Burst response of the Damselfish, Acanthochromis polyacanthus to visual stimulus ABSTRACT: Burst performance determines the ability of fish to avoid predators and capture prey, especially in their larval or juvenile stages. Since most coral reef fishes are sedentary, burst performance has the potential to determine survival and abundance of adult fish within a habitat. Previous studies on burst performance of vertebrates focused on commercial freshwater and marine fishes. This study focuses on the burst performance of a coral reef fish, Spiny chromis (Acanthochromis polyacanthus), which is abundant common species on the Great Barrier Reef, Australia. Furthermore, this species of damselfish has the unique characteristic that it does not have a pelagic larvae stage, but rather hatches as a juvenile. This study examines the effect of early ontogenetic development of juveniles and the effect caused by batch variation to burst performance. This study was done by identifying the pattern of various variables such as maximum burst speed, directionality and responsiveness in burst performance. Parameters of burst performance examined included the maximum burst speed, mean burst speed, directionality and response latency. Visual stimulus, a looming silhouette triggered by a pendulum was used to imitate a predator attack to initiate a startle response from the juvenile. As predicted, larger and older juveniles had higher maximum burst speed and mean burst speed. Both of these variables were highly correlated. Generally, juveniles showed improvement of burst response variables such as higher maximum burst speed, mean burst speed, directionality and responsiveness as they grow. However, no parental effect was found on burst performance variables and there was no ontogenetic effect on the response latency of juveniles. Interestingly, the response latency of the juveniles was weakly correlated with their maximum burst speed. More research is needed to clarify the relationship between the kinematic and sensory receptors during the burst performance of this species. This study shows that the importance of burst performance behaviour during the early life stage of a coral reef damselfish in predator avoidance. KEY WORDS: Juvenile fish – Coral reef – Burst Performance – Growth and Development-Batch effect

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Burst response of the Damselfish, Acanthochromis polyacanthus to visual
stimulus
ABSTRACT: Burst performance determines the ability of fish to avoid predators and capture prey,
especially in their larval or juvenile stages. Since most coral reef fishes are sedentary, burst
performance has the potential to determine survival and abundance of adult fish within a habitat.
Previous studies on burst performance of vertebrates focused on commercial freshwater and
marine fishes. This study focuses on the burst performance of a coral reef fish, Spiny chromis
(Acanthochromis polyacanthus), which is abundant common species on the Great Barrier Reef,
Australia. Furthermore, this species of damselfish has the unique characteristic that it does not
have a pelagic larvae stage, but rather hatches as a juvenile. This study examines the effect of
early ontogenetic development of juveniles and the effect caused by batch variation to burst
performance. This study was done by identifying the pattern of various variables such as
maximum burst speed, directionality and responsiveness in burst performance. Parameters of
burst performance examined included the maximum burst speed, mean burst speed, directionality
and response latency. Visual stimulus, a looming silhouette triggered by a pendulum was used to
imitate a predator attack to initiate a startle response from the juvenile. As predicted, larger and
older juveniles had higher maximum burst speed and mean burst speed. Both of these variables
were highly correlated. Generally, juveniles showed improvement of burst response variables
such as higher maximum burst speed, mean burst speed, directionality and responsiveness as
they grow. However, no parental effect was found on burst performance variables and there was
no ontogenetic effect on the response latency of juveniles. Interestingly, the response latency of
the juveniles was weakly correlated with their maximum burst speed. More research is needed to
clarify the relationship between the kinematic and sensory receptors during the burst performance
of this species. This study shows that the importance of burst performance behaviour during the
early life stage of a coral reef damselfish in predator avoidance.
KEY WORDS: Juvenile fish – Coral reef – Burst Performance – Growth and Development-Batch
effect

INTRODUCTION
Burst performance of fish is an important mechanism for predation avoidance and for
obtaining food in the early life stages of fish (Eaton and DiDomenico, 1986). Previous studies
have found that the food availability is less important source of mortality during the larval phase
than predation (Martin et al, 1985; Paradis et al., 1996). Hence, predator avoidance influences
fish survival and for this reason burst performance is a key mechanism in population dynamic
study. Burst performance is also often known as escape or fast-start response to external
stimulus or threats (Webb, 1976).
Body size and ontogenetic development of fish have been proposed as the major factors
that influence burst performance (Bailey, 1984; Bailey and Batty, 1984; Blaxter and Batty, 1985;
Margulies, 1989; Cowan et al, 1996). Body size of mackerel (Scomber scombus), white seabass
larvae (Atractoscion nobilis), and fish length northern Anchovy (Engraulis mordax) were found to
determine the burst swimming speed (Margulies, 1989; Wardle and He, 1988; Webb and Corolla,
1981). Age is correlated with size and ontogenetic development and these are also known to
influence escape performance. The development of sense organs (Blaxter, 1986, Blaxter, 1988;
Blaxter and Fuiman, 1990; Blaxter, 1991), visual neurological responses (Anderson, 1988) and
locomotor mechanics (Hale, 1996) all seem to improve the fast start performance of fish. Size
and age correlates with larval survival, such that smaller individuals often have a higher
probability of dying (the ‘bigger is better’ hypothesis, Miller et al. 1988). There is however,
considerable debate on the general applicability of this hypothesis (Litvak & Leggett 1992;
McCormick & Molony 1993).
Parental effects can be profound during the early life stage of teleosts fish (Kamler,
2006). Batches of juveniles from different parents or clutch effect showed different behavioural
performance for red drum (Sciaenops ocellatus) larvae on the burst performance (Fuiman et al.
2005) especially during their early life stage. However, previous field studies on batch effects on
larval quality in term of burst performance were highly confounded by environmental effects, such
as water temperature (marine temperate fish, Batty et al, 1993; freshwater fish, Lyon et al, 2008)
and timing of spawn within a breeding season ( Fuiman et al, 2005). In addition, nutrition levels in
the water or food supply after hatching also influence the success of a larval batch (Yin and
Blaxter, 1987). The present study investigates the effect of different parents on the larval burst
speed.
Other factors, such as previous experience, phenotypic plasticity and rearing condition,
influence the burst performance of larvae. A previous study showed that the reactive distance of
the zebra danio increased as a function of past experiences (Dill, 1974a). It showed repetitive
stimulus to a single subject will cause uncertainty or noise in burst performance. Phenotypic

plasticity is another noise factor in behavioural study (Wiedenmayer, 2009). The rearing condition
variation such as temperature (Batty et al, 1993) and food (Yin and Blaxter, 1987) can cause
developmental change of larval. Herring larval, Clupea herengus had higher burst speed in higher
post-hatching environment temperature (Batty et al, 1993). Well fed marine fish larvae such as
Clyde and Baltic-herring, cod and flounder had higher burst speed than others (Yin and Blaxter,
1987).
A fast-start burst has three kinematic stages (Anderson, 1988). In stage one, the fish
bend their body, orienting themselves away from the stimulus and making a profound tailed flip,
but the center of mass of the fish remain stationary (Foreman and Eaton, 1993; Hale, 2000). In
the second stage, the fish display a reflexive tail flip resulting in the rapid propulsion of the center
of mass (Foreman and Eaton, 1993; Hale, 2000). In the third and final stage, the fish burst and
cruise alternatively away from the stimulus. Other fast start performance variables have included
the burst distance, speed and acceleration, latency of response to the stimulus, the duration of
each kinematic stage, and the change of directionality (Domenici and Blake, 1997). Generally,
smaller fish larvae had shorter duration of the kinematic stages compared to larger fish (Domenici
and Blake, 1997; Hale, 2000). The directionality of fish during burst performance includes the
turning angle and turning radius. The turning angle of fish during escape can span about 180° to
either side of the fish (Domenici and Blake, 1991; Foreman and Eaton, 1993). On the other hand,
the turning radius is important in predator prey interaction and it is always in circular path around
the centre of mass of the fish (Webb, 1976; Domenici and Blake, 1991).The turning radius was
found to be highly related to fish body length (Webb, 1976; Domenici and Blake, 1991).
A variety of methods have been used to study the burst performance of larval fish. The
earliest methods consisted of using the presence of a real predator to feed on the fish larvae.
This is a ‘black box’ experiment, where the gut content of the predator determines the success of
prey during the predator-prey encounter. However, this method had the disadvantage because
the actual acceleration or burst performance of the fish larvae is not measured. However, it was
not until the development of high-speed cinematology in recording the kinematic of fish burst
performance when scientists were able to measure the burst response of fish. Then, the visual
stimulus and acoustic stimulus had been used to trigger the burst performance largely due to it
high repeatability (Dill, 1974b). The most recent developed method is the used of water flow to
trigger the predator strike (McHenry et al, 2009). A previous study was done to compare
response from visual stimuli and acoustic stimuli for red drum (Fuiman et al, 1999).The used of
pendulum with the silhouette that represent the cross section of the fish had most promising
result.

Most studies on burst performance of fish have focused on temperate freshwater species
and pelagic marine species with commercial value (Leis et al, 2006). Early studies include
flounder, plaice, herring and cod larvae (Bailey, 1984), northern anchovy (Webb and Corolla,
1981), salmon, red drum (Fuiman et al, 1999) and zebra Danio (Dill, 1974a,b). There has been
little research tropical reef species despite reef fish being different from temperate fish in many
aspects. For example, similar length reef fish larvae swim faster compared to temperate fish (Leis
and McCormick, 2006). In physiological aspect, fish are able to swim better in warmer tropic (25-
29°C) than in temperate condition (10-20°C) (Fuiman and Batty, 1997). The present study
collected some of the first information for the burst performance of a coral reef fish.
The overall objective of this project was to study the burst performance of a common reef
fish, the spiny Chromis (Acanthochromis polyacanthus). This objective will be achieved by: 1)
comparing the mean and maximum swimming speed during the startle response through the
early development stage of juvenile fish; 2) examining the effect of clutch identity on burst
performance; 3) examining the effect of growth and development on the response latency of
juvenile fish to visual stimulus.; and 4) identifying the change of direction juvenile fish during the
burst performance.

METHODS AND MATERIALS
Study species and maintenance: This study was done in February 2010 using laboratory
reared spiny damselfish, Acanthochromis polyacanthus, a fish lacking a larval dispersal phase
(Doherty et al, 1994; Miller-sims et al, 2008). The reason for choosing this species was to
compare the immediate response of the juvenile to previous studies on larvae. Juveniles of this
species have significant morphological differences from other Pomacentrid larvae (Murphy et al.,
2007), hence the burst performance of the juvenile may be unique. To obtain juveniles, the adult
fish were captured from a few locations within the Northern and Southern regions of the Great
Barrier Reef, because a previous study found that this species had genetically differentiated
subspecies within the Great Barrier Reef (Doherty et al, 1994). Hence, the subjects of this study
represent the whole breed stock of this species. Adult breeding pairs were kept in different tanks
at MARFU (JCU). The tanks were equipped with continuous flow of filtered salt water. Three
bricks were arranged like an arch in the tanks to allow the adult fish to attach the eggs.
Laboratory protocol: Juveniles were separated from the adult fish on the day they
hatched and reared in an air-conditioned environment maintained at constant temperature of 25
°C. The study was conducted in the laboratory and daily observations were taken before juveniles
were fed. A juvenile fish was randomly selected from a batch of fish and left in an aquarium
(10x10x10cm) that had been covered with black and dull fabric on 3 sides for 30 minutes to
acclimate. After the acclimation, an electro-magnetised pendulum was triggered to release a
transparent plate with darken oval, which acted as the burst stimulus. The shape of the oval was
chosen to represent the cross section silhouette of a common predator, the dottyback
Pseudochromis fuscus. A previous study showed that predation was maximal highest when fish
larvae were 10% of the length of the predator (Paradis et al, 1996). Hence, the size of the oval
was approximately the cross section of a dottyback that had 10 times the length of the average
body length of juvenile Acanthochromis. The burst performance of the juvenile before and after
the visual stimulus was video recorded (Fuiman & Cowan, 2003). A soft sponge was used to stop
the swinging stimulus and minimising the noise and vibration from the pendulum to avoid
triggering acoustic and tactile responses from the juvenile.
Video recording and analysis: The entire burst response of juvenile fish was recorded
with a high speed camera at 300fps. The camera was placed parallel to a (30cmx30cm) mirror
with a 45° to the experimental aquarium (Fuiman and Cowan, 2003; Fuiman et al., 2006). The
video was replayed, trimmed and transformed to image sequence (60Hz) using a Quicktime Pro.
Measurements including the burst speed and response latency were made using Image-J with
MtrackJ software. The maximum burst speed of a juvenile is the highest speed performed by it for
3 trials. The mean burst speed of a juvenile is the average of the highest speed performed by it

for 3 trials. Another high resolution digital stills camera was used to record the standard and total
length of the fish and the morphological differences (caudal fin grows) during its development
were noted by observation.
Data analysis: The responsiveness of juvenile fish to the pendulum image was recorded
as a binary variable, 0 as no response and 1 as positive response (Fuiman, 1989; Fuiman et al.,
1999). The response latency was defined as the t ime after the stimulus was triggered until the
first detectable movement observed from the juvenile fish (Domenici and Batty, 1997). The
directionality of the juvenile fish during the escape response was recorded as binary data. “Away
response” and “towards response” each comprised of 180° where the axis line is perpendicular to
the stimulus. One-factorANOVAs were used to examine the effect of size and age on the burst
speed to size and age respectively. Regression line was drawn to illustrate the response latency
to the age (day) and body size of the juvenile fish. Two-factor ANOVA was done on the size and
the batch effect to the maximum burst speed.

RESULTS
The overall mean of burst performance variables, such as the response latency (ms),
Square maximum , maximum burst speed (mm s-1), mean burst speed (mm s-1 )of all 177
juveniles is recorded in Table 1. The descriptive data such as the minimum, maximum and
mean of the variables is recorded.
Ontogenetic trends of maximum burst speed: The maximum burst speed (mm/s)
increased linearly with the length of the fish. The correlation both total length and standard length
of the fish to the maximum burst speed were statistically significantly (Fig. 1). However, the
standard length (r2 =0.149, p<0.001) of the fish showed a stronger burst response compared to
the total length (r2 =0.131, p<0.001).
Batch or clutch effects on juvenile performance: No systematic effect of clutch identity
was observed, where each batch of the juvenile spread within the range of maximum burst speed
within a range of body size (Fig 2).
Juveniles’ responsiveness and response latency: The responsiveness of the juvenile
fish increased as their body size increased and juveniles grew older. Of a total of 228 trials, 177
juveniles (77.6%) responded positively (Table 2 and 3). Juveniles one week after hatching (1wph)
had the lowest positive response (59.5%), followed by two-week post-hatching fish (2wph)
(74.8%), 3wph (82.4%) and 4wph juveniles which had 100% positive response (χ²= 17.705
p<0.001, df =3, Table 2). Juveniles with small standard body length had the lowest positive
response (59.02%), followed by the medium length (76.67%), large (91.94%) and extra large
(100%) juveniles which had 100% positive response (χ²= 23.848 p<0.001, df =3, Table 3). Chi-
squared test was done on the standard body length of the juvenile fish and age of the juvenile
fish. Both body size and age of the juvenile had a significant effect on the response score of the
fish (P<0.001, Chi-squared test).
The response latency had no relationship with the age of the juveniles and their body
size. Latency did not decrease for older juvenile for either total length or standard length of the
fish. However, fish standard length better represented the response time of the fish compared to
total length (r2 =0.10, p<0.001; r2 =0.08, p<0.001 respectively) . There was a negative correlation
between the maximum burst speed and the response latency of juvenile spiny Chromis (r=-0.396,
with 95% CI, Figure 3).
Directionality of the fish: From all the positive response, the initial startle directionality
was showed by binary angular variables. The Away or Towards (A/T) response which each

constitute 180° (Blaxter and Batty, 1985) depend on the change of directionality of fish during the
first stage of burst performance. Hence, the result was as expected where most of the juveniles
(99%) turned away from the stimuli with a few exceptions.

DICUSSION
Development of a startle response : Four of the five variables measured as part of the
burst response variables improved ontogenetically as expected. Maximum escape speed and
mean burst speed both significantly increased as juveniles grew. This result agrees with those of
previous studies on temperate species such as cod, flounder plaice and herring larval (Bailey and
Batty, 1984). However, contrary to previous study showing that the larvae required a shorter time
to respond as they grew larger (Fuiman et al, 1999), our study did not show an ontogenetic effect
on response latency. The 100 % away response showed by A. polyacanthus indicates a good
directionality in the predator prey interaction.
Ontogenetic trends of performance in A. polyacanthus :The maximum swimming
speed of juvenile Acanthochromis was highly dependent on the total length and standard length
of the juvenile fish. This result agrees with a previous study, which derived an equation for
average routine swimming speed as a function of larval total length for 9 species of fish (Miller et
al., 1988). The slope of our function in Figure 1 is slightly steeper than other studies which focus
on Northern anchovy larvae (Webb and Corolla, 1981; { max (cm/s)=1.95+20.8(length(mm)} and
temperate marine fish such as Atlantic Cod (Gadus morhua), Atlantic herring (Clupea harengus),
radiated shanny (Ulvaria subbifurcata) , capelin (Mallotus villosus) and winter flounder
(Pleuronectes americanus) William et al, 1996) {log (max speed) = 0.054 (length) +0.826. The
slope for Acanthochromis was much higher compared to both previous generalisations. This may
be due to a more advanced developmental stage of Acanthochromis, where the juvenile can
swim and feed immediately after they hatch.
Mean burst speed of juvenile Acanthochromis was also highly dependent on the standard
length of the juvenile indicating the importance of body length in predator avoidance. The slope of
the relationship of the mean burst speed against the standard size was smaller than for the
maximum burst speed, but still had a steeper slope compared to a previous studies (William et al,
1996) {log (mean speed) = 0.059 (length) +0.562}. This indicates that comparing the swimming
speed of different species based on length alone is not appropriate because fish undergo
developmental stages at different size. Generally, species which had similar body form perform
similarly.
As expected mean burst speed was highly correlated with the maximum burst speed.
This result agrees to that of a previous study, which also examined the effect of development on
the escape response in marine fishes (William et al, 1996). Since both variables are highly
correlated with body size, hence our study supports the ‘bigger –better hypothesis’ indirectly. This
is based on the hypothesis that higher burst speed enhances survival probability in the predator-
prey encounter. However, the linear increase found in the present study is contra to a previous

study, which included the encounter probability and searching effort as determining factors
(Bailey and Houde, 1989 in Paradis et al., 1996). These studies found that the vulnerability of fish
larvae to invertebrates’ predation and predatory fish is dome shape, where the fish seem to have
its optimum escaping size then the ability decrease through time (Bailey and Houde, 1989 in
Paradis et al., 1996). However, our study only involved the immediate post hatched juvenile,
which is the earliest life stage, hence survival of fish in different stages needs to be examine in
further research.
The response latency of juveniles did not reduce as age and/or body size increased.
These results differ from a previous study, which showed that red drum larvae had a shorter
response time as the larvae grew bigger levelling off as they reached 8 mm total length (Fuiman
et al, 1999). There may be other factors besides age and body size affecting response latency.
One factor may be eye development of the fish (Jones and Blaxter, 1967), where the rod
formations in the retina determine the response latency . Previous study had showed that the size
of eye is one of the determining factors in response time. Besides sensory organ, mechanical
factors, such as muscle constriction that has negative relationship with body length, also may
affect the response latency in this study (Wardle, 1967). The actual distance of the fish from the
stimulus might also affect the response latency. A study showed that the fish responded earlier
when they were nearer to the source of stimulus. The depth of the juvenile when the stimulus is
triggered was founds to be influential in response latency. This was due to the velocity of the
looming silhouette from the pendulum, known as the approaching speed of the dark oval, to the
juvenile. Since the stimulus is a two-dimensional image, due to the refraction of light the visual
image viewed by the juvenile is different at different depths in the aquarium.
However, our study found that the response latency of juveniles showed an interesting
negative relationship with maximum burst speed. Juveniles that reacted faster also responded
with a higher speed, or they had a faster burst response. This indicates that the magnitude of the
innate response by the juvenile is highly related to the effect of stimulus but, more research is
needed to understand the relationship of these two burst performance variables. A correlation
between these two variables, as showed in this study, would have important implications in our
understanding of the predator-prey encounter process.
Ontogenetic trends in juvenile responsiveness: The responsiveness of the juvenile fish
to the stimulus also increased as the size of the fish increased. This finding agrees with those of
previous studies on red drum larvae (Fuiman et al, 1999) and herring larvae (Blaxter and Fuiman,
1990). The increasing trend in this study was significant for larvae less than 6mm in body length,
but then the probability of responsiveness levelled off. Acanthochromis had a better
developmental stage compared to red drum larval and herring larvae, which each levelled off

after reaching 8mm (Fuiman et al., 1999) and 26 mm (Blaxter and Fuiman, 1990) respectively.
Hence, reef fish juvenile had better responsiveness compared to marine temperate fishes with
respect to body size.
An increase in the probability that a fish would respond to a visual stimulus indicates the
increase of active predator avoidance mechanism in these fish. They utilised more active
mechanisms for predator avoidance as their size increased. The newly hatched juvenile might
utilise other forms of predator avoidance mechanism, such as remain static and sink ing to the
bottom to camouflage themselves from the predator. Since they are small and less efficient
swimmers, their only mechanism to survive is to hide from the predator. This might indicate a shift
of mechanism from early to late juvenile stage from stationary camouflage to active escape.
Since their burst speed increases with body size, they can use more effective mechanism in
predator-prey encounter with increasing size.
Batch or clutch effects on juvenile performance: The maximum burst speed of juvenile
fish varied over a similar range with growth and development. This study showed no batch or
clutch effect on the burst performance, hence it support the risk spreading reproductive strategy
indirectly. This theory states that females try to vary the condition their offspring to maximise the
chance of batch survival. In other words, the individual survival is not as important as batch
survival in this reproductive resource allocation strategy. Although the Spiny Chromis lacks a
pelagic larvae stage, females may still practise this way of maximising the success of a
population through its evolution.
Kinematics of the burst performance: The usual swimming burst speed and the
stimulated burst speed were found to be highly correlated to one another (Holmes and
McCormick, 2009). This suggests that the improvement of propulsive surface area as the fins and
caudal tail grow helps the fish to burst in both circumstances. We observed that the surface area
of the caudal tail in the present study increased as the size of the juvenile reached 9 mm. This
result agrees with those of previous studies on many species of fish larvae (Fuiman, 1983;
Fuiman et al, 1988). Red drum larvae start to grow their caudal fin rays at a size of 4.5mm and
finish the process before they reach 6.0 mm, after which the burst speed increment reduces
(Fuiman et al., 1998). Hence, swim-related morphological features do influence on burst
performance kinetically.
Directionality during the first stage of the burst response (‘away response’): Only the
newly-hatched juvenile fish did not show a positive away response in Acanthochromis. This
suggests that that their visual sensory organ had not developed yet (at the body length of ~6mm).
For temperate fish, proliferation of rod in the peripheral retina occurred at a threshold of 25-30mm
body length (Blaxter and Jones, 1967). The result of the present study agrees with previous

studies where larger larvae showed a better away response to the stimulus with increasing age
(Yin and Blaxter, 1987; Domenici and Batty, 1997). An improvement in visual acuity might be the
cause of these directionality changes in escape response.
Limitations of the study : There were some limitations in recording the fish movement
such as the container volume, temperature and the duration of the experiment (Paradis et al.,
1996). The size of the container limits the distance travelled by the juvenile, because they reach
the side of the aquaria within a second forcing it to change direction. However, a previous study
on herring larvae showed that the response distance is no associated with sensory capability
(Blaxter and Fuiman, 1990). A second limitation of this study is the burst performance variables
only included the two dimension analysis (horizontal image only), hence the vertical movement of
the juvenile was ignored. To include this it is necessary to develop a new image analysing
software and a recording method to measure the change in depth by juvenile fish in burst
response. Since two out of the five variables measured depend on the distance travelled, it will be
more accurate if we included the vertical distance travel during the burst.
CONCLUSION
This study examined the burst performance variables for a common coral reef fish,
Acanthochromis polyacanthus. Measurements of burst performance, including mean and
maximum burst speed, directionality and response latency represent the first dataset for a coral
reef fish. These data can be used as reference for further studies on reef fish in survival during
the post-hatching stage. The present study found that the burst performance of very young
juveniles was less active compared to older juveniles. Their transparent body and static reaction
upon hatching may be due to underdeveloped sense organs or an innate response. On the other
hand, the older juveniles (more than 7 days after hatching) actively burst away from the stimulus
source. Future studies should examine the impact of parental condition in influencing the
development of burst performance in this interesting coral reef fish species.

REFERENCE LIST
Anderson JJ (1988) A neural model for visual activation of startle behavior in fish. Journal of
Theoretical Biology 131(3):279-288
Bailey KM (1984) Comparison of laboratory rates of predation of five species of marine fish larvae
by three planktonic invertebrates: effects of larval size on vulnerability. Marine Biology 79(3):303-
309
Bailey KM, Batty RS (1984) Laboratory study of predation by Aurelia aurita on larvae of cod,
flounder, plaice and herring: development and vulnerability to capture. Marine Biology 83 (3):
287-291
Batty S, Blaxter JHS, Fretwell K (1993) Effect of temperature on the escape responses of larval
herring, Clupea herengus. Marine Biology 115:523-528
Bertram DF (1994) Predation risk during the early life history periods of fishes - Separating the
effects of size and age. Marine Ecology Progress Series (Halstenbek) 109 (2-3):105-114
Blaxter JHS (1986) Development of sense organs and behaviour of teleosts larvae with special
reference to feeding and predator avoidance. Transactions of the American Fisheries Society
115: 98-114
Blaxter JHS (1991) Sensory systems and behaviour of larval fish. Marine Biology Its
accomplishment and future prospect (EDs) John Mauchline and Takahisa Nemoto. Amsterdam,
NY:15-38
Blaxter JHS (1988) Pattern and variety in development. In W. S. Hoas, D.J. Randall (eds). Vol XI.
The physiology of developing fish. Part A Eggs and larvae. Academic Press Inc. N.Y. London.
Blaxter JHS, Batty RS (1985) The development of startle responses in herring larvae. Journal
Marine Biology Association U.K. 65:737-750
Blaxter JHS, Fuiman LA (1990) The role of the sensory systems of herring larvae in evading
predatory fishes. Journal Marine Biology Association U.K. 70:413-427
Beamish FWH (1984) Swimming performance of three southwest Pacific fishes. Marine Biology
79:311-313
Claireaux G, Handeisman C, Standen E, and Nelson JA (2007)Thermal and temporal stability of
swimming performance in the European sea bass. Physiological and biochemical zoology 80 (2):
186-196

Cowan JH Jr, Houde ED, and Rose KA (1996) Size-dependent vulnerability of marine fish larvae
to predation: an individual-based numerical experiment. ICES Journal Marine Science 53:23–37
D’eath RB (1998) Can video images imitate real stimuli in animal behaviour experiments?
Biological Reviews 73:267-292
Dayton GH, Saenz D, Baum KA, Langerhas B, DeWitt TJ (2005) Body shape, burst speed and
escape behaviour of larval anurans. OIKOS 111:582-591
Dill LM (1974) The escape response of the zebra danio (Brachydanio rerio). II The effect of
experience. Animal Behaviour 22 (3):723-730
Dill LM (1974) The escape response of the zebra danio (Brachydanio rerio). I. The stimulus for
escape. Animal Behaviour 22 (3):711-722
Doherty PJ, Mather P, Planes S (1994) Acanthochromis polyacanthus, a fish lacking larval
dispersal, has genetically differentiated populations at local and regional scales on the Great
Barrier Reef. Marine Biology 121:11-21
Domenici P, Blake RW (1997)The kinematics and performance of fish fast-start swimming.
Journal of Experimental Biology 200 (8):1165-1178
Domenici P, Batty RS (1997) Escape behaviour of solitary herring (Clupea harengus) and
comparisons with schooling individuals. Marine Biology 128 (1):29-38
Dulcic J (2005) Biometric properties of damselfish, Chromis chromis (Osteichthyes:
Pomacentridae) from the middle Adriatic. Acta Adriat 46 (1):91-98
Eaton RC, Bombardieri RA, Meyer DL (1977) The mauthner-initiated startle response in teleosts
fish. Journal of Experimental Biology 66:65-81
Eaton RC, Didomenico R (1986) Role of the teleosts escape response during development.
Transactions of the American Fisheries Society 115:128-142
Eaton RC, Emberley DS (1991) How stimulus direction determines the trajectory of the mauthner-
initiated escape response in a teleosts fish. Journal of Experimental Biology 161:469-487
Ehrlich KF, Blaxter JHS, Pemberton R (1976) Morphological and histological changes during the
growth and starvation of herring and plaice larvae. Marine Biology 35 (2):105-118

Faria AM, Ojanguren AF, Fuiman LA, Goncalves EJ (2009) Ontogeny of critical swimming speed
of wild-caught and laboratory-reared red drum Sciaenops ocellatus larvae. Marine Ecology
Progress Series 384:221-230
Farrell AP (2008) Comparisons of swimming performance in rainbow trout using constant
acceleration and critical swimming speed tests. Journal of Fish Biology 72:693-710
Fisher R, Bellwood DR (2002) The influence of swimming speed on sustained swimming
performance of late-stage reef fish larvae. Marine Biology 140:801-807
Fisher R, Bellwood DR (2003) Undisturbed swimming behaviour and nocturnal activity of coral
reef fish larvae. Marine Ecology Progress Series 263:177-188
Fisher R, Hogan JD (2007) Morphological predictors of swimming speed: a case study of pre-
settlement juvenile coral reef fishes. The Journal of Experimental Biology 210:2436-2443
Franklin CE, Wilson RS, Davison W (2003) Locomotion at -1.0°C: burst swimming performance of
five species of Antarctic fish. Journal of Thermal Biology 28:59-65
Fuiman LA (1983) Growth gradients in fish larvae. Journal of Fish Biology 23, 117-123
Fuiman LA (1994) The interplay of ontogeny and scaling in the interactions of fish larvae and their
predators. Journal of Fish Biology 45 (suppl. a): 55
Fuiman LA, Cowan JH (2003) Behavior and recruitment success in fish larvae: repeatability and
covariation of survival skills. Ecology 84(1): 53-67
Fuiman LA, Cowan JH, Smith ME, O’Neal JP (2005) Behavior and recruitment success in fish
larvae: variation with growth rate and the batch effect. Canadian Journal of Fisheries and Aquatic
Sciences 62:1337-1349
Fuiman LA, Poling KR, Higgs DM (1998) Quantifying developmental progress for comparative
studies of larval fishes. Copeia 3: 602-611
Fuiman LA, Rose KA, Cowan JR JH, Smith EP (2006) Survival skills required for predator
evasion by fish larvae and their relation to laboratory measures of performance. Animal
Behaviour 71: 1389-1399

Fuiman LA, Smith ME, Malley VN (1999) Ontogeny of routine swimming speed and startle
responses in red drum, with a comparison of responses to acoustic and visual stimuli. Journal of
Fish Biology 55 sup A: 215-226
Fuiman LA, Webb PW (1988) Ontogeny of routine swimming activity and performance in zebra
danios (Teleostei: Cyprinidae). Animal behaviour 36 (1): 250-261
Gregory TR, Wood CM (1998) Individual variation and interrelationships between swimming
performance, growth rate, and feeding in juvenile rainbow trout (Oncorhynchus mykiss).
Canadian Journal of Fisheries and Aquatic Science 7:1583-1590
Greenwood AK, Peichel CL, Zottoli SJ (2010) Distinct startle responses are associated with
neuroanatomical differences in pufferfishes. The Journal of Experimental Biology 213:613-620.
Hale ME (1996) The development of fast-start performance in fishes: Escape kinematics of the
chinook salmon (Oncorhynchus tshawytscha). American Zoology 36: 695-709
Hale ME (1999) Locomotor mechanics during early life history: Effects of size and ontogeny on
fast-start performance of salmonid fishes. Journal of Experimental Biology 202 (11): 1465-1479
Hale ME (2000) Startle responses of fish without mauthner neurons: Escape behavior of the
lumpfish (Cyclopterus lumpus). The Biological Bulletin 199(2):180-182
Harper DG, Blake RW (1989) A critical analysis of the use of high-speed film to determine
maximum accelerations of fish. Journal of Experimental Biology 142:465-471
Harper DG, Blake RW (1990) Fast-start performance of rainbow trout Salmo gairdneri and
Northern pike Esox lucius. Journal of Experimental Biology 150:321-342
Harper DG, Blake RW (1991) Prey capture and the fast-start performance of Northern Pike, Esox
lucius. Journal of Experimental Biology 155:175-192
Hogan JD, Fisher R, Nolan C (2007) Critical swimming speed of settlement-stage coral reef
fishes from the Caribbean: A methodological and geographical comparison. Bulletin of Marine
science, 80(1):219-231
Holmes TH, McCormick M (2009) Influence of prey body characteristics and performance on
predator selection. Oecologia 159:401-413
Hunt von Herbing I, Boutilier RG (1996) Activity and metabolism of larval Atlantic cod (Gadus
morhua) from Scotian Chelf and Newfoundland source population. Marine Biology 124:607-617

Hunt von Herbing I, Boutilier RG, Miyake T, Hall BK (1996) Effects of temperature on
morphological landmarks critical to growth and survival in larval Atlantic cod (Gadus morhua).
Marine Biology 124:593-606
Johnson TP, Johnston IA (1991) Power output of fish muscle fibres performing oscillatory work:
Effects of acute and seasonal temperature change. Journal of Experimental Biology 157:409-423
Kamler E (2006) Parent-egg-progeny relationships in teleosts fishes: an energetic perspective.
Reviews in Fish Biology and Fisheries 15:399-421
Kendall Jr AW, Matarese AC (1994) Status of early life history descriptions of marine teleosts.
Fisheries Bulletin 92: 725-736
Kimmel CB, Patterson J, Kimmel RO (1972) The development and behavioural characteristics of
the startle response in the zebra fish. Developmental Psychobiology 7(1):47-60
Kolok AS (1992) Morphological and physiological correlates with swimming performance in
juvenile largemouth bass. American Journal of Physiology-Regulatory, Integrative and
Comparative Physiology 263:1042-1048
Laurence GC (1972) Comparative swimming abilities of fed and starved larval largemouth bass
(Micropterus salmoides). Journal of Fisheries Biology 4:73-78
Leis JM, Hay AC, Trnski T (2006) In situ ontogeny of behaviour in pelagic larvae of three
temperate, marine, demersal fishes. Marine Biology 148:655-669
Litvak MK, Leggett WC (1992) Age and size-selective predation on larval fishes: the bigger-is-
better hypothesis revisited. Marine Ecology Progress Series (Halstenbek) 81(1):13-24
Lyon JP, Ryan TJ, Scroggie MP (2008) Effects of temperature on the fast-start swimming
performance of an Australian freshwater fish. Ecology of Freshwater Fish 17:184-188
Margulies D (1989) Size-specific vulnerability of predation and sensory system development of
white seabass, Atractoscion nobilis larvae. Fishery bulletin (Washington, D.C.) 87 (3):537-552
Marras S, Claireaux G, Mckenzie DJ, Nelson JA (2010) Individual variation and repeatabili ty in
aerobic and anaerobic swimming performance of European sea bass, Dicentrarchus labrax. The
Journal of Experimental Biology 213:26-32.
Martinez M, Bedard M, Dutil JD, Guderley H (2004) Does condition of Atlantic cod (Gadus
morhua) have a greater impact upon swimming performance at Ucrit or sprint speeds? The
Journal of Experimental Biology 207:2979-2990

McCormick M, Molony BW (1993) Quality of the reef fish Upenus tragula (Mullidae) at settlement:
is size a good indicator of condition? Marine Ecology Progress Series 98:45-54
McHenry MJ, Feiti KE, Strother J, Van Trump WJ (2009) Larval zebrafish rapidly sense the water
flow of a predator’s strike. Biology Letter 5:477-479
McPeek MA, Grace M, Richardson JML (2001) Physiological and behavioural responses to
predators shape the growth/ predation risk trade-off in damselflies. Ecology 82(6):1535-1545
McPeek MA, Schrot AK, Brown JM (1996) Adaptation to predators in a New Community:
Swimming Performance and Predator avoidance in damselflies. Ecology 77 (2): 617-629
Meager JJ, Domenici P, Shingles A, Utne-Palm AC (2006) Escape responses in juvenile Atlantic
cod Gadus morhua L.: the effects of turbidity and predator speed. The Journal of Experimental
Biology 209: 4174-4184
Miller TJ (1993) Ontogenetic changes in behavioural and histological measures of visual acuity in
three species of fish. Environmental Biology of Fishes 37(1):1-8
Miller-sims VC, Gerlach G, Kingsford MJ, and Atema J (2008) Dispersal in the spiny damselfish,
Acanthochromis polyacanthus, a coral reef fish species without a larval pelagic stage. Molecular
Ecology 17:5036-5048
Muller, U.K. 2008 Swimming and Muscle. In Fish larval physiology. R.N. Finn, Kapor, B.G. (eds).
Science publiasher, USA.
Muller UK, Stamhuis EJ, Videler JJ (2000) Hydrodynamics of unsteady fish swimming and the
effects of the body size: comparing the flow fields of fish larvae and adults. The Journal of
Experimental Biology 203:193-206
Murphy BF, Leis JM, Kavanagh KD (2007) larval development of the ambon damselfish
Pomacentrus amboinensis, with a summary of pomacentrid development. Journal of Fish Biology
71:569-584
Nelson JA, Gotwalt PS and Snodgrass JW (2003) Swimming performance of blacknose dace
(Rhinichthys atratulus) mirrors home-stream current velocity. Canadian Journal Fish Aquatic
Science 60:301-308
Ott M (2006)Visual accommodation in vertebrates: Mechanisms, physiological response and
stimuli. Journal of comparative physiology. A, Neuroethology, sensory, neural, and behavioral
physiology, 192 (2):97

Oufiero CE, Garland Jr T (2009) Repeatability and correlation of swimming performances and
size over varying time-scales in the guppy (Poecilia reticulate). Functional Ecology 23:969-978
Paradis, AR, Pepin P, Brown JA (1996) Vulnerability of fish eggs and larvae to predation: review
of the influence of the relative size of prey and predator. Canadian Journal Fish Aquatic Science
53: 1226-1235
Penfold R, Grutter AS, Kuris AM, McCormick M, Jones CM (2008) Interactions between juvenile
marine fish and gnathiid isopods: predation versus micropredation. Marine Ecology Progress
series 357:111-119
Poling KR, Fuiman LA (1998) Sensory Development and Its Relation to Habitat Change in Three
Species of Sciaenids. Brain Behaviour Evolution 52: 270-284.
Portugues R, Engert F (2009) The neural basis of visual behaviors in the larval zebrafish. Current
Opinion in Neurobiology 19:644-647.
Post JR, Lee JA (1996) Metabolic ontogeny of teleost fishes. Canadian Journal of Fisheries and
Aquatic Sciences 53 (4):910
Robertson DR (1972) Field observations on the reproductive behaviour of a pomacentrid Fish,
Acanthochromis polyacanthus. Z. Tierpsychol 32: 319-324.
Royle NJ, Metcalfe NB, Lindstrom J (2006) Sexual selection, growth compensation and fast -start
swimming performance in green Swordtails, Xiphophorus helleri. Functional Ecology 20: 662-669
Skajaa K, Browman HI (2007)The escape response of food-deprived cod larvae (Gadus morhua
L.). Journal of Experimental Marine Biology and Ecology 353:135-144
Taylor EB, McPhail JD (1985) Variation in burst and prolonged swimming performance among
British Columbia populations of coho salmon, Oncorhynchus kisutch. Canadian Journal of
Fisheries and Aquatic Sciences 42(12):2029-2033
Taylor EB, McPhail JD (1985) Variation in body morphology among British Columbia populations
of coho salmon, Oncorhynchus kisutch. Canadian Journal of Fisheries and Aquatic Sciences 42
(12):2020-2028
Tudorache C, Blust R, De Boeck G (2008) Social interactions, predation behaviour and fast start
performance are affected by ammonia exposure in brown trout (Salmo trutta L.). Aquatic
Toxicology 90:145-153

Turesson H, Satta A, Domenici P (2009) Preparing for escape: anti-predator posture and fast-
start performance in gobies. The Journal of Experimental Biology 212:2925-2933
Wardle CS (1977) Effects of size on the swimming speeds of fish. In: Scale effects in animal
locomotion, (Ed) T.J. Pedle:299-313 NY Academic Press.
Wardle CS, He P (1988) Burst swimming speeds of markerel, Scomber scombrus L. Journal of
Fisheries Biology 32:471-478
Watkins TR, Vraspir J (2005) Both incubation temperature and posthatching temperature affect
swimming performance and morphology of wood frog tadpoles (Rana sylvatica). Physiological
and Biochemical Zoology 79(1):140-149
Webb PW (1983) Body and fin form and strike tactics of four teleosts predators attacking fathead
Minnow (Pimephales promelas) Prey. Canadian Journal Fishieries Aquatic Science 41:157-165
Webb PW, Corolla RT (1981) Burst swimming performance of northern anchovy, Engraulis
mordax, larvae. Fishery Bulletin 79(1):143-150
Webb PW, Fairchild AG (2001) Performance and maneuverability of three species of teleostean
fishes. Canadian Journal of Zoology 79(10): 1866-1877.
Weihs D (1974) Energetic advantages of burst swimming of fish. Journal Theory Biology 48:215-
229
Weihs D, Webb PW (1984) Optimal avoidance and evasion tactics in predator-prey interactions.
Journal Theoritical Biology 106, 189-206
Wiedenmayer CP (2009) Plasticity of defensive bahavior and fear in early development.
Neuroscience and Biobehavioral Reviews 33:432-441
William PJ, Brown JA, Gotceitas V, Pepin P (1996) Developmental changes in escape response
performance of five species of marine larval fish. Canadian Journal Fisheries Aquatic Sciences
53:1246-1253
Yamashita M, Naitoh T, Wassersug RJ (2000) Startle response and turning bias in Midrohyla
Tadpoles. Zoological Science 17:185-189
Yin MC, Blaxter JHS (1987) Escape speeds of marine fish larvae during early development and
starvation. Marine Biology 96:459-468
Zeddies DG, Fay RR (2005) Development of the acoustically evoked bahavioral response in
zebrafish to pure tones. The Journal of Experimental Biology 208:1363-1372
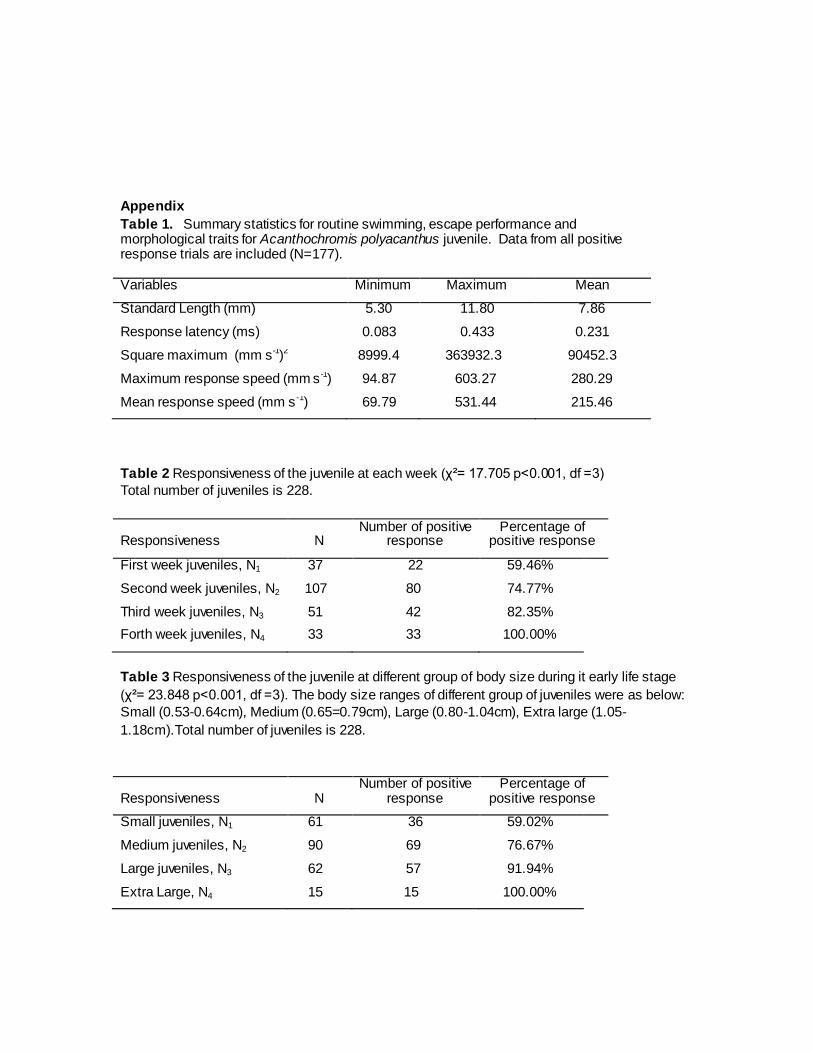
Appendix
Table 1. Summary statistics for routine swimming, escape performance and morphological traits for Acanthochromis polyacanthus juvenile. Data from all positive response trials are included (N=177). Variables Minimum Maximum Mean
Standard Length (mm) 5.30 11.80 7.86
Response latency (ms) 0.083 0.433 0.231
Square maximum (mm s-1)2 8999.4 363932.3 90452.3
Maximum response speed (mm s -1) 94.87 603.27 280.29
Mean response speed (mm s -1) 69.79 531.44 215.46
Table 2 Responsiveness of the juvenile at each week (χ²= 17.705 p<0.001, df =3)
Total number of juveniles is 228.
Responsiveness N Number of positive
response Percentage of
positive response
First week juveniles, N1 37 22 59.46%
Second week juveniles, N2 107 80 74.77%
Third week juveniles, N3 51 42 82.35%
Forth week juveniles, N4 33 33 100.00%
Table 3 Responsiveness of the juvenile at different group of body size during it early life stage
(χ²= 23.848 p<0.001, df =3). The body size ranges of different group of juveniles were as below:
Small (0.53-0.64cm), Medium (0.65=0.79cm), Large (0.80-1.04cm), Extra large (1.05-
1.18cm).Total number of juveniles is 228.
Responsiveness N Number of positive
response Percentage of
positive response
Small juveniles, N1 61 36 59.02%
Medium juveniles, N2 90 69 76.67%
Large juveniles, N3 62 57 91.94%
Extra Large, N4 15 15 100.00%
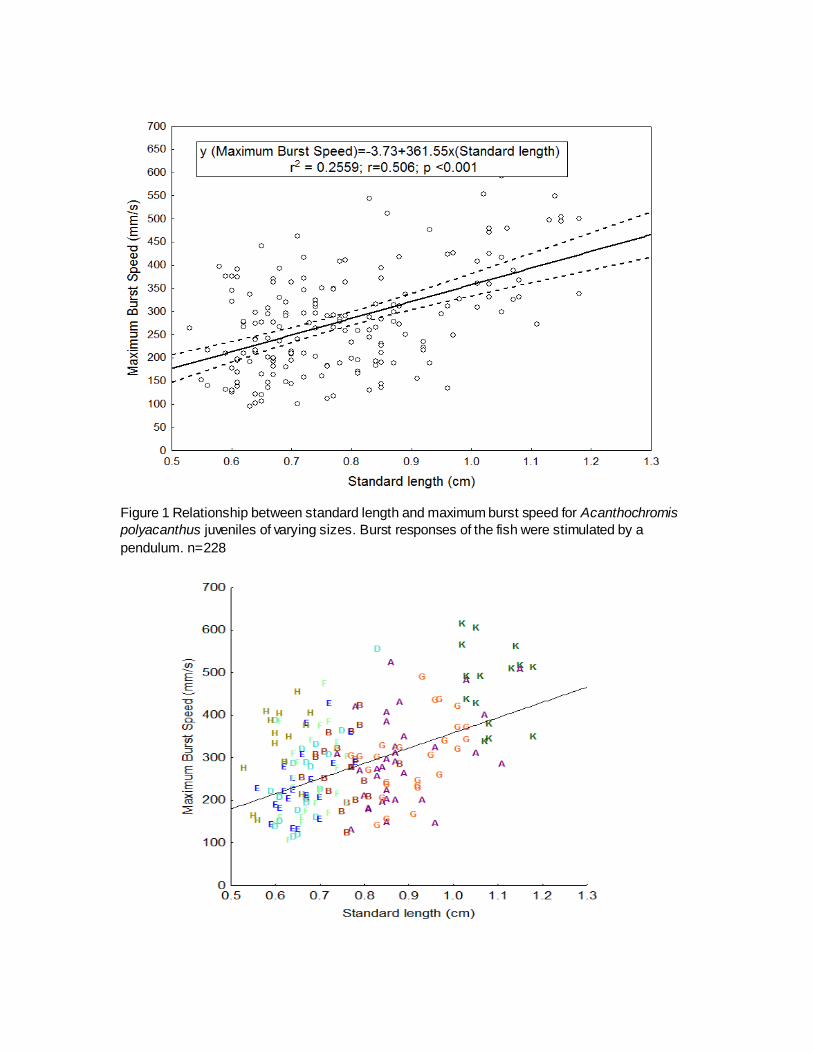
Figure 1 Relationship between standard length and maximum burst speed for Acanthochromis
polyacanthus juveniles of varying sizes. Burst responses of the fish were stimulated by a
pendulum. n=228
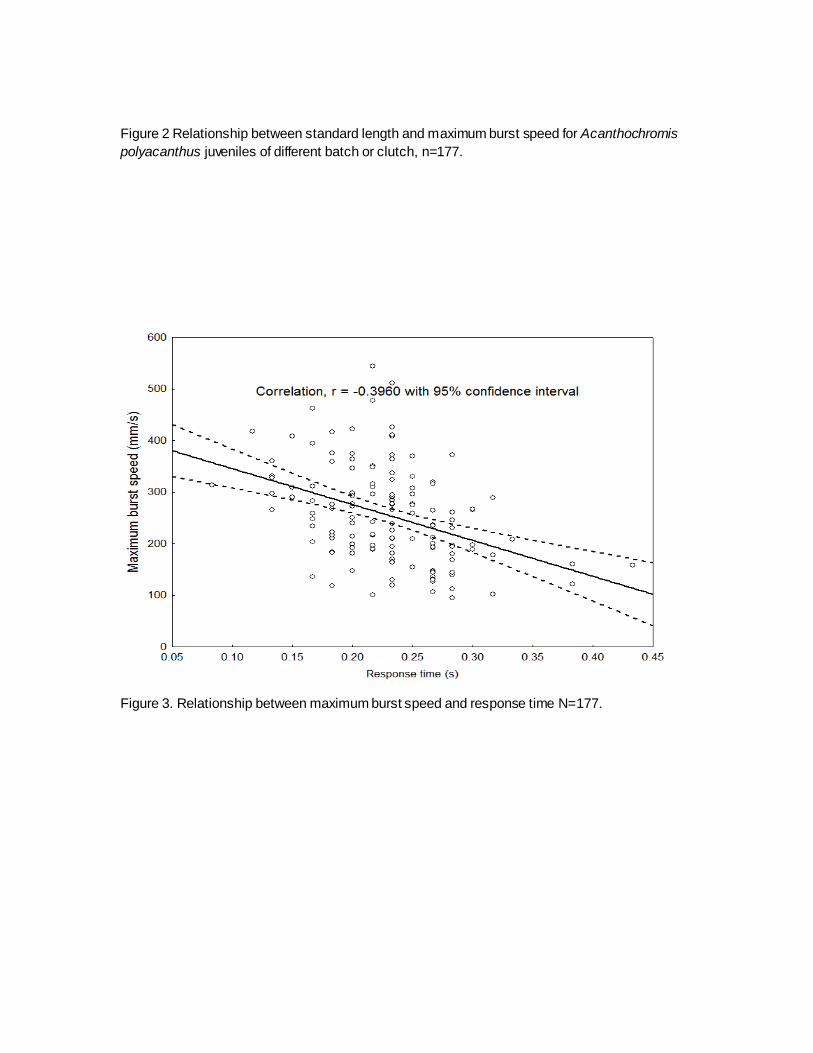
Figure 2 Relationship between standard length and maximum burst speed for Acanthochromis
polyacanthus juveniles of different batch or clutch, n=177.
Figure 3. Relationship between maximum burst speed and response time N=177.
Related Documents











