ORIGINAL ARTICLE Cadmium-induced biochemical responses of Vallisneria spiralis Ragini Singh & R. D. Tripathi & Sanjay Dwivedi & Munna Singh & P. K. Trivedi & D. Chakrabarty Received: 24 December 2009 / Accepted: 7 April 2010 / Published online: 6 May 2010 # Springer-Verlag 2010 Abstract The following study was carried out to investigate the cadmium (Cd) accumulating potential of Vallisneria. After subjecting plants to different concentrations of Cd, it was observed that plants are able to accumulate ample amount of metal in their roots (5,542 μgg −1 dw) and leaves (4,368 μg g −1 dw) in a concentration- and duration- dependent manner. Thus, it is evident that the accumulation in roots was 1.3 times higher than the shoots. It was also noted that with increasing Cd accumulation, roots of the plant appeared darker in color and harder in texture. In response to metal exposure, amount of low molecular weight antioxidants such as cysteine and nonprotein thiols (NP-SH) and activity of enzymes such as APX and GPX were significantly enhanced at lower concentrations of Cd, followed by decline at higher doses. It was also observed that in exposed plants, activity of APX enzyme was higher in roots (ca. 3 times) as compared to leaves. However, chlorophyll and protein content was found to decline significantly in a dose-dependent manner. Results suggested that due to its high accumulation potential, Vallisneria may be effectively grown in water bodies moderately contami- nated with Cd. Keywords Accumulation . Antioxidant . Cadmium . Cysteine . Vallisneria spiralis Introduction In the course of industrialization, levels of toxic metals have risen tremendously and significantly exceeded those from natural sources for practically all metals. Due to mobilization of metals into the biosphere, their circulation through soil, water, and air has increased manifolds (Singh et al. 2006; Mishra et al. 2009). Among the heavy metals, cadmium (Cd) is one which is a nonessential, toxic metal that enters aquatic environment from natural (weathering) as well as anthropogenic sources (industrial effluents and agricultural runoff; Singh et al. 2006; Wang et al. 2009). It causes easily identifiable toxicity symptoms in plants at very low concentration. Submerged aquatic plants take up Cd by both adsorption and energy-dependent transport systems (Tripathi et al. 1996) and incorporate them into their own system disturbing various metabolic processes. Besides, this metal also forms a complex with the natural ligands which affects its bioaccumulation and toxicity potential. Cd is reported as a human carcinogen. Its accumulation in plants causes various symptoms such as stunted growth, chlorosis, leaf epinasty, disturbs uptake of nutrients, inhibits photosynthesis, inactivates enzymes in CO 2 fixation, induces lipid peroxidation and, also inhibits pollen germi- nation and tube growth (Mishra et al. 2006, Wang et al. 2009). Cd is a nonredox-active metal, however, it induces reactive oxygen species (ROS) which has to be kept under tight control because their accumulation result in cell death due to oxidative processes such as membrane lipid peroxidation, protein oxidation, enzyme inhibition, and Handling Editor: Bumi Nath R. Singh : R. D. Tripathi (*) : S. Dwivedi : P. K. Trivedi : D. Chakrabarty Ecotoxicology and Bioremediation Group, National Botanical Research Institute (Council of Scientific and Industrial Research), Rana Pratap Marg, Lucknow 226 001, India e-mail: [email protected] M. Singh Department of Botany, University of Lucknow, Lucknow 226 001, India Protoplasma (2010) 245:97–103 DOI 10.1007/s00709-010-0146-4

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL ARTICLE
Cadmium-induced biochemical responsesof Vallisneria spiralis
Ragini Singh & R. D. Tripathi & Sanjay Dwivedi &Munna Singh & P. K. Trivedi & D. Chakrabarty
Received: 24 December 2009 /Accepted: 7 April 2010 /Published online: 6 May 2010# Springer-Verlag 2010
Abstract The following study was carried out to investigatethe cadmium (Cd) accumulating potential of Vallisneria.After subjecting plants to different concentrations of Cd, itwas observed that plants are able to accumulate ampleamount of metal in their roots (5,542 μg g−1 dw) and leaves(4,368 μg g−1 dw) in a concentration- and duration-dependent manner. Thus, it is evident that the accumulationin roots was 1.3 times higher than the shoots. It was alsonoted that with increasing Cd accumulation, roots of theplant appeared darker in color and harder in texture. Inresponse to metal exposure, amount of low molecular weightantioxidants such as cysteine and nonprotein thiols (NP-SH)and activity of enzymes such as APX and GPX weresignificantly enhanced at lower concentrations of Cd,followed by decline at higher doses. It was also observedthat in exposed plants, activity of APX enzyme was higher inroots (ca. 3 times) as compared to leaves. However,chlorophyll and protein content was found to declinesignificantly in a dose-dependent manner. Results suggestedthat due to its high accumulation potential, Vallisneria maybe effectively grown in water bodies moderately contami-nated with Cd.
Keywords Accumulation . Antioxidant . Cadmium .
Cysteine . Vallisneria spiralis
Introduction
In the course of industrialization, levels of toxic metalshave risen tremendously and significantly exceeded thosefrom natural sources for practically all metals. Due tomobilization of metals into the biosphere, their circulationthrough soil, water, and air has increased manifolds (Singhet al. 2006; Mishra et al. 2009). Among the heavy metals,cadmium (Cd) is one which is a nonessential, toxic metalthat enters aquatic environment from natural (weathering)as well as anthropogenic sources (industrial effluents andagricultural runoff; Singh et al. 2006; Wang et al. 2009). Itcauses easily identifiable toxicity symptoms in plants atvery low concentration. Submerged aquatic plants take upCd by both adsorption and energy-dependent transportsystems (Tripathi et al. 1996) and incorporate them intotheir own system disturbing various metabolic processes.Besides, this metal also forms a complex with the naturalligands which affects its bioaccumulation and toxicitypotential.
Cd is reported as a human carcinogen. Its accumulationin plants causes various symptoms such as stunted growth,chlorosis, leaf epinasty, disturbs uptake of nutrients, inhibitsphotosynthesis, inactivates enzymes in CO2 fixation,induces lipid peroxidation and, also inhibits pollen germi-nation and tube growth (Mishra et al. 2006, Wang et al.2009). Cd is a nonredox-active metal, however, it inducesreactive oxygen species (ROS) which has to be kept undertight control because their accumulation result in cell deathdue to oxidative processes such as membrane lipidperoxidation, protein oxidation, enzyme inhibition, and
Handling Editor: Bumi Nath
R. Singh : R. D. Tripathi (*) : S. Dwivedi : P. K. Trivedi :D. ChakrabartyEcotoxicology and Bioremediation Group, National BotanicalResearch Institute (Council of Scientific and Industrial Research),Rana Pratap Marg,Lucknow 226 001, Indiae-mail: [email protected]
M. SinghDepartment of Botany, University of Lucknow,Lucknow 226 001, India
Protoplasma (2010) 245:97–103DOI 10.1007/s00709-010-0146-4

DNA and RNA damage (Ruley et al. 2004; Mishra et al.2008). To counteract these effects, plants are equipped withROS-detoxifying enzymes such as peroxidases (APX andGPX), superoxide dismutases (SOD), and catalase (CAT)for protecting potential cell injury causing tissue dysfunc-tion (Halliwell 1987; Srivastava et al. 2006).
Phytoremediation is a cleanup technology where plantsare used to degrade, extract, or immobilize contaminantsfrom soils and water (Wang et al. 2009). For effectivephytoremediation of metals, they must be translocated andaccumulated in the aerial parts of the plants. The submergedplants may be of great use for the phytoremediationstrategies of heavy metal-polluted water bodies as they donot migrate and attain equilibrium with their surroundingswithin a short period (Guilizzoni 1991) and possess betteraccumulation potential due to large exposed surface area.Therefore, in the present study Vallisneria spiralis, a rootedsubmerged plant was used to evaluate its potential foraccumulating Cd under laboratory conditions. Effect ofmetal ion on some metabolic parameters, such as chloro-phyll, protein, cysteine, and nonprotein thiols (NP-SH), aswell as the activity of enzymes like APX and GPX in theroots and leaves of treated plants, was also studied.
Materials and methods
Plant material and treatment conditions
Plants were collected from water bodies of Unnao BirdSanctuary [26.53°N 80.5°E], Unnao (Uttar Pradesh) andwere grown in large tubs in the field in natural condition forhealthy growth. For experimental purpose, small newplantlets (ca. 2–4 in. in length) were separated from themother plant, transferred to laboratory, and grown in cultureroom in 10% Hoagland solution (Hoagland and Arnon1950) under controlled conditions (16 h light usingfluorescent tube light, 114 μ mol m−2 s−1 at 23 ± 2°C) insmall plastic tub for 5 days to acclimatize in laboratoryconditions. After acclimatization, plants were treated withdifferent concentrations of Cd.
The acclimatized plants were transferred to 250 mlbeakers containing various concentrations of metals intriplicates and were harvested at 24, 48, 72, and 96 hexposures. The different metal concentrations of Cd (0.1, 1,10, 25, 50, and 100 μM) were prepared by diluting 1 mMstock solution of the metal salt CdCl2 with 10% Hoaglandsolution.
Metal accumulation
Harvested plants were thoroughly washed in distilled waterand oven-dried at 80°C till constant weight was obtained.
Dried plant material (root and leaves separately) (100 mg)was powdered and wet digested in HNO3:HClO4 (3:1, v/v)at 70°C. Digested material was diluted with milli-Q water,and metal content was determined using atomic absorptionspectrophotometer (GBC Avanta Σ, Australia).
The standard reference material of Cd (E-Merck,Germany) was used for the calibration and qualityassurance for each analytical batch. Analytical data qualityof metal was ensured with three repeated analysis (n=6) ofEPA quality control samples (lot TMA 989), and the resultswere found within (±2.820) the certified values. Recoveryof Cd from the plant tissue was found to be more than98.5% as determined by spiking samples with a knownamount of metal. The blanks were run in triplicate to checkthe precision of the method with each set of samples. Thedetection limit of Cd was found to be 0.1 μg l−1.
Plant growth parameters
For the estimation of photosynthetic pigments, plantmaterial (300 mg) was ground in chilled 80% acetone indark. After centrifugation at 10,000 g for 10 min at 4°C,absorbance of the supernatant was taken at 480, 510, 645,and 663 nm. The content of chlorophylls was estimated bythe method of Arnon (1949) and that of carotenoid by usingthe formula given by Duxbury and Yentsch (1956). Proteincontent was estimated following the method of Lowry et al.(1951).
Estimation of nonenzymatic antioxidants
Fresh plant material (500 mg) was homogenized in 5%chilled perchloric acid and centrifuged at 10,000 g for10 min at 4°C. Cysteine content was measured insupernatant using acid-ninhydrin reagent and absorbancewas recorded at 560 nm (Gaitonde 1967).
NP-SH content was measured following the method ofEllman (1959). Fresh plant material (700 mg) washomogenized in 6.67% 5'-sulfosalicylic acid. After centri-fugation at 10,000 g for 10 min at 4°C, NP-SH content wasmeasured in the supernatant by reaction with Ellmanreagent, and absorbance was recorded at 412 nm.
Estimation of antioxidant enzymes
Plant material (500 mg) was homogenized in 100 mMpotassium phosphate buffer (pH 7.0) containing 0.1 mMEDTA and 1% polyvinylpyrrolidone (w/v) at 4°C. Homog-enate was filtered through four layers of cheese cloth andcentrifuged at 15,000 g for 15 min at 4°C. Supernatant wasused to measure the activities of enzymes.
98 R. Singh et al.

The activity of APX was measured according to themethod of Nakano and Asada (1981) by estimating the rate ofascorbate oxidation (extinction coefficient 2.8 mM−1 cm−1).The 3 ml reaction mixture contained 50 mM phosphate buffer(pH 7.0), 0.1 mM H2O2, 0.5 mM sodium ascorbate, 0.1 mMEDTA, and a suitable aliquot of enzyme extract. Thechange in absorbance was monitored at 290 nm, andenzyme activity was expressed as micromoles of ascorbateoxidized min−1 g−1 fw.
GPX activity was assayed according to the method ofHemeda and Klein (1990). A 100 ml of reaction mixturewas prepared by adding 10 ml of 1% guaiacol (v/v), 10 mlof 0.3% H2O2, and 80 ml of 50 mM phosphate buffer(pH 6.6). Enzyme extract (75 μl) was added to reactionmixture in a final volume of 3 ml. The increase inabsorbance due to oxidation of guaiacol (extinction coeffi-cient 26.6 mM−1 cm−1) was monitored at 470 nm. Enzymeactivity was expressed as micromoles of guaiacol oxidizedmin−1 g−1 fw.
Statistical analysis
The experiment was done following a randomized blockdesign. Two-way analysis of variance was done with all thedata to confirm the variability of data and validity ofresults, and Duncan’s multiple range test was performed todetermine the significant difference between treatments.Correlation analysis was performed which has been givenwithin text at relevant places (p<0.001***; p< 0.01**;p < 0.1*; NS non significant; Gomez and Gomez 1984).
Results
Results showed that treated plants of Vallisneria accumu-lated very high amounts of Cd in both roots and leaveswhich increased in a dose-dependent manner but accumu-lation rate decreased with the increasing duration. Accu-mulation was found more in the roots (56%; Table 1) thanleaves (44%; Table 1). Of the total accumulation 46.5%,72%, and 88% in roots and about 47%, 66%, and 85.5% inleaves were accumulated after 24 h, 48 h, and 72 h,respectively. Maximum amount of accumulated metal was5,542 μg g−1 dw (root) and 4,368 μg g−1 dw (leaf) at thehighest exposure dose. Thus, it is evident that theaccumulation in roots was 1.3 times higher than the shoots.
Contents of both total chlorophyll and carotenoidsdecreased in a dose-dependent manner. Maximum decreaseof 72.5% (R=−0.941***) in total chlorophyll (Fig. 1a) and67% (R=−0.928***) in carotenoid (Fig. 1b) was observedat highest exposure dose of Cd in the leaves of treatedplants.
A significant induction in protein content was observedin leaves at all the exposure durations up to 1 μM Cd,followed by a significant decline at higher doses; whereasin roots, there was no change in the protein content up to24 h, but with increasing duration, a significant inductionwas observed up to 1 μM till 96 h, followed by a gradualdecline with increasing Cd concentrations. Maximuminduction of 62% (leaf) and 67% (root) at1 μM for 72 hand maximum reduction of 47% (R=−0.837**; leaf) and59% (R=−0.817**; root) at highest treatment dose were
Cd content (μgg−1dw)
Exposure period (h)
24h 48h 72h 96h
Root Cd concentration (μM)
0 ND ND ND ND
0.1 55.5±16f 66±20f 134±19f 208±20e
1 162.5±22e 255±66e 304.5±75e 464±121d
10 668±232d 786±155d 859±166d 917.5±215c
25 1,289±199c 1,491±213c 1,571.5±178c 1,658.5±195b
50 1,594±185b 2,280±265b 2,569.5±265b 4,656±411a
100 2,575±251a 3,978±333a 4,890±425a 5,542±550a
Leaf Cd concentration (μM)
0 ND ND ND ND
0.1 20±10f 42.5±16f 69.5±20f 94.5±25f
1 69±20e 145±19e 156±20e 420±100e
10 166.5±23d 381±85d 461.5±112d 770±145d
25 572.5±182c 621±223d 1,003±225c 1,332±223c
50 1,157.5±197b 1,356.5±213b 1,964.5±150b 3,089±296b
100 2,039±259a 2,978.5±225a 3,731±323a 4,368±400a
Table 1 Accumulation ofcadmium in roots and leaves ofVallisneria spiralis exposed todifferent concentration andexposure periods
All the values are mean oftriplicates ± SD. ANOVAsignificant at p≤0.01. Differentletters indicate significantlydifferent values at a particularduration (DMRT, p≤0.05)
Cadmium-induced biochemical responses of Vallisneria spiralis 99
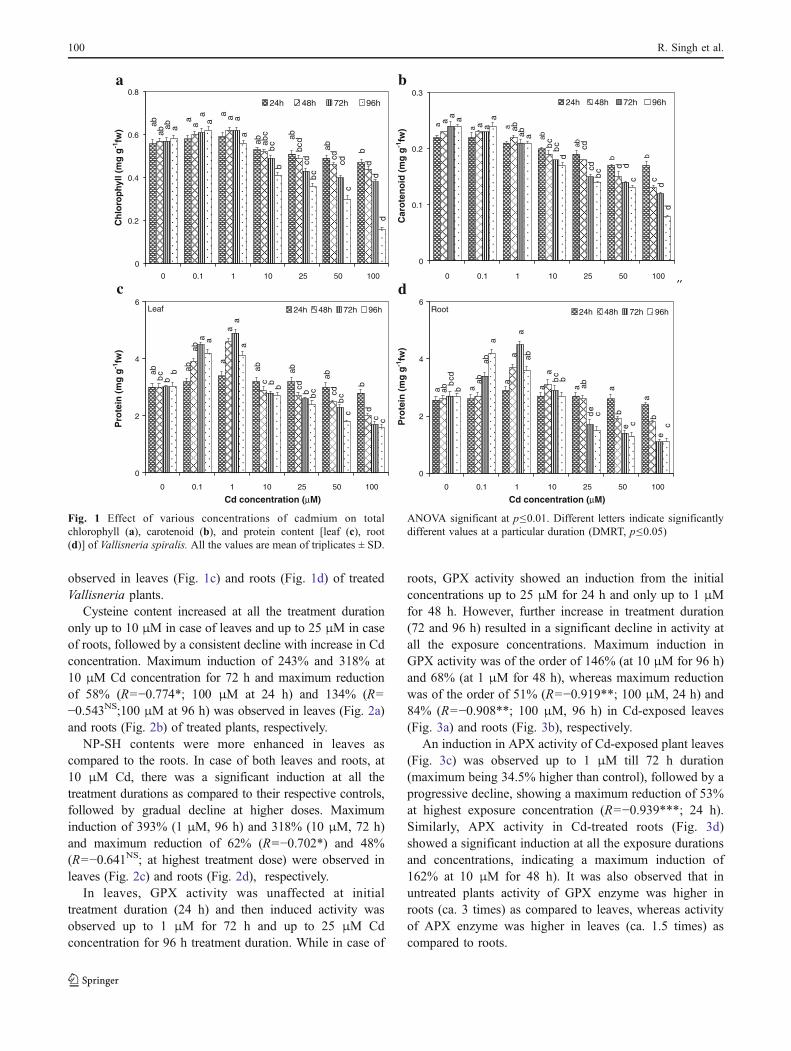
observed in leaves (Fig. 1c) and roots (Fig. 1d) of treatedVallisneria plants.
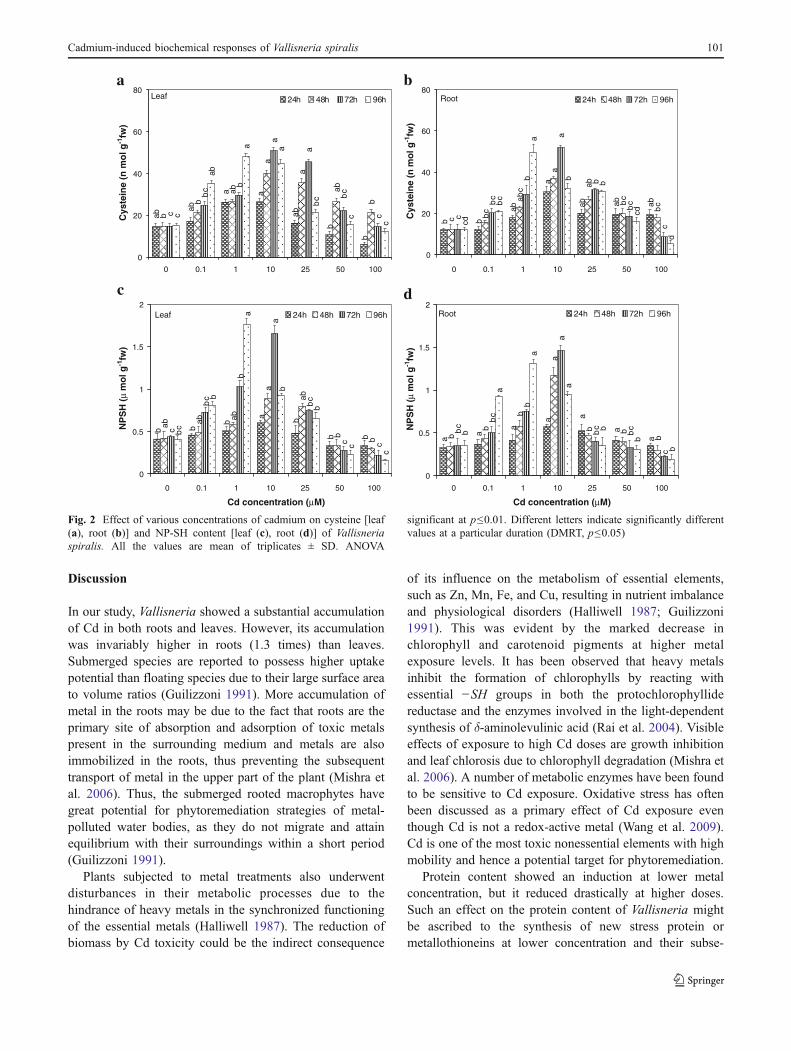
Cysteine content increased at all the treatment durationonly up to 10 μM in case of leaves and up to 25 μM in caseof roots, followed by a consistent decline with increase in Cdconcentration. Maximum induction of 243% and 318% at10 μM Cd concentration for 72 h and maximum reductionof 58% (R=−0.774*; 100 μM at 24 h) and 134% (R=−0.543NS;100 μM at 96 h) was observed in leaves (Fig. 2a)and roots (Fig. 2b) of treated plants, respectively.
NP-SH contents were more enhanced in leaves ascompared to the roots. In case of both leaves and roots, at10 μM Cd, there was a significant induction at all thetreatment durations as compared to their respective controls,followed by gradual decline at higher doses. Maximuminduction of 393% (1 μM, 96 h) and 318% (10 μM, 72 h)and maximum reduction of 62% (R=−0.702*) and 48%(R=−0.641NS; at highest treatment dose) were observed inleaves (Fig. 2c) and roots (Fig. 2d), respectively.
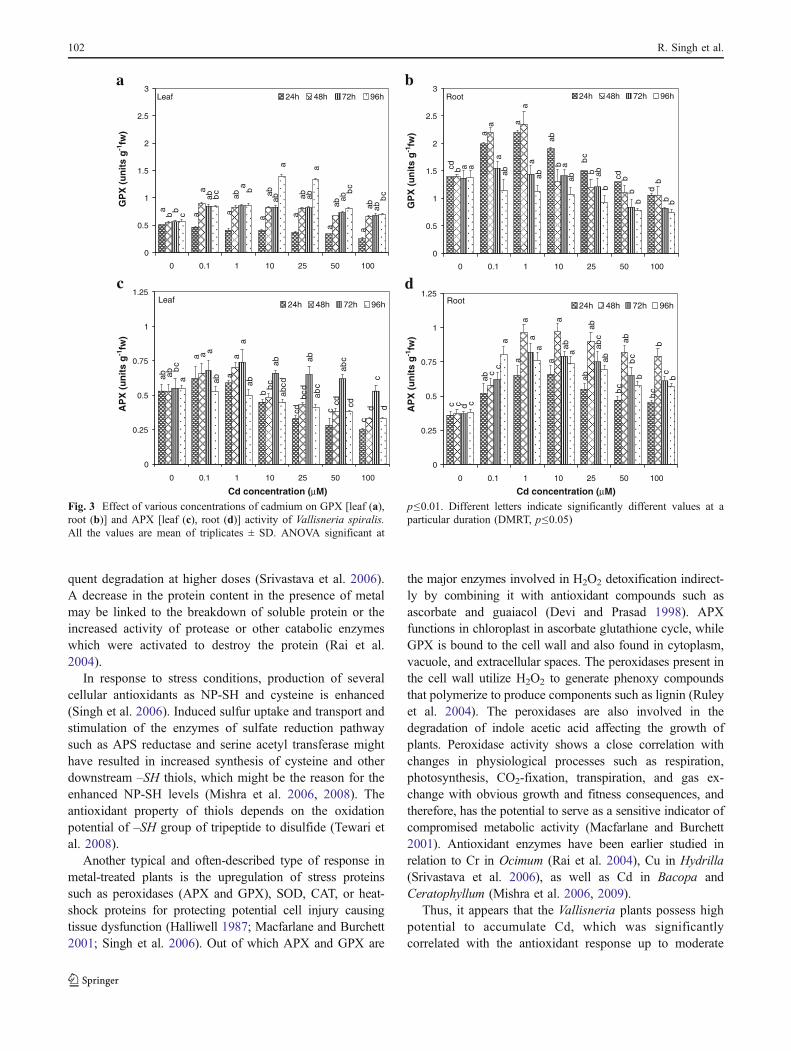
In leaves, GPX activity was unaffected at initialtreatment duration (24 h) and then induced activity wasobserved up to 1 μM for 72 h and up to 25 μM Cdconcentration for 96 h treatment duration. While in case of
roots, GPX activity showed an induction from the initialconcentrations up to 25 μM for 24 h and only up to 1 μMfor 48 h. However, further increase in treatment duration(72 and 96 h) resulted in a significant decline in activity atall the exposure concentrations. Maximum induction inGPX activity was of the order of 146% (at 10 μM for 96 h)and 68% (at 1 μM for 48 h), whereas maximum reductionwas of the order of 51% (R=−0.919**; 100 μM, 24 h) and84% (R=−0.908**; 100 μM, 96 h) in Cd-exposed leaves(Fig. 3a) and roots (Fig. 3b), respectively.
An induction in APX activity of Cd-exposed plant leaves(Fig. 3c) was observed up to 1 μM till 72 h duration(maximum being 34.5% higher than control), followed by aprogressive decline, showing a maximum reduction of 53%at highest exposure concentration (R=−0.939***; 24 h).Similarly, APX activity in Cd-treated roots (Fig. 3d)showed a significant induction at all the exposure durationsand concentrations, indicating a maximum induction of162% at 10 μM for 48 h). It was also observed that inuntreated plants activity of GPX enzyme was higher inroots (ca. 3 times) as compared to leaves, whereas activityof APX enzyme was higher in leaves (ca. 1.5 times) ascompared to roots.
ab a
a
ab ab
ab b
ab
a
a
abc
bcd
cd
d
ab
a a
bc
cd cd
dd
c
bc
b
a
a
a
0
0.2
0.4
0.6
0.8
0 0.1 1 10 25 50 100
Ch
loro
phy
ll (m
g g
-1fw
)
24h 48h 72h 96h
bb
ab
ab
a a a
c
d
cdbc
a a ab
d
dcd
bc
ab
a
a
d
cbc
d
a
a a
0
0.1
0.2
0.3
0 0.1 1 10 25 50 100
Car
ote
no
id (
mg
g-1
fw)
24h 48h 72h 96h
b
ab
ababab ab
a
d
cd
cd
cbc
ab
a
c
bcb
b
a
b
a
c
c
bcb
a
b
a
0
2
4
6
0 0.1 1 10 25 50 100
Cd concentration (µM)
Pro
tein
(m
g g
-1fw
)
24h 48h 72h 96hLeaf
a
aaaa a
a
b
b
ab
a
ab
ab
a
e
e
de
bc
a
bcd
ab
cc
c
b
ab
b
a
0
2
4
6
0 0.1 1 10 25 50 100
Cd concentration (µM)
Pro
tein
(m
g g
-1fw
)
24h 48h 72h 96hRoot
a b
c d }
Fig. 1 Effect of various concentrations of cadmium on totalchlorophyll (a), carotenoid (b), and protein content [leaf (c), root(d)] of Vallisneria spiralis. All the values are mean of triplicates ± SD.
ANOVA significant at p≤0.01. Different letters indicate significantlydifferent values at a particular duration (DMRT, p≤0.05)
100 R. Singh et al.
Discussion
In our study, Vallisneria showed a substantial accumulationof Cd in both roots and leaves. However, its accumulationwas invariably higher in roots (1.3 times) than leaves.Submerged species are reported to possess higher uptakepotential than floating species due to their large surface areato volume ratios (Guilizzoni 1991). More accumulation ofmetal in the roots may be due to the fact that roots are theprimary site of absorption and adsorption of toxic metalspresent in the surrounding medium and metals are alsoimmobilized in the roots, thus preventing the subsequenttransport of metal in the upper part of the plant (Mishra etal. 2006). Thus, the submerged rooted macrophytes havegreat potential for phytoremediation strategies of metal-polluted water bodies, as they do not migrate and attainequilibrium with their surroundings within a short period(Guilizzoni 1991).
Plants subjected to metal treatments also underwentdisturbances in their metabolic processes due to thehindrance of heavy metals in the synchronized functioningof the essential metals (Halliwell 1987). The reduction ofbiomass by Cd toxicity could be the indirect consequence
of its influence on the metabolism of essential elements,such as Zn, Mn, Fe, and Cu, resulting in nutrient imbalanceand physiological disorders (Halliwell 1987; Guilizzoni1991). This was evident by the marked decrease inchlorophyll and carotenoid pigments at higher metalexposure levels. It has been observed that heavy metalsinhibit the formation of chlorophylls by reacting withessential −SH groups in both the protochlorophyllidereductase and the enzymes involved in the light-dependentsynthesis of δ-aminolevulinic acid (Rai et al. 2004). Visibleeffects of exposure to high Cd doses are growth inhibitionand leaf chlorosis due to chlorophyll degradation (Mishra etal. 2006). A number of metabolic enzymes have been foundto be sensitive to Cd exposure. Oxidative stress has oftenbeen discussed as a primary effect of Cd exposure eventhough Cd is not a redox-active metal (Wang et al. 2009).Cd is one of the most toxic nonessential elements with highmobility and hence a potential target for phytoremediation.
Protein content showed an induction at lower metalconcentration, but it reduced drastically at higher doses.Such an effect on the protein content of Vallisneria mightbe ascribed to the synthesis of new stress protein ormetallothioneins at lower concentration and their subse-
b
b
ab
a
ab
ab
a
b
ab
a
a
b
b
ab
c
bc
a
a
b
c
bc
c
c
bc
aa
c
ab
0
20
40
60
80
0 0.1 1 10 25 50 100
Cys
tein
e (n
mo
l g-1
fw)
24h 48h 72h 96hLeaf
ab
bb
a
ab ab ab
abc
bc
c
a
ab
bc
bcb
c
c
b
a
b
bc
c
bc
cd
a
b
b
cd
d
0
20
40
60
80
0 0.1 1 10 25 50 100
Cys
tein
e (n
mo
l g-1
fw)
24h 48h 72h 96hRoot
bb
ba
b b
b
bb
ab
a
ab ab ab
cc
bc
a
b
c
bc
c
c
b
b
a
bc
b
0
0.5
1
1.5
2
0 0.1 1 10 25 50 100
Cd concentration (µM)
NP
SH
(µ
mo
l g-1
fw)
24h 48h 72h 96hLeaf
a
a
a
a a
a
a
b
b
b
a
b b
b
bc
bc
b
a
bc
bc
c
a
b
a
a
b
b
b
0
0.5
1
1.5
2
0 0.1 1 10 25 50 100
Cd concentration (µM)
NP
SH
(µ
mo
l g-1
fw)
24h 48h 72h 96hRoot
a b
c d
Fig. 2 Effect of various concentrations of cadmium on cysteine [leaf(a), root (b)] and NP-SH content [leaf (c), root (d)] of Vallisneriaspiralis. All the values are mean of triplicates ± SD. ANOVA
significant at p≤0.01. Different letters indicate significantly differentvalues at a particular duration (DMRT, p≤0.05)
Cadmium-induced biochemical responses of Vallisneria spiralis 101
quent degradation at higher doses (Srivastava et al. 2006).A decrease in the protein content in the presence of metalmay be linked to the breakdown of soluble protein or theincreased activity of protease or other catabolic enzymeswhich were activated to destroy the protein (Rai et al.2004).
In response to stress conditions, production of severalcellular antioxidants as NP-SH and cysteine is enhanced(Singh et al. 2006). Induced sulfur uptake and transport andstimulation of the enzymes of sulfate reduction pathwaysuch as APS reductase and serine acetyl transferase mighthave resulted in increased synthesis of cysteine and otherdownstream –SH thiols, which might be the reason for theenhanced NP-SH levels (Mishra et al. 2006, 2008). Theantioxidant property of thiols depends on the oxidationpotential of –SH group of tripeptide to disulfide (Tewari etal. 2008).
Another typical and often-described type of response inmetal-treated plants is the upregulation of stress proteinssuch as peroxidases (APX and GPX), SOD, CAT, or heat-shock proteins for protecting potential cell injury causingtissue dysfunction (Halliwell 1987; Macfarlane and Burchett2001; Singh et al. 2006). Out of which APX and GPX are
the major enzymes involved in H2O2 detoxification indirect-ly by combining it with antioxidant compounds such asascorbate and guaiacol (Devi and Prasad 1998). APXfunctions in chloroplast in ascorbate glutathione cycle, whileGPX is bound to the cell wall and also found in cytoplasm,vacuole, and extracellular spaces. The peroxidases present inthe cell wall utilize H2O2 to generate phenoxy compoundsthat polymerize to produce components such as lignin (Ruleyet al. 2004). The peroxidases are also involved in thedegradation of indole acetic acid affecting the growth ofplants. Peroxidase activity shows a close correlation withchanges in physiological processes such as respiration,photosynthesis, CO2-fixation, transpiration, and gas ex-change with obvious growth and fitness consequences, andtherefore, has the potential to serve as a sensitive indicator ofcompromised metabolic activity (Macfarlane and Burchett2001). Antioxidant enzymes have been earlier studied inrelation to Cr in Ocimum (Rai et al. 2004), Cu in Hydrilla(Srivastava et al. 2006), as well as Cd in Bacopa andCeratophyllum (Mishra et al. 2006, 2009).
Thus, it appears that the Vallisneria plants possess highpotential to accumulate Cd, which was significantlycorrelated with the antioxidant response up to moderate
aa
aa
aa
a abab
ababab
a
b
ab
ababab
a
ab
b
bcb
c
a
bc
c
b
a
0
0.5
1
1.5
2
2.5
3
0 0.1 1 10 25 50 100
GP
X (
un
its
g-1
fw)
24h 48h 72h 96hLeaf
d
cd
bc
ab
a
a
cd
bb
b
b
a
a
b
b
b
ab
aaa
a
bb
ab
ab
a
ab
b
0
0.5
1
1.5
2
2.5
3
0 0.1 1 10 25 50 100
GP
X (
un
its
g-1
fw)
24h 48h 72h 96hRoot
c
ccd
b
a
a
ab
d
cd
bcdb
c
aa
ab c
abcabab
a
a
bc
dcd
abcdaba ab
abc
0
0.25
0.5
0.75
1
1.25
0 0.1 1 10 25 50 100
Cd concentration (µM)
AP
X (
un
its
g-1
fw)
24h 48h 72h 96hLeaf
bcbc
ab
aa
ab
c
bab
ab
aa
c
c
c
bc
abc
ab
a
c
d
bb
a
a
c
a
ab
0
0.25
0.5
0.75
1
1.25
0 0.1 1 10 25 50 100
Cd concentration (µM)
AP
X (
un
its
g-1
fw)
24h 48h 72h 96hRoot
a b
c d
Fig. 3 Effect of various concentrations of cadmium on GPX [leaf (a),root (b)] and APX [leaf (c), root (d)] activity of Vallisneria spiralis.All the values are mean of triplicates ± SD. ANOVA significant at
p≤0.01. Different letters indicate significantly different values at aparticular duration (DMRT, p≤0.05)
102 R. Singh et al.

exposure concentrations. However, at higher doses, thedefense responses became insufficient to cope up withincreasing metal toxicity leading to suppressed growth. Itmay be inferred that Cd is efficiently tolerated byVallisneria plants through activation of antioxidant systemand thiolic pathways and thus, the plant may be useful indecreasing the metal load of moderately contaminatedwater bodies.
Acknowledgments The authors are thankful to Director, NationalBotanical Research Institute, Lucknow for the facilities provided.Ragini Singh is grateful to Council of Scientific and IndustrialResearch (CSIR), New Delhi, India for the award of Senior ResearchFellowship (SRF).
Conflict of interest The authors declare that they have no conflict ofinterest.
References
Arnon DI (1949) Copper enzyme in isolated chloroplast polyphenoloxidase in Beta vulgaris. Plant Physiol 24:1–15
Devi SR, Prasad MNV (1998) Copper toxicity in Ceratophyllumdemersum L. (Coontail), a free floating macrophyte: responseof antioxidant enzymes and antioxidants. Plant Sci 138:157–165
Duxbury AC, Yentsch CS (1956) Plantkton pigment monograph. JMar Res 15:93–101
Ellman GL (1959) Tissue sulphydryl groups. Arch Biochem Biophys82:70–77
Gaitonde MK (1967) A spectrophotometric method for the directdetermination of cysteine in the presence of other naturallyoccurring amino acids. Biochem J 104:627–633
Gomez KA, Gomez AA (1984) Statistical Procedures for AgriculturalResearch. John Wiley, New York
Guilizzoni P (1991) The role of heavy metal and toxic materials inphysiological ecology of submerged macrophytes. Aquat Bot87:87–109
Halliwell B (1987) Oxidative damage, lipid peroxidation and antioxidantprotection in chloroplasts. Chem Phys Lipids 44:327–340
Hemeda HM, Klein BP (1990) Effects of naturally occurringantioxidants on peroxidase activity of vegetable extracts. J FoodSci 55:184–185
Hoagland DR, Arnon DI (1950) The water-culture method for growingplants without soil. California Agri Exp Station Circ 347:1–32
Lowry OH, Roenbrough NJ, Farr AL, Randal EJ (1951) Proteinmeasurement with the folin phenol reagent. J Biol Chem193:265–275
Macfarlane GR, Burchett MD (2001) Photosynthetic pigments andperoxidase activity as indicators of heavy metal stress in the greymangrove, Avicennia marina (Forsk.) Vierh. Mar Pollut Bull42:233–240
Mishra S, Srivastava S, Tripathi RD, Govindarajan R, Kuriakose SV,Prasad MNV (2006) Phytochelatin synthesis and response ofantioxidants during cadmium stress in Bacopa monnieri L. PlantPhysiol Biochem 44:25–37
Mishra S, Srivastava S, Tripathi RD, Dwivedi S, Shukla MK (2008)Responses of antioxidant enzymes in coontail (Ceratophyllumdemursum L.) plants under cadmium stress. Environ Toxicol23:294–301
Mishra S, Tripathi RD, Srivastava S, Dwivedi S, Trivedi PK,Dhankher OP, Khare A (2009) Thiol metabolism play significantrole during cadmium detoxification by Ceratophyllum demersumL. Bio Technol 100:2155–2161
Nakano Y, Asada K (1981) Hydrogen peroxide is scavenged byascorbate-specific peroxidase in spinach chloroplasts. Plant CellPhysiol 22:867–880
Rai V, Vajpayee P, Singh SN, Mehrotra S (2004) Effect of chromiumaccumulation on photosynthetic pigments, oxidative stressdefense system, nitrate reduction, proline level and eugenolcontent of Ocimum tenuiflorum L. Plant Sci 167:1159–1169
Ruley AT, Sharma NC, Sahi SV (2004) Antioxidant defense in a leadaccumulating plant, Sesbania drummondii. Plant Physiol Biochem42:899–906
Singh S, Eapen S, D’Souza SF (2006) Cadmium accumulation and itsinfluence on lipid peroxidation and antioxidative system in anaquatic plant, Bacopa monnieri L. Chemo 62:233–246
Srivastava S, Mishra S, Tripathi RD, Dwivedi S, Gupta DK (2006)Copper-induced oxidative stress and responses of antioxidantsand phytochelatins in Hydrilla verticillata (L.f.) Royle. AquaToxicol 80:405–415
Tewari A, Singh R, Singh NK, Rai UN (2008) Amelioration ofmunicipal sludge by Pistia stratiotes L.: role of antioxidantenzymes in detoxification of metals. Bio Technol 99:8715–8721
Tripathi RD, Rai UN, Gupta M, Chandra P (1996) Induction ofphytochelatins in Hydrilla verticillata (l.f.) Royle under cadmiumstress. Bull Environ Contam Toxicol 56:505–512
Wang C, Sun Q, Wang L (2009) Cadmium toxicity and phytochelatinproduction in a rooted-submerged macrophyte Vallisneria spiralisexposed to low concentrations of cadmium. Environ Toxicol24:271–278
Cadmium-induced biochemical responses of Vallisneria spiralis 103
Related Documents











