Accepted Preprint This article is protected by copyright. All rights reserved Environmental Toxicology CADMIUM ACCUMULATION AND TOXICITY IN THE UNICELLULAR ALGA PSEUDOKIRCHNERIELLA SUBCAPITATA : INFLUENCE OF METAL-BINDING EXUDATES AND EXPOSURE TIME NATHALIE PAQUET, MICHEL LAVOIE , FRÉDÉRIC MALONEY , JÉRÔME F.L. DUVAL , P ETER G.C. CAMPBELL , and C LAUDE FORTIN Environ Toxicol Chem. , Accepted Article • DOI: 10.1002/etc.2927 Accepted Article "Accepted Articles" are peer-reviewed, accepted manuscripts that have not been edited, formatted, or in any way altered by the authors since acceptance. They are citable by the Digital Object Identifier (DOI). After the manusc ript is edited and formatted, it will be removed from the “Accepted Articles” Web site and published as an Early View article. Note that editing may introduce changes to the manuscript text and/or graphics which could affect content, and all legal disclaimers and ethical guidelines that apply to the journal pertain. SETAC cannot be held responsible for errors or consequences arising from the use of information contained in these manuscripts.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
Environmental Toxicology
CADMIUM ACCUMULATION AND TOXICITY IN THE UNICELLULAR ALGA
PSEUDOKIRCHNERIELLA SUBCAPITATA: INFLUENCE OF METAL-BINDING
EXUDATES AND EXPOSURE TIME
NATHALIE PAQUET, MICHEL LAVOIE, FRÉDÉRIC MALONEY, JÉRÔME F.L. DUVAL, PETER G.C.
CAMPBELL, and CLAUDE FORTIN
Environ Toxicol Chem., Accepted Article • DOI: 10.1002/etc.2927
Accepted Article "Accepted Articles" are peer-reviewed, accepted manuscripts that have not been edited, formatted, or in any way altered by the authors since acceptance. They are citable by the Digital Object Identifier
(DOI). After the manuscript is edited and formatted, it will be removed from the “Accepted Articles” Web site and published as an Early View article. Note that editing may introduce changes to the manuscript text and/or graphics which could affect content, and all legal disclaimers and ethical
guidelines that apply to the journal pertain. SETAC cannot be held responsible for errors or consequences arising from the use of information contained in these manuscripts.

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
Environmental Toxicology Environmental Toxicology and Chemistry DOI 10.1002/etc.2927
CADMIUM ACCUMULATION AND TOXICITY IN THE UNICELLULAR ALGA
PSEUDOKIRCHNERIELLA SUBCAPITATA: INFLUENCE OF METAL-BINDING EXUDATES AND EXPOSURE TIME
Running title: Evolution of cadmium accumulation and toxicity over time
NATHALIE PAQUET, †‡ MICHEL LAVOIE, †§ FRÉDÉRIC MALONEY, †|| JÉRÔME F.L. DUVAL, #
PETER G.C. CAMPBELL, † and CLAUDE FORTIN†
† Institut national de la recherche scientifique, Centre Eau Terre Environnement (INRS-ETE), Québec, Québec, Canada
‡ Centre d’expertise en analyse environnementale du Québec, Ministère du développement
durable, de l’Environnement et de la Lutte contre les changements climatiques, Québec, Québec,
Canada
§ Québec-Océan and Takuvik Joint Université Laval/CNRS Research Units, Département de Biologie, Université Laval, Québec, Québec, Canada
|| Administration portuaire de Québec, Québec, Québec, Canada
# Laboratoire interdisciplinaire des Environnements Continentaux (LIEC), CNRS-Université de Lorraine, Vandoeuvre- lès-Nancy cedex, France
*Address correspondence to [email protected]
Additional Supporting Information may be found in the online version of this article.
This article is protected by copyright. All rights reserved
Submitted 14 October 2014; Returned for Revision 31 January 2015; Accepted 2 February 2015

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
Abstract: Predicting metal availability and toxicity for chronic (several hours or days) metal
exposure scenarios, even for unicellular algae, is a major challenge to existing toxicity models.
This is because several factors affecting metal uptake/toxicity, such as the release of metal-
binding exudates, changes in the kinetics of metal uptake/toxicity over time and algal
physiological acclimation to internalized metals, are still poorly understood. The present study
assessed the influence of these factors on Cd uptake and toxicity in laboratory batch cultures of
the freshwater alga, Pseudokircheneriella subcapitata. To do so, changes in the free Cd2+
concentrations due to the release of metal-binding algal exudates were monitored, 109Cd
accumulation in algal cells was measured and Cd-induced inhibition of algal growth as a
function of exposure time (from 12 to 96 h) was followed. Our results indicate that metal-binding
exudates may decrease the proportion of the free Cd2+ ion in solution up to twofold, a decrease
that affect Cd uptake and toxicity. Pseudokirchneriella subcapitata has the capacity to decrease
net Cd uptake rate on short-time scales (< 24 h), but this reduction in the Cd uptake rate
disappeared after 24 h and Cd toxicity occurred at relatively high Cd concentrations in solution.
These data illustrate some of the pitfalls of standard algal toxicity assays, which were designed
for short-term exposures, and suggest how robust chronic bioassays might be developed.
This article is protected by copyright. All rights reserved
Keywords: Phytoplankton, Ecological risk assessment, Metal uptake, Toxicity testing, Green
alga

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
INTRODUCTION
The biotic ligand model (BLM) has been shown to accurately predict short-term uptake
(< 1 h) of cationic metals and their acute toxicity in algae, although some exceptions exist [1, 2].
However, predicting metal accumulation and toxicity in algae under chronic metal exposure
conditions (several hours or days) is still challenging and cannot be easily achieved [3-6]. In
chronic metal exposure, poorly understood complicating factors need to be taken into account in
order to model metal uptake and toxicity. Such factors include growth biodilution of internalized
metal [7], acclimation of the targeted organisms to the presence of the toxic metal [8], metal
efflux [9], possible depletion of metals from bulk solution due to the necessarily finite size of the
metal-supplying medium [10, 11], and production of metal-binding exudates [12]. The latter
situation can be particularly problematic in weakly buffered culture media without strong ligands
(e.g. NTA, EDTA) in which metal-binding exudates can significantly affect metal speciation,
uptake and toxicity [13-16]. Toxicity assays, which often need to be performed on natural waters
without addition of strong metal chelators in order to evaluate metal toxicity in situ [17, 18],
might then be biased by the release of metal-binding exudates by the algae.
Until now, studies about the effect of metal-binding exudates on metal uptake and
toxicity have focused mainly on marine algae [13-15, 19] with fewer reports dealing with
freshwater algal species [16, 20, 21]. In many of these studies, the free metal ion concentration
was not measured and the effect of metal-binding exudates on metal speciation thus remains
poorly documented, particularly in freshwater algal cultures. Other complications in chronic
exposures such as changes in the kinetics of metal uptake in algae and the time scale over which
Cd exerts its toxicity have not received much attention in the literature. Therefore, improvement
in our fundamental knowledge of metal uptake/toxicity in algae over chronic exposure periods is

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
critically needed to refine current chronic toxicity models as well as to evaluate potential
weaknesses in toxicity assay procedures.
In the present study, the freshwater alga Pseudokirchneriella subcapitata was used as the
test organism. This species is one of the most commonly used bioassay algae around the world
[17, 18, 22, 23] and is widely distributed across nearly all types of inland waters [24, 25]. The
objectives of this research were to: 1) evaluate the effect of metal-binding exudates produced by
the freshwater alga P. subcapitata on Cd speciation; 2) evaluate Cd accumulation kinetics in P.
subcapitata in chronic (96 h) exposures to Cd and; 3) investigate the link between Cd speciation,
Cd accumulation and toxicity in P. subcapitata as a function of exposure time.
MATERIALS AND METHODS
Test organisms and culture conditions
Pure agar cultures of the unicellular green alga Pseudokirchneriella subcapitata
(Korshikov) Hindak (CPCC 37) were purchased from the University of Toronto Culture
Collection (UTCC), now known as the Canadian Phycological Culture Center (CPCC).
Asynchronous batch cultures were grown axenically (periodically checked by culture plating) in
100 mL of CEAEQ (Centre d’expertise en analyse environnemental du Québec) culture medium
with the 3-(N-morpholino)-propanesulfonic acid (MOPS) pH buffer but without any metal
chelating agent (Table 1). Stock algae cultures were maintained in 250 mL Erlenmeyer glass
flasks under constant rotary agitation (60 rpm), illumination (100 μmol m-2 s-1) and temperature
(20 ± 1 °C). A 2-mL sub-sample of algal culture was transferred to a sterile culture medium
every week using a sterile serological pipette. Culture media (without trace metals) as well as
material (Erlenmeyer flasks and filtration apparatus) destined for algal cultures were autoclaved
at 121 °C for 15 min. Culture media were then supplemented with a filter-sterilized trace metal

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
mix (0.2 µm polycarbonate membrane, 47 mm). All plasticware and glassware were soaked for
at least 24 h in 10% (v/v) HNO3 and rinsed seven times with ultrapure water (18 MΩ cm;
Nanopure grade) and then dried under a class 100 laminar flow hood where all manipulations
that required precautions against possible contamination by airborne particulates were
performed.
Fe oxyhydroxides and trace metal speciation in the CEAEQ medium
Since the CEAEQ medium did not contain a trace metal buffer to stabilize iron in
solution, the predicted amount of trace metals (Cu, Co, Zn, Cd) adsorbed on amorphous
ferrihydrite (Feox) was calculated with the two- layer surface complexation model [26]. The
MINEQL+ chemical speciation programme version 4.6 [27] with updated formation constants for
aqueous inorganic complexes and intrinsic surface complexation constants for cations on Feox
[28] was used. Even when it is assumed that virtually all the added Fe precipitated in the
medium, the modeled adsorption of essential (Co, Zn, Cu) and non-essential (Cd) metal ions on
Feox was quantitatively unimportant (<10 %; except for Cu, for which up to 50% of Cu may be
adsorbed to Fe colloids). Moreover, at the highest [Cd] used in the present study, MINEQL+
simulations show that competition between Cd and Cu remains low, such that Cu adsorption on
Feox decreases by only 17%. Our calculations thus suggest that Fe colloids will not appreciably
affect Cd accumulation and toxicity in our experiments at constant [Cu2+].
Preparation of exposure media
A stock of acidified 109Cd radioisotope solution (3.45 mCi mg-1 Cd, Amersham) was
prepared in fluorinated ethylene propylene copolymer (Teflon®) flasks. The exposure media
consisting of CEAEQ medium spiked with 109Cd were left to equilibrate 24 h after addition of
109Cd and pH adjustment with small additions of NaOH or 0.1 M HCl. In all exposure media, the

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
Cd addition step (targeted initial total Cd concentrations from 8 to 270 nM) did not increase the
ionic strength significantly (i.e., it remained constant at 7.5 meq L-1). Each exposure
concentration was tested in triplicate in 70 mL of CEAEQ medium using polycarbonate 250 mL
Erlenmeyer flasks. The pH was buffered at 7.0 ± 0.1 by the addition of MOPS (10 mM; the pH
fluctuated by less than 0.08 units after the 96-hour exposure). Cadmium speciation in the
exposure solutions was calculated with the chemical speciation model MINEQL+ (version 4.6;
[29]) with updated equilibrium constants [28], using total measured metal concentrations (at t = 0
or 96 h) as input data. All calculations were constrained to a fixed input pH (7.0) at equilibrium
with the atmosphere (pCO2 = 10-3.5 atm).. MINEQL+ simulations indicated that around 98% of
the total initial Cd in the CEAEQ exposure solutions is present as free Cd2+ ions.
Measurements of cadmium accumulation, cell density and cell size
Exponentially growing cells from the stock cultures were gently harvested on 2 µm
polycarbonate filters (Poretics, Fisher Scientific) using a vacuum pressure of <13 kPa. Harvested
cells were rinsed five times with CEAEQ-R medium (simplified culture medium; Table 1) and
resuspended in 20 mL of CEAEQ medium. Cell diameter, surface and volume (assuming
spherical cell shape) were determined in the algal concentrate resuspension with a Multisizer 3
Coulter Counter (70-µm aperture, Beckman). Afterwards, algal cells were inoculated into Cd
exposure media under a laminar flow hood. We decreased the initial cell density from 10,000 to
2,500 cells mL-1 to potentially minimize metal depletion over time and mimic more closely the
cell densities generally encountered in oligotrophic freshwater ecosystems [21]. Immediately
after inoculation, algal cultures were placed in the environmental chamber. Subsamples of the
algal cultures inoculated at an initial cell density of 2,500 cells mL-1 were taken after 0, 12, 24,
48, 72 and 96 h in order to measure cell sizes with the Coulter Counter and cell densities with a

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
FacsCalibur flow cytometer (see details of measurement using flow cytometry in the next
section). Algal cultures inoculated at an initial cell density of 10,000 cells mL-1 were only
subsampled at 96 h.
At each time step where cell density and size were analyzed, 10-mL aliquots of the algal
cultures were gently filtered through two superimposed 2-µm polycarbonate membranes. The
filtrates were recovered to analyse the dissolved cadmium concentration remaining in the culture
medium. Afterwards, the filters were soaked for 10 min with 5 mL of 2 × 10-4 M EDTA solution
(in simplified CEAEQ-R medium); the EDTA solutions were then gently filtered and another 5
mL of EDTA was rapidly passed through the filters. The EDTA rinse removes the 109Cd bound
to the surface of the algal cells and the Cd remaining on the filter membrane is thus operationally
defined as intracellular [30, 31]. Filter radioactivity (intracellular algal Cd content) was measured
with a scintillation counter (Wallac 1414; Perkin Elmer Life Sciences) by placing the filters in
borosilicate vials containing 20 mL of Ecolume scintillation cocktail (ICN). The activity of the
lower filter was subtracted from that measured on the upper filter to correct for passive retention
of 109Cd by the polycarbonate filters. 109Cadmium activities in the filtrates of exposure media
(dissolved Cd in the exposure media) or after the EDTA rinsing step (cadmium adsorbed on the
algae) were also measured by scintillation counting. Mass balance calculations indicate that
losses of Cd on the filtration apparatus, pipette tips and due to handling of filters were 11 ± (SD)
6%.
Ion-exchange technique
Free [Cd2+] in initial exposure media (before inoculation of algal cells) as well as in
filtered samples from algal cultures at the end of the exposure (96 h; 0.2 µm, polycarbonate
filters) was measured using an ion-exchange technique (IET) [32] in algal cultures inoculated at

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
2,500 cells mL-1. For the cultures inoculated at 10,000 cells mL-1, the free Cd2+ concentrations
were measured at the end of the exposure but not at the beginning. Briefly, the technique consists
of bringing a calibrated ion-exchange resin to equilibrium with the test solution. The metal
bound to the resin is then eluted and the original free metal ion concentration is back calculated
from a previous metal- resin calibration (See the SI.1 section of Supplemental Data,for details
about the IET technique).
Flow cytometry
Algal cell density was analyzed using a FACSCalibur (BD BioSciences) flow cytometer.
Algae subsamples (1 mL) were first vortexed for 5 minutes and then injected into the flow
cytometer. The algal cells were then hydrodynamically focused into a fluid stream so that the
cells passed individually through the light source. The pigments of the algal cells were excited by
a laser emitted by a 488 nm xenon- ion excitation lamp and the autofluorescence of the algae was
detected as red fluorescence (660-700 nm) by a FL3 fluorescence detector equipped with
fluorescence emission filters. Nonalgal particles and dead cells were excluded from the analysis
by setting an acquisition threshold on FL3 (positioned below the distribution of healthy control
cells). All parameters were collected as logarithmic signals and analyses were performed at a
high flow rate (60 µL/min), with acquisition over 120 s. The stability of the rate of aspiration
was measured after each 10 samples using TRUCount tubes (BioScience, # 340334).
Concentrations of algal cells (Cpop in cell/mL) were determined using the following equation 6
[33]:

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
where “occurrence number” represents the number of fluorescence signals detected, “Vtot” is the
total sample volume comprising the fixing solution (mL), “Valgae” is the volume of culture
medium add in the cytometer (mL), “Raspiration” is the aspiration rate of the flow cytometer
(mL/min ) and “Treading” is the time for analysing one sample (min).
Growth rate calculation
Cell densities of algal cultures exposed to various total [Cd] and monitored at each time
points by flow cytometry were multiplied by the mean biovolume per cell (µm3 cell-1) measured
with the Coulter counter yielding total cell volume concentrations per mL (µm3 of alga mL-1).
The total cell volume concentrations per mL were then used to calculate the specific growth rate
(µ) of algal cultures exposed to various total [Cd] with the following equation (7):
where t0 and t represent the beginning and end of the experiment and Nt and Nt0 are total cell
volume concentrations (µm3 of alga mL-1) at time t and t0 respectively.
Cadmium toxicity
The toxicity of Cd was evaluated by measuring the culture yield (cell number or total cell
volume) at each exposure time step (12 to 96 h). The culture yield was then plotted as a function
of the total dissolved [Cd] (initial and mean exposure concentration), free final measured [Cd2+],
intracellular Cd concentrations or Cd cell quotas. The data were fitted with the macro REGTOX
(http://www.normalesup.org/~vindimian/en_index.html) to calculate the EC50 with the Hill
approach. The 95% confidence intervals (95%CI) around the mean EC50 values were estimated
by a bootstrap non parametric simulation.
Statistics

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
Preliminary conditions of parametric statistical tests were evaluated and, if these
conditions were violated, parametric tests were performed on Box-Cox transformed data.
Statistical analyses were performed using Systat software (Version 10.0) and results were
considered to be significant if p < 0.05. Unless otherwise indicated, errors are given as ± one
standard deviation. Differences among effective Cd concentrations inhibiting algal growth by
50%, as determined with the cell yield or the cell volume yield, were conservatively evaluated by
comparing the 95%CI of the EC50.
RESULTS
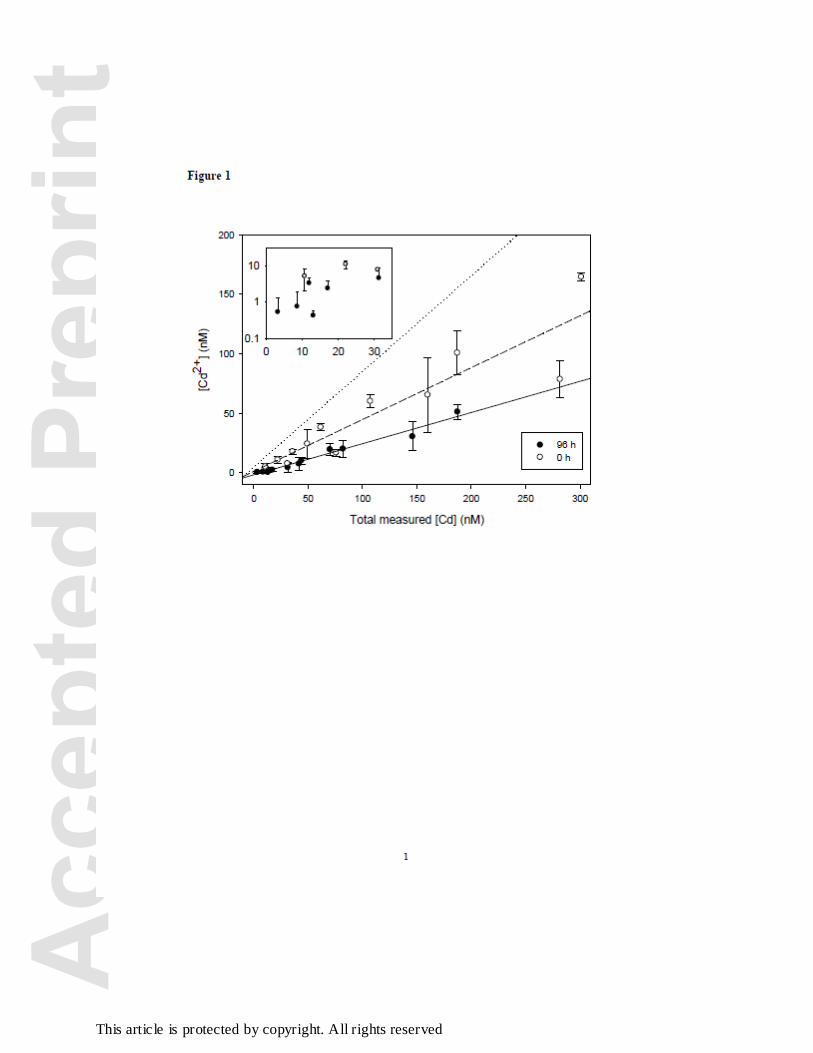
Cadmium speciation: complexation by metal-binding exudates
The slope of the theoretical regression representing the expected initial Cd2+
concentrations in solution modeled with MINEQL+ (I = 200 meq L-1, CEAEQ medium, ionic
strength adjusted with NaNO3) as a function of the total initial Cd concentration was of 0.83
(dotted line, Figure 1). At mean measured total initial [Cd] < 62 nM, the slope of the regression
of the free measured [Cd2+] as a function of total measured initial [Cd] was close to the dotted
theoretical line (slope = 0.64 ± (SE) 0.11, regression not shown) as expected. However, when
including all the measured Cd concentrations at t=0, the slope of the regression of the free
measured [Cd2+] as a function of the measured total [Cd] was of 0.44 ± (SE) 0.06 (Figure 1, open
circles, regression not shown), which is almost two times less than the expected slope based on
MINEQL+ calculations. At t = 96 h, the slope of the regression of measured Cd2+ as a function
of total Cd (including all the Cd concentrations) further decreased to 0.26 ± (SE) 0.01 (Figure 1,
close circles, regression not shown). At total measured [Cd] < 20 nM, the mean measured free
[Cd2+] was up to an order of magnitude lower after the 96 h exposure compared to the initial
exposure conditions, although this difference was not statistically significant because of the high

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
variability in the measured free [Cd2+] at t = 96 h. However, for total measured [Cd] > 40 nM,
the mean measured free [Cd2+] decreased significantly by about 50% at the end of the exposure.
Total dissolved cadmium concentrations
Over the first 12 h, the total dissolved [Cd] in solution showed an apparent decrease of 15
to 40% compared to the initial total [Cd] (t = 0), for which there was only one replicate (n = 1).
In the interval between 12 and 96 h, the total dissolved [Cd] decreased further but by no more
than 20%. For the experiments performed at an initial cell density of 2,500 cells mL-1, we thus
decided to average all measured total dissolved [Cd] between 12 and 96 h (denoted “mean
exposure total [Cd]”) when studying the relationship between Cd accumulation or toxicity over
time and the total [Cd] in solution. For the 96-h Cd exposure experiment performed at an initial
cell density of 2,500 cells mL-1 and 10,000 cells mL-1; the total dissolved [Cd] decreased by 54 ±
(SD) 9% and 18 ± (SD) 3% respectively.
Cadmium accumulation as a function of time and dissolved [Cd]
Intracellular cadmium concentration in P. subcapitata cells exposed to relatively low
mean exposure total [Cd] (< 50 nM) remained stable (p > 0.05) after 24 h of exposure (see insert
in Figure 2A). However, at relatively high mean exposure total [Cd] (> 56 nM), intracellular Cd
concentrations diverged appreciably as the exposure time was extended. Generally, Cd
accumulation increased as mean exposure total [Cd] increased, except for t = 12 h, where
intracellular Cd concentrations were surprisingly constant across a wide range of mean exposure
total [Cd] (Figure 2B). This constancy in intracellular Cd concentrations was confirmed
statistically since the slope of the regression relating the intracellular Cd concentration to the
mean exposure total [Cd] between 89 and 202 nM was not significantly different from 0 (t-test
analysis).

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
Cadmium toxicity
Algal cells inoculated at 2,500 cells mL-1 grew exponentially (µ = 0.058 ± SE 0.003 h-1)
over the 96 h exposure in the control medium (no Cd added; full circles, Figure 3). At a mean
exposure total [Cd] of 18 nM, P. subcapitata cells also grew exponentially over the 96 h
exposure, but the specific cellular growth rate decreased by approximately 26% (µ = 0.046 ± SE
0.0009 h-1) with respect to control cells. For mean exposure total [Cd] 56 nM, the specific
growth rate changed over the exposure period; the algal cells grew at the maximal rate for the
first 24 h and then continued to grow at a much lower rate (5.3 times less than the control at 56
nM mean exposure total [Cd]) from 48 to 96 h (mean µ = 0.011 ± SE 0.001 h-1; solid line in
Figure 3). Note that the algal cells inoculated at 10,000 cells mL-1 in control medium with no
added Cd grew exponentially over the 96 h exposure at a specific growth rate of 0.035 ± SE 0.1
h-1, which was significantly slower than the specific growth rate of algal cells inoculated at 2,500
mL-1.
Figure 4 and Table SI.1 show the EC50 values calculated for the cell yield (panels A to
D) or the total cellular volume yield (panels E to H) as a function of the exposure time; the EC50
values were calculated in terms of total initial [Cd], final free [Cd2+], mean exposure total [Cd],
intracellular [Cd] or Cd quotas. No effect on cell growth was observed after 12 or 24 h across the
whole range of Cd concentrations and thus no EC50s could be computed at these time steps. The
EC50s expressed as initial total [Cd], mean exposure total [Cd] or final free [Cd2+] all tended to
decrease over the exposure time, but this decrease was less clear when total cellular volume yield
was chosen as an endpoint instead of the cell number yield. When considering the cell number
yield as an endpoint, this decrease in EC50s over time was statistically significant as
conservatively evaluated on the basis of the absence of overlapping 95%CI for the EC50 values

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
at t = 48, 72 and 96 h. However, for the EC50s at 48, 72 and 96 h calculated on the basis of the
total cellular volume yield instead of the cell number yield, the 95%CI overlapped and the
statistical significance of the decrease in EC50 values over time could not be demonstrated.
For the toxicity assays performed at very low initial cell densities (2,500 cells mL-1), the
EC50 values at 96 h calculated from the effect of total initial [Cd] or Cd accumulation on cell
number yield or total cell volume yield tended to be lower than the EC50s measured at higher
initial cell density (10,000 cells mL-1). The differences in these EC50s were statistically
significant (except when considering the intracellular Cd concentrations) on the basis of the
absence of overlap between the 95%CI at high or low initial cell density. The intracellular Cd
concentrations inhibiting by 50% the total cell volume yield at 96 h of exposure for the cultures
inoculated at a low initial cell density were particularly variable (1.09 × 10-19 mol Cd µm-3;
95%CI = 6.0 × 10-20 to 6.8 × 10-19 mol Cd µm-3) and the 95%CI of this EC50 overlapped with
the 95%CI of the EC50 at high initial cell density; hence, the significance of this difference
could not be demonstrated conservatively.
DISCUSS ION
Cadmium speciation and metal-binding exudates
The free measured initial Cd2+ concentrations at total measured [Cd] < 62 nM was close
(although the data variability was high) to the modeled free Cd2+ concentrations suggesting that
no Cd complexation occurred initially in the culture medium before algal inoculation (Figure 1,
open circles). However, when taking into account all total measured [Cd] at t = 0 Figure, the
slope of the regression of the free measured Cd2+ concentration as a function of the total
measured [Cd] (Figure 1, open circles) was almost two times less than the expected slope based
on MINEQL+ calculations (Figure1, dotted line). This discrepancy could be due to the low
number of replicates (n = 1) used for the measurements of total [Cd] at t = 0 or to an unexpected

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
Cd complexation at t = 0. Although there are some uncertainties on the degree of Cd
complexation at t = 0, the data clearly show that Cd complexation was more pronounced at t = 96
h (Figure1, solid line) than at t = 0 h (Figure1, dashed line), which strongly suggests that metal-
binding exudates produced by the algae complexed a significant portion of total Cd in solution.
In a natural freshwater environment, metals such as Cd, Co and Zn are typically found in
large proportions as the free ion species [34-36] due to their relatively low affinity for dissolved
organic matter, which poorly buffers the free metal ion concentrations. Hence, one might expect
that metal-binding exudates produced by phytoplankton in oligotrophic but particularly in
eutrophic systems would sometimes affect metal speciation, as suggested by several free metal
ion measurements in ecosystems in different trophic states [37-41]. Koukal et al. [16] also
suggested that metal-binding exudates released by P. subcapitata could decrease Cd, Cu, Pb and
Zn uptake and toxicity due to metal complexation in solution and/or to an indirect effect of
organic exudates on algae. The present study affords additional information about the interaction
between metal-binding ligands of algal origin and Cd speciation. For laboratory experiments in
synthetic media, the results show that algal exudates secreted by the freshwater algae P.
subcapitata may significantly decrease the free Cd2+ concentration, particularly at Cd
concentrations > 40 nM for which Cd complexation significantly increased at t = 96 h relative to
t = 0 (Figure1). Such a decrease would be expected to affect Cd accumulation and toxicity in
algae as well as in other aquatic organisms. The results also suggest that toxicity assays
performed in natural waters without adding strong metal chelators might be biased by the release
of metal-binding exudates by the algae and subsequent Cd complexation. The results thus
demonstrate the importance of keeping cellular density as low as possible in toxicity assays using
P. subcapitata to better reflect the metal sensitivity of the cells.

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
Effect of exposure time on steady-state Cd accumulation
Contrary to acute (<1 h) metal exposure experiments, chronic algal experiments (i.e.,
several hours or a few days) bring into play several poorly understood factors such as growth
kinetics (and metal biodilution), algae acclimation to varying metal concentrations, release of
metal-binding exudates, and metal efflux. All of these physiological responses of algae to metals
may potentially affect metal accumulation and toxicity in phytoplankton. When algae are
acclimated to the exposure medium, a steady-state can be attained or approximated in which
metal uptake is balanced by metal loss. In the present study, chronic Cd accumulation in P.
subcapitata cells exposed to relatively low [Cd] (i.e. mean exposure [Cd] < 60 nM) tended to
reach a steady-state in Cd accumulation after an exposure time of 24 h (Figure 2A). In batch
cultures of P. subcapitata where depletion in dissolved [Cd] (and free Cd2+) is negligible and
specific growth rate is constant, Cd uptake is expected to be balanced only by growth biodilution
[7, 8, 42] since Cd efflux in P. subcapitata was shown to be negligible in an earlier study [42].
The specific growth rate of P. subcapitata cells exposed to a low mean exposure total [Cd] of 18
nM was constant over the 96-hour exposure (Figure 4). However, at higher mean exposure total
[Cd] (56 nM and above), specific algal growth rates decreased after 24 h of exposure (Figure 3).
Such a decrease in growth rate after 24 h suggests that Cd slowly exerts its toxic effect within the
algal cells (see the last section of the discussion about Cd toxicity for details). This delay in the
expression of Cd toxicity on growth rate will tend to postpone reaching a steady-state in
intracellular Cd contents. Figure 2A effectively demonstrates that at the mean exposure Cd
concentrations of 56 nM and 89 nM (i.e. highly toxic concentrations, Figure 4), intracellular Cd
concentrations level off only after 72 h whereas at mean exposure Cd concentrations of 108 nM
or greater, intracellular Cd concentrations did not reach a steady-state over the 96-h exposure.

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
Note that an increase in Cd complexation by algal exudates over time (Figure 1) will tend to
decrease Cd uptake and should also affect whether or not a steady-state in Cd accumulation may
be achieved over the 96-h Cd exposure experiment.
Cadmium uptake as a function of mean exposure total [Cd] and exposure time
Cadmium accumulation as a function of the mean measured exposure total [Cd] shows an
upward curvature as toxic Cd concentrations are reached (Figure 2B). Positive Cd feedback on
Cd accumulation (i.e., Cd uptake inhibits growth rate, which decreases growth biodilution) is a
plausible hypothesis that could explain the non-linear relationship between Cd accumulation and
[Cd] observed in Figure 2. If true, one might expect that the extent of this possible positive
feedback on Cd accumulation, as well as the sensitivity of Cd uptake to variations in [Cd2+], will
depend on the number, affinity and capacity of Cd transport system(s) involved.
Another interesting result is the capacity of P. subcapitata to maintain intracellular Cd
concentrations at nearly stable levels after an exposure time of 12 h across a wide range of mean
exposure total [Cd] (from 89 to 202 nM; Figure 2, full circles). These results suggest that algal
cells can reduce net Cd uptake rates on a short-time scale. Such a response of P. subcapitata was
also reported by Errécalde and Campbell [42], who demonstrated that Cd uptake in the presence
of high [Cd2+] (280 and 600 nM) induced a decrease in Cd uptake rates after an exposure of just
10 min.
Cadmium toxicity (EC50) as a function of the initial cell density
Standard toxicity assays such as those performed by the CEAEQ, Environment Canada
and the USEPA measure metal toxicity using 96 h exposures of P. subcapitata cells grown in
batch cultures with initial algal inoculums of 10,000 cells mL-1 [17, 18, 43]. In the present study,
Cd toxicity based on cell yields (expressed as total initial Cd) using the standardized initial cell

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
density used by several environmental agency (10,000 cells mL-1) was around three times higher
(EC50 three times lower) when a low initial cell density (2,500 cells mL-1) was used. The present
study thus demonstrates that the sensitivity of standardized toxicity assays may be improved by
decreasing the initial algal density. Higher algal sensitivity to total initial Cd as the initial cell
density decreased cannot be explained by lower metal depletion at lower initial cell density
because metal depletion at t = 96 h increased, not decreased, as the initial cell density decreased,
i.e. 54 ± (SD) 9% and 18 ± (SD) 3% of total dissolved Cd was lost at 2,500 cells mL-1 and
10,000 cells mL-1 respectively. Higher metal depletion observed at lower initial cell density
might be explained by inter-experimental differences in specific growth rates. For instance,
control algal cells inoculated at 10,000 cells mL-1 grew more slowly than control algal cells
inoculated at 2,500 cells mL-1, and this variation in growth rate, presumably due to slight
changes in the physiological state of algal cells, may lead to large changes in the amount of Cd
taken up by the cells (and depleted from the culture medium) after a long-term Cd exposure of
four days. Instead, we propose that the higher reported algal sensitivity to total initial Cd as the
initial cell density decreased (Figure 4) might be at least partly explained by a lower release of
metal-binding exudates (explained by the four times lower initial cell density) and, thus a higher
proportion of free Cd2+ at a given total [Cd]. Such effects of the initial cell density on metal-
binding exudate production, metal complexation and toxicity have also been suggested by
Franklin et al. [21] to explain the inverse relationship that they observed between initial cell
density (from 103 cells mL-1 to 105 cells mL-1) and Cu toxicity in P. subcapitata. However, the
effect of initial cell density on Cd toxicity was not a simple function of the final measured Cd2+
concentration since the EC50 values (expressed in terms of the final measured Cd2+
concentration) calculated on the basis of total cell volume and cell number yield were

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
approximately 20 and 15 times higher when the cultures were inoculated at 10,000 cells mL-1
than at 2,500 cells mL-1. We hypothesize that variations in algal physiological states (related to
differences in the algal density and/or inter-experimental variability) could disproportionately
affect the sensitivity of P. subcapitata to the free Cd2+ ions. However, other experiments are
needed to confirm or infirm the above hypothesis.
Cadmium toxicity (EC50) as a function of time and endpoint
The present study showed that EC50 values calculated on the basis of total cell volume
yields of P. subcapitata and expressed in terms of initial total [Cd] or mean exposure total [Cd]
tended to decrease (toxicity increased) with time. For exposure periods of 48, 72 and 96 h, the
decrease in EC50 with time was mainly due to an increasing divergence between cellular volume
yields of control and Cd-exposed cells as the exposure time increased. Such a result occurred
with a constant growth rate over time (at mean exposure total [Cd] ≤ 18 nM; Figure 3) or with a
decreasing growth rate over time (at mean exposure total [Cd] 56 nM; Figure 3). This
increasing trend in Cd toxicity over time was more pronounced when the cell number yield was
used as the endpoint rather than the total cell volume yield. The difference in the decreasing
trend in the EC50s over time between both endpoints can be explained by the increase in
biovolume per algal cell at toxic Cd concentrations; this increase in biovolume per cell
compensated in part for the decrease in cell yields. For similar reasons, toxicity expressed in
terms of total Cd or final free Cd2+ also were consistently greater when the cell number yield was
used as the endpoint. Cellular volume yield as an endpoint for Cd2+ toxicity was also shown to be
less sensitive than the cell number yield in another recent study investigating Cd toxicity in a
different green algal species, Chlamydomonas reinhardtii [44].

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
Cadmium effects on cell number or total cell volume yield (Figure 4) as well as on
growth rate (Figure 3) remained undetectable after exposure times of 12 to 24 h, even in the very
highly contaminated cultures (i.e., for high mean exposure total [Cd] reaching up to 202 nM and
leading to high intracellular Cd concentrations; Figure 2). These results strongly suggest that Cd
exerts its toxicity slowly within the cells, leading to little growth inhibition over the first 24 h of
exposure. Small inhibitions of algal growth rate (e.g. by < 20%) will not lead to a large decrease
in cell yield after 12 or 24 h; such small decreases in cell yield will remain difficult to measure
accurately. For the two above reasons (i.e., toxicity delay and difficulty of measuring growth
yield inhibition after 12 or 24 h in the presence of a relatively small growth rate inhibition),
EC5024 hours values based on growth rates or cell yields should not be considered as reliable
toxicity endpoints in Cd toxicity assays using P. subcapitata.
CONCLUSION
Cadmium accumulation and toxicity in the freshwater alga P. subcapitata can be
influenced not only by the free Cd2+ ion, but also by the Cd exposure time (i.e., expression of
toxicity may be delayed after initial Cd accumulation). While P. subcapitata may actively
decrease Cd uptake on short-time scales (< 24 h), Cd toxicity inevitably occurred over chronic (>
24 h) exposures to relatively high Cd concentrations in solution. The present study also
demonstrates that metal-binding exudates released by P. subcapitata may significantly decrease
the proportion of total Cd present as the free Cd2+ ion. Such a decrease in Cd2+ concentration is
expected to decrease Cd uptake and toxicity in freshwater organisms. The use of flow cytometry
to follow algal growth at very low cellular densities can be helpful to minimize such exudation
and also possibly metal depletion over time. The present results afford insights about chronic Cd
toxicity mechanisms in a freshwater alga species commonly used in standard toxicity assays.

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
Such information should be helpful for the elaboration of robust algal bioassay as well as for the
derivation of water quality guidelines.
SUPPLEMENTAL DATA
Table S1. (52 KB. PDF).
Acknowledgement—We gratefully thank L. Martel, R. Cardin and C. Bastien from the Centre
d’expertise en analyse environnementale du Québec (CEAEQ) for their collaboration. We also
acknowledge the technical assistance provided by BD Bioscience, L. Rancourt, M. Bordeleau,
and S. Duval. ML held a scholarship from the Fonds Québecois de recherche sur la nature et les
technologies (FQRNT). CF and PGCC are supported by the Canada Research Chair programme.
REFERENCES
1. Campbell PGC, Errécalde O, Fortin C, Hiriart-Baer VP, Vigneault B. 2002. Metal
bioavailability to phytoplankton - Applicability of the biotic ligand model. Comp Biochem
Physiol C 133:189-206.
2. Campbell PGC. 1995. Interactions between trace metals and aquatic organisms: A
critique of the free-ion activity model. In Tessier A, Turner DR, eds, Metal Speciation and
Bioavailability in Aquatic Systems. John Wiley & Sons, New York, NY, USA, pp 45-102.
3. Paquin PR, Gorsuch JW, Apte S, Batley GE, Bowles KC, Campbell PGC, Delos CG, Di
Toro DM, Dwyer RL, Galvez F, Gensemer RW, Goss GG, Hogstrand C, Janssen CR, McGeer
JC, Naddy RB, Playle RC, Santore RC, Schneider U, Stubblefield WA, Wood CM, Wu KB.
2002. The biotic ligand model: A historical overview. Comp Biochem Physiol C 133:3-35.
4. Paquin PR, Zotay V, Winfield RP, Wu KB, Mathew R, Santore RC, Di Toro DM. 2002.
Extension of the biotic ligand model of acute toxicity to a physiologically-based model of the

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
survival time of rainbow trout (Oncorhynchus mykiss) exposed to silver. Comp Biochem Physiol
C 133:305-343.
5. Slaveykova VI, Wilkinson KJ. 2005. Predicting the bioavailability of metals and metal
complexes: Critical review of the biotic ligand model. Environ Chem 2:9-24.
6. Janssen CR, Heijerick DG. 2003. Algal toxicity tests for environmental risk assessments
of metals. Rev Environ Contam Toxicol 178:23-52.
7. Sunda WG, Huntsman SA. 1998. Processes regulating cellular metal accumulation and
physiological effects: Phytoplankton as model systems. Sci Total Environ 219:165-181.
8. Lavoie M, Campbell PGC, Fortin C. 2012. Extending the biotic ligand model to account
for positive and negative feedback interactions between cadmium and zinc in a freshwater alga.
Environ Sci Technol 46:12129-12136.
9. Sunda WG, Huntsman SA. 1996. Antagonisms between cadmium and zinc toxicity and
manganese limitation in a coastal diatom. Limnol Oceanogr 41:373-387.
10. Duval JFL, Rotureau E. 2014. Dynamics of metal uptake by charged soft biointerphases:
impacts of depletion, internalisation, adsorption and excretion. Phys Chem Chem Phys 16:7401-
7416.
11. Pinheiro JP, Galceran J, Van Leeuwen HP. 2004. Metal speciation dynamics and
bioavailability: Bulk depletion effects. Environ Sci Technol 38:2397-2405.
12. Vasconcelos MTSD, Leal MFC, van den Berg CMG. 2002. Influence of the nature of the
exudates released by different marine algae on the growth, trace metal uptake, and exudation of
Emiliania huxleyi in natural seawater. Mar Chem 77:187-210.

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
13. Hardstedt-Romeo M, Gnassia-Barelli M. 1980. Effect of complexation by natural
phytoplankton exudates on the accumulation of cadmium and copper by the haptophyceae
Cricosphaera elongata. Mar Biol 59:79-84.
14. Vasconcelos MTSD, Leal MFC, Van Den Berg CMG. 2002. Influence of the nature of
the exudates released by different marine algae on the growth, trace metal uptake and exudation
of Emiliania huxleyi in natural seawater. Mar Chem 77:187-210.
15. Croot PL, Moffett JW, Brand LE. 2000. Production of extracellular Cu complexing
ligands by eucaryotic phytoplankton in response to Cu stress. Limnol Oceanogr 45:619-627.
16. Koukal B, Rossé P, Reinhardt A, Ferrari B, Wilkinson KJ, Loizeau JL, Dominik J. 2007.
Effect of Pseudokirchneriella subcapitata (Chlorophyceae) exudates on metal toxicity and
colloid aggregation. Water Res 41:63-70.
17. USEPA. 2002. Short-term methods for estimating the chronic toxicity of effluents and
receiving waters to freshwater organisms. US Environmental Protection Agency, Washington
DC.
18. CEAEQ. 2007. Détermination de la toxicité : Inhibition de la croissance chez l'algue
Pseudokirchneriella subcapitata. Centre d'expertise en analyse environnementale du Québec
(CEAEQ), Québec.
19. Vasconcelos MT, Leal MF. 2008. Exudates of different marine algae promote growth and
mediate trace metal binding in Phaeodactylum tricornutum. Mar Environ Res 66:499-507.
20. Franklin NM, Stauber JL, Lim RP, Petocz P. 2002. Toxicity of metal mixtures to a
tropical freshwater alga (Chlorella sp.): The effect of interactions between copper, cadmium, and
zinc on metal cell binding and uptake. Environ Toxicol Chem 21:2412-2422.

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
21. Franklin NM, Stauber JL, Apte SC, Lim RP. 2002. Effect of initial cell density on the
bioavailability and toxicity of copper in microalgal bioassays. Environ Toxicol Chem 21:742-
751.
22. Postma JF, de Valk S, Dubbeldam M, Maas JL, Tonkes M, Schipper CA, Kater BJ. 2002.
Confounding factors in bioassays with freshwater and marine organisms. Ecotoxocol Environ Saf
53:226-237.
23. Leischmann A, Greene J, Miller E. 1979. Bibliography of Literature Pertaining to the
Genus Selenastrum. US Environmental Protection Agency, Corvallis, Oregon.
24. Komarek J, Comas AG. 1982. Taxonomical definition of the genera and several species
of Ankistrodesmus and Selenastrum (Chlorococcales). Algol Stud 32:259-277.
25. Krienitz L, Bock C. 2011. SSU rRNA gene phylogeny of morphospecies affiliated to the
bioassay alga "Selenastrum capricornutum" recovered the polyphyletic origin of crescent-shaped
Chlorophyta. J Phycol 47:880-893.
26. Dzombak DA, Morel FMM. 1990. Surface complexation modeling: Hydrous ferric oxide.
John Wiley and Sons, USA.
27. Schecher WD, McAvoy D. 1998. MINEQL+: A Chemical Equilibrium Modeling
System. 4.5 ed. Environmental Research Software, Hallowell, ME, USA.
28. Martell AE, Smith RM, Motekaitis RJ. 2004. NIST critical stability constants of metal
complexes, version 8. National Institute of Standards and Technology. Gaithersburg, MD.
Gaithersburg, MD.
29. Schecher WD, McAvoy DC. 1992. MINEQL+: A software environment for chemical
equilibrium modeling. Comput Environ Urban Syst 16:65-76.

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
30. Vigneault B, Campbell PGC. 2005. Uptake of cadmium by freshwater green algae:
Effects of pH and aquatic humic substances. J Phycol 41:55-61.
31. Bates SS, Tessier A, Campbell PGC, Buffle J. 1982. Zinc adsorption and transport by
Chlamydomonas variabilis and Scenedesmus subspicatus (Chlorophyceae) grown in
semicontinuous culture. J Phycol 18:521-529.
32. Fortin C, Campbell PGC. 1998. An ion-exchange technique for free-metal ion
measurements (Cd2+, Zn2+): applications to complex aqueous media. Int J Environ Anal Chem
72:173-194.
33. Marie D, Simon N, Vaulot D. 2005. Phytoplankton cell counting by flow cytometry. In
Anderson RA, ed, Algal culturing techniques. Academic Press, Elsevier, pp 253-267.
34. Fortin C, Couillard Y, Vigneault B, Campbell PGC. 2010. Determination of free Cd, Cu
and Zn concentrations in lake waters by in situ diffusion followed by column equilibration ion-
exchange. Aquat Geochem 16:151-172.
35. Mueller KK, Lofts S, Fortin C, Campbell PGC. 2012. Trace metal speciation predictions
in natural aquatic systems: incorporation of dissolved organic matter (DOM) spectroscopic
quality. Environ Chem 9:356-368.
36. Crémazy A, Leclair S, Mueller KK, Vigneault B, Campbell PGC, Fortin C. 2014.
Development of an in situ ion-exchange technique for the determination of free Cd, Co, Ni and
Zn concentrations in freshwaters. Submitted to Aquatic Geochemistry.
37. Xue H, Sigg L. 1998. Cadmium speciation and complexation by natural organic ligands
in fresh water. Anal Chim Acta 363:249-259.
38. Qian J, Xue HB, Sigg L, Albrecht A. 1998. Complexation of cobalt by natural ligands in
freshwater. Environ Sci Technol 32:2043-2050.

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
39. Xue H, Kistler D, Sigg L. 1995. Competition of copper and zinc for strong ligands in a
eutrophic lake. Limnol Oceanogr 40:1142-1152.
40. Kalis EJJ, Weng L, Dousma F, Temminghoff EJM, Van Riemsdijk WH. 2006.
Measuring free metal ion concentrations in situ in natural waters using the Donnan Membrane
Technique. Environ Sci Technol 40:955-961.
41. Sigg L, Black F, Buffle J, Cao J, Cleven R, Davison W, Galceran J, Gunkel P, Kalis E,
Kistler D, Martin M, Noël S, Nur Y, Odzak N, Puy J, Van Riemsdijk W, Temminghoff E,
Tercier-Waeber ML, Toepperwien S, Town RM, Unsworth E, Warnken KW, Weng L, Xue H,
Zhang H. 2006. Comparison of analytical techniques for dynamic trace metal speciation in
natural freshwaters. Environ Sci Technol 40:1934-1941.
42. Errécalde O, Campbell PGC. 2000. Cadmium and zinc bioavailability to Selenastrum
capricornutum (Chlorophyceae): Accidental metal uptake and toxicity in the presence of citrate.
J Phycol 36:473-483.
43. EC. 2007. Biological test method: Growth inhibition test using a freshwater alga, Method
Development and Applications Section, Environmental Science and Technology Centre,
Environment Canada.
44. Lavoie M, Fortin C, Campbell PGC. 2012. Influence of essential elements on cadmium
uptake and toxicity in a unicellular green alga: the protective effect of trace zinc and cobalt
concentrations. Environ Toxicol Chem 31:1445-1452.
Figure 1. Free Cd2+ ion concentrations as measured by the ion-exchange technique at t = 0 (open
circles) or t = 96 h (full circles) as a function of total measured Cd concentrations. The dotted
line represents the expected initial Cd2+ concentrations modeled with MINEQL+ (I = 200 meq L-

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved
1, CEAEQ medium, ionic strength adjusted with NaNO3) (slope of 0.83). The short dashed line
and the solid line are least-square best fits of linear regressions for the data at t = 0 and t = 96 h
respectively. Insert shows data on a finer scale. The initial cell density was 2,500 cells mL-1.
Error bars represent the standard deviation of two to four replicates. In the insert, only the upper
error bars are shown for the data at t = 96 h.
Figure 2. Intracellular Cd concentrations (mol Cd µm-3) as a function of exposure time for
different mean exposure total [Cd] (Panel A) and as a function of the measured mean exposure
total [Cd], for different exposure times (Panel B). Insert shows results on a smaller scale. Error
bars represent the standard deviation of three replicates.
Figure 3. Natural logarithm of the total cellular volume per mL (µm3 of alga mL-1) as a function
of exposure time to various measured mean exposure total Cd concentrations (between t = 12 h
and t = 96 h). The dotted, small dash and solid lines represent linear regressions for the control
(R2 = 0.97), the 18 nM (R2 = 0.99) and the 56 nM (from 48 to 96 h) (R2 = 0.98) Cd
concentrations. The initial cell density was 2,500 cells mL-1. Error bars represent the standard
deviation of three replicates.
Figure 4. Effective Cd concentrations (A, E: initial measured total [Cd]; B, F: measured mean
exposure total [Cd] between 12 and 96 h; C, G: final measured free [Cd2+]; D, H: intracellular
Cd) inhibiting by 50% the cell yield (cell number mL-1) (A to D) or the total cell volume (µm-3 of
alga mL-1) (E to H) measured after 48, 72 and 96 h of exposure to Cd. The EC50s (except EC50s
expressed as measured mean exposure total [Cd]) at 96 h were calculated for cultures inoculated
at two different initial cell density (2,500 cells mL-1 or 10,000 cells mL-1); all other time points
are at an initial cell density of 2,500 cells mL-1. No significant Cd toxicity was measured after 12
or 24 h of exposure. Error bars represent the confidence intervals at 95% (95%CI).
Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved

Acc
epte
d Pr
eprin
t
This article is protected by copyright. All rights reserved

1
Supporting information for
Cadmium accumulation and toxicity in Pseudokirchneriella subcapitata: influence of
metal-binding exudates and exposure time
Authors: Nathalie Paquet†‡, Michel Lavoie†§, Frédéric Maloney†||, Jérôme F.L. Duval#,
Peter G.C. Campbell†, Claude Fortin†1
† Institut national de la recherche scientifique, Centre Eau Terre Environnement (INRS-
ETE), 490 de la Couronne, Québec (Québec), Canada G1K 9A9.
‡ Present address: Centre d’expertise en analyse environnementale du Québec, Ministère
du développement durable, de l’Environnement et de la Lutte contre les changements
climatiques, 2700 rue Einstein, Québec (Québec), Canada G1P 3W8.
§ Present address: Québec-Océan and Takuvik Joint Université Laval/CNRS Research
Units, Département de Biologie, Université Laval, Québec (Québec), Canada G1K 7P4.
|| Present address: Administration portuaire de Québec, P.O. Box 80, Haute-Ville,
Québec (Québec), Canada G1R 4M8.
# Laboratoire interdisciplinaire des Environnements Continentaux (LIEC), CNRS-
Université de Lorraine, B.P. 40, Vandoeuvre-lès-Nancy cedex, France
1 Corresponding author: [email protected]

2
1. Description of the ion-exchange technique
In this technique, the free Cd2+ ion concentrations equilibrate with a sulfonic acid-type resin that has been pre-calibrated with Cd standard solutions.
↔ . 1 This reaction is described by a conditional equilibrium constant :
. 2
where R = resin, = resin-binding sites occupied mainly by Na, the predominant major cations (mol L-1), and = resin-binding sites occupied by metal M (mol L-1). In the presence of a sufficiently high concentration of counter-ion (Na+), the Na+ concentration in solution and adsorbed on the resin is negligibly affected by the free metal ion which is present at trace levels (i.e. [RNa+] >> [RMz+]). Then, eq. 2 can be rearranged yielding a distribution coefficient , , valid at fixed ionic strength (200 meq
L-1 adjusted with 1 M NaNO3) and pH (7.0).
, , . 3
Before analysing samples of culture media, the resin is firstly calibrated with solutions of known [Cd2+] (e.g. without significant complexation by ligands or in the presence of significant metal-ligand complexes with well-known stability constants) in order to compute , , for Cd. This coefficient can be calculated by measuring the amount of
metal bound to the resin , which is obtained after elution of the resin with a volume V (4 mL) of strong acid (1.5 M HNO3). According to the quantity of resin used (mr = 7 mg; weighed on a micro-balance; Sartorius, Edgewood, NY, USA) and the concentration of metal measured in the standard eluate ( ) (mol L-1), can be calculated with Eq.4.
. 4
Analyses of CEAEQ culture media adjusted at an ionic strength of 200 meq L-1 with NaNO3 and spiked with 100 nM total Cd (MINEQL+ simulation predict 83 nM Cd2+ due to Cd complexation by high nitrate concentrations) yielded a , , of 0.41 ± 0.05 L g-1
(n = 9).

3
After calibration of the resin, the ionic strength of sample solutions was adjusted to that of the standard IET matrix (200 meq L-1) with NaNO3 1 M (dilution did not exceed 1% of the sample volume). Then, samples were passed through the resin for 4 min (V = 20 mL), after which equilibration between the ion-exchange resin and the solution is achieved (i.e. the metal concentration entering and exiting the resin column are equal). Combining Eq. 3 and 4 yields Eq. 5:
, ,
. 5
To determine free Cd2+ concentrations in culture medium samples, cadmium bound to the resin is first eluted with 2 ml of 1.5 M HNO3 in pre-weighted borosilicate vials (Opticlear, Fisher Scientific, Ottawa, ON, Canada). Thereafter, the eluates collected in the vials were weighted again and the exact volume of elution was computed using the density of the nitric acid eluent solution (1.055 g mL-1). Cadmium concentrations in
sample eluates ( ) were then measured by counting the 109Cd radioisotope
with a scintillation counter (Wallac 1414; Perkin Elmer Life Sciences, Turku, Finland) using Ecolume scintillation cocktail (ICN, Costa Mesa, CA, USA). Finally, the free [Cd2+] of the samples was computed with eq. 5. Analytical blanks were also run in parallel of each sample to evaluate background contamination of samples from the nitric acid used for elution of the resin column and were subtracted from the sample values.

4
2. Tabulated EC50 values obtained in the present study The EC50 values that are plotted in Fig. 4 of the main text body are shown in the Table SI.1 below.
Cd toxicity based on the cell yield (cell number mL-1) EC5048h-2,500 EC5072h-2,500 EC5096h-2,500 EC5096h-10,000 Initial measured total [Cd] (nM) 74 (59-89) 44 (39-50) 23 (16-32) 82 (75-89) Mean measured exposure [Cd] (nM) 44 (35-55) 25 (21-29) 11 (7.1-16) - Final measured free [Cd2+] (nM) - - 1.3 (0.21-2.3) 35 (33-37) Intracellular Cd quotas (× 10-17 mol Cd cell-1) 2.8 (1.9-3.8) 0.85 (0.67-1.1) 0.35 (0.025-0.79) 4.9 (4.4-5.3)
Cd toxicity based on the total cell volume (µm-3 of alga mL-1)
EC5048h-2,500 EC5072h-2,500 EC5096h-2,500 EC5096h-10,000 Initial measured total [Cd] (nM) 99 (73-128) 73 (59-88) 66 (52-80) 112 (100-119) Mean measured exposure [Cd] (nM) 59 (45-76) 46 (36-56) 47 (36-58) - Final measured free [Cd2+] (nM) - - 1.4 (0.42-2.9) 50 (42-59) Intracellular Cd quotas (× 10-19 mol Cd µm-3) 4.3 (2.6-6.6) 2.4 (0.84-5.4) 1.1 (0.60-6.8) 7.5 (6.2-8.5) Table SI.1: Effective Cd concentrations (expressed as initial measured total [Cd], measured mean exposure total [Cd] between 12 and 96 h, final measured free [Cd2+], and intracellular Cd) inhibiting by 50% the cell yield (cell number mL-1) or the total cell volume (µm-3 of alga mL-1) measured after 48, 72 and 96 h of exposure to Cd. The EC50s at 96 h were calculated for cultures inoculated at two different initial cell density (2,500 cells mL-1 or 10,000 cells mL-1) except for EC50s expressed as a function of measured mean total exposure [Cd]; all other time points are at an initial cell density of 2,500 cells mL-1. No significant Cd toxicity was observed after 12 or 24 h of exposure. Data are presented as mean EC50time-initial cell density ± 95%CI.
Related Documents











