41 4, o -» I1 - 2 H , r Tiijli’.ii;;- hvimca 4 o 3s ■ I 1-1'! i:n< . !;•(•!! F* *.»&/. ?,)}< i'vC-ll'.JJ i7.‘l( • f. 5!/ FiffUre 1,41 CorT-ar » ;^Flv.ovoi-vaHt (5 X Mix “ v X ) o >r no-cn, i .. cm X /» i;» o n . (\ Iif;'' "C ■ i ) 1 ,f, S H U "1/ jj / M 'x i ! i !> it r • ‘ !.,i • I .. Figure L42 ('} W ti'M H v 5- ■^rs'tndiyu: { k ~ a ) 5-fl’dr inhibits the ORzyuo thy.ai<:ly1nte synthetase, This is the enzyiw which in the production of endogenous thy,«i dine converts dUMP (uridinn Boncphorjphats} to dTMP (thymidine monophosphate)» indicated in Figure l.ffj below. In the noniuil tnctuhol 1c cycle the thyisidine monophosphate is then converted to thymidine triphosphate (nucleotide) and in this form enters the DMA, I t can be seen therefore that i f thymidylciU* synthetase is inhibited* endogenous thymidine is not produced and in the absence of thymidine DMA cannot rep licate, Thiss of com sc» prevents ni to?»i s > as has been observed experimentally (ToHvet tuiu Sitnon 1967), Addition of v \ternu1 thymidine can reverse the effects of THUdK because DMA replication (requiring thymidine) can again take place.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

41
4,o
-» I1 -
2 H ,rTiijli’.ii;;-
h v i m c a
4o
3s
■ I1-1'! i:n< . !;•(•!!
F * * .» & / .
?,)}< i'vC- ll'.JJ i7.‘l( • f. 5!/
FiffUre 1,41 CorT-ar » ;^Flv.ovoi-vaHt
(5XMix “v
X )o >r
no-cn,
i..cm
X/»
i;»
on. (\I if;'' "C ■ i ) 1 , f , S H U "1/
jj/M 'x i ! i !> it
r • ‘! . , i • I ..
Figure L 4 2 ('}
W t i ' M H v 5- ■ ^rs'tnd iyu : { k ~a )
5-fl’dr inhibits the ORzyuo thy.ai<:ly1nte synthetase, This is the enzyiw which in the production of endogenous thy,«i dine converts dUMP (uridinn Boncphorjphats} to dTMP (thymidine monophosphate)» indicated in Figure l.ffj below. In the noniuil tnctuhol 1c cycle the thyisidine monophosphate is then converted to thymidine triphosphate (nucleotide) and in this form enters the DMA, I t can be seen therefore that i f thymidylciU* synthetase is inhibited* endogenous thymidine is not produced and in the absence of thymidine DMA cannot replicate, Thiss of com sc» prevents ni to?»i s > as has been observed experimentally (ToHvet tuiu Sitnon 1967), Addition of v \ternu1 thymidine can reverse the effects of THUdK because DMA replication ( requiring thymidine) can again take place.

. The actui.l reaction taking place during the formation of dTMP from dUMP is shown below and in Figure 1.43 at (1), TIk methyl group in the reaction below is transferred from r. 10 methylene tetra- hydrofoltite (CH.JF) to the 5 position of the pyrimidine ring of dUMP forming cITMF-' + dihydrofolato.
dUMP + CH.JF " • • x dTf# + DHF£ ---- - * ■ ■
tet” -ihyc!ro - dihydrofolatefolate.
The reaction is shown i*; Its context in the Metabol ic; scheme inFigure L43 below.

The actual bloi*f;<vj*< of th*1 activity of the enyynKi thyialdyloto
synthetase .occurs as foil owe. ;
dllMP normally binds the en/ynie thymidylatc synlhrtaso fK'ape) (1
mole cult, of dUHP to one snolocuH of thymidyluto synthetase), The
■inhibitor 5-4'tMR bind*, with this enzyme in the ratio two molecules
of 5-Ftk!R to one t.iolocuie of thyridylate r.ynthetsso (Jteldelbenjfir
1965), Tins is fhown in Fiouro 1,44 bo low.
TSasn + 1 dUM P-^ T fn iS ' v (d !., : r ) 1
TSase + 2FdUMr TSase - {rd lfr iP i *. ■ \ ' ” ■ ■ e.
Figure 3 .44 *.v.,v■*,' *r.V /•,. j o n i!n,:.iL'?ulaU
Once the enzy*-? has bean Huarf by dttT (normal reaction) It is used
to convert diKP to dl'IP by faeiM tal i tig the transfer of a mothvl
9**‘owl) 1 ro;»i 5» 10 f;’fct.hylc*:;o totraby dr**?r<lqte to the 6 position cf the
pyrinridine r1»K: of dUflF (K< rUran cud l!cidalboru?r 15G1). Methylene
tefcrabydrofo<»;£« is mj-MIr^d for the? inhibition of this reaction by
5-rUtid, {in the forr,! FdUItf*) a s s In ils alnerce, FdU»MP binds poorly to
tho The transfer of the Cb’5 uroup from tho mu thy low*
tetrahyrirofelate to the ddMP is shown in Figure 1.45 on page 44

K,:
d i'h y d r o f o i a t eV
o
"Clf.
5, iO-CK,«if< Foiatc/y ' methylene
teUfihydiofQlatii
Methyl Gioap0
.■A. IsShould ir. ;>), * •
this
0J p.-0-CIf , 0 {s' u
OH
t'.u,'.:?
HN ,j ^ cr^K-;.^
■Cit»<5
/ s-r-u*;:th^m idy tote*
"01F ~ < 0 ~ C H ^ O
b loct; a '*>* ” t hvt.»;•1*
ViOJ
..„Y'
0 dTMP■V
O'
Ai'~o-c;i. ^0V
7m ft It 0 s 'his
OHF-dUMJ!
Finure 1.45 :">vo;.vr,->> a/’ CH.. from to dVMP,
A further schemn of the3 5-ftldR block is shown below with 5-FUdR
entering tho cycle at (1) and forming FdUMP Instead of thymine being
able to form dTMP at (2). The FUdR enters the cycle as 5-FJdR
and is motabnl iscni to FHg as shown in Figure 1.46 on pago 45

n
eyele* 2'ke inhibition €»ic in inav1:cd I.
In conclusion* 5-FUdR inhibits the enzyme thyniidylate synthetase by
competing for its binding site and in so doing prevents the
formation of thymidine, thus preventing DNA replication. This causes
an inhibition of mitosis,
1.8.4. S-Bromodooxyuridine
5-Bromodeoxyuridine differs from 5-FUdR i,i that a nromine atom
rather than a fluorine atom is found at position 5 (see Finn re 1.41)
os shown in Figure 1.47 below,
5-BUdR replaces thymidine in the DNAS making the DNA heavier than
normal DuA. riguro 1,48 below shows the mode of entry of the 5-BUdR
into the DNA at the position marked 2.
NdR
Figure 1,47 Stmwluvc of &-l>t>m->deaxyuvid-inv (5-BUdR)

For comparison the position marked 1 again shows the 5-FUdR
block. DNA containing 5-BUdR prevents differentiation in cells in
culture although the cells g n w normally. This will be further
discussed in section *.3.5. below.
B U d K P P P
tlB U d R P P
B l i d R P
A
B U d R
Ii
BU
Bl oc ! ; in T h y m l d y l a t e S y n t h e s i s
F i g u r e _ L 48 E n t r y o f S -B U dR i n t o DNA . ( s r y b a l s k i 1 9 6 2 )
1,8.5. Effects of 5-BUdR and 5-FUdR on.cells in culture
Although cells vn ■ h 5-BUdR in their DNA are fully functional i.e.
they continue growing and dividing, the production of "luxury
proteins (proteins for the specialised functioning of the cell e.g.
myosin in muscle) is prevented. This can be reversed, as when the
5-BUdR is removed the cells proceed to differentiate normally.
Likewise the effects may be reversed if thymidine is added to
replace the shortage of endogenous thymidine. Stockdale et al (1964)
showed that myogenic cells (prospective muscle cells) in the
presence of 5-BUdR failed to fuse and form multinucleated myotubes
prior to becoming muscle.

Simon (1963) reported that HeLa cells in culture with both 5-BUdR
and 5-FUdR divided only once and incorporated slightly more 5~BUdR
than did cells in 5-BUdR alone. It could be suggested that this is
due to the lack of thymidine caused by 5-FUdR in the DN'A, (which the
cells replace with 5-BUdR). The one cell division is the final one
in the presence of 5-FUdR.
1.8.5.1. The effects of 5-FUdR on cells and organisms
when administered alone, with special
reference to concentration
Owing to the cost of 5-FUdR, prudent use was made in the administration
of this drug. As a first approximation to set a working concentration
of 5-FUdR, it was decided to follow the dosage concentrations for
Chinese Hamster cells which were worked out by Conrad and Ruddle
(1972). They showed that when Chinese Hamster cells were grown in
concentrations of 0,00005 f-ig/ml 5-FUdR, there was no reduction in
the rate of mitosis and hence no reduction in thymidylate activity.
In concentrations of 0,0001 pg/mT 5-FUdR, the rate of mitosis
initially slowed down, stabilised and then returned to normal,
showing an initial reduction in thymidylate activity. When these
workers used a concentration of 0,001 pg/ml 5-FUdR, the rate of cell
division was slowed down permanently and did not return to normal.
Thymidylate synthetase activity was reduced to 20% of its original
level, ,
Another worker (Ferguson 1978) who worked on the teratological
effects of 5-FUdR in Wister rats, injected 0,1 pg 5-FUdR per mg. of ’
body weight. ,
In the present study, tadpoles were maintained continuously in
water, and concentrations ranging from 0,2 - 20 yg/ml of 5-FUdR were
added to the water. These concentrations were 2000 times greater
than the concentrations of 0,001 jjg/ml that were effective in tissue
culture and 200 times greater than the concentrations injected into
the Wister rats. Furthermore, while the rate of uptake of the 5-FUdR
may have been relatively lower, this was compensated for by the
continuous application of 5-FUdR.

1.8.5.2. Effacts of 5-BUdR on rills and organisms
when administered alone, with special
reference to concentration
In the present project a concentration of 10 ng/ml 5-BUdR was ucid.
This was higher than that used in most tissue culture.experiments as
the tadpoles 'fere swum in the solution and it was assumed that the
chemical could be less toxic with this method than Jn tissue culture
as the tadpoles were only getting a fresh solution of 5-BUdR once
weekly.
While 5-BUdR affects differentiation, it does not affect cell
division and growth as shown by Bishoff and Holtzer (1970). The fact
that thymidine competes with 5-BUdR to enter the DMA was shown by
Stockdale et al (1964) where excess thymidine added to the culture
medium prevented the uptake of 5-BUdR, -
Abbott and Holtzer (1968) examined the effects of 5~BUdR on
chondrocytes taken from chick vertebral cartilage. The concentra
tion used was 20 ng/nil. The resulting chondrocytes had bizarre
shapes and the clones consisted of widely scattered fibroblastic
cells i.e. true condensations required for cartilage formation were
not seen. The mucopolysaccharide required for the matrix was
destroyed.
U s h e r and Cahn (1969) in their studies of cartilage cells
found that a concentration of 10~4M was toxic to the cells, while
10 M was not toxic, but prevented differentiation.
Agnish and Kochhar (1976) showed a relationship between the
developmental stagt- of mice embryos and their sensitivity to 5-BUdR.
From each embryo they removed the limbs and grew them in cell
culture, one on normal medium and one on 5-BUdR. They found the
following:
1. At a concentration of 2 ug/ml 5-BUdP, stage 26 to 29 embryos
(eleven day) showed a complete suppression of chondrogenesis.
2. Mid-eleventh day embryos needed 10 to 25 qg/ml 5-BUdR to
achieve the same effect.

3. If twelve and thirteen day embryos were used the effects were
milder no matter how high the concentration of 5-BUdR was.
The effectiveness of the drug coincides with the various stages
of cartilage differentiation, as the first signs of differentiation
•were apparent only in the late eleventh day embryo and after
differentiation has taken place, the drug has no more effect.
Abbott and Holtzer (1968), Coleman et al (1970), (1958), Lasher
and Cahn (1969) found that 5-FUdR had a proximo-distal effect. The
scapula, first to differentiate7became increasingly resistant to the
drug, while the more distal radius and ulna retained sensitivity
throughout. This is in keeping with Tschumi (1957) who shows that
differentiation proceeds in a proximo-distal direction.
Sal a and Rizotti (1975) used stage 40 or 47 (NF stage) tadpoles
(Xenopus laevis) and exposed them to 5-BUdR by injecting it into the
coelomic cavity (2ifl of 5 x 10'2M ) . The most severe effects were
noticed on stage 40 tadpoles reduction in pigmentation,
swelling in the anterior portion of the head, curvature of the tail
and accelerated cardiac rhythm. The effects were less severe in the
stage 47 tadpoles and at stage 51 no external modifications 'were
seen. Histologically, however, all the animals had suffered
abnormalities to some degree.
In conclusion 5-BUdR is found to affect the differentiation of
cells in culture and the severity of the effect is related to
concentration.
1 *9 Analysis of DNA by density gradient
aflalytical ultracentrifugation , '
In the present study DNA was made heavy by swimming tadpoles in a
solution of 5-BUdR. This heavy DNA was analysed by Density Gradient
Ultracentrifugati on. This technique is a means of measuring the
amount of 5-BUdR that has entereu the DNA due to the fact that the
5-BUdR makes the DNA heavy. Meselson and Stahl (1958) performed the
classic experiment using this technique on DNA labelled with 15N
heavy hydrogen. If several species of DNA of different density are
placed in an ultracentrifuge cell in a solution of caesium chloride
and spun for twelve hours or more at 20,000 to 50,000 revolutions
per second. As the DNA absorbs ultraviolet light at 260um ,
Ultraviolet photographs will display these various bands.

Hanawalt ^1968) has shown that the tjoyant density, of DNA containing
5~BUdR increases by 0,9 gem while W^i 1 et al (1935) have shown
that in the presence of 5~FUcH even more s-iiUdR is taken up as chown
by the fact tha'; the buoyant density of the DNA increases,. Further,
by this method one can distinguish between very heavy DNA which has
two strands substituted with 5-BUdR and hybrid DNA, which is
lighter as only one strand has been substituted with 5-BUdR.
The hybrid bands of DNA are found between the normal DNA and the
heavy DNA on the photographs. Vi nograd (1963) has postulated the use
of marker DNA bands using DNA of known density to determine the
density of the various bands of DNA. Each DNA species forms a band
at the position where the CsCl (caesium chloride) density equals the
buoyant density of that species in the ultracentrifuge cell. The
CsCl spins down forming a varied density solution throughout the
cell, the densest solution being at the base of the cell. The
various solutions of DNA stabilise in position in the cell when
their density is counterbalanced by the density of the CsCl
at a particular point in the cell. This takes up to 12 hours to
achieve. At this point bands can be seen on the ultraviolet
photographs.
Figure 1.49 below shows a cell with the various densities of
CsCl.
I n s i d e of c e l l
w i t h C s C l s o l u t i o n
D N A
A i r
M e n i s c u s Lowe r
d e n s i t y
L o w e r d e n s i t y H I g h e r d e n s i t y
Figure 1.49 Co Cl in the u ttmcantvifuge cell
XH i g h e r
density
Figure *].50dh p,5l shows-a typical result of an ultracentrifuge
ultraviolet photograph.

yDMA band D N A b a n d DMA bemd
Figure 1.50 Sketch of ultraviolet photograph showing
various DNA hands
' *
1.10 Objectives of this study .
The Hindiimb of Xenopus laevis appears to develop in a proximo-
distal direction (Tschumi 1957} under the influence of the apical
ectodermal ridge. The zone of polarising activity (ZPA) appears to
influence the antero-posterior patterning of the limb during its
development (Saunders anti Gasseling 1968}. Of the three limb axes,
proximo-distal, antero-posterior and dorso-ventralythe dorso-ventral
axis is established first, followed by the antero-posterior axis and
lastly the proximo-distal axis is determined.
The differentiation of the hind]imb tissues, muscle, cartilage
and connective tissue^appears to be determined by the cell's
position which the cell determines according to a gradient of a
particular chemical or it could determine its position according to
the amount of time it has spent in the “progress zone” (Wolpert
1981}.
The present study used 5-BUdR and 5-FUdR to study the above
processes further. As 5-BUdR prevents differentiation and 5-FUdR
prevents cell division, the hind!imb development was likely to be
affected in various ways. These effects could increase our
knowledge of the processes of cell differentiation and cell
patterning In the Xenopus laevis hind!imb.

52
2. METHODS AND MATERIALS
2.1 Experiments investigating the effects of 5-BUdR and 5-FUdR
on the shape and patterning of the Xenopus laevis hindlinth
2.■ 1.X. Tadpole breeding and rearing
For the experiments in which the tadpoles were swum in 5-BUdR and 5-
FUdR a reliable, regular supply.of tadpoles was required. Xenopus laevis
responds well to life in captivity, in that it breeds all year round
when kept at warm temperatures and treated with a regular dose of
breeding hormone (Pregnyl). The tadpoles respond equally well by
growing and metamorphosing as long as the temperature is kept
constantly warm. For these reasons Xenopus laevis was chosen as the
amphibian for this study.
Xenopus laevis tadpoles were bred from adults supplied by
Jonkershoek Island Fish Hatchery, Stellenbosch. Females and males
were isolated and fed on chopped beef liver twice weekly in summer.
When breeding became more difficult, in winter, they were fed daily.
The water was changed after feeding and the frogs were kept at a
temperature of 25°C->v/hich is conducive to breeding. Breeding was
induced by Pregnyl (Organon). A solution was made up of 4500 units
in 9m 1 water, about 500 units/ml. The male was given 2 doses of
0,3ml, about 300 units in total. As breeding became more difficult
to induce during the winter, it was necessary to prime the males
daily for two weeks with the above dosage until the black nuptial
pads appeared. The females did not require increased doses. The
hormone was injected through the dorsal lymph sac (Gurdon 1967). The
frogs were then placed in a laying tank in shallow water on a mesh.
The eggs dropped through the mesh and were collected the following
morning and placed in shallow trays at 25°C, At this temperature
they developed to stage 43 in three days (Nieuwkoop and Faber 1967).
At about this stage they started feeding and were fed on Liquifry
No. 2 (Liquifry Company Ltd, Dorking), about ten drops daily to
sixty tadpoles in two litres of water, The tadpoles were thinned out
constantly to prevent growth inhibition by overcrowding. The water
was changed regularly.

2.1.2. In vivo experiments ii. which Xertopus laevis
tadp^les were swum in 5-BUrlR and 5-FUdR and
analysed for growth deformities_ •
The following experiments were sot up to test the effects of 5~pJdR
and 5-FUdR on Xenopus laevis tadpole growth, A weekly analysis was
carried out under the dissecting microscope and hindlimb deformities
were noted. (See section 3.3 for pictures.) The hi'rJlimb was
therefore focussed on for further experimentation.
2.1.2.1. Pilot experiments „
The following initial experiments were set up :
a) Experiment started at stage 43 (NF stage)
Solution Volume (ml) Number of tadpoles
(2 dishes, 6 tadpoles
per dish)____________
5-BUdR 10 ug/ml +
5-FUdr 0,2 ug/ml 200 2 x 6
5-FUdR 0,2 ug/ml 200 2 x 6
5-BUdR 100 ug/ml 200 2 x 6
5-BUdR 10 ug/ml 200 2 x 6
5-0UdR 1 ug/ml 200 2 X 6
5-BUdR 0,1 ug/ml 200 2 x 6
Water 200 2 x 6
b) Experiment started at stage 46 (NF stage)
Solution
5-BUdR 10 ug/ml
"5-BUdR 10 M,g/ml ■
J5~FUdR 0,2 ug/ml
5-FUdR 20 qg/ml
Water
Volume (ml)
200
200
200
200
Number of tadpoles
(2 dishes, 6 tadpoles
per dish) ___
2 x 6
2 x 6
2 x 6
2 x 6

54
2.1.2.2. Further1 experiments using Xenopus
laevis tadpoles swum in 5-FUclR
On the basis of the results of the pilot experiments in 2.1,2,1.
further experiments were set up in which the tadpoles were swum only
in 5-FUdR as 5-BUdR was noted not to have had any externally visible
deforming effects on the hindlimb.
The following experiments were set up :
Stage Solution Cons, entrati on Volume Number of tadpoles
(Expenment tig/ml ml (2 dishes, 6 tadpoles
begun) ^ per dish)
0 FUdR 10 200 6 x 2
46 FUdR 20 200 6 x 1
46 FUdR 10 200 6 x 1
46 FUdR Id 200 6 x 2
47 FUdt* 20 200 6 x 2
4B FUdR 20 200 5 x 1
48 FUdR 10 to o o 6 x 1
49 FUuR 20 200 6 x 1
49 FUdR 10 200 6 x 1 .SO FUdR 20 200 6 x 2
52 FUdR 20 200 6 x 2
43 Water 200 6 x 4
(The tadpole stages used can be seen in Appendix A.)
In order to study the deformed limbs in further detail, and
for the purposes of clearer photography, the tadpoles were fixed and
the nmbs stained, removed and mounted for photography (whole
The tadpoles were fixed in Karnovsky's fixative (Kernovsky
1S6£>) as this was found to be less damaging than the formaldehyde
mentioned in the same article.
Kri rnov s ky1 s FI xat i ve .4g of parufmmalriwhyde are dissolved in 46ml water, The temperature
is raised to 60oC in u water bath and 1M NaOH is added dropwise
until thi* solution clears,
Al'-1 60 1:11 i w C «nd W m glutaraldeliydu. Titrate ,t
toofti t f . i n rr-s to pH 7>2 ** 7j4 usinrj IN HC'l
f

55
Buffer C -- 41,5ml soln A + 8j5m1 soln B.
Soln A « Nr.HgPO^ HgO (mouosodium phosphate) 2?26%
Soln D = -NaOH (sodium hydroxide) ?.,52$.
Washing solution 50^ buffer + 50!' glutaraldehyde solution.
The tadpoles were fixed for 24 hours and then washed for 24 hours.
They were then transferred to acid alcohol for 24 hours (1% HOT in
70% ethyl alcohol) (Mahoney 1973), They were then stained for one
week in 1% Victoria Blue in acid alcohol, They were then differen
tiated in several chances of acid alcohol until the desired colour
was obtained. When differentiation was complete they were transferred
to 90ri alcohol for twelve hours. They were then dehydrated thoroughly
in several changes of absolute alcohol for a few hours. When
completely dehydrated they v/sre cleared in methyl benzoate and
stored in fresh methyl benzoate, inis method stained the cartilage
blue and mode the limb transparent so that the cartilage could be
clearly seen, The limbs wort1 no»» whole mounted and photographed
having been severed iroa the animal. The preliminary experiment?
were photographed on the whole aniv.al (see 3,1). These whole mounts
were analysed by r,*oa?ts of a deformity index (see section 3.2),
2.2 Radioactive thymidine uptake* in the presence of
S-FUdK by _X;f;na;*us laevis tadpole hind!imbs
As 6-FUdR is known to depress the uptake of thymidine (by decreasing
DNA replicatioi which reduces the requirement for thymidine), it was
decided to test this by si/mming Xenqpus laevis tadpoles in
radioactive thynidinc* (^IMiiynridina). The experiments were set up as
follows ;
Control
One dish of 200»tl capacity with six stage 49 tadpoles which were
swum in the solution of 20 ug/ml 5-FUdR for one week. They ware
unfed for throe days then fed on liquifry No. 2 for four days.
Experiment
One dish of 200;al capacity with six stage 49 tadpoles which were
swum in tirj solution of 20 j j/ml 5-FUdR + 5 fjCi/ml thymidine for one
week, They v.vru unfad for three days, then fed on liquifry No. 2 for
fOur d;iyF..

The limbs of the tadpoles were removed and weighed as were the
tails. They were than placed singly into clean glass scintillation
vials containing 1ml of soluene which was used to solubilise the
tissue. The vials were immediately capped and incubated at 45°C in a
water bath for eight hours to assist the solubilisation process.
10ml of a specially prepared Toluene based mixture was added to each
vial. The vials were recapped and thoroughly shaken. The vials were
then placed in the dark in a refrigerator at approximately 6°C for
twelve hours.
The vials were then counted in a Packard Tri-Carb Scintill
ation Counter (Series 3*000) for fifty minutes each. Counts of
radioactivity were registered on all three channels., each channel
having a window setting of 50 - 1000 and a gain setting of 53% for
■tritium. The couits per fifty minutes were then reduced to counts/
min/mg. (Price 1973.) The results are analysed in section 3.3.
2.3 Buoyant density gradient analysis of 5-BUdR DNA
It was decided to analyse the amount of 5-BUdR entering the DNA
using buoyant density gradient analysis and to compare this to the
uptake of 5-BUdR by the DNA in the presence of 5-FUdR. In the
presence of 5~BUdR DNA should become heavy and in the presence of 5-
BUdR and 5-FUdR DNA should become heavier still as the lack of
thymidine should cause the uptake of more 5-BUdR. The tadpoles were
swum in the solutions as follows :
a) 100 stage 41 (NF stage) Xenopus laevis tadpoles in 200ml water
for three days at 21°C. These tadpoles were killed at stage 43,
b) 100 stage 43 (NF stage) Xenopus laevis tadpoles in 200ml of 0,1
mg/ml 5-BUdR for five hours at 21°C.
c) 100 stage 43 (NF stage) Xenopus laevis tadpoles in 200ml
0,1 mg/ml 5-BUdR and 0,0002 mg/ml 5-FUdR (0,2 ng/ml as 0,001 mg
= 1 ug) for five hours at 21°C.
The DNA was then extracted from the tadpoles and analysed by buoyant
density ultracentrifugation. '

57
2.3,1. DNA extraction
DNA extraction was done essentially according to the method of
Marmur (1961) and Grossman, Goldring and Marmur (1969),
a) 100 tadpoles were killed in 15 ml of ice cold EDTA solution
(Ethylene Diamine Tetra-acetic Acid* Disodium Salt, Merck
Analytical Reagent). EDTA solution : 0,1M EDTA + Q J M NaCl. The
solution was adjusted to pH 8. The tadpoles were homogenised in
this sol u Li on by nujans of an ultra-turrax homogeniser, During
this process, the test tube with the solution was placed in a
beaker of ice (Ledoux and Huart 1967).
b) The cells were lysed in Sarkosyl NL 30, Sarkosyl NL 97 or
Sarkosyl 0 (samples by kind donation of Ciba-Geigy), The
Sarkosyl was used at a concentration of 1,3^. The mixture was
placed in a 50WC water bath for ten minutes and then allowed
to cool to room temperature (Marmur 1961), Sarkosyl is a new
detergent used by Grossman, Goldring and Marmur (1%9) instead
of SDS (sodiws dodecyl sulphate) used by most earlier workers.
SDS is usually used to dissociate the protein from the nucleic
acid but the ar.nds derivative, sodium dodecyl sarcosinate, is
now used due to its high solubility in concentrated CsCl.
Probably the hydrocarbon chain of sodium dodecyl sulphate
competes f*r hydrophobic bonds. The anionic detergent appears
to inhibit nucleases and the negative charge prevents inter
action with nucleic acids (Noll and Stutz 1968). On comparing
DNA yields for tho different detergents, Sarkosyl was found to
give a higher yield than SDS. Of the three Sarkosyl NL 30 was
used instead of the Sarkosyl NL 97 used by both of the above
groups of research workers, as it gave a better DNA yield.
Sarkosyl surfactants are high molecular weight carboxylic acids
or their own sodium salts of the form : OIL (CiL) CON (CH )
CH2 ~ COCH (or COOKii). 3
They are modified fatty acids in which the hydrocarbon chain is
interrupted by an ami dome thy] group (-CONCIIg-). They are the
amide derivatives of SDS (also known as sodium lauryl
sulphate). This modification improves the solubility and
crystal!inity of the molecule, enhances the acidity of the
carboxylic acid group and increases the absorption characteristics.

58
They are soluble in most organic solvents and are appreciably
soluble at mildly acidic pH's. They foam less than SDS
(Handbook - Geigy Industrial Chemicals),
c) Sodium perchlorate (pure reagent) was added at a high concen
tration to separate the protein from the nucleic acid (Harmur
1961). 1*838 g were added to the lysate thus forming a 5M
solution which was then adjusted to pH 5.
d) The solution was deprote'?niscd by shaking in 30ml chloroform r
isoatuyl alcohol 24 ; 1 (isoamyl alcohol 3 methyl butanol
CgHijOH pure reagt.it, Merck), The solution was shaken gently
for thirty minutes in a stoppered flask.
The chloroform causes surface denaturation of the protein,
while isoa.r.j 1 alcohol reduces foaming, aids the separation and
maintains the stability of the layers of the centrifuged
deproteinised solution {tfarmur 1961).
b) The emulsion was separated into three layers by a ten minute
centrifugation at 2000 rp:n, The upper aqueous layer containing
the nucleic acids was removed with a Pasteur pipette. The
protein interface and chloroform layer were discarded.
f) Further deprotelnisation was accomplished with pronase
griseus, Wiles-Seravac (Grossman, Goluring and
Harmur lOC'J)),
It was made up as follows :
300ag dissolved in 300«1 water and placed in a water bath at
37 C for forty-five minutes. The solution was then adjusted to
pH 5 with concentrated MCI and left a t t e m p e r a t u r e ^
forty-five minutes. Solid Tris Buffer (Seravac) was th, ,ed
to adjust the pH to 7 and the enzyme was used at a final
concentration of 1 mg/ml and allowed to act for three hours at
37 C (Stern 19G8). As well as digesting protein, pronase partly
digests itself (McCarthy 1968),
g) The lysate was dialysed against SSC for three hours at room
temperature and then overnight at 4°C (Grossman, Goldring and
Ha ray r 1963), SSC (standard saline citrate) - 0,1514 NaCl +
0,015M Trisodium Citrate (?~Hydrate) (Analytical Reagent,
llorck). 5

h) The lysate was treated with Ribonuclease (RNase) (Bovine
Ribonuclease Grade II 5 x crystallised Mi'les-Seravac). The
enzyme was used at a final concentration of 50 u,g/ml and
allowed to act at 37°C for three hours (Marmur 1961). RNase
also partly digests itself (McCarthy 1968). The RNase was made
up as follows (Kalf and Greee 1968) :
At a concentration of 1 mg/ml in acetate buffer pH 4,5 and
incubated at 37°C for thirty minutes. Acetate buffer - 0,001M
sodium acetate (trihydrate) (Analytical Reagent BDH) + 0,001M
EDTA. pH was adjusted with concentrated glacial acetic acid. The
solution was heated at 80°C for ten minutes to inactivate any
DNase.
i) The lysate was dialysed at room temperature for four hours and
then overnight at 4°C (Marmur 1961). It was dialysed against
0,2% SSC + Q,15M NaCl at a pH of 5.
The dialysis tubing was handled only with rubber gloves, as
enzymes on the hands could denature the DNA. Dialysis removes
the detergent, enzymes and the prodi • if enzyme degradation
(Kalf and Grece 1968).
j) The nucleic acids were precipated by addition of absolute ethyl
alcohol (Kalf and Grece 1968). The DNA was then centrifuged
down and dried in a vacuum for two hours and redissolved in 3ml
SSC. A few drops of chloroform were added and it was stored at
4°C. •
2.3,2. Buoyant analytical density
gradient ultracentrifugation "
The extracted DNA was analysed by buoyant density ultracentrifuqation.
2.3.2.1. Theory of analytical density
• ■' dient ul tracentri fugati on
As 5-BUdR has been found to enter the DNA, and as bromine has a
higher molecular weight than the methyl group which it replaces, the
buoyant density of the substituted DNA has been found to increase
and this principle has been used by Meselson, Stahl and Vinograd '
(1957) to study molecular weights and partial specific volumes of
macromolecules.

h) The lysate was treated with Ribonuclease (RNase) (Bovine
Ribonuclease Grade II 5 x crystallised Miles-Seravac). The
enzyme was used at a final concentration of 50 i^g/ml and
allowed to act at 37°C for three hours (Marmur 1961). RNase
also partly digests itself (McCarthy 1968). The RNase was made
up as follows (Kalf and Grece 1968) :
At a concentration of 1 mg/ml in acetate buffer pH 4,5 and
incubated at 37°C for thirty minutes. Acetate buffer - Q,001M
sodium acetate (trihydrate) (Analytical Reagent BDH) + 0,001M
EDTA, pH was adjusted with concentrated glacial acetic acid. The
solution was heated at 80°C for ten minutes to inactivate any
DNase.
i) The lysate was dialysed at room temperature for four hours and
then overnight at 4°C (Marmur 1961). It was dialysed against
0,2% SSC + 0,15M NaCl at a pH of 5. ,
The dialysis tubing was handled only with rubber gloves, as
enzymes on the hands could denature the DNA. Dialysis removes
the detergent, enzymes and the products of enzyme degradation
(Kalf and Grece 1968).
j) The nucleic acids were precipated by addition of absolute ethyl
alcohol (Kalf and Grece 1968). The DNA was then centrifuged
down and dried in a vacuum for two hours and redissolved in 3ml
SSC. A few drops of chloroform were added and it was stored at
4°C. •
2 -3>2‘ Buoyant analytical density
gradTsnt ultracentn'fugation *
The extracted DNA was analysed by buoyant density ultracentrifugation.
2.3.2.1. Theory of analytical density
, gradient ultracentrifuoation
As 5-BUdR has been found to enter the DNA4 and as bromine has a
higher molecular weight than the methyl group which it replaces, the
buoyant density of the substituted DNA has been found to increase
and this principle has been used by Meselson, Stahl and Vinograd '
(1957) to study molecular weights and partial specific volumes of
macromolecules.

The method involves an observation of the equilibrium
distribution of the macromolecu! ar material in a. density gradient,
itself at equilibrium. The density gradient is established by the
sedimentation of a low molecular weight solute in a solution subject
to a constant centrifugal field. A solution of low molecular weight
is centrifuged until equilibrium is closely approached. The
opposing tendencies of sedimentation and diffusion have then
produced a stable concentration gradient of Lhc low molccular wpight
solute. The initial concentration of low molecular weight solute,
the eentrifug field strength and the length cf the liquid column
are chosen so that the range of density at equilibrium encompasses
the effective density of the macromolecu!nr mate. ial. At equilib
rium, the reacromolecules reach positions at which the density of the
surrounding liquid is equal to their own, thus m a c m o l e c u l e s of
different easil y band at different positions in the ultra centrifuge
cell (Meselson, Stahl and Vi nograd 1557, and Heckman Review).
As the molecules sediment towards this bottom of the cell a trans
parent solvent solution region is created and the image of this
portion of the cell becomes darter. Where the ultraviolet light is
absorbed by the DNA light bands will be seen. By means of a
densitometer the image on the film can be converted into a plot of
optical density versus distance from the axis of rotation (Meselson
and Stahl 1958 Beckman 1962).
The advantage of this optical system is that concentrations of
0,0012 of D M can be detected which is of the order of maqnitude
likely to be found in the experimental samples (Beckman Instruction
Manual E~ 114-3).
2.3.2.?,. ^ethod Jised
One of the most effective low molecular weight substances found for
DNA is CsCl, which combines with it to form the cncsiu... salt.
Meselson, Stahl and VinogrM (1957) chose it due to its high
solubility, high molecular weight and high density which allows the
obtaining of steep gradients, 1,25 to 1,90 g/ml. It also has a low
viscosity suitable (or rapid sedimentation, is non-toxic to DNA and
is soluble in water (Dirk::) and salt solutions (Szyhalski 1968).

The average buoyant density of UNA in CsCl is +/- 1,7 g/ml
and thus a caesium chloride solution was <„ade up at average density
1 ,699 g/ml at 25°C, t’e temperature used the ultracentrifuge
runs (Szybalski 1968).
'rhe experimental work was done: on a Beckman Model E Analytical
Ultrac^ntrifuge, A 30mm charcoal-filled Epon centrepiece was used as
CsCl is corrosive to aluminium (Vinograd and Hearst 1962). The
longer centrepiece was chosen, as low volumes of DNA of a certain
density were more likely to be detected. 0,2 to 0,5 ug enn be used
(Vinograd and Bruner 1966), A counter-balance of 30 mm v/as used.
The optical density of the sample v/as read on a Beckman
Ultraviolet Spectrophotometer at 260
.The sample was diluted to an optical density of approximately
0,65 with SSC. 2,85 g of CsCl (Optical Grade - Suprapur Merck) were
added and the sample made up to a weight of 5,09 g with SSC, The
refractive index of the solution was read off art Abbe Refractometer
and the density read off from t ties (Chervcnka 1969).
The upper plain window of the cell was replaced with a 2°
negative wedge window to correct for the light scatter caused by the
density of the solution (Vinograd and Hearst 1962).
The AN.E Rotor was used and the machine was run at 44 770 rpm
in order to allow the sedimentation of the macromolecules in the
dense gradient (Meselson, Stahl and Vinograd 1957, Grossman,
Goldring and Marmur 1969).
The duration of the run was eighteen hours and photographs were
taken at two hourly intervals with an exposure of two minutes.
The photographs were analysed in a Beckman RB Analytrol with a
Film Densitometer attachment.
The following three runs were performed :
Optical Refractive Density
Density Index CsCl
DNA CsCl
M m l )
(9/ml)
Volume
used
M L1. Water 3 hrs. 0,58 1,3991
2. 5-BUdR 5 hrs. 0,52 1,4000
3. 5-BUdR h* 5-FUdR 0,46 1,4005
5 hrs.
The photographs and densitometer traces are shown in section 3.4,
1.699
1.699
1,720
1.3
1,8
1.3

2.4 Investigation into the growth of tadpoles
‘ under different methods of feeding
It was decided .to investigate the -optimum growth conditions for the
tadpoles, as far as feeding was concerned, in both large trays (2
litre capacity normally used) and small dishes (200 ml capacity used
in this study due to scarcity of 5-FUdR).
Tadpoles were taken at stage 47 (NF stage) and placed into
trays of 2 litre capacity and large surface area. Sixty tadpoles
were placed in two litres of water as this was the concentration of
tadpoles used in the small (200 ml) dishes.
Experiment A
Food Pose Number of
Tadpoles
Commercial
baby food
(liver and
vegets
Complan
Yeast
Liquifry
1 9/day for 2 weeks
2 g/'de.y for rest
60
60
60
CO
Volume of
Water (litres)
2
22
Experiment B
Commercial
baby food
(liver and
vegctaMos)
Fish fond
(Tetram In)
Yeast
Liquifry
Lettuce
Dose
1 g/day for 2 weeks
2 9/day for rest
Number of
Tadpoles
60
60
60
60
60
Volume of
Water (litres)
2
2
2
2
2

63
Experiment C
Food
Commercial
baby food
(liver and
v e g e t
Yeast
Fish Food
(T c t r a m in )
Lettuce
Liquifry
Dose Number of
Tadpoles
0,16 g/day - V weeks
0 S32 g/day for rest
6
6
6
Volume of
Water (mis)
2C0
200
200
200
200
The tadpole'" were analysed for rate of growth once weekly by
counting thv, nuober of arrivals at any one stage (NF stage). The
results are shown in section 3.5.

64
£.5 Details of the chemicals used
and source -11st of suppliers
1. BLIdR, 5-Brcmo 2‘deoxyuridin?, 5 1gma Chemical Corporation,
2. 5-FUdR, 5-fluoro 2'deoxyuricHne kindly donated by Dr, Raw of
Roche Products (Pty) Limited,
3. Victoria Blue, 4R Merck, Art 8539.
4. ■■Tetramin, Lopis* Johannesburg, containing fish meat, fish roe,
fish liver, crayfish, water plants, mussel meat* brine shrimp*
wheat germ, cod liver* insect larvae, kelp,
5. Liver and vegetables, Licence Gerber Prod. Company, USA,
Purity Ricket and Column SA Ltd, containing modified maize
starch, liver, soya protein, tomatoes, yeast, onion extract,
caramel, iodised salt.
6. -Yeast* Anchor Yeast (Pty) Limited, Vlatt Street, Industrie,
OohnnnesburQ, .
7. Comp!an, Glaxo Allenbury's $A (Pty) Limited, Manchester Road,
K'adeville, containing protein 3lg» fat 16g, carbohydrate 44gs
calcium 850,'Jig, phosphorus 7S0isig, sodium 400mg, chloride 750sng,
potassium llOGawt, iron 7,7nig> iodine 44ug, Vit A 1100 units,
Vit Bj i,0mg.
8. CsCl, Art 2041 Merck 2525019 extrapur tm, 168,36,
9. Soluene 350 (Packard) is an extremely efficient solubiliser of
animal tissue. Since most of the Biological samples containing
radioactivity measured by liquid scintillation counting are not
readily soluble in the aromatic hydrocarbon based scintillation
solution (Toluene) special solubilisers are required to obtain
a hort'ogenous system for reproducible measurement of radioactivity.
10. Toluene scintillant is composed, of 100ml Permafluor III
(Packard) in 1 litre of folueno (Packard), The resulting
solution contains 5 ,0y PPO and 0,bg bi s~MSB per litre, PPO and
bis-MSB are highly efficient scintillators which will not form
quenching adducts with tissue solubilisers, aminos and acids,
11. Toluene mixture. The solubilising mixture was made up as
follows :
Porniafluor 111 Packard : 70 •>
Triton X 100 Packard : 3d.: ,

65
Perntafluor is a concentrated solution of stable efficient
scintillators (PPO and bis-MSB), It does not form quenching
adducts with tissue solubiliser, amines and acids, Triton X
100, especially purified for liquid scintillation counting,
is an efficient tissue solubiliser* so that the mixture of
Permafluor and Triton is an extremely efficient scintillant.
This mixture modified from Thornley 1971» was found to be the
most efficfc»;t method of detecting radioactivity in aqueous
biological samples. Samples were* incubated in the dark to allow
them to equilibrate and to reduce cherniluminescence to a
minimum (Packard handbook), .
12. Methyl 3H Ihytr.idino specific activity 18 Ci/mmol labels DNA.,3
13. 5 }6 H Uridine (methyl) specific activity 4U,0 Ci/mmol label A,
14. Liquifry tlo, 2 (liquifry Company Limited, Dorking),
15. Pregnyl (Orgr.non), This is a mammalian gonadotropin which in
the male fro:; stimulates the growth of the testes* sexual
maturityand release of the sperm9 as well as the formation of
dark nuptial pads on the insides of the fore-limbs. The
formation of the nuptial pads indicates that the males are in
mating condition,
16. EDTA, Ethyl one Diamine Tctra - acetic acid, di sodium salt,
Merck analytical reagent,
17. Sarkosyl tit. 30* sarkosyl NL97* sarkosyl 0, by kind donation of
Ci ha Go i
18. Isoamyl alcc-hol - 3 methyl butanol CgH^OH pure reagent Merck.
19. Pronase, Sjjhscus Miles.Soravac.
20. ftibomicler.., t Bovine Ribonucleaso Grade II 5X crystal! H o d
Miles Seravac.
21* Sodium Acetate, Analytical Reagent BOH.

3. RESULTS
The hind!itnb buds of the tadpoles swum in 5-FUdR were found to be
deforr J (see section 2.1) either in shape or lacking the full
number of digits or both. A method was devised to approximate the
degree of deformity of each limb, In the limb each defect was given
equal weighting as described on page 73. For convenience, the limbs
were divided into three types: adult limbs (56 - 65) were given a
count of 9.1 when normal, medium mature limbs (54 - 5b) were given a
count of 32 when normal and immature limbs were given a count of 24
when normal. From these normal counts points were removed depending
on the type of deformity and the result is referred to as the
deformity index for each limb.
3.1 Results of pilot experiments in which Xenopus laevis
tadpoles were swun in solutions of 5-FUdR and 5-BUdR
A series of initial exp r ments revealed the hindlimb deformities on
observation of the tadpoles under the dissecting microscope. These
tadpoles were subsequently photographed to show the deformities. The
photographs of these tadpoles are shown in the next few pagt#*. In
section 2.1 the limbs were further analysed by staining the
cartilage and taking magnified photographs of the result. Below, in
Figure 3.1, is a picture of a tadpole (NF stag-' <f0) with three toes
on the left hindlimb and two toes on the right hindlimb.

Figure 3.1 Dorsal view of stage 60 tadpole swim in O^uglmi
S-FUdR. A claw is visible on the third toe oj
the left foot.
In Figure 3.2 below are sketches of the hindlimbs of the above
tadpole compared to the sketch of the normal tadpole hindlimb for
that stage.
Fiqure 3.2 Sketch of the hindlimbs of the tadpole in Figuve----r.------------ - & '3.1 conrpaved to that of tthe normal stage GO hindlimb.

The tadpole in Fi g u r e 3.1 can be compared to the normal stage 60
tadpole shown in Figure 3.3 below.
Figure 3.3 Photograph of normal stage 60 tadpole.
This type of result was found consistently throughout the various
preliminary experiments as will be seen in the following photo
graphs (Figures 3.4 to. 3.11),. A stage 53 tadpole was seen to have a
bifurcated hindiimb bud instead of the normal division into five
toes at this stage. See Figure 3.4 below.
Figure 3.4 L a t e r a l v iew o f phage 58 tad p o le swum i n ,. lOfcg/ml
5-BudR + %^{j/mi s-nm.

The tadpole in Figure 3.1 can be compared to the normal stage 60
tadpole shown in Figure 3.3 below.
1 1
*; 1 J ! «jf' -
Figure 3.3 Photograph of normal stage 60 tadpole.
This type of result was found consistently throughout the various
preliminary experiments as will be seen in the following photo
graphs (Figures 3.4 to 3.31). A stage 53 tadpole was seen to have a
bifurcated hindiimb bud instead of the normal division into five
toes at this stag*. See Figure 3,4 below.
ti
Figure 3.4 Lateral view of stage 53 tadpole miun in .. 10/-<g/ml
5-BUdR •/- lf^g/ml 5-Fl

The same tadpole was-enlarged to show only the abdominal region so
that the hindlimb was clearly visible (see Figure 3.5 below).
Figure 3.5 Abdominal region of stage 53 tadpole enlarged to
show detail of bifurcated hindlimb.
Figure 3.6 Sketch of the above hindlimb with a normal
limb for ooinparison.

70
A photograph of the normal stage 53 tadpole is shown below
comparison to the photograph in Figure 3.4.
Figure 3.7 Photograph of normal- stage S3 tadpole
In Figure 3.8 below is a photograph of a stage 57 tadpole with
a hindlimb having three toes.
Figure 3.8 Lateral view of stage 5? tadpole swum in lOojJg/ml
5~BUdR + , OjJjHg/ml 5-FUdR. Only three toe $ c m
be seen.

71
The photograph in Figure 3.8 has been enlarged to show the
hindlimb more clearly.
Figure 3.9 Enlargement of the hindlimb region of the tadpole
in Figure 5.8 on page 70. Three toes are clearly seen.
A sketch is shown in Figure 3,10 below comparing the three-toed
stage 57 hindlimb to the normal stage 57 hindlimb.
Figure 3.10 Sketch showing the hindlimb from Figure 3.9 above
* compared to normal tadpole hindlimb.

In Figure 3.11 below an enlarged hindiimb of a normal stage 57
tadpole hindiimb'is seen for comparison to Figure 3.9.
Figure 3.11 Enlarged "hindiimb region of stage 5? tadpole
to show normal hindiimb
These preliminary results provided the basis for further exhaustive
experimentation. The hind/limbs were studied in greater detail by the
more refined method of cartilage staining (see section 2.1.2.2.) and
the limbs were analysed in detail in section 3.2.
3.2 Effect of 5-BUdR and 5-FUdR on tadpole
hindiimb development - a detailed analysis
The tadpoles were immersed in solutions of 5-BUdR and 5-FUdR* which
were renewed weekly (see section 2.1). The tadpoles were staged and
studied each week under the dissecting microscope, in order to
detect any deformities in the development of the hindiimb. The
progress of the deformity was traced until no further deformity
appeared. Subsequently the tadpoles were killed and the Hindiimbs
stained to make the cartilage clearly visible (see section 2.1)

These stained hindlimbs were photographed. The photographs were
analysed in a semi-quantitative manner, according to the degree of
the deformity. A penalty number was given which reflected the degree
of this deformity.
In order to attain a norv-weighted figure (overall) a measure of
the degree of deformity with regard to the temporal aspect of the
formation of the deformity was adopted. In this approach a score was
allocated which reflected the absence or magnitude of the deformity.
For convenience the developing hindlimb was classifed into three
stages :
1. The most mature limb (adult limb) in which the digits are
clearly visible and demarcated and all the cartilaginous
elements as would be seen in the adult limb are present, Stages
56 to 65. (See Figures 3.IE and 3.15.)
2. The medium mature limb in which the cartilage elements are not
clearly seen but demarcation of toes was visible. Stages 55 to
56. (See Figures 3.13 and 3.17.)
3. Immature limbs in which only a paddle was visible. Only
deformities in shape could be detected in these limbs. Stages
54.and younger. (See Figures 3.14 and 3.18.)
In this approach each element was awarded a point of 4 and from this
number a penalty of 2 v/as imposed on a deformed element, while a
penalty of 4 was imposed on an absent element. (Toescould be visible)
3.2,1. Adult hindlimb -
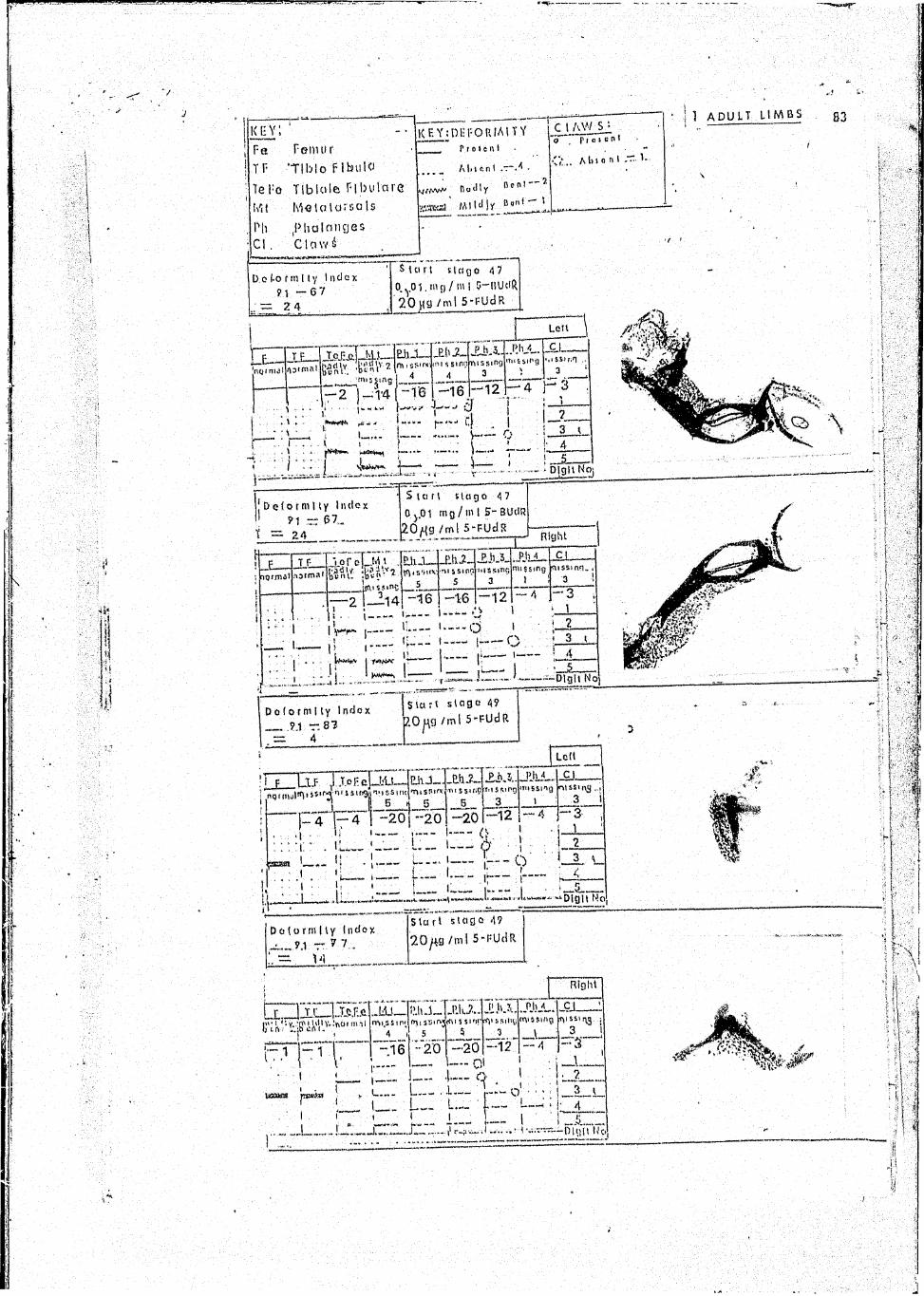
In this limb according to the point system above a total score of 91
was accumulated in the normal adult limb as follows :
Femur 4 .
Ti bio-fibula 4
Tibi ale fibiale 4
Metatarsals 20 for 5 elements = 20
Phalanges 56 2(2x4) + 2(3x4) -I- 4(4) * 16 + 24 + 16 = 56
Claws 3 (1 per claw)
Total 91

In a typical deformed adult limb one could calculate a penalty
index as follows :
bent (4-2)
bent (4-2)
mildly bent (4-1)
two absent (20-2(4))
digits 4 and 5 absent (56-4(4)-4(3)
Femur
Tibi0-fibula
Tibiale fibulare
Metatarsals
Phalanges
Total
22
3
12
28
47
This indicates approximately 50$ deformity. '
To illustrate these analyses more clearly a series of graphs
has been drawn alongside each limb photograph, next to the penalty
index analysis. In Figure 3.12 below a normal adult limb is shown on
this graph and below it a typical deformed limb as described above
with a penalty index of 47. ______
Deformity Indor.
= 91.
Kj:Y;
Pe FemurTF 'Tiblo F ibu laToFe Tibicile F ibu la reMi M e ta ta rsa l sP|t .Phalong-ecCl .Claw*' .
K IT T p e p o r m IT YPro lent
„'‘j Abiftnl — .4.*-.- M.svv/ B ad ly B tn t
fracas M11 <1! y - & '■<'' '
T V AW S •o PI ei crvl
A b ie n I
j_p .!IE-Wtmslnormi! rtotnui
i
• ; ’1 „ , i ■
J ? 1 'X -C.|....HoiVrM
’ • 1•
I !1
3 JX-. m l t o
N o r m a 1.1 imb
j D c ( o r m11 y Index i » 91 -11 :' = 69 '
noffnfll >,;"»•>* rnr/m 5 ^|{!iVlV| fei)«Lv Pm s
................................................. W ,
f 1bent?/ ;—.
1 teswS!
,4„j - 6 1-2 — V -
— r ! : ■ t \»wim■■»«» | Mtwu
....... i !, - + *- M * f t
sinpj,
JL__
4
Ulyii No
D e f o r m e d l i mb
Figure 3.12 Gmphiaal roproBoniaiion of novml adult
Hindiimb and a typical defonned hindiimb

3.2.2. Medium mature hindlimb
In this stage of limb development, although the metatarsals and
phalanges were present they were not clearly demarcated and could
not easily be counted. For this reason penalties in the distal
region of the hindlimb were only given for the presence or absence
of toes.
According to the point system a total score of 32 was
accumulated for the normal medium mature limb as follows :
Femur 4
Tibio-fibula 4
Tibiale fibulare 4
Toes 20 (5x4)
Total 32 •
In a typical deformed medium mature limb one could calculate a
deformity index as follows :
Femur 4
Tibio-fibula 4
Tibiale fibulare 2 slightly bent
T°es 12 (3x4) 2 toes missing
Total 22
This indicates approximately 33% deformity.
To illustrate these analyses more clearly, a series of graphs
has been drawn alongside each limb photographs next to the deformity
analysis. In Figure 3.13 below is a graphical representation of a
normal medium mature limb and below it the typical deformity
analysed above.

.Deform Ity liulox .32 7~ 0.= 3 2 .. - .
,52 - J= ?.5.
k e y : 1 - •
Te Femur TF ’ T 1 b I o F ibula Tefe Y lb la le Flbulcire Mt M e ta ta r s a l s
. '
K E Y j D E r O t U A l T Y
. P r o i o n t • • » '
............. / i b s o n I . r r . ' t — • -
I W i t H y B o u t — *
v m t ' M i i J ! y - C c n l !
C L A W S i -
o i* r c j o n I — - •
A b i o n 1 -------
N o r m a l I i m b
D e f o r m e d l i m b
Figure 3.13 Graphical representation of the normal medium
mature hindlini) and a typical deformed kindlimb
3,2.3. Immature hindlimb
In this stage of limb development deformities in early limb shape (
were noticed, which often went on to form the typically reduced
number of digits in the adult limb. Penalties were given only for
shape and possible absence of toes judged by indentations on the
paddle at stage 53 only. According to the point system a total of 24
was accumulated for the normal immature limb as follows :
Shape 4 ,
Toes 20 (5x4)
Total 24 -
In a typical deformed immature limb one could calculate a deformity
index as follows :
Shape 1 1imb very bent
Toes 16 (4x4) stage 53 smaller paddle in width i,e*
one toe less
Total 17
A 28% deformity. '

To illustr&te these analyses more clearly a series of g/aphs^
has been drawn alongside each limb photograph, next to the deformity
analysis. In Figure 3.14 below is a sketch of a normal immature limb
and below it the'deformity analysed above.
D o fo rm i!y In d o *. . . . ..2 4 0 „
„S . Sh«pe
T,..ToesST
Wtsiing | 3
s -2
T - 8
Figure 3.14 Graphical representation of the normal immature
hindlimb mid a typical deformed hindlimb
This detailed analysis is summarised as follows :
There are three classes of limb development for this analysis,
The earlier two classes were analysed to show the on-going progress
of the deformity while the adult class was a detailed analysis of
the deformities themselves. This semi-quantitative approach gave
interesting results even though the deformities were not found to be
consistent either with stage or concentration.
The detailed analysis of the limb deformities is carried out in
3.2.5. for adult limbs, 3.2.6. for medium mature limbs and 3.2.7.
for immature limbs.
The results show limbs with one or more digits missing. In the
thirty cases of adult limbs analysed in detail :
Digit 1 missing
Digit 2 missing
Digit 3 missing
Digit 4 missing
Digit 5 missing
23
13
6
16
16
30 cases
30 cases
30 cases
30 cases
30 cases.

3.2.4. Analysis of normal hindlimbs
Figure 3.15 Normal hindlimbs

79i
1 s j iopa
I 1 O 0 s’. 'I,— '
Normal S tage 50
. ■
■t-w
iN.ormal Stage 51
Nqi m. o I Slag o 5 2

T t o o sf-
Normcii S tage . 50
|N.ormal Stage 51
AKtU.iU.flJ S ta g e 5 2
'^psr

k e y :— -------------- j
1C C Y :pEFORMl T YFe Femur _______ P K M e n i
TF ’Tlbio F ibu la ____ "j A b t o n l - t . 4 .
Te Fe Tibia lo F lb u la r e , f B o o ’ l y Bonl 2
Mt M e ta ta r s a l s r r e r w t M i l d l y - C e n t 1
Morm oi. Stoge 54
n Q f n t a l
XLoof mnlX?£a J U _ L £ i
normal«normal
, i1 i 1
ii- 23 \A
>Dl9t( Ho
.Normal S to g g 55
Norm a] Stage 57
JE—
igrm.ii noiin.ilT£_ Ic.Lfi
no f Hint normal,normalJJ.U.norma)
J U j X
r s o / m a j
” p h >t 1 c i .
n o r m a l * n o , r m a 1 ;
r > . - ............................. K !0 .
1 '* *
1 ■ 3 <
n i • !i 4
'Digit No|

KE'FaI F
Fern ii r'T ib !® F i b u l a KEY:DEF0RIAITY
TeF© T lb la lc r-lbulare t Z T * P'* * * "1Mt M e ta l arsa Is jPh .Phctlnngas C l , Cl aw 4 JESSrf
Absent
Dotlly Ml |d ly . n*n< "" 1
B e n t 1
N.ojipg} Stag^ _S8.
_UL_ Jste- P h J I I Jill-2 J- ti'- S- normal norma! normaj
. . P h 4 _
j l f l o V m a i normal
Ifcior ma I,..Stags 5 9
a —Jncitmal
T P Tor-f t (4 I __r t o r n ' s l ■ y o f r n . i l normal
i i
EhJ—normal normal
P h Jnormal
rr>- I*
_PJiAnormal norma)
X " DTgjt^o

3.2.5. Detailed analysis of adult hindlimbs
Figure 3.16 Adult hindlimbs
k e y : - • KEYsFa Pom u r ■
1 t r- •T lb lo f lbulo ......; Te l:o Tlb lo le r*lbu are W>V’w
Wt M e to la rs a l s ?;*kzl, Ph ph a la n g es • -; c i . C1 a w ^ •
P I o I c n I •
Alii nn I
B a d l y ' n o " 1— 7
C t AW-SP | 0 I B 1 *
A In o n I .t : '■
1 ADULT LIMBS 83*
S ta r t vlctgo 4 7 0 01 , m t) /in | C-llUdft 20 jta /mI 5-FUdR
Lt—nQimal
EU—i 4Hannnl •ijilnl' 7
2 J —:T4 ( —IB —16 1 • * J*...... |»* w J.
4 t . , *■«***«* . L... J— ’»'*** i,|' . ; i _ . ’
ill' 2.i» i s smn
A
P.Ji.3..riissii'O
3
PM_.unssii’g
'12
Leu
X ! ______issir.n 3 ____‘
■* 3
i . \
' I
6 6
...... v-
t
0(
. A - .
JLDigit Not
D e fo rm ity indexS tti r I Hugo <1 7 Oj.01 mg / in I 5- BUdt?
l o ]\9 /ml 5-FUdR Right
normalJL jr-
aatmai'i.o I" e I„.M t
t-z'U l4
i----«:
ELb.Ji___ EJiA
' 5-16
mssino 5
_PJl_3_iuS5(H<i
3
- uy-r- j—
wwVWW**,
li___'U---I]___
_ - 1-6 i — ■ 0
-o
-12
I’Mmissing
Imssinn.
3
I 2
o ♦ 3 t, J 4
I --Digit No
D o I o r m I ly IruJo x — .9.1 " 8 1
=z 4
S ta rt stage 49 7,0^9 /ml 5-FUdR
Loft
E z n r i i ^ i U uj HQ fm ilts5• nT.'»s« n«,sSin.
-20
ELhJ—*ru$fl!/x
-2 0
mmi s S»f*P
5
L.M,tit 5 smg
3
J P J l J —missing
X
-20— 0
-12
OISS1I13,3 I
I
U
j. I**
9 ■ V - oi;--- i"
Iu .«*«♦*»»y *Vtw ‘ Digit No
}
D o ( o r m 11 y I n cf o % 7.1 t * 7 .
Stci rt sU tgo <1?20/^cj /m l 5-FUdR
\
1 Z I E P 3 5 S
r i= T - ( ;
Highl
j 5u Z u u J I k £ wp jt j L ;»ni,ssin» •nus'M <<\l S si'll' if)1 s.sidy miosit'o
4 i 5 5 3“ l r ~” : .1 6 "*•*20* — 2 0 - 1 T
♦ _ — — O ' . i
pa*M jI— — 4“" ^I___ L--- • -U-r
Is z : ■r r-?“
o
lyssiry ; 3 i
“ 3_ j _■ J L ._ L _ l ' t.
--01 git No
\
*J

k_p y ; . .
Fe Femur T!: *T|blo N b u la TePe Tib la Id f l bulcire Mi M o I a I a r s a 1 s 4Ph /Miolcmges C l. .C la w i •
l( E V : D E |! 0 R M I T Y ___P io i on i . ...
AbtenI .rr.'fv/vWr'’ fiotlly Ban I *“2
jssass M11 <J | y. I iU —* i
Deformity I ndox 91 “ 73
' ~ 18
S,t a r t . s I o g 0 4 £ I0 i|cj/m l (>•* PU ilR
X L___________ Top.nl M i__
^—«<»»’ sjiAg
*2 i-16
Left
Elut.tnissiia
4'16
I:
---ll_ . . >11----J.
Deformity Index .9 1 " 7 3
: 18
>«i s 5in'j5
i ’J 'A .ttlssing
3
- M J . .ttifS isis/1
1, c . l _rn ss> rig ,
2 i“ 20 -12 — 4 — ST , *■
/*: - » 1WW***. $ • - ■ ■ - j. ?
■ I . J L LI . . . j 4I
r t— L ''' •
j 15
f
nil
Sl a f |. stage 4,5 j ,0p « / m l 5 ~ f U ciR I
x Z J l ^ i X M j J ^ Z L C u i -lnQiniag’c'{idt!r’l>“i'iV” *B«nly2'H,5S"ltn1'Sam9
Wght-Cl .hissing
s i V. ,
,1 1: 2
3 t,...4 .
Digit No!«?w;wtpw*D e fo rm ity I it d o >c
?1 ~ 3 1 .60
S t a r t stag a 43
° ) 2 ^ g / , n l 5-PtJdR
JL JJL Jiulcl-Mlocjfftiat <roim,iI g'gjfl* Wfln.j Ju,j JihX, f* It 1
«> i 5«n*.tiissidii2 1
—8 - 4
D 0 1 o t m 11 y ! n d x . 5>,1 .“ 4 6
S 1 n r t tu ifja /IS 0)2MS/ml 5‘PUr)R

- ■. ..J L J .L - .-lJ !^ ■....... . U
X K E Y : DH l: 0 R W ! T Y
p ( C, 10 It I . *
4 .A b t c n I
iv y ;(: o " FemurTP ' Tlblo F i b u l a ^Tero Tlblcile F lbu iaro L j , Dc.diy Ben' ' ~ 7l
~ i <-i ( m rr.Cl I S ' iffflEKMl IA l i ( l ! y - l ic n l_____ . 1
C LAW 5ji" I' I C1 0 n <\V.. , A b 1 O f> l — I

k e y : ' *,Fe> F e m u rTl: ‘Tlblo F ibu la , to !:e T Ib lc i l e P f b u l a r eMl M o t a t a i ft ti I sf’l) (P Ik i tangosC l . ,CIaw
D o { o r m 11 y Index 91. — 43
= 42
K E Y : Di- FO R M IT Vf‘ / o i o n t
. . . » A 1j i o n ( c tA ..VWV*1 Hail I y Bon I “ " 2
3SKTJ M i l d l y Bcnl — 1
C LA W s ! o I' f 0 I n n l
U A b t o n I ~ I ,
86
Start s t ci y o 4 8 20 US/ml 5-FUdR
■ r X L03 f fU«< I
IAE js. J jLL- fib J.J.P.I).?..normal (iSS'iyj 8^HiL,«'lssin3
__ m I i MI«V \ 3- Il i Q . l _ J r
I MWy | --- * |• • I
t i ' Xnussirg
3 1 2 ~
Lettf»M_.
A
£ 1 _ , rjijsing.
■ A>
Hanoi | ttmnsr 0IiT
J
I
J ■L.5— .- D i t| 11 Ui>
D.atorml ty Intlcx 9.1 — 4 9 „
= 4 1 .
Stcirt s l ct g ft 4 ii 2 0 /Ag /m l 5 ‘ PU d R
____ —- . r
. ■ ! 'I . . * i * * > ,! * :
i ;= 1
:•
1
! . ■ - |
“ 11 - 1
U l - . J : ....... 1
D o l o t m l l y lnc>.‘ )- .9.1 ” 57.
"- ■T"'1 ; , 3 4
Right
i t t A ,t n !VU .1K'-"' 1 ’ I «Mllmi lljty Vt fll 'l .3____?
-12’I4 tntJSina ’
I H i x <
Ph.-t\ l Hints ftilssing
2 1~ e — 4
(Y>7rA
|*tyw>v
t A f * .....I.■*"**. I —
1-Sl.p <1 u'aijD tj, \ i l i\d /ml S-PlicJR! ’
..5....♦Wan No
iQbn.ilX L J tf.&i*. hw t at*
t . I
H2. i t* >;yv.*v Ifewww t
“CJblL
3 1 !
z::n :=T “
I jmnw ; rmsm ]»* 1I ), ©ass* |
o
Loftii
n>«ma 3__
~3
JL.
•o 3 _ t
..Ij
& o f o r m 11 y Index5> 1 — 56
.. ~ 35 *
■ fcAw'.J'fct ^
„S la r t clugo 4<52 0 fU) /ml 5 -r-UdR
\yrn.>»ii j : „ .
di 6S r-'U5S\V(— Ip M !* . . « * M ^ U i~ .
? . . "1 2 ^ 1.2 ! ! z l 6 a;- ;■ * * I >A*Am | ft8tf5W»i | — f . JI /VVWv . laftocsK ( * I ’1 1 1 r' . 11 . Bmnvu 1 hBSB9
| w — ' ^ L _ ,— : 1—
I ...1“ '" irr*:.!— ,.i;.... J : . . :
A L l .flissmg
JLr - V
C — 111 rii nrnight |
.Ci.Oiiiii^g.,,
3 I "3
J ^J i-.iin, , 1, .,ft,
!— ?t—
:■ A..
: ()|tiU No
>

k e y :1
r a Foin 11 rT l: •Tlblo P l b u U )
!To I:g Tlblct lo r i b u l c o o
■Ml M 0. (ct I a r s a Is
n (Phc i lc inoosc . .C l o w i ,
K E Y t PE r -OKMI TYi ,i;. p f <r t c n i .
A b ie n l --‘.'J -
nodiy 1,0,11'P5?»!) Ml NUy. tlent r
[Do (o r m 1 iy Index ? j - 2 6 ~ 65
C l AW S[■ r 11 f n I
. ,A 1)1 «i.n I
TF To Fo M t
■format piss mg )
Ph1
fni ssi nq l
Pit 2
miss int I
Ph3
nissmg 2
S I 0 r I s I a g q 4 7
0,01 mg/ml 5-BUdR 10ug/ ml 5 - f U d R
LettPh ‘1
■tissing\
Cl
Nssmij?
I i 2i i
I ,
D o l o r m l t y h itl«v .. 9.1 ~ ,2,6.
.:■==• 65
normalTP.
nstmalTcFe
riOftrlfilML.nvissin ■ !
5 l a r i via go »1 7O.JM.mtj/m l.ir-fludR
5 - F U d R *
‘t 3— JG Z" 1 BfgTl No
Right
EJxJL.fni53trr*
£1) J2„
\
I h.5,.r.tSRintj
i8
Ph;l..missmij1
£L
<33Ifr
•V
.I/—1;i.
D e f o r m i t y Index 7.1 ™ 26
JL
JL
■ * 65JLsssas „ . * -
r Y i L L j I l *rxjfmai fuimnt riormat
i ; ; . j : 1I ' - : . - L _ _ i
Sior l . Mage 47“ ' mi 6— ru tiR
Digit No
'Xcft ]
m 1.1 Sir. 1
i; 4
eiurniss1 miitiijg 4 1 - 5
I l i X(01l iSing iflusi«<3
7 <- a — 4
i!L M
1 i
r1 lln 0
% I.,
!
J— J—l
I ifWOHj
5 tor i Mago 47
iO^g/ml 5— ru UR
Digit No
jgr"**
87

—>»t—riV.mxn.0".
k k y : 1
V " l:0im/rr r ' TIb 1 o F ibu laTe Po Tlblcile IMbulareMl M o I a 1 d rs a 15
fJ h h ci 1 a n {1 e sCl . .Claws
KGYfPE r ORMl TY----.--P t o i on I
. _ A b i o n I •tt.'S
WAV* U tiJly B o n l“
WKEtf M l |tl )y . (lent ~
D o ( o r m I ( \> I n tl o x ...?1 - - 3 4 . . .
< = 57 _
C LAW S i .O' I’loionl . . .
w '.. A 1) i a n I ]...
S iu n Etcirjo 5220 qg /ml 5-PUdR
88
[ U f t
X X L XsJLa.ftO(rn.il
,,.L.
iI <_ I !I (___
Mj . i ..
, M j . J R l i J __ tM> 2_{H}aivi:m,.«.ra«r-miSSir«. -
I
6fifty*
8(THssir;^
£ i'A .missing
1
'JU L l ,ITnssing
14 j
I. - d
Cl__rt 1901 nQ
3~3 " J ____
ii •« foam ,
j I
D e f o r m i t y Intlox . 30 .* = 53
’ 3 i . . A
_5■Ltlcill (4oj
S (a .r .L s ttig o 52 2 0 ju$ / m 5-PUdR
nqimaiTF
rdirl&l-U‘J>
cie»f m«J
I Right
tvn s1 - .7l»ra«s(} tuannr L
H J3 L . 8
a.- 8‘
£ k L-t>, ssmg
?a
njtSSing_ J _■— 4 “
Iod
i •O'
' | i; ., i... •'**• ja* ■ <-»*,, ««*» >t
D e f o r m H f In t lo x „ ,9,1 - r.4 5
: = . 4 6
fltSMriJ,.;3 t
•3~~-1___ J23 t
4— , I- • -1
--Dig ,S"fo r i ,s I o g o 52 ,
2.0po /ml 5-FUdR
ri>qirua<
X L
1!i * i!
j .
mii iiijiji
- 12-' i--~l/vwv j- |/Vw !*■ /W*A* , •
I AVMVt
E X r t T J J i l a ”lic'nl't mtSMf-g
sj-
s
miiSipmisfioji
6 1-14 ‘
D eform ity Intlox . P.1 45 .
'= i 46**>«» **>»
ngtmolX L ™fiormAl
i^.llanor mrtl
f-
|V1 <11.V, is,M(>3|m i iM ^
■12 i“ ~6
V., I .
| AVVA 1.. ... .| j»-—_I tow * I*——_<
I /VVN\ I
lk*wv l*» j—* W«#
S iu r I .s ia^ o G2 20 pg /in 1 5-PUdU
Ph, i iMt iT l j ’T ?• n i H v . h n d i u 1 1
2

I*JI
:
f
k e y : I — —
Fe FemurTF ' T! b I o F1 b v 1 clToFe Tlbldl-e f:|butar-eMl M c 1 a l a i s a i sn ,P h a I a i] (j-e sCl. ,C 1 ci w si
K 1: V : DEFORM I T YI't o 11- n I , ...
___ A b i on I ~ t A ... ,
HUA^ Dudly Beni--?.wasa Milfljy. &»«< ~ l
Detor l i i l t v Index S>1 - 3 9
— 52 ...
_E— JJ ... ..UP.Uu.il floim
unsung lOisiigj ! -8 1-6
i .
2 -- 1 0 1- o... «.... rr-k.. . __i,-
C LAW 5 : © Pioicnl .
A b t o n I !.
89
S.tci r t s l a p j o_ 0 ... ID uy / ni I 5— I’U lU!
left
ii •I -
. , VIV/W1 I
ftQ
j : j .__rniMif'g n < svt ng.,...1 ' 3 i
“ 3,.J.__j
■ ; . __■ ; 3 v.■ . . A
j | /iiwm, | ftsw. [ ««*(\
D oto i ni I ty Index .9 1 .” .39.
• — 52 . ..
MoS.Ut.r t „ static-. 0 .
1 0 ^ ,/ m l 5 ~ TU ilR
l i _ Tr-I.uf| Ofmil
.: i.
f •lL _1?
laiJE.uZDiD7-hastyn.tvdiy iBtmi rf.htntT /;,benr J '
“ 8 I “ 6 1-10—__J ^ ... f*v_____i _____ u,
C)ii
Right
S,J>A. Mi •(oi**nr«5, i t l lS H '.g
? f
— 8 — 4
I i ' , i:
* t i i
• o i
t f 1 «•
Deformity Index .9.1 - 30.
61
S fa.r.t . slag c. 0,.10;.ui / m l S - P U t l R
oztrnz f ncm.n l & t l f e >•»»**^ *** 91 n>t s5«r mi v s»ucJtfSv & j >nq
. 1“ 7.6' j-S 1-6 1-5>•1a
. ; . i.
D e Io r m11y Index „9 ,1 — .3 0
.-61.-
Lo't
M )£ .. .CJ_____n ssmg ,
1 ' 3 !-■4 - 3 ;>. ...... r '. . . . • 7
•O
: 45•■* ...J•Digit No
.a cart , sioqe. 0 N/Jici / m I S - r - U i l R
J:._nyuuolj_r__(ISffi’fl asJLe.not mn\
i ' ‘( ’ *
R'Ciw
IL.
1. 1,1’h 7 n » T s IT h 7 " tiflilty, ywdiv. 'M'aoly. ’—bum.I Ibuni i bunt.I p.irtij KHiMmn*yi j. s i * a | ^
6 I 5 1 ’ 5 I 4
— ' ~~_i~~~~ I l ___|._____j___ _■; i .1
j Wift | fWVW’
C L ._ ..
3 !:'3~ ”
!__]___ 'L 2 _
<1 ”.... ___
••• f1 i fj i! Is'n
' ;L

khy; .f'« Fem ur Tl: ' T l h l o F i b u l a
To Fe> T lb la l f t F| bular-e Mi M o l a ( a re a Is Pli JMia lcuu je ' j C !. ,Cla\vi * •
K E V i D E F O R M l T Y | C L A W 5~~— , f*( o ion I . .
Abienl “ <4____*W*Vtv 11 a <! I y Bon! 2
IlSEtSEl M t l d i f Si oil I ]
> P ( c m n l
(1™ A b i c.n I 1,,
90
D e f o r m i t y I » cl e x ?,1 “ 34
.57..
S.tci u .,.s tago 52 , <0/.^/ m l 5- FU ii K
■r Right

' 3.2.
Figure 3.
3. Detailed analysis of medium mature Hindiimbs
L7 Medium mature hindlimks
\
f
ft

2 M E D I U M M A T U R j
k u y :t:-e Femur TP 'T lblo Fibulu ToFo Tlb ia le Flbulcne Ml M e t a t a r s a l s
Dofo r tn ( ty Index.12 ~ U
J'.o. .. .
KEY: DEFORMITY ______ I’ i •> i o n I
_____ Mis cii I. 77A —
tloJly Bont-
355tifd fAi Id !y* ,u‘n* "
S ( a r i s logo 4 5 0,01 mtj/ 111)
■ ’s-nucJR
Led
.Deform 11 y Index .32 .-.12
_“ 20_. ■
lr t fnptnjisl normal no* mo-*
• { . .
S l " t l s t ago 43 O./.OI.mtj/inl &-IlUrfRt O/jljng/m.1 5"FUdR
Right
....... l
t ' T
D Cto tm t ty i title x,5 2 J 3
19 ._____ j i . J - 'nuti»o)S!c}i i.tnoK,>!l1
S t a r t Mc)t}0 4 7IO^ci / ml C—_ru cfl-f
Left
„De(o rm I t y Index i 3 2 ~ r . t4 .! = 18 .
S t a r t t i <i a o 4 1 lQ^tt/ mI 5— I U ill?
Right
J.ef «miid I y.
e,n i - - mi :• s i 3
t.J
n

2 M ED lUfA M ATUR E L I M B S92
k i3 y : 1 -
F'tt !:c mu r
TP ’TIbIo !:lljulci
To no T I! j 1 a I e l: l b u l a r e Mt Molctlcirsuls
D clo r m 11 y 1 ncto x . 1 2—12
. . 2 0 . ......
K E Y : P C i : O R M I T Y. ™ ~ , (’ t '■ i (i n 1
...... A lii on I. rr.4- ,CJ I y B o o t “ 2
SH5ST3 (A i I i! ! y ■ I1 cn1 ~ '
; S t ci r i stciga <13 o.,.01. mg/ml &-«Ur)R 0/2 / nil 5-FUdR
Loft
•' F J T E _ „ J S I A
nsjim al r>of m al
* *" I
J t ' * .
J Lit? r rua I
T Fnotma.1
JjgJLeno/mM
J jLL-L o..<r|(.siin<r
_ 3 _ ?-12
' _L
___'Oicjh Noi
1
,D o lo rni H y I ndex . S 2 ,~ .1 2
*i=. 20
S t a r t stag a 43 O.j.Ol.mtj/m I 5-SiUtlR) 0,'Zjt g/ml S'FUciR
Right
j :r_ . i
Deform l l y 1 ntlox .3 2 .- 1 3
— 1_9 _
_L__ JX r j_le.6.eHutr’iolffco r .lrr>0 "» ° I
rt i -
t il!
_M £ „J.. _____
. . . ________ : 3 t
— 'Digit HoS t a r t stcigo A 7 ‘
10jut,/ m l d— ru uk
-,1
m a J-% S f tu,3 1 2 ~
in « '3 (
‘i JL.feaaeagt l !
J . ______
, 3 . , 1
4
Ojgit No
Loft
P e(p r m It y I ndox 3 2 ~.-.14 .
18
M o i l vt( igo 4 7 10 f.\g /_m I 5 *- )■ U tl f!
L . Riohl
1 1 T.F, ( ( ( J | y.ni)itnalb-1" o l„...
■J.ft P cv 11(11V. > on t,L.
1 .
mi** inn’'3 i*12
i • ' i .
| fcw»?fc&i I —)
1 ftKOtfUl
! L _

ti
B3
ii k e y :
Pci F emu r TP T lb lo i; IbulciTe Te Tibialo F ibula re sodiy non 1 — 2Ml M e t a t a r s a l s ' " ~ ‘ 'Deform 11 y I nde x
. S 2 ~ .1 3 = 19
l( E Y : DE T OR M I TV 1* I 0 * 0 !• J
..... A ti 10 ri I -~A6
M ild j y B cf> * 1
S ta r,i ,.s tog 0 52,,20 /ml 5-FUdR
Left
1 , r... TF 1taUeM-yu.ojmisl ■aJ-. rr12.
r -
J . ___X __3 (
■ .. ;_j Mo
u d o i m 11 y I ncicx , . ,52 8.,= '24
S t a r t staao 43 Ot 2 pcj/'ml 5-FUdR 0- ;0Lm y/ id! S-ftUcJR,
Left□
i-LL...
natmiii-sL-
l-*8
: I;,_ t ______ [•..(L.. f-
JLX
*-*v?___‘intjii No
,D o(o r m 11 y t ode x .5 2 r : . 8
24
Start tuigo <15
0*2 fig/m l 5"FUttR/ml C“ BUtt«
Riflht

34
k e y : ‘ ••rft Femur TP ‘Tlblo Plbulo
;Vol;« Tlbiulo Pi bulare •M» Metutuisab
K E Y : D E F O R M I T Y„ „ P r c i o n J . .
. . . A l> i e n I --Otttily I ^
W 5SS*' M l l ' U y . Dcnl - I
D o to r m i I y I nciox .5 2 .”10 ...
.= 16
Sta rt si ago 4?2 0 /mI S-FUdR
Left
L E LTEjJe.r.eBan |V--.no/n><ll
-2t
V*rt«s ftVn
T JL
Dcto i m I ty Index .5 2 ~ 7
= 25
Sta rt s t age 49 AiS /ml 5-FUdR
Right
“ ■-saw-srDole rm I t y Index
, S 2 — .6 „= 26
s ! a r i 11 a f| o <1 y 2 0 j.(ci /ml 5-FUdR
u, Left
! - V ‘ F J I F I TeFff
,-1 r-,1 .
' I
\tk4*. i-..
I J ._ J . _ _ L 3. »
rrulitTyj j. on i,,;..j ;.lT .fo il
■porffl.il
'*1;■ it.!-1 ....... 1 . . . .
Jslfc.fftissmi ■ I > - 4
l J U .
1 A
igps*
1 J ...*i ^
L l — . .
!0TrjT?Ho •
e
D ofo t m I t y 1 iicJ dx , .5 2 “ .6 .~ 2 6 , . .
' S t a l l i.(ao o 47 '20 }tg /m 1 5-FUdR
.............
Right

K
k e y : 1I'o Femur
T F ’ Tlblo r-1 bu ln ,To|‘e Tiblcile F ibu la reM t M o t a t ci r s ci I s
K t: Y : D F. r 0 R / A1 T Y, I’ I 0 I 0 ft I
. . „ _ A ti ( o n I , T , ‘1 . ■
yWANV Bvdly I ' " I I1— 7
msms* i/ .lN ly - CH' 1 “ ’
D o l o r m i l y I ndex ..,3 2 . “ 14 .
. ~ . 18 _____
M.i.•>QC
.1 i..
S Ui r t s ( a tj o 54 , ; ! i ° | i i j/ m i 5 - r-u.iiR
712
D cto
.. .. "i
r
rm 11 y 2 -r .1
r r |met «K-S'.y *
t : * •~2 ‘
* ,J i
i: i 1l ; _ 2 ____i • 3 l
" ™
JliCLlU iQ1 ' '
S.t o x 1 s Ui g o mi 5-
- X,,,, „ .ru ciR
' '-, * 0 |_ ___ i
I -.12 r , _ _ _
lilL
WrA>(V>VrA *-.3,.(
■*01«11 Ue
° o ( o rm I l y I ltd ox >32 — 4.
J = 28
SlOri slqyo/jp20j^g /nil 5-FUtir
* ‘ " ■* * ■■
y . J r__ -Ur.c.no, my.
'.V,v i:;. j
. :v 1:::■LLL 1
.::;: I.; ■: r— j . . . . . . L _
D o(o rm I ly I ndox 32 — 4.
— 28
* "—•Olyil J\'o____
S t ci r 1 1 1 ct p u 4 0 ,20||g /ml 'i-FUdR
'Urtwir„ F __ JJF .-.
----- ----- -—------
fiyinml rwrrfltti mifiTify ; ..1 1 * 1
I, " T ~ “
■ . 1 , . ; ~ 7 | ,1... '
• ■ .. . [ w ^__ , U lL
-J. J' Digit Koj
eft
H"- 111
Hight

k e y : -Pe Femur TP 'Tlblo F ibu la To Fe Tlblcilo. F lbu la re Ml Me let tars ci IsD q I o rm 11y I ik I ox
, 3 2 ~ 4 .= 28
IC E Yi PEf-OKMI T YPromnl
A b t on l .rr .4 -
Radiy bent-"Mildly H on I ~
Star t sUig o <1 8 2 0 (./a /ml 5 -FUdR
Led
D o 1o r m 11 y I ndex . 5 2 — .4
= 28
Stor i s l ot ) o4B 2 0 ff j /r,i I 5-PUdR
r
F T F J S lE a MjL-.iQimai fO«» I rvorm,\l r\ i,j »iivj
- 4 j
■' ' '
---- - :
- ch r r . J
x
DHilt Mo,,«-v— J —

Author Christie C AName of thesis The effect of 5-bromodeoxyuridine and 5-fluorodeoxyuridine on differentiation and metamorphosis in Xenopus Laevis Tadpoles 1982
PUBLISHER:University of the Witwatersrand, Johannesburg
©2013
LEGAL NOTICES:
Copyright Notice: All materials on the U n i v e r s i t y o f t he W i t w a t e r s r a n d , J o h a n n e s b u r g L i b r a r y website are protected by South African copyright law and may not be distributed, transmitted, displayed, or otherwise published in any format, without the prior written permission of the copyright owner.
Disclaimer and Terms of Use: Provided that you maintain all copyright and other notices contained therein, you may download material (one machine readable copy and one print copy per page) for your personal and/or educational non-commercial use only.
The University o f the W itw atersrand, Johannesburg , is not responsible for any errors or omissions and excludes any and all liability for any errors in or omissions from the information on the Library website.
Related Documents


![o -3 o o c: g: o a a a X cn c: a a o g cn tri Z o o: 1.0 N ...€¦ · o -3 o o c: g: o a a a X cn c: a a o g cn tri Z o o: 1.0 N p: o Bau-Schalldämm-Maß R' [dB] I I I I i I I 00](https://static.cupdf.com/doc/110x72/60823909957b3b3f5a030db3/o-3-o-o-c-g-o-a-a-a-x-cn-c-a-a-o-g-cn-tri-z-o-o-10-n-o-3-o-o-c-g-o.jpg)








