with CADPE. CADPE (10mg/kg) or
dimethylsulfoxide (DMSO) was injected (intraperitoneally [i.p.])
into the ovariectomized mice every 2 days. After 3 months, all
mice were euthanized with excess amounts of anesthetic. The
body weight of these mice was measured every 2 days. Sections
of lumbar vertebrae (L3) were obtained for histomorphometric
measurements using the OsteoMeasure Analysis System
(Osteometrics, Decatur, GA, USA) according to standard criteria.
For osteoclast TRAP staining, calvariae were isolated and fixed in
10% paraformaldehyde fixation buffer (PFA) and decalcification
performed with 10% EDTA for 2 weeks. The samples were
embedded in paraffin for TRAP staining (Sigma).
Real-time RT-PCR
For the real-time RT-PCR analysis, we extracted total RNA
from cells with TRIZOL (Invitrogen) as described.(31) NFATc1,
cathepsin K, and TRAP transcripts were quantified on a Mx 3005P
(Stratagene, Santa Clara, CA, USA) using SYBR green dye and
normalized with b-actin. The following primer sets were used:
mouse TRAP: forward, 50-GCTGGAAACCATGATCACCT-30; reverse,50-GAGTTGCCACACAGCATCAC-30; mouse Cathepsin K: forward,
50-CTTCCAATACGTGCAGCAGA-30; reverse, 50-TCTTCAGGGCTTT-CTCGTTC-30; mouse NFATc1: forward, 50-TGGAGAAGCAGAGC-ACAGAC-30; reverse, 50-GCGGAAAGGTGGTATCTCAA-30; and
mouse b-actin: forward, 50-GTACGCCAACACAGTGCTG-30;reverse, 50-CGTCATACTCCTGCTTGCTG-30.
Luciferase reporter gene assay
The effect of CADPE on the RANKL-induced AP-1-luciferase
reporter assay was determined as described.(32,33)
Intracellular calcium imaging
We cultured BMMs in 35-mm cell culture dishes with or without
CADPE for 60 hours in the presence of RANKL (30 ng/mL) and
M-CSF (20 ng/mL). After washing with PBS, cells were incubated
with 5mM Fura-2/AM at 378C, and then imaged at 340
and 380 nM excitation wavelengths to detect intracellular free
calcium (with an Olympus IX71 and LAMBDA DG-4) and recorded
by InVivo software; analysis was by Image-Pro Analyzer 6.2
software (Media Cybernetics, Bethesda, MD, USA).(19)
Statistical analysis
All experimental data are presented as the mean� SD, with
values frommore than three experiments. Statistical significance
was determined by the Student t test.
Results
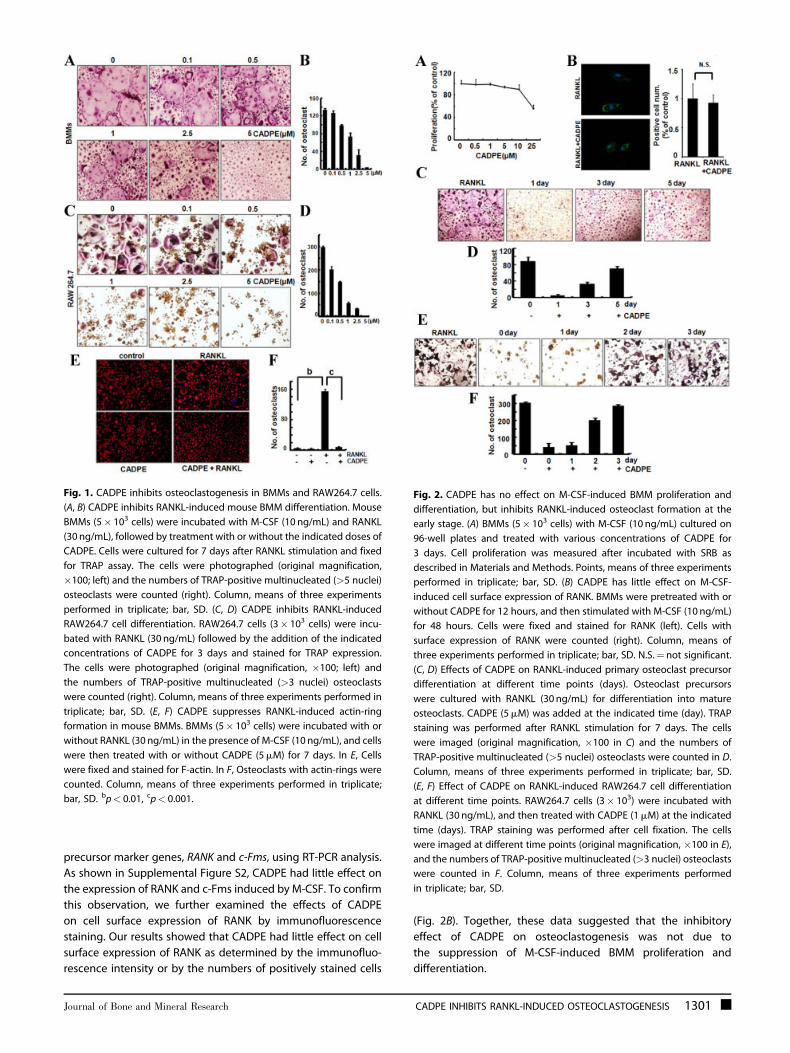
CADPE inhibits osteoclastogenesis in mouse BMMs andRAW264.7 cells
To examine the effects of CADPE on osteoclastogenesis, we used
two standard in vitro osteoclast differentiation models. The first
consisted of BMMs with RANKL and M-CSF stimulation. The
second was a mouse osteoclast precursor cell line RAW264.7
with RANKL stimulation.(28) BMMs were incubated with various
concentrations of CADPE during differentiation for 7 days, and
RAW264.7 cells with CADPE for 3 to 5 days. As shown in Fig. 1A–D,
CADPE inhibited osteoclast formation in a dose-dependent
manner in both BMMs and RAW264.7 cells, suggesting that
administration of CADPE in the differentiation cell models
inhibited osteoclast differentiation; with an one-half maximal
inhibitory concentration (IC50) of 0.5 to 1mM in BMMs and
RAW264.7 cells.
CADPE inhibits RANKL-induced actin-ring formationin osteoclasts
To further explore the effect of CADPE on osteoclastogenesis, we
tested whether CADPE could affect the actin-ring formation,
which is a prerequisite for osteoclast bone resorption and is the
most obvious character of mature osteoclasts during osteoclas-
togenesis.(34) In the presence of RANKL stimulation, BMMs can
differentiate into mature osteoclasts and form obvious actin-ring
structures by FITC-phalloidin staining (Fig. 1E). However, the size
and the number of actin-ring structures were significantly
decreased when cells were treated with 2.5 to 5mM CADPE
(Fig. 1E), suggesting that CADPE suppressed the formation of the
actin-ring in osteoclasts.
To investigate whether the blockage of osteoclastogenesis
and actin-ring formation by CADPE was due to potential toxicity
of this compound, we further examined the cytotoxicity of
CADPE by SRB assay on both BMMs and RAW264.7 cells
(Supplemental Fig. S1A and S1B). Although cytotoxicity was
observed when higher doses of CADPE were used, CADPE at
5mM (which completely inhibits osteoclastogenesis), did not
induce cytotoxicity significantly in both BMMs and RAW264.7
cells. These data indicate that the inhibitory effect of CADPE
on osteoclast differentiation was not due to the cytotoxicity of
the compound.
CADPE has no effect on M-CSF-induced proliferationand differentiation of BMMs
Osteoclastogenesis is a multistep process that can be separated
into two major events: the proliferation of BMMs and their
differentiation into osteoclast precursors induced by M-CSF;
and the subsequent differentiation of osteoclast precursors into
mature osteoclasts induced by RANKL.(12) To examine which step
CADPE affects in osteoclastogenesis, we first investigated the
effect of CADPE on BMM proliferation induced by M-CSF. Similar
to the cytotoxicity assay results, CADPE had little effect on
M-CSF-induced proliferation of BMMs at low concentrations (less
than 10mM) (Fig. 2A). To examine the effects of CADPE on BMM
differentiation into osteoclast precursors induced by M-CSF, we
investigated the expression levels of M-CSF–induced osteoclast
1300 WU ET AL. Journal of Bone and Mineral Research
precursor marker genes, RANK and c-Fms, using RT-PCR analysis.
As shown in Supplemental Figure S2, CADPE had little effect on
the expression of RANK and c-Fms induced by M-CSF. To confirm
this observation, we further examined the effects of CADPE
on cell surface expression of RANK by immunofluorescence
staining. Our results showed that CADPE had little effect on cell
surface expression of RANK as determined by the immunofluo-
rescence intensity or by the numbers of positively stained cells
(Fig. 2B). Together, these data suggested that the inhibitory
effect of CADPE on osteoclastogenesis was not due to
the suppression of M-CSF-induced BMM proliferation and
differentiation.
Fig. 1. CADPE inhibits osteoclastogenesis in BMMs and RAW264.7 cells.
(A, B) CADPE inhibits RANKL-induced mouse BMM differentiation. Mouse
BMMs (5� 103 cells) were incubated with M-CSF (10 ng/mL) and RANKL
(30 ng/mL), followed by treatment with or without the indicated doses of
CADPE. Cells were cultured for 7 days after RANKL stimulation and fixed
for TRAP assay. The cells were photographed (original magnification,
�100; left) and the numbers of TRAP-positive multinucleated (>5 nuclei)
osteoclasts were counted (right). Column, means of three experiments
performed in triplicate; bar, SD. (C, D) CADPE inhibits RANKL-induced
RAW264.7 cell differentiation. RAW264.7 cells (3� 103 cells) were incu-
bated with RANKL (30 ng/mL) followed by the addition of the indicated
concentrations of CADPE for 3 days and stained for TRAP expression.
The cells were photographed (original magnification, �100; left) and
the numbers of TRAP-positive multinucleated (>3 nuclei) osteoclasts
were counted (right). Column, means of three experiments performed in
triplicate; bar, SD. (E, F) CADPE suppresses RANKL-induced actin-ring
formation in mouse BMMs. BMMs (5� 103 cells) were incubated with or
without RANKL (30 ng/mL) in the presence of M-CSF (10 ng/mL), and cells
were then treated with or without CADPE (5mM) for 7 days. In E, Cells
were fixed and stained for F-actin. In F, Osteoclasts with actin-rings were
counted. Column, means of three experiments performed in triplicate;
bar, SD. bp< 0.01, cp< 0.001.
Fig. 2. CADPE has no effect on M-CSF-induced BMM proliferation and
differentiation, but inhibits RANKL-induced osteoclast formation at the
early stage. (A) BMMs (5� 103 cells) with M-CSF (10 ng/mL) cultured on
96-well plates and treated with various concentrations of CADPE for
3 days. Cell proliferation was measured after incubated with SRB as
described in Materials and Methods. Points, means of three experiments
performed in triplicate; bar, SD. (B) CADPE has little effect on M-CSF-
induced cell surface expression of RANK. BMMs were pretreated with or
without CADPE for 12 hours, and then stimulated with M-CSF (10 ng/mL)
for 48 hours. Cells were fixed and stained for RANK (left). Cells with
surface expression of RANK were counted (right). Column, means of
three experiments performed in triplicate; bar, SD. N.S.¼not significant.
(C, D) Effects of CADPE on RANKL-induced primary osteoclast precursor
differentiation at different time points (days). Osteoclast precursors
were cultured with RANKL (30 ng/mL) for differentiation into mature
osteoclasts. CADPE (5mM) was added at the indicated time (day). TRAP
staining was performed after RANKL stimulation for 7 days. The cells
were imaged (original magnification, �100 in C) and the numbers of
TRAP-positive multinucleated (>5 nuclei) osteoclasts were counted in D.
Column, means of three experiments performed in triplicate; bar, SD.
(E, F) Effect of CADPE on RANKL-induced RAW264.7 cell differentiation
at different time points. RAW264.7 cells (3� 103) were incubated with
RANKL (30 ng/mL), and then treated with CADPE (1mM) at the indicated
time (days). TRAP staining was performed after cell fixation. The cells
were imaged at different time points (original magnification, �100 in E),
and the numbers of TRAP-positive multinucleated (>3 nuclei) osteoclasts
were counted in F. Column, means of three experiments performed
in triplicate; bar, SD.
Journal of Bone and Mineral Research CADPE INHIBITS RANKL-INDUCED OSTEOCLASTOGENESIS 1301

CADPE inhibits RANKL-induced osteoclast formation atthe early stage, but has little effect on mature osteoclastformation and resorption
To determine the effects of CADPE on RANKL-induced osteoclast
precursor differentiation into mature osteoclasts, we examined
the differentiation process of primary osteoclast precursors
beginning on day 0 to day 5 with RANKL stimulation in the
absence or presence of CADPE (Fig. 2C). Our results showed that
administration of CADPE strongly inhibited RANKL-induced
osteoclast differentiation during the first day of differentiation
(Fig. 2C, 1 day). However, CADPE was not effective in the
inhibition of osteoclast differentiation at later stages, especially
at day 5 (Fig. 2D). Similar results were obtained in RANKL-induced
RAW264.7 cell differentiation (Fig. 2E, F), suggesting that CADPE
inhibited RANKL-induced osteoclast differentiation at an early
stage.
To further explore whether CADPE inhibited osteoclast bone
resorption, we performed bone resorption assays using BMMs.
When BMMs were cultured on a dentin slice, mature osteoclasts
caused the resorption of lacunae and the formation of pits in the
present of M-SCF and RANKL, as compared to blank slice controls
(Fig. 3A, top left and middle). The number and area of pits on the
surface of the dentin slices were markedly decreased by the
addition of CADPE at the beginning of the assays (Fig. 3A, top
right). Similar results were obtained by confocal microscopy
scanning (Fig. 3A, bottom). These data were consistent with
previous results that CADPE inhibited osteoclast differentiation
and actin-ring formation, and as a result, CADPE inhibited
osteoclast activity. However, after RANKL stimulation for 6 days,
matured osteoclasts were formed and then seeded on dentin
slices, followed by incubation with or without CADPE for an
additional 2 days. The bone resorption lacunae pits and erosion
areas showed no significant differences between the control
slices and CADPE-treated slices (Fig. 3B), suggesting that CADPE
had little effect on mature osteoclast bone resorption. To further
investigate the effect of CADPE on mature osteoclasts, we
examined the sealing zone formation, which is essential to
osteoclast attachment to the bone and its resorption.(35) Our data
showed that CADPE exerted little effect on the sealing zone
dimensions, including the sealing zone width, perimeter, and
height (Supplemental Fig. S3C). Together, our data suggested
that CADPE exerted no effect on M-CSF-induced BMM
proliferation and differentiation, but inhibited RANKL-induced
osteoclast formation at the early stages.
CADPE prevents ovariectomy-induced bone loss byinhibiting osteoclast activity and stimulating osteoblastdifferentiation in vivo
Activation of osteoclasts plays an essential role in bone-related
diseases, including postmenopausal osteoporosis, in which
estrogen deficiency enhances the genesis and activity of
osteoclasts; and results in an unbalanced increase in bone
resorption.(36) To examine whether CADPE inhibits osteoclasto-
genesis in vivo, we used the ovariectomized mouse model
to mimic menopause-induced bone loss in women.(36) The
uterine weight data showed that the ovariectomy operation
was successful (Supplemental Fig. 4). Von Kossa staining and
histomorphometric analysis of lumbar vertebrae also indicated
that bone volume (BV/TV) and trabecular number (Tb.N) in the
ovariectomized (OVX) mice decreased dramatically, whereas
trabecular spacing (Tb.Sp) was dramatically increased when
compared with sham-operated controls (SHAM). Treatment of
CADPE (10mg/kg) in ovariectomized mice (OVXþCADPE)
significantly inhibited the ovariectomy-induced bone loss
as measured in these parameters (Fig. 4A, B). To investigate
whether CADPE prevented bone loss through inhibition of
Fig. 3. CADPE inhibits osteoclast bone resorption at early stages, but has
little effect on mature osteoclast bone resorption. (A) CADPE suppresses
osteoclast bone resorption at the early stage. BMMs (5� 103 cells) were
induced by M-CSF (10 ng/mL) and RANKL (30 ng/mL) with or without
CADPE (5mM) on dentine slices for 7 days. Dentine slices were stained
with Mayer’s hematoxylin after removal of cells. The resorption pits were
visualized with light microscopy (original magnification, �10; top) and
confocal microscopy (original magnification, �40; bottom). (B) CADPE
has little effect on bone resorption by matured osteoclasts. BMMs
(5� 103 cells) were induced by M-CSF (10 ng/mL) and RANKL (30 ng/
mL) for 6 days. Mature osteoclast cells were digested with collagenase
and seeded on dentine slices followed by treatment with or without
CADPE (5mM) for an additional 2 days. Dentine slices were stained with
Mayer’s hematoxylin after removal of cells. The resorption pits were
visualized with light microscopy (original magnification, �10) and con-
focal microscopy (original magnification, �40). The numbers of pits and
erosion areas were analyzed with Image-Pro Plus software (bottom).
Column, means of three experiments performed in triplicate; bar, SD.
N.S.¼not significant.
1302 WU ET AL. Journal of Bone and Mineral Research
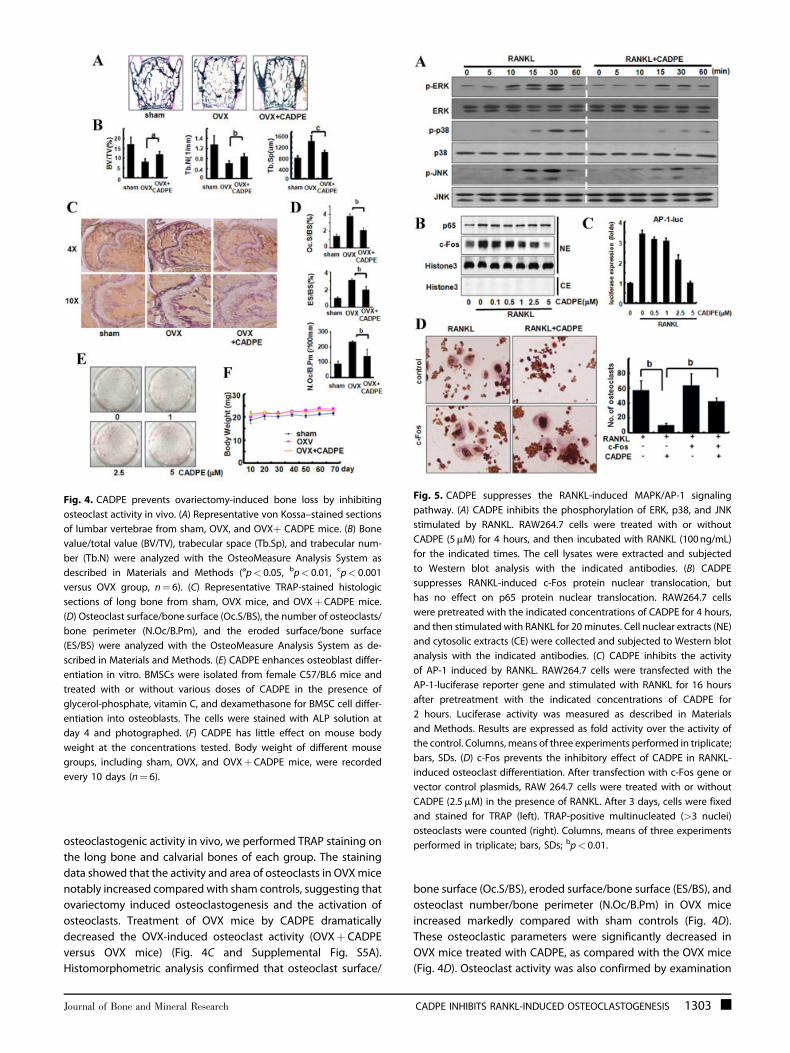
osteoclastogenic activity in vivo, we performed TRAP staining on
the long bone and calvarial bones of each group. The staining
data showed that the activity and area of osteoclasts in OVXmice
notably increased compared with sham controls, suggesting that
ovariectomy induced osteoclastogenesis and the activation of
osteoclasts. Treatment of OVX mice by CADPE dramatically
decreased the OVX-induced osteoclast activity (OVXþ CADPE
versus OVX mice) (Fig. 4C and Supplemental Fig. S5A).
Histomorphometric analysis confirmed that osteoclast surface/
bone surface (Oc.S/BS), eroded surface/bone surface (ES/BS), and
osteoclast number/bone perimeter (N.Oc/B.Pm) in OVX mice
increased markedly compared with sham controls (Fig. 4D).
These osteoclastic parameters were significantly decreased in
OVX mice treated with CADPE, as compared with the OVX mice
(Fig. 4D). Osteoclast activity was also confirmed by examination
Fig. 4. CADPE prevents ovariectomy-induced bone loss by inhibiting
osteoclast activity in vivo. (A) Representative von Kossa–stained sections
of lumbar vertebrae from sham, OVX, and OVXþ CADPE mice. (B) Bone
value/total value (BV/TV), trabecular space (Tb.Sp), and trabecular num-
ber (Tb.N) were analyzed with the OsteoMeasure Analysis System as
described in Materials and Methods (ap< 0.05, bp< 0.01, cp< 0.001
versus OVX group, n¼ 6). (C) Representative TRAP-stained histologic
sections of long bone from sham, OVX mice, and OVXþCADPE mice.
(D) Osteoclast surface/bone surface (Oc.S/BS), the number of osteoclasts/
bone perimeter (N.Oc/B.Pm), and the eroded surface/bone surface
(ES/BS) were analyzed with the OsteoMeasure Analysis System as de-
scribed in Materials and Methods. (E) CADPE enhances osteoblast differ-
entiation in vitro. BMSCs were isolated from female C57/BL6 mice and
treated with or without various doses of CADPE in the presence of
glycerol-phosphate, vitamin C, and dexamethasone for BMSC cell differ-
entiation into osteoblasts. The cells were stained with ALP solution at
day 4 and photographed. (F) CADPE has little effect on mouse body
weight at the concentrations tested. Body weight of different mouse
groups, including sham, OVX, and OVXþCADPE mice, were recorded
every 10 days (n¼ 6).
Fig. 5. CADPE suppresses the RANKL-induced MAPK/AP-1 signaling
pathway. (A) CADPE inhibits the phosphorylation of ERK, p38, and JNK
stimulated by RANKL. RAW264.7 cells were treated with or without
CADPE (5mM) for 4 hours, and then incubated with RANKL (100ng/mL)
for the indicated times. The cell lysates were extracted and subjected
to Western blot analysis with the indicated antibodies. (B) CADPE
suppresses RANKL-induced c-Fos protein nuclear translocation, but
has no effect on p65 protein nuclear translocation. RAW264.7 cells
were pretreated with the indicated concentrations of CADPE for 4 hours,
and then stimulated with RANKL for 20minutes. Cell nuclear extracts (NE)
and cytosolic extracts (CE) were collected and subjected to Western blot
analysis with the indicated antibodies. (C) CADPE inhibits the activity
of AP-1 induced by RANKL. RAW264.7 cells were transfected with the
AP-1-luciferase reporter gene and stimulated with RANKL for 16 hours
after pretreatment with the indicated concentrations of CADPE for
2 hours. Luciferase activity was measured as described in Materials
and Methods. Results are expressed as fold activity over the activity of
the control. Columns,means of three experiments performed in triplicate;
bars, SDs. (D) c-Fos prevents the inhibitory effect of CADPE in RANKL-
induced osteoclast differentiation. After transfection with c-Fos gene or
vector control plasmids, RAW 264.7 cells were treated with or without
CADPE (2.5mM) in the presence of RANKL. After 3 days, cells were fixed
and stained for TRAP (left). TRAP-positive multinucleated (>3 nuclei)
osteoclasts were counted (right). Columns, means of three experiments
performed in triplicate; bars, SDs; bp< 0.01.
Journal of Bone and Mineral Research CADPE INHIBITS RANKL-INDUCED OSTEOCLASTOGENESIS 1303

for serum TRAP5b and cross-linked C-telopeptide 1 (CTX-1),
which reflects osteoclast activity in vivo. As shown in
Supplemental Fig. S5B, the serum TRAP5b and CTX-1 levels
were increased in OVX mice compared with sham-operated
control mice, whereas CADPE treatment significantly decreased
the levels of TRAP5b and CTX-1 induced by OVX, suggesting
that CADPE could inhibit ovariectomy-induced osteoclast activity
in vivo.
Because bone remodeling is regulated by both osteoblasts
and osteoclasts, we also investigated the effect of CADPE on
osteogenesis. CADPE strikingly enhanced osteogenic differenti-
ation of bone marrow stromal cells (BMSCs) in a concentration-
dependent manner as evaluated by alkaline phosphatase (ALP)
staining (Fig. 4E and Supplemental Fig. S5C). To comprehensively
evaluate the effect of CADPE on osteogenesis, in vivo ALP
staining was performed. As shown in Supplemental Fig. S5D,
CADPE markedly augmented osteoblast activity in OVX mice,
which is consistent with the in vitro osteogenic differentiation
assay.
Furthermore, to examine whether CADPE had a specific effect
on ovariectomy-induced bone loss, the effect of CADPE on sham-
operated mice was explored. Our data indicated that CADPE had
no significant effect on the bone parameters including BV/TV,
Tb.N, and Tb.Sp between the sham control mice and the CADPE-
treated sham mice (Supplemental Fig. S6), Moreover, in the
CADPE-treated groups, CADPE had little effect on body weight
at the concentrations tested at 3 months (Fig. 4F), which is
consistent with a previous report,(24) suggesting little toxicity of
CADPE at the tested concentration.
CADPE inhibits RANKL-mediated MAPK/AP-1signaling pathway
RANKL-induced MAPK/AP-1 activation is among the very early
molecular events induced by RANKL,(2,37) and our observations
showed that CADPE inhibited RANKL-induced osteoclast forma-
tion at an early stage of differentiation. To understand the
signaling pathways mediated by CADPE in osteoclastogenesis,
we examined whether CADPE suppressed RANKL-induced
phosphorylation of extracellular signal-regulated kinase (ERK),
p38, and c-Jun N-terminal kinase (JNK) by Western blot analysis.
The phosphorylation of MAPKs reached its peak within 30min
of RANKL stimulation (Fig. 5A, left). Treatment with CADPE
significantly decreased the phosphorylation of ERK, p38, and JNK
when compared with the RANKL-stimulated control (Fig. 5A). To
further investigate whether the formation of transcription factor
AP-1 was affected by CADPE, we examined the c-Fos nuclear
translocation in RAW264.7 cells with CADPE treatment. Our data
showed that nuclear translocation of c-Fos was upregulated with
RANKL stimulation, whereas the addition of CADPE inhibited the
nuclear translocation in a dose-dependent manner (Fig. 5B). To
confirm the data generated by Western blot assays, we also
performed an AP-1-luciferase reporter gene assay. CADPE
inhibited RANKL-induced AP-1 activity in a concentration-
dependent manner (Fig. 5C). To further confirm that CADPE
inhibited osteoclast differentiation by suppressing RANKL-
induced AP-1 activity, we investigated whether overexpression
of c-Fos in RAW264.7 cells could prevent the inhibition of CADPE
on osteoclast differentiation. CADPE inhibited osteoclast differ-
entiation in control RAW264.7 cells (Fig. 5D, top, and
supplemental Fig. S7A). However, CADPE had little inhibitory
effect on osteoclastogenesis in c-Fos-overexpressed RAW264.7
cells (Fig. 5D, bottom). To further examine whether CADPE
selectively targets the MAPK signaling pathway in osteoclasts,
we investigated the activation of MAPKs in mouse adipocyte
3T3-L1 cells with CADPE treatment using the same conditions.
Our data showed that CADPE had little effect on MAPK activation
in 3T3-L1 cells (Supplemental Fig. S7B), suggesting that CADPE
may specifically target the MAPK signaling pathway in
osteoclasts.
CADPE has little effect on RANKL-induced NF-kBsignaling pathway
RANKL-induced NF-kB signaling pathway is another pivotal early
molecular event induced by RANKL.(38) To determine whether
CADPE affects NF-kB activation, we investigated NF-kB activation
by Western blot assay using specific antibodies. Interestingly,
RANKL-induced phosphorylation of p65 and degradation of IkBa
were not affected with CADPE treatment (Fig. 6A). Furthermore,
nuclear translocation and DNA-binding activity of p65 were
not affected by CADPE treatment in nuclear translocation assay
(Fig. 5B) and electrophoretic mobility gel shift assay (EMSA)
(Supplemental Fig. S8A), suggesting that CADPE has little effect
on the RANKL-induced NF-kB signaling pathway.
CADPE suppresses RANKL-induced Ca2þ oscillation
Besides the MAPK/AP-1 and NF-kB pathways, RANKL-induced
Ca2þ oscillations constitute another early molecular event during
osteoclast differentiation.(11) To further understand the mecha-
nism of CADPE’s inhibitory effects on osteoclast differentiation,
cytosolic Ca2þ signals were investigated in BMMs. Our results
showed that RANKL markedly induced Ca2þ oscillations (Fig. 6B,
middle). However, addition of CADPE significantly inhibited
the amplitude and frequency of Ca2þ oscillations, and kept
the oscillations at a relatively low level (Fig. 6B, right, and
Fig. 6C).
CADPE inhibits RANKL-induced TRAF6-c-Src associationand AKT phosphorylation
Because release of intracellular Ca2þ requires activation of the
PI3K/AKT signaling pathway,(17–19) and RANKL activates PI3K/AKT
through the TRAF6 and c-Src complex.(15,16) we then assessed the
effect of CADPE on the RANKL-induced association of TRAF6 with
c-Src by endogenous coimmunoprecipitation assay and the
effect of CADPE on the RANKL-induced phosphorylation of
AKT by Western blot analysis. Our data showed that CADPE
suppressed the RANKL-induced interaction between TRAF6 and
c-Src in a dose-dependent manner (Supplemental Fig. S9A).
Similarly, CADPE dose-dependently inhibited RANKL-induced
AKT phosphorylation (Supplemental Fig. S9B). All of our results
suggest that CADPE suppresses RANKL-induced Ca2þ oscillation
by inhibition of the TRAF6/c-Src/PI3K/AKT pathway.
1304 WU ET AL. Journal of Bone and Mineral Research
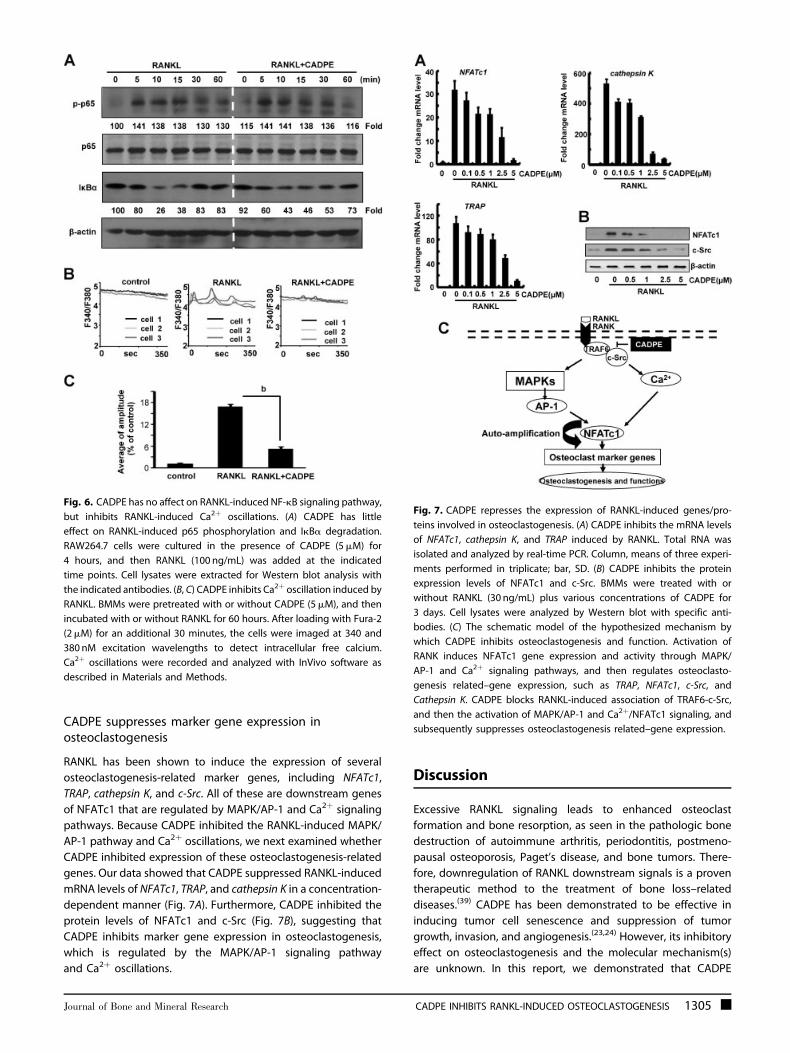
CADPE suppresses marker gene expression inosteoclastogenesis
RANKL has been shown to induce the expression of several
osteoclastogenesis-related marker genes, including NFATc1,
TRAP, cathepsin K, and c-Src. All of these are downstream genes
of NFATc1 that are regulated by MAPK/AP-1 and Ca2þ signaling
pathways. Because CADPE inhibited the RANKL-induced MAPK/
AP-1 pathway and Ca2þ oscillations, we next examined whether
CADPE inhibited expression of these osteoclastogenesis-related
genes. Our data showed that CADPE suppressed RANKL-induced
mRNA levels of NFATc1, TRAP, and cathepsin K in a concentration-
dependent manner (Fig. 7A). Furthermore, CADPE inhibited the
protein levels of NFATc1 and c-Src (Fig. 7B), suggesting that
CADPE inhibits marker gene expression in osteoclastogenesis,
which is regulated by the MAPK/AP-1 signaling pathway
and Ca2þ oscillations.
Discussion
Excessive RANKL signaling leads to enhanced osteoclast
formation and bone resorption, as seen in the pathologic bone
destruction of autoimmune arthritis, periodontitis, postmeno-
pausal osteoporosis, Paget’s disease, and bone tumors. There-
fore, downregulation of RANKL downstream signals is a proven
therapeutic method to the treatment of bone loss–related
diseases.(39) CADPE has been demonstrated to be effective in
inducing tumor cell senescence and suppression of tumor
growth, invasion, and angiogenesis.(23,24) However, its inhibitory
effect on osteoclastogenesis and the molecular mechanism(s)
are unknown. In this report, we demonstrated that CADPE
Fig. 6. CADPE has no affect on RANKL-induced NF-kB signaling pathway,
but inhibits RANKL-induced Ca2þ oscillations. (A) CADPE has little
effect on RANKL-induced p65 phosphorylation and IkBa degradation.
RAW264.7 cells were cultured in the presence of CADPE (5mM) for
4 hours, and then RANKL (100 ng/mL) was added at the indicated
time points. Cell lysates were extracted for Western blot analysis with
the indicated antibodies. (B, C) CADPE inhibits Ca2þ oscillation induced by
RANKL. BMMs were pretreated with or without CADPE (5mM), and then
incubated with or without RANKL for 60 hours. After loading with Fura-2
(2mM) for an additional 30 minutes, the cells were imaged at 340 and
380 nM excitation wavelengths to detect intracellular free calcium.
Ca2þ oscillations were recorded and analyzed with InVivo software as
described in Materials and Methods.
Fig. 7. CADPE represses the expression of RANKL-induced genes/pro-
teins involved in osteoclastogenesis. (A) CADPE inhibits the mRNA levels
of NFATc1, cathepsin K, and TRAP induced by RANKL. Total RNA was
isolated and analyzed by real-time PCR. Column, means of three experi-
ments performed in triplicate; bar, SD. (B) CADPE inhibits the protein
expression levels of NFATc1 and c-Src. BMMs were treated with or
without RANKL (30 ng/mL) plus various concentrations of CADPE for
3 days. Cell lysates were analyzed by Western blot with specific anti-
bodies. (C) The schematic model of the hypothesized mechanism by
which CADPE inhibits osteoclastogenesis and function. Activation of
RANK induces NFATc1 gene expression and activity through MAPK/
AP-1 and Ca2þ signaling pathways, and then regulates osteoclasto-
genesis related–gene expression, such as TRAP, NFATc1, c-Src, and
Cathepsin K. CADPE blocks RANKL-induced association of TRAF6-c-Src,
and then the activation of MAPK/AP-1 and Ca2þ/NFATc1 signaling, and
subsequently suppresses osteoclastogenesis related–gene expression.
Journal of Bone and Mineral Research CADPE INHIBITS RANKL-INDUCED OSTEOCLASTOGENESIS 1305

inhibited RANKL-induced osteoclast formation and actin-ring
at the early stages but had no effect on M-CSF-induced BMM
proliferation and differentiation. CADPE also prevented ovariec-
tomy-induced bone loss through the suppression of osteoclast
activity in a mouse model. At the molecular level, CADPE
suppressed RANKL-induced phosphorylation of MAPKs, includ-
ing ERK, p38, and JNK, and the subsequent nuclear translocation
and activation of AP-1/c-Fos, as well as RANKL-induced Ca2þ
oscillations by inhibition of the TRAF6/c-Src/PI3K/AKT pathway;
however, CADPE had no effect on the NF-kB signaling pathway.
Finally, CADPE inhibited expression of osteoclastogenesis-
related marker genes, such as NFATc1, TRAP, cathepsin K, and
c-Src.
It is well established that M-CSF induces the proliferation of
BMMs and their differentiation into the osteoclast precursor,
whereas RANKL induces subsequent differentiation of osteoclast
precursors into mature osteoclasts. In our study, we demonstrat-
ed that CADPE had little effect on M-CSF–induced BMM
proliferation and osteoclast precursor marker gene expression,
whereas CADPE inhibited the RANKL-induced differentiation of
osteoclast precursors into osteoclasts. These data indicates that
CADPE suppressed RANKL-induced osteoclastogenesis at the
step of osteoclast precursor differentiation into mature osteo-
clasts, and suggests that CADPE modulated the RANKL signaling
pathway.
Binding of RANKL to its receptor RANK results in the
recruitment of the adaptor molecule’s tumor necrosis factor
receptor-associated factors (TRAFs), including TRAF1, 2, 3, 5,
and 6.(11) The NF-kB signaling pathway is one of the key
downstream signaling pathways from the complex.(1) Our
research indicates that CADPE had no effect on the degradation
of nuclear factor of kappa light polypeptide gene enhancer in
B-cells inhibitor, alpha (IkBa) and the phosphorylation, nuclear
translocation, and DNA-binding activity of NF-kB subunit p65. All
of the results suggest that CADPE had little effect on the RANKL-
induced NF-kB signaling pathway.
RANKL-induced intracellular Ca2þ oscillation requires the PI3K/
AKT signaling pathway(11,19,40) and RANKL activates PI3K/AKT
through the TRAF6 and c-Src complex in osteoclastogen-
esis.(15,16) In our study, we verified that CADPE inhibited the
RANKL-induced interaction between TRAF6 and c-Src, sup-
pressed the phosphorylation of AKT, and abrogated the
oscillation of Ca2þ, suggesting that CADPE modulated the
TRAF/c-Src/PI3K/Ca2þ signaling pathway in osteoclastogenesis.
Moreover, genetic and pharmacological evidence shows that
PI3K can regulate the activation of MAPKs induced by
RANKL.(16,41) In our results, CADPE suppressed the phosphoryla-
tion of MAPKs. Furthermore, CADPE inhibited AP-1/c-Fos nuclear
translocation and activity. When c-Fos was overexpressed in
RAW264.7 cells, the inhibitory effect of CADPE in osteoclasto-
genesis was partially prevented. These results suggested that
CADPE suppression of MAPKs signaling pathway was due to
CADPE regulation of the TRAF/c-Src/PI3K signaling pathway in
osteoclastogenesis.
Previous research showed that in ovariectomized mice,
a decrease in circulating estrogen levels led to production
of osteoclastogenic cytokines, such as TNFa and RANKL in
osteoblasts and T cells, in turn leading to rapid bone loss.(42)
Using an ovariectomized mouse model, we have demonstrated
through our histomorphometric analysis and TRAP staining
analysis that CADPE prevented bone loss by suppressing
osteoclastogenesis and osteoclast activity. We also examined
the effects of CADPE on bone homeostasis by investigating
osteoblast differentiation and osteoblast activity using OVX mice
and ALP staining, and demonstrated that CADPE increased
osteoblast differentiation. Therefore, CADPE prevented ovariec-
tomy-induced bone loss by inhibiting osteoclastogenesis and
increasing the formation of osteoblasts. The effects of CADPE in
bone homeostasis are similar to the widely used osteoporosis
drugs, bisphosphonates, which also inhibit bone digestion
by osteoclasts and stimulate the formation of osteoblast
precursors.(43) Therefore, further investigation will be needed
to understand the mechanism of CADPE’s involvement in
osteoblast differentiation.
Collectively, our data demonstrate for the first time, that
CADPE can suppress osteoclastogenesis both in vitro and in vivo.
Furthermore, we show that CADPE inhibits osteoclastogenesis
through inhibition of the MAPK/AP-1 signaling pathway
and Ca2þ oscillations, which lead to the activation of NFATc1
and the expression of downstream genes. These results suggest
that CADPE could be a potential therapeutic candidate in
treating osteoclast-related diseases such as osteoporosis.
Disclosures
All authors state that they have no conflicts of interest.
Acknowledgments
This work is partially supported by the grants from the National
Basic Research Program of China (2012CB910400, 2010CB529704),
National Natural Science Foundation of China (30930055,
30800653, 81071437, and 20802020), and the Science and Tech-
nology Commission of Shanghai Municipality (11DZ2260300).
Authors’ roles: Study design: XW, ML, and JL. Study conduct:
XW, ZL, ZY, CZ, and JJ. Data analysis: XW, ZL, ZY, YC, XY, XL, JX, ML,
and JL. Chemical compound synthesis: WQ, FY, and JT. Drafting
manuscript: XW. Revising manuscript content: ML, and JL. JL
takes responsibility for the integrity of the data analysis.
References
1. Boyle WJ, Simonet WS, Lacey DL. Osteoclast differentiation and
activation. Nature. 2003;423(6937):337–42.
2. Takayanagi H. Osteoimmunology: shared mechanisms and crosstalk
between the immune and bone systems. Nat Rev Immunol. 2007;7(4):292–304.
3. Teitelbaum SL, Ross FP. Genetic regulation of osteoclast develop-
ment and function. Nat Rev Genet. 2003;4(8):638–49.
4. Seeman E. Reduced bone formation and increased bone resorption:
rational targets for the treatment of osteoporosis. Osteoporos Int.
2003;14(Suppl 3):S2–8.
5. Zhang Z, Xiao B, Chen Q, Lian XY. Synthesis and biological evaluationof caffeic acid 3,4-dihydroxyphenethyl ester. J Nat Prod. 2010;73(2):
252–4.
6. Cappuzzo KA, Delafuente JC. Teriparatide for severe osteoporosis.
Ann Pharmacother. 2004;38(2):294–302.
1306 WU ET AL. Journal of Bone and Mineral Research

7. Pazianas M, Abrahamsen B. Safety of bisphosphonates. Bone.2011;49(1):103–10.
8. Qiu SX, Dan C, Ding LS, Peng S, Chen SN, Farnsworth NR, Nolta J,
Gross ML, Zhou P. A triterpene glycoside from black cohosh that
inhibits osteoclastogenesis by modulating RANKL and TNFalphasignaling pathways. Chem Biol. 2007;14(7):860–9.
9. D’Amelio P, Grimaldi A, Di Bella S, Brianza SZ, Cristofaro MA, Tamone
C, Giribaldi G, Ulliers D, Pescarmona GP, Isaia G. Estrogen deficiencyincreases osteoclastogenesis up-regulating T cells activity: a key
mechanism in osteoporosis. Bone. 2008;43(1):92–100.
10. Cenci S, Toraldo G, Weitzmann MN, Roggia C, Gao Y, Qian WP,
Sierra O, Pacifici R. Estrogen deficiency induces bone loss by increas-ing T cell proliferation and lifespan through IFN-gamma-induced
class II transactivator. Proc Natl Acad Sci U S A. 2003;100(18):
10405–10.
11. Negishi-Koga T, Takayanagi H. Ca2þ-NFATc1 signaling is an essentialaxis of osteoclast differentiation. Immunol Rev. 2009;231(1):
241–56.
12. Takahashi N, Udagawa N, Tanaka S, Suda T. Generating murine
osteoclasts from bone marrow. Methods Mol Med. 2003;80:129–44.
13. Matsumoto M, Sudo T, Saito T, Osada H, Tsujimoto M. Involvement of
p38 mitogen-activated protein kinase signaling pathway in osteo-clastogenesis mediated by receptor activator of NF-kappa B ligand
(RANKL). J Biol Chem. 2000;275(40):31155–61.
14. Hotokezaka H, Sakai E, Kanaoka K, Saito K, Matsuo K, Kitaura H,
Yoshida N, Nakayama K. U0126 and PD98059, specific inhibitors ofMEK, accelerate differentiation of RAW264.7 cells into osteoclast-like
cells. J Biol Chem. 2002;277(49):47366–72.
15. Wong BR, Besser D, Kim N, Arron JR, Vologodskaia M, Hanafusa H,
Choi Y. TRANCE, a TNF family member, activates Akt/PKB througha signaling complex involving TRAF6 and c-Src. Mol Cell. 1999;4(6):
1041–9.
16. Lee SE, Woo KM, Kim SY, Kim HM, Kwack K, Lee ZH, Kim HH. Thephosphatidylinositol 3-kinase, p38, and extracellular signal-regulated
kinase pathways are involved in osteoclast differentiation. Bone.
2002;30(1):71–7.
17. Peng Q, Malhotra S, Torchia JA, Kerr WG, Coggeshall KM,Humphrey MB. TREM2- and DAP12-dependent activation of PI3K
requires DAP10 and is inhibited by SHIP1. Sci Signal. 2010;3(122):
ra38.
18. Boudot C, Saidak Z, Boulanouar AK, Petit L, Gouilleux F, Massy Z,Brazier M, Mentaverri R, Kamel S. Implication of the calcium sensing
receptor and the phosphoinositide 3-kinase/Akt pathway in the
extracellular calcium-mediated migration of RAW 264.7 osteoclast
precursor cells. Bone. 2010;46(5):1416–23.
19. Kuroda Y, Hisatsune C, Nakamura T, Matsuo K, Mikoshiba K.
Osteoblasts induce Ca2þ oscillation-independent NFATc1 activation
during osteoclastogenesis. Proc Natl Acad Sci U S A. 2008;105(25):8643–8.
20. Asagiri M, Sato K, Usami T, Ochi S, Nishina H, Yoshida H, Morita I,
Wagner EF, Mak TW, Serfling E, Takayanagi H. Autoamplification of
NFATc1 expression determines its essential role in bone homeostasis.J Exp Med. 2005;202(9):1261–9.
21. Takeshita S, Kaji K, Kudo A. Identification and characterization of the
new osteoclast progenitor with macrophage phenotypes being able
to differentiate into mature osteoclasts. J Bone Miner Res. 2000;15(8):1477–88.
22. El-Mousallamy AM, Hawas UW, Hussein SA. Teucrol, a decarboxyr-
osmarinic acid and its 40-O-triglycoside, teucroside from Teucrium
pilosum. Phytochemistry. 2000;55(8):927–31.
23. Dong A, Fang Y, Zhang L, Xie J, Wu X, Lian X, Chen Y, Luo J, Liu M.
Caffeic acid 3,4-dihydroxy-phenethyl ester induces cancer cell senes-
cence by suppressing twist expression. J Pharmacol Exp Ther. 2011;
339(1):238–47.
24. Jung JE, Kim HS, Lee CS, Park DH, Kim YN, Lee MJ, Lee JW, Park JW,Kim MS, Ye SK, Chung MH. Caffeic acid and its synthetic derivative
CADPE suppress tumor angiogenesis by blocking STAT3-mediated
VEGF expression in human renal carcinoma cells. Carcinogenesis.
2007;28(8):1780–7.
25. Won C, Lee CS, Lee JK, Kim TJ, Lee KH, Yang YM, Kim YN, Ye SK, Chung
MH. CADPE suppresses cyclin D1 expression in hepatocellular carci-
noma by blocking IL-6-induced STAT3 activation. Anticancer Res.2010;30(2):481–8.
26. Han H, Du B, Pan X, Liu J, Zhao Q, Lian X, QianM, LiuM. CADPE Inhibits
PMA-stimulated gastric carcinoma cell invasion and matrix metallo-
proteinase-9 expression by FAK/MEK/ERK-mediated AP-1 activation.Mol Cancer Res. 2010;8(11):1477–88.
27. Mesa-Siverio D, Machin RP, Estevez-Braun A, Ravelo AG, Lock O.
Structure and estrogenic activity of new lignans from Iryanthera
lancifolia. Bioorg Med Chem. 2008;16(6):3387–94.
28. Li C, Yang Z, Li Z, Ma Y, Zhang L, Zheng C, Qiu W, Wu X, Wang X, Li H,
Tang J, Qian M, Li D, Wang P, Luo J, Liu M. Maslinic acid suppresses
osteoclastogenesis and prevents ovariectomy-induced bone loss by
regulating RANKL-mediated NF-kappaB and MAPK signaling path-ways. J Bone Miner Res. 2011;26(3):644–56.
29. Koide M, Kinugawa S, Ninomiya T, Mizoguchi T, Yamashita T, Maeda
K, Yasuda H, Kobayashi Y, Nakamura H, Takahashi N, Udagawa N.Diphenylhydantoin inhibits osteoclast differentiation and function
through suppression of NFATc1 signaling. J Bone Miner Res. 2009;
24(8):1469–80.
30. Luo J, Zhou W, Zhou X, Li D, Weng J, Yi Z, Cho SG, Li C, Yi T, Wu X, LiXY, de Crombrugghe B, Hook M, Liu M. Regulation of bone formation
and remodeling by G-protein-coupled receptor 48. Development.
2009;136(16):2747–56.
31. Yang Z, Li C, Wang X, Zhai C, Yi Z, Wang L, Liu B, Du B, Wu H, Guo X, LiuM, Li D, Luo J. Dauricine induces apoptosis, inhibits proliferation and
invasion through inhibiting NF-kappaB signaling pathway in colon
cancer cells. J Cell Physiol. 2010;225(1):266–75.
32. Ikeda F, Nishimura R, Matsubara T, Tanaka S, Inoue J, Reddy SV, Hata
K, Yamashita K, Hiraga T, Watanabe T, Kukita T, Yoshioka K, Rao A,
Yoneda T. Critical roles of c-Jun signaling in regulation of NFAT
family and RANKL-regulated osteoclast differentiation. J Clin Invest.2004;114(4):475–84.
33. Li C, Yang Z, Zhai C, QiuW, Li D, Yi Z, Wang L, Tang J, QianM, Luo J, Liu
M. Maslinic acid potentiates the anti-tumor activity of tumor necrosis
factor alpha by inhibiting NF-kappaB signaling pathway. Mol Cancer.2010;9:73.
34. Wilson SR, Peters C, Saftig P, Bromme D. Cathepsin K activity-
dependent regulation of osteoclast actin ring formation and bone
resorption. J Biol Chem. 2009;284(4):2584–92.
35. McMichael BK, Meyer SM, Lee BS. c-Src-mediated phosphorylation of
thyroid hormone receptor-interacting protein 6 (TRIP6) promotes
osteoclast sealing zone formation. J Biol Chem. 2010;285(34):26641–51.
36. Sun L, Vukicevic S, Baliram R, Yang G, Sendak R, McPherson J, Zhu LL,
Iqbal J, Latif R, Natrajan A, Arabi A, Yamoah K, Moonga BS, Gabet Y,
Davies TF, Bab I, Abe E, Sampath K, Zaidi M. Intermittent recombinantTSH injections prevent ovariectomy-induced bone loss. Proc Natl
Acad Sci U S A. 2008;105(11):4289–94.
37. Rosenberger SF, Finch JS, Gupta A, Bowden GT. Extracellular signal-
regulated kinase 1/2-mediated phosphorylation of JunD and FosB isrequired for okadaic acid-induced activator protein 1 activation. J Biol
Chem. 1999;274(2):1124–30.
38. Iotsova V, Caamano J, Loy J, Yang Y, Lewin A, Bravo R. Osteopetrosis
in mice lacking NF-kappaB1 and NF-kappaB2. Nat Med. 1997;3(11):1285–9.
39. Leibbrandt A, Penninger JM. RANK/RANKL: regulators of immune
responses and bone physiology. Ann N Y Acad Sci. 2008;1143:
123–50.
Journal of Bone and Mineral Research CADPE INHIBITS RANKL-INDUCED OSTEOCLASTOGENESIS 1307

40. Zou W, Kitaura H, Reeve J, Long F, Tybulewicz VL, Shattil SJ, GinsbergMH, Ross FP, Teitelbaum SL. Syk, c-Src, the alphavbeta3 integrin, and
ITAM immunoreceptors, in concert, regulate osteoclastic bone
resorption. J Cell Biol. 2007;176(6):877–88.
41. Munugalavadla V, Vemula S, Sims EC, Krishnan S, Chen S, Yan J, Li H,
Niziolek PJ, Takemoto C, Robling AG, Yang FC, Kapur R. The p85alphasubunit of class IA phosphatidylinositol 3-kinase regulates the
expression of multiple genes involved in osteoclast maturationand migration. Mol Cell Biol. 2008;28(23):7182–98.
42. Krum SA, Brown M. Unraveling estrogen action in osteoporosis. Cell
Cycle. 2008;7(10):1348–52.
43. Jin H, Dai J, Chen X, Liu J, Zhong D, Gu Y, Zheng J. Pulmonary toxicity
and metabolic activation of dauricine in CD-1 mice. J Pharmacol ExpTher. 2010;332(3):738–46.
1308 WU ET AL. Journal of Bone and Mineral Research
Related Documents







![Methyl trans-([plus-minus sign])-1-oxo-2-phenethyl-3 ... · Methyl trans-( )-1-oxo-2-phenethyl-3-(thiophen-2-yl)-1,2,3,4-tetrahydro-isoquinoline-4-carboxylate Mehmet Akkurt,a* Selvi](https://static.cupdf.com/doc/110x72/5f8f753362af594ccf1f8cae/methyl-trans-plus-minus-sign-1-oxo-2-phenethyl-3-methyl-trans-1-oxo-2-phenethyl-3-thiophen-2-yl-1234-tetrahydro-isoquinoline-4-carboxylate.jpg)



