
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript





Dengue Bulletin – Volume 33, 2009 iii
AcknowledgementsEditor, Dengue Bulletin, WHO/SEARO, gratefully thanks the following for peer reviewing manuscripts submitted for publication.
In-house review:
Nand L. Kalra: Reviewed the manuscripts in respect of format check, content, conclusions drawn, including condensation of tabular and illustrative materials for clear, concise and focused presentation and bibliographic references. He was also involved in the final stages of printing of the Bulletin.
Peer reviewers
Raman Velayudhan 1. Vector Ecology and Management Department of Control of Neglected Tropical Diseases World Health Organization 20 Avenue Appia CH-1211 Geneva 27 Switzerland E-mail: [email protected]
Michael Johansson 2. Centers for Disease Control and Prevention Division of Vector-Borne Infectious Diseases 1324 Calle Canada Urb Puerto Nuevo San Juan, PR 00920 USA E-mail: [email protected]
To Setha 3. National Dengue Control Programme Ministry of Health Kingdom of Cambodia Phnom Penh, Cambodia E-mail: [email protected]
Zairi Jaal 4. Vector Control Research Unit School of Biological Sciences Universiti Sains Malaysia 11800 Penang Malaysia E-mail: [email protected]
Christophe Lagneau 5. Entente Interdépartementale de Démoustication du Littoral Méditerranéen Montpellier France E-mail: [email protected]
Sander Koenraadt 6. Laboratory of Entomology Wageningen University P.O. Box 8031 6700 EH Wageningen The Netherlands E-mail: [email protected]
Brian Kay 7. Australian Centre for International and Tropical Health Queensland Institute of Medical Research Brisbane Queensland, Australia E-mail: [email protected]
V.K. Saxena 8. Centre for Medical Entomology and Vector Management National Centre for Disease Control 22 Shamnath Marg Delhi 110054 E-mail: [email protected]

iv Dengue Bulletin – Volume 33, 2009
Denise Valle 9. Laboratory of Physiology and Control of Arthropod Vectors Instituto Oswaldo Cruz (IOC/FIOCRUZ) Av. Brasil 4365, Manguinhos Rio de Janeiro, RJ, Brazil E-mail: [email protected]
Thomas W. Scott 10. Mosquito Research Laboratory Department of Entomology One Shields Ave University of California Davis, CA 95616, USA E-mail: [email protected]
Melissa C. Hardstone 11. Cornell University Department of Entomology Ithaca, NY 14853, USA E-mail: [email protected]
Aruna Srivastava 12. National Institute of Malaria Research Delhi – 110092, India E-mail: [email protected]
Lars Eisen 13. Department of Microbiology, Immunology and Pathology Colorado State University, Fort Collins, CO 80523, USA E-mail: [email protected]
Lourdes Esteva 14. Departamento de Matemáticas Facultad de Ciencias, UNAM Circuito Exterior México, D.F. 04510 E-mail: [email protected]
Andrew Falconar 15. London School of Hygiene & Tropical Medicine Keppel Street London WC1E 7HT UK E-mail: [email protected]
Dave Chadee 16. Department of Life Sciences University of the West Indies St Augustine, Trinidad and Tobago E-mail: [email protected]
Jennifer Kyle 17. Division of Infectious Diseases and Vaccinology School of Public Health University of California, Berkeley, Berkeley, California, United States of America E-mail: [email protected]
Martin Grobusch 18. Infectious Diseases Unit Division of Clinical Microbiology and Infectious Diseases National Health Laboratory Service and School of Pathology Faculty of Health Sciences University of the Witwatersrand 7 York Road, Parktown 2196 Johannesburg, South Africa E-mail: [email protected]
Scott Ritchie 19. School of Public Health Tropical Medicine and Rehabilitation Sciences James Cook University Cairns, QLD 4870 Australia E-mail: [email protected]
Sebastien Marcombe 20. Institut de Recherche pour le Développement Centre Collaborateur OMS UR 016 Caracterisation et controle des populations de vecteurs LIN, IRD, BP 64501 34394 Montpellier Cedex 5 France E-mail: [email protected]

Dengue Bulletin – Volume 33, 2009 v
Héctor Masuh 21. Pests and Insecticides Research Centre Buenos Aires Argentina E-mail: [email protected]
Hilary Ranson 22. Vector Group Liverpool School of Tropical Medicine Liverpool, L35QA UK E-mail: [email protected]
Chang Moh Seng 23. The World Health Organization Regional Office for the Western Pacific (WPRO) P.O. Box 2932 1000 Manila Philippines E-mail: [email protected]
T.D.K. Thai 24. Division of Infectious Diseases, Tropical Medicine and AIDS Academic Medical Center, F4-106 Meibergdreef 9 1105 AZ Amsterdam The Netherlands E-mail: [email protected]
John D. Edman 25. Center for Vector-borne Diseases University of California Davis, CA 95616, USA E-mail: [email protected]
Siripen Kalyanarooj 26. Queen Sirikit National Institute of Child Health Bangkok Thailand E-mail: [email protected]
Jacqueline Deen 27. E-mail: [email protected]
Javier Mota-Sanchez 28. Virology and Immunology Southwest Foundation for Biomedical Research San Antonio, TX, USA E-mail: [email protected]
S Satish Appoo 29. Environmental Health Department National Environment Agency Singapore E-mail: [email protected]
Donald Yee 30. Department of Biological Sciences The University of Southern Mississippi 118 College Drive # 5018 Hattiesburg, MS 39406-0001, USA E-mail: [email protected]
Thomas Burkot 31. Centers for Disease Control and Prevention Division of Parasitic Diseases 4770 Buford Highway, NE Mailstop F-42 Bldg 102, Room 2118 Atlanta, GA 30341-3724, USA E-mail: [email protected]
Ole Wichmann 32. Dept. for Infectious Disease Epidemiology Robert Koch-Institute DGZ-Ring 1 13086 Berlin Germany E-Mail: [email protected]
John McBride 33. Infectious Diseases Physician and Clinical Microbiologist School of Medicine and Dentistry James Cook University, Cairns Campus Cairns Base Hospital PO Box 902 Cairns, Queensland Australia E-mail: [email protected]
Nguyen Thanh Hung 34. Department of Dengue Haemorrhagic Fever Children’s Hospital #1 Ho Chi Minh City Viet Nam E-mail: [email protected]
Eric Martinez 35. Pedro Kouri Institute of Tropical Medicine Havana City Cuba E-mail: [email protected]

vi Dengue Bulletin – Volume 33, 2009
Robert V Gibbons 36. Department of Virology Armed Forces Research Institute of Medical Research 315/6 Rajvithi Road Bangkok 10400 Thailand E-mail: [email protected]
Scott Halstead 37. Research Program Pediatric Dengue Vaccine Initiative Rockville, Maryland 20852, USA E-mail: [email protected]
Anon Srikiatkhachorn 38. Center for Infectious Disease and Vaccine Research University of Massachusetts Medical School Worcester, Massachusetts, USA E-mail: [email protected]
Elizabeth Hunsperger 39. Serology Diagnostics and Viral Pathogenesis Research Laboratory Centers for Disease Control and Prevention Dengue Branch 1324 Calle Canada San Juan, PR 00920, USA E-mail: [email protected]
Veerle Vanlerberghe 40. Unit of Epidemiology and Disease Control Public Health Department, Institute of Tropical Medicine Nationalestraat 155, Antwerp Belgium E-mail: [email protected]
Linda Lloyd 41. 3443 Whittier St. San Diego, CA 92106, USA. E-mail: [email protected]
Peter Ryan 42. Mosquito Control Laboratory Queensland Institute of Medical Research PO Royal Brisbane Hospital Brisbane QLD 4029 Australia E-mail: [email protected]
Michael Nathan 43. E-mail: [email protected]
Ng Lee-Ching 44. Environmental Health Institute The National Environment Agency Singapore E-mail: [email protected]

Dengue Bulletin – Volume 33, 2009 vii
Contents
Implementing predictive models for domestic decision-making 1. against dengue haemorrhagic fever epidemics ..................................................1Halmar Halide
Use of geographical information system (GIS) and 2. global positioning system (GPS) for dengue and dengue haemorrhagic fever control in Sri Lanka ................................................................................11G.A.J.S.K. Jayasooriya, S.M.L. Senaratne, W.M.C.M. Wijesinghe, P.H.D. Kusumawathie, J. Gunatilake
Estimating the basic reproduction number of dengue transmission 3. during 2002-2007 outbreaks in Bandung, Indonesia .......................................21Asep K. Supriatna
Dengue fever in a tertiary hospital in Makkah, Saudi Arabia4. ............................34W. Shahin, A. Nassar, M. Kalkattawi and H. Bokhari
Genotypic and phenotypic characteristics of DENV-3 isolated 5. from patients with different disease severities in Indonesia ..............................45Beti Ernawati Dewi, Tomohiko Takasaki, Shigeru Tajima, T Mirawati Sudiro, R.P. Larasati, Andrew Lee Corwin and Ichiro Kurane
Dengue fever among ill-returned travellers and concurrent infection 6. by two dengue virus serotypes ........................................................................60Khoa T.D. Thai, Josta A. Wismeijer, Michèle van Vugt, Katja C. Wolthers and Peter J. de Vries
Acute abdominal pain in dengue haemorrhagic fever: 7. A study in Sri Lanka, 2009 ..............................................................................70K.G.A.D. Weerakoon, S. Chandrasekaram, J.P.S.N.K. Jayabahu, S. Gunasena and S.A.M. Kularatne
Involvement of the liver in dengue infections8. ..................................................75Duncan R. Smith and Atefeh Khakpoor
Improving dengue virus diagnosis in rural areas of Mexico9. ..............................87Moreno-Altamirano M.M.B., Sánchez-García F.J., López-Martínez I., Rosales-Jiménez C., Vázquez-Pichardo M., Arriaga-Valona L.J. and Capitan-Ortega F.

viii Dengue Bulletin – Volume 33, 2009
Contents
Dengue vector surveillance and control in Hong Kongin 2008 and 200910. ........95K.Y. Cheung and M.Y. Fok
Comparative life parameters of transgenic and wild strain 11. of Aedes aegypti in the laboratory .................................................................103H.L. Lee, H. Joko, W.A. Nazni and S.S. Vasan
Protein profiles of dengue-infected 12. Aedes aegypti (L) ....................................115H.L. Lee, Y.C. Wong and A. Rohani
Susceptibility status of transgenic 13. Aedes aegypti (L.) against insecticides .........124W.A. Nazni, S. Selvi, H.L. Lee, I. Sadiyah, H. Azahari, N. Derric and S.S. Vasan
Epidemiological analysis of hospitalized cases of 14. dengue fever/dengue haemorrhagic fever and extent of breeding of Aedes aegypti in major hospitals in the National Capital Territory of Delhi (NCT Delhi), 2005–2009 ................................................................130J. Nandi, R.S. Sharma, R.K. Dasgupta, R. Katyal, P.K. Dutta and G.P.S. Dhillon
Studies on the efficacy of 15. Toxorhynchites larvae and three larvivorous fish species for the control of Aedes larval populations in water-storage tanks in the Matale district of Sri Lanka ...............................140W.M.G.S. Wijesinghe, M.B. Wickramasinghe, P.H.D. Kusumawathie, G.A.J.S.K. Jayasooriya and B.G.D.N.K. De Silva
A novel method of controlling a dengue mosquito vector, 16. Aedes aegypti (Diptera: Culicidae) using an aquatic mosquito predator, Diplonychus indicus (Hemiptera: Belostomatidae) in tyres .............................148N. Sivagnaname
Effect of water supply system installation on distribution of 17. water storage containers and abundance of Aedes aegypti immatures in urban premises of Ho Chi Minh City, Viet Nam ............................................161Ataru Tsuzuki, Trang Huynh, Loan Luu, Takashi Tsunoda and Masahiro Takagi
Evaluation of premise condition index in the context of 18. Aedes aegypti control in Marília, São Paulo, Brazil ........................................167Maria Teresa Macoris Andrighetti, Karen Cristina Galvani and Maria de Lourdes da Graça Macoris

Dengue Bulletin – Volume 33, 2009 ix
Contents
The control of 19. Aedes aegypti for water access in households: Case studies towards a school-based education programme through the use of net covers ........................................................................176João Bosco Jardim, Héliton da Silva Barros, Caroline Macedo Gonçalves, Paulo Filemon Paolucci Pimenta and Virgínia T. Schall
Container survey of mosquito breeding sites in a university campus 20. in Kuala Lumpur, Malaysia ............................................................................187C.D. Chen, H.L. Lee, S.P. Stella-Wong, K.W. Lau and M. Sofian-Azirun
Detection of insecticide resistance in 21. Aedes aegypti to organophosphate and synthetic pyrethroid compounds in the north-east of Thailand ...............194S. Pimsamarn, W. Sornpeng, S. Akksilp, P. Paepornand M. Limpawitthayakul
Evaluation of a “fogging” canister for indoor elimination of 22. adult Aedes aegypti .......................................................................................203Pang Sook Cheng, Foo Siew Yoong, Png Ah Bah, Deng Lu, Lam-Phua Sai Gek, Tang Choon Siang and Ng Lee Ching
Oviposition behaviour of 23. Aedes albopictus in temephos and Bacillus thuringiensis israelensis-treated ovitraps ........................................................209W.A. Nazni, H.L. Lee, W.M. Wan Rozita, A.C. Lian, C.D. Chen,A.H. Azahari and I. Sadiyah
Book reviews
Dengue guidelines for diagnosis, treatment, 24. prevention and control, 2009 (WHO/HTM/NTD/DEN/2009.1) .....................218
Instructions for contributors25. ..........................................................................220

Dengue Bulletin – Volume 33, 2009 1
Implementing predictive models for domestic decision-making against
dengue haemorrhagic fever epidemicsHalmar Halide#
Physics Department, FMIPA, Hasanuddin University, Makassar 90245, Indonesia
Abstract
The efficacy of two simple models for predicting dengue haemorrhagic fever (DHF) epidemics are evaluated. One model uses persistence while the other uses past dengue cases and climate factors to make predictions. It is shown that the efficacy of the models is not significantly different. The value of the prediction is also investigated when it is used to decide whether it can protect a household from epidemics. When the model predicts that a DHF epidemic is forthcoming, a highly effective but low-cost DEET product is applied by the whole family as protection against mosquito bites. It is found that the cost of implementing such a model for prediction is much cheaper than other options such as: (i) using protection without any forecast; and (ii) neglecting any protection. It is also found that the value of a forecast depends on the forecast skill and the cost-to-loss ratio.
Keywords: DHF epidemics; predictive model; forecast value; decision-making; DEET.
#E-mail: [email protected]
IntroductionDengue haemorrhagic fever (DHF) causes a substantial burden to a family in terms of loss of life and economic impact.[1,2,3,4,5] The number of people suffering from the illness is also predicted to increase in the years ahead due to global warming.[6,7,8] Therefore, an early warning system (EWS), even with a one-month lead prediction for an upcoming dengue haemorrhagic fever (DHF) epidemic,[9] is urgently needed.[10,11,12] Such a system can be used to make an informed decision to
prevent the occurrence of an epidemic at a family scale.
There are a few models that could serve as an EWS. The models range in complexities and use biotic and abiotic factors to make dengue predictions. More recently, a simple statistical model, HR2008, has been able to give a useful epidemic prediction up to six months in advance.[9]
In this study, the HR2008 model and a persistence model are implemented in a

2 Dengue Bulletin – Volume 33, 2009
Implementing predictive models for domestic decision-making against DHF epidemics
decision-making problem as an attempt to prevent an epidemic in the city of Makassar, Indonesia (5.1°S, 119.6°E). The decision of whether or not a family applies a protective measure is made based on the model’s prediction. The value of a forecast is assessed through expenses resulting from several decision options.
Methods
Data
The monthly number of confirmed DHF cases was recorded by the Public Health Division at the city of Makassar, Indonesia. Predictive models were developed using these cases. Length of stay (LoS) and cost to patients were obtained at a regional hospital, RS Wahidin Sudirohusodo, at Makassar during DHF epidemics, i.e. the months of January and April. The focus was on patients who occupied rooms with the least expensive rates. Other demographic data such as household size was obtained from the Makassar Bureau of Statistics.
Model and predictions
The two models used to give a one-month lead prediction of DHF epidemics are briefly described. An epidemic is defined when the number of DHF cases exceeds the 75th percentiles.[13] The models are:
a persistence model which states (1) that the number of DHF cases in the following month is the same as that of the present month, i.e.
N(t+1)=N(t) [1]
where N(t) is the number of cases at time t (measured in months).
a DHF predictive model HR2008 (2) developed earlier.[9] This model uses both past DHF cases and local meteorological variables such as relative humidity h and average temperature Tave to predict cases in the following month. The model was run on DHF data from the period January 1999 to December 2005 and gives the following closed-form formula for predicting the number of cases a month in advance:
N(t+1)=0.73N(t)–3.44h(t-4)-16.43Tave(t-5) +732.45 [2]
Note that the HR2008 model is capable of producing a useful prediction skill up to six months in advance against a no-skill random forecast.[9]
Prediction skill assessment
In order to assess the prediction skill of these two models, we use predictions covering the period from February 1999 to December 2005, i.e. 83 months. The skill of each model is determined by its Peirce score using a contingency table as in Table 1. In this table a, b, c and d refer respectively to the number of times the epidemic is forecast and also observed, the epidemic is forecast but did not occur, the epidemic is not forecast but did occur, and the epidemic is neither forecast nor observed. The score and its error estimate are calculated using data from Table 1 and the following formulas below.[14]
Peirce skill score PSS = (ad-bc)/(a+c)(b+d) [3]
Standard error ePSS = [(n2-4(a+c)(b+d)PSS2)/4n(a+c)(b+d)]1/2 [4]
where the total number of predictions and observations n = a+b+c+d.

Dengue Bulletin – Volume 33, 2009 3
Implementing predictive models for domestic decision-making against DHF epidemics
Table 1: Contingency table for the Yes/No of DHF epidemic forecast[9]
DHF epidemic predicted
DHF epidemic observed
Yes No
Yes a (hit) b (false alarm)
No c (miss) d (correct rejection)
The prediction skill of a model is usually compared against a random no-skill forecast by first transforming the above a, b, c, d values as:
ar=(a+c)(a+b)/n [5]
br=(b+d)(a+b)/n [6]
cr=(a+c)(c+d)/n [7]
dr=(b+d)(c+d)/n [8],
and then the transformed values (5–8) are substituted into (3) and (4) to obtain score and error for the random forecast.
Decision-making problem
A household based its decision whether or not to take any protective measures depending on a model forecast. The family will only take protective measures against an epidemic when a model predicts that the event is forthcoming. In this case, the family member applies a highly effective but low-cost DEET product daily for personal protection.[15] Note that this mode of protection is selected from among other forms of domestic interventions[16,17,18,19] because it directly protects a person both in and outside the house from mosquito bites. The economic value of using such a model forecast for taking a decision is examined below.
Forecast value evaluation
The value of a decision is examined in terms of cost C and loss L. The former occurs when the family uses a daily protection method and the loss is incurred when the unprotected family suffers from an epidemic. Note that one could also perform a cost-benefit analysis, i.e. a benefit is the savings resulting from taking a protection. Beside a forecast-led decision, there are also other options to consider. They are: the family applies a daily protection regardless of any forecast and the family does not use any protection at all.
The expense E for each decision is calculated using Thorne and Stephenson (2002) formulation.[20]
E1 (for not using any protection) = (a+c) × L [9]
E2 (for a daily protection regardless of any forecast) = (a+b+c+d) × C [10]
E3 (for using a predictive model) = ((a+b) × C) + (c × L) [11]
E4 (for using perfect forecast) = (a+c) × C [12]
The value of a forecast is presented as a value index and calculated using the above expenses as:
VI = (E2-E3)/(E2-E4) [13]
Results
Models skill
Observed DHF cases (circled) and out-of-sample predictions (lined) of cases for both predictive “HR2008” and “persistence” models are presented in Figure 1. We also

4 Dengue Bulletin – Volume 33, 2009
Implementing predictive models for domestic decision-making against DHF epidemics
plot a horizontal dotted-line at dengue cases equalling to 134 at 75% percentiles to assign epidemic events. Figure 1 shows that the HR2008 model correctly predicts the moderately severe epidemic peaks from 2001 to 2005. These epidemics, however, are predicted to occur one month later by the persistence model as expected. It was also found that the HR2008 wrongly predicted higher cases in 1999 and 2000 and a few negative cases in 1999. None of the latter problems are found in the persistence model.
The contingency parameters and forecast skills for both models are presented in Table 2 and Figure 2. The one-month delay in predicting these epidemics seems to lower the number of hit rates a, and the correct rejections d obtained by the persistence model compared with that of the HR2008
model. The Peirce skill score, however, is not significantly different from each model. Both models have a much higher skill than that of the random forecast.
Table 2: Prediction skill of the HR2008 and persistence models and their corresponding
no-skill forecasts (in parenthesis)
ParametersModel
HR2008 Persistence
a 18 (7) 16 (6)
b 5 (6) 7 (17)
c 7 (8) 6 (16)
d 53 (42) 54 (44)
Peirce skill score
0.63±0.10 (0.0±0.12)
0.61±0.10 (-0.01±0.12)
Figure 1: Data (observed DHF cases) and the out-of-sample predictions of DHF cases at one month in advance for the HR2008 and persistence models
(The horizontal dotted line represents the 75% percentiles of DHF cases)

Dengue Bulletin – Volume 33, 2009 5
Implementing predictive models for domestic decision-making against DHF epidemics
Models’ forecast value
Cost of protection
The household size in Makassar ranges from 3.16 to 5.26 persons, with an average of 4.26 in a total population of about 1 223 540.[21] The minimum monthly regional wage in 2006 was US$55.64[21] (US$ 1=11 000 Indonesian Rupiahs). Let us suppose a family of four is to be protected against an epidemic. The mode of protection uses an insect-repellent called AUTAN. This product comes in a lotion which contains 12.5% DEET. It is packed in a sachet weighing 10 g. Each person applies the product twice a day, i.e. two sachets, for 12-hour protection during daytime according to an efficacy test.[22] One sachet of AUTAN costs 4.5 cents. The total cost of protecting a family of four for 30 days, therefore, equals US$ 10.9.
Loss due to DHF epidemics
If a member of the family is not protected against dengue-carrying mosquito bites, he/she has the risk of getting hospitalized due to DHF. The length of stay (LoS) (in terms of nights) of a DHF patient during the 2008 epidemic in Wahidin Sudirohusodo Hospital ranges from one to eight days, with an average of 4.8 days. The economic loss for each night spent in the least expensive room is presented in Table 3. The cost includes: blood examination, treatment, meals, visits by physicians and nurses, and room rent. The cost-to-loss ratio (C/L), expenses and the value index of the two predictive models are also presented in Table 3 and Figure 3.
In Table 3, the expense resulting from implementing a forecast E3 is cheaper than that of the no-protection E1 and protection
Figure 2: Peirce skill scores including the error estimates (error bar) for both predictive models HR2008 (circle) and persistence model (upper triangle) and their associated no-skill random
models in crosses (×), respectively

6 Dengue Bulletin – Volume 33, 2009
Implementing predictive models for domestic decision-making against DHF epidemics
Table 3: Forecast value of the HR2008 and persistence models (expenses and value index for their corresponding no-skill forecasts in brackets. The cost C for protecting a family of four
people is US$ 10.9. E2 and E4 are the same for all nights. Note that the figures in squared-brackets are the number
of patients with corresponding LoS)
Model
Parameters
Length of stay in hospital LoS (nights)
1 [3]
2 [13]
3 [8]
4 [9]
5 [3]
6 [0]
7 [2]
8 [1]
Loss (L) (US$) 15.0 23.2 31.4 39.5 47.7 55.9 64.1 72.3
C/L 0.73 0.47 0.35 0.28 0.23 0.20 0.17 0.15
HR2008 E1 (US$) 375.0 (375.0)
579.5 (579.5)
784.1 (784.1)
988.6 (988.6)
1193.2 (1193.2)
1397.7 (1397.7)
1602.3 (1602.3)
1806.8 (1806.8)
E2 (US$) 905.5 (905.5)
E3 (US$) 355.9 (520.9)
413.2 (668.2)
470.4 (815.4)
527.7 (962.7)
585.0 (1110.0)
642.3 (1257.3)
699.5 (1404.5)
756.8 (1551.8)
E4 (US$) 272.7 (272.7)
VI 0.87 (0.61)
0.78 (0.38)
0.69 (0.14)
0.60 (-0.09)
0.51 (-0.32)
0.42 (-0.56)
0.32 (-0.79)
0.23 (-1.02)
Persistence E1 (US$) 330.0 (330.0)
510.0 (510.0)
690.0 (690.0)
870.0 (870.0)
1050.0 (1050.0)
1230.0 (1230.0)
1410.0 (1410.0)
1590.0 (1590.0)
E2 (US$) 905.5 (905.5)
E3 (US$) 340.9 (490.9)
390.0 (621.8)
439.1 (752.7)
488.2 (883.6)
537.3 (1014.5)
586.4 (1145.5)
635.5 (1276.4)
684.5 (1407.3)
E4 (US$) 240.0 (240.0)
VI 0.85 (0.62)
0.77 (0.43)
0.70 (0.23)
0.63 (0.03)
0.55 (-0.16)
0.48 (-0.36)
0.41 (-0.56)
0.33 (-0.75)
without any forecast E2 option. Table 3 also shows that both models give similar forecast values. Their corresponding no-skill random forecasts have lower forecast values due to their low skill (Table 2). It is also found that as the C/L ratio gets smaller, the forecast value decreases (Figure 3). Note that the value index (VI) of the no-skill forecast contains some non-positive value. In such a case, the forecast has no value, i.e. it is better just to use a daily protection regardless of any prediction.
Discussion
This study is the first to implement and determine the value of a prediction by using a single mode of protection against DHF epidemics with an insect repellent. It is shown that the forecast implementation has an economic value. The value depends on factors such as forecast skills and the cost-to-loss ratio. Simple protection using a DEET-based repellent is rarely used as a means for community protection against epidemics.

Dengue Bulletin – Volume 33, 2009 7
Implementing predictive models for domestic decision-making against DHF epidemics
The DEET-based product is highly effective and offers a broad-spectrum protection against mosquitoes, ticks, flies and insect bite.[23,24] Depending on application dosage and DEET concentrations, the product is able to give protection up to seven hours.[23,25] This product is also safe for adults and children provided the dose is correctly applied.[26] It is not surprising that DEET has been considered the single-most effective personal protection for many years.[27] However, this mode of protection has not been widely used in a population against DHF epidemics.
There are at least two reasons why the population at large is still reluctant to use a DEET product against epidemics. First, it might affect the human skin since the product is known to be corrosive to fabrics, plastic and vinyl.[28] Secondly, skin irritation, poisoning and toxicity occurrence have been reported in cases of excessive dosage.[29,30] Therefore, it is important to
ensure that the product is used properly. The recommendations to be followed are: there should be a six-hour interval between DEET applications, and the repellent should not be orally ingested.[31] In addition, for infants aged above two months, the product is limited to one application per day and the maximum DEET concentration should be 30%.[31]
ConclusionThe skill of two simple models for predicting DHF epidemics is assessed using a Peirce score. The skill of HR2008 model is not significantly different than that of a persistence model. Both models have a much higher skill than that of their corresponding no-skill random forecast. Both model predictions are also applied to determine whether or not a family should take protective measures against mosquito bites.
Figure 3: Calculated forecast values of predictive models including the no-skill random forecasts for a DHF patient at the hospital

8 Dengue Bulletin – Volume 33, 2009
Implementing predictive models for domestic decision-making against DHF epidemics
In order to avoid mosquito bites, use of a DEET-based repellent is proposed and simulated. It is found that the cost of implementing DEET application based on model predictions is lower than that of other options such as: never using any protection and never using any forecast when applying a protection. It is also shown that both models have a similar forecast value and they have a much higher economic value than that of no-skill forecast. The forecast value gets smaller as the C/L ratio decreases.
Acknowledgements
We thank Mr Suherman, a medical staff at Wahidin Sudirohusodo Public Hospital in Makassar, who provided us with the expenses and LoS data of DHF patients in the hospital. We also thank an anonymous reviewer for the constructive comments, and Dr Peter Ridd of James Cook University and Dr David McKinnon of the Australian Institute of Marine Science for proof-reading the manuscript.
ReferencesGubler DJ. Epidemic dengue/dengue [1] hemorrhagic fever as a public health, social and economic problem in the 21st century. Trends in Microbiology. 2002;10(2):100-103.
van Damme W, van Leemput L, Por Ir, [2] Hardeman W, Meessen B. Out-of-pocket health expenditure and debt in poor households: evidence from Cambodia. Tropical Medicine and International Health. 2004;9(2):273-280.
Shepard DS, Suaya JA, Halstead SB, Nathan [3] MB, Gubler DJ, Mahoney RT, et al. Cost-effectiveness of a pediatric dengue vaccine. Vaccine. 2004; 42, 1275-1280.
Anderson KB, Chunsuttiwat S, Nisalak [4] A, Mammen MP, Libraty DH, Rothman AL, et al. Burden of symptomatic dengue infection in children at primary school in Thailand: a prospective study. Lancet. 2007;369:1452-1459.
Mimura N, Nurse L, McLean RF, Agard J, [5] Briguglio L, Lefale P, et al. Small islands. In: Parry ML, Canziani OF, Palutikof JP, van der Linden PJ, Hanson CE. Eds. Climate change 2007: impacts, adaptation and vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, 2007. p. 687-716.
Hales S, de Wet N, Maindonald J, Woodward [6] A. Potential effect of population and climate changes on global distribution of dengue fever: an empirical model. Lancet. 2002;360:830-834.
Confalonieri U, Menne B, Akhtar R, Ebi [7] KL, Hauengue M, Kovats RS, et al. Human health. In: Parry ML, Canziani OF, Palutikof JP, van der Linden PJ, Hanson CE. Eds. Climate change 2007: impacts, adaptation and vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, 2007. p. 391-431.

Dengue Bulletin – Volume 33, 2009 9
Implementing predictive models for domestic decision-making against DHF epidemics
Barclay E. Is climate change affecting dengue in [8] the Americas? Lancet. 2008;371:973-4.
Halide H, Ridd P. A predictive model for [9] dengue haemorrhagic fever (DHF) epidemics. Int J Environ Health Res. 2008;18(4):253-65.
Drake JM. Fundamental limits to the precision [10] of early warning systems for epidemics of infectious diseases. PLoS Medicine. 2005;2(5):e144.
Farrar J, Focks D, Gubler D, Barrera R, Guzman [11] MG, Simmons C, et al. Towards a global dengue research agenda. Trop Med Int Health. 2007;12:695-9.
Runge-Ranzinger S, Horstick O, Marx M, [12] Kroeger A. What does dengue disease surveillance contribute to predicting and detecting outbreaks and describing trends? Trop Med Int Health. 2008;13(8):1022-1041.
Nisalak A, Endy TP, Nimmannitya S, [13] Kalayanarooj S, Thisayakorn U, Scott RM, et al. Serotype-specific dengue virus circulation and dengue in Bangkok, Thailand from 1973-1999. Am J Trop Med Hyg. 2003; 68: 191-202.
Stephenson DB. Use of the “odds ratio” [14] for diagnosing forecast skill. Weather and Forecasting. 2000;15:221-232.
Klun JA, Strickman D, Rowton E, Williams [15] J, Kramer M, Roberts D, et al. Comparative resistance of Anopheles albimanus and Aedes aegypti to N,N-Diethyl-3-methylbenzamide (Deet) and 2-Methylpiperidinyl-3-cyclohexen-1-carboxamide (AI3-37220) in laboratory human-volunteer repellent assays. J Med Entomol. 2004;41(3):418-22.
Kay BH, Nam VS, Tien TV, Yen NT, Phong TV, [16] Diep VTB, et al. Control of Aedes vectors of dengue in three provinces of Vietnam by use of Mesocyclops (Copepoda) and community-based methods validated by entomologic, clinical, and serological surveillance. Am J Trop Med Hyg. 2002;66(1):40-48.
McConnell KJ, Gubler DJ. Guidelines on the [17] cost-effectiveness of larval control programs to reduce dengue transmission in Puerto Rico. Rev Panam Salud Publica. 2003;14(1):9-16.
Paeporn P, Komalamisra N, Deesin V, [18] Rongsriyam Y, Eshita Y, Thongrungkiat S. Temephos resistance in two forms of Aedes aegypti and its significance for the resistance mechanism. Southeast Asian J Trop Med Public Health. 2003;34(4):786-792.
Suaya JA, Shepard DS, Chang M-S, Caram [19] M, Hoyer S, Socheat D, Chantha N, Nathan MB. Cost-effectiveness of annual targeted larviciding campaigns in Cambodia against the dengue vector Aedes aegypti. Trop Med Int Health. 2007;12(9):1026-1036.
Thornes JE, Stephenson DB. How to judge [20] the quality and value of weather forecast products. Meteorological Application. 2001;8:307-314.
Central Board of Statistics (BPS). Makassar in [21] Figures, 2006 and South Sulawesi in Figures, 2006.
Costantini C, Badolo A, Ilboudo-Sanogo E. Field [22] evaluation and the efficacy and persistence of insect repellents DEET, IR3535, and KBR3023 against Anopheles gambiae complex and other Afrotropical vector mosquitoes. Trans R Soc Trop Med Hyg. 2004;98(11):644-52.
Fradin MS, Day JF. Comparative efficacy of [23] insect repellents against mosquito bites. N Engl J Med. 2002 July; 347 (1): 13-18.
Klun JA, Khrimian A, Debboun M. Repellent [24] and deterrent effects of SS220, Picaridin and DEET suppress human blood feeding by Aedes aegypti, Anopheles stephensi, and Phlebotomus papatasi. J Med Entomol. 2006;43(1):34-39.
Kalyanasundaram M, Mathew N. [25] N,N-diethyl phenylacetamide (DEPA): a safe and effective repellent for personal protection against hematophagous arthropods. J Med Entomol. 2006;43(3):518-525.

10 Dengue Bulletin – Volume 33, 2009
Implementing predictive models for domestic decision-making against DHF epidemics
Katz TM, Miller JH, Hebert AA. Insect [26] repellents: Historical perspectives and new developments. J Am Acad Dermatol. 2008;58(5):865-871.
W-Smith A, Schwartz E. Dengue in travellers. [27] New England Journal of Medicine 2005 Sept; 353 (9): 924-932. Brown M, Hebert AA. Insect repellents: An overview. J Am Acad Dermatol. 1997;36:243-249.
A-Donia M, Dechkovskaia AM, Goldstein [28] LB, A-Rahman A, Bullman SL, Khan WA. Co-exposure to pyridostigmine bromide, DEET, and/or permethrin causes sensorimotor deficit and alterations in brain acetylcholinesterase activity. Pharmacology, Biochemistry and Behavior. 2004;77:253-262.
Vijayaraghavan R, Rao SS, Suryanarayana MVS, [29] Swamy RV. Acute and subacute inhalation toxicity studies of a new broad spectrum insect repellent, N, N-diethylphenylacetamide. Toxicology. 1991;67:85-96.
Schaefer C, Peters PWJ. Intrauterine [30] Diethyltoluamide exposure and fetal outcome. Reprod Toxicol. 1992;6:175-176.
Hexsel CL, Bangert SD, Hebert AA, Lim [31] HW. Current sunscreen issues: 2007 Food and Drug Administration sunscreen labelling recommendations and combination sunscreen/insect repellent products. J Am Acad Dermatol. 2008;59(2):316-323.

Dengue Bulletin – Volume 33, 2009 11
Use of geographical information system (GIS) and global positioning system (GPS) for dengue and dengue
haemorrhagic fever control in Sri LankaG.A.J.S.K. Jayasooriyaa, S.M.L. Senaratnea, W.M.C.M. Wijesingheb,
P.H.D. Kusumawathiea#, J. Gunatilakec
aRegional Office, Anti Malaria Campaign, Dutugemunu Mawatha, Watapuluwa, Kandy, Sri Lanka
bEpidemiology Unit, Office of the Regional Director of Health Services, Kandy, Sri Lanka
cDepartment of Geology, University of Peradeniya, Peradeniya, Sri Lanka Physics Department, FMIPA, Hasanuddin University, Makassar 90245, Indonesia
Abstract
The dengue virus causing dengue fever (DF) and dengue haemorrhagic fever (DHF) is transmitted by the female mosquitoes – Aedes aegypti and Ae. albopictus. Because DF/DHF is a local and focal disease, identification of finer-scale risk areas and application of vector control interventions in these areas are important actions for disease prevention and control. The present study was carried out to: (a) identify DF/DHF risk levels of different Grama Niladari (GN) areas under the jurisdiction of the Medical Officer of Health (MOH), Kadugannawa area, Kandy district; and (b) determine the impact of Aedes larval control in DF/DHF high-risk GN areas on the overall DF/DHF burden in the MOH area. Ae. aegypti and Ae. albopictus density (Breteau index) in each GN area of MOH Kadugannawa was determined by immature (larvae and pupae) surveys. Details of suspected and serologically confirmed DF/DHF cases were collected from MOH Kadugannawa and georeferenced using global positioning system (GPS) receivers. Data on Ae. aegypti and Ae. albopictus density and DF/DHF cases were analysed and mapped using the geographical information system (GIS) to identify the DF/DHF risk levels in different GN areas of the MOH. With reference to risk mapping, health education and source reduction (interventions) were carried out in high-risk GN areas (areas with DF/DHF cases and Ae. aegypti prevalence) in July 2008. Kandy district showed an increasing trend of DF/DHF since 2001. The MOH area Kadugannawa also followed the same trend from January 2004 to July 2008, contributing 18.8%–37.5% of the monthly case load in the district in the period January–July 2008. Following the intervention in July 2008, MOH Kadugannawa showed a decreasing trend of DF/DHF during August–December 2008 and contributed 22.7%–8.8% of monthly DF/DHF cases in Kandy district. We conclude that identification of finer-scale DF/DHF risk areas using GIS and GPS and application of vector control interventions in high-risk GN areas is very useful for DF/DHF prevention and control.
Keywords: GIS and GPS; DF/DHF control; Sri Lanka.
#E-mail: [email protected]

12 Dengue Bulletin – Volume 33, 2009
Use of GIS and GPS for DF/DHF control in Sri Lanka
Introduction
Dengue fever (DF), dengue haemorrhagic fever (DHF) and dengue shock syndrome (DSS) are part of a disease complex caused by four serotypes – DENV 1-4. The virus is transmitted by female mosquitoes of Aedes aegypti and Ae. albopictus.[1] Thus, the occurrence of DF/DHF depends on the presence of the dengue virus, the vector mosquito species and a susceptible human population.
In Sri Lanka, DF was first reported in the early 1960s.[2] Since then, sporadic, progressively larger and more frequent DF/DHF outbreaks have occurred in the country. The morbidity, mortality and spatial distribution of the disease have increased considerably since 1995 with 15 434 suspected or serologically-confirmed DF/DHF cases and 88 deaths in the year 2004 alone. At present, many urban and semi-urban areas are endemic for DF/DHF while new areas are being invaded, making DF/DHF a major public health problem in the country.[3]
In the absence of a specific treatment or vaccine for DF/DHF, vector control remains the only option for disease prevention and control. Application of targeted vector control measures requires information on the geographical distribution and breeding habitats of the vector mosquito species. Because DF/DHF is a local and focal disease, identification of finer scale (Grama Niladari (GN) = smallest administrative unit in Sri Lanka) DF/DHF risk areas would be very helpful for undertaking cost-effective vector control measures. This study was carried out to (a) identify DF/DHF risk levels of different GN areas under the jurisdiction of the Medical Officer of Health (MOH), Kadugannawa, Kandy district; and (b) determine the impact of health education and source reduction in DF/DHF “high-risk” GN areas on the overall DF/DHF burden.
Materials and methods
Study area
The area selected for the study was the MOH Kadugannawa area in the Kandy district of Sri Lanka. The MOH area consists of 95 GN areas with an estimated mid-year population of 101 677 for the year 2006.[4] Kadugannawa area is endemic for DF/DHF with 49–194 cases reported annually during 2004–2007, contributing 7.9%–13.2% of the total annual case burden in Kandy district. The study area showed an increasing trend of DF/DHF from January 2004 to July 2008, based on both monthly and annual trends of DF/DHF in the district (Record at the Office of the Regional Director of Health Services in Kandy).
Prevalence of Ae. aegypti and Ae. albopictus in different GN areas of MOH, Kadugannawa
Aedes immature (larvae and pupae) surveys were carried out from January 2004 to December 2007 to detect the prevalence of Ae. aegypti and Ae. albopictus in each GN area of MOH Kadugannawa. During the surveys, a representative sample of 100 houses in each GN area was examined. All indoor and outdoor potential breeding habitats for Ae. aegypti and Ae. albopictus were examined, and up to 10 Aedes larvae and 10 pupae were randomly collected from each larvae/pupae positive container by dipping or pipetting, depending on the nature of the breeding habitat, for identification of the vector species.
If a particular container had <10 Aedes larvae/pupae, all larvae/pupae were collected. If a particular GN area did not report Ae. aegypti and Ae. albopictus in the first survey, two more surveys with 3–4 month intervals in

Dengue Bulletin – Volume 33, 2009 13
Use of GIS and GPS for DF/DHF control in Sri Lanka
high dengue transmission periods were carried out in order to confirm the presence/absence of Ae. aegypti and Ae. albopictus in that area. Aedes larvae were identified at the larval stage and pupae were allowed to develop to adult stage and identified using standard keys.[5] Ae. aegypti and Ae. albopictus density in each GN area was determined by calculation of the Breteau index (BI) for each species (number of Ae. aegypti/Ae. albopictus-positive containers per 100 houses) using multiple samples.
Determination of GN area-level distribution of DF/DHF cases
Details (name, age, sex and address) of all reported, suspected or serologically confirmed DF/DHF cases from 2004–2007 in the study area were collected in consultation with MOH Kadugannawa. These cases were geo-referenced, using global positioning system (GPS) receivers, at the point of residence of the patient.
Development of maps to identify the GN level-distribution of DF/DHF, Ae. aegypti, Ae. albopictus, and the DF/DHF risk levels of different GN areas in MOH Kadugannawa area
Data for DF/DHF cases and Breteau indices of Ae. aegypti and Ae. albopictus at the GN area spatial scale were analysed using a geographical information system (GIS), ArcView 9.0 software. Jayasooriya et al. (2008) reported that there was a strong correlation between the Breteau index of Ae. aegypti and DF/DHF in the study area, but no similar correlation between BI of Ae. albopictus and DF/DHF cases was observed.[6] Based on this
information, MOH area Kadugannawa was stratified as high-risk, medium-risk and low-risk and mapped accordingly. The high-risk GN areas were the areas with DF/DHF cases and with Ae. aegypti prevalence. The GN areas that were situated adjacent to Ae. aegypti-positive GN areas would undoubtedly have been infested with Ae. aegypti adults having emerged from the adjacent GN areas. Thus, the GN areas that were situated adjacent to the Ae. aegypti-positive GN areas were classified as medium-risk. The low-risk GN areas were the areas with DF/DHF cases and with Ae. albopictus but without Ae. aegypti in either the GN area itself or in adjacent GN areas.
Application of intervention (health education and elimination of vector breeding sites) in DF/DHF high-risk GN areas in MOH Kadugannawa area
With reference to the DF/DHF risk map, health education and elimination of breeding sites of Ae. aegypti and Ae. albopictus (intervention) were carried out in July 2008 in the DF/DHF high-risk GN areas. Health education was carried out through house-to-house visits by a team comprising a trained entomological assistant, public health staff and village volunteers. During these visits, each household in the area was made aware of the DF/DHF situation in the area, types of Ae. aegypti and Ae. albopictus breeding sites, and suitable measures to be taken for the elimination of vector breeding sites on their premises. Mosquito breeding sites encountered during these visits were promptly eliminated by emptying water storage containers, burning and/or burying discarded containers and cleaning domestic appliances such as refrigerators.

14 Dengue Bulletin – Volume 33, 2009
Use of GIS and GPS for DF/DHF control in Sri Lanka
Determination of the impact of intervention on overall DF/DHF incidence in the MOH Kadugannawa area
Annual data on DF/DHF from the MOH Kadugannawa area and the Kandy district area were plotted from 2004 to 2007 to identify the trends of DF/DHF in MOH Kadugannawa area and Kandy district. Monthly data on (a) DF/DHF cases in MOH Kadugannawa and Kandy district, and (b) per cent contribution of MOH Kadugannawa to the total disease burden in Kandy district were determined for the year 2008 to compare the (a) monthly trends of DF/DHF in MOH Kadugannawa and Kandy district, and (b) monthly percentage contribution of MOH Kadugannawa to the total DF/DHF burden in Kandy district, before and after the vector control intervention.
Results
Of the 95 GN areas in MOH Kadugannawa, neither Ae. aegypti nor Ae. albopictus were reported from three GN areas, namely, Haliyadda, Kirimetiya Estate and Pahala-yatigammana. Only Ae. aegypti was found in one GN area (Walgampaya) and both Ae. aegypti and Ae. albopictus were found in 17 GN areas, namely, Arambegama-west, Danture, Edanduwawa-east, Edaduwawa-west, Gannoruwa-east, Gannoruwa-central, Gurugama, Ihala-alagalla, Kadawatgama, Kadugannawa, Kiribathkumbura-east, K i r ibathkumbura-west , Kotabogoda, Munwathugoda, Pilimatalawa, Udaeriyagama-west and Waturakumbura. The remainder of the GN areas (n=74) reported only Ae. albopictus (Figures 1 and 2).
Figure 1: Distribution of Ae. aegypti and DF/DHF cases within the MOH Kadugannawa area by finer scale GN area

Dengue Bulletin – Volume 33, 2009 15
Use of GIS and GPS for DF/DHF control in Sri Lanka
Figure 2: Distribution of Ae. albopictus and DF/DHF cases within the MOH Kadugannawa area by finer scale GN area
DF/DHF cases were reported from both GN areas where Ae. aegypti was present and where Ae. aegypti was absent, although there is a correlation (r=0.5473) between Breteau index of Ae. aegypti and DF/DHF cases.[6] In our case, GN areas with Ae. aegypti contributed 44.9% to the total DF/DHF cases in MOH Kadugannawa with 11.3–12.0 cases per GN area, making these areas at high risk of DF/DHF (Table, Figures 1 and 3).
Apart from GN areas with Ae. aegypti present, the GN areas adjacent to areas with Ae. aegypti presence contributed 23.2% of DF/DHF cases, placing these areas at medium risk for DF/DHF. These areas, together with the Ae. aegypti-positive areas, contributed 68.1% of the total DF/DHF burden in the MOH area despite the fact that they constituted only 35 (36.8%) of the 95 GN areas (Figure 3).
Table: Number and percentage of DF/DHF cases from GN areas where Ae. aegypti/Ae. albopictus were present versus areas
where not collected
Aedes species
GN areas with
Aedes species
DF/DHF cases (%)
GN area without Aedes
species (%)
Ae. aegypti 1 12 (2.6) 12.0
Ae. aegypti + Ae. albopictus
17 192 (42.3) 11.3
Ae. albopictus 74 241 (53.1) 3.3
None 3 9 (2.0) 3.0
Total 95 454 (100)

16 Dengue Bulletin – Volume 33, 2009
Use of GIS and GPS for DF/DHF control in Sri Lanka
Of the 454 DF/DHF cases reported in MOH Kadugannawa from 2004–2007, 142 (31.3%) cases were from GN areas where Ae. aegypti was absent but with Ae. albopictus presence, and that were situated outside the typical flying range (<200 m) of Ae. aegypti. Because there is a very poor correlation between BI of Ae. albopictus and DF/DHF cases (r=–0.0469), based on the study by Jayasooriya, et al. (2008),[6] these GN areas were classified at low risk of DF/DHF (Figure 3).
Kandy district experienced DF/DHF epidemics in 2002, 2004, 2006 and 2008. MOH Kadugannawa area showed an increasing trend from 2001–2004. Since 2004, Kadugannawa has followed the same trend of DF/DHF as in Kandy district with epidemics in 2004 and 2006 (Figure 4). However, with the
application of intervention (health education and elimination of breeding sites of Ae. aegypti and Ae. albopictus in 18 high-risk GN areas) in July 2008, MOH Kadugannawa showed a downward trend of DF/DHF. This was in contrast to the upward trend in Kandy district (Kandy district included the data for MOH Kadugannawa). The percentage contribution of MOH Kadugannawa to the total DF/DHF incidence in Kandy district was comparatively high and fluctuated over the months from January to July 2008, ranging from 18.8% to 37.5% (before intervention). However, with the intervention in July 2008, the percentage contribution of MOH Kadugannawa to the total DF/DHF incidence in Kandy district decreased over the months July–December 2008, ranging from 22.7% to 8.8% (after intervention) (Figure 5).
Figure 3: Risk map of DF/DHF transmission of MOH Kadugannawa area

Dengue Bulletin – Volume 33, 2009 17
Use of GIS and GPS for DF/DHF control in Sri Lanka
Figure 4: Trend of DF/DHF in MOH Kadugannawa and in Kandy district
Figure 5: Pre- and post-intervention trends of DF/DHF in MOH Kadugannawa
and Kandy district (including MOH Kadugannawa) and the percentage
contribution of MOH Kadugannawa to the total DF/DHF incidence in
Kandy district
Num
ber o
f DF/D
HFca
ses
60
50
40
30
20
10
0
Jan-08
Months
Feb Mar
AprMay Jun Jul Au
g Sep OctNovDec-08
DF/DHF in Kandy districtDF/DHF in MOHKadugannawa
% contribution of MOHKadugannawa to the totalincidence of DF/DHF inKandy district
Discussion
The incidence of DF/DHF in an area depends on the occurrence and density of the vector species and serotype-specific herd immunity. In the present study, a high percentage (44.93%) of DF/DHF cases was observed in the GN areas where Ae. aegypti was present. The presence of a high correlation between BI of Ae. aegypti and DF/DHF cases indicated that 18 GN areas in MOH Kadugannawa were at high risk of DF/DHF transmission. Thus, application of uninterrupted larval control measures in these 18 GN areas was of utmost importance for the prevention and control of DF/DHF.
High incidence of DF/DHF was also observed in the GN areas that were situated adjacent to the Ae. aegypti-prevalent GN areas. These GN areas contributed 23.2% cases to the total DF/DHF cases in MOH Kadugannawa, making these GN areas – together with Ae. aegypti-prevalent GN areas – contributing 68.13% to the total DF/DHF cases in the MOH area. The GN areas that are situated adjacent to the Ae. aegypti (immatures)-positive areas are within the typical flight range (<200 m) of Ae. aegypti, thus enabling Ae. aegypti adults to invade these GN areas.
This resulted in a high DF/DHF transmission in the adjacent GN areas of Ae. aegypti immatures-positive GN areas, even in the absence of Ae. aegypti breeding sites, making these areas vulnerable to DF/DHF.
This indicated the necessity of adult vector control measures for DF/DHF prevention and control in such areas irrespective of the absence of Ae. aegypti breeding sites. With the impending socioeconomic changes, the spread of Ae. aegypti is most imminent in

18 Dengue Bulletin – Volume 33, 2009
Use of GIS and GPS for DF/DHF control in Sri Lanka
such GN areas. Thus, periodic entomological investigations in these GN areas are necessary to monitor the potential breeding sites of Ae. aegypti and for taking remedial measures.
Since there is a poor and negative correlation between BI of Ae. albopictus and DF/DHF cases, this species is less important as a vector of DF/DHF in MOH Kadugannawa. However, Ae. albopictus was reported to be naturally infected with dengue virus.[7,8] Apart from being a vector of DF/DHF, Ae. albopictus is an important vector of chikungunya[9] that emerged in an epidemic form in the country in 2006-2007.[10] Some GN areas of MOH Kadugannawa were also affected by the recent outbreak of chikungunya in 2007 (Regional Epidemiologist, Kandy, pers. com.). This shows that MOH Kadugannawa area is highly vulnerable to chikungunya as well as that 92 GN areas (96.8%) were infested with Ae. albopictus and/or Ae. aegypti. Thus, continuous disease and vector surveillance and community-based vector control interventions are necessary for the elimination of potential breeding sites of Ae. albopictus and Ae. aegypti in order to prevent and control the occurrence of chikungunya in this MOH area.
From 2001-2007, the MOH Kadugannawa area contributed 7.73%–21.35% cases to the total DF/DHF cases in Kandy district. MOH Kadugannawa area always followed the trend of DF/DHF in Kandy district, especially during the epidemic periods. Since mid-2008, Kandy district showed an increasing trend of DF/DHF; accordingly, MOH Kadugannawa also showed an increase of DF/DHF in July 2008. Application of health education and source reduction in the high-risk 18 GN areas resulted in (a) downward trend of DF/DHF in Kadugannawa area as against the upward trend in Kandy district; and (b) a reduction in percentage contribution of MOH
Kadugannawa to the total DF/DHF cases in Kandy district for the rest of the year.
During this intervention the GN area-wise distribution of DF/DHF, the presence of Ae. aegypti/Ae. albopictus, and the risk levels of DF/DHF in different GN areas were identified using GIS and GPS technology. These maps were very convenient in the identification of DF/DHF risk areas because: (a) they gave visual information on the DF/DHF risk levels of different GN areas that could be understood even by a non-technical person; and (b) these maps could be updated easily.
GIS-based dengue risk maps were developed in dengue endemic areas of South-East Asia and the Americas at spatial scales ranging from village, town, district to country.[11,12,13] Some of these studies reported the feasibility of using Aedes larval indices for prediction of spatial risk for dengue transmission[14,15] while others failed to find significant associations between Aedes larval density and spatial patterns of dengue incidence.[16,17] However, the present study shows the feasibility of using epidemiological and entomological data for finer scale spatial risk mapping and control of DF/DHF in Sri Lanka. Incorporation of entomological indices (Breteau indices of Ae. aegypti and Ae. albopictus) gave an advantage over the use of only DF/DHF case data in spatial risk mapping.
In MOH Kadugannawa, there were 33 GN areas with DF/DHF cases and Ae. albopictus. The DF/DHF cases in these GN areas were sporadic and were reported with longer time intervals, indicating that these cases were not of indigenous origin (imported cases from other dengue transmission areas). Another 27 GN areas reported Ae. albopictus but there was no DF/DHF case. Incorporation of entomological data in risk mapping helped

Dengue Bulletin – Volume 33, 2009 19
Use of GIS and GPS for DF/DHF control in Sri Lanka
to classify these GN areas as at “low risk” of DF/DHF. However, if only DF/DHF case data were used in risk mapping, these GN areas would have been identified as “high-risk” and recommended for more costly DF/DHF control measures such as space spraying and larviciding with insecticides. Incorporation of entomological data helped to identify the actual risk of DF/DHF transmission in each GN area, thus facilitating the application of DF/DHF control measures in a cost-effective manner.
In the preparat ion of r isk maps, accuracy of entomological data is of utmost importance. During the present study, trained entomological teams were deployed for entomological surveillance. These teams surveyed a representative sample of houses in each GN area and included all high-risk premises/institutions of vector breeding, such as schools, offices, religious places, tyre shops, city transport bus depots, etc.[18] Furthermore, if Ae. aegypti/Ae. albopictus immatures were not encountered during the first survey, two more such surveys were carried out in that particular GN area in order to confirm the presence/absence of Ae. aegypti/Ae. albopictus. Thus, it is very unlikely to get a negative result for a GN area that has the presence of Ae. aegypti/Ae. albopictus. However, regular entomological investigation in GN areas that are hitherto
negative for Ae. aegypti and updating the risk maps accordingly is important as vector mosquitoes infest new GN areas when environmental conditions become conducive for vector breeding.
In conclusion, GIS and GPS is a useful tool in DF/DHF prevention and control. These technologies can be used not only for risk mapping but also for spatial and space-time modelling to visualize and analyse mosquito vector and epidemiological data in operational dengue vector control programmes. Geographical information system softwares are becoming more user-friendly and now are complemented by free mapping software that provide access to satellite imagery and basic feature-making tools facilitating the generation of static maps as well as dynamic time-series maps. This will also enable disease control programmes to generate risk maps for other parameters such as exposure to dengue virus, to develop priority area classifications for vector control, and explore the socioeconomic associations of dengue risk.[19]
Acknowledgements
Financial support provided by the Small Grants Programme of the TDR/WHO (Grant No. SN 1176) is gratefully acknowledged.
ReferencesChavasse DC, Yap HH. [1] Chemical methods for the control of vectors and pests of public health importance. Document no. WHO/CTD/WHOPES/97.2. Geneva: World Health Organization, 1997.
World Health Organization. [2] Arthropod borne and rodent borne viral diseases: report of a WHO Scientific Group. Technical Report Series, 719. Geneva, WHO, 1985.

20 Dengue Bulletin – Volume 33, 2009
Use of GIS and GPS for DF/DHF control in Sri Lanka
Ministry of Health, Sri Lanka. [3] Framework for plan of action for prevention and control of dengue fever in Sri Lanka. Draft prepared by the Epidemiology Unit, Ministry of Health. Colombo: Ministry of Health, 2005.
Ministry of Health, Sri Lanka. [4] Mid year population estimates 2006. Colombo: Medical Statistics Unit, Ministry of Healthcare and Nutrition, 2006.
Barraud PJ. [5] The fauna of British India. Vol 5. London: Taylor and Francis, 1934. p. 138-143.
Jayasooriya GAJSK, Kusumawathie PHD, [6] Wijesinghe WMCW, Gunatilake J. Use of GIS to determine the GN level risk of dengue transmission in Kadugannawa MOH area, Sri Lanka. Proceedings of the Fifth National Symposium on Geo-Informatics. In: Jagath Gunatilake. Eds. Geo-Informatics Society of Sri Lanka and Post Graduate Institute of Science, University of Peradeniya, Sri Lanka, 2008. p. 149-157.
Rudnick A, Chan YC. Dengue type 2 virus in [7] naturally infected Aedes albopictus Mosquitoes in Singapore. Science. 1965;149:638-639.
Sergio Ibáñez-bernal, Baltasar Briseño, [8] John Paul Mutebi, Enid Argot , Guadalupe Rodríguez , Carmen Martinez-campos, Rafael Paz , Pedro de la Fuente-san Román, Roberto Tapia-conyer and Ana Flisser. First record in America of Aedes albopictus naturally infected with dengue virus during the 1995 outbreak at Reynosa, Mexico. Medical and Veterinary Entomology. 2008;11:305-309.
Ministry of Healthcare and Nutrition. [9] Fact sheet on Chikungunya. Colombo: Epidemiological Unit, Ministry of Health, 2009. http://www.epid.gov.lk/outbreak.htm – accessed 22 June 2010.
Ministry of Healthcare and Nutrition. [10] Investigation of the outbreak of chikungunya fever 2006/7. Colombo: Epidemiological Unit, Ministry of Health. http://www.epid.gov.lk/outbreak.htm - accessed 22 June 2010.
Rotela C, Fouque F, Lamfri M, Sabatier P, [11] Introini V, et al. Space-time analysis of the dengue spreading dynamics in the 2004 Tartagal outbreak, Northern Argentina. Acta Trop. 2007;103:1-13.
Ali M, Wagatsuma Y, Emch M, Breiman RF. [12] Use of a geographic information system for defining spatial risk for dengue transmission in Bangladesh: role for Aedes albopictus in an urban outbreak. Am J Trop Med Hyg. 2003;69:634-640.
Mondini A, Chiaravalloti-Neto F, Gallo Y [13] Sanches M, Lopes JCC. Spatial analysis of dengue transmission in a medium-sized city in Brazil. Rev Saude Publica. 2005;39:444-451.
Sanchez L, Vanlerberghe V, Alfonso L, Marquetti [14] MC, Guzman MG, et al. Aedes aegypti larval indices and risk for dengue epidemics. Emerg Infect Dis. 2006;12:800-806.
Chadee DD, Williams FLR, Kitron UD. Impact [15] of vector control on a dengue fever outbreak in Trinidad, West Indies, in 1998. Trop Med Int Health. 2005;10:748-754.
Sulaiman S, Pawanchee ZA, Arifin Z, Wahab [16] A. Relationship between Breteau and house indices and cases of dengue/dengue hemorrhagic fever in Kuala Lumpur, Malaysia. J Am Mosq Contr Assoc. 1996;12:494-496.
Teixeira MG, Barreto ML, Costa MCN, Ferreira [17] LDA, Vasconcelos PF, et al. Dynamics of dengue virus circulation: A silent epidemic in a complex urban area. Trop Med Int Health. 2002;7:757-762.
Kusumawathie PHD. Larval infestation of [18] Aedes aegypti and Ae. albopictus in six types of institutions in a dengue transmission area in Kandy, Sri Lanka. Dengue Bulletin. 2005;29:165-168.
Eisen L, Lozano-Fuentes S. Use of mapping and [19] spatial and space-time modelling approaches in operational control of Aedes aegypti and dengue. PLos Neglected Tropical Diseases. 2009;3(4):e 411.

Dengue Bulletin – Volume 33, 2009 21
Estimating the basic reproduction number of dengue transmission during
2002-2007 outbreaks in Bandung, IndonesiaAsep K. Supriatna#
Department of Mathematics, Padjadjaran University, Jl. Raya Bandung-Sumedang Km 21, Sumedang 45363, Indonesia
Abstract
In this paper a dengue transmission mathematical model has been developed. The model assumes that human population is divided into three sub-populations: susceptible, infective and recovered sub-population, while the mosquito population is divided into two sub-populations: susceptible and infective. It is also assumed that the survival rates for humans and mosquitoes decrease with age. Two basic reproduction numbers are derived, namely, the host-to-vector basic reproduction number and the host-to-host basic reproduction number. An illustration on how to estimate the basic reproduction numbers from dengue incidence data is presented using the dengue incidence data during the 2002–2007 dengue outbreaks in Bandung, Indonesia.
Keywords: Dengue transmission; mathematical model; basic reproduction number; Bandung, Indonesia.
#E-mail: [email protected], [email protected]
Introduction
Indonesia is an archipelago comprising of more than 13 660 islands and stretching from 06° 08’ N to 11° 15’ S latitude and 94° 45’ to 141° 65’ E longitude. It has tropical climate in all its regions. Indonesia experiences hot, humid weather throughout the year and is, therefore, endemic for vector-borne diseases including dengue transmitted by Aedes aegypti and Aedes albopictus. Since its first appearance in Jakarta and Surabaya in 1968, dengue fever (DF) and its more severe form,
dengue haemorrhagic fever (DHF), have been growing steadily, with low incidence rates initially,[1,2] to a significant high incidence rate. Other changing patterns of dengue epidemiology have now been recognized which pose challenges for the control of this infection and have made difficult the understanding and control of the disease, let alone eliminating it.[3]
Although the disease is endemic throughout the country, most dengue infections have occurred in urban areas where approximately

22 Dengue Bulletin – Volume 33, 2009
Estimating the basic reproduction number of dengue transmission in Bandung, Indonesia
one third of the 240 million people of Indonesia live. Almost 60% of them live on the island of Java. Bandung, as one of the biggest cities in Java has been experiencing severe dengue infection ever since it was reported for the first time in 1969.[4] A recent study in a cohort of adult population shows that all the four known serotypes are circulating in Bandung, with DENV-2 predominating.[5]
For the prevention and control of dengue epidemic in Indonesia, in general, there are three critical strategies: (i) DF/DHF case and vector surveillance; (ii) disease management; and (iii) changing behaviours of communities and building partnerships in order to reduce the presence, or improve the management, of larval habitats.[6] Various applications of insecticides, including the application of ultra-low volume (ULV), have been used in order to reduce the larval and adult populations of the vectors.[7,8] Nonetheless, the dengue incidence rates, number of reported cases and deaths, and affected areas under DHF during the period 1992–2004 were alarming.
Simulation of the application of ULV shows that the abrupt change of vector mortality due to ULV will result in a delay of the endemic stage but will only slightly reduce the severity of the infection.[9] A similar result is also shown for a different scenario in which dengue re-infection is possible.[10] This indicates that different strategies for control are needed to improve the current situation for combating the disease. One possible control strategy is through vaccination.[11]
Some scientists argue that in the preparation for vaccine trials it is important to understand the epidemiology of the disease in an adult population where most of the individuals have been exposed to one or more DENV infections.[5] Other theoretical studies have been made to anticipate the application of a tetravalent vaccine currently being
developed[12,13] in an attempt to eliminate the disease, such as the study on the effects of vaccination in single serotype and in multiple serotype dengue infections[14-16] and in an age-structured population.[17]
One important epidemiological concept related to minimum vaccination coverage is the basic reproduction number.[18] The present paper discusses a method of how to compute this basic reproduction number, which, in dengue cases, is slightly difficult due to the presence of the indirect transmission of the disease and other complications such as different serotypes of the agents. We then apply the theory to estimate the value of the basic reproduction number during the period of dengue outbreaks (2002–2007) in Bandung. In the next section we develop a mathematical model for dengue transmission as a basis for the computation of the basic reproduction number used in the subsequent sections.
Mathematical model
It is well known in literatures that in a direct-transmission disease model the threshold number 0R can be estimated by the ratio of life expectancy and mean age at infection.[19,20] Examples are abundant, including estimation that considers a direct transmission in an age-structured model[21]. In this paper we will show that, if the survival functions of humans and mosquitoes are exponentially negative, then this estimation in the direct-transmission model generalizes into the indirect-transmission model. The method used in this paper gives a practical way to obtain the estimate of the basic reproduction number. This finding is important since the minimal effort to eliminate the disease depends on the basic reproduction number through the relation 01 1/cp R> − .[18,19,22]

Dengue Bulletin – Volume 33, 2009 23
Estimating the basic reproduction number of dengue transmission in Bandung, Indonesia
The model discussed here is the generalization of the model in a study by Fred Brauer[21] to include recovery compartment and indirect transmission of the disease, such as in dengue disease transmission. To formulate the model, let us assume that the host population
HN is divided into three compartments, namely, susceptible ( HS ), infective ( HI ) and recovered assumed immune individuals ( HR ). We assume that vectors remain infective for life due to their short life period compared with the duration of the disease, and hence the vector population only comprises two compartments, namely, susceptible ( VS ) and infective ( VI ). Recruitment rate for the host and the vector is HB and VB , with the death rate Hµ and Vµ , respectively. The disease transmission probability from vector to host and from host to vector is given by
Hb and Vb . Suppose
there exist non-negative monotonically non-increasing functions of age, ( )HQ a and
( )VQ a , describing the fraction of host and vector population, respectively, who survive to the age of a or more, such that (0) 1HQ = and (0) 1VQ = . Since the host life expectancy
is finite, such that 0( )
H H Q a da L
∞=∫ and
0( )
H aQ a da
∞< ∞∫ , and assuming the initial
conditions for the host population is given by (0)HN , then we have:
0
0( ) ( ) ( )
t
H H H H N t N t B Q a da= + < ∞∫ , (1)
with 0 ( ) (0) ( )H H HN t N Q t= . Furthermore, since the per capita rate of infection in the population at time t is ( )H VI tb , then the number of those susceptible at time t is given by:
( )0
0( ) ( ) ( )
tH Vt a
t I s dsH H H H
S t S t B Q a e dab
−−∫= + ∫ , (2)
with 0( )0 ( ) (0) ( )
tH VI s ds
H H HS t S Q t eb−∫= . And if we
assume that the rate of recovery is γ , then the number of the infective host at time t is given by:
( )0
0( ) ( ) ( ) 1
t tH Vt a t a
t I s ds dsH H H H
I t I t B Q a e e dab γ
− −− − ∫ ∫= + −
∫ , (3)
with 00 ( ) (0) ( )t
dsH H HI t I Q t e
γ−∫= . Furthermore, since ( ) ( ) ( ) ( )H H H HR t N t S t I t= − − , then we have:
( )0
0( ) ( ) ( ) 1 1
t tH Vt a t a
t I s ds dsH H H H
R t R t B Q a e e dab γ
− −− − ∫ ∫= + − −
∫ , (4)
with 0 0 0 0( ) ( ) ( ) ( )H H H HR t N t S t I t= − − . Equations (2) to (4) constitute transmission dynamics in the host population. Analogously, the transmission dynamics in the vector population is governed by the following equations:
0
0( ) ( ) ( )
V V V V N t N t B Q a da
∞= + ∫ , (5)
with 0 ( ) (0) ( )V V VN t N Q t= ,
( )0
0( ) ( ) ( )
tV Ht a
t I s dsV V V V
S t S t B Q a e dab
−−∫= + ∫ , (6)
with 0( )0 ( ) (0) ( )
tV HI s ds
V V VS t S Q t eb−∫= , and
( )0
0( ) ( ) ( ) 1
tV Ht a
t I s dsV V V V
I t I t B Q a e dab
−− ∫= + −
∫ , (7)
with 0 ( ) (0) ( )V V VI t I Q t= .
It is clear that 0 ( )t Hlim N t→∞ , 0 ( )t Hlim S t→∞ 0 ( )t Hlim I t→∞ and 0 ( )t Hlim R t→∞ are all equal to
zero, so does the same thing for the vector. The full model for the host-vector SIR

24 Dengue Bulletin – Volume 33, 2009
Estimating the basic reproduction number of dengue transmission in Bandung, Indonesia
(susceptible-infective-recover) model is given by equations (2) to (4) and (6) to (7). If 0γ = , then the model reduces to a host-vector SI (susceptible-infective) model. It is shown in Appendix A that there is a non-trivial endemic equilibrium * *( , )H VI I satisfying:
**
0( ) 1 H V
I a aH H H
I B Q a e e dab γ∞ − − = − ∫ , (8)
**
0( ) 1 V H
I aV V V
I B Q a e dab∞ − = − ∫ , (9)
if 0 1Rγ > , in which:
( )( )0 0 0( ) ( )
aH H H V V V
R B aQ a e da B aQ a daγ γb b∞ ∞−= ∫ ∫ . (10)
It is also shown that the threshold number 0Rγ also determins the stability of the resulting
endemic equilibrium point.[23] In the following section we show that the value of the basic reproduction number can be estimated as the ratio of life expectancy ( HL and VL ) and mean age at infection ( Ha and Va ), which usually are available in the field or literature.
The estimation of the basic reproduction number
We assume that the survival rates for humans and mosquitoes decrease with age, satisfying
( ) H aHQ a e µ−= and ( ) V a
VQ a e µ−= respectively. SI model is not relevant for dengue infection, but we do the calculation of SI model for certain reasons in the next section. To facilitate calculation for both SIR and SI models, we def ine ( )( ) ( ) H Ha aa
H HQ a Q a e e eµ γγ γ − + −Μ−= = = . Following [19] and [20], the mean age at
infection is given by the inverse of the force of
infection, hence *
1H
H V
aIb
= and
*
1V
V H
aIb
=.
Considering (8) and (9), consequently the threshold number in (10) can be written as:
( )( )
( )( )( )( ) ( )( )
* *
* *
0 00
* *0 0
* *
0 0
0 0
( ) ( )
1 1( ) ( )
( ) ( )
( ) 1 ( ) 1
H V V H
H V V H
H H H V V V
I a I a
H H V V H V
H H H V V V
I a I aH V
B aQ a da B aQ a daR
e eB Q a da B Q a daI I
I aQ a da I aQ a da
Q a e da Q a e da
γ
γ
b bγ
γ
b bγ
b b
b b
∞ ∞
− −∞ ∞
∞ ∞
∞ ∞− −
= − −
=− −
∫ ∫
∫ ∫
∫ ∫∫ ∫
which can be further simplified as:
2 2 2 2
0
1 1 1 1 1 1 1 1
111 1 1 1
1 1 1 1
1 1 1 1
H V H VH V H V
V VH HVH
H VH V
H H V VH VH V
H VH VH
a a a aR
aa
a a a a
a
γ µ µ
µ µ
µ µ µ µ
µµ
Μ Μ = =
+ −Μ + −Μ − −
Μ Μ + + Μ Μ + +
= Μ + + Μ
1 11 1V H H V Va a aµ
= + +
Μ
Finally we have:
0 1 1 VH
H V
LLRa a
γ = + +
, (11)
where 1H
H
L =Μ
and 1V
V
Lµ
= .
Next, we define 0 0R Rγγ ≡ . We note
that 0R γ and 0Rγ have the same threshold number, i.e. 0 01 1R Rγ
γ> ⇔ > . Observe also that 0R γ conforms more to the definition of the basic reproduction number in [19, eq. 5.9 p. 75]. In this regard, the basic reproduction number is the multiplication factor in n generation measured on a “per generation” basis. Alternatively, 0R γ can be interpreted as

Dengue Bulletin – Volume 33, 2009 25
Estimating the basic reproduction number of dengue transmission in Bandung, Indonesia
the geometric average of the expected number of new cases in human population caused by one infected mosquito during its entire period of infectiousness and the expected number of new cases in the mosquito population caused by one infected human during his/her entire period of infectiousness.
However, some authors have pointed out that the basic reproduction number measured per generation basis is less useful when the control effort needed to eliminate a host-vector disease (such as dengue) is to be targeted at only the host population.[24] They argue that the expected number of secondarily infected hosts that results from a single infected host is the square of the host-vector basic reproduction number, since two generations are required to transmit an infection from host to host: the first generation due to infection from a host to the vector population and the second generation due to infection from infected vectors back
to the host population. Hence, 0 0R Rγγ ≡ is
the host-to-vector basic reproduction number and 0Rγ is the host-to-host basic reproduction number. We note that using host-vector basic reproduction certainly underestimates the control effort needed to eliminate the disease, if the only target of control is host population, such as in host vaccination programme, since the resulting minimum vaccination coverage
is given by: 0 0
1 11 1cpR Rγ
γ
= − < − .[22]
Note that i f we derive the basic reproduction number directly from (10) then we have:
0 2 2( )V VH H
H V
BBRγ bbµ γ µ
= +
. (12)
However, i f we derive the basic reproduction number using the next generation matrix in [19, eq. 5.9 p. 75; see also Appendix B], then we have:
01 1
( )G VH
H VV H H V
BBR b bµ µ µ γ µ
= +
. (13)
It can be verified that 0GR is comparable
to the known form of the basic reproduction number.[10] If we denote 0
0R as the basic reproduction number for the SI model, that is 0Rγ with 0γ = , then we have the following relation:
00 0 0
GR R Rγ < < , (14)
with
00 0
0 0
1G
HG
H
R RR Rγ
µ γµ+
= = > , (15)
which consequently is,
00 0 0GR R Rγ= . (14)
The next section will illustrate the implementation of the theory developed in this section with the data during the 2002–2007 dengue outbreaks in Bandung.
The basic reproduction number in Bandung, Indonesia
We collected data consisting of the number of suspected and/or confirmed dengue patients from three major hospitals in Bandung, Advent, Muhammadiyah and St. Yusuf

26 Dengue Bulletin – Volume 33, 2009
Estimating the basic reproduction number of dengue transmission in Bandung, Indonesia
Hospitals, during the dengue outbreaks from 2002 up to 2007. We also collected data from all existing puskesmas (community health centres). Both data sets consist of patients’ name and age at the time they registered at the hospital or puskesmas due to dengue illness symptoms.
A total of 22 981 patients, comprising 12 030 patients from hospitals and 10 951 patients from puskesmas, were analysed.
The yearly patients’ distribution is shown in Figure 1. During the period 2002–2005, this yearly number of patients was approximately 0.15% of the total population of the city for the respective year.
Figure 2 shows that during the period 2002–2007, the dengue incidence steadily increased for three different classes: babies/toddlers, teenagers and adults. This figure reveals that marginal increase of the older
Figure 1: Total number of patients per year from hospitals (left) and from puskesmas (right)
Figure 2: Total number of patients per year for different classes: babies/toddlers, teenagers and adults

Dengue Bulletin – Volume 33, 2009 27
Estimating the basic reproduction number of dengue transmission in Bandung, Indonesia
class is slightly greater that those for the younger classes. A similar trend was also observed in other regions of Indonesia[25] and was consistent with those reported on a nationwide scale that there is a shifting of age of infection to older classes.[3,26] This is true for other countries as well.[27,28]
It remains unclear whether the data from hospitals and puskesmas overlap or are mutually exclusive. To handle this, we consider those data sets as samplings taken from two different places. We will compare the resulting basic reproduction numbers computed using those different data sets and our formula. To compute the basic reproduction number, our formula needs the value of the human’s average age at infection. This can be done directly from the data sets we collected from the hospitals and puskesmas as follows.
We model the survival of the human after catching the disease by the Weibull distribution
function given by 1
( )ttf t e
bbab
a a
− − =
, with
mean 1 1µ ab
= Γ +
. Statistical analysis
shows that our data fit to this distribution with the resulting parameters given in Table 2
along with the corresponding curves in Figures 3 and 4. The left part of Figure 3 shows the Weibull curves from hospital data sets 2002–2006. The curves from bottom to top, in increasing order, indicate the outbreaks in 2002, 2003, 2004, 2005 and 2006, consecutively. The figure reveals that there is a shifting of peaks from higher age at infection to lower age at infection and an increase of peaks during the periods 2002 to 2004, slight shifts to a higher age at infection in 2005 and then shifts again to a lower age at infection in 2006. The data from the puskesmas does not give a clear pattern.
Figure 3: The distribution of suspect and confirmed dengue patients
Figure 4: Mean age at infection decreases during the outbreak period 2002–2005
followed by increase in the periods afterwards

28 Dengue Bulletin – Volume 33, 2009
Estimating the basic reproduction number of dengue transmission in Bandung, Indonesia
Table 1: The parameters of the Weibull human survival model during the outbreak period 2002–2007
Source of data set Year a b µ
Hospitals
2002 25.56 1.62 22.89
2003 24.88 1.49 22.48
2004 23.25 1.34 21.35
2005 20.90 1.29 19.33
2006 22.35 1.28 20.70
Puskesmas
2004 22.68 1.21 21.29
2005 19.83 1.25 18.46
2006 21.18 1.32 19.50
2007 23.04 1.21 21.63
Detail computation has been described by Rubiana.[29]
Another human parameter we need to find the estimation of the basic reproduction number is the rate of recovery γ . The value of the average infectious period, that is, the inverse of the rate of recovery, is believed to be between three and eight.[30] Other scientists suggest that this average infectious period is six days,[31] and hence human recovery rate is 1/6 per unit time.
We also need the Ae. aegypti life expectancy. There are numerous values of life expectancy used in literatures, ranging from three days – in the field – to more than 90 days – when reared in laboratory.[32] Maciel-de-Freita et al.[32] also pointed out that Ae. aegypti must survive for periods longer than the sum of the initial non-feeding period plus the virus’ extrinsic incubation period in order to be able to transmit the disease to another human. Nulliparous females usually do not blood-feed for ≥2 days and the extrinsic incubation period of dengue virus is at least
10 days.[33,34] Taking into account those facts, many authors argued that the life expectancy of the mosquitoes must be at least 12 days. We will assume that the value of this life expectancy in the field is approximately 14 days as many scientists believe.[35,36,37] We also assume that to enable transmission, the average at infection for the mosquitoes is as early as the second day of their adult life stage. The resulting basic reproduction numbers for various times of outbreak using different sampling data sets are figured out in Table 2.
Concluding remarks
The estimation of dengue basic reproduction number in Bandung presented in this paper is considered to be among the first attempts to be made for such a study. The resulting basic reproduction numbers derived by the present method, 0Rγ and 0
GR in Table 2, are noticeably higher than the known estimated basic

Dengue Bulletin – Volume 33, 2009 29
Estimating the basic reproduction number of dengue transmission in Bandung, Indonesia
Table 2: The basic reproduction number during the outbreak period 2002–2007
Source of data set Year Ha
HL
Host-to-vector reproduction number
Host-to-host reproduction number
00R 0R 0GR 00R 0R 0
GR
Hospitals
2002 22.89 72.50 0.1667 5.7740 2.8294 4.0419 33.3386 8.0057 16.3371
2003 22.48 72.52 0.1667 5.8145 2.8295 4.0561 33.8078 8.0059 16.4518
2004 21.35 72.54 0.1667 5.9314 2.8295 4.0967 35.1813 8.0062 16.7829
2005 19.33 72.56 0.1667 6.1669 2.8296 4.1773 38.0300 8.0068 17.4499
2006 20.70 72.58 0.1667 6.0042 2.8295 4.1218 36.0502 8.0064 16.9891
Puskesmas
2004 21.29 72.54 0.1667 5.9378 2.8295 4.0989 35.2579 8.0062 16.8012
2005 18.46 72.56 0.1667 6.2806 2.8297 4.2157 39.4453 8.0071 17.7719
2006 19.50 72.58 0.1667 6.1463 2.8296 4.1703 37.7764 8.0067 17.3916
2007 21.63 73.39 0.1667 5.9282 2.8295 4.0956 35.1438 8.0061 16.7739
Ha = Human’s average age at infection are computed from the data sets
HL = Human’s life expectancy is provided by the Indonesian Bureau of Statistics
Va = 2 days
VL = 14 days
reproduction numbers for dengue infection from a neighbouring country, Singapore,[38] and from other countries.[31,39,40,41] In general, the estimates of dengue reproduction numbers vary considerably between studies while the reasons behind this variability are still not well-explained.[42] However, we can argue that our results are certainly higher than those of others since we follow the argument by Roberts MG, et al.[24] that the host-vector basic reproduction number should be computed as a second-generation host-to-host basic reproduction number when it is only host population that is to be controlled.
The result presented here is a crude approximation of the true value of the basic reproduction number since we ignored some details of the structure and transmission mechanism of the disease. Other factors that are worthy of being taken into account are the intrinsic and extrinsic incubation times. By taking these factors into consideration we predict that the resulting basic reproduction number will be discounted by the delay time of incubation periods (see [17] as an illustration). Other factors, such as the presence of multiple strains, may also increase the realism of the model, and hence, refine the estimate of the basic reproduction number.

30 Dengue Bulletin – Volume 33, 2009
Estimating the basic reproduction number of dengue transmission in Bandung, Indonesia
Acknowledgements
This work was supported financially by the Indonesian Government (the Directorate of Higher Education) through Penelitian Hibah Kompetensi – Batch 1, 2008–2010. The author would like to thank an anonymous referee who read the earlier version of the paper and made constructive suggestions to improve the quality of the paper.
Appendix A
Since we are interested in the long-term behaviour of the model, let us consider the limiting condition of the system (1) to (7). Whenever t →∞ , the system has the same dynamic behaviour as the following system:
( )
0( ) ( )
tH Vt a
I s dsH H H
S t B Q a e dab
−∞ −∫= ∫ , (A.1)
( )
0( ) ( ) 1
tH Vt a
I s ds aH H H
I t B Q a e e dab γ−
∞ − − ∫= −
∫ , (A.2)
( )
0( ) ( ) 1 1
tH Vt a
I s ds aH H H
R t B Q a e e dab γ−
∞ − − ∫ = − −
∫ ,
(A.3)
( )
0( ) ( )
tV Ht a
I s dsV V V
S t B Q a e dab
−∞ −∫= ∫ , (A.4)
( )
0( ) ( ) 1
tV Ht a
I s dsV V V
I t B Q a e dab
−∞ − ∫= −
∫ . (A.5)
Considering that we are only interested in the infective compartments without loss of generality, we can concentrate only on the sub-system (A.2) and (A.5), in which the equilibrium of the sub-system is given by
* *( , )H VI I with *HI and *
VI satisfying
** *10
( ) 1 ( )H V I a a
H H H V I B Q a e e da F Ib γ∞ − − = − ≡ ∫ , (A.6)
** *20
( ) 1 ( )V H I a
V V V H I B Q a e da F Ib∞ − = − ≡ ∫ . (A.7)
Note that (A.6) can be written as a composition,
*
0( ) 1
* *1 2 0
( ) ( ) 1 I sV H
H V V B Q s e ds a a
H H H H I F F I B Q a e e da
bbγ
∞ − − −∞ − ∫= = −
∫ . (A.8)
It is easy to see that (0,0) is the trivial or disease-free equilibrium. To find the non-trivial endemic equilibrium we could observe that
1 2F F is bounded, satisfying 1 2( ) 0H
d F FdI
>
and 2
1 22
( ) 0H
d F FdI
< , meaning that it is increasing
and concave down. Therefore, a unique non-trivial value of *
HI exists if and only
if 1 2( )(0) 1H
d F FdI
> , which is equivalent to
( )( )0 0( ) ( ) 1
aH H H V V V
B aQ a e da B aQ a daγb b∞ ∞− >∫ ∫
or simply 0 1Rγ > , with 0Rγ being the threshold number as in equation (10).
Appendix B
Let ijk denote the expected number of new cases of type i , caused by one infected individual of type j , during the entire period of infectiousness. In this case,
{ }, 1, 2i j∈ . Moreover, let type 1 be infection in host and type 2 be infection in vector, and by considering the exponential survival rates
( ) H aHQ a e µ−= and ( ) V a
VQ a e µ−= , then we have:
11 0k = , *12
1H H
V
k Sbµ
= , *
211
( )V VH H
k Sbµ γ
=+
,

Dengue Bulletin – Volume 33, 2009 31
Estimating the basic reproduction number of dengue transmission in Bandung, Indonesia
22 0k = . In a completely virgin population, the
steady states population are given by * HH
H
BSµ
=
and * VV
V
BSµ
= . Theorem 5.3 of [19] implies that
( ) 21
12
00ij
kK k
k
= =
is the next generation
matrix, with the dominant eigen value given by:
0 12 211 1
( )VH
G H VH V V H H
BBR k k b bµ µ µ µ γ
= = +
, (B.1)
which is obviously the square root of the basic reproduction number in (13).
ReferencesHotta S, Miyasaki K, Takehara M, Matsumoto [1] Y, Ishihama Y, Tokuchi M et al. Clinical and laboratory examinations on a case of “hemorrhagic fever” found in Surabaja, Indonesia, in 1968. Kobe J Med Sci. 1970;16(4):203-210.
Kho LK, Wulur H, Himawan T, Thaib S. Dengue [2] haemorrhagic fever in Jakarta (follow up study). Paediatr Indones. 1972;12(1):1-14.
Setiati TE, Wagenaar JFP, de Kruif MD, Mairuhu [3] ATA, van Gorp ECM, Soemantri A. Changing epidemiology of dengue haemorrhagic fever in Indonesia. Dengue Bulletin. 2006;30:1-14.
Rivai A, Hamzah S, Rahman O, Thaib S. Dengue [4] and dengue haemorrhagic fever in Bandung. Paediatr Indones. 1972;12(1):40-48.
Porter KR, Beckett CG, Kosasih H, Tan [5] RI, Alisjahbana B, Rudiman PI, Widjaja S, Listiyaningsih E, Ma’Roef CN, McArdle JL, Parwati I, Sudjana P, Jusuf H, Yuwono D, Wuryadi S. Epidemiology of dengue and dengue hemorrhagic fever in a cohort of adults living in Bandung, West Java, Indonesia. Am J Trop Med Hyg. 2005;72(1):60-66.
Kusriastuti R, Sutomo S. Evolution of dengue [6] prevention and control programme in Indonesia. Dengue Bulletin. 2005;29:1-6.
Suroso T. Dengue haemorrhagic fever in [7] Indonesia – epidemiological trends and development of control strategy. Dengue Bulletin. 1996;20:35-40.
World Health Organization, Regional Office for [8] South-East Asia. Dengue/dengue haemorrhagic fever prevention and control programme in Indonesia: report of an external review, Jakarta, 5-19 June 2000. Document SEA-Haem. Fever-73/SEA-VBC-79. New Delhi: WHO/SEARO, 2001.
Esteva L, Vargas C. Analysis of a dengue [9] disease transmission model. Math Biosci. 1998;150:131.
Soewono E, Supriatna AK. A Two-dimensional [10] model for the transmission of dengue fever disease. Bull Malays Math Sc Soc. (Second Series) 2001;24:49-57.
Clarke T. Breakbone fever. [11] Nature. 2002;416:672.
Kinney RM, Huang CY. Development of [12] new vaccines against dengue fever and Japanese encephal i t i s . Inter v i rology. 2001;44:176-191.
Konishi E, Kosugi S, Imoto J. Dengue tetravalent [13] DNA vaccine inducing neutralizing antibody and anamnestic responses to four serotypes in mice. Vaccine. 2006;24(12):2200-2207.

32 Dengue Bulletin – Volume 33, 2009
Estimating the basic reproduction number of dengue transmission in Bandung, Indonesia
Soewono E, Supriatna AK. [14] Paradox of vaccination predicted by a simple dengue disease model. In: M.C. Joshi et al. Eds. Industrial Mathematics. New Delhi: Narosa, 2006. p. 459.
Nuraini N. [15] Mathematical models for external and internal dengue disease transmission. Ph.D. Dissertation, ITB Bandung – Indonesia (unpublished, in Indonesian), 2008.
Supriatna AK, Nuraini N, Soewono E. [16] Mathematical models of dengue transmission and control: a survey. In: Basak Ganim and Adam Reis (eds.), Dengue Virus: Detection, Diagnosis and Control. New York: Nova Science Publishers. (in press).
Supriatna AK, Soewono E, van Gils SA. A two-[17] age-classes dengue transmission model. Math Biosci. 2008;216:114-121.
Anderson RM, May RM. [18] Infectious diseases of humans. Oxford: Oxford University Press 1991.
Diekmann O, Heesterbeek JAP, [19] Mathematical epidemiology of infectious diseases. New York: John Wiley and Sons, 2000.
Nishiura H. Mathematical and statistical [20] analyses of the spread of dengue. Dengue Bulletin. 2006;30:51-67.
Brauer F. A model for an SI disease in an age-[21] structured population. Disc Cont Dyn Sys-Ser B. 2002;2:257-264.
Scherer A, McLean A. Mathematical model [22] of vaccination. British Medical Bulletin. 2002;62:187-199.
Supriatna AK, Soewono E. Analysis of an [23] SIR dengue transmission model with age-dependent survival rates (in prep.)
Roberts MG, Heesterbeek JAP, A new method [24] for estimating the effort required to control an infectious disease. Proc Roy Soc Lond. 2003;B270:1359.
Corwin AL, Larasati RP, Bangs MJ, Wuryadi [25] S, Arjoso S, Sukri N, Listyaningsih E, Hartati S, Namursa R, Anwar Z, Chandra S, Loho B, Ahmad H, Campbell JR, Porter KR, Epidemic dengue transmission in southern Sumatra, Indonesia. Trans R Soc Trop Med Hyg. 2001;95:257.
Suwandono A, Kosasih H, Nurhayati, [26] Kusriastuti R, Harun S, Ma’roef C, Wuryadi S, Herianto B, Yuwono D, Porter KR, Beckett CG, Blair PJ. Four dengue virus serotypes found circulating during an outbreak of dengue fever and dengue haemorrhagic fever in Jakarta, Indonesia, during 2004. Trans R Soc Trop Med Hyg. 2006;100(9):855-62.
Ooi EE, Goh KT, Gubler DJ, Dengue prevention [27] and 35 years of vector control in Singapore. Emerg Infect Dis. 2006;12:887.
Patumamond J, Tawichasri C, Nopparat S, [28] Dengue hemorrhagic fever in Uttaradict, Thailand. Emerg Infect Dis. 2003;9:1348.
Rubiana I. Age-structured model of dengue [29] patients in Bandung during 2002-2007 dengue outbreaks. S1 Thesis, Dept. Mathematics, University Padjadjaran 2008 (unpublished, in Indonesian).
Halstead SB. [30] Dengue. In: Tropical and geographical medicine. Warren KS, Mahmoud AAF. Eds. New York: McGraw-Hill, 1990. p. 675-684.
Marques CA, Forattini 0P, Massad E. The basic [31] reproduction number for dengue fever in Sao Paulo state, Brazil: 1990- 1991 epidemic. Trans R Soc Trop Med Hyg. 1994;88:58-59.
Maciel-de-Freita R, Codeco CT, Lourenco-de-[32] Oliveira R. Body size-associated survival and dispersal rates of Aedes aegypti in Rio de Janeiro. Med Vet Entomol. 2007;21:284-292.
Kuno G. Review of the factors modulating [33] dengue transmission. Epidemiological Review. 1995;17:321-335 .

Dengue Bulletin – Volume 33, 2009 33
Estimating the basic reproduction number of dengue transmission in Bandung, Indonesia
Salazar MI, Richardson JH, Sánchez-Vargas [34] I, Olson KE, Beaty BJ. Dengue virus type 2: replication and tropisms in orally infected Aedes aegypti mosquitoes. BMC Microbiology. 2007;7:9.
Feng Z, Velasco-Hernandez JX . Competitive [35] exclusion in a vector-host model for the dengue fever. J Math Biol. 1997;35:523-544.
Nuraini N, Soewono E, Sidarto KA. A [36] mathematical model of dengue internal transmission process. J Indones Math Soc. 2007;13:123-132.
Nuraini N, Soewono E, Sidarto KA. Mathematical [37] model of dengue disease transmission with severe DHF compartment. Bull Malays Math Sci Soc. 2007;(2)30(2):143-157.
Hsieh YH, Ma S. Intervention measures, [38] turning point, and reproduction number for dengue, Singapore, 2005. Am J Trop Med Hyg. 2009;80(1):66-71.
Koopman JS, Prevots DR, Vaca Marin MA, [39] Gomez Dantes H, Zarate Aquino ML, Longini IM, Sapulveda Amor J. Determinations and predictors of dengue infection in Mexico. Am J Epidemiology. 1991;133:1168-1178.
Massad E, Coutinho FAB, Burattini MN, Lopez [40] LF. The risk of yellow fever in a dengue-infested area. Trans R Soc Trop Med Hyg. 2001;95:370-374.
Favier C, Degallier N, Rosa-Freitas MG, [41] Boulanger JP, Costa Lima JR, Luitgards-Moura JF, Menkes CE, Mondet B, Oliveira C, Weimann ETS, Tsouris P. Early determination of the reproductive number for vector-borne diseases: the case of dengue in Brazil. Trop Med Internat Health. 2006;11(4):343-351.
Chowell G, Diaz-Duenas P, Miller JC, Alcazar-[42] Velasco A, Hyman JM, Fenimore PW, Castillo-Chavez C. Estimation of the reproduction number of dengue fever from spatial epidemic data. Math Biosci. 2007;208:571-589.

34 Dengue Bulletin – Volume 33, 2009
Dengue fever in a tertiary hospital in Makkah, Saudi Arabia
W. Shahina,b#, A. Nassara,c, M. Kalkattawia and H. Bokharia
aInternal Medicine Department, Al Noor Specialist Hospital, King Abdullah Medical City, Makkah, Saudi Arabia
bTropical Medicine Department, Benha University, Benha, Egypt
cDermatology Department, Tanta University, Tanta, Egypt
Abstract
Dengue fever is endemic in the western part of Saudi Arabia. This study aimed at describing the clinical and laboratory profiles of dengue fever patients admitted to a tertiary hospital in Makkah, Saudi Arabia, from 2006 to 2008.
A total of 159 dengue fever patients were admitted during the spring and early summer. Their mean age was 25.6±16.1 years. Males outnumbered females by a ratio of 2:1. Of them, 143 patients (89.9%) had classic dengue fever and 16 patients (10.1%) had dengue haemorrhagic fever (DHF); one of them developed severe dengue shock syndrome (DSS) and died (0.6%). The common symptoms were high-grade fever, headache and body aches (100%), nausea and vomiting (27%), retro-orbital pain (25%), skin rash (16.4%), dry cough (8.2%) and haemorrhagic manifestations (3.14%). The main laboratory abnormalities were leukopenia (WBCs less than 4000/cmm) in 53.7% of patients, thrombocytopenia (platelet count less than 100 000/cmm) in 36.2% of patients and prolonged partial thromboplastin time (PTT) (>1.5 times of control value) in 33% of patients. Aspartate aminotransferase (AST) and alanine aminotransferase (ALT) were elevated more than five-fold in 35.2% of patients. The mean AST/ALT were 226.7/164 U in DF while in DHF these were 555/314 U. Twenty-six patients (16.4%) developed skin rash and had a significantly lower ALT value and a higher platelet count than those without rash.
Keywords: Dengue virus; dengue haemorrhagic fever; Makkah; Saudi Arabia; clinical symptoms; AST/ALT ratio.
#E-mail: [email protected]; Tel.: 00966507971162; Fax: 0096625668276
Introduction
Dengue fever (DF) is one of the world’s major re-emerging infections. In recent decades, there has been an expanded geographical distribution of the virus and the mosquito vector, increased
epidemic activity, and the emergence of dengue haemorrhagic fever (DHF) in new geographical regions.[1] The disease is endemic in more than 100 countries and around 2500 million people are at risk. WHO estimates that there may be 50 million cases of dengue infection worldwide

Dengue Bulletin – Volume 33, 2009 35
Dengue fever in a tertiary hospital in Makkah, Saudi Arabia
every year.[2,3] The reasons for the emergence of dengue haemorrhagic fever are complex and not fully understood but demographic, social and public health infrastructure changes in the past decades have contributed greatly to this phenomenon[4].
Dengue virus belongs to the genus Flavivirus, family Flaviviridae. It is composed of single-stranded RNA and has four antigenically-related but distinct serotypes (DENV-1, DENV-2, DENV-3 and DENV-4). It is transmitted by the bite of Aedes aegypti mosquito.[5] According to WHO, dengue virus can cause classic dengue fever, dengue haemorrhagic fever and dengue shock syndrome (DSS).[6] Outbreaks have been more common in West Asia in the 1990s, with a major epidemic occurring in Jeddah, Saudi Arabia, in 1994.[7] A few reports on the epidemic were published after 2001.[8,9,10] Many factors contribute to the recognition of such outbreaks, which include increased awareness on the part of medical authorities and more exposure of populations to the mosquito vector, especially in low-standard areas and in the peripheral region of towns where solid waste disposal is suboptimal.
This study aimed at reporting the demographic, clinical and laboratory data along with the disease outcome of all dengue patients admitted to Al Noor Hospital, Makkah, during the period 2006–2008, and comparing the characteristics between those with simple and complicated disease, which may aid in improved recognition of the disease in the area.
Materials and methods
Study site and population
This study was conducted on 159 patients admitted with diagnosis of dengue infection
to Al Noor Specialist Hospital, Makkah, from 2006 to 2008. Makkah is a holy city for Muslims from all over the world. It is located in the western province of Saudi Arabia, about 70 km from the Red Sea (Jeddah city). It is the third largest city in Saudi Arabia after Riyadh and Jeddah. It has a population of about 3 million and receives about four million visitors a year. Al Noor Specialist Hospital is a 600-bed, well-equipped, tertiary-level hospital. It is the main hospital in Makkah. It actually serves the entire local community as well as visitors as it is only 3 km away from the holy mosque. All age groups, including paediatric patients, are admitted there.
Study design
All patients presented to the emergency room with high fever, bone pains and bicytopenia were admitted as cases of “fever and bicytopenia for investigation” and were fully investigated. The diagnosis was revised on discharge and the final diagnosis was implemented on the Hospital Information System (HIS) according to the disease coding system ICD-10 AM (International Classification of Disease-10, Australian Modification), version 2006. Data were collected retrospectively by reviewing the HIS and the patient discharge summary.
The data studied included age, sex, nationality and the presence of fever, constitutional symptoms, skin rashes and bleeding tendency. Investigations included complete blood count, liver function tests and coagulation profile (PT, PTT and INR). Tests for fever of unknown origin were conducted and these included: bacterial cultures, serology for Salmonella and Brucella and serology for viruses (hepatitis viruses A, B and C, cytomegalovirus and infectious mononucleosis virus). Thin and thick blood films were examined for malaria parasites.

36 Dengue Bulletin – Volume 33, 2009
Dengue fever in a tertiary hospital in Makkah, Saudi Arabia
Sera from all suspected cases were tested in the Central Laboratory of the Ministry of Health in Jeddah for anti-dengue immunoglobulins (IgM) by enzyme-linked immunoassay (ELISA) and for the dengue virus RNA by polymerase chain reaction (RT-PCR). The results were either positive or negative for dengue fever. It must, however, be mentioned that serotyping was not done in this study.
The diagnosis of dengue fever depended on the clinical and laboratory findings and positive serology according to WHO criteria.[6]
Statistical analysis
All data were entered and analysed using Microsoft Office Excel 2007.
Results
All patients tested positive for anti-dengue immunoglobulins (IgM-ELISA) and/or dengue virus RNA by polymerase chain reaction (RT-PCR). They had negative cultures and negative serology for Salmonella, Brucella, viral hepatitis, cytomegalovirus and infectious mononucleosis virus infections. Also, thin and thick blood films for malaria were negative.
Seasonality
Thirty nine patients were admitted during 2006, 97 during 2007 and 23 during 2008. Most of the patients were admitted during the spring and early summer (April, May and June) (123/159, 77.4%) (see Figure).
Figure: Monthly distribution of dengue fever patients admitted during 2006–2008

Dengue Bulletin – Volume 33, 2009 37
Dengue fever in a tertiary hospital in Makkah, Saudi Arabia
Two thirds of the patients were Saudi (67%), only two patients were visitors (pilgrims for Hajj), while all others were residents of Makkah. The mean age was 25.6±16.1 years (range 4 to 81 years). All age groups, including children, were admitted; the percentage of children below the age of 12 years was only 24%. The age distribution is shown in Table 1. The male to female ratio was 2:1 (107 M:52 F).
Table 1: Age distribution of dengue fever patients
Age (Years)
Number of patients (%) (Total =159)
Up to 12 38 (24.0%)
13–20 47 (30.0%)
21–30 32 (20.0%)
31–40 17 (10.5%)
41–50 17 (10.5%)
>50 8 (5.0%)
According to WHO criteria,[6] 143 patients (89.9%) were diagnosed as classic DF and 16 patients as DHF (10.1%), five of them had clinically significant bleeding (3.1%) and one patient died because of severe DSS (0.6%). Plasma leakage, which is the hallmark of DHF, could not be fully assessed since it is a retrospective study and no routine serial follow-up of complete blood count (CBC), chest X-ray (CXR) or ultrasound to document plasma leakage was undertaken unless it was clinically significant.
Clinical manifestations
All patients had headache, bodyaches and high-grade fever. Fever was more than
38.5 °C for an average of 4.83±2.48 days before admission (range: 1 to 14 days). Two male patients presented with fever and coma on top of chronic liver disease and were diagnosed as hepatic encephalopathy and dengue fever; they improved on supportive treatment. One patient presented with shock and gastrointestinal bleeding; the endoscopy showed haemorrhagic gastritis and the mucosa was oozing blood. This patient died after two days in the intensive care unit because of irreversible shock (Table 2).
It is not possible to revise the clinical data according to the new WHO guidelines (2009)[11] for severe and non-severe dengue as these were published after the end of the study.
Laboratory investigations
The haematological abnormalities were thrombocytopenia and leukopenia. Platelet count less than 100 000/cmm was seen in 36.2% of patients and less than 50 000/cmm in 6.9% of patients. The white blood cell count was less than 4000/cmm in 53.7% of patients and less than 2000/cmm in 9.3% of patients. Partial thromboplastin time was 1.5-fold higher than the upper normal level in 33.3% of patients, while prothrombin time and INR were normal in all patients. The AST and ALT values were five-fold more than the upper normal levels in 56 patients (35.2%). The AST value was ten-fold more than the upper normal level in 23 patients (14.5%), while ALT value was ten-fold more than the upper normal level in 14 patients (8.8%), and AST/ALT ratio was 1.38:1.
Five patients presented with DHF and with clinically significant levels of bleeding; they were four males and one female and all were non-Saudi, the average age being 20.8±9.8 (8 to 37). Haemoglobin was lower than 12 gm/dl

38 Dengue Bulletin – Volume 33, 2009
Dengue fever in a tertiary hospital in Makkah, Saudi Arabia
and WBC count was above 4000/cmm in all patients. The platelet count was below 100 000/cmm in three patients (60%) and PTT was above normal in all patients. In all patients, ALT was above five-fold, and AST was above ten-fold. Table 3 shows that the patients presented with bleeding had less platelet count and more prolonged PTT, and significantly higher WBCs, AST and ALT values.
Table 2: Clinical manifestations of patients with dengue fever
Clinical manifestations n (%)
Fever 159 (100%)
Headache/bodyache 159 (100%)
Retro-orbital pain 40 (25%)
Chills and rigours 34 (21.4%)
Gastrointestinal symptoms
Upper Nausea and vomiting
Lower Abdominal pain and diarrhoea
43 (27.0%)
39 (24.5%)
Skin rash: Total
Morbilliform
White islands in a sea of red
Sunburn-like erythema of the face
26 (16.4%)
15/26 (57.7%)
8/26 (30.8%)
3/26 (11.5%)
Respiratory symptoms (dry cough and sore throat) 13 (8.2%)
Haemorrhagic manifestations (five patients)
Haematemesis and melena
Epistaxis and hemoptysis
Haematuria
Puerperal haemorrhage
5(3.14%)
2 (40%)
1 (20%)
1 (20%)
1 (20%)
Others (hepatic encephalopathy) 2 (1.3%)
Mortality (one patient) 1 (0.6%)
n=number of patients (159) %=percentage of patients with studied clinical manifestation to the total number of patients and the group.
Twenty-six patients had skin rashes and dengue fever; the skin rash ranged from morbilliform (15 patients), white islands in a sea of red (eight patients) to sunburn-like erythema of the face (three patients) (Table 2). It was found that the patients who presented with rash had a statistically insignificant lower WBCs count and AST values while the platelet count was significantly higher and ALT value was significantly lower (Table 4).

Dengue Bulletin – Volume 33, 2009 39
Dengue fever in a tertiary hospital in Makkah, Saudi Arabia
Table 3: Comparison of laboratory findings between DF and DHF patients presented with bleeding
Parameter DF patientsMean±SD (range)
DHF patients with bleeding
(5 patients)Mean±SD (range)
P value
Platelets (150–400x103/cumm)
124±72.8x103
(6–340)101±54 (45–161)
>0.05
WBCs (4–11x 103/cumm)
3.94±2.05x103
(0.9–9.9)8±2.6
(5.2–11.3)<0.05
PTT (28.6–38.2 sec) 45.69±9.6 (32–71 )
47.6±8.2 (39.3–55.6)
>0.05
AST (15–37 IU/L) 226.7±190 (45–980)
555±183 (425–685)
<0.05
ALT (17–41 IU/L) 164±119.5 (31–458)
314±26 (295–333)
<0.05
SD=standard deviation ALT=alanine aminotransferase AST=aspartate aminotransferase PTT=partial thromboplastin time WBCs=white blood cells
Table 4: Comparison between laboratory results in dengue fever patients with and without skin rash
Without skin rash (133) Mean±SD (range)
With skin rash (26) Mean±SD (range) Parameter P value
4.15±2.13 (0.9–9.9)
3.9±2.29 (1.9 to 9.9)
WBCs (4–11x103/cumm)
>0.05 (0.717)
112.34±65.5 (6–340)
171±86.3 (6–330)
Platelets (150–400x103/cumm)
<0.05 (0.010)
257.96±207 (47–980)
119±25.49 (45–165)
AST (15–37 IU/L)
>0.05 (0.072)
180±126.5 (31–458)
84.2±45.1 (22–182)
ALT (17–41 IU/L)
<0.05 (0.026)
SD=standard deviation ALT=alanine aminotransferase AST=aspartate aminotransferase PTT=partial thromboplastin time WBCs=white blood cells

40 Dengue Bulletin – Volume 33, 2009
Dengue fever in a tertiary hospital in Makkah, Saudi Arabia
Course of the illness
All patients were treated symptomatically with intravenous fluids and analgesics. Fresh frozen plasma, platelet and blood transfusion were given only when needed, according to WHO criteria. The length of stay in the hospital varied from one to nine days with an average of 4.07±1.48 days. All patients improved, except one male, non-Saudi patient, who developed severe DSS and died. Patients were discharged from hospital, according to WHO criteria for discharging patients which include: absence of fever for at least 24 hours without the use of anti-fever therapy, return of appetite, visible clinical improvement, good urine output, minimum of three days after recovery from shock, no respiratory distress from pleural effusion, and no ascites and platelet count of more than 50 000/cmm.[6]
Discussion
This study signifies the special pattern of dengue fever in the western part of Saudi Arabia, which is different from other parts of Asia; it happens more in adults and males, has different seasonality, has less morbidity and mortality, and does not constitute a major health problem for visitors to holy places.
Epidemics of dengue fever have been reported from the Arabian Peninsula since the late 19th century, affecting many major cities such as Aden, Jeddah, Makkah and Madinah.[7] Dengue fever virus was first isolated in Jeddah in 1994 from a fatal case of DHF; since that time, all medical personnel in the area were alerted and a surveillance system was established by the Ministry of Health. From 1994 to 1999, a total of 207 cases of dengue fever were reported;[8] since then, sporadic cases have continued to be reported from Jeddah and Makkah.
In the present study, most of the cases were admitted in the spring and early summer (77%); cases were also reported from Jeddah.[10] While in tropical areas most of the epidemics happen in the post-rainy season in the autumn months,[12] it is important to mention that it seldom rains in Makkah and Jeddah but there are swamps and inadequate sanitary facilities in some areas. While dengue fever is an important health problem for travellers to all endemic areas, including Makkah, only two of the patients were visitors (for Hajj) and the others were non-Saudi patients resident in Makkah. The reason for a lesser number of Hajj patients could be because of good sanitation and mosquito control in Hajj residency areas. (The risk of exposure to dengue fever can be minimized in modern, air-conditioned hotels with well-kept grounds.[13])
The mean age of patients was 25.6±16.1 years and the percentage of infected children was 24%; in Jeddah the mean age was 27.6±11.2 years and the percentage of children was 6%.[10] Internationally, dengue fever is an infectious disease of children. Reports from India, Sri Lanka, Indonesia and Thailand indicate that children may constitute up to 95% of cases and the peak age of contracting dengue infection is between 5 and 10 years.[14-18]
It is clear that the age pattern of dengue fever infection in Saudi Arabia is different from that of South-East Asia, and is closer to the age pattern of dengue fever infection in Brazil, where it is the highest in adults. This may be related to the genotypes present. The reports from Brazil indicate that DENV-1 and DENV-2 viruses happen more in adults,[19,20] and in Jeddah, DENV-1 and DENV-2 viruses constitute 93% of the cases[9]. Males are affected more than females (ratio of 2 to 1); the same is the case in Jeddah[10] while reports from other endemic areas show that males and

Dengue Bulletin – Volume 33, 2009 41
Dengue fever in a tertiary hospital in Makkah, Saudi Arabia
females are affected equally.[16,17,21] The less number of females affected in Saudi Arabia may be due to their limited outdoor exposure (for social reasons) and to their wearing the veil (hijab) outdoors when the whole body is covered.
As regards the patients’ symptoms, high-grade fever, headache and bone pains were the most common symptoms, followed by gastrointestinal complications, chills and rigours, skin rash and respiratory problems. Similar findings have been reported by national[10] and international researchers.[22] Haemorrhagic manifestations happened in five patients (3.14%), similar to other reports in Saudi Arabia and internationally.[10,23] In a study from Indonesia, out of 1300 cases severe bleeding was recorded in 76 of the dengue cases (6%) and included haematemesis (58%), melena (21%), haematemesis-melena (16%) and DIC (5%).[20] Two patients presented with hepatic encephalopathy (1.3%), and both had chronic liver disease. A report from Thailand shows that hepatic encephalopathy was considered an unusual manifestation of dengue fever and they attributed this to toxic substance, drugs or underlying liver conditions.[24] In rare cases dengue fever may present as an acute liver failure.[25] One patient presented with DSS and GI bleeding and died (0.6%). In the Indonesian study, out of 1300 patients, 102 patients (8%) had DSS and 17 patients died (1.3%).[20] About 36.2% of patients had a platelet count of less than 100 000/cmm, which is less than the international figure (58%)[26]. The lower percentage of DSS and thrombocytopenia in this study compared to the international figures perhaps because of the limited number of cases; also, it is necessary to do more frequent investigations to detect early and mild cases of plasma leakage (such as CBC, abdominal ultrasound and CXR).
Patients who developed bleeding were not necessarily those who had the lowest platelet number, which means that bleeding is a multifactorial process; bleeding may result from a combination of factors such as thrombocytopenia, coagulation defects and vasculopathy.[20] The average number of white cell count was 3940±2050 and 53.7% of the patients had WBCs less than 4000/cmm; this was close to a local study[10] where 48.72% of patients had WBC below 4000/cmm. Also, in a study from South-East Asia, the mean WBCs was 3834±2216 and the percentage of leukopenia below 5000/cmm was 89.77%[26]. Prolonged PTT more than 1.5-fold the upper normal limit was found in one third of the patients; this was similar to reports from Jeddah, Thailand and India.[10,15,22]
The liver is a target organ of dengue infection; hepatic involvement ranged from mildly elevated aminotransferases to fulminant hepatic failure leading to death.[27-30] The pathology included severe, diffuse hepatitis, focal necrosis of hepatic cells and hyaline necrosis of Kupffer cells.[31,32,33] Liver function tests showed elevated liver enzymes with normal bilirubin and alkaline phosphatase, liver enzymes were higher than five-fold the upper normal limit in 56 patients (35.2%); it was also elevated as reported from Saudi Arabia and Thailand in 66.7%, 63%, 67% and 34.2% of patients.[10,27,34,35] It is worth noting that the level of AST elevation was higher than that of ALT (ALT/AST ratio of 1:1.38) in contrast to acute viral hepatitis; this finding was also reported from South-East Asia[24].
Sixteen patients had DHF (10.1%), five of them presented with overt bleeding; the average age was 20.8±9.8 (8 to 37), similar to reports from Brazil[20] but different from South-East Asian patients where most of them were children.[18] Liver enzymes were higher than those with classic dengue fever and the

42 Dengue Bulletin – Volume 33, 2009
Dengue fever in a tertiary hospital in Makkah, Saudi Arabia
AST/ALT ratio was higher at 1.77:1 while it was 1.38:1 in dengue fever patients. In South-East Asia, 92% of patients with DHF had AST more than 60U, and in patients who presented with encephalopathy, AST and ALT values usually exceeded 200U.[24] The frequency of hepatic dysfunction depends on dengue fever disease severity. There was a correlation between liver enzymes and the severity of dengue infection.[36] The appearance of rash may be associated with the milder illness of classic DF.
The prevalence of dengue fever in Saudi Arabia is not unique. WHO has described the presence of dengue in other areas of the Eastern Mediterranean Region – in Egypt as early as in 1979, in Sudan (1985) and in Djibouti (1991). In Saudi Arabia, three major epidemics have been reported: a DENV-2 epidemic in 1994 with 469 cases of dengue,
23 cases of DHF, two cases of DSS and two deaths; a DENV-1 epidemic in 2006 with 1269 cases of dengue, 27 cases of DHF, 12 cases of DSS and six deaths; and a DENV-3 epidemic in 2008 with 775 cases of dengue, nine cases of DHF, four cases of DSS and four deaths[36].
The limitations of this study are due mainly to its retrospective type, limited number of cases, inability to know the total number of suspected cases, and lack of frequent investigations to pick up early cases of plasma leakage. These issues should be taken into consideration in later studies.
In conclusion, dengue fever is an endemic disease in Makkah, the holy city for Muslims. It is rare in visitors who come to Makkah for the Hajj pilgrimage, but all measures to prevent the spread of the disease should be taken.
ReferencesGubler DJ. The global emergence/resurgence [1] of arboviral diseases as public health problems. Arch Med Res. 2002;33:330-342.
World Health Organization. [2] Dengue and dengue haemorrhagic fever. Fact sheet N°117, Revised April 2009. Geneva: WHO, 2002.
Gibbons RV, Vaughn DW. Dengue: an escalating [3] problem. BMJ. 2002;324:1563-1566.
Gubler DJ. The global pandemic of dengue/[4] dengue haemorrhagic fever: current status and prospects for the future. Ann Acad Med Singapore. 1998;27(2):227-34.
Calisher CH, Karasbarsos N, Dalrymple [5] JD, Shope RE, Porterfield JS, Weataway EG, Brandt WE. Antigenic relationships between flaviviruses as determined by cross-neutralization tests with polyclonal antisera. J Gen Virol. 1989;70:37-43.
World Health Organization.[6] Guidelines for treatment of dengue fever/dengue haemorrhagic fever in small hospitals. New Delhi: World Health Organization, Regional Office for South-East Asia, 1999. p. 2-3.
Gubler DJ. [7] Dengue and dengue haemorrhagic fever: its history and resurgence as a global public health problem. In: Gubler DJ. Kuno G. Editors. Dengue and dengue haemorrhagic fever. London: CAB International, 1997. p. 1-22.
Fakeeh M, Zaki AM. Virologic and serologic [8] surveillance for dengue fever in Jeddah, Saudi Arabia, 1994-1999. Am J Trop Med Hyg. 2001;65(6):764-67.
Fakeeh M, Zaki AM. Dengue in Jeddah, [9] Saudi Arabia, 1994-2002. Dengue Bulletin. 2003;27:13-18.

Dengue Bulletin – Volume 33, 2009 43
Dengue fever in a tertiary hospital in Makkah, Saudi Arabia
Ayyub M, Khazindar AM, Lubbad EH, Barlas [10] S, Alfi AY, Al-Ukayli S. Characteristics of dengue fever in a large public hospital, Jeddah, Saudi Arabia. J Ayub Med Coll Abbottabad. 2006;18(2):9-13.
World Health Organization. [11] Dengue guidelines for diagnosis, treatment, prevention and control. Geneva: WHO, 2009.
Pancharoen C, Thisyakorn U. Dengue virus [12] infection during infancy. Trans R Soc Trop Med Hyg. 2001;95(3):307-8.
G u b l e r D J . D e n g u e a n d d e n g u e [13] haemorrhagic fever. Clin Microbiol Rev. 1998;11(3):480-496.
Aggarwal A, Chandra J, Aneja S, Patwari [14] AK, Dutta AK. An epidemic of dengue haemorrhagic fever and dengue shock syndrome in children in Delhi. Indian Pediatr. 1998;35(8):727-32.
Witayathawornwong P. Dengue haemorrhagic [15] fever in infancy at Petchabun Hospital, Thailand. Southeast Asian J Trop Med Public Health. 2001;32(3):481-7.
Nimmannitya S. Clinical spectrum and [16] management of dengue haemorrhagic fever. Southeast Asian J Trop Med Pub Hlth. 1987;18(3):392-97.
Nimmannitya S. [17] Dengue haemorrhagic fever: diagnosis and treatment. In: Gubler DJ and Kuno G. Eds. Dengue and dengue haemmorrhagic fever. Wallingford: CAB International, 1997. p. 133-45.
Halstead SB. Dengue in the Americas and [18] Southeast Asia: do they differ? Rev Panam Salud Publica. 2006;20:407-15.
Teixeira Mda G, Costa Mda C, Barreto ML, [19] Mota E. Dengue and dengue haemorrhagic fever epidemics in Brazil: what research is needed based on trends, surveillance, and control experiences? Cad Saude Publica. 2005;21:1307-15.
Teixeira MG, Costa M, Coelho G, Barreto [20] M. Recent shift in age pattern of dengue haemorrhagic fever, Brazil. Emerging Infectious Diseases. 2008;14(10):1663.
Chairulfatah A, Setiabudi D, Agoes R, [21] Colebunders R. Thrombocytopenia and platelet transfusions in dengue haemorrhagic fever and dengue shock syndrome. Dengue Bulletin. 2003;27:138-143.
Kabra SK, Jain Y, Singhal T, Ratageri VH. [22] Dengue haemorrhagic fever: cl inical manifestations and management. Indian J Pediatr. 1999;66(1):93-101.
Sung V, O’Brien DP, Matchett E, Brown [23] GV, Torresi J. Dengue Fever in travelers returning from southeast Asia. J Travel Med. 2003;10(4):208-13.
Nimmannitya S, Thisyakorn U, Hemsrichart [24] V. Dengue haemorrhagic fever with unusual manifestations. Southeast Asian J Trop Med Public Health. 1987;18(3):398-406.
Vinodh BN, Bammigatti C, Kumar A, Mittal [25] V. Dengue fever with acute liver failure. Case reports. 2005,51(4):322-323.
Gomber S, Ramachandran VG, Kumar [26] S, Agarwal KN, Gupta P, Gupta P, Dewan DK. Hemato log ica l observa t ions as diagnostic markers in dengue haemorrhagic fever – a reappraisal. Indian Pediatr . 2001;38(5):477-81.
Kalayanarooj S, Nimmannitya S, Suntayakorn [27] S, Vaughn DW, Nisalak A, Green S, Chansiriwongs V, Rothman A, Ennis FA. Can doctors make an accurate diagnosis of dengue infections at an early stage? Dengue Bulletin. 1999;23:39-43.
Lum LC, Lam SK, George R, Devi S. [28] Fulminant hepatitis in dengue infection. Southeast Asian J Trop Med Public Health. 1993;24(3):467-471.

44 Dengue Bulletin – Volume 33, 2009
Dengue fever in a tertiary hospital in Makkah, Saudi Arabia
de Souza LJ, Goncalves Carneiro H, Souto [29] Filho JT, Ferreira de Souza T, Azevedo Cortes V, Neto CG, Bastos DA, da Silva Siqueira EW. Hepatitis in dengue shock syndrome. Braz J Infect Dis. 2002;6(6):322-327.
Lawn SD, Tilley R, Lloyd G, Finlayson [30] C, Tolley H, Newman P, Rice P, Harrison TS. Dengue haemorrhagic fever with fulminant hepatic failure in an immigrant returning to Bangladesh. Clin Infect Dis. 2003;37(1):e1-4.
Lin YL, Liu CC, Lei HY, Yeh TM, Lin YS, [31] Chen RM, Liu HS. Infection of five human liver cell lines by dengue-2 virus. J Med Virol. 2000;60(4):425-431.
World Health Organization. [32] Dengue haemorrhagic fever: diagnosis, treatment, prevention and control. 2nd edn. Geneva: WHO, 1997.
Huerre MR, Lan NT, Marianneau P, Hue [33] NB, Khun H, Hung NT, Khen NT, Drouet MT, Huong VT, Ha DQ, Buisson Y, Deubel V. Liver histopathology and biological correlates in five cases of fatal dengue fever in Vietnamese children. Virchows Arch. 2001;438(2):107-115.
Pancharoen C, Rungsarannont A, [34] Thisyakorn U. Hepatic dysfunction in dengue patients with various severity. J Med Assoc Thai. 2002;85 Suppl 1:S298-301.
Petdachai W. Hepatic dysfunction in [35] children with dengue shock syndrome. Dengue Bulletin. 2005;29,112-118.
Mohan B, Patwari AK, Anand VK. Hepatic [36] dysfunction in childhood dengue infection. J Trop Pediatr. 2000;46(1):40-43.

Dengue Bulletin – Volume 33, 2009 45
Genotypic and phenotypic characteristics of DENV-3 isolated from patients with
different disease severities in IndonesiaBeti Ernawati Dewia,b#, Tomohiko Takasakia, Shigeru Tajimaa,
T Mirawati Sudirob, R.P. Larasatic, Andrew Lee Corwinc and Ichiro Kuranea
aLaboratory of Vector-Borne Viruses, Department of Virology 1, National Institute of Infectious Diseases, 1-23-1 Toyama, Shinjuku-ku, Tokyo, 163-8640, Japan
bDepartment of Microbiology, Medical Faculty, University of Indonesia, Jalan Pegangsaan Timur no. 16, Jakarta 10320, Indonesia
cU.S. Naval Medical Research Unit No. 2, Jakarta, Indonesia
Abstract
We have investigated the genetic diversity of DENV-3 isolated during an outbreak of dengue haemorrhagic fever (DHF) in the city of Palembang, south Sumatra, Indonesia. Four viruses were isolated from patients with different disease severities. Amino acid differences were detected at 40 positions throughout the structural and non-structural proteins, except for the preM protein. Of the 40 positions, amino acids were conserved at 24 positions among three DSS/DHF strains, and the DF strain possessed different amino acids at these positions. Five nucleotide substitutions were identified on 3’ UTR. In monocyte cell line, U937, the three DSS/DHF strains were able to propagate to higher levels compared to the DF strain. These preliminary results showed that the DSS/DHF strains have unique amino acid residues that are different from the DF strain and have better ability to propagate to a higher level in human monocytes. These characteristics of the DSS/DHF strains may contribute to enhanced infection and induction of severity of the disease.
Keywords: DENV-3; genotypic characteristics; phenotypic characteristics; Indonesia.
#E-mail: [email protected]
Introduction
Dengue virus infection continues to increase in tropical and sub-tropical countries of the world, and presently, more than 2.5 billion people now live in regions with the
risk of dengue infection.[1] The pathological mechanisms of dengue haemorrhagic fever (DHF) are still poorly understood. The lack of animal models for severe dengue illness has hampered the elucidation of mechanisms by which dengue virus infection leads to DHF.

46 Dengue Bulletin – Volume 33, 2009
Genotypic and phenotypic characteristics of DENV-3 in Indonesia
A number of models have been proposed, based on epidemiological and experimental data, to explain the pathogenesis of severe dengue illness.
The hypothesis that various kinds of cytokines are responsible for plasma leakage, the hallmark of DHF, has been supported by many workers.[2,3,4] The other mechanism, antibody-dependent enhancement (ADE), arose from the observation that secondary infection with heterotypic dengue virus is associated with an increased risk of DHF and from the experiments demonstrating that sub-neutralizing the concentration of antibodies increased virus titres in vivo and in vitro.[5,6] However, secondary infection by some dengue viruses does not produce DHF/DSS.[7,8] DHF may also be due to the intrinsic biological properties of dengue virus strains.[9]
Advances in technology of gene cloning, nucleotide sequencing and gene expression have facilitated the understanding of the molecular biology of dengue viruses. Dengue virus genome is composed of approximately 10 600 nucleotides, single-stranded positive sense RNA.[10] It contains a single open reading frame that is flanked by two un-translated regions; 5’ and 3’ un-translated region (UTR). The amino acid differences of dengue viruses have been implicated to the pathogenesis of DHF.[11,12] Dengue virus virion is composed of three structural proteins: core protein (C); membrane protein (M); and envelope protein (E). Seven non-structural proteins are also present: NS1, NS2a, NS2b, NS3, NS4a, NS4b and NS5.[10,13]
Although dengue epidemic occurred at regular intervals in Indonesia since it was first recognized in Java, there have been limited reports of the diversity of dengue viruses isolated in Indonesia. Analysis of the viruses
isolated from DHF patients from 1975 to 1978 demonstrated that all four dengue viruses were endemic in Jakarta, but DENV-3 was the most predominant.[14] DENV-3 was also the most frequently isolated virus outside Jakarta and had the widest distribution in the country.[14,15]
In the present study, we defined the amino acid substitutions that may contribute to the disease severity using DENV-3 isolated from patients with different disease severities. We also analysed the phenotypic characteristics; growth kinetic in Vero, C6/36 and U937 Cell. In addition, we also analysed the genetic relationship of these four isolates among other DENV-3 isolates.
Materials and methods
Source of the viruses
The dengue viruses used in this study were isolated from patients during an outbreak of DHF in the city of Palembang, South Sumatra, from January to April 1998.[15] The viruses were isolated from patients with different disease severities according to the criteria fixed by the World Health Organization (Table 1). Serum samples were serologically examined by anti-dengue IgM/IgG antibody-capture ELISA[16] and rapid immunochromatographic assay (PanBio Inc., Brisbane, Australia). Serological tests were done at the US Naval Medical Research Unit-2 (US NAMRU-2), Jakarta.
Virus isolation
Serum samples were collected from the hospitalized, suspected dengue cases and analyzed for the presence of dengue virus at

Dengue Bulletin – Volume 33, 2009 47
Genotypic and phenotypic characteristics of DENV-3 in Indonesia
US NAMRU-2.[15] Virus isolation was done by inoculating 50 ul of 1:10 dilution serum samples onto C6/36 cell line in 24 well plates.[17] Dengue viruses were propagated in C6/36 cells at 28 °C for seven days. The culture supernatants were harvested and centrifuged at 900 g for five minutes and then filtered through the syringe driven millex GV with 0.22 um filter unit (Millipore, Co., Bedford, MA, USA). Culture supernatants were collected and checked for the presence of dengue virus by plaque titration onto confluent Vero cells.[17] The types of the isolated dengue viruses were determined by real-time RT-PCR.[18] Four DENV-3 isolates were extensively used in the present study (Table 1). These viruses included two isolates from DSS patients, one from DHF patient and one from a DF patient.
Plaque titration assay in Vero cells
Briefly, a 10-fold serial dilution of DENV-2 was inoculated onto Vero cell monolayer in duplicate wells. Absorption was carried out at 37 °C in 5% CO2 for two hours with agitation at intervals of 30 minutes. Methylcellulose overlay medium was added and infected Vero cells were incubated at 37 °C in 5% CO2 for seven to nine days. Plaque numbers were counted after methylene blue staining.
Determination of mutations
RNA was extracted from 200 ul of the culture supernatants of infected C6/36 cells using High Pure Viral RNA kit (Roche Applied Science, GmbH, Germany) according to the manufacturer’s instructions. Complementary DNA (cDNA) strands were reverse-transcribed using Super Script II First Strand Synthesis System with random hexanucleotide primer according to the manufacturer’s instructions (invitrogen). The PCR amplification of the entire genome was performed using the cDNA products in PTC-100TM programmable thermal cycles (MJ Research Inc.). PCR amplification reaction included: 2 µl of cDNA, 3 µl of 10X PCR buffer containing 1.5 mM MgCl, 1.5 µl of 0.2 mM of each dNTPs, 0.3 µl of 2.5 U Taq polymerase, 0.5 µl of each primer 100 pmol in a total reaction mixture of 30 µl. PCR was performed with initial denaturation at 94 °C for one minute followed by 30 cycles of denaturation at 94 °C for 30 seconds, annealing at 60 °C for 30 seconds and extension at 72 °C for 60 seconds, and a final single cycle extension at 72 °C for two minutes. After amplification, PCR products were analysed by using electrophoresis. Five micro litre of PCR product was loaded on 2% agarose gel in TAE buffer and stained ethidium bromide, and visualized under ultraviolet (UV) light.
Table 1: The strain code, disease severities, patients’ demography, serological responses and GeneBank accession number of DENV-3 isolates
Code of isolates Serotype Clinical
gradeSerological response Sex Age
(years)
Date of onset of
fever
Date of sampling
GeneBank accession
no.
98902890 DENV-3 DF Secondary F 8 7 Apr 98 9 Apr 98 AB189128
98901517 DENV-3 DHF Secondary M 2 7 Apr 98 9 Apr 98 AB189127
98901403 DENV-3 DSS Secondary M 13 8 Apr 98 9 Apr 98 AB189125
98901437 DENV-3 DSS Secondary M 11 5 Apr 98 9 Apr 98 AB189126

48 Dengue Bulletin – Volume 33, 2009
Genotypic and phenotypic characteristics of DENV-3 in Indonesia
Twenty-five micro litres of PCR amplification products were purified using QIAquick PCR purification kits (Qiagen GmbH, Hilden, Germany) as outlined by the manufacturer and used for direct sequencing.
Primer sequences were obtained and designed from the published data of DENV-3 (GenBank accession no. AY648961) using Oligo 6 programme (Table 2). The unknown sequences at either 5’ and 3’ UTR were determined using rapid amplification of cDNA ends (RACE) according to the manufacturer’s instructions (invitrogen).
The sequences were determined using Taq Big Dye Deoxy Terminator Cycle sequencing kits (Applied Biosystems, Foster City, California, USA). Three micro litres (±50ng) of purified products was sequenced directly by adding 3.2 pmol of primer, 2 µl Big Dye Terminator v. 3.0 and 4 µl of 5X Big Dye Terminator v. 3.0 buffer (Applied Biosystems, Foster City, CA USA) and sterile distilled water up to 20 ul. Following thermal cycling as described in the manufacturer’s instructions, reaction products were purified by column purification (CentriSeps, USA) and vacuum-dried for 30 minutes. The pellet was resuspended in 15 µl of Hi DiTM Formamide (Applied Biosystems, USA) and analysed by using an ABI PRISM
3100-avant genetic analyser (data collection software ver. 1.0 sequence analysis software ver. 3.7). Nucleotide and amino acid sequence analysis was performed using GENETIC MAC- and BLAST programme. Codon positions used in the present study were based on the GenBank accession number described earlier.
Pylogenetic analysis
The envelope protein of DENV-3 from GenBank was used to develop a phylogenetic tree for comparison among the isolates.
Construction of phylogenetic trees and homology analysis were performed using the Genetyx-Win version 6.1 programme.
Growth kinetics in Vero cell and C6/36 cell
Vero cells and C6/36 cells were infected with DENV-3 at an m.o.i of 0.001 pfu/cell and 0.01 pfu/cell respectively in 12 well plates. After two hours of incubation at 37 °C in 5% CO2, cells were washed with MEM supplemented with 2% FBS. Two ml of MEM supplemented with 2% FBS medium were added and this point of time was designed as 0 hour. At indicated time points after infection, 400 ul of culture supernatant fluids were removed, centrifuged to remove debris and frozen at –80 °C. The level of virus replication was assayed by plaque titration in Vero cells.
Growth kinetics in U937 cells
A total of 1 x 106 U937 cells were infected with 100 ul of DENV-3 at an m.o.i of 0.2 pfu/cell with the presence of 1:10 000 diluted DENV-2-infected patient’s sera. The infection was carried out for two hours at 37 °C in 5% CO2. After two hours of incubation, cells were washed with RPMI 1640 medium without FBS. Subsequently, U937 cells were diluted at the concentration of 1 x 106 cell/ml in RPMI 1640 medium (Sigma) supplemented with 10% heat inactivated FBS (Sigma, USA), L-glutamine and NaHCO3 (Gibco), and antibiotics 100 ug of streptomycin and 100 units of penicillin (Gibco).[19] At various points of time after infection, 0.2 ml of U937 cells culture supernatant was harvested and centrifuged at 1.500 rpm for five minutes at 4 °C. Supernatants were collected and assayed for virus titres using plaque assay in Vero cells. The heat-inactivated virus was used as

Dengue Bulletin – Volume 33, 2009 49
Genotypic and phenotypic characteristics of DENV-3 in Indonesia
Table 2: Primer sequence, nucleotide position in the dengue genome and RT-PCR product size [Primer sequences were obtained and designed from the published data of DENV-3
(GenBank accession no. AY648961) using Oligo 6 programme]
Nucleotide position Sequence 5’-3’ Size (bp) Information
1-22461-482
AGTTGTTAGTCTACGTGGACCGGATTTTCCTCTTTCATTCTT
481 ForwardReverse
390-411666-687
GACAGCAAGGTAGCCCATAGCCTTATCGCGTCTATGCTCTCC
297 ForwardReverse
412-433965-986
GACAGCAAGGTAGCCCATAGCCACCACGTCAACCCACGTGGC
574 ForwardReverse
843-8641041-1062
TAGCCCTATTTCTTGCCCATTCCTCGGTCTTCTGGAGCTCTA
219 ForwardReverse
985-10061438-1459
AGCCACGTGGGTTGACGTGGTTCTAGCCCAAGGGTTCCATAT
474 ForwardReverse
1381-14021906-1927
CCAGGTGGGAAATGAAACGCATCCGTGGAGAAGGGAATCTTG
546 ForwardReverse
1725-17462381-2401
GAGCTACAGAGATCCAAACCTCCTTTCCAGTTTATGACACAC
676 ForwardReverse
2342-23632973-2994
AACACTTCTATGTCATTTTCAAATAGCCCATGTCGGCGTGTA
652 ForwardReverse
2810-18313366-3387
TTTATAAATAGATGGGCCAAACCGTACCAGCAGCCGTCTTCTC
577 ForwardReverse
3164-31853812-3833
ATTTCGCAACACAACCACAGGAGCAATTCCATTCGCCATTTG
669 ForwardReverse
3653-36744280-4301
TCCAACGCCTCTGACAGAATGTCCTGTTTGCTCAGCCTCTTC
648 ForwardReverse
3857-38784437-4458
GGGCTCATGGCTCTTAAATTGAAAGTGTTGCAGGTATGGAGT
601 ForwardReverse
4308-43294996-5017
AGGAAGAGGCTGAGCAAACAGGGTCCATCTGGCTCTGCGTTT
709 ForwardReverse
4808-48295267-5288
GCCGTAGAGCCTGGGAAGAACCGTTGCGTGACACATTAGATC
480 ForwardReverse
5104-51255775-5796
GTCAGGAAAGACGCGGAAATAAAATGACTGGCTTGAGACATC
692 ForwardReverse
5616-56376268-6289
CTGGGAATGTCATAGCAAACTGCATCAAGCCACCTAGGCCTC
673 ForwardReverse
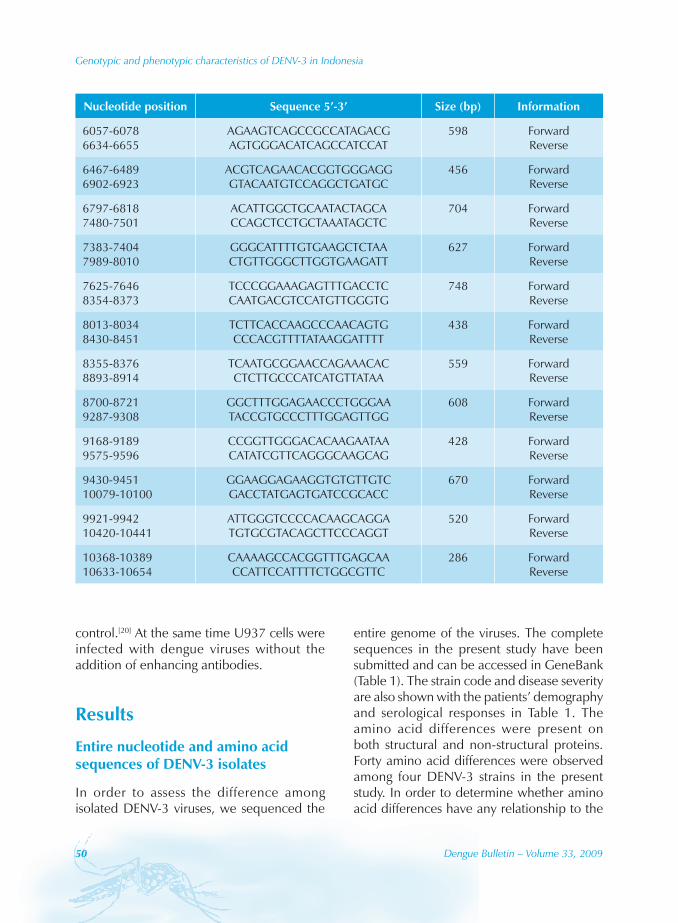
50 Dengue Bulletin – Volume 33, 2009
Genotypic and phenotypic characteristics of DENV-3 in Indonesia
Nucleotide position Sequence 5’-3’ Size (bp) Information
6057-60786634-6655
AGAAGTCAGCCGCCATAGACGAGTGGGACATCAGCCATCCAT
598 ForwardReverse
6467-64896902-6923
ACGTCAGAACACGGTGGGAGGGTACAATGTCCAGGCTGATGC
456 ForwardReverse
6797-68187480-7501
ACATTGGCTGCAATACTAGCACCAGCTCCTGCTAAATAGCTC
704 ForwardReverse
7383-74047989-8010
GGGCATTTTGTGAAGCTCTAACTGTTGGGCTTGGTGAAGATT
627 ForwardReverse
7625-76468354-8373
TCCCGGAAAGAGTTTGACCTCCAATGACGTCCATGTTGGGTG
748 ForwardReverse
8013-80348430-8451
TCTTCACCAAGCCCAACAGTGCCCACGTTTTATAAGGATTTT
438 ForwardReverse
8355-83768893-8914
TCAATGCGGAACCAGAAACACCTCTTGCCCATCATGTTATAA
559 ForwardReverse
8700-87219287-9308
GGCTTTGGAGAACCCTGGGAATACCGTGCCCTTTGGAGTTGG
608 ForwardReverse
9168-91899575-9596
CCGGTTGGGACACAAGAATAACATATCGTTCAGGGCAAGCAG
428 ForwardReverse
9430-945110079-10100
GGAAGGAGAAGGTGTGTTGTCGACCTATGAGTGATCCGCACC
670 ForwardReverse
9921-994210420-10441
ATTGGGTCCCCACAAGCAGGATGTGCGTACAGCTTCCCAGGT
520 ForwardReverse
10368-1038910633-10654
CAAAAGCCACGGTTTGAGCAACCATTCCATTTTCTGGCGTTC
286 ForwardReverse
control.[20] At the same time U937 cells were infected with dengue viruses without the addition of enhancing antibodies.
Results
Entire nucleotide and amino acid sequences of DENV-3 isolates
In order to assess the difference among isolated DENV-3 viruses, we sequenced the
entire genome of the viruses. The complete sequences in the present study have been submitted and can be accessed in GeneBank (Table 1). The strain code and disease severity are also shown with the patients’ demography and serological responses in Table 1. The amino acid differences were present on both structural and non-structural proteins. Forty amino acid differences were observed among four DENV-3 strains in the present study. In order to determine whether amino acid differences have any relationship to the

Dengue Bulletin – Volume 33, 2009 51
Genotypic and phenotypic characteristics of DENV-3 in Indonesia
disease severity, we compared three isolates from DSS (98901403 and 98901437) and DHF (98901517) patients defined as DSS/DHF strains with that from a DF (98902890) patient defined as DF strain. Twenty-four of a total of 40 amino acid differences were present between three DSS/DHF strains and one DF strain. In the M, E and NS-2b proteins, all the amino acid differences were the same among the three DSS/DHF strains.
Among the DSS/DHF strains, strain 98901403 demonstrated a high level of homology with strain 98901437 (99.35%) and strain 98901517 (99.33%). The homologies among these three DSS/DHF strains were over 99%. On the contrary, the homology between these three isolates and the DF strain 98902890 was approximately 97%.
Amino acid differences on structural and non-structural proteins
The amino acid differences were detected at eight positions in the structural proteins. Seven of eight amino acid substitutions among four DENV-3 isolates were found in the three strains analysed from DSS/DHF patients.
In the seven non-structural proteins, 32 amino acid substitutions were found. Among these four strains, eight and nine amino acids substitutions were found in the NS-1 and NS-5 regions respectively. The DSS/DHF strains had 18 amino acid substitutions that were differed from the DF strain: NS1 (2, 109, 128, 145, 157), NS2B (58), NS3 (20, 60, 350), NS4A (78, 100), NS4B (25, 175) and NS5 (364, 671, 689, 670, and 873).
Nucleotide substitutions on untranslated regions
Nucleotide substitutions at 3’UTR are listed in Table 3. No nucleotide substitutions were found in 5’UTR among these four DENV-3 isolates. Five nucleotide substitutions were identified at 3’UTR.
The predicted secondary RNA structure formed by complete 3’UTR is shown in Figure 1. The nucleotide differences at positions 10472 (A to G), 10512 (T to C) and 10596 (T to C) between strain 98901403 and strain 98902890 have no apparent effect on the predicted secondary structures (Figures 1A and 1C). However, the nucleotide differences
Table 3: Summary of amino acid replacement on full-length sequences of DENV-3 isolates from patients with different disease severities
[The amino acid position number corresponds to the position in the respective proteins]
DENV-3 strains
Nucleotide position at
10271 10305 10472 10512 10596
98901403 C C A T T
98901437 T T A C T
98901517 T T A C T
98902890 C C G C C

52 Dengue Bulletin – Volume 33, 2009
Genotypic and phenotypic characteristics of DENV-3 in Indonesia
at positions 10271 (C to T) and 10305 (C to T) between strain 98901403 and strain 98901437 or strain 98901517 induced a minor change in the secondary structures (Figures 1A and 1B).
Figure 1: Predicted RNA secondary structures formed by the complete 3’UTR [The prediction of secondary structures was analysed by RNA-Fold programme
(A) 98901403 DSS isolate; (B) 98901437 DSS and 98901517 DHF isolates;
(C) 98902890 DF isolate]
Phylogenetic analysis
To determine the relationship among these dengue viruses, the nucleotide sequences of the E genes were compared along with other published E gene sequences available in the GeneBank. A phylogenetic tree was constructed on the basis of partial sequences 933 to 2413 of the E gene. Four DENV-3 isolates were clustered with viruses from Malaysia 1974, in genotype I (Figure 2). Genotype II comprises viruses isolated in Thailand and Malaysia 1993. DENV-3 from Sri Lanka, India, and Samoa were included in genotype III. Genotype IV comprises DENV-3
strains isolated in Puerto Rico. Those isolated from China in the 1980s and the Philippines in 1956 were clustered in the genotype V. Although all the four isolates were included in the genotype I, three of DSS/DHF strains and the DF strain were separated into different clusters.
Growth kinetics of the four DENV-3 isolates in cell lines
We compared the biological activities of these viruses. The growth kinetics of the DENV-3 isolates were examined with Vero, C6/36 and U937 cells as described in the “Materials and methods” section.
The DSS/DHF strains and DF strain showed a similar growth pattern in Vero cells and the viruses were detected as early as two days after infection. The growth kinetic in C6/36 mosquito cell line also showed a similarity among DHF and DF strains (data not shown). However, the growth kinetics in U937 cells in the presence of enhancing antibody was different between DSS/DHF strains and DF strain (Figure 3). In addition, we also examined the growth kinetic in U937 cell without the presence of enhancing antibody (ADE). The growth levels of viruses in the absence of ADE were lower than the presence of ADE in U937 cells, but different between DSS/DHF strains and DF strain (Figure 4). The DSS/DHF strains were detected as early as Day 1 after infection, and comparable levels of peak titres were detected on Days 2 and 3. The DF strain demonstrated one or one-and-a-half-log lower peak titres on Days 2 and 3. This result suggests that the DSS/DHF strains grow to higher levels than the DF strain in a monocyte cell line U937.

Dengue Bulletin – Volume 33, 2009 53
Genotypic and phenotypic characteristics of DENV-3 in Indonesia
Figure 2: Maximum likelihood trees showing the phylogenetic relationships among a global sample of 27 strains of DENV-3
[The analysis was based on nucleotide sequences of the E gene. The genotype clusters are labelled according to the scheme of Wittke et al., 2002. Roman numerals denote the different genotypes of DENV-3. An asterisk (*) indicates the strains that were sequenced in this study. The name of isolates refers to country of origin, year of isolation and also disease severity if
available in GeneBank]

54 Dengue Bulletin – Volume 33, 2009
Genotypic and phenotypic characteristics of DENV-3 in Indonesia
Figure 3: Growth kinetic of DENV-3 isolates in U937 cell line
[U937 cell line was infected at an m.o.i of 0.2 pfu/cell. Supernatants were collected
every day for five days. Virus titre was determined by plaque assay in Vero cells]
Discussion
Nucleotide and amino acid changes may contribute to the difference in growth, transmission and virulence of dengue viruses. In order to determine whether the amino acid and nucleotide differences contribute to the severity of illness, we analysed two DSS strains, one DHF strain and one DF strain replication in three cell lines. In the present study the DSS/DHF strains demonstrated higher levels of growth than the DF strain in U937 cells. This may suggest that DF strain has lower mean viraemia level in vivo. It has been reported that the level of DENV-3 viraemia was greater in DHF cases than DF cases[21]. Other studies also demonstrated that the level of viraemia[22] and viral RNA copies[7] are higher in DHF cases
Figure 4: Growth kinetic of DENV-3 isolates in U937 cell line with and without presence of enhancing antibody (ADE)
(A) 98901403 DSS isolate; (B) 98901437 DSS isolate; (C) 98901517 DHF isolate; (D) 98902890 DF isolate. : with ADE; : without ADE]

Dengue Bulletin – Volume 33, 2009 55
Genotypic and phenotypic characteristics of DENV-3 in Indonesia
than in DF cases. These results suggest that dengue virus with a high growing capability might cause more severe manifestations. Cologne and Rico-Hesse explained the higher mean viraemia observed in patients with DHF: (i) more cells are infected in DHF patients, thus generating more virus; (ii) more virus is produced per infected cell; or (iii) the virus is cleared at a slower rate compared to patients with DF, resulting in an increase in blood-borne virus.[23] Our results that DSS/DHF strains are propagated to a higher level than DF strain in U937 may partly explain why this DF strain induced mild disease. Epidemiological and clinical studies have demonstrated that strain differences between DENV-3 viruses contributed to transmission and disease severities in Indonesia.[14,24]
Antibody-dependent enhancement of infection (ADE) concept, which is believed to play a role in the pathogenesis of DHF/DSS, arose from the observation that secondary infection with heterotypic dengue virus is associated with an increased risk of DHF.[6,26] The present study showed that enhancing antibody augments the propagation of DENV-3 isolates.
Mos t ep i topes invo l ved in the neutralization of dengue viruses are located on the E protein.[27] Multiple antigenic sites have been discovered in the E protein as non-neutralizing linear epitope E37-46, B and T cell epitope E35-55 and E79-99 in domain I, and E333-368 in Domain I as well as E375-399 dengue-complex epitope.[28,29] The E protein also contains determinants responsible for virus attachment to host cells and for fusion of virions with host cell membranes.[30] The E gene of DENV-3 is relatively conserved at the amino acid levels.[31] We demonstrated amino acid differences at three positions in the E protein (E-124, E-301, E-377) between DSS/DHF strains and DF strain. Bray and Gualano
reported that amino acid at E-124 and E-132 affected the neurovirulence in mice.[32,33] These regions also contain significant linear and, possibly, conformational epitope and fusion domain.[34,35]
The amino acid difference at E-377 has not been reported in any DENV-3 strains.[31,36,37] This difference is within the position E375-399 which contains dengue-complex epitope. Amino acid substitution at E-390 has been suggested to affect the virulence of DENV-2 virus in mice[38] and humans,[39] and reduction in virus output from monocyte-derived macrophage.[40] We hypothesize that the amino acid replacements in E protein, especially at the conserved region of E-377 (valine to isoleucine), may have some effect on the phenotype of DENV-3 and this will be examined in a future study.
The activation of dengue virus replication complex is composed of NS1, NS2A, NS3, NS4A, NS4B and the viral RNA template.[41] We found nine amino acid replacements in NS1 among four DENV-3 isolates. The NS1 glycoprotein is involved in the early step of viral replication.[42] The amino acid at the position 109 (DSS/DHF strains were methionine and DF strain was threonin) is located immediately upstream from the consensus linear epitope (NS1-111).[43]
The amino acid replacements were also observed in NS3 protein. Amino acid at the position 579 is located at the C-terminal region of NS3. NS3 protein demonstrates several activities associated with virus replication and some functions are located within a large C-terminal domain.[44] Duarte dos Santos et al. showed that amino acid replacement in the helicases domain of NS3 and E protein altered the efficiency of viral RNA production by the replication complexes, and thereby affect viral growth.[45] NS5 was the common nonstructural

56 Dengue Bulletin – Volume 33, 2009
Genotypic and phenotypic characteristics of DENV-3 in Indonesia
protein which contained high numbers of amino acid replacements between DHF and DF strains in our study.
The nucleotide substitutions at positions 10472 and 10596 at 3’UTR did not introduce a change in the predicted RNA secondary structure for the 98901403 and 98902890 strains, but those at position 10271 and 10305 introduced minor change in the predicted RNA secondary structure. The 3’UTR mutation with those in E gene and 5’UTR synergistically decreased dengue virus replication in human cells[23]. However, correlation among mutation at 3’region, RNA secondary structure and phenotype characteristic in dengue viruses is not defined yet.
Genetic evolution among the preM/M and E protein of dengue viruses has occurred independently within geographical regions where the viruses are endemic.[37] The alignment of nucleotide sequences followed by parsimony analysis allowed the generation of phylogenetic trees, demonstrating that geographically-independent evolution of DENV-3 had occurred.[37] Nucleotide sequence data and phylogenetic methods were widely used to understand a genetic relationship among dengue viruses as well as the epidemiology of viral disease. The phylogenetic analysis demonstrated that DENV-3 isolates in Indonesia clustered with DENV-3 strains from Malaysia 1974, Philippines, Fiji, Tahiti and the South Pacific in genotype I. A close genetic relationship was demonstrated between recent isolates with other previous isolates from Indonesia. The DF strain (98902890) showed a close relationship with those isolated in Indonesia over 20 years ago. Although this is not an intensive horizontal epidemiological study, the identified nucleotide substitutions between the reference strain (DENV-3 1973) and those isolated in 1998 suggest that some evolution of DENV-3 occurred in Indonesia.
Full-length sequencing studies of dengue viruses and comparison with different genotypes and distinct clinical phenotypes demonstrated some relationship between disease severity and particular sequence in structural and non-structural proteins, 5’ and 3’UTR.[11,39] Previous studies in vitro suggested that different viral genotypes exhibit different phenotypic characteristics. DENV-2 isolated from patients with different disease severities demonstrated different levels of growth in LLK-MK2 cell,[11] peripheral blood leukocytes and C6/36 cells[46]. In the present study, DENV-3 isolates from different disease severities also demonstrated unique amino acid substitutions and different levels of growth in U937 cell line.
The mechanisms involved in the pathogenesis of DHF are complex and are likely to include multiple viral and host determinants. The result in the present study may provide a clue to the contribution of viral determinants to the pathogenesis of DHF. We did not focus on identifying amino acid substitution that is responsible for the phenotypic change; we plan to address this question by using infectious clone technology in future studies.
Acknowledgements
We thank Dr S. Tajima, Mr A. Kotaki, Dr M. Ito and Ms R. Nerome, Laboratory of Vector-Borne Viruses, Department of Virology 1, National Institute of Infectious Diseases, for their technical help. This study was supported by the grant-in-aid (No. 12877045) from the Ministry of Education, Sciences, Sports and Culture, Japan, and the grant for Research on Emerging and Re-Emerging Infectious Diseases from the Ministry of Health, Labour and Welfare, Japan.

Dengue Bulletin – Volume 33, 2009 57
Genotypic and phenotypic characteristics of DENV-3 in Indonesia
ReferencesBosch I, Xhaja K, Estevez L, Raines G, [1] Melichar H, Warke RV, Fournier MV, Ennis FA, Rothman AL. Increased production of interleukin-8 in primary human monocytes and in human epithelial and endothelial cell lines after dengue virus challenge. J Virol 2002;76:5588-5597.
Bray M, Men R, Tokimatsu I, Lai CJ. Genetic [2] determinants responsible for acquisition of dengue type 2 virus mouse neurovirulence. J Virol 1998;72(2):1647-1651.
Chambers TJ, Hahn CS, Galler R, Rice CM. [3] Flavivirus genome organization, expression, and replication. Annu Rev Microbiol 1990;44:649-688.
Chen CJ, Kuo MD, Chien LJ, Hsu SL, Wang YM, [4] Lin JH. RNA-protein interactions: involvement of NS3, NS5, and 3’ noncoding regions of Japanese encephalitis virus genomic RNA. J Virol 1997;71(5): 3466-3473.
Cologna R, Rico-Hesse R. American genotype [5] structures decrease dengue virus output from human monocytes and dendritic cells. J Virol 2003;77(7):3929-3938.
Corwin AL, Larasati RP, Bangs MJ, Wuryadi [6] S, Arjoso N, Sukri, Listyaningsih E, Hartati S. Namursa R, Anwar Z, Chandra S, Loho B, Ahmad H, Campbell JR, Porter KR. Epidemic dengue transmission in southern Sumatra, Indonesia. Trans R Soc Trop Med Hyg 2001;95:257-265.
Crill WD, Roehrig JT. Monoclonal antibodies [7] that bind to domain III of dengue virus E glycoprotein are the most efficient blockers of virus adsorption to Vero cells. J Virol 2001;75(16):7769-7773.
Diamond MS, Edgil D, Roberts TG, Lu B, Harris [8] E. Infection of human cells by dengue virus is modulated by different cell types and viral strains. J. Virol 2000;74:7814-7823.
Duarte dos Santos CN, Frenkiel MP, Courageot [9] MP, Rocha CF, Vazeille-Falcoz MC, Wien MW, Rey FA, Deubel V, Despres P. Determinants in the envelope E protein and viral RNA helicase NS3 that influence the induction of apoptosis in response to infection with dengue type 1 virus. Virology 2000;274(2):292-308.
Falconar AK. Identification of an epitope [10] on the dengue virus membrane (M) protein defined by cross-protective monoclonal antibodies: design of an improved epitope sequence based on common determinants present in both envelope (E and M) proteins. Arch Virol 1999;144(12):2313-2330.
Gualano RC, Pryor MJ, Cauchi MR, Wright [11] PJ, Davidson AD. Identification of a major determinant of mouse neurovirulence of dengue virus type 2 using stably cloned genomic-length cDNA. J Gen Virol 1998; 9(Pt 3):437-446.
Gubler DJ, Suharyono W, Lubis I, Eram S, [12] Sulianti Saroso J. Epidemic dengue hemorrhagic fever in rural Indonesia. I. Virological and epidemiological studies. Am J Trop Med Hyg 1979;28(4):701-710.
Guirakhoo F, Hunt AR, Lewis JG, Roehrig [13] JT. Selection and partial characterization of dengue 2 virus mutants that induce fusion at elevated pH. Virology 1993;194(1):219-223.
Halstead SB. In vivo enhancement of [14] dengue virus infection in rhesus monkeys by passively transferred antibody. J Infect Dis 1979;140:527-533.
Halstead SB, O’Rourke EJ. Antibody-enhanced [15] dengue virus infection in primate leukocytes. Nature 1977;265(5596):739-741.
Ho LJ, Wang JJ, Shaio MF, Kao CL, Chang [16] DM, Han SW, Lai JH. Infection of human dendritic cells by dengue virus causes cell maturation and cytokine production. J Immunol 2001;166(3):1499-1506.

58 Dengue Bulletin – Volume 33, 2009
Genotypic and phenotypic characteristics of DENV-3 in Indonesia
Innis BL, Nisalak A, Nimmannitya S, [17] Kusalerdchariya S, Chongswasdi V, Suntayakorn S, Puttisri P, Hoke CH. An enzyme-linked immunosorbent assay to characterize dengue infections where dengue and Japanese encephalitis co-circulate. Am J Trop Med Hyg 1989;40(4):418-427.
It[18] o M, Takasaki T, Yamada K, Nerome R, Tajima S, Kurane I. Development and evaluation of fluorogenic TaqMan reverse transcriptase PCR assays for detection of dengue virus types 1 to 4. J Clin Microbiol 2004;42(12):5935-5937.
Khromykh AA, Sedlak PL, Guyatt KJ, Hall RA, [19] Westaway EG. Efficient trans-complementation of the flavivirus kunjin NS5 protein but not of the NS1 protein requires its coexpression with other components of the viral replicase. J Virol 1999;73(12):10272-10280.
Kliks SC, Nisalak A, Brandt WE, Wahl L, Burke [20] DS. Antibody-dependent enhancement of dengue virus growth in human monocytes as a risk factor for dengue hemorrhagic fever. Am J Trop Med Hyg 1989;40(4):444-451.
Lanciotti RS, Lewis JG, Gubler DJ, Trent DW. [21] Molecular evolution and epidemiology of dengue-3 viruses. J Gen Virol 1994;75(Pt 1):65-75.
Lee E, Gubler DJ, Weir RC, Dalgarno L. [22] Genetic and biological differentiation of dengue 3 isolates obtained from clinical cases in Java, Indonesia, 1976-1978. Archives of Virology 1993:133;112-125.
Leitmeyer KC, Vaughn DW, Watts DM, Salas [23] R, Villalobos I, de Chacon, Ramos C, Rico-Hesse R. Dengue virus structural differences that correlate with pathogenesis. J Virol 1999;73:4738-4747.
Libraty DH, Young PR, Pickering D, Endy TP, [24] Kalayanarooj S, Green S, Vaughn DW, Nisalak A, Ennis FA, Rothman AL. High circulating levels of the dengue virus nonstructural protein NS1 early in dengue illness correlate with the development of dengue hemorrhagic fever. J Infect Dis 2002;186(8):1165-1168.
Lindenbach BD, Rice CM. Genetic interaction [25] of flavivirus nonstructural proteins NS1 and NS4A as a determinant of replicase function. J Virol 1999;73(6):4611-4621.
Lok SM, Ng ML, Aaskov J. Amino acid and [26] phenotypic changes in dengue 2 virus associated with escape from neutralisation by IgM antibody. J Med Virol 2001;65(2):315-323.
Mangada MN, Igarashi A. Molecular and in vitro [27] analysis of eight dengue type 2 viruses isolated from patients exhibiting different disease severities. Virology 1998;244(2):458-466.
Morens DM, Marchette NJ, Chu MC, Halstead [28] SB. Growth of dengue type 2 virus isolates in human peripheral blood leukocytes correlates with severe and mild dengue disease. Am J Trop Med Hyg 1991;45(5):644-651.
Murgue B, Roche C, Chungue E, Deparis X. [29] Prospective study of the duration and magnitude of viraemia in children hospitalised during the 1996-1997 dengue-2 outbreak in French Polynesia. J Med Virol 2000;60(4):432-438.
Pandey BD, Igarashi A. Severity-related [30] molecular differences among nineteen strains of dengue type 2 viruses. Microbiol Immunol 2000;44(3):179-188.
R Putnak, Feighny R, Burrous R, Cochran [31] M, Hackett M, Smith G, Hoke C. Dengue-1 virus envelope glycoprotein gene expressed in recombinant baculovirus elicits virus-neutralizing antibody in mice and protects them from virus challenge. Am J Trop Med Hyg 1991;45:159-167.
Pryor MJ, Carr JM, Hocking H, Davidson AD, [32] Li P, Wright PJ. Replication of dengue virus type 2 in human monocyte-derived macrophages: comparisons of isolates and recombinant viruses with substitutions at amino acid 390 in the envelope glycoprotein. Am J Trop Med Hyg 2001:65(5):427-434.
Rice CM, Lenches EM, Eddy SR, Shin SJ, [33] Sheets RL, Strauss JH. Nucleotide sequence of yellow fever virus: implications for flavivirus gene expression and evolution. Science 1985;229(4715):726-733.

Dengue Bulletin – Volume 33, 2009 59
Genotypic and phenotypic characteristics of DENV-3 in Indonesia
Rico-Hesse R, Harrison LM, Salas RA, Tovar D, [34] Nisalak A, Ramos C, Boshell J, de Mesa MT, Nogueira RM, da Rosa AT. Origins of dengue type 2 viruses associated with increased pathogenicity in the Americas. Virology 1997;230:244-251.
Roehrig JT, Bolin RA, Kelly RG. Monoclonal [35] antibody mapping of the envelope glycoprotein of the dengue 2 virus, Jamaica. Virology 1998;246(2):317-328.
Roehrig JT, Risi PA, Brubaker JR, Hunt AR, [36] Beaty BJ, Trent DW, Mathews JH. T-helper cell epitopes on the E-glycoprotein of dengue 2 Jamaica virus. Virology 1994;198(1):31-38.
Sanchez IJ, Ruiz BH. A single nucleotide [37] change in the E protein gene of dengue virus 2 Mexican strain affects neurovirulence in mice. J Gen Virol 1996;77:2541-2545.
Scott RM, Nisalak A, Cheamudon U, [38] Seridhoranakul S, Nimmannitya S. Isolation of dengue viruses from peripheral blood leukocytes of patients with hemorrhagic fever. J Infect Dis 1980;141:1-6.
Uzcategui NY, Comach G, Camacho D, Salcedo [39] M, Cabello de Quintana M, Jimenez M, Sierra G, Cuello de Uzcategui R, James WS, Turner S, Holmes EC, Gould EA. Molecular epidemiology of dengue virus type 3 in Venezuela. J Gen Virol 2003;84(Pt 6):1569-1575.
Vaughn DW, Green S, Kalayanarooj S, [40] Innis BL, Nimmannitya S, Suntayakorn S, Endy TP, Raengsakulrach B, Rothman AL, Ennis FA, Nisalak A. Dengue viremia titer, antibody response pattern, and virus serotype correlate with disease severity. J Infect Dis 2000;181(1):2-9.
Watts DM, Porter KR, Putvatana P, Vasquez B, [41] Calampa C, Hayes CG, Halstead SB. Failure of secondary infection with American genotype dengue 2 to cause dengue haemorrhagic fever. Lancet 1999;354(9188):1431-1434.
Wills BA, Oragui EE, Stephens AC, Daramola [42] OA, Dung NM, Loan HT, Chau NV, Chambers M, Stepniewska K, Farrar JJ, Levin M. Coagulation abnormalities in dengue hemorrhagic Fever: serial investigations in 167 Vietnamese children with dengue shock syndrome. Clin Infect Dis 2002;35(3):277-285.
Wittke V, Robb TE, Thu HM, Nisalak A, [43] Nimmannitya S, Kalayanrooj S, Vaughn DW, Endy TP, Holmes EC, Aaskov JG. Extinction and rapid emergence of strains of dengue 3 virus during an interepidemic period. Virology 2002;301(1):148-156.
World Health Organization. [44] Dengue and Dengue hemorrhagic fever. Fact sheet no. 117. Geneva: WHO, 2002.
Wu HC, Huang YL, Chao TT, Jan JT, Huang JL, [45] Chiang HY, King CC, Shaio MF. Identification of B-cell epitope of dengue virus type 1 and its application in diagnosis of patients. J Clin Microbiol 2001;39(3):977-982.
Yamada K, Takasaki T, Nawa M, Kurane I. Virus [46] isolation as one of the diagnostic methods for dengue virus infection. J Clin Virol 2002;24, 203-209.

60 Dengue Bulletin – Volume 33, 2009
Dengue fever among ill-returned travellers and concurrent infection by two dengue virus serotypes
Khoa T.D. Thaia,b#, Josta A. Wismeijera, Michèle van Vugta,b, Katja C. Wolthersc and Peter J. de Vriesa,b
aDivision of Infectious Diseases, Tropical Medicine & AIDS, Academic Medical Center, Amsterdam, The Netherlands
bCenter for Infection and Immunity (CINIMA), Academic Medical Center, University of Amsterdam, Amsterdam, The Netherlands
cDepartment of Medical Microbiology, Laboratory of Clinical Virology, Academic Medical Center, Amsterdam, The Netherlands
Abstract
A sudden increase in dengue virus (DENV)-infected returned travellers was observed at the outpatient department of Tropical Medicine, Academic Medical Center, Amsterdam, The Netherlands.
A descriptive observational study was conducted to analyse the epidemiology, clinical manifestations and laboratory features of imported DENV-infected patients. From September 2008 to June 2009, a total of 45 ill-returned travellers suspected for dengue were prospectively and four ill-returned travellers retrospectively were included. The majority (32 out of 49, 65%) of patients returned after a visit to the Dutch Antilles or Suriname. DENV-1, DENV-2 and DENV-3 were found in 27 viraemic patients. We identified four patients with a concurrent DENV infection with DENV-1 and DENV-2 serotypes and described their clinical and laboratory features. The clinical signs and symptoms in DENV-infected patients were mild and variable. Leukopenia and thrombocytopenia were observed between three to six days after the onset of illness. The majority of the patients had elevated serum transaminases levels between 7 to 10 days after the onset of illness. Within the first six months of 2009, ~10% were diagnosed with dengue infection. DENV infection at our hospital is not a rare imported viral disease. Increased international travel with changing epidemiology and increasing frequency of dengue in the sub-tropics will induce imported DENV infections in Western countries, including The Netherlands.
Keywords: Dengue; epidemiology; concurrent; travellers; The Netherlands.
# E-mail: [email protected]; Tel.: +31 20 5664380; Fax: +31 20 6972286

Dengue Bulletin – Volume 33, 2009 61
Dengue virus infection in Dutch travellers
Introduction
The dengue virus (DENV) transmission takes place primarily through bites by the mosquito vectors, Aedes aegypti and Aedes albopictus. They feed preferentially on human blood and are often found in and around human dwellings.[1,2] Infection with any of the four DENV serotypes does not always cause symptoms. The most common clinical presentations are an undifferentiated febrile illness or more recognizable dengue fever (DF), but life-threatening manifestations such as dengue haemorrhagic fever (DHF) and dengue shock syndrome (DSS) may also occur.[3]
Dengue has become a major international public health problem due to its increasing geographical distribution. Dengue transition varies between countries, ranging from epidemic transmission with long inter-epidemic intervals to endemic with seasonal fluctuation.[4,5]
Parallel to the global expansion of dengue is an increasingly common diagnosis among travellers who consult the Department of Tropical Medicine of the Academic Medical Center, Amsterdam, The Netherlands. Between 1985 and 1994, 34 travellers, 4% of all travellers with fever, were diagnosed with dengue by serological testing.[6,7] The incidence among short-term travellers to south-east Asia was 30 per 1000 person months. It was also found that not every DENV infection among travellers caused the clinical form of the disease and the majority of DENV infections in travellers remained mild.[8]
From September 2008 onwards, a sudden increase in DENV infections was observed in travellers returning to The Netherlands from the Caribbean, mostly the Dutch Antilles and Suriname. Here we present the epidemiology,
clinical manifestations and laboratory findings of imported DENV infections among these travellers.
Methods
Study site and study population
Returning travellers presenting themselves at the Department of Tropical Medicine of the Academic Medical Center, with a history of ≤14 days of illness and who were suspected for dengue (based on signs, symptoms and travel history), were included in the study. We also included patients diagnosed with dengue retrospectively. (All patients attending our clinic are routinely informed that routinely collected clinical data may be used for scientific reporting. Patients are given the choice to not have their data included. None of the patients objected).
Blood samples were collected from each patient for routine diagnostic procedures. All samples were tested by enzyme-linked immunosorbent assay (ELISA) and/or rapid immunochromatographic test (RDT) for anti-DENV IgM and IgG antibodies and real-time polymerase chain reaction (RT-PCR). Patients with DENV infection, confirmed by serological and/or RT-PCR testing, were included in this study.
Dengue diagnostics
Serum samples were tested for dengue with direct IgG enzyme-linked immunosorbent assay (ELISA) and IgM-Capture ELISA and/or rapid immunochromatographic test (RDT) (Panbio Tech Co., Brisbane, Australia) according to the manufacturer’s instructions. RNA was isolated from plasma or serum as described elsewhere.[9] RNA was reverse transcribed, and DENV viraemia levels were assessed using

62 Dengue Bulletin – Volume 33, 2009
Dengue virus infection in Dutch travellers
an internally controlled, serotype-specific, real-time reverse-transcriptase polymerase chain reaction (RT-PCR) assay that has been described elsewhere; the results were expressed as cDNA equivalents per ml of serum.[10] The RT-PCR and ELISA results were used for the diagnosis and classification of primary and secondary dengue infections.
Classification of primary and secondary dengue
Acute primary DENV infections were confirmed by RT-PCR and/or dengue serum-specific IgM antibodies detection in acute samples, in the absence of both dengue serum-specific IgG antibodies. A negative first acute sample for dengue serum specific IgM and IgG antibodies with seroconversion for dengue serum-specific IgM antibodies within 7 to 14 days was also considered as primary dengue. Acute secondary DENV infections were confirmed on acute samples (≤14 days) with positive IgG results and detection of viral genome, either with or without detectable dengue serum-specific IgM antibodies. Patients who presented with both positive IgM and IgG results in the first acute sample, without viral genome detection by PCR, were assessed by the IgM/IgG ratio. A ratio of anti-dengue IgM to IgG ≥1.8 was the criterion for primary dengue infection and a ratio of IgM to IgG <1.8 was considered as secondary infection.[11]
Statistical analysis
All calculations were performed using SPSS (version 17.0, SPSS Inc., Illinois) and SigmaPlot (version 11.0, Systat Software Inc., San Jose). The results were summarized in terms of means for continuous variables. For dichotomous variables, Fisher’s exact test was performed. A P-value of <0.05 was considered as statistically significant.
Results
Study population and travel history
From September 2008 to June 2009, a total of 45 ill-returned travellers were prospectively included. In addition, four such travellers were retrospectively included based on their clinical presentation, duration of illness and travel history. The demographic information and travel history in those 49 ill-returned travellers by diagnosis are shown in Table 1. Figure 1A shows a sudden increase in DENV-infected patients. The majority (32 out of 49) had returned from a visit to the Dutch Antilles or Suriname.
Diagnosis of DENV infection parity and virological features
Viral genomes were detectable in 27 out of 40 patients, of which 18 were classified as primary and 9 as secondary DENV infection. The viraemia levels ranged from 2.4 × 102/mL to 1.6 × 109/mL (mean of 8.7 x 107/mL). Figure 1B shows the distribution of viraemia levels in ill-returned travellers.
Clinical and laboratory features
The clinical manifestations of DENV infection in returned travellers are summarized in Table 2.
Figures 2A and 2B show the laboratory features in the acute phase of illness. Leukopenia (<3.1 x 103/µL) and thrombocytopenia (<1.0 x 105/µL) was observed in 12 and 13 of the 40 DENV-infected patients in the acute phase (≤10 days of illness), respectively, and two patients had marked thrombocytopenia with a platelet count <5.0 x 104/µL. Among those patients tested, most had increased liver enzyme levels.

Dengue Bulletin – Volume 33, 2009 63
Dengue virus infection in Dutch travellers
Table 1: Demographic data of ill-returned travellers
DF (n = 40) OFI (n = 9) P
Male/female 17/23 6/3 NS
Age* 38.2 (17.4-73.6)
40.8 (26.3-64.0)
NS
Days ill at presentation* 6.1 (0-13) 4.0 (0-9) NS
Dengue classification
PrimarySecondary
2416
——
Serotype
DENV-1DENV-2DENV-3DENV-1/DENV-2
17424
————
Travel history Africa1
Asia2
Central America/Caribbean3
South America4
—101911
2232
* mean (range) DF: Dengue fever; OFI: Other febrile illness; NS: not significant 1Mauritius; 2Indonesia, Thailand, Cambodia; 3Aruba, Bonaire, Curacao, Dutch Antilles, Panama; 4Suriname
Figure 1: DENV-infected returned travellers with their virological features [(A) Number of confirmed DENV-infected patients by month presented at the Academic
Medical Center, Amsterdam, The Netherlands, February 2008 – June 2009. (B) Distribution of viral loads of 27 DENV-infected patients with a single serotype by DENV infection parity.
Four with dual DENV infection with DENV-1 and DENV-2, shown as and DENV-2 as , respectively]

64 Dengue Bulletin – Volume 33, 2009
Dengue virus infection in Dutch travellers
The mean WBC and platelet counts over the course of the disease decreased up to six days after the onset of illness and increased rapidly to normal values. The most abnormal values of WBC were observed between three and six days after the onset of illness, whereas the most abnormal aspartate transaminase (ASAT) and alanine transaminase (ALAT) values were found beyond seven days of illness. After 14 days of illness, the ASAT and ALAT values decreased, although the majority of values still exceeded the reference values.
Concurrent DENV infection by two serotypesAmong the 27 viraemic DENV-infected returned travellers, there were four patients with a dual DENV-1 and DENV-2 infection. All had a primary DENV infection immune response and the viraemia levels of DENV-1 were higher than in DENV-2 (Figure 1B). Clinical and laboratory features are shown in
Table 2: Clinical presentations in 36 returned travellers with dengue fever
Signs/symptoms Frequency (%) (n=36*)
SymptomsFeverHeadacheMuscle painRetro-orbital painArthralgiaAnorexiaNauseaDiarrhoeaVomitingFatigue Cough
1006967534231281111178
SignsRashPetechiaeSwollen red handsHepatomegaly
17171714
*four patients with concurrent DENV-1 and DENV-2 infection are presented in Table 3.
Figure 2: DENV-infected returned travellers with laboratory features [(A) WBC counts and platelets counts distribution by days after onset of illness. Data show
mean value and standard error with upper and lower limits. (B) Distribution of ASAT and ALAT values by days after onset of illness. Data show mean value and standard error with upper and
lower limits, 1 number of samples per time point.]
Days of illness (groups)Maximum valueMinimum valueUpper reference value
Maximum valueMinimum valueUpper & lower reference value
Days of illness (groups)0-2 3-6 7-10 11-14 >15 0-2 3-6 7-10 11-14 >15
10
10
10
3
2
1
10
5
0
600500400300200100
A B
Cellsx10
/µL
3
U/L
Platelet1 5 22 20 9 10
5 21 18 8 13WBC1
ALAT1 5 18 14 8 115 21 15 8 13ASAT1

Dengue Bulletin – Volume 33, 2009 65
Dengue virus infection in Dutch travellers
Table 3: Clinical and laboratory data of four DENV-infected patients with concurrent DENV-1 and DENV-2
Patients 1 2 3 4
Age/sex 38/M 43/M 45/M 41/F
Clinical symptoms Fever Fever, headache,
myalgia
Fever, retro-ortibal pain, myalgia, examthema
Fever, retro-ortibal pain, headache, arthralgia, myalgia, erythema
Platelets (150-400 x 103/µL)
198 202 151 128
White blood cell (2-7.2 x 103/µL)
4.2 2.9 4.1 1.5
Neutrophil (50-60 %)
45.4 60.3 59.1 65.4
Lymphocyte (25-30 %)
35.0 23.2 22.7 26.8
Monocyte (2-10 %)
19.4 15.4 18.0 7.8
Basophil (<1 %)
0.2 1.1 0.2 0.0
ASAT (<40 U/L)
43 42 22 31
ALAT (<50 U/L)
61 46 15 19
C-reactive protein (<10 mg/L)
7.5 20 8.2 4.7
Haemaglobulin (M: 8.5-11.0 mmol/L F: 7.5-10.0 mmol/L)
9.9 9.4 9.2 8.1
Haematocritt (M: 0.41-0.51 L/L F: 0.36-0.47 L/L)
ND 0.45 0.44 0.38
Days after of illness 2 1 1 2
M: male; F: female; ASAT: aspartate aminotransferase; ALAT: alanine aminotransferase.

66 Dengue Bulletin – Volume 33, 2009
Dengue virus infection in Dutch travellers
Table 3. Leukopenia was found in two patients and the WBC count differentials shows monocytosis (>6%) in all patients. Platelet counts were normal and transaminases were slightly elevated in three patients.
Discussion
This study reports a sudden increase of DENV infection in returning travellers who presented at the Academic Medical Center, which has not been documented before in The Netherlands. The majority (65%) of patients returned after a visit to the Dutch Antilles or Suriname. DENV-1, DENV-2 and DENV-3 were found in travellers returning from this area, which is suggestive of co-circulation of multiple DENV serotypes in those areas. Clinical signs and symptoms in the DENV-infected patients were mild and variable. The majority of them had elevated transaminases (ASAT and ALAT) with the most abnormal levels between 7 to 10 days post onset of illness. Leukopenia and thrombocytopenia were observed between three to six days after onset of illness. Among the 27 viraemic patients, we identified four patients with a concurrent DENV infection with DENV-1 and DENV-2 serotypes and described their clinical and laboratory features.
Due to the spread of the vector, the geographical distribution of dengue is expanding rapidly in tropical and subtropical countries.[12] Growing international travel coupled with increasing transmission or re-emergence and changing epidemiology of dengue in various sub-tropical countries may have resulted in a steady rise in confirmed DENV infection among ill-returned travellers who presented at our hospital. In this present study, we have shown a sudden increase in DENV infection in travellers returning from the
Caribbean, including Aruba, Bonaire, Curacao, Dutch Antilles, Panama and Suriname. Reports of dengue infection in travellers from south-east Asia are by far more frequent than from the Americas.[13] This difference may reflect different travel patterns and destinations but also differences in local transmission intensities. Depending on the study design, the study population and the destinations, the prevalence of dengue in travellers varies.[14-16] A previous prospective study at our hospital showed that the incidence rate among Dutch short-term travellers with destinations in endemic areas in Asia was 30 per 1000 person-months, with a clinical/subclinical infection ratio of 1:3.3.[8] In long-term travelers, with a median of 5.3 months’ stay in endemic countries, it showed a seroconversion rate of 6.7%.[17] The incidence rates may differ during epidemics. A high attack rate (69%) during a DENV epidemic among aid workers was reported on a Caribbean island.[18] Variable attack rates and clinical/subclinical ratios may occur, possibly reflecting the role of different virulent virus strains, the importance of initial viral load or entomological or host factors.
Most of the clinical manifestations and laboratory features presented in our study agree with the previous reports.[14,15,19,20] Fever, headache (mostly frontal or retro-orbital), arthralgia, rash and myalgia were important clinical characteristics; however, swollen red hands have never been observed. The most helpful laboratory results were: leukopenia, thrombocytopenia and increased transaminases. All patients were managed as outpatients and elevated transaminases were found in the majority, including a few patients who showed values higher than ten times the upper limit.
The rapid spread of dengue, coupled with co-circulation of multiple DENV serotypes,

Dengue Bulletin – Volume 33, 2009 67
Dengue virus infection in Dutch travellers
have been increasingly reported in the last six decades.[5] Despite the geographical expansion, reports of simultaneous infection of patients with more than one DENV serotype remain low in comparison with the frequency of DENV infections. Concurrent infection with multiple DENV serotypes has been reported in several countries, mainly from south-east Asia and South America.[21-25] Among the DENV-infected patients in this study, four patients with concurrent DENV infection with DENV-1 and DENV-2 serotypes were detected by RT-PCR assay. All had a primary immune response against DENV. Clinical presentation and laboratory features (data not shown) were similar to other DENV-infected returned travellers. The RT-PCR assay used in this study detected all four DENV serotypes and is used extensively in many other studies. However, we did not attempt other techniques, e.g. virus isolation, to support our findings because RT-PCR techniques have been shown to be the most robust methods and are more specific than any other viral detection method.[26]
Conclusion
This descriptive study shows the sudden rise in the number of DENV-infected travellers returning from the Dutch Antilles and Suriname. It also records the co-circulation of multiple DENV by detecting DENV-1, DENV-2 and DENV-3 in the ill-returned travelers. Most interestingly, we showed the concurrent infection with DENV-1 and DENV-2 serotypes. The growth in international travel, with changing epidemiology and increasing incidence of dengue in the tropics, will lead to an increase in imported DENV infections in Western countries, including The Netherlands. DENV infection at our hospital, therefore, is not a rare occurrence of imported viral disease.
Acknowledgements
K.T.D. Thai is supported by a ‘Mosaic’ fellowship from The Netherlands Organization for Scientific Research (NWO).
ReferencesHarrington LC, Edman JD, Scott TW. Why do [1] female Aedes aegypti (Diptera: Culicidae) feed preferentially and frequently on human blood? J Med Entomol 2001;38(3):411-422.
Scott TW, Amerasinghe PH, Morrison AC, [2] Lorenz LH, Clark GG, Strickman D, Kittayapong P, Edman JD. Longitudinal studies of Aedes aegypti (Diptera: Culicidae) In Thailand and Puerto Rico: Blood feeding frequency. J Med Entomol 2000;37(1):89-101.
Rigau-Perez JG, Clark GG, Gubler DJ, [3] Reiter P, Sanders EJ, Vorndam AV. Dengue and dengue haemorrhagic fever. Lancet 1998;352(9132):971-977.
Gibbons RV, Vaughn DW. Dengue: an escalating [4] problem. BMJ. 2002;324(7353):1563-1566.
Guzman MG, Kouri G. Dengue: An update. [5] Lancet Infect Dis 2002;2(1):33-42.
Bakker RC, Veenstra J, Dingemans-Dumas [6] AM, Wetsteyn JCFM, Kager PA. Imported dengue in the Netherlands. J Travel Med 1996;3(4):204-208.
Wetsteyn JC, Driessen SO, de Vries PJ, [7] Cobelens FG, Kager PA. [Tropical diseases and imported disorders in 1763 patients seen at the Outpatient Clinic for Tropical Diseases, Academic Medical Center, Amsterdam (1996-1997)]. Ned Tijdschr Geneeskd. 2000;144(45):2152-2156.

68 Dengue Bulletin – Volume 33, 2009
Dengue virus infection in Dutch travellers
Cobelens FG, Groen J, Osterhaus AD, [8] Leentvaar-Kuipers A, Wertheim-van Dillen PM, Kager PA. Incidence and risk factors of probable dengue virus infection among Dutch travellers to Asia. Trop Med Int Health 2002;7(4):331-338.
Boom R, Sol C, Beld M, Weel J, Goudsmit [9] J, Wertheim-van Dillen P. I Improved silica-guanidiniumthiocyanate DNA isolation procedure based on selective binding of bovine alpha-casein to silica particles. J Clin Microbiol 1999;37(3):615-619.
Laue T, Emmerich P, Schmitz H. Detection of [10] dengue virus RNA in patients after primary or secondary dengue infection by using the TaqMan automated amplification system. J Clin Microbiol 1999;37(8):2543-2547.
Innis BL, Nisalak A, Nimmannitya S, [11] Kusalerdchariya S, Chongswasdi V, Suntayakorn S, Puttisri P, Hoke CH. An enzyme-linked immunosorbent assay to characterize dengue infections where dengue and Japanese encephalitis co-circulate. Am J Trop Med Hyg 1989;40(4):418-427.
Kroeger A, Nathan MB. Dengue: setting [12] the g lobal research agenda. Lancet 2006;368(9554):2193-2195.
Schwartz E, Weld LH, Wilder-Smith A, von [13] Sonnenburg F, Keystone JS, Kain KC, Torresi J, Freedman DO; GeoSentinel Surveillance Network. Seasonality, annual trends, and characteristics of dengue among ill returned travelers, 1997-2006. Emerg Infect Dis 2008;14(7):1081-1088.
Jelinek T, Dobler G, Hölscher M, Löscher T, [14] Nothdurft HD. Prevalence of infection with dengue virus among international travelers. Arch Intern Med 1997;157(20):2367-2370.
López-Vélez R, Pérez-Casas C, Vorndam AV, [15] Rigau J. Dengue in Spanish travelers returning from the tropics. Eur J Clin Microbiol Infect Dis 1996;15(10):823-826.
Schwar t z E , Mende l son E , S id i Y. [16] Dengue fever among travelers. Am J Med 1996;101(5):516-520.
Potasman I, Srugo I, Schwartz E. Dengue [17] seroconversion among Israeli travelers to tropical countries. Emerg Infect Dis 1999;5(6):824-827.
Lyerla R, Rigau-Pérez JG, Vorndam AV, [18] Reiter P, George AM, Potter IM, Gubler DJ. A dengue outbreak among camp participants in a Caribbean island, 1995. J Travel Med 2000;7(2):59-63.
Badiaga S, Delmont J, Brouqui P, Janbon F, [19] Durant J, Bosseray A, Malvy D, Bonnet E, Sotto A, Dydymski S, Peyramond D. [Imported dengue: study of 44 cases observed from 1994 to 1997 in 9 university hospital centers. Infectio-Sud-France group]. Pathol Biol (Paris) 1999;47(5):539-542.
Badiaga S, Barrau K, Brouqui P, Durant J, [20] Malvy D, Janbon F, Bonnet E, Bosseray A, Sotto A, Peyramont D, Dydymski S, Cazorla C, Tolou H, Durant JP, Delmont J; Infectio-Sud Group. Imported Dengue in French University Hospitals: a 6-year survey. J Travel Med 2003;10(5):286-289.
Gubler DJ, Kuno G, Sather GE, Waterman SH. [21] A case of natural concurrent human infection with two dengue viruses. Am J Trop Med Hyg 1985;34(1):170-173.
Kukreti H, Chaudhary A, Rautela RS, Anand [22] R, Mittal V, Chhabra M, Bhattacharya D, Lal S, Rai A. Emergence of an independent lineage of dengue virus type 1 (DENV-1) and its co-circulation with predominant DENV-3 during the 2006 dengue fever outbreak in Delhi. Int J Infect Dis 2008;12(5):542-549.
Wang WK, Chao DY, Lin SR, King CC, Chang [23] SC. Concurrent infections by two dengue virus serotypes among dengue patients in Taiwan. J Microbiol Immunol Infect 2003;36(2):89-95.

Dengue Bulletin – Volume 33, 2009 69
Dengue virus infection in Dutch travellers
Wenming P, Man Y, Baochang F, Yongqiang [24] D, Tao J, Hongyuan D, Ede Q. Simultaneous infection with dengue 2 and 3 viruses in a Chinese patient return from Sri Lanka. J Clin Virol 2005;32(3):194-198.
Wilder-Smith A, Yoksan S, Earnest A, [25] Subramaniam R, Paton NI. Serological
evidence for the co-circulation of multiple dengue virus serotypes in Singapore. Epidemiol Infect 2005;133(4):667-671.
Wilder-Smith A, Schwartz E. Dengue in travelers. [26] N Engl J Med 2005;353(9):924-932.

70 Dengue Bulletin – Volume 33, 2009
Acute abdominal pain in dengue haemorrhagic fever: A study in Sri Lanka, 2009
K.G.A.D. Weerakoona, S. Chandrasekaramb, J.P.S.N.K. Jayabahub, S. Gunasenac and S.A.M. Kularatnea#
aDepartment of Medicine, Faculty of Medicine, University of Peradeniya, Sri Lanka
bTeaching Hospital, Peradeniya, Sri Lanka
cDepartment of Virology, Medical Research Institute, Colombo 08, Sri Lanka
Abstract
We attempted to unravel the association of acute severe abdominal pain in 14 patients with dengue haemorrhagic fever during the epidemic in 2009. All had secondary dengue infection. The findings were severe thrombocytopenia (the mean platelet count 18x109/l, range 12–48), high liver enzymes (mean ALT 374U/l, range 82–2692), ascites in all cases, acute hepatitis in 11 cases (79%), acalculous cholecystitis in five cases (42%), and renal involvement in three cases (25%). All had normal serum amylase level and normal-looking pancreas in ultrasound scan. The abdominal pain in dengue infection warrants investigation to find a specific cause.
Keywords: Dengue haemorrhagic fever; secondary infection; abdominal pain; severe thrombocytopenia; Sri Lanka.
# E-mail: [email protected]; Tel.: +94773420771
Introduction
Dengue, an endemic infection in the tropical and subtropical regions of the world, is widely distributed in many countries in Asia, including Sri Lanka.[1,2] Over the past two decades, changing epidemiology with regular epidemics of dengue fever (DF) and dengue haemorrhagic fever (DHF) have been reported in Sri Lanka.[2,3] All four serotypes, DENV-1 to DENV-4, are in circulation and are responsible for the changing face of epidemics.[2,3]
The protean character of dengue fever ranges from mild febrile illness to profound shock; thus, it remains a challenge for saving lives. The common symptoms in dengue infection are fever, malaise, headache, musculoskeletal pain, nausea and vomiting. Nonetheless, a significant number of patients develop one or more complications that include bleeding, dengue shock syndrome (DSS), acute renal failure and seizures.[4,5] Furthermore, other complications such as acute myocarditis,[6,7] effusions, acute hepatic

Dengue Bulletin – Volume 33, 2009 71
Acute abdominal pain in dengue haemorrhagic fever
failure[8] and dengue encephalitis have also been reported.[9,10] The abdominal pain in dengue infection is a dilemma which could be either specific or non-specific. Of the specific cases, surgical emergencies like acute pancreatitis,[11] acute acalculous cholecystitis[12] and gastrointestinal bleeding[13] are found in literature. In addition, there are reports of dengue enteritis mimicking appendicitis.[14,15] However, in many cases, even in severe abdominal pain, the causes could not be found.
In mid-2009, Sri Lanka faced a severe epidemic of dengue infection with the highest case-load and deaths. In the midst of this epidemic, we observed frequent occurrence of abdominal pain in patients presenting to the Teaching Hospital, Peradeniya (THP), in the Central Province of the country. We investigated 14 such patients with dengue haemorrhagic fever, who presented with acute severe abdominal pain.
Materials and methods
We possess rich clinical records of all dengue infections admitted to the THP during May–August 2009. Of the total 337 patients with diagnosed dengue infections, those with severe abdominal pain were included in the analysis. In each patient, details such as symptoms, signs and medical management were recorded. All of them were managed according to the routine protocol of the Unit. Investigations such as regular measurement of full blood counts, blood urea, serum creatinine level, aspartate aminotransferase (AST), alanine aminotransferase (ALT), serum amylase, blood culture, ascetic fluid culture, ECG, chest radiograph and ultrasound scan of the abdomen were done to detect complications. The ultrasound scan was
performed meticulously by the same radiologist to avoid personal bias.
Confirmation of the diagnosis was made by doing dengue-specific IgM and haemagglutination inhibiting (HI) antibodies test, and in selected cases, by viral identification with reverse transcriptase polymerase chain reaction agarose gel electrophoresis (RT-PCR-AGE) at Medical Research Institute, Colombo, Sri Lanka. The RT-PCR-AGE was done for patients who presented within four days of onset of fever. Paired sera were tested by the HIA and for IgM antibody using IgM antibody capture ELISA. If only dengue virus-specific IgM antibodies were detectable in the test sample, the patient was considered to have primary dengue infection, whereas the presence of both IgM and IgG or HIA titre above 1:2560, or both together, was considered as secondary dengue infection.
Results
The analysis included 14 patients with acute abdominal pain severe enough to keep them only on intravenous fluids until they were able to tolerate oral fluids. The series comprised males and females in equal numbers, with a median age of 28 years (range 12–47 years). All qualified the WHO criteria of DHF and, of them, four were included in the category of DSS.
Apart from fever, headache and bleeding tendency, many patients had vomiting (Table 1). The abdominal pain was more or less generalized in four patients and the others had pain located mainly in the epigastric region. The mean date of the onset of the abdominal pain was four days, with a range of two to five days. The mean duration of the abdominal pain was three days ranging from one to five days. Serologically, all had secondary dengue infection.

72 Dengue Bulletin – Volume 33, 2009
Acute abdominal pain in dengue haemorrhagic fever
Table 1: Other clinical features of 14 patients with abdominal pain
Clinical feature No (%)
Fever 14(100)
Headache 13 (93)
Arthralgia 8 (57 )
Myalgia 13 (93)
Vomiting 11 (79)
Bleeding tendency 14 (100)
Skin rash 2 (14)
A significant thrombocytopenia was the hallmark in the series with spontaneous bleeding tendency (Table 2). Thirteen patients had elevated levels of AST and all 14 had elevated ALT. Interestingly, the serum amylase levels were normal in all patients (Table 2).
Table 2: Investigation results of 14 patients
Investigation Mean Range
Platelet count (x109/l)
18 12–48
SGPT (IU/l) 374 82–2692
SGOT (IU/l) 418 163–2382
Serum amylase (IU/l)
57 34–97
The ultrasound scan of the abdomen showed ascitis, hepatomegaly, splenomegaly, acute liver parenchymal disease, acute aclaculous cholecystitis with pericholecystic fluid collection, gall bladder distension and wall oedema and renal paranchymal involvement (Table 3). The renal involvements were acute renal parenchymal changes, pyelonephritis and perinephric fluid collection. Extra attention was paid to the pancreas, but all had normal-looking pancreas.
Table 3: Ultrasound scans findings of 14 patients
Finding No (%)
Ascitis 14 (100)
Hepatomegaly 12 (86)
Hepatitis 11 (79)
Splenomegaly 3 (25)
Acalculous cholecystitis
5 (42)
Renal paranchymal changes
3 (25)
Pancreatitis Nil
Blood cultures and ascetic fluid cultures were negative and a few patients had ECG changes. The RT-PCR-AGE gave negative results of a limited number of cases where it was done. All these patients were managed with antibiotics and supportive care and they recovered completely.
Discussion
We found that abdominal pain was non-specific in these cases, except in five cases where it could be attributed to acalculous cholecystitis. The cardinal finding was the presence of ascites and none had any form of pancreatitis. The pathogenesis of abdominal pain in dengue fever is not clearly understood; however, lymphoid follicular hyperplasia seems to play an important role and plasma leakage through damaged capillary endothelium had also been proposed.[14,15] It could be the possible explanation for subserosal fluid collection and thickened gall bladder associated with dengue fever. The specific organ involvement, like hepatitis, acalculous cholecystitis, pancreatitis, appendicitis and peptic ulcer disease, could also produce abdominal pain.[15,16]

Dengue Bulletin – Volume 33, 2009 73
Acute abdominal pain in dengue haemorrhagic fever
Acute liver involvement with hepatitis and hepatomegaly was detected in more than 78% of the current case series. They all had elevated liver enzymes and, in some cases, the rise was alarmingly high. However, none of these patients went into acute liver failure. The rise of liver enzyme in DHF due to acute liver parenchymal disease had been discussed.[17] Acute cholecystitis is an increasingly reported manifestation in DHF. The pathogenesis of acute cholecystitis in DHF is not fully understood, but may result from localized microangiopathy in the gall bladder wall.[18] The role of secondary bacterial infection causing acute cholecystitis is not clear, but administration of broad-spectrum antibiotics helped our patients.
Even though pancreatitis has been reported in dengue haemorrhagic fever,[16] none of these 14 patients had any evidence of pancreatitis such as elevation of serum amylase or ultrasound scan findings. But a pregnant mother, who succumbed to dengue infection at the beginning of the epidemic at THP showed florid pancreatitis at necropsy (unpublished
data). The three patients in the series who had renal involvement showed acute renal parenchymal disease and perinephric fluid collection in the ultrasound examination. Of them, the diagnosis of pyelonepritis was made in one patient who had pus cells and white cell casts in the urine full report. This patient was treated with antibiotics considering secondary bacterial infection, and the patient recovered. Furthermore, acute renal failure associated with DHF has been reported and one explanation given is the myoglobinuria in DHF causing glomerular damage.[19,20]
All cases in this series had a severe form of the disease (DHF and DSS) caused by secondary exposure to dengue virus. Identification of the causative serotype was not possible in these cases, but DENV-1 was detected in five other patients during the current epidemic. Broad-spectrum antibiotics were given to all these patients as a desperate measure to prevent fatalities.
In conclusion, the abdominal pain in dengue infection warrants investigation to find out the specific cause.
ReferencesWilder-Smith A, Schwartz E. Dengue in [1] travelers. N Engl J Med 2005;353:924-32.
Kularatne SAM, Seneviratne SL, Malavige GN, [2] Fernando S, Velanthiri VGNS, Ranathunga PK, Wijewickrama ES, Gurugama PN, Karunathilaka DH, Aaskov JG, Jayaratne SD. Synopsis of findings from recent studies on dengue in Sri Lanka. Dengue Bulletin 2006;30:80-6.
Kanakaratne, Wahala MPB, Messer WB, [3] Tissera HA, Shahani A, Abeysinghe A, de Silva A, Gunasekera M. Severe Dengue Epidemics in Sri Lanka, 2003–2006. Emerg Infect Dis 2009;15(2):192-9.
Harris E, Videa E, Pérez L, Sandoval E, Téllez [4] Y, Pérez ML, Cuadra R, Rocha J, Idiaquez W, Alonso RE, Delgado MA, Campo LA, Acevedo F, Gonzalez A, Amador JJ, Balmaseda A. Clinical, epidemiologic, and virologic features of dengue in the 1998 epidemic in Nicaragua. Am J Trop Med Hyg 2000;63:5-11.
World Health Organizat ion. [5] Dengue haemorrhagic fever: diagnosis, treatment, prevention and control. Geneva: WHO, 2009.

74 Dengue Bulletin – Volume 33, 2009
Acute abdominal pain in dengue haemorrhagic fever
Promphan W, Sopontammarak S, Pruekprasert [6] P, Kajornwattanakul W, Kongpattanayothin A. Dengue myocarditis. Southeast Asian J Trop Med Public Health 2004;35:611-3.
Kularatne SAM, Pathirage MMK, Kumarasiri [7] PVR, Gunasena S, Mahindawanse SI. Cardiac complications of a dengue fever outbreak in Sri Lanka, 2005. Trans R Soc Trop Med Hyg 2007;101:804-8.
Vinodh BN, Bammigatti C, Kumar A, Mittal V. [8] Dengue fever with acute liver failure. J Postgrad Med 2005;51:322-3.
Muzaffar J, Venkata Krishnan P, Gupta N, Kar P. [9] Dengue encephalitis: why we need to identify this entity in a dengue-prone region. Singapore Med J 2006;47:975-7.
Kularatne SAM, Pathirage MMK, Gunasena [10] S. A case series of dengue fever with altered consciousness and electroencephalogram changes in Sri Lanka. Trans R Soc Trop Med Hyg 2008;102(10):1053-4.
Derycke T, Levy P, Genelle B, Ruszniewski [11] P, Merzeau C. Acute pancreatitis secondary to dengue. Gast roentero l C l in B io l 2005;29:85-6.
Goh BK, Tan SG. Case of dengue virus infection [12] presenting with acute acalculous cholecystitis. J Gastroenterol Hepatol 2006;21:923-4.
Tsai CJ, Kuo CH, Chen PC, Changcheng CS. [13] Upper gastrointestinal bleeding in dengue fever. Am J Gastroenterol 1999;86:33-5.
Premaratna R, Bailey MS, Ratnasena BG, [14] de Silva HJ. Dengue fever mimicking acute appendicitis. Trans R Soc Trop Med Hyg 2007;101:683-5.
Boon-Siang Khor, Jien-Wei liu, Ing-Kit lee, [15] and Kuender d. Yang. Dengue hemorrhagic fever patients with acute abdomen: clinical experience of 14 cases. Am J Trop Med Hyg 2006;74(5):901-904.
Khanna S, Vij JC, Kumar A, Singal D, Tandon [16] R. Dengue fever is a differential diagnosis in patients with fever and abdominal pain in an endemic area. Ann Trop Med Parasitol 2004;98:757-60.
Kuo CH, Tai DI, Chang-Chien CS, Lan CK, [17] Chiou SS, Liaw YF. Liver biochemical tests and dengue fever. Am J Trop Med Hyg 1992;47:265-70.
Hakala T, Nuutinen PJ, Ruokonen ET, Alhava [18] E. Microangiopathy in acute acalculous cholecystitis. Br J Surg 1997;84:1249-1252.
Gunasekera HH, Adikaram AV, Herath CA, [19] Samarasinghe HH. Myoglobinuric acute renal failure following dengue viral infection. Ceylon Med J 2000;45:181.
Wichmann D, Kularatne SAM, Ehrhardt S, [20] Wijesinghe S, Brattig NW, Abel W, Burchard GD. Cardiac involvement in dengue virus infections during the 2004/2005 dengue fever season in Sri Lanka. Southeast Asian J Trop Med Public Health 20

Dengue Bulletin – Volume 33, 2009 75
Involvement of the liver in dengue infectionsDuncan R. Smith# and Atefeh Khakpoor
Molecular Pathology Laboratory, Institute of Molecular Biosciences, Mahidol University, Thailand
Abstract
Dengue virus, the causative agent of dengue fever (DF) and more severe forms of the disease – dengue haemorrhagic fever (DHF) and dengue shock syndrome (DSS) – can infect a number of different types of cells. While monocytes and macrophages are considered to be primary target cells driving the pathology of the disease, numerous studies have implicated the liver as a site of dengue virus replication and both clinical observations and experimental data support a role of the liver in dengue disease. This review aims to provide a brief overview of the data supporting a role of the liver in the pathology of dengue disease.
Keywords: Liver; dengue virus; dengue fever; dengue haemorrhagic fever; dengue shock syndrome.
# E-mail: [email protected]; Tel.: (662) 800 3624; Fax (662) 441 9906
Introduction
With an estimated 75–500 million infections per year and 3.6 billion people living at the risk of infection,[1] dengue is one of the most important public health problems in most tropical and subtropical countries. The causative agent of dengue, the dengue viruses (DEN-V), are transmitted to humans by the bite of infected female mosquitoes belonging to the Aedes family, most commonly Aedes aegypti and Aedes albopticus.[2]
Infection with DENV can be asymptomatic or can result in a wide spectrum of disease, varying from a mild, non-specific viral illness to dengue fever (DF) to the more severe
forms of the disease – dengue haemorrhagic fever (DHF) and dengue shock syndrome (DSS). DF usually starts with a high fever and is often accompanied by a rash, headache and abdominal pains lasting 2–7 days. Frank haemorrhage is uncommon during DF but gastrointestinal bleeding, gingival bleeding and petechiae have been reported in some patients.[3] DHF can occur in primary infections but is more common in secondary infections and also starts with a high fever that is not significantly different from that observed in DF patients. Haemorrhagic phenomenon may be minimal, but around the time of defervescence, patients develop the hallmark of DHF, namely, significant plasma leakage, which in DSS is further characterized by

76 Dengue Bulletin – Volume 33, 2009
Involvement of the liver in dengue infections
tachycardia and hypotension.[4] If the plasma leakage is severe and remains untreated, patients may develop shock, which can be fatal.[4,5]
While the main manifestations of severe dengue disease are primarily haemorrhagic in nature, a significant body of work has been accumulated which implicates the liver as a critical part of the disease pathology.
Hepatomegaly
Perhaps the most obvious sign of the involvement of the liver in dengue infections is the high proportion of dengue cases with liver enlargement. In one of the earliest large-scale series of clinical investigations into dengue, Halstead and colleagues observed that the frequency of liver enlargement was similar in both primary and secondary dengue infections, and the authors proposed that a moderate liver enlargement may be a part of the “normal” pathological response to dengue infection.[6] More recent studies have been somewhat divided, with some reports suggesting that hepatomegaly is present at between 50–100% of cases[7-14] while others document a significantly lower rate of hepatomegaly.[15-17] In addition, some studies support a higher rate of hepatomegaly in DHF/DSS cases as opposed to DF cases[13,14] although this may not reach statistical significance.[14] On balance, the studies tend to support a high level of hepatomegaly in dengue cases, with perhaps a slightly higher rate in the more severe cases, although this may depend somewhat upon the exact case definitions used.
Liver enzymes
Upon injury to the liver, the enzymes, aspartate aminotransferase (AST) and alanine aminotransferase (ALT), are released into the
bloodstream, and as a consequence these enzymes are believed to be sensitive indicators of liver damage. Perhaps, unsurprisingly, these enzymes are frequently elevated in dengue patients, as has been shown in numerous studies.[7,9,12,14,16,18-22] In one large series of patients examined for both AST and ALT levels, Kuo and colleagues evaluated 240 dengue patients from the 1987–1988 outbreak in Taiwan.[20] Elevated levels of AST and ALT were found in 93.3% and 82.2% of cases respectively. While the majority of patients had only mildly or moderately elevated levels of these transaminases, some 10% (11% and 7% for AST and ALT respectively) of patients had levels elevated by 10-fold or greater. Somewhat lower levels of liver enzyme disorder were noted by de Souza and colleagues in their study of 1585 dengue patients, and they observed alterations of AST and ALT levels in 63% and 45% of patients respectively.[22] Interestingly, however, the authors noted that the average levels of AST and ALT were significantly higher in DHF patients than in DF patients, an observation supported by other studies.[9,14] Several authors have noted that the levels of serum AST are greater than serum ALT,[18,20,21] which is in contrast to the normal finding with viral hepatitis.[23] Some evidence has suggested that there is a greater degree of involvement of the liver in infections with DENV-3 and DENV-4.[24] Overall, the studies are consistent with elevated levels of liver enzymes being a common characteristic of dengue disease, and as such possibly represent a discriminating factor in differentiating dengue from other febrile diseases,[19] but is of less use in differentiating DF from DHF.
Liver specimen studies
While both hepatomegaly and alterations in liver enzymes point to the involvement of the liver in the disease, they are unable to distinguish between the result of a bystander

Dengue Bulletin – Volume 33, 2009 77
Involvement of the liver in dengue infections
or secondary effect.[25] As such, a number of studies have sought to provide direct evidence for the involvement of the liver in the disease process. The earliest of these studies undertook direct histological investigations of specimens from the livers of fatal cases of dengue infection.[26-28] The predominant findings in these studies were microvesicular steatosis and small foci of hepatocellular necrosis in addition to the presence of councilman bodies, Kupffer cell hyperplasia and mononuclear cell infiltrates at the portal tract.[26-28] In this respect, the liver damage seen in fatal dengue cases is significantly less severe than that seen in fatal cases of yellow fever virus infection.[29] While relatively uncommon, cases of fulminant hepatitis have also been documented.[30-32]
S e v e r a l s t u d i e s h a v e u s e d a n immunohistochemical approach to detect the presence of dengue ant igens in liver specimens.[33-40] These studies have predominantly used antibodies directed against dengue E protein,[34-40] although one recent study used an antibody directed against dengue NS3 protein,[33] which gives a greater degree of certainty that infected cells are undergoing viral replication and do not reflect the presence of endocytosed or phagocytosed virus particles without viral replication. The majority of the studies detect, to a greater or lesser extent, the presence of dengue antigen in hepatocytes, the major cell type composing the liver.[33-38,40] Interestingly, while some studies have suggested that 80–90% of hepatocytes show immunoreactivity,[36] other studies fail to detect dengue antigen in hepatocytes at all.[39] Whether this extreme difference is due to methodological or sample-preparation differences, or truly reflects a different tissue tropism of some dengue virus lineages remains unclear at this point. While some studies have detected the presence of dengue antigen in Kupffer cells,[36,38,39] the
study by Balsitis and colleagues did not detect the presence of immunoreactive NS3 in these cells,[33] suggesting thereby that Kupffer cells do not support replication of the dengue virus, and immunoreactivity to dengue structural proteins noted by others[36,38,39] may reflect phagocytosed virus. This would be consistent with the studies by Marianneau and colleagues who showed that dengue was efficiently taken up by isolated primary human Kupffer cells, but that the infection was non-productive.[41]
Dengue-specific RT-PCR and in situ hybridization has been employed in several studies to detect the presence of the dengue genome.[36,38,39,42-44] As with the studies utilizing immunohistochemistry, while the majority of the studies detect the presence of the dengue genome in liver samples, and more specifically in hepatocytes and to a lesser extent in Kupffer cells,[36,38,43,44] other studies do not detect the presence of the dengue genome in either hepatocytes or Kupffer cells.[39] Again, this may well reflect either methodological differences or inherent differences in virus tropism.
Several studies have recovered infectious dengue virus from liver specimens[27,38,45,46] and, in particular, Rosen and colleagues recovered infectious dengue serotype 2 or 3 from 5 out of 17 liver specimens from fatal cases of dengue infection through the mosquito inoculation technique[46]. However, these studies do not use defined populations of cells from the liver and as such, no information as to the cell types involved in the disease process is available.
Mouse model studies
A number of studies have utilized various mouse model systems to understand the pathogenesis of dengue infections.[33,47-56] Perhaps the simplest model is the intraperitoneal and intravenous injection of DENV-2 into BALB/c mice.[49,55,56] Histopathological analysis of these

78 Dengue Bulletin – Volume 33, 2009
Involvement of the liver in dengue infections
animals showed severe liver injury, including hepatocyte swelling and vacuolization, necrosis and steatosis. Liver damage was more severe, and occurred earlier in the intravenous injection system as compared to the intraperitoneal injection system.[49,55,56] Ultrastructural studies showed the accumulation of cytoplasmic lipid droplets as well as mitochondrial swelling in hepatocytes, possibly related to the induction of apoptosis.[55,56] Evaluation of liver enzymes showed a peak of both AST and ALT at seven days post-infection, which was correlated to the extent of liver injury.[55,56] Dengue virus antigens were detected not only in hepatocytes but also in Kupffer cells.[55,56]
While SCID mice are apparently resistant to dengue infection,[54] SCID mice transplanted with human liver cell lines HepG2 or Huh-7[47,57] or with K562 (a erythroleukemia cell line) cells[54] and intraperitoneally injected with DENV-2[47,54] or DENV-4[57] have all been used as model systems, with somewhat conflicting results. For example, while An and colleagues report high levels of the virus in the liver of SCID mice with transplanted HepG2 cells[47] in the early stage post-infection, Lin and colleagues report little or no involvement of the liver in the same SCID mice engrafted with human K562 cells.[54]
Immunocompetent C57BL/6 mice injected with a high titre of DENV-2[51] showed elevated levels of AST and ALT on days 3, 5 and 7 post-infection, which was elevated further after a second inoculation. Dengue RNA was detected in the liver of infected mice and a strong correlation was found between T cell activation and hepatic cellular infiltration. Dengue infection of interferon receptor-deficient (AG129) mice is uniformly lethal,[58] and the presence of immunoreactive NS3 was detected in hepatocytes, implying dengue virus replication in these cells[33]
being consistent with reports of dengue virus infection of human primary hepatocytes.[59]
While some of these studies failed to detect the involvement of the liver,[51,54] other studies have demonstrated a clear evidence of the involvement of the liver and of hepatocytes specifically.[33,55,56] Where liver involvement is seen in the model systems, the changes observed reflect the pathology of human disease.
Isolated primary liver cells
To date, only two studies have investigated the susceptibility to dengue infection of primary (untransformed) human liver cells.[41,59] The first study isolated Kupffer cells from liver sections obtained during partial hepatectomy for liver cancer[41] and demonstrated that while dengue virus entered into Kupffer cells efficiently, no viral progeny were produced and that infected cells underwent cell death by apoptosis. The second study[59] utilized commercially obtained hepatocytes purified from liver transplantation “cut-downs” and demonstrated productive infection with DENV-2 strain 16681 as assessed by plaque assay and cellular expression of both structural and non-structural proteins (E and NS1). A significant cytokine response was observed which included the up-regulation of TRAIL, MIP-1a, MIP-1b, IFN-b, IL-8 and RANTES. The profile of cytokine induction was similar, but not identical to the profile generated by DENV-2-infected HepG2 cells undertaken in parallel with the infection of the primary hepatocytes. Although not formally evaluated, evidence of the induction of apoptosis in response to DENV-2 infection was observed in the infected primary hepatocytes which showed extensive nuclear fragmentation.[59] Infected primary hepatocytes additionally

Dengue Bulletin – Volume 33, 2009 79
Involvement of the liver in dengue infections
showed the up-regulation of TRAIL,[59] a type-II transmembrane protein that has significant proapoptotic effect in liver cells,[60] and that has been proposed to be the primary mediator of apoptosis induction in dengue-infected transformed hepatocytes.[61]
Studies on transformed liver cells
Transformed liver cells are broadly permissive to dengue infection,[62,63] and a large number of studies have investigated cellular responses to dengue infection in transformed liver cells. These studies have investigated virus attachment, entry, replication and the consequent liver cell death. Heparin sulphate has been implicated in dengue virus attachment to liver cells, either as a direct virus receptor[64] or, more probably, as either a low affinity-binding molecule[65] or receptor co-factor.[66] Two cell surface-expressed proteins have been implicated as liver cell-expressed dengue virus receptors, and it has been proposed that these proteins act in a serotype-specific manner.[66,67]
The first protein implicated, GRP78 or BiP, was proposed to act as a receptor for DENV-2[67] while the 37/67kda high-affinity laminin receptor protein has been implicated as a DENV-1 receptor.[66] A critical role for GRP78 as a chaperone protein in the replication of dengue has also been suggested.[68,69] The 37/67kda high-affinity laminin receptor protein has additionally been implicated as a receptor for Sindbis virus,[70] adeno-associated virus,[71] tick-borne encephalitis virus, and Venezuelan equine encephalitis virus,[72] while GRP78 has been implicated as a co-receptor for coxsackie virus A9.[73] The issue of serotype-specific entry of the dengue virus into liver cells is somewhat controversial, as other identified dengue virus receptors such
as DC-SIGN[74]and Hsp79/90[75] in different cell types do not show such a specificity, although in some cases, not all serotypes have been investigated.[75] Dengue virus entry to liver cells has been suggested to occur predominantly, but not solely, by clathrin-coated pit-mediated endocytosis and it has been proposed that as much as 20% of virus entry to liver cells may occur through alternate pathways.[76] While somewhat controversial, support for the concept of multiple entry pathways has also been presented in studies on dengue virus entry into Vero (monkey kidney) cells.[77] One study has suggested that dengue virus entry into HepG2 cells is modulated at least in part by the cell cycle,[78] as has been found with dengue entry into insect cells.[79]
Two studies have investigated global gene expression changes in HepG2 cells by either microarray analysis[80] or cDNA-AFLP.[81] Both studies estimate that some 500 gene transcripts belonging to a number of different pathways are differentially regulated in response to dengue infection. The microarray analysis identified a number of genes involved in the innate immune response and, in particular, pattern recognition genes such as TLR3, TLR8, RIG-1 and MDA5 were identified as being up-regulated.[80] Some genes identified such as IL-6, IL-8, RANTES and IFN-b[80] were consistent with data generated from analysis of dengue-infected primary hepatocytes.[59] Perhaps significantly, the microarray analysis of dengue virus-infected HepG2 cells also detected the up-regulation of caspases 8 and 10 (see next section).
In contrast to the data which suggests that some 500 genes have altered transcriptional regulation,[80,81] proteomic analysis of dengue-infected HepG2 cells have only identified some 17 proteins as being differentially expressed, and these proteins were primarily

80 Dengue Bulletin – Volume 33, 2009
Involvement of the liver in dengue infections
involved in the regulation of transcription and translation.[82] Interestingly, a secretome analysis of dengue-infected HepG2 cells identified significantly more proteins as being differentially regulated by dengue virus infection, with 35 proteins being down-regulated and 24 proteins being up-regulated in infected HepG2 cells.[83] Several proteins, including a-enolase, superoxide dismutase (SOD), peptidyl-prolyl isomerases A and B (cyclophilins A and B), tissue inhibitor of metalloproteinases 1 and 2 (TIMP-1 and 2) and macrophage migration inhibitory factor (MIF) identified in the secreteome of dengue virus-infected HepG2 cells (as opposed to the secretome of uninfected HepG2 cells), have been previously identified in other virus infections or inflammation situations.[83]
Two groups have shown that autophagy, the lysosomal degradation pathway, is activated in response to dengue virus infection of liver cells[84-86] and it has been suggested that in liver cells, autophagic vesicles act as sites for dengue virus replication.[86] Again, as with dengue virus receptor usage,[66,67,87] it has been proposed that interactions between the dengue virus and the autophagy pathway are serotype-specific.[84,86] Given that the endocytosis pathway interacts with the autophagic pathway, a model has been proposed to link dengue virus entry and replication in liver cells in terms of a continuing interaction with membranes of an endosomal-autophagosomal lineage.[88]
Apoptosis
Cellular apoptosis in the liver upon dengue virus infection has been reported both in vivo and in vitro.[35,40,63,89,90] Histological examination of the livers of fatal cases of dengue virus infection note the presence of councilman
bodies which are believed to be the remains of cells undergoing apoptosis.[38] Dengue virus infection of primary cultures of human Kupffer cells and hepatoma cell lines induces apoptosis as evidenced by DNA laddering,[63,89-91] and infection of primary human hepatocytes produces morphological changes characteristic of apoptosis.[59] The mechanism by which the dengue virus induces apoptosis remains to be clearly elucidated. While some authors have proposed that the apoptosis of liver cells occurs by a p53-independent mechanism, based in part on the high level of apoptosis in the p53-null cell line Hep3B,[63] other authors have proposed that apoptosis occurs by a p53-dependent mechanism.[92] In 2005, Matsuda and colleagues[61] presented evidence that Apo2L or TRAIL (tumour necrosis factor-related apoptosis-inducing ligand) is induced by dengue infection and that this interacts with the Apo2L/TRAIL receptor DR5/TRAIL-R2 expressed on the surface of liver cells[61] inducing apoptosis. This would suggest that apoptosis was induced primarily through an extrinsic apoptosis pathway. In contrast Nasirudeen and Liu, who have suggested apoptosis primarily results through an intrinsic, mitochondrially-mediated pathway.[92] However, more recently, Nasirudeen and Liu also suggested that apoptosis in liver cells may be mediated through caspase 1.[93] Other authors have proposed that the critical event in triggering apoptosis in dengue-infected hepatocytes is the interaction between the dengue virus capsid protein and the human death domain-associated protein Daxx,[94] and that nuclear localization of the dengue capsid protein is essential for the interaction with Daxx and subsequent apoptosis.[95] Further research is required to integrate the various suggested models of induction of apoptosis in liver cells as a consequence of dengue virus infection.

Dengue Bulletin – Volume 33, 2009 81
Involvement of the liver in dengue infections
Alternate mechanisms
A number of alternatives to direct infection of liver cells have been suggested to account for the involvement of the liver in dengue disease. These mechanisms include damage induced by the infiltration of activated lymphocytes, especially CD8+ T cells,[51] bystander lysis mediated by CD4+ cytotoxic T cells after activation by dengue-infected Kupffer cells,[96] as well as the action of antibodies against dengue proteins cross-reacting with host cell proteins.[97] Overall, the theories of an alternate mechanism of liver damage in dengue infection are somewhat less well supported than models proposing direct infection of cells. However, there is no evidence ruling out either model, and it is possible that both mechanisms, direct and indirect liver involvement, may occur together in dengue infections.
Conclusions
A large body of clinical and experimental evidence points to the involvement of the liver in the pathobiology of dengue virus infections of humans. The balance of evidence suggests that hepatocytes are directly involved in the infection as sites of dengue replication, possibly adding to the total viral burden. It remains to be seen whether the increasing body of information on the involvement of the liver can be translated into hepato-protective treatments, and whether this would reduce either the severity of the disease or the likelihood of patients developing more severe forms of the disease.
AcknowledgementDRS is supported by the Thailand Research Fund and Mahidol University.
References
PVDI:[1] global burden of dengue. http://www.pdvi.org/about_dengue/GBD.asp – accessed 29 June 2010.
Weaver SC. Host range, amplification and [2] arboviral disease emergence. Arch Virol Suppl 2005:33-44.
Martina BE, Koraka P, Osterhaus AD. Dengue [3] virus pathogenesis: an integrated view. Clin Microbiol Rev 2009;22:564-581.
Srikiatkhachorn A, Green S. Markers of dengue [4] disease severity. Curr Top Microbiol Immunol 2010;338:67-82.
Srichaikul T, Nimmannitya S. Haematology [5] in dengue and dengue haemorrhagic fever. Baillieres Best Pract Res Clin Haematol 2000;13:261-276.
Halstead SB, Nimmannitya S, Margiotta MR. [6] Dengue and chikungunya virus infection in man in Thailand, 1962-1964. II. Observations on disease in outpatients. Am J Trop Med Hyg 1969;18:972-983.
Butt N, Abbassi A, Munir SM, Ahmad SM, [7] Sheikh QH. Haematological and biochemical indicators for the early diagnosis of dengue viral infection. J Coll Physicians Surg Pak 2008;18:282-285.
Faridi MM, Aggarwal A, Kumar M, Sarafrazul [8] A. Clinical and biochemical profile of dengue haemorrhagic fever in children in Delhi. Trop Doct 2008;38:28-30.
Mohan B, Patwari AK, Anand VK. Hepatic [9] dysfunction in childhood dengue infection. J Trop Pediatr 2000;46:40-43.

82 Dengue Bulletin – Volume 33, 2009
Involvement of the liver in dengue infections
Nguyen TH, Lei HY, Nguyen TL, Lin YS, Huang [10] KJ, Le BL, et al. Dengue hemorrhagic fever in infants: a study of clinical and cytokine profiles. J Infect Dis 2004;189:221-232.
Pancharoen C, Mekmullica J, Thisyakorn [11] U. Primary dengue infection: what are the clinical distinctions from secondary infection? Southeast Asian J Trop Med Public Health 2001;32:476-480.
Pancharoen C, Rungsarannont A, Thisyakorn [12] U. Hepatic dysfunction in dengue patients with various severity. J Med Assoc Thai 2002;85 Suppl 1:S298-301.
Shah GS, Islam S, Das BK. Clinical and [13] laboratory profile of dengue infection in children. Kathmandu Univ Med J (KUMJ) 2006;4:40-43.
Wahid SF, Sanusi S, Zawawi MM, Ali RA. A [14] comparison of the pattern of liver involvement in dengue hemorrhagic fever with classic dengue fever. Southeast Asian J Trop Med Public Health 2000;31:259-263.
Ahmed S, Ali N, Ashraf S, Ilyas M, Tariq WU, [15] Chotani RA. Dengue fever outbreak: a clinical management experience. J Coll Physicians Surg Pak 2008;18: 8-12.
Itha S, Kashyap R, Krishnani N, Saraswat VA, [16] Choudhuri G, Aggarwal R. Profile of liver involvement in dengue virus infection. Natl Med J India 2005;18:127-130.
Venkata Sai PM, Dev B, Krishnan R. Role [17] of ultrasound in dengue fever. Br J Radiol 2005;78:416-418.
Chongsrisawat V, Hutagalung Y, Poovorawan [18] Y. Liver function test results and outcomes in children with acute liver failure due to dengue infection. Southeast Asian J Trop Med Public Health 2009;40:47-53.
Faisal T, Taib MN, Ibrahim F. Reexamination [19] of risk criteria in dengue patients using the self-organizing map. Med Biol Eng Comput 2010;48(3):293-301.
Kuo CH, Tai DI, Chang-Chien CS, Lan CK, [20] Chiou SS, Liaw YF. Liver biochemical tests and dengue fever. Am J Trop Med Hyg 1992;47:265-270.
Nguyen TL, Nguyen TH, Tieu NT. The impact [21] of dengue haemorrhagic fever on liver function. Res Virol 1997;148:273-277.
de Souza LJ, Nogueira RM, Soares LC, Soares [22] CE, Ribas BF, Alves FP, et al. The impact of dengue on liver function as evaluated by aminotransferase levels. Braz J Infect Dis 2007;11:407-410.
Gholson CF, Provenza JM, Bacon BR. [23] Hepatologic considerations in patients with parenchymal liver disease undergoing surgery. Am J Gastroenterol 1990;85:487-496.
Kalayanarooj S, Nimmannitya S. Clinical [24] presentations of dengue hemorrhagic fever in infants compared to children. J Med Assoc Thai 2003;86 Suppl 3:S673-680.
Seneviratne SL, Malavige GN, de Silva HJ. [25] Pathogenesis of liver involvement during dengue viral infections. Trans R Soc Trop Med Hyg 2006;100:608-614.
Bhamarapravati N, Tuchinda P, Boonyapaknavik [26] V. Pathology of Thailand haemorrhagic fever: a study of 100 autopsy cases. Ann Trop Med Parasitol 1967;61:500-510.
Burke T. Dengue haemorrhagic fever: a [27] pathological study. Trans R Soc Trop Med Hyg 1968;62:682-692.
Fresh JW, Reyes V, Clarke EJ, Uylangco CV. [28] Philippine hemorrhagic fever: a clinical, laboratory, and necropsy study. J Lab Clin Med 1969;73:451-458.
Marianneau P, Steffan AM, Royer C, Drouet [29] MT, Kirn A, Deubel V. Differing infection patterns of dengue and yellow fever viruses in a human hepatoma cell line. J Infect Dis 1998;178:1270-1278.

Dengue Bulletin – Volume 33, 2009 83
Involvement of the liver in dengue infections
Fabre A, Couvelard A, Degott C, Lagorce-[30] Pages C, Bruneel F, Bouvet E, et al. Dengue virus induced hepatitis with chronic calcific changes. Gut 2001;49:864-865.
Ling LM, Wilder-Smith A, Leo YS. Fulminant [31] hepatitis in dengue haemorrhagic fever. J Clin Virol 2007;38:265-268.
Thein S, Aung MM, Shwe TN, Aye M, [32] Zaw A, Aye K, et al. Risk factors in dengue shock syndrome. Am J Trop Med Hyg 1997;56:566-572.
Balsitis SJ, Coloma J, Castro G, Alava A, Flores D, [33] McKerrow JH, et al. Tropism of dengue virus in mice and humans defined by viral nonstructural protein 3-specific immunostaining. Am J Trop Med Hyg 2009;80:416-424.
Basilio-de-Oliveira CA, Aguiar GR, Baldanza [34] MS, Barth OM, Eyer-Silva WA, Paes MV. Pathologic study of a fatal case of dengue-3 virus infection in Rio de Janeiro, Brazil. Braz J Infect Dis 2005;9:341-347.
Couvelard A, Marianneau P, Bedel C, Drouet [35] MT, Vachon F, Henin D, et al. Report of a fatal case of dengue infection with hepatitis: demonstration of dengue antigens in hepatocytes and liver apoptosis. Hum Pathol 1999;30:1106-1110.
de Macedo FC, Nicol AF, Cooper LD, Yearsley [36] M, Pires AR, Nuovo GJ. Histologic, viral, and molecular correlates of dengue fever infection of the liver using highly sensitive immunohistochemistry. Diagn Mol Pathol 2006;15:223-228.
Hall WC, Crowell TP, Watts DM, Barros VL, [37] Kruger H, Pinheiro F, et al. Demonstration of yellow fever and dengue antigens in formalin-fixed paraffin-embedded human liver by immunohistochemical analysis. Am J Trop Med Hyg 1991;45:408-417.
Huerre MR, Lan NT, Marianneau P, Hue NB, [38] Khun H, Hung NT, et al. Liver histopathology and biological correlates in five cases of fatal dengue fever in Vietnamese children. Virchows Arch 2001;438:107-115.
Jessie K, Fong MY, Devi S, Lam SK, Wong KT. [39] Localization of dengue virus in naturally infected human tissues, by immunohistochemistry and in situ hybridization. J Infect Dis 2004;189:1411-1418.
Limonta D, Capo V, Torres G, Perez AB, [40] Guzman MG. Apoptosis in tissues from fatal dengue shock syndrome. J Clin Virol 2007;40:50-54.
Marianneau P, Steffan AM, Royer C, Drouet [41] MT, Jaeck D, Kirn A, et al. Infection of primary cultures of human Kupffer cells by dengue virus: no viral progeny synthesis, but cytokine production is evident. J Virol 1999;73:5201-5206.
Kangwanpong D, Bhamarapravati N, Lucia [42] HL. Diagnosing dengue virus infection in archived autopsy tissues by means of the in situ PCR method: a case report. Clin Diagn Virol 1995;3:165-172.
Rosen L, Drouet MT, Deubel V. Detection of [43] dengue virus RNA by reverse transcription-polymerase chain reaction in the liver and lymphoid organs but not in the brain in fatal human infection. Am J Trop Med Hyg 1999;61:720-724.
Sariol CA, Pelegrino JL, Martinez A, Arteaga [44] E, Kouri G, Guzman MG. Detection and genetic relationship of dengue virus sequences in seventeen-year-old paraffin-embedded samples from Cuba. Am J Trop Med Hyg 1999;61:994-1000.
Kho LK, Sumarmo, Wulur H, Jahja EC, Gubler [45] DJ. Dengue hemorrhagic fever accompanied by encephalopathy in Jakarta. Southeast Asian J Trop Med Public Health 1981;12:83-86.
Rosen L, Khin MM, U T. Recovery of virus [46] from the liver of children with fatal dengue: reflections on the pathogenesis of the disease and its possible analogy with that of yellow fever. Res Virol 1989;140:351-360.

84 Dengue Bulletin – Volume 33, 2009
Involvement of the liver in dengue infections
An J, Kimura-Kuroda J, Hirabayashi Y, [47] Yasui K. Development of a novel mouse model for dengue virus infection. Virology 1999;263:70-77.
Barreto DF, Takiya CM, Paes MV, Farias-Filho J, [48] Pinhao AT, Alves AM, et al. Histopathological aspects of dengue-2 virus infected mice tissues and complementary virus isolation. J Submicrosc Cytol Pathol 2004;36:121-130.
Barth OM, Barreto DF, Paes MV, Takiya CM, [49] Pinhao AT, Schatzmayr HG. Morphological studies in a model for dengue-2 virus infection in mice. Mem Inst Oswaldo Cruz 2006;101:905-915.
Boonpucknavig S, Vuttiviroj O, Boonpucknavig [50] V. Infection of young adult mice with dengue virus type 2. Trans R Soc Trop Med Hyg 1981;75:647-653.
Chen HC, Lai SY, Sung JM, Lee SH, Lin YC, Wang [51] WK, et al. Lymphocyte activation and hepatic cellular infiltration in immunocompetent mice infected by dengue virus. J Med Virol 2004;73:419-431.
Hotta H, Murakami I, Miyasaki K, Takeda Y, [52] Shirane H, Hotta S. Inoculation of dengue virus into nude mice. J Gen Virol 1981;52:71-76.
Hotta H, Murakami I, Miyasaki K, Takeda Y, [53] Shirane H, Hotta S. Localization of dengue virus in nude mice. Microbiol Immunol 1981;25:89-93.
Lin YL, Liao CL, Chen LK, Yeh CT, Liu CI, Ma [54] SH, et al. Study of dengue virus infection in SCID mice engrafted with human K562 cells. J Virol 1998;72:9729-9737.
Paes MV, Lenzi HL, Nogueira AC, Nuovo GJ, [55] Pinhao AT, Mota EM, et al. Hepatic damage associated with dengue-2 virus replication in liver cells of BALB/c mice. Lab Invest 2009;89:1140-1151.
Paes MV, Pinhao AT, Barreto DF, Costa SM, [56] Oliveira MP, Nogueira AC, et al. Liver injury and viremia in mice infected with dengue-2 virus. Virology 2005;338:236-246.
Blaney JE, Jr., Johnson DH, Manipon GG, [57] Firestone CY, Hanson CT, Murphy BR, et al. Genetic basis of attenuation of dengue virus type 4 small plaque mutants with restricted replication in suckling mice and in SCID mice transplanted with human liver cells. Virology 2002;300:125-139.
Johnson AJ, Roehrig JT. New mouse model [58] for dengue virus vaccine testing. J Virol 1999;73:783-786.
Suksanpaisan L, Cabrera-Hernandez A, Smith [59] DR. Infection of human primary hepatocytes with dengue virus serotype 2. J Med Virol 2007;79:300-307.
Jo M, Kim TH, Seol DW, Esplen JE, Dorko K, [60] Billiar TR, et al. Apoptosis induced in normal human hepatocytes by tumor necrosis factor-related apoptosis-inducing ligand. Nat Med 2000;6:564-567.
Matsuda T, Almasan A, Tomita M, Tamaki [61] K, Saito M, Tadano M, et al. Dengue virus-induced apoptosis in hepatic cells is partly mediated by Apo2 ligand/tumour necrosis factor-related apoptosis-inducing ligand. J Gen Virol 2005;86:1055-1065.
Lin YL, Liu CC, Lei HY, Yeh TM, Lin YS, Chen [62] RM, et al. Infection of five human liver cell lines by dengue-2 virus. J Med Virol 2000;60:425-431.
Thongtan T, Panyim S, Smith DR. Apoptosis in [63] dengue virus infected liver cell lines HepG2 and Hep3B. J Med Virol 2004;72:436-444.
Chen Y, Maguire T, Hileman RE, Fromm JR, [64] Esko JD, Linhardt RJ, et al. Dengue virus infectivity depends on envelope protein binding to target cell heparan sulfate. Nat Med 1997;3:866-871.
Hilgard P, Stockert R. Heparan sulfate [65] proteoglycans initiate dengue virus infection of hepatocytes. Hepatology 2000;32:1069-1077.

Dengue Bulletin – Volume 33, 2009 85
Involvement of the liver in dengue infections
Thepparit C, Smith DR. Serotype-specific entry [66] of dengue virus into liver cells: identification of the 37-kilodalton/67-kilodalton high-affinity laminin receptor as a dengue virus serotype 1 receptor. J Virol 2004;78:12647-12656.
Jindadamrongwech S, Thepparit C, Smith DR. [67] Identification of GRP 78 (BiP) as a liver cell expressed receptor element for dengue virus serotype 2. Arch Virol 2004;149:915-927.
Limjindaporn T, Wongwiwat W, Noisakran [68] S, Srisawat C, Netsawang J, Puttikhunt C, et al. Interaction of dengue virus envelope protein with endoplasmic reticulum-resident chaperones facilitates dengue virus production. Biochem Biophys Res Commun 2009;379:196-200.
Wati S, Soo ML, Zilm P, Li P, Paton AW, [69] Burrell CJ, et al. Dengue virus infection induces upregulation of GRP78, which acts to chaperone viral antigen production. J Virol 2009;83:12871-12880.
Wang KS, Kuhn RJ, Strauss EG, Ou S, Strauss [70] JH. High-affinity laminin receptor is a receptor for Sindbis virus in mammalian cells. J Virol 1992;66:4992-5001.
Akache B, Grimm D, Pandey K, Yant SR, Xu H, [71] Kay MA. The 37/67-kilodalton laminin receptor is a receptor for adeno-associated virus serotypes 8, 2, 3, and 9. J Virol 2006;80:9831-9836.
Ludwig GV, Kondig JP, Smith JF. A putative [72] receptor for Venezuelan equine encephalitis virus from mosquito cells. J Virol 1996;70:5592-5599.
Triantafilou K, Fradelizi D, Wilson K, Triantafilou [73] M. GRP78, a coreceptor for coxsackievirus A9, interacts with major histocompatibility complex class I molecules which mediate virus internalization. J Virol 2002;76:633-643.
Tassaneetrithep B, Burgess TH, Granelli-[74] Piperno A, Trumpfheller C, Finke J, Sun W, et al. DC-SIGN (CD209) mediates dengue virus infection of human dendritic cells. J Exp Med 2003;197:823-829.
Reyes-del Valle J, Chavez-Salinas S, Medina F, [75] Del Angel RM. Heat shock protein 90 and heat shock protein 70 are components of dengue virus receptor complex in human cells. J Virol 2005;79:4557-4567.
Suksanpaisan L, Susantad T, Smith DR. [76] Characterization of dengue virus entry into HepG2 cells. J Biomed Sci 2009;16:17.
Acosta EG, Castilla V, Damonte EB. Alternative [77] infectious entry pathways for dengue virus serotypes into mammalian cells. Cell Microbiol 2009;11:1533-1549.
Phoolcharoen W, Smith DR. Internalization [78] of the dengue virus is cell cycle modulated in HepG2, but not Vero cells. J Med Virol 2004;74:434-441.
Helt AM, Harris E. S-phase-dependent [79] enhancement of dengue virus 2 replication in mosquito cells, but not in human cells. J Virol 2005;79:13218-13230.
C Conceição TM, El-Bacha T, Villas-Bôas CS, [80] Coello G, Ramírez J, Montero-Lomeli M, et al. Gene expression analysis during dengue virus infection in HepG2 cells reveals virus control of innate immune response. J Infect 2010;60(1):65-75.
Ekkapongpisit M, Wannatung T, Susantad [81] T, Triwitayakorn K, Smith DR. cDNA-AFLP analysis of differential gene expression in human hepatoma cells (HepG2) upon dengue virus infection. J Med Virol 2007;79:552-561.
Pattanakitsakul SN, Rungrojcharoenkit K, [82] Kanlaya R, Sinchaikul S, Noisakran S, Chen ST, et al. Proteomic analysis of host responses in HepG2 cells during dengue virus infection. J Proteome Res 2007;6:4592-4600.
Higa LM, Caruso MB, Canellas F, Soares MR, [83] Oliveira-Carvalho AL, Chapeaurouge DA, et al. Secretome of HepG2 cells infected with dengue virus: implications for pathogenesis. Biochim Biophys Acta 2008;1784:1607-1616.

86 Dengue Bulletin – Volume 33, 2009
Involvement of the liver in dengue infections
Khakpoor A, Panyasrivanit M, Wikan N, Smith [84] DR. A role for autophagolysosomes in dengue virus 3 production in HepG2 cells. J Gen Virol 2009;90:1093-1103.
Lee YR, Lei HY, Liu MT, Wang JR, Chen SH, [85] Jiang-Shieh YF, et al. Autophagic machinery activated by dengue virus enhances virus replication. Virology 2008;374:240-248.
Panyasrivanit M, Khakpoor A, Wikan N, [86] Smith DR. Co-localization of constituents of the dengue virus translation and replication machinery with amphisomes. J Gen Virol 2009;90:448-456.
Jindadamrongwech S, Smith DR. Virus [87] overlay protein binding assay (VOPBA) reveals serotype specific heterogeneity of dengue virus binding proteins on HepG2 human liver cells. Intervirology 2004;47:370-373.
Panyasrivanit M, Khakpoor A, Wikan N, Smith [88] DR. Linking dengue virus entry and translation/replication through amphisomes. Autophagy 2009;5:434-435.
Marianneau P, Flamand M, Deubel V, Despres [89] P. Apoptotic cell death in response to dengue virus infection: the pathogenesis of dengue haemorrhagic fever revisited. Clin Diagn Virol 1998;10:113-119.
Marianneau P, Flamand M, Deubel V, Despres [90] P. Induction of programmed cell death (apoptosis) by dengue virus in vitro and in vivo. Acta Cient Venez 1998;49 Suppl 1:13-17.
Marianneau P, Cardona A, Edelman L, Deubel [91] V, Despres P. Dengue virus replication in
human hepatoma cells activates NF-kappaB which in turn induces apoptotic cell death. J Virol 1997;71:3244-3249.
Nasirudeen AM, Wang L, Liu DX. Induction of [92] p53-dependent and mitochondria-mediated cell death pathway by dengue virus infection of human and animal cells. Microbes Infect 2008;10(10-11):1124-32.
Nasirudeen AM, Liu DX. Gene expression [93] profiling by microarray analysis reveals an important role for caspase-1 in dengue virus-induced p53-mediated apoptosis. J Med Virol 2009;81:1069-1081.
Limjindaporn T, Netsawang J, Noisakran S, [94] Thiemmeca S, Wongwiwat W, Sudsaward S, et al. Sensitization to fas-mediated apoptosis by dengue virus capsid protein. Biochem Biophys Res Commun 2007;362:334-339.
Netsawang J, Noisakran S, Puttikhunt C, [95] Kasinrerk W, Wongwiwat W, Malasit P, et al. Nuclear localization of dengue virus capsid protein is required for DAXX interaction and apoptosis. Virus Res 147:275-283.
Gagnon SJ, Ennis FA, Rothman AL. Bystander [96] target cell lysis and cytokine production by dengue virus-specific human CD4(+) cytotoxic T-lymphocyte clones. J Virol 1999;73:3623-3629.
Lin CF, Wan SW, Chen MC, Lin SC, Cheng [97] CC, Chiu SC, et al. Liver injury caused by antibodies against dengue virus nonstructural protein 1 in a murine model. Lab Invest 2008;88:1079-1089.

Dengue Bulletin – Volume 33, 2009 87
Improving dengue virus diagnosis in rural areas of Mexico
Moreno-Altamirano M.M.B.a#, Sánchez-García F.J.a, López-Martínez I.b, Rosales-Jiménez C.b, Vázquez-Pichardo M.b, Arriaga-Valona L.J.a and Capitan-Ortega F.a
aDepartamento de Inmunología, Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional, Prol. de Carpio y Plan de Ayala, Col. Santo. Tomás, México D.F. 11340, Mexico
bInstituto de Diagnóstico y Referencia Epidemiológicos (InDRE), SS, Prol. de Carpio 470, Col. Santo Tomás, México 11340, D.F., México
Abstract
Evaluation of an “in-house system” for the diagnosis of dengue infection by detection of specific IgM and IgG antibodies showed that 25 out of 34 (73.53%) serum samples were positive for IgM antibodies; 6 (17.64%) were positive for IgG and 3 (8.8%) were negative for both IgM and IgG anti-DENV antibodies. Ten samples from “non-symptomatic” people were all negative. In order to evaluate the anti-DENV ELISA, 20 serum samples obtained from healthy individuals from a non-endemic region (Mexico City) and 20 serum samples previously classified as positive were tested. All 20 samples from healthy individuals proved to be negative for both IgM and IgG anti-DENV antibodies, whereas not all positive samples resulted as positive in our assay.
Keywords: In-house diagnosis kit; dengue; surveillance; endemic; Tamiahua; Mexico.
# E-mail: [email protected], [email protected]; Tel.: (5255) 55729-6000 ext. 62370; Fax: (5255) 55396-3503
Introduction
Dengue virus infection in the Americas, as in the rest of the world, is increasing dramatically. Currently, Mexico could be considered as an endemic region for dengue since the mosquito vector, Aedes aegypti, is present in more than 85% of the country.[1] Dengue viruses are members of the Flaviviridae family, genus Flavivirus. Infection can lead to dengue fever (DF), a self-limiting flu-like illness, or to dengue haemorrhagic fever (DHF) with fatal
consequences. Dengue fever is caused by any of the four dengue virus serotypes (DENV-1-4).[2] It is well-documented that primary infection with one of the four serotypes confers long-lasting immunity to that specific serotype. However, secondary infection with a different serotype is associated with an increased risk of developing DHF where an antibody-dependent enhancement (ADE) of infection is associated with the pathophysiological mechanisms of DHF.[3,4] In addition to DF and DHF, some other clinical outcomes, such as

88 Dengue Bulletin – Volume 33, 2009
Alternative diagnosis for dengue in rural villages of Mexico
neurological manifestations and liver damage have also been reported[5-8].
Since all the four serotypes are circulating in Mexico, there is a need to develop an efficient diagnosis system to improve case management of the patients. Until now, the incidence of dengue infection has been underestimated since most cases are not properly diagnosed, especially in small towns or villages where private or state laboratories for diagnosis are lacking. The diagnosis problem exists not only at this level, since several regional hospitals outside Mexico City have inadequate budgets to buy the required number of diagnostic kits to match the demand, making epidemiological surveillance, case management and outbreak investigation difficult. In order to set up a rapid and reliable diagnosis, some reference laboratories in Mexico use the Panbio Dengue Duo IgM and IgG Rapid Cassette test kits or the ELISA Dengue IgM capture kits. However, it is sometimes difficult to have access to these kits due to stock shortage in the market, import difficulties, and of course, high cost.
Dengue infection diagnosis can be achieved by several assays such as RT-PCR,[9] real time-PCR,[10] virus isolation[11] and NS-1 protein detection.[12,13] However, the enzyme-linked immune assay (ELISA) has for a while been, due to its practicality, the routine diagnostic system for dengue infection serological confirmation.[14-17] Different kits are commercially available, such as Dengue IgM capture ELISA (MAC-ELISA) and IgG ELISA, which, in terms of sensitivity and specificity, are reliable with regard to anti-dengue antibodies detection, although they do not clearly distinguish between different Flaviviruses. There are some other commercial assays that use antigens from each dengue virus serotype (DENV-1 to DENV-4); these assays have a high sensitivity and specificity, typically ranging from 90% to 100%.[18]
The distinction between primary and secondary infections is currently assessed by measuring IgM and IgG responses to dengue antigens in paired serum samples taken from a febrile patient in the acute stage of disease and after convalescence.[19] Clearly, the availability of systems for the detection of dengue infections is a public health priority. In this study, we report a reliable “in-house system” for the diagnosis of dengue infection that was field-tested in a small village (Tamiahua) in the state of Veracruz, Mexico.
Materials and methods
Serum samples
A total of 84 serum samples were included in the evaluation of the “in-house system” for the diagnosis of dengue virus infection. Forty-four samples were collected from a dengue endemic area in Tamiahua, which is located in the Gulf of Mexico, in the state of Veracruz between the Pánuco and Tuxpan rivers, at 21°06’ latitude north and 97°46’ longitude (Figure 1); and classified as shown in Table 1. Blood samples were collected from the 44 Tamiahua participants after obtaining parental consent in cases where it was needed. The participants answered a short list of questions that included whether they suffered from higher than 37.5 °C fever, headache, retro-orbital and abdominal pain, vomiting, skin rash, nose or any other type of haemorrhage over the last three months.
Children who neither had any of the above-mentioned manifestations nor had any clinical manifestation of dengue over the previous three months but who had said that someone in their household did have some of the clinical manifestations were also included in the study (Table 1). In addition, 20 dengue virus-negative serum samples

Dengue Bulletin – Volume 33, 2009 89
Alternative diagnosis for dengue in rural villages of Mexico
were obtained from healthy undergraduate students from Mexico City, a city officially reported as a non-endemic area for dengue. Their negative quality was confirmed by the absence of cytopathic effect on C6/36 cell culture. Twenty DENV-2-positive serum samples, previously classified as such by the Instituto de Diagnóstico y Referencia Epidemiológicos (InDRE) on the basis of virus isolation, RT-PCR, NS-1 detection (Bio-Rad kit) and IgM/IgG determination (PanBio), following the manufacturers’ standard protocols,[20] were also included in the “in-house” dengue diagnostic system.
Sample collection and transport from Tamiahua to Mexico City
Five ml of blood were collected by venopuncture in 13×100 vacutainer tubes. After a 60-minute incubation period at room temperature, the tubes were centrifuged in a small portable centrifuge at 2000 rpm for 10 minutes. The serum was transferred to propylene eppendorf vials and stored in a fridge (4 °C). The sera were transported in ice boxes back to the research laboratory in Mexico City (travel time of about seven hours) where samples were stored at –70 °C until testing.
Evaluation of the in-house dengue diagnostic system
To evaluate the in-house-made diagnostic system, a WHO serum panel for dengue, composed of DENV-1, 2, 3, 4 and negative sera (anti-dengue, WHO Reference Panel, Code: 05/248, NIBSC, Potters Bar Hertfordshire, UK), as well as the positive and negative controls included in the Platelia dengue NS1 commercial kit (Bio-Rad) were used, along with the 84 serum samples mentioned in the serum samples section.
Table 1: Characterization of 44 individuals from Tamiahua included in the IgM/IgG anti-dengue virus detection trial by using the in-house ELISA system
Average age
Group 1Elementary school (n=17)
Group IISecondary school (n=18)
Group IIIAdults (n=9)
10 years 14 years 29 years
Gender Female=11 Male=6 Female=10 Male=8 Female=7 Male=2
High fever Female=8 Male=6 Female=8 Male=6 Female=5 Male=1
Aches and pain Female=8 Male=6 Female=8 Male=6 Female=5 Male=1
Nausea and vomiting
Female=3 Male=2 Female=4 Male=2 Female=3
Bleeding Female=2 Male=2 Female=3 Male=1 Female=2
“Healthy” Female=3 Female=2 Male=2 Female=2 Male=1
Figure 1: Location of Tamiahua

90 Dengue Bulletin – Volume 33, 2009
Alternative diagnosis for dengue in rural villages of Mexico
Dengue diagnosis by an in-house system
All serum samples were tested by an indirect ELISA method performed as follows: 96 well plates were coated with 100 µl of carbonate buffer (pH 9.6) containing the antigen (DENV-2, New Guinea strain, propagated in C6/36 cells). After an overnight 4 °C incubation, the plates were washed five times with PBS-T (0.01% Tween-20 in phosphate buffer solution), five minutes per wash, and blocked with 3% skimmed milk in PBS for 45 minutes at room temperature. Serum samples were serially diluted; the dilution which generated an OD value three times higher than that from negative samples was thoroughly used in the rest of the study. All serum samples were diluted 1:50, and 100 µl of each one was added to individual wells by triplicate and incubated for two hours at 37 °C. After washing three times with PBS-T, 100 µL of peroxidase-conjugated protein-A (invitrogen; 1:10 000 dilution) or 100 µL of anti-human IgM-peroxidase (serotec; 1:2000 dilution) was added per well. Plates were incubated for one hour at 37 °C, and after washing three times, 200 µL of ortophenylenediamine (OPD) solution (0.020 g of OPD dissolved in 10 ml of citrate solution, pH 5.0 and 10 µL 30% H2O2) was added to each well. Plates were incubated for 30 minutes (kept away from light); the reaction was stopped by adding 30 µl of 0.2 N H2SO4, and ODs were read in an ELISA plate reader with a 492 nm filter (Labsystems).
Cross-reactivity test for other Flaviviridae family viruses
In order to test the in-house dengue virus ELISA for cross-reactivity with other flaviviruses, four human serum samples positive for West Nile virus (virus isolation and PCR), kindly provided by the InDRE, were tested (it proved
impossible to procure more samples). It is worth noting that yellow fever and Japanese encephalitis are diseases not found in Mexico; so, samples of these two infections were not included in the study due to non-availability of the serum samples.
Data analysis
Mean and confidence intervals (95% CI) from OD ELISA data were calculated using GraphPad Prism version 5.02 (GraphPad software, Inc).
Results
The in-house ELISA system for the detection of IgM and IgG dengue-specific antibodies allowed us to distinguish between the likely primary and secondary dengue infections using serum samples collected in a dengue endemic region in Mexico’s countryside. 25 out of 34 (73.53%) samples obtained from both children and adults that reported having symptoms comparable with a dengue virus infection within the previous three months to the time of sampling presented high OD values for IgM dengue-specific antibodies, whereas six (17.64%) did so for IgG dengue-specific antibodies and three (8.8%) scored negative for both IgM and IgG anti-dengue as shown in Figure 2 and Table 2. Taken together, these results indicate that 91% of the selected (based on referral symptoms) serum donors were positive for anti-dengue antibodies; this correlates with the endemicity of the geographical region. The 10 Tamiahua samples obtained from people that reported no symptoms were negative both for IgM and IgM anti-dengue antibodies (Table 2). The evaluation results of the in-house system showed that it scored 100% for specificity and 77% sensitivity for IgM and 83% for IgG,

Dengue Bulletin – Volume 33, 2009 91
Alternative diagnosis for dengue in rural villages of Mexico
and was able to discriminate well between primary (IgM) and secondary (IgG) dengue infections (Figure 3 and Table 2) in relation to the 20 negative and 20 positive serum samples provided by the InDRE.
There was no cross-reactivity (data not shown) with West Nile virus samples, suggesting a high specificity for our in-house anti-dengue ELISA. However, a higher number of West Nile-positive samples would have to be tested for a more reliable sensitivity and specificity assessment of the dengue virus diagnostic system.
In-house dengue system evaluation
The evaluation of our in-house system with the anti-dengue WHO panel resulted in a clear difference between the negative serum
and the positive for DENV-1, 2, 3, and 4 sera. Also, the OD values for DENV-2 were higher than those for the other serotypes. However, some cross-reactivity was observed between the four serotypes (data not shown). Positive and negative controls included in the Platelia dengue NS1 commercial kit (Bio-Rad) were clearly positive and negative, respectively, when tested by the in-house ELISA system (data not shown).
Discussion
Dengue infection is a growing public health concern in endemic areas all over the world. Hyperendemic geographical areas have been defined as those with continuous presence of multiple viral serotypes and competent vectors, and a large population of susceptible hosts, as seems to be the case for Mexico,[1] so diagnostic systems for dengue are badly needed. Several serological diagnostic kits are commercially available; however, in practice they are not always accessible due to high cost and other constraints. In addition, some technical drawbacks have been reported.[17] As a result there is poor epidemiological surveillance, the number of cases is underestimated, and infected people do not always receive appropriate medical treatment.
Taking all this into account, we are working to improve the development of an in-house ELISA system for the detection of IgM and IgG antibodies specific for dengue virus. After testing it with samples known to be positive (by means of viral isolation, RT-PCR and ELISA) or negative for dengue, further field-testing was carried out in a small village in Veracruz, Mexico. Although paired samples (in the acute stage of the disease and after convalescence) are recommended for the diagnosis of dengue by IgM and IgG ELISAs, in
Figure 2: IgM and IgG anti-dengue virus antibodies in serum samples from Tamiahua, Veracruz, Mexico [Ten samples from healthy people and 34 serum samples from children
and adults who reported having dengue symptoms over the past three months before
sampling were tested for the presence of IgM and IgG antibodies specific for dengue virus using an in-house ELISA system. The
distribution, mean, and confidence intervals for OD values for IgM and IgG are shown. The hatched line represents the cut-off OD values.]
IgM
2.5
2.0
1.5
1.0
0.5
0.0
In-h
ouse
ELIS
AO
Dva
lue
IgG

92 Dengue Bulletin – Volume 33, 2009
Alternative diagnosis for dengue in rural villages of Mexico
the present study this was not possible since donors did not show any clinical symptoms at the time of blood sampling but rather symptoms were reported during a period of time between two and three months before sampling. The threshold for an IgM- or IgG-positive test result was established at an OD value two times (IgM) or three times (IgG) higher than that of the negative controls taken from PanBio IgM and IgG capture ELISA kits. To evaluate the specificity and sensitivity of the in-house ELISA system, 20 dengue-positive samples (laboratory characterized as such two years earlier) along with 20 samples from healthy donors that proved to be negative in C6/36 cytopathic effect assays were assayed.
All in all, the in-house detection system for dengue viruses seemed to perform well in terms of specificity, and seemed suitable
Table 2: Serological diagnosis of dengue infection, based on the in-house ELISA system
Characteristics of the samples tested with the in-house-made
system
Positive results for Negative results for IgM or IgG
Specificity of the
in-house-made assay
Sensitivity of the in-
house-made assay
IgM IgG
Samples from individuals with dengue-like symptoms within three months before sampling (Tamiahua) n=34
25/34 (73.53%)
6/34 (17.64%)
3/34 (8.8%)
Samples from healthy individuals from endemic region (Tamiahua) n=10
0/10 0/10 10/10 10/10 (100%)
Samples from healthy individuals from non-endemic region (Mexico City) n=20
20/20 20/20 20/20 20/20 (100%)
True positive samples for primary dengue infection (laboratory diagnosed in InDRE) n=20
14/20 0/20 6/20 77%
True positive samples for secondary dengue infection (laboratory diagnosed in InDRE) n=20
0/20 16/20 4/20 (83%)
Figure 3: Assessment of IgM and IgG anti-dengue virus antibodies in positive and negative serum samples by using an
in-house ELISA system [20 positive and 20 negative samples characterized as such by
virus isolation, NS-1 detection and RT-PCR were used. Distribution, mean and confidence intervals for OD values for
IgM and IgG are shown. The hatched line represents the cut-off OD values.]
IgMDENV+ DENV+DENV- DENV-
2.5
2.0
1.5
1.0
0.5
0.0
In-h
ouse
ELIS
AO
Dva
lue
IgG

Dengue Bulletin – Volume 33, 2009 93
Alternative diagnosis for dengue in rural villages of Mexico
for in-field diagnosis. We propose the use of the in-house IgM/IgG anti-dengue detection systems as a viable alternative in Mexico when commercial kits are not available, at least in terms of a first screening. Even if this system needs to be improved by increasing the number of samples, for example, especially from asymptomatic individuals from the endemic regions in Mexico, to increase its accuracy in the dengue virus diagnosis, it is important to keep working on the development of reliable diagnostic tools in
order to establish an efficient surveillance system in dengue endemic areas.
Acknowledgements We thank the health authorities of Tamiahua for making the field study possible. We also thank the NIBSC from the United Kingdom for kindly providing the WHO reference panel (through LEI, S.A de C.V). This work was supported by CONACYT grants (103353 and 46767).
ReferencesCENAVECE. [1] Dengue; distribución del vector 2008. http://www.cenave.gob.mx/Dengue
Gubler DJ. The global emergence/resurgence [2] of arboviral disease as public health problems. Arch Med Res 2002;33:330-342.
Halstead SB, O’Rourke EJ. Dengue viruses [3] and mononuclear phagocytes. I. infection enhancement by non-neutralizing antibody. J Exp Med 1977;146:201-217.
Halstead SB. Pathogenesis of dengue: [4] challenges to molecular biology. Science 1988;239:476-481.
Muzaffar J, Venkata KP, Gupta N. Dengue [5] encephalitis: why we need to identify this entity in a dengue-prone region. Singapore Med J 2006;47(11):975-977.
Innis BL, Myint KSA, Nisalak A, Ishak KG, [6] Nimmannitya S, Laohapand T, Tranprasertsuk S, Pongritsakda V, Thisyakom U. Acute liver failure is one important cause of fatal dengue infection. Southeast Asian J Trop Med Public Health 1990;21:695-696.
Paes MV, Lenzi HL, Nogueira AC, Nuovo GJ, [7] Pinhao AT, Mota EM, Basilio-de-Oliveira CA.
Schatzmayr H, Barth OM, Alves AM. Hepatic damage associated with dengue-2 virus replication in liver cells of BALB/c mice. Lab Invest 2009;89(10):1140-51.
Vàzquez-Pichardo M, Rosalez-Jimènez C, [8] Rojas-Espinosa O, Lòpez-Martìnez I, Moreno-Altamirano MMB. Is liver damage dependent on the serotype of dengue virus? – a study in Mexico. Dengue Bull 2006;30:114-120.
Yamada K. Virus isolation as one of the [9] diagnostic methods for dengue virus infection. J Clin Virol 2007;24(3):203.
Velathanthiri VGNS, Fernando S, Fernando [10] R, Neelika Malavige G, Peelawaththage M, Jayaratne SD, Aaskov J. Comparison of serology, virus isolation and RT-PCR in the diagnosis of dengue viral infections in Sri Lanka. Dengue Bull 2006;30:191-196.
Yee-LingL, Youne-Kow Ch, Hwee-Cheng [11] T,Hoon-Fang Y, Grace Y, Eng-Fong O, Lee-Ching N. Cost-effective real-time reverse transcriptase PCR (RT-PCR) to screen for dengue virus followed by rapid single-tube multiplex RT-PCR for serotyping of the virus. J Clin Microb 2007;45(3):935-941.

94 Dengue Bulletin – Volume 33, 2009
Alternative diagnosis for dengue in rural villages of Mexico
Alcon S, TalarminA, Debruyne M, Falconar [12] A, Deubel V, Flamand M. Enzyme-linked immunosorbent assay specific to dengue virus type 1 nonstructural protein NS1 reveals circulation of the antigen in the blood during the acute phase of disease in patients experiencing primary or secondary infections. J Clin Microb 2002;40 (2):376-381.
Falconar AK. Antibody responses are generated [13] to immunodominant ELK/KLE-type motifs on the nonstructural-1 glycoprotein during live dengue virus infections in mice and humans: implications for diagnosis, pathogenesis, and vaccine design. Clin Vaccine Immunol 2007;14(5):493-504.
Vaughn DW, Nisa lak A, Solomon T, [14] Kalayanarooj S, Nguyen MD, Kneen R, Cuzzubbo A, Devine PL. Rapid serologic diagnosis of dengue virus infection using a commercial capture ELISA that distinguishes primary and secondary infections. Am J Trop Med Hyg 1999;60(4):693-698.
Vajpayee M, Singh UB, Seth P, Broor S. [15] Comparative evaluation of various commercial assays for diagnosis of dengue fever. South East Asian J Trop Med Public Health 2001;332(3):472-475.
Thanh TT, Thai KTD, Phuong HL, Giao PT, Hung [16] LQ, Binh T, Mai VT, Nam NV, Vries P. Evaluation of two tapid immunochromatographic assays
for diagnosis of dengue among Vietnamese febrile patients. Clin Vaccine Immunol 2007;14(6):799-801.
Hunsperger EA, Yoksan S, Buchy P, Nguyen VC, [17] Sekaran SD, Enria DA, Pelegrino JL, Vázquez S, Artsob H, Drebot M, Gubler DJ, Halstead SB, Guzmán MG, Margolis HS, Nathanson CM, Rizzo Lic NR, Bessoff KE, Kliks S, Peeling RW. Evaluation of commercially available anti-dengue virus immunoglobulin M tests. Emerg Infect Dis 2009;15(3):436-440.
Groen J, Koraka P, Veltzing J, Copra C, Osterhaus [18] AD. Evaluation of six immunoassays for detection of dengue virus-specificimmunoglobulin M and G antibodies. Clin Diagn Lab Immunol 2000;7:867-871.
Tran TN, de Vries PJ, Hoang LP, Phan GT, Le [19] HQ, Tran BQ, Vo CM, Nguyen NV, Kager PA, Nagelkerke N, Groen J. Enzyme-linked immunoassay for dengue virus IgM and IgG antibodies in serum and filter paper blood. BMC Infect Dis 2006;6:13.
Cuzzubbo AJ, Vaughn DW, Nisalak A, Solomon [20] T, Kalayanarooj S, Aaskov J, Dung NM, Devine PL. Comparison of PanBio Dengue Duo Enzyme-Linked Immunosorbent Assay (ELISA) and MRL Dengue Fever Virus Immunoglobulin M Capture ELISA for Diagnosis of Dengue Virus infections in Southeast Asia. Clin Diagn Lab Immunol 1999;6(5):705-12.

Dengue Bulletin – Volume 33, 2009 95
Dengue vector surveillance and control in Hong Kong in 2008 and 2009K.Y. Cheung and M.Y. Fok
Food and Environmental Hygiene Department, China, Hong Kong Special Administrative Region
Abstract
Dengue vector surveillance programmes were implemented in the community and port areas in Hong Kong Special Administrative Region. Aedes albopictus was the only Aedes vector detected and its distribution was extensive in various areas during the summer months. Aedes aegypti was, however, not detected in any area under surveillance. For community surveillance, the monthly ovitrap indices (MOI) of 2008 and 2009 followed a similar trend as in previous years but were generally lower. The highest ovitrap index recorded in 2008 and 2009 was 37.5% and 48.3% respectively, which were much lower than the highest recorded of 70.9% in 2007. Only two surveyed areas in 2009 had ovitrap indices greater than 20.0% for more than two consecutive months, which was more often seen in 2007. Improvement in the dengue vector situation in the past two years could be attributed to the swift response of the agencies concerned upon getting information on surveillance results via the geographic information system. The public was also informed of the results regularly through the Internet and press releases to create awareness to prevent and control the dengue vector. Health education programmes targeted at different organizations/parties were arranged to promote knowledge about dengue vector prevention and control.
Keywords: Dengue vector surveillance; ovitrap indices; Aedes albopictus; vector control; Hong Kong SAR.
IntroductionDengue fever has statutorily been made notifiable in China, Hong Kong Special Administrative Region (Hong Kong SAR) since 1994.[1] Between 1994 and 2001, the annual number of notifications ranged from 3 to 17 imported cases. In 2002, for the first time, there were 44 confirmed cases recorded, of which 20 were locally infected. There was another local case recorded in 2003 but none since 2004. The number of imported cases
remained at 31 from 2004 to 2006, increased to 58 in 2007, and then dropped to 42 in 2008 and 43 in 2009 (Table 1).[2]
Aedes albopictus, one of the dengue vectors, is commonly found in the Hong Kong Special Administrative Region, and has wide distribution both in urban and rural areas. Ae. aegypti, on the other hand, probably has not been an indigenous species in Hong Kong SAR, though it was once discovered on board a vessel from another city in the mid-1950s.[3,4]

96 Dengue Bulletin – Volume 33, 2009
Dengue vector surveillance and control in Hong Kong in 2008 and 2009
The dengue vector surveillance programme using oviposition traps (ovitraps) in Hong Kong SAR was first introduced in 1998 at the Hong Kong International Airport. Since then, the programme has been gradually extended to cover the community areas and all the port areas. It aims to detect the presence and distribution of the dengue vectors, to monitor and evaluate the effectiveness of dengue vector control work carried out by various agencies, and to make timely adjustments to dengue vector control strategies and measures.
Methods and materials
Ovitraps were used in this surveillance programme as a tool to detect the prevalence and distribution of aedine mosquitoes. The device was locally manufactured as per specifications. It comprised a simple black plastic container with a straight and slightly tapered side of approximately 200 ml capacity. The opening measured 6.5 cm in diameter, the base diameter was 5.0 cm and the container was 10.0 cm in height. The ovitrap was covered by a black cap with four round holes and a grey umbrella-shaped raised cover to protect the contents in the ovitrap from contamination by unwanted materials, such as pesticidal chemicals, litter, cigarette butts, etc. A brownish wooden tongue depressor was placed diagonally inside the container as an oviposition paddle. For surveys in community areas, tap water which had been dechlorinated for one night was used as the breeding medium in the ovitraps. For surveys in port areas, 10% hay infusion solution, which had been prepared by steeping 125 g of dried grass in 15 litres of dechlorinated tap water in a tightly closed plastic container for seven days, was used as the breeding medium in the ovitraps. A new batch of hay infusion was started seven days in advance as followed in Cambodia.[5]
Community areas
Thir ty eight areas with high human concentration, such as housing estates, schools and hospitals, were selected. All the 38 areas were surveyed every month by four teams of paired staff to closely monitor the situation in each area and to obtain a territory-wide picture of the vectorial situation. On average, 55 ovitraps were placed at each selected site. Every month, the ovitraps were set at a distance of at least 100 m from one another
Table 1: Number of imported and indigenous dengue fever cases
from 1994 to 2009[2]
Year Imported cases
Indigenous cases
Total
1994 3 0 3
1995 6 0 6
1996 5 0 5
1997 10 0 10
1998 15 0 15
1999 5 0 5
2000 11 0 11
2001 17 0 17
2002 24 20 44
2003 48 1 49
2004 31 0 31
2005 31 0 31
2006 31 0 31
2007 58 0 58
2008 42 0 42
2009 43 0 43

Dengue Bulletin – Volume 33, 2009 97
Dengue vector surveillance and control in Hong Kong in 2008 and 2009
for one week and then collected and brought back to the laboratory. To serve as a quick reference for taking prompt follow-up action for mosquito control, each of the ovitraps was examined immediately for the presence of mosquito larvae/eggs. The ovitraps were then incubated at room temperature for one week for the eggs, if any, to hatch. All the larvae found in the ovitraps were identified under a compound microscope to species level. The percentage of positive ovitraps retrieved from the field was recorded as the “Ovitrap Index”. “Area Ovitrap Index” (AOI) refers to the ovitrap index of an area whereas “Monthly Ovitrap Index” (MOI) refers to the pooled results of all the ovitraps retrieved from the 38 areas in the same month. MOI reflects the overall vector situation for the month.
For operational purposes, the ovitrap indices were classified into four different categories – Level One: indices less than 5.0%; Level Two: indices between 5.0% and less than 20.0%; Level Three: indices between 20.0% and less than 40.0%; and Level Four: indices at 40% or above. Different actions were taken based on the levels recorded.
Port areas
A total of 33 ports, which were categorized into seven groups according to their nature, were also surveyed using the methodology similar to community surveillance. These seven groups were categorized into Hong Kong International Airport, Railway Cargoes Handling Areas (three ports), Cross Boundary Check Points on Land (six ports), Cross Boundary Ferry Terminals (three ports), Container Terminals (nine ports), Public Cargoes Working Area (eight ports) and Private Cargoes Working Area (three ports). Twenty ovitraps were used at all ports, except for Hong Kong International Airport where 727 ovitraps filled with hay infusion were set.
Results
Community surveillance
The MOIs of 2008 and 2009 followed a similar trend as previous years but were generally lower (Figures 1 and 2). The MOIs in the first quarters were maintained at a rather low level of 0.0% to 0.2%. However, the indices rose sharply in the second quarter from 2.9% in April to the peak of 14.2% in June 2008 and from 4.8% in April to 19.0% in June 2009. The highest MOIs recorded in 2008 and 2009 were lower than the averages of previous years (23.4% in June for 2000–2007; 22.4% in June for 2000–2008). The MOIs decreased gradually from 14.2% in June to 7.3% in October 2008 and from 19.0% in June to 5.7% in October 2009, and then dropped to 0.1% in December 2008 and 0.0% in December 2009.
In 2008, there were three AOIs that exceeded 20.0% in May. The number of AOIs greater than 20.0% increased sharply to seven in June and further to eight in July. There were two locations found to have AOIs greater than 30.0%, one in June and the other in July. Their AOIs were brought down to below 20.0% in the following month in both cases. All AOIs recorded in August and September were lower than 30.0%. The number of AOIs reaching 20.0% also decreased from eight in July to four in August and further to three in September. The indices remained at a lower level in the last quarter. No activity of aedine mosquitoes was detected in most of the survey areas after November.
In 2009 there were two and one AOIs that exceeded 20.0% in April and May respectively. The number abruptly rose to 12 in June, among which eight AOIs exceeded 30.0% and one AOI even exceeded 40.0%. The number of AOIs that exceeded 20.0% dropped down

98 Dengue Bulletin – Volume 33, 2009
Dengue vector surveillance and control in Hong Kong in 2008 and 2009
Figure 1: Comparison of Monthly Ovitrap Index of 2008 with the average of previous years (2000 to 2007)
11.7 9.8 7.31.9 0.1
3.00.8
21.8
1.53.9
19.923.4
0.3
13.616.5 13.9
9.5
0.0 0.1 0.2 2.9
11.8 14.2 13.1
0.0
10.0
20.0
30.0
40.0
50.0
60.0
70.0
80.0
90.0
100.0
1 2 3 4 5 6 7 8 9 10 11 12(month)
Ovi
trap
Inde
x(%
)
Average of Monthly Ovitrap Index from 2000 to 2007Monthly Ovitrap Index for 2008
Figure 2: Comparison of Monthly Ovitrap Index of 2009 with the average of previous years (2000 to 2008)
0.0
10.0
20.0
30.0
40.0
50.0
60.0
70.0
80.0
90.0
100.0
1 2 3 4 5 6 7 8 9 10 11 12(month)
Ovi
trap
Inde
x(%
)
Average of Monthly Ovitrap Index from 2000 to 2008Monthly Ovitrap Index for 2009
0.0
10.0
20.0
30.0
40.0
50.0
60.0
70.0
80.0
90.0
100.0
1 2 3 4 5 6 7 8 9 10 11 12(month)
Ovi
trap
Inde
x(%
)
4.88.2
19.0
12.8 13.78.4 5.7 1.6 0.0
9.312.516.0
12.4
0.3
22.4 19.1
3.61.3
20.6
0.72.7
0.20.20.0

Dengue Bulletin – Volume 33, 2009 99
Dengue vector surveillance and control in Hong Kong in 2008 and 2009
to four in July and six in August, and none of them were higher than 30.0%. No more AOI exceeding 20.0% was recorded thereafter.
The highest ovitrap index recorded in 2008 and 2009 was 37.5% and 48.3% respectively, which were much lower than the highest recorded of 70.9% in 2007. None in 2008 and only two in 2009 of the surveyed areas had ovitrap indices greater than 20.0% for more than two consecutive months, which was seen more often in 2007. In general, areas more often recorded with high ovitrap indices ( 20.0%) during summer months were those with vegetated slopes present; whereas for areas where such landscape was lacking/minimal, the ovitrap indices recorded throughout the whole year were usually less than 20.0%.
Port surveillance
In 2008, the Port Monthly Ovitrap Index (PMOI) ranged from 0.0% in January through March to 1.6% in June. The variation in PMOIs showed a similar trend as in previous years
(Figure 3). The ovitrap indices of all the seven port groups were below 20.0%. The highest index of the port groups (8.3%) was recorded in the Railway Cargoes Handling Areas in July. The ovitrap indices of two other port groups (the Cross Boundary Check-Points on Land and Public Cargoes Working Area) reached the highest in June, and were 7.2% and 5.8% respectively. In 2008, the average PMOI was 0.4%, which was lower than that in 2007 (0.8%).
In 2009, the PMOI ranged from 0.0% in January through March to 1.5% in June. The variation in PMOIs showed a similar trend as in previous years (Figure 4). The ovitrap indices of all port groups were below 20.0%. The highest index of the port groups (11.7%) was again recorded in the Railway Cargoes Handling Areas in June. The ovitrap indices of two other port groups (the Cross Boundary Check-Points on Land and Private Cargoes Working Area) reached the highest in June, and were 7.9% and 8.3% respectively. In 2009, the average PMOI was 0.3%, which was lower than that in 2008 (0.4%).
Figure 3: Comparison of Port Area Ovitrap Index: 2004-2007 and 2008
0 0 0 0.2
0.9
1.6
0.8 0.6 0.20.1
0 00.1 0.1 0.1
0.7
2.5
3.93.3
2.42.9
1.00.4 0.1
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
8.0
9.0
10.0
Jan. Feb. Mar. Apr. May June July Aug. Sep. Oct. Nov. Dec.Month
Ovi
trap
Inde
x(%
)
2008 Port Area Monthly Ovitrap Index
Average figure of Port Area Monthly –Ovitrap Index (2004 2007)

100 Dengue Bulletin – Volume 33, 2009
Dengue vector surveillance and control in Hong Kong in 2008 and 2009
Discussion
The results of the community and port surveillances indicated that Ae. albopictus existed in various areas in Hong Kong SAR and its distribution was wide, particularly in summer. High ovitrap indices were recorded repeatedly in some of the areas covered by the surveillance programme, indicating the presence of persistent breeding grounds that needed continuous attention. Ae. aegypti, the important vector for the transmission of dengue fever and yellow fever, was, however, not detected in all the areas covered by the community and port surveillance programmes.
The dengue vector surve i l l ance programmes served as a tool not only to monitor the local dengue vector distribution but also to provide objective information to take
appropriate actions by the community against dengue vectors. Government departments were able to access detailed information and the results of the surveillances, including locations of positive ovitraps through a geographic information system (GIS) which was accessible by registered users through the government Intranet. They were able to target mosquito control action at venues that fell within the 100 m radius of all positive ovitraps under their purview. The AOIs, MOIs and PMOIs were released to the public through press releases and the Internet to raise their awareness levels about preventing the spread of mosquitoes. Detailed and comprehensive advice on mosquito prevention and control was issued together with the press release. The public were also able to access the information through the Internet. People were advised to pay particular attention to any water accumulation in and near their residences.
Figure 4: Comparison of Port Area Ovitrap Index: 2004-2008 and 2009
0 0 00.1 0.6
1.5
10.6 0.2
0.10 0
0.0 0.1 0.10.6
2.2
3.4
2.8
2.02.3
0.80.3 0.1
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
8.0
9.0
10.0
Jan. Feb. Mar. Apr. May June July Aug. Sep. Oct. Nov. Dec.
Month
2009 Port Area Monthly Ovitrap Index
Average figure of Port Area Monthly –Ovitrap Index (2004 2007)
Ovi
trap
Inde
x(%
)

Dengue Bulletin – Volume 33, 2009 101
Dengue vector surveillance and control in Hong Kong in 2008 and 2009
At lower levels of the ovitrap index (Levels One–Two) control measures mainly relied on source reduction, e.g . proper disposal of disused articles, empty lunch boxes, containers, etc. Potential breeding sites such as saucers underneath plant pots, surface drainage channels, roadside gully traps or keyholes of manhole covers were inspected weekly and any accumulated water was removed promptly. Larvicides were applied whenever immediate elimination of breeding sources was not feasible. For areas with indices at Level Three or above, an inter-departmental anti-mosquito task force was set up at the district level to coordinate efforts in mosquito control. Relevant nongovernmental organizations, including housing estate management companies and schools, were also invited to participate in special control operations. When the ovitrap index reached Level Four, space spraying of insecticides was carried out at the resting places of adult mosquitoes to contain the mosquito problem.
As in previous years, the government organized annual territory-wide anti-mosquito campaigns in 2008 and 2009 to promote community participation and forge close partnerships between government depar tment s and nongovernmenta l organizations to control the breeding and spread of mosquitoes.
On health education, health talks were organized for schoolchildren, managements of estates, construction sites, as well as local
organizations such as area committees at the district level to disseminate the message of mosquito prevention and control. Training was also organized for pest control personnel in the government. Operatives of pest control contractors providing mosquito control services funded by the government were required to receive proper training on general pest control, including mosquito control and dengue fever.
ConclusionsAccording to the results of the dengue vector surveillance in 2008 and 2009 in Hong Kong SAR, Ae. aegypti was not detected and the activity of Ae. albopictus was, in general, under control. The MOIs during this period were all lower than the averages of the past few years. This indicated that the vector problem had been brought under control in 2008 and 2009.
Active participation of the government, local organizations and the public at large was the key to success in controlling the dengue vector. The timely release of the results of dengue vector surveillance to all parties concerned through the GIS and to the public by press releases facilitated prompt remedial actions on vector control. Health education for sustaining public participation in the prevention and control of the vector continued to be one of the key elements in the mosquito prevention programme.

102 Dengue Bulletin – Volume 33, 2009
Dengue vector surveillance and control in Hong Kong in 2008 and 2009
ReferencesChuang VWM, Wong TY, Leung YH, Ma ESK, [1] Law YL, Tsang OTY, Chan KM, Tsang IHL, Que TL, Yung RWH, Liu SH. Review of dengue fever cases in Hong Kong during 1998 to 2. Hong Kong Med J 2008;14:170-7.
Department of Health, The Government of [2] the Hong Kong Special Administrative Region. Centre for Health Protection. http://www.chp.gov.hk/en/notifiable1/10/26/43.html - accessed 29 June 2010.
Food and Environmental Hygiene Department, [3] Hong Kong SAR Government. 2005. Mosquitoes of Hong Kong. Government Logistics Department.
Chau G.W. [4] An illustrated guide to the identification of the mosquitoes of Hong Kong. Hong Kong, Urban Council Publication, 1982.
Polson KA, Curtis C, Seng Chang Moh, Olson [5] JG, Chantha N, Rawlins SC. The use of ovitraps baited with hay infusion as a surveillance tool for Aedes aegypti mosquitoes in Cambodia. Dengue Bull 2002,2

Dengue Bulletin – Volume 33, 2009 103
Comparative life parameters of transgenic and wild strain of Aedes aegypti in the laboratory
H.L. Leea#, H. Jokob, W.A. Naznia and S.S. Vasanc
aMedical Entomology Unit, Infectious Diseases Research Centre, Institute for Medical Research, Jalan Pahang, 50588 Kuala Lumpur, Malaysia
bDepartment of Community Medicine & Public Health, Faculty of Medicine, Hasanuddin University Makassar, South Sulawesi Province, Indonesia
cCEBAR, IPS Building, Level 5, Block B, University of Malaya, Kuala Lumpur, 50603, Malaysia and Malaysia Oxitec S/B, Plaza See Hoy Chan, Suite 1502, Jalan Raja Chulan, Kuala Lumpur, 50200, Malaysia
Abstract
The release of genetically-modified Aedes aegypti males is considered an effective method of suppressing this vector of dengue. One of the prerequisites is the capability to mass produce the genetically-modified mosquito in the laboratory before release. To do so would necessitate the understanding of the lifecycle and parameters of the transformed strain. In this regard, comparative life-history parameters of a wild-type laboratory strain of Ae. aegypti (WT) and a genetically transformed Ae. aegypti (LA513) were studied in the laboratory. The following parameters were statistically indistinguishable in both strains: the number of eggs laid, the number of unhatched eggs, the egg-hatching rate, the duration of larval period in all four instars, larval survivorship, pupation, adult eclosion rate, gonotrophic cycle, adult fecundity, adult lifespan and offspring sex ratio. The basic life-history and growth rate of the genetically-modified Ae. aegypti was not affected by genetic manipulation and that its mating competitiveness will remain intact, which is important for the successful use of this technology.
Keywords: Transgenic Aedes aegypti; RIDL Ae. aegypti; bionomics; life parameters.
IntroductionAedes aegypti is the main vector of arboviral diseases such as dengue fever and yellow fever. Dengue, which is mainly transmitted by Ae. aegypti, has been recognized in over 100 countries. There are more than 2.5 billion people living in areas where dengue is endemic, and 50–100 million cases of dengue occur
every year. About 250 000–500 000 cases of dengue are officially notified annually.[1] Yellow fever, another disease which is transmitted by Ae. aegypti, is still a problem in Africa and South America, even though a vaccine for yellow fever was found many years ago[2].
Currently, neither a vaccine nor antiviral drugs are available against dengue. Vector
# E-mail: [email protected]

104 Dengue Bulletin – Volume 33, 2009
Comparative life parameters of transgenic and wild strain of Ae. aegypti in the laboratory
control still remains the most important approach to reduce the occurrence of dengue fever (DF) and dengue haemorrhagic fever (DHF).[2] The main strategy directed at controlling the vector of dengue is the use of chemical insecticides. However, this method has not been effective so far in curbing the disease. Therefore, other alternative control methods need to be looked into.
Genetic vector control offers possibilities for controlling vector-borne diseases such as dengue and malaria. Sterile Insect Technique (SIT) is the most prominent form of population suppression.[3] The deleterious effect of radiation imposes a fitness penalty, rendering the sterile male mosquitoes less competitive to wild male mosquitoes and, therefore, a less effective agent of SIT.[4]
Release of Insect Carrying Dominant Lethality (RIDL) is one other alternative method for population suppression. RIDL has so far been exemplified in Ae. aegypti and other mosquito species.[5,6,7] RIDL could be used to avoid the need for irradiation. RIDL is based on the development of transgenic insect with a genetic construct that confers repressible dominant lethality.[8] In its simplest form, RIDL operates in a non-sex-specific manner, killing all insects carrying at least one copy of the construct unless these insects are provided with a specific repressor of the lethality. The principle is simple: if the released insects are homozygous for a dominant lethal and mate with wild insects, all the progeny are heterozygous for a dominant lethal and so they die.[9]
Assessing the fitness of genetically modified mosquito (GMM) will be a critical component of programmes for the genetic control of disease vectors and prevention of vector-borne diseases. It is assumed that in most cases genetic modification will incur fitness costs. This could undermine a population-reduction
strategy by rendering the released insect non-competitive for wild-type mates.
A RIDL strain of Ae. aegypti has now been established in our insectarium. Prior to undertaking further experiments using this strain, its bionomics needs to be studied in detail in order to ascertain its fitness in reproductive potential and suitability for field-release experiments in the future.
Materials and methods
Mosquito strains
The mosquito strain used in this study was the Malaysian wild strain Ae. aegypti which has been maintained in the insectarium and has not been revitalized with field-collected materials. The transgenic Ae. aegypti strain originated from Oxford LA513 strain and has been maintained in an ACL-2 (Arthropod Containment Level 2) laboratory since January 2007 as F1. It was originally generated in a Rockefeller strain background. Rockefeller is a laboratory strain, originally of Caribbean origin, colonized in the early 1930s.
Rearing conditions
All experiment containers were maintained at a mean temperature of 25±1 °C with a 12:12 hr (light/dark) photoperiod and mean relative humidity of 75%±10% in a secured insect-rearing room in the ACL-2 laboratory in the insectarium. The temperature and relative humidity were recorded daily.
Establishment of experimental cohorts
Thirty pupae of each experimental strain (IMR strain and LA513 transgenic) were placed individually into a glass tube containing 10 ml

Dengue Bulletin – Volume 33, 2009 105
Comparative life parameters of transgenic and wild strain of Ae. aegypti in the laboratory
of tap water seasoned overnight and 5 mg of diet (Tetramin®). The glass vial was plugged with cotton wool. Adult mosquitoes that emerged from these pupae were designated as F0, and male and female pairs from each experiment strain that emerged were set up in small cages (15x15x15 cm) for mating. There were fifteen pairs of F0. Five days after emergence, the female mosquitoes were blood-fed through the mesh using a white mouse. The mouse was removed after 24 hours or until the mosquitoes had fully engorged.
Two days after blood feeding, oviposition cups of moistened filter paper were introduced into each cage. Every 24 hours the oviposition cups were removed and replaced with a new moistened filter paper. Filter paper containing mosquito eggs was labelled with a female identification number that indicated the mosquito’s strain and date of oviposition. The number of eggs laid was counted under a dissecting microscope using a counter and recorded daily for each pair.
The eggs were allowed to embryonate by air-drying. Individual egg paper was submerged into 150 ml of dechlorinated tap water. The transgenic mosquito eggs were submerged in 50 µg/ml concentration of tetracycline in 500 ml distilled water. Tetracycline is required to repress the lethal gene so that the transgenic larvae are able to develop to full term. There were 15 individual trays for each strain and every tray was covered with the mesh. After 24 hours, 150 1st instar larvae designated as F1 generation (15 larvae from each of 10 females) were placed individually into small tubes with 10 ml overnight tap water and 5 mg diet (Tetramin®).
Preimaginal development times
Individual F1 larvae that emerged from the eggs were placed in 15 ml glass vials
and monitored every 24 hours. Glass vials containing larvae were labelled with a female identification number that indicated the larvae strain. The number of days spent in each preimaginal life and stage (i.e. 1st, 2nd, 3rd and 4th) and pupae and numbers surviving in each stage were observed and recorded daily. Each stage of larvae was determined by floating moulted skin on the water surface. The mean number of days in each preimaginal stage was determined and compared between two mosquito strains with t-independent test. The first 40 female and male mosquitoes to emerge for each strain were paired and placed in cages. To reduce possible inbreeding effects, adults within experimental strains were paired only if they originated from different F0 females.
Adult longevity, female fecundity and offspring sex ratio
The F1 adult survivorship rates were determined every 24 hours. Female mosquitoes were blood-fed for five days post-emergence. Daily fecundity and the percentage of eggs hatched after 48 hours and one week were recorded for each F1 female that produced eggs from each blood-feeding for four consecutive gonotrophic cycles. One week after each oviposition cycle, the blood meal was offered to the surviving females in each experimental strain and the fecundity rates were recorded.
The percentage of females either failing to lay eggs or laying eggs which failed to hatch in each gonotrophic cycle was considered sterile. The sterility rates were calculated and compared across experimental strains using chi-square test in SPSS (version 14). The mean lifetime fecundity (viable eggs across all gonotrophic cycles for each female in each strain) was calculated and compared against

106 Dengue Bulletin – Volume 33, 2009
Comparative life parameters of transgenic and wild strain of Ae. aegypti in the laboratory
the mosquito strains with t-independent test in SPSS.
The preimaginal survivorship rate and the sex ratio of offspring were recorded for each of the four consecutive gonotrophic cycles for each reproductive female in each experimental strain. Preimaginal survivorship rates for each experimental strain were used to construct partial life tables. The sex of the adults resulting from each gonotrophic cycle for each experimental strain was recorded.
Results
Numbers of eggs laid by F0 transgenic and wild strain of Ae. aegypti
All 15 pairs of transgenic and wild strain Ae. aegypti oviposited. The mean number of eggs laid by F0 transgenic was 74.86±10.95 per female while the number of eggs laid by F0 wild strain was 63.07±6.86 per female. Statistical analysis showed that there was no significant difference between the number of eggs laid by F0 generation for transgenic strain and wild strain Ae. aegypti (P>0.05). All the 15 pairs of female transgenic strain ceased oviposition on Day 6, whereas the Malaysian wild strain completely ceased egg-laying on the 7th day after the blood meal.
Comparison of F0 egg sterility of transgenic and wild strain Ae. aegypti
The number of sterile eggs in transgenic strain Ae. aegypti was less than wild strain Ae. aegypti. The mean number of sterile eggs in the transgenic strain was 25.00±3.35 per female while in the wild strain it was 31.47±5.57. There was no significant difference between the number of sterile eggs for F0 the transgenic strain and the Malaysian wild strain (P>0.005).
Hatching of F1 larvae of transgenic strain and wild strain Ae. aegypti
All the eggs from the 15 pairs of transgenic mosquitoes hatched, while for the wild strain mosquito only 14 pairs hatched. The mean number of larvae hatched for F1 transgenic was 69.07±8.31, while for F1 wild strain it was 23.79±5.23. There was a significant difference between the numbers of larvae hatched for both strains of Ae. aegypti (P<0.05).
Duration of each life-stage for transgenic F1 generation and wild strain Ae. aegypti
There was no significant difference observed in the developmental time periods of first, second and third instar stages. The number of days spent in L1, L2 and L3 for both transgenic and wild strains was one day in each stage.
For the L4 stage there was no significant difference between the number of days spent in this stage for both strains (Figure 1). The mean number of days for the transgenic strain in the L4 stage was 2.59±0.06 days, while for the wild strain it was 2.57±0.058 days. There was no significant difference in the number of days spent in the L4 stage for both strains (P>0.05).
There was also no significant difference between the number of days spent in the pupae stage for both strains (P>0.05) (Figure 1). The mean number of days for the transgenic strain in the pupae stage was 2.29±0.042 days, while for the wild strain this was 2.36±0.40 days.
The total number of days spent by L1 larvae to become adults was equal for both the strains. The mean number of days spent by the transgenic strain to become adults from L1 larvae was 7.85±0.071, while for the wild strain this was 7.94±0.055 days.

Dengue Bulletin – Volume 33, 2009 107
Comparative life parameters of transgenic and wild strain of Ae. aegypti in the laboratory
Survivorship of F1 generation in each life-stage
The number of larvae surviving in L1, L2, and L3 was equal for both strains. There was no association between the strain and survivorship in L1, L2 and L3 stages (Figure 2). There was no difference between the number of larvae surviving in these stages for both strains (P>0.05).
The number of larvae surviving in the L4 and pupae stages for the transgenic and wild strains was equal. There was no association between the strain and the number of larvae surviving in the L4 and pupae stages (P>0.05).
The number for adult emergence showed a significant difference for both strains (Chi square test, P<0.005). The risk estimate (OR. 3.12) showed that wild strain Ae. aegypti was three times at higher risk to become an adult compared with the transgenic strain.
Partial life tables and preimaginal development
For each gonotrophic cycle, the percentage of eggs to adults was the greatest in the transgenic strain over the Malaysian wild strain for gonotrophic cycle 1 and 2. But in gonotrophic cycle 3, the percentage in the Malaysian wild strain was higher than that in
Figure 1: Number of days spent in each life-stage for transgenic LA513 strain and Malaysian wild strain of Ae. aegypti

108 Dengue Bulletin – Volume 33, 2009
Comparative life parameters of transgenic and wild strain of Ae. aegypti in the laboratory
the transgenic strain. There was a reduction in the percentage of eggs-to-adults for each gonotrophic cycle in the transgenic strain for three gonotrophic cycles and two gonotrophic cycles in the Malaysian wild-type strain.
Across two gonotrophic cycles, the Malaysian wild strain females produced 5%–25% more eggs than the transgenic mosquitoes but the number of adult offspring was higher in the transgenic type for two gonotrophic cycles (gonotrophic cycle 1 and 2) compared with the Malaysian wild type. The percentage of adult offspring in gonotrophic cycle 3 was higher in the wild type than the transgenic type (Table 1).
There was a significant difference in the three gonotrophic cycles of the transgenic strain in the number of larvae hatched, number of pupae, number of adults emerged, number of adult males and females and the number of pupae died prior to emergence as
adult (Kruskal Wallis test, P<0.05). An analysis using the Mann Whitney test showed the difference found between gonotrophic cycle 1 and 2, where all these parameters were higher in the first gonotrophic cycle compared to the second cycle (P<0.05), and gonotrophic cycle 1 and 3, where all these parameters were higher in the first gonotrophic cycle compared to the third cycle (P<0.05), and that there was no significant difference in gonotrophic cycles 2 and 3 (P>0.05) (Table 1).
The result was different for wild strain mosquitoes where almost no differences in all parameters were observed (Kruskall Wallis test, P>0.05), except for the number of sterile eggs. An analysis indicated differences in the number of sterile eggs found between gonotrophic cycle 1 and 3 (P<0.05), and gonotrophic cycle 2 and 3 (P<0.05), and none in gonotrophic cycle 1 and 3 (P>0.05). The number of sterile eggs was found higher
Figure 2: Percentage of survivorship in each stage of life

Dengue Bulletin – Volume 33, 2009 109
Comparative life parameters of transgenic and wild strain of Ae. aegypti in the laboratory
in gonotrophic cycles 1 and 2 as compared with gonotrophic cycle 3.
A comparison between transgenic and wild type mosquitoes in gonotrophic cycle 1 showed that there was a significant difference (Mann Whitney test, P<0.05) between the number of larvae hatched, number of pupae, number of adults emerged, the number of dead larvae, and the number of dead pupae. All these parameters were found higher in the transgenic mosquitoes as compared with the wild type mosquitoes. The number of sterile eggs was significantly different (P<0.05) and was found higher in the wild type than in transgenic mosquitoes. Gonotrophic cycle 2 and gonotrophic cycle 3 showed that there was no difference in all parameters for both strains (Mann Whitney test, P>0.05) (Table 1).
Pre-oviposition periods, lifetime fecundity, offspring sex ratio and sterility
In F1 females, significant differences in the pre-oviposition can only be detected in the first
gonotrophic cycle where the pre-oviposition time for the transgenic strain was much longer than the wild strain (Mann Whitney test, P<0.05). The other two gonotrophic cycles showed that there was no significant difference in the oviposition time (Mann Whitney test, P>0.05) (Table 2).
Adult fecundity showed that there was no significant difference between the numbers of eggs laid by transgenic and wild type Ae. aegypti for all gonotrophic cycles (Mann Whitney test, P>0.05).
The percentage of sterile females for each strain increased concomitantly with the gonotrophic cycle wherein the percentage of sterile females was the highest in the third gonotrophic cycle (Table 2). There was no significant association between the mosquito strains and sterile females in each gonotrophic cycle (Chi square, P>0.05). The transgenic strain produced more viable eggs in gonotrophic cycle 1 while wild strain produced more viable eggs in gonotrophic cycle 3 (Table 1).
Table 1: Stage-specific survivorship of transgenic LA513 strain and Malaysian wild strain Ae. aegypti
Gonotrophic cycle Life-stage
Number entering stage Number dying in stage Proportion dying in stage
Transgenic Wild Transgenic Wild Transgenic Wild
1
LarvaePupaeAdult
% survivorship
103696281156%
33330427418%
74151
2930
0.070.16
0.090.10
2
LarvaePupaeAdult
% survivorship
37333231130%
18516314810%
4121
2215
0.110.06
0.120.09
3
LarvaePupaeAdult
% survivorship
27325418824%
75454849647%
1966
20652
0.070.26
0.270.09

110 Dengue Bulletin – Volume 33, 2009
Comparative life parameters of transgenic and wild strain of Ae. aegypti in the laboratory
The sex ratio for adult progeny produced by each strain across all gonotrophic cycles was similar. The percentage of the females in the transgenic strain was higher than males, while the percentage of males in the wild strain was higher than females (Table 2).
Adult longevity
The adult longevity of the Malaysian wild strain was longer than the transgenic strain. For the 40 days measured, the percentage of male mortality for the transgenic strain (85%) was higher than the Malaysian wild strain (60%). The mean adult longevity for the Malaysian wild male was 25.67±6.53 days while that for transgenic strain it was 20±10.60 days. A similar result was obtained in females whereby the mortality of female transgenic strain (15%) was higher than that of the wild strain (5%). The mean adult longevity for females was 11.33±3.05 days and 8.00±0.00 days for the transgenic and wild strains, respectively.
The life parameters of both strains are summarized in Table 3.
Discussion
This study investigated the bionomic and life-history parameters of a transgenic mosquito strain carrying a repressible lethal gene, relative to a Malaysian wild-type strain. An earlier study on Anopheles stephensi measured the whole-life-cycle fitness of several transgenic strains by monitoring the change in allele frequency over time in mixed populations of transgenic and wild strain mosquitoes in the laboratory.[7] A similar study comparing various bionomic and life-history parameters of four transgenic and one wild type strain of Ae. aegypti was also reported.[10] These studies were focused on transgenic mosquitoes designed to be released to replace the natural Ae. aegypti population and thereby to establish stable and persistent populations of transgenic mosquitoes in the wild. In contrast, RIDL strains are intended to be used to suppress the target population in a method analogous to the Sterile Insect Technique (SIT). Therefore, the transgene is not required or expected to persist in the wild, and different aspects of fitness and performance become significant. Parameters such as female fecundity, egg hatch, larval development, etc.,
Table 2: Reproductive statistics for LA513 strain and Malaysian wild strain Ae. aegypti
Cycle Parameter Transgenic LA513 strain Malaysian wild strain
1 PreovipositionFecunditySex ratio% sterile females
1.78±0.1776.89±11.08
55.24%10.5%
1.32±0.1177.25±9.00
51.33%10%
2 PreovipositionFecunditySex ratio% sterile females
1.47±0.1958.44±11.14
52.09% 16.70%
1.76±0.21974.55±11.19
43.24%35%
3 PreovipositionFecunditySex ratio% sterile females
1.50±0.2246.65±12.08
59.57%41.20%
1.71±0.2254.95±11.09
48.39%36.80%

Dengue Bulletin – Volume 33, 2009 111
Comparative life parameters of transgenic and wild strain of Ae. aegypti in the laboratory
relate primarily to the ease and efficiency of mass production, whereas adult male performance parameters, such as mating competitiveness, relate to field performance.
The fitness of the transgenic 513A strain and wild strain of Ae. aegypti used in this study was not significantly different in several parameters. There was no significant difference in the number of eggs laid and the number of pupae in F1 generations, nor in the number of days in each stage of life (developmental period). The number of larvae, pupae surviving and adults emerging in each stage of life for both strains were not significantly different.
The results we obtained were different from the previous study.[10] The transgenic strains they had investigated showed significant reduction in many fitness parameters relative to the wild type strain. In essence, the transgenic strains were severely compromised and were uncompetitive with the wild type. In contrast, in our study, most parameters measured showed no significant difference between
the wild type and transgenic strains; others (e.g. egg production or hatch rate in different gonotrophic cycles) showed differences but not so much as to imply a major difference in fitness levels between the two strains.
There are several potential reasons for this apparent discrepancy. Firstly, the RIDL-513A strain was pre-selected by Oxitec using some basic experimental determinants of fitness, although detailed bionomic measurements had not been performed. Secondly, and more significantly, the data of Irvin et al.[10] merely show that the transgenic lines can have reduced fitness, but not that this is inevitable. That insertional mutants can be deleterious is obvious, but the question is whether strong negative effects are unavoidable. Marelli et al.[11] discussed this question, taking into account also the larger literature from Drosophila on insertional mutations, and concluded that a significant proportion of insertional transgenics should have only ‘modest’ (but probably non-zero) fitness penalties relative to the wild type.
Table 3: Summary of bionomics study on transgenic and wild strain Ae. aegypti
Parameter Strain Mean Mean rank P-value
Number of eggs laid 513AWild type
74. 861±16.9463.07 ± 6.855
16.0314.97
0.740
Number of larvae hatched 513AWild type
69.07 ± 8.30623.79 ± 5.237
17.7013.30
0.001
Number of sterile eggs 513AWild type
25.00 ± 3.34931.47 ± 5.57
14.3316.67
0.468
Number of days spent in L4 513AWild type
2.59 ± 0.062.57 ± 0.58
142.84141.20
0.851
Number of days spent in pupae stage
513AWild type
2.29 ± 0.0422.36 ± 0.40
133.52144.99
0.141
Number of days spent from L1 to adult
513AWild type
7.85±0.0717.49±0.055
130.19141.28
0.200

112 Dengue Bulletin – Volume 33, 2009
Comparative life parameters of transgenic and wild strain of Ae. aegypti in the laboratory
A further issue confounding some previous studies of fitness relates to inbreeding. Catteruccia et al.[7] found significant fitness defects in their transgenic An. stephensi, but concluded that much or all of this was due to inbreeding. Moreira et al.[12] avoided inbreeding effects by using out-crossed heterozygotes, and found that the fitness penalty differed depending on the expressed gene-zero for SM1 but significant for PLA2. Prior to transfer to the IMR, considerable efforts were made by Oxitec to minimize inbreeding effects during the construction of the RIDL-513A strain. Similarly, after transfer to IMR, the strain was kept with a minimum population size of 200 individuals, and egg storage used to minimize the number of effective generations – both measures used to minimize genetic drift and bottlenecking.
It is hardly surprising that some genes might be deleterious when ectopically expressed, especially when they are supposed to cause the death of that organism. However, in the specific case of 513A, the gene is supposed to be lethal when de-repressed. Therefore, any significant uncontrolled leakiness in the control of gene expression might reasonably be expected to lead to mortality or loss of fitness. Therefore, there would need to be tight regulation of the gene. That significant reductions in fitness were not observed validates the proposal of Gong et al.[13] that the “positive feedback” design incorporated into 513A[14] may give superior control over gene expression and minimize adverse fitness effects.
The transgenic strain 513A was initially constructed using transposon-mediated transformation with a non-automonous piggyBac-based transformation vector.[14] piggyBac has several advantages for this
purpose. One is that its insertion and excision is unusually precise for this class of elements, with no evidence of imprecise insertions and excisions during transformation. This effect, well known for P elements in Drosophila, can lead to the creation of additional deleterious mutations in the course of transformation or remobilization of P elements; the advantage of piggyBac in this respect has been noted previously.[15] A second advantage of piggyBac, though not specifically related to fitness, is that piggyBac insertions in Ae. aegypti are extremely stable, even in the presence of exogenous piggyBac transposase.[16] The single report of an apparently unstable piggyBac insertion in Aedes represented an aberrant insertion event with multiple copies of the entire plasmid in an array.[17] Such an array is susceptible to recombination, without any need to invoke a transposase-mediated mechanism.
In summary, we found that for multiple parameters (pre-oviposition period, lifetime fecundity, offspring sex ratio and female sterility) the transgenic 513A strain was not significantly different from the wild strain. This contrasts with previous studies by Irvin et al.[10] and Cateruccia et al.[7] These previous studies reported that transgenic-modified mosquito, especially Ae. aegypti and An. stephensi, was not competitive for all measured parameters compared to the wild strain of Ae. aegypti and An. stephensi. Instead, we share the conclusion of Moreira et al.[12] and Marelli et al.[11] that transgenic lines can be constructed which have little or no fitness penalty relative to wild type; RIDL-513A seems to be an example of such a line.
Finally, it should be noted that all these experiments were conducted under permissive conditions. RIDL-513A larvae reared in

Dengue Bulletin – Volume 33, 2009 113
Comparative life parameters of transgenic and wild strain of Ae. aegypti in the laboratory
restrictive conditions die as late larvae or pupae; therefore, the strain has a fitness of approximately zero under restrictive conditions. This is done by design so that the offspring wild type mosquitoes that mate with RIDL will not survive.
Acknowledgements
The authors thank the Director-General of Health, Malaysia, for permission to publish these findings. This study was partially supported by the SEAMEO-TROPMED Programme.
References
World Health Organization. [1] The world health report 2007: a safer future: global public health security in the 21st century. Geneva: WHO, 2007. http://whqlibdoc.who.int/publications/2007/9789241563444_eng.pdf – accessed 30 June 2010.
Adelman ZN, Jasinskie N, James AA. [2] Development and application of transgenesis in the yellow fever mosquito, Aedes aegypti. Mol & Biochem Parasitol 2002;121:1-10.
Coleman PG, Alphey L. Genetic control of [3] vector populations: an imminent prospect. Tropical Med & Int Hlth 2004;9:433-7.
Benedict MQ, Robinson AS. The first release [4] of transgenic mosquitoes: an argument for the sterile insect technique. Trends in Parasitol 2003;19(8):349-53.
Thomas DD, Donelly CA, Wood RJ, Alphey L. [5] Insect population control using a dominant, repressible, lethal genetic system. Science 2000;287:2474-76.
Asman SM., Nelson RL, MacDonald PT. Pilot [6] release of a sex linked multiple translocations into a Culex tarsalis field population in Kern County, California. Mosq Systematics 1979;39:248-58.
Cateruccia F, Godfray HCJ, Crisanti A. Impact [7] of genetic manipulation on the fitness of Anopheles stephensi mosquitoes. Science 2003;299:1225-27.
Alphey L. Re-engineering the steri le [8] insect technique. Insect Biochem Mol Biol 2000;32:1243-1247.
Alphey L, Andreasen M. Dominant lethality [9] and insect population control. Mol Biochem Parasitol 2002;121:173-8.
Irvin N, Hoddle MS, O’Brochta DA, Carey [10] B, Atkinson PW. Assessing fitness cost for transgenic Ae. aegypti expressing the GFP marker and transposase genes. PNAS 2003;101(3):891-6.
Marrelli MT, Moreira CK, Kelly D, Alphey [11] L, Jacobs-Lorena M. Mosquito transgenesis: what is the fitness cost? Trends in Parasitol 2006;22:197-202.
Morreira LA, Jacobs-Lorena M. Transgenic [12] mosquitoes for malaria control: progress and cha l l enge s . Neo t rop En tomo l 2003;32(4):513-36.
Gong P, Epton MJ, Fu G, Scaife S, Hiscox A, [13] Condon KC, Condon GC, Morrison NI, Kelly DW, Dafa’alla T, Coleman PG, Alphey L. A dominant lethal genetic system for autocidal control of the Mediterranean fruit fly. Nature Biotech 2005;23:453-6.
Phuc HK, Andreasen A, Burton RS, Vass C, [14] Epton JE, Alphey L. Late acting dominant lethal genetic system and mosquito control. Biomed Central Biology 2007;5:1-11.

114 Dengue Bulletin – Volume 33, 2009
Comparative life parameters of transgenic and wild strain of Ae. aegypti in the laboratory
Thibault ST, Singer MA, Miyazaki WY, Milash B, [15] Dompe NA, Singh CM, et al. A complementary transposon tool kit for Drosophila melanogaster using P and piggyback. Nature Genetics 2004;36:283-7.
Sethuraman N, Fraser MJ Jr, Eggleston P, [16] O’Brochta DA. Post-integration stability of piggyBac in Aedes aegypti. Insect Biochem Mol Biol 2007;37:941-51.
Adelman ZN, Jasinskiene N, Vally KJ, Peek C, [17] Travanty EA, Olson KE, Brown SE, Stephens JL, Knudson DL, Coates CJ, James AA. Formation and loss of large, unstable tandem arrays of the piggyBac transposable element in the yellow fever mosquito Aedes aegypti. Transgenic Res 2004;13:411-25.

Dengue Bulletin – Volume 33, 2009 115
Protein profiles of dengue-infected Aedes aegypti (L)H.L. Lee#, Y.C. Wong and A. Rohani
Medical Entomology Unit/Infectious Disease Research Centre and WHO Collaborating Center for Vectors, Institute for Medical Research,
Jalan Pahang, 50588 Kuala Lumpur, Malaysia
Abstract
The protein profiles of DENV-2 and DENV-4 viruses-infected adult Aedes aegypti were analysed using Coomassie-stained SDS-PAGE samples. For this study, adult female mosquitoes were infected orally with either DENV-2 or DENV-4 virus using spiked human blood samples. The infections in the individual mosquitoes were confirmed using the reverse transcription-polymerase chain reaction (RT-PCR). From the SDS-PAGE analyses, a total of 11 proteins with molecular weights of 22 kDa to greater than 181.8 kDa were identified in DENV-2 and DENV-4-infected mosquito homogenates. Three proteins (Mr 49, 64 and 200 kDa) from DENV-2-infected mosquitoes and five proteins (Mr 49, 50, 60, 135 and 200 kDa) from DENV-4-infected mosquitoes were identified at higher concentrations than in uninfected controls. Further work is now needed to specifically identify these proteins and study their biological roles in DENV infections.
Keywords: Dengue; Aedes aegypti; infection proteins; protein profiles.
# E-mail: [email protected]
IntroductionIn Malaysia, classical dengue fever was first documented in 1901-1902,[1] while the first reported outbreak of dengue haemorrhagic fever (DHF) occurred in 1962.[2] Since then dengue has remained endemic, with one of the four dengue serotypes circulating in the country and outbreaks of DHF being reported periodically. At present, neither an effective vaccine nor a specific drug treatment for DF/DHF is available, making the laboratory-based surveillance system an important tool to provide an early warning of an impending outbreak of the disease.
Vector survei l lance al lows t imely implementation of emergency mosquito control measures such as space application of chemical insecticides against adult mosquitoes and destruction of their breeding places to contain an outbreak from spreading. However, when the adult mosquito density is low, direct entomological monitoring is not a sensitive enough indicator to serve as an early warning surveillance system for outbreak prevention. It is in this particular situation that detection of dengue viruses in vector population becomes an important element as part of an early alert system.[3,4,5]

116 Dengue Bulletin – Volume 33, 2009
Protein profiles of dengue-infected Ae. aegypti (L)
The current “gold s tandard” for dengue detection in mosquitoes involves isolation of the virus followed by indirect immunofluorescence.[5] However, this requires cell culture facilities which are difficult to maintain and the technique is not sensitive. The reverse transcriptase-polymerase reaction (RT-PCR) technique is sensitive and specific but very costly, especially during an outbreak where large-scale screening needs to be performed. Hence, alternative methods of detection are needed.
It is observed that dengue is less likely to cause mortality to the Aedes vector, leading to the belief that the mosquitoes may synthesize certain proteins (known as “infection proteins”) to work against the dengue viruses during invasion. Detection of these proteins, therefore, not only provides further understanding of the interaction between both the dengue virus and its vector but also could be the potential antigen used for detecting the presence of the virus in the mosquito.
Materials and methods
Preparation of dengue virus source for membrane feeding
The Aedes albopictus cell line C6/36, maintained with 10% fetal bovine serum in the minimum essential medium (MEM), was used to propagate dengue virus (DENV). DENV-2 and DENV-4-infected, as well as non-infected, C6/36 Ae. albopictus cell-culture supernatants (DENV-2/-4 ICS and NICS, respectively) were used to spike normal human blood for membrane feeding. The DENV-2/-4 ICS and NICS were clarified by centrifugation, and confirmed to contain either DENV-2/-4 ICS or to be uninfected (NICS) using the DENV serotype-specific RT-PCR (reference) and stored at –70 °C.
Preparation of blood meal for artificial membrane feeding
Initially, equal volumes of blood and phosphate buffered saline (PBS) were mixed in a 10 ml culture tube and centrifuged at 500 x g for 10 minutes at room temperature. The supernatant was then removed and the large blood cells (erythrocytes and leukocytes) were further washed five times with PBS using this method until the supernatant became clear. The blood cells were then transferred to fresh culture tubes.
Artificial membrane feeding
An artificial membrane feeding technique employed was modified from Graves (1980). Two hundred four- to seven-day-old adult female Ae. aegypti mosquitoes were collected into paper cups and starved overnight prior to blood feeding. A total of 30 female mosquitoes were placed into each paper cup covered with netting. A glass feeder fitted with a water jacket at 37 °C was covered at the bottom by wrapping a small piece of membrane made from chicken skin moistened with normal saline.
In the preparation of the artificial infective blood meal, 6 ml of washed red blood cells was mixed with 3 ml of D2/4-ICF. One ml of adenosine 5′-triphosphate (ATP) was added to the final concentration of 0.02 M to stimulate mosquitoes to engorge.[6,7] For negative control, the same ratio of washed red blood cells, normal CF and ATP was used. The blood meals were heated to 37 °C before being presented to the mosquitoes. Blood was presented to the mosquitoes by placing the cup containing the mosquitoes below the feeder, with the surface of the nylon netting of the cup in contact with the membrane of the feeder. The mosquitoes were then allowed to feed for approximately 10 to 30 minutes.

Dengue Bulletin – Volume 33, 2009 117
Protein profiles of dengue-infected Ae. aegypti (L)
Rearing of infected mosquitoes
Fully-engorged mosquitoes were selected and transferred to a small cage. The dengue-infected mosquitoes were reared at 28 °C and 70%–80% relative humidity in ACL-2 insectarium for 15 days before being killed. A single mosquito was transferred into a nuclease-free 1.5 ml microcentrifuge tube and kept at –70 °C in a freezer for further analysis.
Detection of dengue virus using RT-PCR
The single mosquito was homogenized on ice using a homogenizer attached to a Pellet Pestle Motor (Kontes, Japan) in a nuclease-free 1.5 ml microcentrifuge tube containing 210 µl of nuclease-free, double-distilled water. The homogenized samples were then centrifuged at 5000 x g for 10 minutes at 4 °C. A similar procedure was performed for non-infected mosquito homogenates. The rest of the samples were thawed and put on ice. High pure viral nucleic acid kit (Roche Diagnostic Corp., USA) was used to extract the viral RNA from the mosquito homogenates following the manufacturer’s guidelines. Extracted RNA was kept at –20 °C until used.
RT-PCR
The RT-PCR method of Lanciotti et al. (1992) was employed. DENV-2-specific primers (GTTCCTCTGCAAACACTCCA and GTGTTATTTTGATTTCCTTG) or DENV-4-specific primers (CCATTATGGCTGTGTTGTTT and CTTCATCCTGCTTCACTTCT) were used. The Access RT-PCR System (Promega, USA) was used to trans cribe the DENV RNA to cDNA. Each reaction contained 10.25 µl of double-distilled water, 2 µl of dNTP mixture, 1.25 µl of dithiotreitol, 0.5 µl of RNAse inhibitor, 5.0 µl of RT-PCR buffer, 0.5µl of enzyme mixture, 0.5 µl of each dengue primer and 0.5µl of RNA.
For dengue virus detection, the reaction was carried out at 51 °C for 30 minutes to produce cDNA, which was then amplified by the following PCR steps: initial denaturation at 92 °C for three minutes, 41 cycles of 92 °C for 30 seconds, 51 °C for 45 seconds and 72 °C for one minute; followed by 72 °C for five minutes. For every RT-PCR, a positive control and a negative control were included.
PCR products were analysed by performing electrophoresis in 2.0% Nusieve PCR gel (FC Bio, USA) at 100 volts and staining with ethidium bromide. The gel was viewed under ultraviolet illuminator (Ultra Lum Inc, California, USA) and the resulting bands were photographed with a polaroid camera.
Protein analysis of DENV-2- and DENV-4-infected mosquitoes
SDS-PAGE gel was prepared using Dual Slab Gel system. Both RT-PCR-detected infective mosquito homogenates and non-infected mosquito homogenates (negative control) were homogenized in double-distilled water and the extract was separated on SDS-polyacrylamide slab gel using the discontinuous system consisting of 4% acrylamide stacking gel and 12% acrylamide separating gel. Approximately 10 ul of the mosquito samples were boiled at 100 °C for five minutes before loading onto the gel. The separated protein bands were visualized by staining with Coomassie brilliant blue.
Results
The results obtained in a previous study[8] showed that the extrinsic incubation period for Aedes mosquitoes (DENV-2-infected) decreased from Day 11 to Day 17 and (DENV-4-infected) from Day 7 to Day 19 as the temperature increased from 26 °C to

118 Dengue Bulletin – Volume 33, 2009
Protein profiles of dengue-infected Ae. aegypti (L)
30 °C. The mortality rate of mosquitoes was found to be high at 30 °C compared with that at 28 °C or 26 °C. Based on these results, the extrinsic incubation temperature of 28 °C and incubation period of 15 days was, therefore, selected in this experiment in order to obtain a high number of live infected mosquitoes and a shorter viral replication time.
Protein synthesis in DENV-2-infected Ae. aegypti
Figure 1 shows the agarose gel electrophoresis of individual DENV-2-infected Ae. aegypti after RT-PCR assay. Lane 1, 2, 4, 5, 6, 7 and 9 represent the dengue-infected Aedes mosquito.
The protein profiles of individual DENV-2-infected Ae. aegypti are shown in Figures 2 and 3. There are at least 11 protein bands, which fall in the range of approximately 200 kDa to less than 24 kDa for both experimental and control mosquito. The predominant
Figure 1: Result of agarose gel electrophoresis of individual DENV-2-infected Ae. aegypti collected on 15th day after dengue viral amplification through RT-PCR assay
Figure 2: Protein profiles of DENV-2-infected Ae. aegypti sacrificed on 15th day

Dengue Bulletin – Volume 33, 2009 119
Protein profiles of dengue-infected Ae. aegypti (L)
polypeptides are approximately: 181.8 kDa, 135 kDa, 115.5 kDa, 110 kDa, 64.2 kDa, 60 kDa, 50 kDa, 49 kDa, 45 kDa, 30 kDa and 24 kDa.
Three polypeptides out of the 11 were found intensely expressed in DENV-2-infected Ae. aegypti compared to the control sample. These included proteins with an approximate molecular weight of 200 kDa, 64 kDa and 22 kDa. In contrast, one protein with molecular weight of not less than 60 kDa was found suppressed compared with the control sample.
Protein synthesis in DENV-4-infected Ae. aegypti
Figure 3 represents the agarose gel electrophoresis of individual DENV-4-infected Ae. aegypti after RT-PCR assay. Lane 1, 3, 4, 6 and 7 represent the dengue-infected Aedes mosquito.
Figure 3: Schematic diagram of protein profiles of DENV-2-infected Ae. aegypti
executed on 15th day Bandwidth indicates intensity (semi-quantitative)
Figure 4: Result of agarose gel electrophoresis of individual DENV-4-infected Ae. aegypti collected on 15th day after dengue viral amplification through RT-PCR assay

120 Dengue Bulletin – Volume 33, 2009
Protein profiles of dengue-infected Ae. aegypti (L)
The protein profiles of individual DENV-4-infected Ae. aegypti are shown in Figure 5. The predominant protein bands observed were in the range of approximately 200 kDa to less than 24 kDa in both experimental and control mosquito (Figure 6). These proteins are having molecular weight of approximately 181.8 kDa, 135 kDa, 115.5 kDa, 110 kDa, 64.2 kDa, 60 kDa, 50 kDa, 49 kDa, 45 kDa, 30 kDa and 24 kDa.
Out of the 11 proteins observed, there were five proteins with approximate molecular weight of 181.8 kDa, 135 kDa, 60 kDa, 50 kDa and 49 kDa that were highly expressed in DENV-4-infected Aedes mosquito.
The Table summarizes the protein profiles obtained in both DENV-2- and DENV-4-infected Ae. aegypti. It is obvious that uninfected mosquitoes also produced similar protein profiles, although in both
Figure 5: Protein profiles of DENV-4-infected Ae. aegypti sacrificed on 15th day
Figure 6: Schematic diagram of protein profiles of DENV-4-infected Ae. aegypti
executed on 15th day
Table: Molecular weight of proteins synthesized in both experimental and
control mosquito
No.Molecular weight, kDa
Control DENV-2 DENV-4
1 >181.8 >181.8* >181.8*
2 >135 >135 >135*
3 115.5 115.5 115.5
4 >110 <110 >110
5 64.2 64.2* 64.2
6 >60 >60** >60*
7 >50 >50 >50*
8 >49 >49* >49*
9 >45 >45 >45
10 >30 >30 >30
11 >24 >24 >24
* highly expressed protein ** suppressed protein

Dengue Bulletin – Volume 33, 2009 121
Protein profiles of dengue-infected Ae. aegypti (L)
DENV-2- and DENV-4-infected mosquitoes some of the similar proteins were over-expressed.
DiscussionAs mentioned earlier, RT-PCR is a powerful tool which can be used in the detection of infection due to virus such as dengue. This technique, however, needs to be used in the laboratory as it is difficult to carry out in the field. Therefore, development of an ELISA-based antigen detection system may be necessary if anti-dengue viral “proteins” (or “infection proteins”) are to be obtained from the dengue virus-infected mosquitoes, and to be used as an ELISA-based detection kit to detect dengue infection in mosquitoes.
The “infection proteins” produced in a vector may act as a kind of defence mechanism to protect itself from a detrimental condition. For example, innate immune response is activated against the infection of plasmodium in Anopheles gambiae[9] as defensive and putative Gram-negative bacteria-binding proteins are synthesized mainly to block the parasite from entering into the mosquito. Chee and AbuBakar[10] had identified a tubulin or tubulin-like C6/36 mosquito cell protein which was able to bind to DENV-2 virus. It is believed that Aedes mosquito may also elicit the defense component (or “infection proteins”) to protect itself from the invaded dengue viruses. Yunus[11] found the presence of “infection proteins” in dengue-infected Ae. albopictus with molecular weight of 18 kDa, 27 kDa, 28 kDa and 70 kDa, while Rohani et al.[12] found that there are four such proteins having molecular weight of 24 kDa, 25 kDa, 31 kDa and 76 kDa from the DENV-2-infected Ae. aegypti.
There are 11 predominant polypeptides observed in the control with molecular weight of not less than 181.8 kDa, 135 kDa, 115.5 kDa, 110 kDa, 64.2 kDa, 60 kDa, 50 kDa, 49 kDa, 45 kDa, 30 kDa and 24 kDa. The result reported herein, however, is not identical to the findings reported by Rohani et al.[12] In their study, there were seven conspicuous proteins in the range of 72 kDa to 17 kDa detected in the normal blood-fed Ae. aegypti. The difference could be due to the concentration of the SDS-PAGE gel used since 10% of separating gel was used to separate the proteins present in the control mosquito in this study, as compared with 12% separating gel employed in Rohani et al.[12] study. Therefore, the major proteins determined in this study are relatively higher in molecular weight compared with the study done by Rohani et al.[12] where lower molecular weight of polypeptides was observed. On the other hand, Lee et al.[13] reported that 29 protein bands were observed in the sugar-fed Ae. aegypti. This shows that the protein synthesized in Aedes mosquito could also be closely related to the food consumed by the mosquitoes.
The protein profiles of the dengue-infected mosquito were found similar to that of the control, with an overall of 11 conspicuous polypeptides found. These proteins, however, exhibited dif ferent expression levels represented by the bandwidth. Proteins with molecular weight of not less than 181.8 kDa, 64.2 kDa and not less than 49 kDa were highly expressed in DENV-2-infected mosquito compared with the control sample, while proteins with molecular weight of not less than 181.8 kDa, 135 kDa, 60 kDa, 50 kDa and 49 kDa showed a broader bandwidth in the protein profiles of DENV-4-infected mosquito compared with the control. Such patterns may be due to the high expression

122 Dengue Bulletin – Volume 33, 2009
Protein profiles of dengue-infected Ae. aegypti (L)
of the ordinary proteins in dengue-infected Aedes mosquito during dengue virus invasion in Aedes mosquito. Another possibility could be the different type of protein(s), or “infection protein(s)”, having similar molecular weight(s) to the ordinary protein(s), being synthesized that hence fall in the same distance of the SDS-PAGE gel.
Suppression of protein with molecular weight of not less than 60 kDa was assumed to occur in the DENV-2-infected Aedes mosquito when compared with the control. The molecular weight of 28kDa also seemed to be suppressed in DENV-2-infected Ae. aegypti and such finding was also reported before by Rohani et al.[12] Since no protein suppression was observed in DENV-4-infected mosquito, there is a possibility that the protein synthesis could be serotype-specific.
Alcon et al.[14] reported that NS1 protein, which is not part of the virion and having molecular weight of 48 kDa, was found on the surface of the infected cells or secreted extracellularly into the blood circulation in dengue patients. The function of this protein remains unknown, but it is believed to correlate with the development of dengue haemorrhagic fever.[15,16] Tubulin or tubulin-like mosquito cell protein reported by Chee and AbuBakar[10] is having similar molecular weight of about 48 kDa. Wang et al.[17] reported that there are abundant brush borders found in the midgut of Ae. aegypti, and this element could be the initial interaction site between dengue virus and the mosquito. Hence, it is possible that the
“infection protein” synthesized may appear in the brush border to act as a defence barrier to block the entry of the dengue viruses.
Huang et al.[18] had successfully proved that this viral NS1 protein could be used as an ELISA antigen to detect dengue infection in patients. The protein with a molecular weight of about 49 kDa found in this study is unlikely to be similar to 48 kDa protein reported in cell culture and dengue patient, as this protein was also secreted in DENV-2- and DENV-4-infected and control adult mosquitoes. However, although similar proteins were found in both infected and control Aedes mosquitoes, several over-expressed proteins observed only in infected mosquitoes could be considered potential diagnostic antigens to be used for detecting dengue infection in mosquitoes, based on the quantitative differences in protein concentrations. It is pertinent, therefore, to quantify the various over-expressed proteins in any future study.
AcknowledgementsThe authors thank the Director-General of Health, Malaysia, for his permission to publish this research. Works mentioned in this paper were a part of the Master of Science thesis of the second author at the University of Malaya, Kuala Lumpur, and were partially supported by a research grant No. JPP-05-006 from the National Institute of Health, Ministry of Health, Malaysia.
ReferencesSkae FM. Dengue fever in Penang. [1] Br Med J 1902;2:1581-2.
Lam SK. Two decades of dengue in Malaysia. [2] Trop Med 1993;35(4):195-200.
Liotta DJ, Cabanne G, Campos R , Tonon [3] SA. Molecular detection of dengue viruses in field caught Aedes aegypti mosquitoes from northeastern Argentina. Rev Latinoam Microbiol 2005;47(3-4):82-7.

Dengue Bulletin – Volume 33, 2009 123
Protein profiles of dengue-infected Ae. aegypti (L)
Chow VTK, Chan YC, Rita Yong, Kim LK, [4] Lim LK, Chung Y, Lam-Phua SG. Monitoring of dengue viruses in field-caught Aedes aegypti and Aedes albopictus mosquitoes by a type-specific polymerase chain reaction and cycle sequencing. Am J Trop Med Hyg 1998;58(5):578-86.
Harris E, Roberts TG, Smith L, Selle J, Kramer [5] LD, Valle S, Sandoval E, Balmaseda A. Typing of dengue viruses in clinical specimens and mosquitoes by single-tube multiplex reverse transcriptase PCR. J Clin Microbiol 1998;36(9):2634-9.
Rutledge LC, Ward RA, Gould DJ. Studies on [6] the feeding response of mosquitoes to nutritive solutions in a new membrane feeder. Mosq News 1964;24(4):407-19.
Wirtz RA, Rutledge LC. Reconstituted collagen [7] sausage casings for the blood feeding of mosquitoes. Mosq News 1980;40(2):287-8.
Rohani A, Wong YC, Zamree I, Lee HL, [8] Zurainee MN. The effect of extrinsic incubation temperature on development of dengue serotype 2 and 4 viruses in Aedes aegypti (L.). Southeast Asian J Trop Med Pub Hlth 2009; 40(5):942-50.
Richman AM, Dimopoulos G, Seeley D, [9] Kafatos FC. Plasmodium activates the innate immune response of Anopheles gambiae mosquitoes. EMBO J 1997;16(20):6114-9.
Chee HY, AbuBakar S. Identification of a [10] 48kDa tubulin or tubulin-like C6/36 mosquito cells protein that binds dengue virus 2 using mass spectrometry. Biochem Biophys Res Commun 2004;320(1):11-7.
Yunus W. [11] Protein synthesized by mosquito in response to dengue virus infection. Thesis in Applied Parasitology and Entomology. Kuala Lumpur: Institute for Medical Research, 2000.
Rohani A, Yunus W, Zamree I, Lee HL. Protein [12] synthesized by Aedes aegypti and Aedes albopictus. Trop Biomed 2005;22(2):233-42.
Lee HL, Murahwa FC, Gan SC, Nasuruddin HA. [13] Protein profiles of Malaysian Aedes aegypti and Anopheles maculates and their characterization. Trop Biomed 1994;11:155-60.
Alcon S, Talarmin A, Debruyne M, Falconar [14] A, Deubel V, Flamand M. Enzyme-linked immunosorbent assay specific to dengue virus type 1 nonstructural protein NS1 reveals circulation of the antigen in the blood during the acute phase of disease in patients experiencing primary or secondary infections. J Clin Microbiol 2002;40(2):376-81.
Falconar AK. The dengue virus nonstructural-1 [15] protein (NS1) generates antibodies to common epitopes on human blood clotting, integrin/adhesion proteins and binds to human endothelial cells: potential implication in haemorrhagic fever pathogenesis. Arch Virol 1997;142(5):897-916.
Libraty DH, Young PR, Pickering D, Endy TP, [16] Kalayanarooj S, Green S, Vaughn DW, Nisalak A, Ennis FA, Rothman AL. High circulating levels of the dengue virus non-structural protein NS1 early in dengue illness correlate with the development of dengue hemorrhagic fever. J Infect Dis 2002;186:1165-8.
Wang P, Conrad JT, Shahabuddin M. [17] Localization of midgut-specific protein antigens from Aedes aegypti (Diptera: Culicidae) using monoclonal antibodies. J Med Entomol 2001;38(2):223-30.
Huang JL, Huang JH, Shyu RH, Teng CW, Lin [18] YL, Kuo MD, Y`ao CW, Shaio MF. High level expression of recombinant dengue viral NS1 protein and its potential use as a diagnostic antigen. J Med Virol 2001;65:553-60.

124 Dengue Bulletin – Volume 33, 2009
Susceptibility status of transgenic Aedes aegypti (L.) against insecticides
W.A. Naznia#, S. Selvia, H.L. Leea, I. Sadiyaha, H. Azaharia, N. Derricb and S.S. Vasanc
aMedical Entomology Unit, Infectious Disease Research Centre, Institute for Medical Research, Jalan Pahang 50588 Kuala Lumpur, Malaysia
bOxitec Limited, 71 Milton Park, Abingdon, Oxford OX14 4RX,UK
cCEBAR, IPS Building, Level 5, Block B, University of Malaya, Kuala Lumpur, 50603, Malaysia
Malaysia Oxitec S/B, Plaza See Hoy Chan, Suite 1502, Jalan Raja Chulan, Kuala Lumpur, 50200, Malaysia
Abstract
Two strains of Aedes aegypti, a RIDL® strain (MyRIDL513A) and a laboratory strain (MyWT), were used in the insecticide-susceptibility study. Two-to-five-day-old female mosquitoes from both strains were tested for their susceptibility against seven insecticides from the class of organophosphate, carbamate, pyrethroids and organochlorine. The adult bioassay was performed according to WHO standard procedures. The 50% lethal time (LT50) value was determined for each strain against the seven insecticides. Both the MyRIDL513A and MyWT strains were resistant to DDT, exhibiting mortality of 48% and 33% respectively, but were susceptible to malathion (5%), permethrin (0.75%), cyfluthrin (0.15%) and lambdacyhalothrin (0.05%). This study shows there is no evidence of altered susceptibility to insecticides in the RIDL strain compared to a WT strain of Ae. aegypti.
Keywords: Aedes aegypti; transgenic Ae. aegypti; dengue; insecticides; RIDL Ae. aegypti.
#E-mail: [email protected]
IntroductionDengue is a mosquito-borne infection that in recent decades has become a major international public health concern. Dengue is prevalent in the tropical and sub-tropical regions of the world, predominantly in urban and semi-urban areas. Dengue was
first reported in Malaysia in 1901–1902 on the island of Penang[1] and has since spread nationwide. A total of 30 981 cases and 70 deaths were recorded in Malaysia up to September 2009.[2] Ecological, behavioural and control information on population size, distribution, survivorship, seasonal abundance and insecticide susceptibility is urgently

Dengue Bulletin – Volume 33, 2009 125
Susceptibility status of transgenic Ae. aegypti (L.) against insecticides
required for an understanding of the epidemic potential and for the formulation of new control strategies.
Control and/or elimination of mosquitoes is a complex and difficult challenge but it remains the only viable option in the absence of a vaccine or antiviral treatment. Although space-spraying of insecticides is used widely, this method has so far failed to control the spread of Aedes aegypti, and long-term implications such as resistance[3] and the effects of residues in the environment are important considerations. Recent advances in molecular biology have brought some exciting new control possibilities.[4,5,6]
A particular technology known as RIDL® (Release of Insects carrying Dominant Lethality) has been developed in Ae. aegypti and, while the bionomics of transgenic Ae. aegypti have been evaluated and found to be indistinguishable from the wild type,[3] it is necessary to continue evaluating other aspects of the biology of RIDL Ae. aegypti as Ae. aegypti has been found to be resistant to insecticides in numerous locations throughout the world.[7] This paper reports the susceptibility status of the RIDL Ae. aegypti against insecticides commonly used to control the vector mosquitoes.
Materials and methods
Mosquito strains
Two strains of mosquitoes were used, viz. a laboratory-susceptible strain designated as MyWT strain originating from Penang, Malaysia, and reared in the laboratory for 1014 generations since 1965, and an F2 RIDL strain obtained from Oxitec Limited,
United Kingdom, and reared in the Arthropod Containment Laboratory Facility (ACLF) in the Institute for Medical Research, Kuala Lumpur.
My RIDL-513A strain
The original RIDL Ae. aegypti strain was designated as LA513A and was generated in a Rockefeller strain genetic background. The strain used in this study was generated (laboratory strain of Malaysian origin using 12 homozygous female founder parents) by out-crossing to the MyWT strain.
The RIDL strain was maintained in the Arthropod Containment Level-2 (ACL-2) laboratory at 26±1 °C and 70%–80% relative humidity with a photoperiod of 10 hours of artificial daylight and 14 hours of darkness.[8] Ten drops of Liquifry® No. 1 fish food were added to induce egg hatching in a tray half filled with 1.5 litre of tetracycline water (tet-water) at 30 mg/l to suppress the lethal effect of the RIDL system. Larvae were fed with powdered fish food (Tetramin®) while emerged adults were fed with 10% sucrose supplemented with 1% vitamin B complex solution soaked in lint cloth and placed inside a small plastic bottle. Five days after adult emergence, the females were permitted to blood-feed on mice. Three days after feeding, a piece of moist filter paper in a porcelain bowl half filled with water was introduced for oviposition.
MyWT strain
The origin of the MyWT strain was from Selangor, a state in peninsular Malaysia. The MyWT larvae were reared as above, except that the first and second instar larvae were fed

126 Dengue Bulletin – Volume 33, 2009
Susceptibility status of transgenic Ae. aegypti (L.) against insecticides
on cow liver powder whereas the third and fourth instar larvae were fed on small pieces of partially-cooked liver. Emerged adults were fed with 10% sucrose solution supplemented with 1% vitamin B complex solution soaked in lint cloth and placed inside a small plastic bottle.
Insecticides
The insecticides used in the adult susceptibility test were diagnostic dosages as specified in the WHO standard method. The insecticide-impregnated papers were obtained from the Vector Control Research Centre, Universiti Sains Malaysia. The adults were tested against two organophosphates (5% malathion, 1% fenitrothion), three pyrethroids (0.75% permethrin, 0.05% lambdacyhalothrin, 0.15% cyfluthrin), an organochlorine (4% DDT) and a carbamate (0.1% propoxur). The exposure time for DDT was half an hour and the exposure time for all other insecticides was one hour.
WHO adult bioassay
The bioassay procedure of WHO was used.[9] Sugar-fed, 3–5-days-old adult female mosquitoes were tested. Batches of 25 adults were introduced into pre-holding tubes prior to being exposed to insecticide-impregnated papers in standard WHO test tubes lined with the impregnated papers. Exposed and control tubes of mosquitoes were covered with black cloth during exposure. Equal numbers of control tests were also carried out by exposing mosquitoes to untreated filter paper for one hour. The experiment was replicated four times. All tests were undertaken at 26 °C ± 2 °C and relative humidity of 70%–80% with a photo period of 12-hour darkness and 12-hour light. The mosquitoes were exposed to the
diagnostic dosages at the respective exposure period.[9] Cumulative mortality counts or knockdown were recorded at regular intervals for the respective exposure periods.
After the experiment, all mosquitoes were transferred into a clean paper cup and provided with 10% sugar solution. The test mosquitoes and the controls were held for a 24-hour recovery period and mortality was recorded. If the control mortality was between 5% and 20%, the percentage mortalities were corrected by Abbott’s formula.[10] All data were analysed using a probit analysis computer programme and LT50 and LT90 for each insecticide for both the strains were calculated.[11]
Results and discussion
The RIDL strain was originally developed in a Rockefeller background, which is a laboratory strain originally isolated from the wild over 50 years ago. We wanted to potentially improve this strain by out-crossing to a more recently isolated strain. This was done into a Malaysian strain isolated from the wild more than 45 years ago. In order to determine that the process of out-crossing had no effect on the strain background in terms of insecticide resistance, it was tested against several insecticides commonly used for control.
According to WHO,[12] if the mortality is in the range of 98%–100%, the insects are susceptible to the insecticide; 80%–97% means additional verification is required, and <80% mortality shows that the insects are resistant.
The results of the bioassays on both strains of Ae. aegypti are shown in the Table. The

Dengue Bulletin – Volume 33, 2009 127
Susceptibility status of transgenic Ae. aegypti (L.) against insecticides
Table: Susceptibility status of adults of MyRIDL513A and MyWT Ae. aegypti against various insecticides
Insecticide Dosage
Lethal time, LT (minutes)
Regression line
24 hours post-exposure
mortality (%)
Susceptibility statusLT50
(95% C.L.)LT90
(95% C.L.)
DDT 4%
177.73a
(110.27 – 1109.69)493.54a
(82.79 – 115.72)Y = 2.89x – 30.37 48a Resistant
132.82b
(92.02 – 563.33)298.23b
(152.02 – 4100.77)Y = 3.69x – 39.72 33b Resistant
Fenitrothion 1%
78.44a
(68.19-184.44)104.85a
(80.81-524.47)Y = 10.17x – 116.45 98.7a Susceptible
9.76b
(9.24 – 10.29)21.07b
(19.18 – 23.64)Y = 3.83x – 37.07 96b Tolerant
Malathion 5%
27.65a
(27.32 – 27.97)32.56a
(32.01 – 33.21)Y = 18.05x – 201.56 100a Susceptible
26.05b
(25.66 – 26.45)32.21b
(31.53 – 33.01)Y = 13.92 x – 153.97 100b Susceptible
Propoxur 0.1%
28.53a
(27.94 – 29.15)40.29a
(38.91 – 41.97)Y = 8.55x – 92.93 100a Susceptible
127.99b
(101.99 – 181.47)810.42b
(460.66 – 2016.41)Y = 1.59x – 14.25 88b Tolerant
Permethrin 0.75%
11.75a
(11.41-12.08)19.80a
(18.97-20.81)Y = 12.57x – 11.11 100a Susceptible
14.21b
(13.92-14.52)21.39b
(20.60-22.30)Y = 13.89x – 11.11 100b Susceptible
Lambda-cyhalothrin 0.15%
13.75a
(13.32 – 14.18)23.05a
(22.09 – 24.17)Y = 5.72x – 69.28 100a Susceptible
13.79b
(13.44 – 14.12)18.50b
(17.91 – 19.24)Y = 10.03x – 106.77 100b Susceptible
Cyfluthrin 0.05%
5.12a
(4.93 – 5.31)6.67a
(6.33 – 7.17)Y = 11.15x – 114.46 100a Susceptible
10.12b
(9.88 – 10.36)12.81b
(12.41 – 13.32)Y = 12.51x – 132.73 100b Susceptible
a: MyRIDL513A b: MyWT (Lab strain) C.L.: Confidence Limit

128 Dengue Bulletin – Volume 33, 2009
Susceptibility status of transgenic Ae. aegypti (L.) against insecticides
LT50 value is commonly accepted as the basis for comparison in the investigation of relative toxicities among the insecticides under tests. The results show that both the MyRIDL513A and MyWT strains were resistant to DDT. Resistance to DDT could be due to the genetic origin of the mosquitoes because both possessed a Malaysian genetic background. DDT has been used in the past to control Ae. aegypti, and though its application was stopped in 1957 and substituted by dieldrin,[13] the resistance phenotype has remained in this laboratory strain. Natural pre-existing resistance of Ae. aegypti against DDT has been reported from Malaya[14] (former name for Malaysia), indicating this phenotype is also maintained in the wild.
Generally, against the MyRIDL513A strain, the resistance decreased in the order of: DDT > fenitrothion > propoxur > malathion > lambdacyhalothrin > permethrin > cyfluthrin. The strain MyWT, on the other hand, responded in a slightly different pattern, and the resistance decreased in the order: DDT > propoxur > fenitrothion > malathion > lambdacyhalothrin > permethrin > cyfluthrin. There was a variation in response to fenitrothion and propoxur in both the strains, but the resistance pattern of malathion and pyrethroids in the above order remained the same for both the strains.
However, exposure of RIDL adults to the discriminating dosages of malathion, permethrin, cyfluthrin and lambdacyhalothrin induced 100% mortality 24 hours post-treatment, indicating that the RIDL mosquito was also susceptible to these insecticides. Strains that are resistant to DDT have been shown to have moderate resistance to pyrethroids, whereas permethrin resistance
resulted in strong resistance to both permethrin and DDT.[15] However, our study indicated that there was no cross-resistance to pyrethroids and this was in accordance with a study in Thailand on wild-type mosquitoes.[16]
Fenitrothion and propoxur gave 98.7% and 100% mortality for MyRIDL513A, indicating high susceptibility of this strain. On the other hand, the MyWT strain showed 96% and 88% mortality against fenitrothion and propoxur, indicating possible tolerance of this strain to these insecticides. The development of tolerance could be due to the selection for resistance in the mosquitoes resulting from agricultural application.[17] The MyWT strain, originated from field-caught Ae. aegypti, may have been exposed and selected for resistance against these insecticides. Nevertheless, the trend in the susceptibility status for both the strains was similar.
In summary, both the MyRIDL and MyWT strains of Ae. aegypti exhibited almost identical levels of susceptibility/resistance to insecticides. Therefore, the process of out-crossing RIDL to a Malaysian strain has not changed the insecticide-susceptibility status.
Acknowledgement
We thank the Director-General of Health, Malaysia, and the Director, Institute for Medical Research (IMR), for permission to publish this paper. This study was supported by a grant (No JPP-IMR-053-07) from the National Institutes of Health, Ministry of Health, Malaysia. Thanks are also due to the staff of the Medical Entomology Unit, IMR, for their assistance and to Oxitec Ltd, U.K., for the provision of the RIDL strain.

Dengue Bulletin – Volume 33, 2009 129
Susceptibility status of transgenic Ae. aegypti (L.) against insecticides
References Skae FM. Dengue fever in Penang. [1] BMJ 1902;2:1581–2.
Ministry of Health. [2] Dengue and chikungunya vases 2009; http://www.moh.gov.my/MohPortal/Index.jsp – accessed 30 June 2010.
Brown AWA. Insecticide resistance in [3] mosquitoes: a pragmatic review. J Am Mosq Control Assoc 1986;2:123-40.
Alphey L. Re-engineering the steri le [4] insect technique. Insect Biochem Mol Biol 2002;32(10):1243-1247.
Alphey L, Benedict M, Bellini R, Clark GG, [5] Dame DA, Service MW, Dobson SL. Sterile-insect methods for control of mosquito-borne diseases: an analysis. Vector Borne Zoonotic Dis 2010;10(3):295-311.
Phuc HK, Andreasen MH, Burton RS, Vass C, [6] Epton MJ, Pape G, Fu G, Condon KC, Scaife S, Donnelly CA, Coleman PG, White-Cooper H, Alphey L. Late-acting dominant lethal genetic systems and mosquito control. BMC Biol 2007;5:11.
Lee HL, Joko H, Nazni WA, Seshadri Vasan. [7] Comparative life parameters of transgenic and wild strain of Aedes aegypti (L.) in the laboratory. Dengue Bull 2009 (submitted).
Nimmo D, Gray P. [8] Mosquito rearing protocol. Intensive Workshop on Wild Type and Genetically Sterile Aedes Mosquitoes, 26 Sept – 2 Oct 2007, Kuala Lumpur Malaysia, pp: 27-34.
World Health Organization. Instruction for [9] determining the susceptibility of resistance of adult mosquitoes to organochlorines, o r g a n o p h o s p h a t e s a n d c a r b a m a t e insecticide diagnostic test. World Health Organization Mimeograph 1981; WHO.
VBC/81.8067. World Health Organization. Discriminating concentrations of insecticides for adult mosquitoes. 1998; WHO/CDS/CPC/MAL/98.12.
Abbott WS. A method for computing the [10] effectiveness of an insecticide. J Econ Entomol 1925;18:265-7.
Raymond M. Log-probit analysis basic [11] programme of microcomputer. Cohiers ORSTOM Serie. Entomology Medicale et Parasitologie 1985;23:117-121.
World Health Organization. [12] 10th report of the WHO expert committee on vector biology & control: resistance of vector and reservoir of disease to pesticides. Technical Report Series 737. Geneva: WHO, 1986.
Macdonald WW. Resurvey Of [13] Aedes aegypti at Kuala Lumpur Airport. Medical J Malaya 1958;XIII:179-86.
Shidrawi GR. Laboratory tests on mosquito [14] tolerance to insecticides and the development of resistance by Aedes aegypti. Bull Wld Hlth Org 1957;17:377-411.
Grant DF, Matsumura F. Glutathione [15] S-transferase 1 and 2 in susceptible and insecticide resistant Aedes aegypti. Pestic Biochem Physiol 1989;33:132-43.
La-aied Prapanthadara, Nongkran Promtet, [16] Surangchit Koottathep, Pradya Somboon, Wonnapa Suwonkerd, Lynn McCarroll, Janet Hemingway. Mechanisms of DDT and permethrin resistance in Aedes aegypti from Chiang Mai, Thailand. Dengue Bull 2002;26:185-9.
Lines JD. Do agricultural insecticides select [17] for insecticide resistance in mosquitoes? A look at the evidence. Parasitology Today 1988;4:S17-S20.

130 Dengue Bulletin – Volume 33, 2009
Epidemiological analysis of hospitalized cases of DF/DHF in NCT Delhi, 2005–2009
Epidemiological analysis of hospitalized cases of dengue fever/dengue haemorrhagic fever and extent of breeding
of Aedes aegypti in major hospitals in the National Capital Territory of Delhi (NCT Delhi), 2005–2009
J. Nandia#, R.S. Sharmaa, R.K. Dasguptaa, R. Katyalb, P.K. Duttac and G.P.S. Dhillona
aDirectorate of National Vector Borne Disease Control Programme, 22 Shamnath Marg, Delhi 110054, India
bNational Centre for Disease Control, 22 Shamnath Marg, Delhi 110054, India
cEx-Associate Professor, Armed Forces Medical College, Pune, India
Abstract
Dengue is a notifiable disease in the National Capital Territory of Delhi (NCT Delhi), India. All hospitals, both in the public and private sectors, are under obligation to report serologically confirmed cases of dengue to local health authorities. During the period 2005 to 2009, a total of 7402 serologically confirmed dengue cases were reported from the National Capital Territory of Delhi. Records of 5603 dengue cases (76%) admitted in hospitals were analysed for severity of disease. The trend of dengue has changed from cyclic to annual occurrence. DHF/DSS accounted for 518 (9.2%) of the admitted hospital cases in all age groups. The proportion of males found positive for dengue infection was 68% while females constituted 32%. The transmission season in NCT Delhi is the rainy season (July to October). Container indices monitored in six major hospitals remained persistently high in all the five years (range 1.5 to 23.9) and carried high potential for spatial spread of dengue infection to other parts of the NCT, Delhi region.
Keywords: Dengue haemorrhagic fever (DHF); vulnerability; receptivity; hospitals; Aedes aegypti; NCT Delhi.
#E-mail: [email protected]
IntroductionThe National Capital Territory of Delhi (NCT Delhi), India, has now become an endemic region for dengue. All the four serotypes, i.e. DENV-1–4, and several genotypes including
DENV-3 subtype-III – the most virulent strain known to cause a high incidence of dengue haemorrhagic fever (DHF) and dengue shock syndrome (DSS) – were reported circulating in Delhi and its surrounding areas.[1,2] In 1988, the NCT Delhi recorded an outbreak

Dengue Bulletin – Volume 33, 2009 131
Epidemiological analysis of hospitalized cases of DF/DHF in NCT Delhi, 2005–2009
of dengue with 33% mortality among children admitted in hospitals.[3] In 1996, another severe outbreak in the NCT, Delhi recorded 10 252 cases hospitalized for dengue with 423 deaths.[4] Currently, the trend of the disease has changed from being cyclic to annual incidence.[5] The control of vector-borne diseases, including dengue, in the NCT region vests with multiple public health agencies such as the Municipal Corporation of Delhi (MCD), the New Delhi Municipal Council (NDMC), Defence and Railways.[6] There exist a number of autonomous organizations situated within the jurisdiction of each of these agencies that include major hospitals which are excluded from the purview of public health agencies for undertaking preventive health measures. These autonomous organizations and hospitals are poorly equipped both in terms of trained manpower and skills. As such they are highly prone to breeding of dengue vectors because of their vulnerability to dengue virus as most dengue patients, both asymptomatic and symptomatic, visiting these hospitals are sufficiently viraemic. Thus, these hospitals become major centres for the spatial spread and dissemination of the disease to other parts of the city.[7,8] The present study aimed to focus on the epidemiological information on dengue cases by analysing serologically confirmed hospital admissions during 2005–2009. This included: (i) demographic analysis; (ii) severity of infections; (iii) breeding indices (container index) of Ae. aegypti in major hospitals; and (iv) types of breeding containers of the vector species.
Study area
The NCT region has an area of 1485 sq. km., located between 28° 75’ north latitude and 76° 22’ east longitude. The population, as per the 2001 census, was 13.7 million
and since then it has increased to 15.4 million, an annual increase of 12.2% (Source: Municipal Corporation of Delhi). The NCT Delhi has witnessed phenomenal growth in its population in recent times, and large-scale migration from rural areas into the capital territory resulted in unplanned rapid urbanization that exerted considerable strain on civic amenities, particularly on water supply and solid waste disposal, leading to a predominance of water storage practices. Delhi being a capital city and a tourist centre attracts a large number of visitors/tourists, thus increasing the vulnerability of the city for dengue virus.
Materials and methods
Epidemiological surveillance
Dengue is a notifiable disease in the NCT Delhi. Thirty nine hospitals spread over the entire NCT Delhi follow the under-mentioned case definitions for the purpose of reporting under a mandatory surveillance system.[10] Hospitals report to local bodies (MCD and NDMC) for their records. Local bodies in turn transmit this information to the Directorate of National Vector Borne Disease Control Programme (NVBDCP), the nodal agency for the country.
Patients with clinical symptoms like ••sudden onset of high fever, severe body pain and headache, myalgia, nausea, vomiting and rash with positive dengue-specific IgM capture ELISA (MAC-ELISA) in a single serum specimen were to be considered as serologically-confirmed dengue cases. The IgM capture ELISA is man-ufactured by the National Institute of Virology, (NIV) Pune, India, and are

132 Dengue Bulletin – Volume 33, 2009
Epidemiological analysis of hospitalized cases of DF/DHF in NCT Delhi, 2005–2009
supplied to all hospitals for diagnosis of clinically suspected cases reporting to hospitals. The NIV MAC-ELISA carry 96% sensitivity when compared with Pan Bio IgM ELISA (73%).[11]
Clinical symptoms with low throm-••bocytopenia and leucopenia were also taken as confirmed cases of dengue fever. The presence of both these two criteria with haemorrhagic manifestation and deaths were taken as confirmed deaths due to dengue fever.
High sensitivity to IgM test is impor-••tant to support clinical diagnosis and case management in hospitals.[12]
Entomological surveillance
Entomological surveillance was carried out by the Central Cross-Checking Organization of NVBDCP in six major hospitals periodically during the transmission season for the period 2005 to 2009 due to known high receptivity of hospitals. Hospital compounds were thoroughly searched to detect breeding of Ae. aegypti, the vector of dengue. Infestation of Ae. aegypti was assessed by container index (CI) for measurement of breeding infestation following WHO guidelines.[9] Since the hospitals had several blocks with multiple entry and exit points the House and Breateu Indices were considered insensitive.
Meteorological data
The average annual rainfall (2003–2009) in the NCT Delhi ranged between 1 mm in November to 243.8 mm in July with maximum precipitation occurring during July and August. The average minimum temperature was 7.1 °C in January to 27.3 °C in July. The average
maximum temperature varied from 24.3 °C in January to 46.8 °C in May. Rainy season in the NCT Delhi usually extends from July to October (Source: Indian Meteorological Department, Government of India).
Results
DF/DHF incidence
During the period 2005 to 2009, a total of 7402 cases of DF/DHF were recorded (Figure 1). The annual incidence for the years 2005 to 2009, except 2006 which was an epidemic year, ranged from 548 in 2007 to 1312 cases in 2008. DHF/DSS accounted for 518 (9.2%) of hospital admitted cases in all age groups. Deaths were also few, and varied from one in 2007 to 9 in 2005. The year 2006 was an epidemic year and recorded 3366 cases and 38 deaths. (Source: Health Department, Municipal Corporation of Delhi).
Figure 1: Incidence of DF/DHF and deaths in the NCT Delhi (2005–2009)
Demographic analysis
Records of only 5603 serologically-confirmed hospital admitted dengue cases could be retrieved. Of these 62 cases (1.1%) belonged to the 0-1-year age group, 494 cases (8.8%)

Dengue Bulletin – Volume 33, 2009 133
Epidemiological analysis of hospitalized cases of DF/DHF in NCT Delhi, 2005–2009
to the 2–8-year age group, 982 cases (17.5%) to the 9–15-year age group and 4065 cases (72.6%) were >16 years old (Figure 2). The proportion of males found positive for dengue infection was 68% while females constituted 32% probably indicating a higher rate of transmission at workplaces rather than in households. Classical dengue generally affects older children and adults while DHF/DSS is associated with secondary infections in individuals aged one or more years, the majority in individuals aged 8 to 10 years, or primary infections in infants born to dengue immune mothers.[13] In the NCT region dengue is a mix of DF and DHF/DSS; therefore, it can safely be concluded that the region has gained notoriety for dengue endemicity.[14]
Figure 2: Age profile of 5603 confirmed dengue cases in NCT Delhi (2005–2009)
Figure 3: Month-wise distribution of DF/DHF cases in NCT Delhi (2005–2009)
Transmission season
The month-wise distribution of 5603 dengue cases admitted in hospitals in NCT Delhi is shown in Figure 3. The transmission season in the NCT region starts with the onset of the rainy season (July-October) and cases start appearing in July, peaking in October and then tapering off by November. The favourable temperature regime during the rainy season seems to be the guiding factor in the transmission.
Spatial distribution and incidence rate
The incidence rate per 100 000 population varied from 11.81 in the Narela zone to 57.37 in the south zone under MCD, while the incidence rate was 67.35 in NDMC areas (Table 1, Figure 4). Out of nine urban zones, five zones, namely Shahdara (South), Shahdara (North), Central, South and Civil Lines, border the neighbouring states of Uttar Pradesh and Haryana. The incidence rate in five zones in the central part of the NCT Delhi varied from 12.63 in the West zone to 27.92 in the Karol Bagh zone. The incidence rate in Najafgarh and Narela zone, these two being rural zones bordering the state of Haryana, was 29.0 and

134 Dengue Bulletin – Volume 33, 2009
Epidemiological analysis of hospitalized cases of DF/DHF in NCT Delhi, 2005–2009
Figure 4: Location of zones in NCT regionTable 1: Incidence rate of DF/DHF cases per 100 000 population reported by zones of the Municipal Corporation of Delhi (MCD) and the New Delhi Municipal Council (NDMC)
Zones under MCD & NDMC
Cases per 100 000
population
Karol Bagh 27.92
Sadar Paharganj 24.73
City 15.14
West 12.63
Najafgarh 29.00
Civil Lines 23.25
Shahdara (North) 20.61
Shahdara (South) 15.76
Narela 11.81
Rohini 20.91
South 57.37
Central 16.17
NDMC 67.35
11.81 respectively. The major tertiary level hospitals are located in NDMC zone and four urban zones under MCD also attract a number of patients from adjoining states.
Severity of the disease
The break-up of clinical cases by sign/symptoms and age group of DF/DHF cases as retrieved from hospital records is included in Table 2.
Proportion of DHF/DSS cases
DHF and DSS cases numbered 428 (82.6%) and 90 (17.4%) respectively among all age groups (Table 3).
Dengue haemorrhagic fever (DHF)
DHF cases in infants numbered 5 (1.2%), and 89 (20.8%) in the age group of 2–8 years, 90 (21%) in the age group of 9–15 years and 244 cases (57%) were in the >16-years age group.
Dengue shock syndrome (DSS)
The proportion of infants, children and adults who suffered form DSS was 3 (3.3%) in the 0–1-year age group, 31 (34.4%) in the age group of 2–8 years, 44 (48.9%) in the 9–15-years age group and 12 (13%) in the >16-years age group.

Dengue Bulletin – Volume 33, 2009 135
Epidemiological analysis of hospitalized cases of DF/DHF in NCT Delhi, 2005–2009
Table 2: Signs and symptoms by age group as retrieved from hospital records of DF/DHF cases
Age group/No.
Clinical manifestations
Fever with myalgia, headache
and bodyache
Positive tourniquet
t test
Petechiae ecchymosis or purpura
Bleeding from
mucosa, gum,
injection site or
any site
Haematemesis, melena or epistaxis
(haematological complications)
Thrombo-cytopenia
Evidence of
plasma leakage
Sign of circulatory
failure
0–1 + 0 17 (27.4) 12 (19.4) 4 (6.5) 29 (46.7) 0 0
2–8 + 3 (0.7) 92 (18.6) 44 (8.9) 37 (7.5) 181 (36.6) 74 (14.9) 63 (12.7)
9–15 + 41 (4.2) 109 (11.1) 55 (5.6) 123 (12.5) 436 (44.4) 95 (9.7) 123 (12.5)
16 to above + 3748 163 (4.0) 0 0 77 (1.9) 77 (1.9) 0
*Figures in parentheses indicate percentage
Table 3: Month-wise distribution by age of DHF and DSS cases (2005–2009)
Month
DHF (Figures in parenthesis are %)
DSS (Figures in parenthesis are %)
0–1 2–8 9–15 >16 Total 0–1 2–8 9–15 >16 Total
Jan 0 0 0 0 0 0 0 0 0 0
Feb 0 0 0 0 0 0 0 0 0 0
Mar 0 0 0 0 0 0 0 0 0 0
Apr 0 0 0 0 0 0 0 0 0 0
May 0 0 0 0 0 0 0 0 0 0
June 0 0 0 0 0 0 0 0 0 0
July 0 0 0 0 0 0 0 0 0 0
Aug 1 (0.5) 10 (2.3) 5 (1.2) 33 (7.7) 49 (11.4) 0 1 (1.1) 1 (1.1) 1 (1.1) 3 (3.3)
Sept 2 (0.5) 46 (10.7) 40 (9.3) 100 (23.4) 188 (44.0) 0 2 (2.2) 9 (10.0) 9 (10.0) 20 (22.2)
Oct 2 (0.5) 28 (6.5) 35 (8.2) 104 (24.3) 169 (39.5) 2 (2.2) 19 (21.1) 24 (26.7) 2 (2.2) 47 (52.2)
Nov 0 5 (1.2) 10 (2.3) 7 (1.6) 22 (5.1) 1 (1.1) 9 (10.0) 10 (11.1) 0 20 (22.2)
Dec 0 0 0 0 0 0 0 0 0 0
Total 5 (1.2) 89 (20.8) 90 (21) 244 (57) 428 (82.6) 3 (3.3) 31 (34.4) 44 (48.9) 12 (13) 90 (17.4%)

136 Dengue Bulletin – Volume 33, 2009
Epidemiological analysis of hospitalized cases of DF/DHF in NCT Delhi, 2005–2009
Table 4: Year-wise periodic check for container index in six major hospitals in NCT Delhi during transmission period July to October (2005–2009)
Name of hospital YearMonth
July August September October
All India Institute of Medical Sciences
2005 4.7 – – 4.9
2006 3.7 4.3 2.3 4.5
2007 3.6 3.4 – –
2008 – – 4.2 1.7
2009 – 8.5 5.6 –
Safdarjung Hospital
2005 4.3 – – 5.9
2006 – 3.8 2.0 3.8
2007 3.7 1.5 1.4 –
2008 – – – –
2009 – 9.8 3.8 –
Ram Manohar Lohia Hospital
2005 5.4 – 6.8 3.8
2006 – – 2.1 4.4
2007 5.6 2.4 6.1 –
2008 – – – –
2009 – 23.9 – –
Lady Hardinge Medical College & Associated Hospitals
2005 – – 3.5 –
2006 – – – 4.8
2007 – 6.5 6.7 –
2008 – – – –
2009 – 15.4 – –
Guru Tegh Bahadur Hospital
2005 – 5.0 – –
2006 – – – 3.9
2007 – – 3.1 –
2008 4.8 1.8 – –
2009 – – 12.5 –

Dengue Bulletin – Volume 33, 2009 137
Epidemiological analysis of hospitalized cases of DF/DHF in NCT Delhi, 2005–2009
Entomological surveillance in hospitals
Entomological surveys were conducted to assess the breeding of Ae. aegypti in six major hospitals in the NCT Delhi during the period 2005 to 2009 and the results are shown in Table 4.
Six major hospitals, namely viz. (i) All India Institute of Medical Sciences, (ii) Vardhaman Mahavir Medical College and Hospital (Safdarjung Hospital), (iii) Ram Manohar Lohia Hospital, (iv) Lady Hardinge Medical College and Associated Hospitals (in NDMC zone), (v) Guru Teg Bahadur Hospital (in Shahdara (North) zone) and (vi) Army/Base Hospitals (in Delhi Cantonment zone) were surveyed periodically to detect the breeding of vector species. Breeding of Ae. aegypti was found in all these major hospitals. Persistent breeding was detected in all the hospitals from July to October.
Container indices for July, August, September and October varied from 1.7 to 5.6, 1.6 to 23.9, 1.4 to 12.5 and 1.7 to 5.9 respectively. Major breeding containers were desert coolers, unused containers left in hospital compound, overhead tanks, flower pots and used tyres. Large presence of vectors
in hospitals has already been identified as primary sites for the spatial spread of dengue in Delhi.[8] The present studies highlight that high breeding potential in hospitals still continue to exist.
Discussion
DF/DHF has become endemic with regularity of annual incidence in the NCT region. Breeding of Ae. aegypti in highly vulnerable areas like hospitals continues unabated. Dengue control in the NCT region is based on classical methods of source reduction, larvicidal application in water containers which cannot be emptied, and focal thermal fogging in houses with DHF cases. This is supplemented by health education campaigns through the media to invoke community participation. These efforts apparently seem to have failed to either prevent epidemics or recurrence of disease on annual basis.
Major constraints in the NCT region include multiple health authorities with linked territorial and intersectoral problems. Besides, developmental agencies, namely the Delhi Development Authority (DDA) and the Central Public Works Department (CPWD), work
Name of hospital YearMonth
July August September October
Army/Base Hospital
2005 – 4.5 – 2.5
2006 2.7 – 2.5 2.7
2007 1.7 – – –
2008 – 1.6 – –
2009 – 12.5 1.5 1.9
– Not available CI = container index

138 Dengue Bulletin – Volume 33, 2009
Epidemiological analysis of hospitalized cases of DF/DHF in NCT Delhi, 2005–2009
independently and in isolation and generate a plethora of sites for vector breeding. In addition, the existence of many campuses of several autonomous organizations, namely universities, colleges, chains of hotels and hospitals, industrial estates and commercial complexes, contribute significantly to the build-up of the vector population as these bodies have hardly any skilled manpower or know-how.
Health educational messages to the community are also often incomplete. For example, messages lay a lot of emphasis on cleaning of “evaporation coolers” on a weekly basis, whereas studies in the NCT region have identified several other unconventional sites in the domestic and peri-domestic areas with prolific breeding requiring similar elimination.[15] In view of these findings, there is need for: (i) re-organization of the entire planning process of vector control agencies; (ii) development of intersectoral linkages by roping in all autonomous institutions; and (iii) correct and motivational education messages by informed dialogue for impacting behaviour change of the communities and adoption of evidence-based approaches,
viz. communication for behaviour change (COMBI). Recently SEPA (Socializing Evidence for Participatory Actions) by CIET (Centro de Investigación de Enfermedades Tropicales), a New York-based nongovernmental organization (NGO) has charted a new initiative in this direction. The strategy envisages the involvement of communities right from local/focal research, analysis and building up of control strategies to evaluate vector control by the communities themselves for participatory action (http://www.ciet.org/en/).[16] This model has proved more successful in Nicaragua for control of dengue.
Acknowledgements The authors thankfully acknowledge the guidance from Mr N.L. Kalra, former Deputy Director, NVBDCP, to improve the contents and the manuscript. The most sincere efforts of CCCO staff are highly appreciated. Authors are grateful to Mr J.N. Beniwal, Mr Anil Negi, Mr Raj Kumar, Mr Rakesh Beniwal, Mr Dhirender Singh and Mr V.K. Sood for their contribution in the preparation of figures, tables and the compilation of field data.
ReferencesKumar M, Pasha ST, Mittal V, Rawat DS, Arya [1] SC, Agarwal N, Bhattacharya D, Lal S, Rai Arvind. Unusal emergence of Guate98-like molecular subtype of DEN-3 during 2003 dengue outbreak in Delhi. Dengue Bull 2004,28:161-167.
Saxena P, Parida MM, Dash PK, Santoshi SR, [2] Srivastava A, Tripathi NK, Gupta N, Sahini AK, Bhargava R, Singh CP, Tiwari KN, Sekhar K, Rao PVL. Co-circulation of dengue virus serotypes in Delhi, India. Implication for increased DHF/DSS. Dengue Bull 2006,10:283-287.
Kabra SK, Verma IC, Arora NK, Jain Y, [3] Kalra V. Dengue haemorrhagic fever in children in Delhi. Bull World Health Organ 1992;70(1):105-8.
Kaul SM, Sharma RS, Sharma SN, Panigrahi [4] N, Phukan PK, Lal S. Preventing dengue/dengue haemorrhagic fever outbreaks in the National Capital Territory of Delhi – the role of entomological surveillance. J Commun Dis 1998;30:187-192.

Dengue Bulletin – Volume 33, 2009 139
Epidemiological analysis of hospitalized cases of DF/DHF in NCT Delhi, 2005–2009
Bhattacharya D, Mittal V, Bhardwaj M, Chhabra [5] M, Ichhpujani RL, Lal S. Sero-surveillance in Delhi, India – an early warning signal for timely detection of dengue outbreaks. Dengue Bull 2004;28:207-209.
Kalra NL, Sharma GK. Malaria control in [6] Delhi – past, present and future. J Commun Dis 1987;19(2):91-116.
Paul SD, Dandawate CN, Banerjee K, [7] Krishnamurthy K. Virological and serological studies on an outbreak of dengue-like illness in Visakhapatnam, Andhra Pradesh. Indian J Med Res 1965;53(8):777-89.
Sharma RS, Panigrahi N, Kaul SM. [8] Aedes aegypti prevalence in hospitals and schools, the priority sites for DHF transmission in Delhi, India. Dengue Bull 2001;25:107-108.
World Health Organization, Regional Office [9] for South-East Asia. Prevention and control of dengue and dengue haemorrhagic fever: comprehensive guidelines. WHO Regional Publications, South-East Asia Series No.29. New Delhi: WHO-SEARO, 1999.
World Health Organization. [10] Dengue guidelines for diagnosis, treatment, prevention and control. New Edition. Geneva, WHO, 2009. pp. 116.
Sathish N, Vijayakumar TS, Abraham P, [11] Sridharan G. Dengue fever: its laboratory diagnosis, with special emphasis on IgM detection. Dengue Bull 2003;27:116-125.
World Health Organization, Western Pacific [12] Region. Update on the principles and use of rapid tests in dengue. Prepared by the Malaria, Other Vectorborne and Parasitic Diseases Unit. Manila: WHO-WPRO, 2009.
Healstead SB. Epidemiology of dengue and [13] dengue haemorrhagic fever. In: Gubler DJ, Kuno G. Eds. Dengue and dengue haemorrhagic fever. New York: CAB International, 1997. pp. 23-44.
Srivastava VK, Suri S, Bhasin A, Srivastava [14] L, Bharadwaj M. An epidemic of dengue haemorrhagic fever & dengue shock syndrome in Delhi: A clinical study. Ann Trop Paediatr 1990;10(4):329-34.
Nagpal BN, Srivastava Aruna, Ansari MA, Dash [15] AP. Essentiality of source reduction in both key and amplification breeding containers of Aedes aegypti for control of DF/DHF in Delhi, India. Dengue Bull 2004;28:216-219.
[16] Community intervention trials. Available from http://www.ciet.org/en/documents/methods/2009414105553.asp – accessed 30 June 2010.

140 Dengue Bulletin – Volume 33, 2009
Studies on the efficacy of Toxorhynchites larvae and three larvivorous fish species for the control of
Aedes larval populations in water-storage tanks in the Matale district of Sri Lanka
W.M.G.S. Wijesinghea, M.B. Wickramasingheb, P.H.D. Kusumawathiec, G.A.J.S.K. Jayasooriyac and B.G.D.N.K. De Silvaa#
aDepartment of Zoology, Faculty of Applied Science, University of Sri Jayewardenepura, Sri Lanka
bRetired Entomologist, Anti Malaria Campaign, Colombo 05, Sri Lanka
cRegional Office, Anti Malaria Campaign, Dutugemunu Mawatha, Watapuluwa, Kandy, Sri Lanka
Abstract
A study was conducted to compare the feeding efficacy of Toxorhynchites larvae (L3 & L4) and three larvivorous fish species on Aedes larvae. Ground-level cement water-storage tanks (20%–80%) and water-storing barrels (8.33%–54.55%) formed the majority of Aedes-positive outdoor containers. Ae. albopictus, Ae. macdougali and Ae. vittatus were recorded in water-storage tanks, with Ae. macdougali being dominant.
In the laboratory, the consumption rate (time to devour 10 Ae. albopictus L3 larvae in a vessel of 78.57 cm2 of surface area) for Toxorhynchites was significantly lower (mean time of 330 minutes) than for any of the tested fish species, Poecilia reticulata (Guppy), Puntius bimaculatus (Ipilli Kadaya) and Rasbora caveri (Dandiya), which needed 16.67, 27.33 and 24 minutes respectively. There were no significant differences (P=0.062) between the consumption rates of the three fish species.
A field study was carried out to determine the feeding efficacy of Toxorhynchites larvae, P. reticulata, P. bimaculatus and R. caveri on Aedes larval populations in outdoor cement tanks by noting the percentage reduction of Aedes larvae per 100 cm2 surface area after one week. Toxorhynchites larvae caused a 20%–83.33% reduction with 1–8 larvae per tank. A complete reduction (100%) was achieved with P. bimaculatus and R. caveri with 1–3 fish per tank. P. reticulata showed similar results, but with 90% reduction being achieved once with two fish per tank. There was a higher possibility of losing Tx. larvae than the fish species during the removal of water by the householders. The efficiency of the three fish species for consuming Aedes larvae was greater than that with Tx. larvae. It appears feasible to use Puntius bimaculatus, Rasbora caveri and Poecilia reticulata for controlling Aedes breeding in outdoor cement water-storage tanks in Sri Lanka.
Keywords: Toxorhynchites larvae; larvivorous fish; Aedes control; Sri Lanka.
#E-mail: [email protected]

Dengue Bulletin – Volume 33, 2009 141
Efficacy of Toxorhynchites larvae and larvivorous fish against Aedes larval populations in Sri Lanka
Introduction
Dengue fever (DF) and dengue haemorrhagic fever (DHF) is caused by DENV-1 to 4 serotypes of Flavivirus belonging to the family Flaviviridae. This virus is transmitted by the female mosquitoes of the genus Aedes. Ae. aegypti is the most important epidemic vector while Ae. albopictus, Ae. polynesiensis and Ae. niveus have been incriminated as secondary vectors in some parts of the world.[1]
In Sri Lanka, the first epidemic outbreak of dengue occurred in 1965–1966 during which a few cases of DHF were reported. An outbreak in 1989 caused 203 reported DHF cases with 20 deaths. In recent outbreaks, dengue occurred over a bigger geographical area of the country with multiple serotypes of the virus in circulation. At present, dengue is endemic and an important public health problem in Sri Lanka.[2-3]
Two peaks of dengue fever occur in Sri Lanka annually in conjunction with the south-west monsoon in June-July and the north-east monsoonal rains during October-December. Water collected in man-made containers in domestic and peridomestic environments are important oviposition sites of Ae. aegypti. Ground-level cement water-storage tanks and barrels are reported to be the major larval habitats of Ae. aegypti and Ae. albopictus in both Kandy[4] and Matale districts.[5]
In the absence of a vaccine for the prevention of dengue infection and of a specific treatment for DF/DHF, control of dengue is primarily dependent on the control of Ae. aegypti, the most important vector species, and Ae. albopictus, the secondary vector. Chemical larvicides suitable for use against Aedes breeding in domestic, water-storage tanks, barrels, etc. are extremely limited. Furthermore, frequent use of these
chemicals has the potential of developing vector resistance to insecticides. Thus, greater attention has been paid to the use of biological agents for controlling Aedes breeding in such breeding habitats. Larvivorous fish offer considerable potential for the control of mosquito larvae.[6] The possibility of using Toxorhynchites larvae as a biological agent has also been identified in studies conducted by several investigators.[7] The present study was carried out to compare the efficacy of Toxorhynchites larvae (L3 and L4 stages) and three species of larvivorous fish for the control of Aedes breeding in peridomestic, cement water-storage tanks.
Materials and methods
Study site
The study was conducted in Highlevel gardens, Kaudupelella, in Walliwela Grama Niladhari (GN) division of the Matale district (7° 20’ – 8° 15’ N; 80° 25’ – 81° 00’ W). This area receives piped water originating from a natural water spill. Water is supplied by the Matale Pradeshiya Sabha (urban council). There were frequent interruptions, sometimes for several days, in the supply of unchlorinated water. This encouraged residents to store water for household use in domestic and peridomestic water-storage tanks for several weeks. These storage tanks have been identified as major breeding sites of Ae. aegypti and Ae. albopictus in this area.[8]
Production of Toxorhynchites larvae
Three tyres standing in an upright position and filled to two-thirds of their volume with tap water were kept in vegetated areas. Tyres were inspected after a week for Toxorhynchites larvae. Larvae were identified to species using a standard key.[9]

142 Dengue Bulletin – Volume 33, 2009
Efficacy of Toxorhynchites larvae and larvivorous fish against Aedes larval populations in Sri Lanka
Determination of feeding efficacy of Toxorhynchites larvae and fish in the laboratory
Feeding efficacy tests were carried out under laboratory conditions with three replicates. In each replicate, one L4 Toxorhynchites splendens larva and one Poecilia reticulata, Puntius bimaculatus or Rasbora caveri fish (Figure 1) were placed in separate plastic containers (11 cm height x 10 cm diameter) with well water up to 6 cm deep. They were acclimatized for two hours without food. Then 10 Ae. albopictus larvae were added to each container and the time taken to
consume the introduced larvae was noted. Data were analysed by One-Way ANOVA using Minitab.
Aedes larval surveys to determine larval density in water-storage tanks
Weekly field observations were made from May 2007 to February 2008 in 34 selected houses in the study area to investigate larval breeding in outdoor water-storage containers. During observations, 10 Aedes larvae were collected randomly from each Aedes-positive container using a standard ladle or dropper and placed in separate small plastic bottles. Ten larvae attributed for 100% sensitivity of Aedes surveys[10] were collected. If a particular container had less than 10 larvae, all larvae were collected. Larval identification was carried out using standard keys[9] and larval density was expressed by calculating the House Index (HI) (percentage of houses positive for Ae. aegypti/Ae. albopictus), Container Index (CI) (percentage of containers positive for Ae. aegypti/Ae. albopictus) and Breteau Index (BI) (number of containers positive for Ae. aegypti/Ae. albopictus per 100 houses).[11]
Application of Toxorhynchites larvae and fish species to the water-storage tanks
Aedes larval density per 100 cm2 of surface area of each tank was noted before application of Toxorhynchites larvae or fish (P. reticulata, P. bimaculatus and R. caveri). Toxorhynchites splendens and Tx. minimus L3 and L4 larvae were added to Aedes-positive tanks at the rate of 1–4, 6 and 8 larvae per tank, with 1–3 fish per tank. Aedes larval density per 100 cm2 of surface area of each tank was noted one week after application. Three replicates were carried out for each test.
Figure 1: Fish species used in feeding efficacy tests
Puntius bimaculatus
Rasbora caveri
Poecilia reticulata

Dengue Bulletin – Volume 33, 2009 143
Efficacy of Toxorhynchites larvae and larvivorous fish against Aedes larval populations in Sri Lanka
Results
The mean time taken by Toxorhynchites larvae to consume 10 Aedes albopictus larvae was 330.0 minutes. Mean times for fish to consume 10 Ae. albopictus larvae were: Poecilia reticulate – 16.66 minutes, Puntius bimaculatus – 27.33 minutes, and Rasbora caveri – 24 minutes (Table 1). There was no significant difference (P=0.062) between the mean time taken to consume 10 larvae by the three fish species.
Table 1: Time taken to consume 10 Aedes albopictus larvae by each species of
Toxorhynchites larvae and three species of fish
Species
Time taken to consume 10 Ae. albopictus larvae (minutes)
1 2 3 Mean (minutes)
Toxorhynchites splendens larvae
360 300 330 330.00
Poecilia reticulata 15 14 21 16.67
Puntius bimaculatus
25 25 32 27.33
Rasbora caveri 22 20 30 24.00
Potential outdoor breeding habitats of Aedes species included ground-level cement water-storage tanks, barrels (plastic and metal), plastic buckets, plastic cans, aluminium pots, clay pots, metal pots and coconut shells. Ground-level cement water-storage tanks (20.00%–80.00%) and barrels (8.33%–54.55%) were the major contributors to Aedes-positive outdoor containers in the study area. Plastic buckets (0.00%–33.33%), plastic cans (0.00%–33.33%), aluminium pots (0.00%–12.50%), clay pots (0.00%–16.67%) and metal pots (0.00%–9.52%) were the other important breeding habitats.
Three Aedes species (Ae. macdougali, Ae. albopictus and Ae. vittatus) were found breeding in cement tanks. Ae. macdougali was the dominant species with a contribution of 61.61% of the total collection. Ae. albopictus contributed 37.79% while Ae. vittatus contributed 0.59% to the total collection (Table 2). The selected study area was primarily semi-urban in nature, with more potential breeding sites suitable for Ae. albopictus. All Ae. aegypti breeding sites were treated with Abate by government authorities. These factors contributed to the absence of Ae. aegypti.
Table 2: Species composition of Aedes larvae in water-storage cement tanks
in the study area
Aedes species Number of larvae
Percentage of larvae
Ae. macdougali 724 61.61
Ae. albopictus 444 37.79
Ae. vittatus 7 0.59
Total 1175 100
A House Index (HI) greater than 5% was obtained for Aedes species, including Ae. albopictus, in all 29 field visits. A Container Index (CI) greater than 20% was recorded for Ae. albopictus on seven occasions, and for all Aedes species on 18 occasions.
There was no relationship between the number of Toxorhynchites larvae applied and the reduction of Aedes larvae per 100 cm2
of surface area. In tanks with Toxorhynchites added at the rate of 1, 2, 3, 4, 6 and 8 larvae per tank, 20.00%–83.33% reduction of Aedes larval density was observed. Although a considerable reduction of larvae per tank was obtained, 100% reduction was not achieved even with eight Toxorhynchites larvae (Figures 2 and 4). In the P. reticulata tanks at

144 Dengue Bulletin – Volume 33, 2009
Efficacy of Toxorhynchites larvae and larvivorous fish against Aedes larval populations in Sri Lanka
the rate of 1, 2 and 3 fish per tank, there was a 100% reduction of Aedes larvae, except on one occasion in tanks with two fish, when it was 90% (Figures 3 and 4). A 100% reduction of Aedes larvae was also observed with the application of 1, 2 or 3 Puntius bimaculatus (Ipilli Kadaya) and Rasbora caveri (Dandiya) (Figures 3 and 4).
Figure 2: Reduction of Aedes larval populations using Toxorhynchites larvae
Figure 3: Reduction of Aedes larval populations using three fish species
Figure 4: Percentage reduction of Aedes larval population by each bio-control agent

Dengue Bulletin – Volume 33, 2009 145
Efficacy of Toxorhynchites larvae and larvivorous fish against Aedes larval populations in Sri Lanka
Discussion
Water storage is a common household practice in areas with irregular water supply. In our study area, residents store water in cement tanks and barrels for domestic and peridomestic use due to uncertainty of water supply. These tanks were rarely cleaned at weekly intervals, thus providing ideal developmental sites for both Aedes and other container-breeding mosquitoes. During the present study, a high percentage of ground-level cement water-storage tanks (20%–80%) contained Aedes larvae. Cement water-storage tanks also have been reported as important breeding habitats of Ae. aegypti and Ae. albopictus in dengue transmission areas in Matale district,[5] Kandy and Nuwara Eliya districts[4,10,12,13,14] and in Tangalle.[15]
The presence of Ae. macdougali was reported in the Suduganga area of Matale district[8] and we found Ae. macdougali in cement tanks in high density (61.61% of the total collection) and sharing the habitat with Ae. albopictus and Ae. vittatus. The sharing of water-storage tanks by Ae. macdougali, Ae. aegypti, Ae. albopictus and other non-Aedes species was reported in other studies.[16]
Our data suggest that cement water-storage tanks are important breeding sites of dengue vectors (Ae. aegypti and Ae. albopictus) and other nuisance mosquitoes. Thus, mosquito control in these tanks would help to prevent/control both dengue and mosquito nuisance. The development and presence of other mosquito species along with Ae. aegypti and Ae. albopictus habitats require greater care to be taken during larval collections, and identification and calculation of relevant larval indices. The collection of 10 randomly selected Aedes larvae (or all larvae if the container had <10 larvae) has been adopted for 100% sensitivity of dengue vector surveillance.[10]
Toma and Miyag i repor ted the consumption of 20–26 larvae per day by 1st and 2nd instar Tx. splendens with an average consumption of 389±26 larvae during their entire larval lifespan.[17] However, the success of using Toxorhynchites in Aedes larval control in field settings is controversial.[7] In Malaysia, 2nd instar larvae of Tx. splendens have been used successfully to control Ae. aegypti and Ae. albopictus breeding in domestic water containers,[18] although in Java, Indonesia, introduction of Tx. amboinensis larvae to control Ae. aegypti and Ae. albopictus in the same type of habitats was less successful.[19] We observed that Toxorhynchites larvae only gave 20%–80% reduction of Aedes larval density in the water-storage tanks.
Complete reduction of Aedes larvae was obtained with 1–3 fish of P. reticulata, P. bimaculatus and R. caveri except on one occasion with two P. reticulata per tank in which case it was 90%. Puntius bimaculatus (an endemic species) and Rasbora caveri (a common fresh-water fish in Sri Lanka) are reported to have shown larvivorous activity.[20] Thus, the use of Puntius bimaculatus, Rasbora caveri and Poecilia is recommended for Aedes mosquito control in water-storage tanks in this study area. Danio malabaricus, Oreochromis mossambicus, O. niloticus and Poecilia reticulata are reported to be good larvivorous fish species for anopheline mosquito control.[16] Thus, further studies on the effectiveness of different fish species for Aedes larval control in water-storage tanks in different eco-epidemiological areas are warranted. Such studies would help to identify the most suitable species of fish for a particular eco-epidemiological area as some fish species do not survive well in some areas. Poecilia reticulata is now considered an invasive species and thus great care should be taken when it is applied to natural waters[21] considering the ecology and the epidemiological conditions.

146 Dengue Bulletin – Volume 33, 2009
Efficacy of Toxorhynchites larvae and larvivorous fish against Aedes larval populations in Sri Lanka
Two species of Toxorhynchites (splendens and minimus) have been recorded from Sri Lanka[8,22] and Tx. minimus in the Suduganga area in Matale.[8] During the present study, natural breeding of Tx. splendens was observed in a water-filled barrel in the study area. This indicates the possibility of establishing Tx. splendens and Tx. minimus as part of an integrated vector control programme.
Acknowledgements
The authors wish to thank Dr Ajantha de Alwis, Department of Zoology, University of Sri Jayewardenepura, for helping in the identification of fish species, and Mr P. Dias, Department of Mathematics, University of Sri Jayewardenepura, for his support in the statistical analysis of data.
ReferencesP[1] revention and control of dengue and dengue haemorrhagic fever - comprehensive guidelines. New Delhi: WHO Regional Office for the South-East Asia, 1999. pp. 29; 134.
Kulatilaka TA, Jayakuru WS. Control of dengue/[2] dengue haemorrhagic fever in Sri Lanka. Dengue Bull 1998;22:53-59.
[3] External in-depth review of DF/DHF control programme in Sri Lanka. New Delhi: WHO Regional Office for South-East Asia, 2005. pp. 75.
Kusumawathie PHD, Wickremasinghe [4] AR, Karunaweera ND, Wijeyaratne MJS. Larvivorous potential of fish species found in river bed pools below the major dams in Sri Lanka: J Med Entomol 2006;43(1):79-82.
Yapabandara AMGM, Bandara DMLS. [5] Entomological and sociological investigations in dengue transmission areas in the Matale District. Sri Lanka Association for the Advancement of Science, Proceedings of 62nd annual session, Sri Lanka, 2006. pp.1.
Wickramasinghe MB, Costa HH. Mosquito [6] control with larvivorous fish. Parasitology Today 1986;2(8):228-230.
Collins LE, Blackwell A. The biology of [7] Toxorhynchites mosquitoes and their potential as biocontrol agents. Biocontr News Info 2000;21:105N-116N.
Yapabandara AMGM, Abeykoon AR. [8] Aedes aegypti and Aedes albopictus breeding sites in the Matale Municipal council area. Sri Lanka Association for the Advancement of Science, proceedings of 58th annual sessions, Sri Lanka, 2002. pp. 20.
Sewell RBS, Edward FW. [9] Fauna of British India, including Ceylon & Burma, order: diptera. 1st edition, New Delhi: Today & Tomorrows Printers & Publishers, 1934.
Kusumawathie PHD, Jayasooriya GAJSK, [10] Wickremasinghe R. Sensitivity of different larval collection methods in dengue vector surveillance in the Kandy and Nuwara Eliya districts. Sri Lanka Association for the Advancement of Science: proceedings of the 62nd annual sessions. 2006. pp. 1-2.
Pant CP, Self LS. [11] Vector ecology and bionomics. Monograph on dengue/dengue haemorrhagic fever. WHO Regional Publication, SEARO No 22. New Delhi: WHO-SEARO, 1993. pp. 121-138.
Kusumawathie PHD, Jayasooriya GAJSK, [12] Wickremasinghe AR. Most productive and prevalent breeding habitats of Aedes aegypti and Ae. albopictus (pupae) in the Kandy and Nuwara Eliya districts of Sri Lanka. Sri Lanka Association for the Advancement of Science, Proceedings of the 63 rd Annual Sessions, Sri Lanka 2007. pp. 7-8.

Dengue Bulletin – Volume 33, 2009 147
Efficacy of Toxorhynchites larvae and larvivorous fish against Aedes larval populations in Sri Lanka
Kusumawathie PHD, Wickremasinghe AR, [13] Jayasooriya GAJSK. Disease transmitting mosquito species breeding in water storage tanks in Kandy, Sri Lanka. Sri Lanka Association for the Advancement of Science, Proceedings of the 61st Annual Sessions 2005. pp. 8.
Kusumawathie PHD, Siyambalagoda RRMLR. [14] Distribution and breeding sites of potential dengue vectors in Kandy and Nuwara Eliya districts of Sri Lanka: Ceylon Med J 2005;48(2):43-52.
Ep idemic o f dengue f eve r /dengue [15] haemorrhagic fever in Tangalle. Weekly Epidemiological Report Sri Lanka Ministry of Health 2001;29(50):1-4.
Kusumawathie PHD, Wickramasinghe AR, [16] Karunaweera ND, Wijeyaratne MJS. Use of Sri Lankan larvivorous fish species in mosquito control. In: Karunaratne SHP. Eds. Programme & proceedings national symposium on mosquito control. Colombo: Post Graduate Institute of Science, University of Peradeniya and National Science Foundation, 2005. pp. 58-69.
Toma T, Miyagi I. Laboratory evaluation of [17] Toxorhynchites splendens (Diptera: Culicidae) for predation of Aedes albopictus mosquito larvae. Med Vet Entomol 1992:6;281-289.
Chuah MLK, Yap HH. Studies on biological [18] control potent ia ls of Toxorhynchites splendens (Diptera: Culicidae). Trop Biomed 1984;1:145-150.
Annis B, Nalim S, Widiarthi H, Boewono DT. [19] Toxorhynchites amboinensis larvae released in domestic containers fail to control dengue vectors in a rural village in central Java. J Am Mosq Contr Assoc 1990;6:75-78.
Pethiyagoda R. [20] Fresh water fishes of Sri Lanka. Colombo: Wildlife Heritage Trust of Sri Lanka, Sarvodaya Vishva Lekha, 1991.
Shirantha RRAR, Amarathunga AAD, [21] Weerasekara KAWS. Do feral populations of Poecilia reticulata ready to bring its overall efficacy as a mosquito bio-control agent or pose threats to aquatic biodiversity in Sri Lanka. In: The proceeding of National Symposium on Invasive Alien Species (IAS 2008), Sri Lanka Association for the Advancement of Science (Section D), National Science Foundation and Biodiversity Secretariat, Colombo 7, 2008. pp. 8.
Jayasekera N, Chelliah RV. [22] An annotated checklist of mosquitoes of Sri Lanka UNESCO: man and the biosphere national committee of Sri Lanka. Publication No. 8, 1981. pp. 16.

148 Dengue Bulletin – Volume 33, 2009
A novel method of controlling a dengue mosquito vector, Aedes aegypti (Diptera: Culicidae) using an aquatic mosquito predator, Diplonychus indicus
(Hemiptera: Belostomatidae) in tyresN. Sivagnaname#
Division of Vector Biology and Control, Vector Control Research Centre, Indian Council of Medical Research, Indira Nagar, Puducherry 605 006, India
Abstract
An investigation was undertaken to evaluate the biocontrol potential of a mosquito predator, Diplonychus indicus, against the immatures of dengue vectors breeding in used tyres. Two gardens were selected and ten tyres each filled with water were kept in different sites of both gardens. One was considered as “control” where no predator was used, whereas in the tyres kept in the “experimental” garden, two second instars of D. indicus were released into each tyre. The absolute density of larvae, pupae and man-landing rate of dengue vector were monitored at weekly intervals. After one year, the predators released in the tyres in the experimental garden were removed and it became the control garden whereas two second instars predators were released into the tyres kept in the control garden which then became the experimental garden. When D. indicus was introduced into the tyres kept in the experimental garden, a drastic reduction of 95% and 98% respectively of late instars and pupae of Ae. aegypti was observed. But there was no reduction in the tyres kept in the control garden. The density of early instars of Ae. aegypti fluctuated in the tyres kept in both the gardens during different months of the year. There were significant differences between the densities of early, late instars and pupae of Ae. aegypti in experimental and control gardens (early: p<0.0001; late: p<0.0001; pupae: p<0.0001; df=47). A similar trend was observed even when experimental and control gardens were interchanged during the second round (early: p<0.1; late: p<0.0001; pupae: p<0.0001; df=47). This field trial indicated that D. indicus can be used successfully in reducing the immature stages of Ae. aegypti followed by suppression in adult emergence. This is the first report on the utility of this mosquito predator against dengue vectors in the field.
Keywords: Diplonychus indicus; Aedes aegypti; aquatic mosquito predator; biological control.
#E-mail: [email protected]

Dengue Bulletin – Volume 33, 2009 149
Control of Ae. aegypti breeding in tyres using hemipteran bug – D. indicus
Introduction
Aedes aegypti is the proven and efficient vector of dreadful viral diseases such as dengue and dengue haemorrhagic fever.[1-2] According to a new estimate, 3.6 billion (55% of the world’s population) are at risk of dengue in 124 endemic countries with an estimated 21 000 deaths every year.[3] In the absence of a vaccine and effective drugs, vector control has been considered an important tool in the prevention and control of these diseases. Ae. aegypti breeds in a variety of habitats such as cement tanks, cisterns, tree holes, tyres, mud pots, drums, disposed plastic cups, coconut shells, etc., many of which are inaccessible for larviciding by chemical means.[4-6] Control of the adult vector mosquito is also difficult because of inadequate understanding of its resting behaviour.[7]
The operational utility of biological control methods using bacterial pathogens such as Bacillus thuringiengis var. israelensis and B. sphaericus is not considered for the containment of Aedes immatures because of the high costs involved due to the frequent treatments that are necessary.[8] Other biocontrol agents such as larvivorous fish could be used in wells, water storage containers and cisterns, but their release is unacceptable to the inhabitants and the fish also die due to chlorination during cholera outbreaks.[9] At present, there are no promising biological control agents specific to Ae. aegypti and Ae. albopictus though there are certain instances of successful field trials with cyclopoid copepods.[10-12]
In recent years, the “attract-and-kill method” has drawn attention elsewhere for the control of dengue vectors.[12-17] The rationale is to divert ovipositing females of Ae. aegypti to artificial habitats and destroy the larvae either by emptying, or stocking a predator. It has been well-demonstrated
that Ae. aegypti prefers automobile tyres to any other container for ovipositioning.[6,18-21] Though different species of Toxorhynchites were considered as efficient predators and were exploited for the control of Ae. aegypti, their use in control programmes had met with many setbacks. Several physical and biological factors in nature, variation in the age distribution of the predator and the prey within the habitat and other structural complexities in nature affect the predator-prey interactions.[8,22] Hence, a potential alternative for the control of immatures of Ae. aegypti by biological means is being envisaged.
Diplonychus indicus , an aquat ic hemipteran bug, has been reported to be an efficient predator of mosquito immature in many laboratory studies.[23-26] The advantage of D. indicus over cyclopoid copepods and other mosquito predators is its preferential selection for larger prey, its availability in many of the aquatic habitats, and its tolerance to chlorination, insecticides and biocides.[27-30] Recently, in Australia, the predatorial capacity of Diplonychus sp. has been reported in the natural habitats of An. annulirostris.[31] Though there have been several studies on the predatory potential of D. indicus, there has been no information on the use of this biocontrol agent in the field. The present trial was carried out to explore the utility of D. indicus for the control of Ae. aegypti by introducing it into tyre traps in field conditions.
Materials and methods
Description of study area
A field trial was carried out in Puducherry (formerly known as Pondicherry), a coastal town in South India. It is located between 11° 46’N and 12° 15’N latitude and between

150 Dengue Bulletin – Volume 33, 2009
Control of Ae. aegypti breeding in tyres using hemipteran bug – D. indicus
79° 36’E and 79° 53’E longitude. This region is a flat terrain of an average elevation of about 15 metres above the mean sea level. Tropical climatic conditions without drastic fluctuations in temperature prevail in this region.
Ae. aegypti, the vector of many arboviral diseases including dengue, has been reported to breed in relatively clean rain water collected in containers, cement cisterns, tyre dumps, tree holes, wells, etc.[4,32] It is also reported to breed in septic tanks containing highly polluted water.[33]*1 The dengue vectors have been frequently observed to breed in a variety of other peridomestic habitats such as coconut shells, mud pots and undestroyed disposable plastic cups in and around houses and other places of human agglomeration, especially during the post-monsoon season, which results in difficulties in designing an appropriate control strategy against them.
Study design
The study period lasted for 28 months from September 1994 to December 1996. Preliminary testing for homogeneity between the two selected gardens lasted for four monsoon months (Sept.–Dec. 1994). These gardens were selected to identify the natural breeding habitats of Ae. aegypti. One garden (12 acres) is situated in Uppalam, 0.5 km away from Puducherry town and the other garden (10 acres) is located in Natesan Nagar, which is about 1.5 km away. In both gardens, the major vegetations were mango, papaya, sappotta (edible-fruit yielding) and coconut trees surrounded by human dwellings. These gardens belong to the Aurobindo Ashram, Puducherry, where different kinds of vegetables, flowers and paddy are also cultivated.
* This is an example of extreme ecological compulsion – Editor.
All the breeding sources of this vector such as tree holes, containers, cement tanks, coconut shells and other sources such as disposed plastic cups, broken mud pots, etc., were screened during the rainy months (Sept.–Nov.). The water content of the discarded containers and coconut shells was poured directly into an enamel tray. A siphon was used to collect water from tree holes. An enamel dipper of 300 ml capacity was used to sample water in cement tanks. A strainer and muslin cloth (pore size <0.5 mm) were used to strain the mosquito immatures collected from these habitats. The total of immature stages (larvae + pupae) of dengue vectors were counted and recorded. Sampling was done at weekly intervals for a period of three months from Sept.–Dec. 1994. Samples of pupae were brought to the laboratory to identify the species (n=10).
Introduction of predators and monitoring of immatures of Ae. aegypti in trap tyres
During January 1995, two second instars of D. indicus were introduced into each tyre kept in the experimental garden (at Uppalam). A constant number of predators (2/tyre) was maintained throughout the study period and no additional predators were introduced even though certain tyres supported more density of mosquito immatures. Among the five stages of D. indicus, only the second-instars predator was selected owing to its high attack rate and short handling time.[34] Only two predators were released (instead of many) so as to avoid cannibalism and competition among the predators in such a confined artificial habitat like tyres. The duration required for the second instars of the bug to become an adult is about 50–60 days and so new predators were replenished once in two months as the adult bugs flew away for mating.[35,36] No predators were introduced into the tyres kept in the control garden.

Dengue Bulletin – Volume 33, 2009 151
Control of Ae. aegypti breeding in tyres using hemipteran bug – D. indicus
In 1996, during early January, the predators kept in the experimental garden (at Uppalam) were removed and it became the control garden while two predators were introduced into each tyre kept in the control garden which now became the experimental garden. In this study, the tyres kept in the gardens were the real sampling units and not the gardens as such. The exchange of control and treatment (experimental) during the second year of the study did not have any residual effect in terms of immature and adult densities of Ae. aegypti. The tyres were cleaned out followed by the changing of water before the switching of gardens so as to avoid the chemical cues associated with predation. The effect of other natural mortality factors was ruled out in both the control and experimental gardens. During the whole study period, no other natural predators such as notonecta, damselfly nymphs, etc. or diseased mosquito larvae were encountered in the tyres.
The absolute density of immature stages and man-landing rate of Ae. aegypti were monitored in both experimental and control gardens. The immatures were categorized as early (I and II instars), late (III and IV instars) and pupae. This was done to understand the feeding preference of the predator. Man-landing collection of mosquitoes was also carried out for 15 minutes by partially exposing the hands and legs of the author of this study. The man-landing mosquitoes were carefully collected before they could bite by using a mechanical aspirator.
Statistical analysisOne-way ANOVA was performed by using SPSS (version -16) to find out the variations in the density of the immature stages and pupae of dengue vectors between the control and experimental gardens in different months of each year. Independent t-test was applied to
find out the significant difference between the mean densities of mosquito early instars, late instars, pupae and landing rate of adult females of Ae. aegypti in the control and experimental gardens. The level of significance was taken as p<0.05.
Results The important mosquito species breeding in different habitats of the gardens were identified as Ae. aegypti (89%) and Ae. albopictus (11%) out of the 129 pupae kept for emergence. The major breeding habitats of these vector mosquitoes in both the gardens were tree holes, containers, cement tanks and coconut shells. There was a significant difference in the mean numbers of larvae and pupae in different breeding habitats between the two gardens (F=27.79; p<0.001). Some particular types of containers supported relatively high breeding of Ae. aegypti than the other breeding habitats (22–39 larvae/container/week) in both gardens (Table 1).
The introduction of tyres in both gardens resulted in significant reduction in the densities of immatures in other natural habitats. The tyres received 2–3 times more ovipositing females of Ae. aegypti than the naturally available habitats during the field trial (Tables 2 and 3). In both gardens, the density of mosquito immatures in tyres was significantly higher than in other naturally available breeding habitats. (Garden 1: F=36.41; p<0.0001, Garden 2: F=27.79; p<0.0001).
When D. indicus was introduced into the tyres kept in the experimental garden in 1995, a drastic reduction (95%–98%) of late instars and pupae of Ae. aegypti was observed. But, there was no such reduction in the late and pupal stages of the vector mosquito in the tyres kept in the control garden (Figures 1a, 1b and 1c). Though there was a significant reduction

152 Dengue Bulletin – Volume 33, 2009
Control of Ae. aegypti breeding in tyres using hemipteran bug – D. indicus
of late instars and pupae in the tyres kept in the experimental garden, the early instars showed no such decline in both experimental and control gardens (Figure 1a).
There were significant differences between and among the immature stages of Ae. aegypti in the control and experimental gardens (Table 4). The same trend was exhibited even when the experimental garden became the control garden in the subsequent year. Mean densities of early instars, late instars and pupae when the control and experimental gardens were interchanged are shown in Figures 2a, 2b and 2c.
There were significant differences between the densities of early, late instars and pupae of Ae. aegypti in experimental and control gardens (Tables 4 and 5). The absolute density of pupae in the tyres kept in the experimental garden began to decline immediately within a week following the introduction of the predators. But there was no such reduction in the tyres kept in the control garden where the predators were not introduced. However, the early immature populations fluctuated in all the tyres kept in both experimental and control gardens during different months of the year.
Table 1: Mean density (±SE) of immatures of Ae. aegypti and per cent availability of different breeding habitats before the introduction of tyres (n=10)
Habitats Uppalam Garden 1 % availability Natesan Nagar
Garden 2 % availability
Tree holes 23.64±2.86 25.3 14.7±2.39 20.5
Containers 22.03±2.91 37.8 39.3±3.99 38.6
Cement tanks 16.40±2.19 16.4 11.6±2.15 18.4
Coconut shells 10.42±1.23 12.7 7.9±1.42 13.9
Other sources 5.30±1.28 18.8 10.9±2.09 8.6
Table 2: Mean density (±SE) of immatures of Ae. aegypti in different breeding habitats after introduction of tyres in Garden 2 at Uppalam (n=10)
Habitats Mean±SE Comparison T
Tree holes 10.0±0.69 1x6 6.26*
Containers 9.8±0.96 2x6 5.93*
Cement tank 6.7±1.41 3x6 6.47*
Coconut shells 5.5±1.27 4x6 6.81*
Other sources 3.4±0.87 5x6 7.46*
Tyre 35.3±4.18
*Significant

Dengue Bulletin – Volume 33, 2009 153
Control of Ae. aegypti breeding in tyres using hemipteran bug – D. indicus
Discussion
Source reduction and non-chemical means are the well-known methods advocated for the control of Ae. aegypti through community participation.[36] However, during the rainy season when there is an increase in the number of containers in and around human dwellings, it is difficult to go for source reduction and it would be cumbersome for frequent emptying of tyres and other innumerable breeding sources of Ae. aegypti and Ae. albopictus.[37] Further, there is evidence of failure of source reduction practices for controlling this container breeder due to the reluctance of some householders to discard the most potential mosquito-producing containers. [38] Attracting the ovipositing females of Ae. aegypti to a particular
habitat and controlling them by introducing a biocontrol agent has been attempted in such situations.[9,39-40]
D. indicus is considered an efficient predator even at high density, unlike the backswimmers, Notonecta undualta and Anisops bouverii, where the predation rate is known to diminish from low- to high-prey density. The predator came to the level of satiation only after consuming a large number of mosquito larvae (10–56/hour) unlike the other aquatic predators like Notonecta undulata and Anisops bouverii which consume only 10–20 prey/hour.[41-42]
D. indicus is more efficient than copepods that are currently advocated for the control
Table 3: Mean density (±SE) of immature of Ae. aegypti in different breeding habitats after introduction of tyres in Garden 2 at Natesan Nagar (n=10)
Habitats Mean±SE Comparison T
Tree holes 9.8±1.28 1x6 5.58*
Containers 14.5±1.69 2x6 4.69*
Cement tank 6.1±2.04 3x6 5.99*
Coconut shells 6.3±1.43 4x6 6.14*
Other sources 4.3±1.32 5x6 6.51*
Tyre 42.5±5.72
*Significant
Table 4: Comparison of mean density of immature stages of Ae. aegypti between control and experimental gardens during 1995 and their significance
Immature stagesMean (±SE)
F SignificanceControl Experimental
Early instars 38.98±2.23 21.61±1.18 47.57 0.000
Late instars 16.47±0.83 3.55±0.34 209.94 0.000
Pupae 13.09±0.54 0.36±0.06 554.33 0.000

154 Dengue Bulletin – Volume 33, 2009
Control of Ae. aegypti breeding in tyres using hemipteran bug – D. indicus
Figure 1: Monthly mean densities of immature in control and experimental gardens during 1995
(1a)
(1b)

Dengue Bulletin – Volume 33, 2009 155
Control of Ae. aegypti breeding in tyres using hemipteran bug – D. indicus
(1c)
Figure 2: Monthly mean densities of immature in control and experimental gardens during 1996
(2a)

156 Dengue Bulletin – Volume 33, 2009
Control of Ae. aegypti breeding in tyres using hemipteran bug – D. indicus
(2b)
(2c)

Dengue Bulletin – Volume 33, 2009 157
Control of Ae. aegypti breeding in tyres using hemipteran bug – D. indicus
of Ae. aegypti and Ae. albopictus breeding in phytotelmata including discarded automobile tyres. Cyclops prefer only newly-hatched mosquito larvae and their relative predatory efficiency was less in terms of reduction in immature density of dengue vectors.[43-44] Mean number of Ae. aegypti larvae predated by Mesocyclops leukarti was only 10–15/24 hours[41] when compared with 10–56/hour by D. indicus (VCRC, unpublished report).
In Viet Nam, a Mesocyclops spp. has been reported to devour 16–41 Ae. aegypti larvae/day,[45] but D. indicus could kill >200 larvae/day (VCRC, unpublished report). Killing of more prey could be possible by D. indicus as it feeds on only the soft tissues (not the whole prey) by injecting digestive enzymes into the prey. Further, it could have a high attack rate and less handling time resulting in a high predation rate when compared with other aquatic predators such as predatory fish and Toxorhynchites splendens.[46] More importantly, the cyclopoid copepods seldom predate on larger instars of prey and this may result in pupation followed by emergence of vector mosquitoes.[12]
A study on the biocontrol potential of Macrocyclops albidus revealed that nearly two months were required for the control of immatures of Ae. albopictus in tyre piles.[13] But the release of D. indicus in tyres resulted in a drastic decline in the pupation of Ae. aegypti within a week’s time, with subsequent reduction in adult emergence. Leaf litter in tyres did not affect the predatory potential of D. indicus because of its active foraging behaviour, both on the surface and within the water medium in tyres. The duration of time required for the second instars of the bug to become an adult is about 50–60 days.
The natural habitats of many mosquito predators including D. indicus do not overlap
largely in the field, especially with respect to Ae. aegypti.[47] But the present field trial with D. indicus has indicated that it could be exploited for the control of dengue vectors in tyres though the natural home of this mosquito predator is different. This finding corroborates with the findings related to the biocontrol potential of dragonfly nymphs, Crocothemis servilia, against Ae. aegypti breeding in different types of containers in and around domestic locations which offered a positive impulse on the utility of this mosquito predator though it has different natural homes.[48-49] It also seems to be more ethical as this “new association approach” is helpful for the control of dengue vectors.[47] Further, it has been rightly pointed out that “for artificial containers and some other habitats lacking predators, artificial introduction of local predators can be an option in mosquito control”.[50]
As D. indicus preferred predominantly late instars of the prey population, the emergence of pupae can be easily prevented, which would subsequently reduce the biting density.[29] The present study is an evidence for 98% reduction in pupal density of Ae. aegypti due to the introduction of D. indicus in tyres. This specific reduction may be due to the preferential feeding behaviour of D. indicus against late instars of mosquito prey as supported by certain laboratory studies.[29,51] Similar reduction of Aedes larvae has also been obtained in containers containing most effective species of Cyclops.[11]
ConclusionIn conclusion, the results of this trial have demonstrated the predatory potential of D. indicus in controlling the dengue vectors using tyres. Further study should also be undertaken to assess its efficacy in urban situations where the elimination of the breeding

158 Dengue Bulletin – Volume 33, 2009
Control of Ae. aegypti breeding in tyres using hemipteran bug – D. indicus
sources of the dengue vector is problematic. Health education and demonstration of its efficacy can also attract the awareness and enthusiastic participation of the local community to use this predator against dengue vectors.
Acknowledgements
The author is thankful to the field assistance rendered by Mr A. Nandakumar, Mr K. Sathianathan and Mrs Jessu Mathew, Vector
Biology and Control, Vector Control Research Centre, Puducherry. The author is grateful to Dr M. Kalyanasundaram, then Officer-in-Charge, Vector Control Research Centre, Puducherry, for his keen perusal and comments on the manuscript. The constant encouragement and guidance offered by Dr P.K. Das, former Director, Vector Control Research Centre, Puducherry, is greatly appreciated. Help in statistical analysis provided by Dr P. Vanamail and in literature survey by Mrs R. Sundarammal, Library and Information Officer, VCRC, is gratefully acknowledged.
ReferencesGubler DJ. Dengue and dengue hemorrhagic [1] fever. Clin Microbiol Rev 1998;11:480-496.
Gibbons RV, Vaughn DW. Dengue: an escalating [2] problem. BMJ 2002;324:1563-1566.
Pediatric Dengue Vaccine Initiative. Global [3] burden of dengue. Kwanal. http://www.pdvi.org/about_dengue/GBD.asp - accessed 01 July 2010.
Panicker KN, Rajagopalan PK. A note on tree [4] hole breeding mosquitoes in Pondicherry town. Indian J Med Res 1978;68:610-13.
Mahadev PV, Fulmali PV, Mishra AC. A [5] preliminary study of multilevel geographic distribution of prevalence of Aedes aegypti (Diptera: Culicidae) in the state of Goa, India. Indian J Med Res 2004;120:173-82.
Laojana T, Tauthong P, Phanthumachinda B. [6] Surface preference for oviposition of Aedes aegypti in Thailand. Southeast Asian J Trop Med Public Health 1981;12:209-212.
Bang YH, Tonn RJ. Vector Control and [7] Intervention. In: Monograph on Dengue/Dengue Hemorrhagic Fever. Regional publication, SEARO No: 22. Chapter 9: 139-160. New Delhi: WHO Regional Office for South-East Asia, 1993.
Se rv i ce MW. B io log i ca l con t ro l o f [8] mosquitoes – Has it a future? Mosq News 1983;3:113-115.
Santos LU, Andrade CFS, Carvalho GA. [9] Biological control of Aedes albopictus (Diptera: Culicidae) by Mesocyclops longisetus (Copepoda: Cyclopoida) in two field trials. Mem Inst Oswaldo Cruz 1996;91:161-162.
Lounibos LP, Frank JN, Allison ML, Ocanto [10] P, Navarro JC. Survival, development and predatory effects of mosquito larvae in Venezulean phytotelmata. J Trop Ecol 1987;3:221-242.
Marten GG, Reid JW. Cyclopoid copepods. [11] J Am Mosq Control Assoc 2007;23(2 Suppl):65-92.
Vu SN, Yen NT, Kay BH, Marten GG and [12] Reid JW. Eradication of Aedes aegypti from a village in Vietnam, using copepods and community participation. Am J Trop Med Hyg 1998;59:657-660.
Marten GG. Issues in the development of [13] cyclops for mosquito control. Arbovirus Res Australia 1989;5:159-164.
Marten GG. Elimination of [14] Aedes albopictus from the tyre piles by introducing Macrocyclops albides (Copepoda: Cyclopoidae). J Am Mosq Control Assoc 1990;6:689-693.

Dengue Bulletin – Volume 33, 2009 159
Control of Ae. aegypti breeding in tyres using hemipteran bug – D. indicus
Brown MD, Kay BH, Hendrikz, JK. Evaluation [15] of Australian Mesocyclops (Cyclopoda: Cyclopidae) for mosquito control. J Med Entomol 1991;28:618-623.
Marten GG, Borjas G, Cush M, Fernandez E, [16] Reid JW. Control of larval Aedes aegypti by cyclopoid copepoids in peridomestic breeding containers. J Med Entomol 1994;31:36-43.
Melo AS, Macedo ACC, Andrade CFS. [17] Eff iciency of Dugesia tigrina (Girard) (Turbellaria: Tricladida) as a controlling agent of immature mosquitoes Aedes albopictus (Skuse) in tire-trap. Anns Entomol Soc Brazil 1996;25:321-327.
Rey JR, O’Connell S, Suarez S, Menendez [18] Z, Lounibos LP, Byet G. Laboratory and field studies of Macro Cyclops albidus (Crustaceae: Copepoda) for biological control of mosquitoes in artificial containers in a subtropical environment. J Vector Ecol 2004;29:124-34.
Fay RW, Perry AS. Laboratory studies of [19] ovipositional preference of Aedes aegypti. Mosq News 1965;25:276-281.
Fay RW, Eliason DA. A preferred oviposition [20] site as a surveillance method for Aedes aegypti. Mosq News 1966;26:531-535.
Muir LE, Kay BH, Thorne MJ. [21] Aedes aegypti (Diptera: Culicidae) vision: response to stimuli from the optical environment. J Med Entomol 1992;29:445-450.
Escalante NG, Salas IF, Dantes HG. Field [22] evaluation of Mesocyclops longisetus (Copepoda: Cyclopoidea) for the control of larval Aedes aegypti (Diptera: Culicidae) in Northern Mexico . J Med Entomol 1998;35:699-703.
Baily PCE. The effect of water temperature [23] on the functional response of the water stick insect Ranatra dispar (Heteroptera: Nepidae). Australian J Ecol 1989;14:381-386.
Venkatesan P, Jayachandra CM. Estimation [24] of mosquito predation by the water bug Diplonychus indicus Venkatessan & Rao. Indian J Exp Biol 1985;23:227-229.
Victor R, Ugwoke LI. Preliminary studies [25] on predation by Sphaerodema nepoides Fabricious (Heteroptera: Belostomatidae). Hydrobiologia 1987;154:25-32.
Cloarec A. Predatory versatility in the water [26] bug , Diplonychus indicus. Behav Proc 1991;23:231-242.
Vekatesan P, D’Sylva Tena. Influence of [27] prey size on choice by the water bug , Diplonychus indicus, Vekatesan & Rao (Hemiptera: Belostomatidae). J Entomol Res 1990:14:130-138.
Sivagnaname N. Effects of insecticides and [28] biocides on the biocontrol potential of an aquatic mosquito predator, Diplonychus indicus, Venkatessan & Rao (Hemiptera: Belostomatidae). Anns Med Entomol 2007;16:7-20.
Sivagnaname N. Selective and frequency [29] dependent predation of aquatic mosquito predator, Diplonychus indicus Venkatesan & Rao (Hemiptera: Belostomatidae). Entomol Res 2009;39:356-363.
Aditya G, Bhattacharyya S, Kundu V, Saha [30] GK, Raut SK. Predatory efficiency of the water bug, Sphaerodema annulatum on mosquito larvae (Culex quinquefasciatus) and it’s effect on adult emergence. Bioresource Technology 2004;95:169-172.
Shalam EAA, Canyan DV, Muller R, Younes [31] MWF, Wahab HA, Mansour AH. A mosquito survey in Townville. Australia and an assessment of Diplonychus Sp and Anisops Sp. Predatorial capacity against Cx. annulirostris mosquito immature. J Vector Ecol 2007;32:16-21.
Panicker KN, Geetha Bai, Kalyanasundaram [32] M. The breeding behaviour of Aedes aegypti. Indian J Med Res 1982;76:689-91.
Babu CJ, Panicker KN, Das PK. Breeding of [33] Aedes aegypti in closed septic tanks. Indian J Med Res 1983;77:637.

160 Dengue Bulletin – Volume 33, 2009
Control of Ae. aegypti breeding in tyres using hemipteran bug – D. indicus
Venkatesan P, Sivaraman S. Changes in the [34] functional response of instars of Diplonychus indicus , Venk and Rao (Hemiptera: Belostomatidae) in its predation of two species of mosquito larvae of varied size. Entomon 1984;9:191-196.
Duviard D. Flight activity of Belostomatidae [35] in Cent ra l Ivory Coas t . Ocecolog ia 1974;15:312-28.
World Health Organization. [36] Report of the consultation on: key issues in dengue vector control towards the operationalization of a global strategy. Geneva: WHO, 1995.
Reiter P, Gubler DJ. Surveillance and control [37] of urban dengue vectors. In: Gubler DJ, Kuno G. Eds. Dengue and dengue haemorrhagic fever. Chapter 20. London: CAB International, 1997.
Chan KL, Ho BG, Chan YV. [38] Aedes aegypti (L) and Aedes albopictus (Skuse) in Singapore city. 2. Larval habitats. Bull World Health Organ 1971;44:629-633.
Ellis RA, Borden JH. Predation by [39] Notonecta undulata Say (Hemiptera: Notonectidae) on larvae of yellow fever mosquito. Ann Entomol Soc Amer 1970;63:963-973.
Panicker KN, Rajagopalan PK. Biological-[40] control potential of Anisops bouvieri kirkaldy (Hemiptera:Notonectidae). Indian J Med Res 1977;66:772-776.
Jennings CD, Phommasack B, Sourinadeth [41] B, Kay BH. Aedes aegypti control in the Lao People’s Democratic Republic with reference to copepods. Am J Trop Med Hyg 1995;53:324-330.
Monrique-Saide P, Serigo Ibanez-Bernal, [42] Hugo Delfin-Gonzolez, Victor Parra Tabla. Mesocyclops lingisetus effects on survivorship of Aedes aegypti immature stages in car tyres. Med Vet Entomol 1998;12:386-390.
Riviere F, Sechan Y, Kay BH. The evaluation [43] of predators for mosquito control in French
Polynesia. In: St. George TD, Kay BH, Blok J. Eds. Fourth symposium arbovirus research in Australia, May 6-9, 1986. Brisbane, 1986.
Nam VS, Yen NT, Holynsha M, Resid JW, [44] Kay BH. National progress in dengue vector control in Vietnam: survey for Mesocyclops (Copepoda), Micronecta (Corixidae) and fish as biocontrol agents. Am J Trop Med Hyg 2000:62,5-11.
Cohen AC. Extra-oral digestion in predaceous [45] terrestrial arthropods. Ann Rev Entomol 1995;40:85-103.
Cloarec A. Variations of foraging tactics in [46] a water bug. Diplonychus indicus. J Ethol 1989;7:27-34.
Hokkenen HMT, Pimental D. New associations [47] in biological control: theory and practice. Canadian Entomol 1989;121:829-840.
Sebastian A, Thu MM, Kyaw M, Sein MM. [48] The use of dragonfly nymphs in the control of Aedes aegypti. Southeast Asian J Trop Med Public Health 1980;11:104-107.
Venkatesan P. Prospects for bioethical [49] management of mosquito menace and mosquito-borne diseases. Jayapaul Azariah, Hilda Azaria, Darryl RJ. Eds. In: Bioethics in India: proceedings of the international bioethics workshop in Madras: biomanagement of bioresources 1997; 16-19 Jan 1997, University of Madras. Madras: Ethics Institute, 1997. www.http://eubios.info/index.html.
Mogi M. Insects and other invertebrates [50] predators. In: Floore TG. Ed. Biorational control of mosquitoes. Bulletin of the American Mosquito Control Association 7. 2007;23 (special issue):93-109.
Noeleen R, Venkatesan P. Prey preference [51] and predatory efficiency of the water bug, Diplonychus indicus, Venk & Rao (Hemiptera: Belostomatidae), an effective biocontrol agent for mosquitoes. J Entomol Res 1997;21:267-272.

Dengue Bulletin – Volume 33, 2009 161
Effect of water supply system installation on distribution of water storage containers and abundance of Aedes aegypti immatures in urban premises of
Ho Chi Minh City, Viet Nam Ataru Tsuzukia#, Trang Huynhb, Loan Luub, Takashi Tsunodaa and Masahiro Takagia
aDepartment of Vector Ecology and Environment, Institute of Tropical Medicine, Nagasaki University, 1-12-4 Sakamoto, Nagasaki City 852-8523, Japan
bHo Chi Minh Pasteur Institute, 167 Pasteur Street, 8 Ward, 3 District, Ho Chi Minh City, Viet Nam
Abstract
To assess the effect of installation of a water supply system on the distribution of water storage containers and abundance of Aedes aegypti immatures, we conducted two successive entomological surveys in October 2007 and July 2008 in an urban neighbourhood of Ho Chi Minh City, Viet Nam. In all, 850 water-holding containers, including plastic buckets, water jars, concrete basins, flower vases, ant-traps, outdoor discards, used tyres and plant saucers were observed in 122 premises. Of these, 44 premises already had a water supply system installed at the time of the first survey (i.e. control premises). In the remaining 78 premises, a water supply system was installed 3–6 months before the second survey (i.e. intervention premises). There was a drastic reduction in the number of water storage containers, except concrete basins, between the first and second surveys. We also observed that the distribution of water storage containers in the control and intervention premises was similar. This was probably because the construction of a water supply system before the second survey increased the reliability of the system not only for the residents in intervention premises but also in the control premises (i.e. increased water pressure in the water supply system). The number of plastic buckets and water jars was reduced in the second survey; however, these containers were still an important source of Ae. aegypti pupae. For effective control of dengue infection, it is necessary to conduct entomological monitoring considering the seasonal changes in vector abundance and dengue transmission threshold after installing a water supply system.
Keywords: Dengue; Aedes aegypti; Viet Nam; water supply, vector control.
#E-mail: [email protected]
Introduction
Dengue fever and the associated lethal shock and/or haemorrhage are rapidly growing public health problems in over 100 countries
of the world, and particularly in South-East Asia, North and South America, and the Western Pacific islands. It has been estimated that about 50 × 106 cases of dengue viral infection, including 50 × 104 cases of dengue

162 Dengue Bulletin – Volume 33, 2009
Water supply system installation and distribution of containers and abundance of Ae. aegypti in Viet Nam
fever with shock and/or haemorrhage, occur worldwide annually.[1] An effective vaccine against dengue has not yet been developed.[2] To date, the only option for controlling dengue virus transmission in human population is by reducing the population density of Aedes aegypti—the mosquito primarily responsible for causing dengue—which exclusively feeds on human blood and deposits eggs in a variety of artificial water storage containers in and around residential premises.
An increase in the density and geographical distribution of Ae. aegypti is known to be widely influenced by unprecedented global population growth and unplanned urbanization, which has resulted in substandard housing and inadequate water supply and waste management systems. These conditions increase the number and prevalence of water storage containers that may become the breeding sites for pre-adult Ae. aegypti.[3] However, the effect of installing a water supply system in dengue endemic areas as a measure to reduce the number of potential breeding sites and abundance of Ae. aegypti pre-adults (i.e. larvae and/or pupae) have been evaluated in only a few studies. In order to identify the effect of installation of a water supply system on the distribution of water storage containers and abundance of Ae. aegypti breeding, we conducted two successive entomological surveys in an urban neighbourhood in Ho Chi Minh City in Viet Nam.
Materials and methods
Entomological surveys were conducted in October 2007 (first survey) and July 2008 (second survey) in an urban neighbourhood in District No. 8 of Ho Chi Minh City. The city has a tropical climate with distinct wet and dry seasons. The rainy season usually begins in May and ends in late November;
the dry season lasts from December to April. The surveys were conducted to determine the distribution of water storage containers and prevalence of Ae. aegypti breeding before and after the installation of a water supply system in the study area. The detailed study methods and characteristics of our study area (referred to as area A in the previous study) have been described previously.[4] Containers with more than one Ae. aegypti larva and/or pupa were defined as “positive containers”. We believed that it was reasonable to obtain the Ae. aegypti pupal count, rather than the larval count, to estimate the number of adults because of the high survival rate of pupae[5] and the ease of identification of emerged adult species. All water storage containers situated both indoors and outdoors (including porches and rooftops) were inspected. All the pupae were collected in small plastic vials and subsequently transported to the laboratory. The number of pupae was counted, and the species of the mosquitoes (i.e. Ae. aegypti or others) was confirmed after emergence.
The mosquito density was evaluated in terms of the house index (HI): percentage of houses with containers positive for Ae. aegypti larvae or pupae; container index (CI): percentage of water storage containers positive for Ae. aegypti larvae and pupae; Breteau index (BI): number of containers positive for Ae. aegypti larvae and pupae per 100 houses; and pupae per person (PPP): number of Ae. aegypti pupae per person (i.e. average number of pupae among the number of residents). The data were entered into the Microsoft Office Access 2003 program (Microsoft Corporation, USA) and analysed using Stata™ v 10.0 (Stata Corporation, USA). The average number of containers per premise was expressed as mean (SD). Statistical analysis was performed using the t test. Significance was determined at the 5% level.

Dengue Bulletin – Volume 33, 2009 163
Water supply system installation and distribution of containers and abundance of Ae. aegypti in Viet Nam
Results
Among the 174 residential premises in the study area, 122 (70.1%) were inspected both in the first and the second surveys. When we conducted the first survey, 78 (55.7%) premises did not have a water supply system (intervention premises) and private pump-wells were the main source of water. In the other 44 (44.3%) premises, the water supply system had already been installed (control premises). A water supply system was installed by the Ho Chi Minh City water supply service in 78 premises 3–6 months before the second survey. In comparison with the larval density indexes (CI, HI, BI, and PPP) obtained during the first survey, all, except PPP, were reduced during the second survey in the 78 intervention premises; however, these indices were similarly reduced in the 44 control premises (Table 1).
In all, 850 water-holding containers, including plastic buckets, water jars, concrete basins, flower vases, ant traps, outdoor discards, used tyres and plant saucers, were observed in the
122 premises (Table 2). Almost all the water storage containers, such as plastic buckets, water jars and concrete basins, were located indoors or under eaves; outdoor water-holding containers (e.g. used tyres, plant saucers and outdoor discards) were few in number because most of the surveyed urban residential premises were located adjacent to each other or were connected, leaving little or no outdoor space. Water-storage containers (i.e. plastic buckets, water jars and concrete basins) accounted for 53.8% of the total water-holding containers. There was a drastic reduction in the number of plastic buckets and water jars between the first and second surveys in both the intervention and control premises; however, the number of concrete basins did not change between the survey periods. The above-mentioned water storage containers accounted for the majority of positive containers in the first (55.3% in the intervention premises and 63.7% in the control premises) and second surveys (68.0% in the intervention premises and 86.7% in the control premises). However, the number of positive containers in the second survey was less than that in the first survey. The water
Table 1: Pre-adult Ae. aegypti indices of 122 premises before and after water supply installation in Ho Chi Minh City, Viet Nam*
1st survey (before) 2nd survey (after)
Intervention (n = 78)
Control (n = 44)
Intervention (n = 78)
Control (n = 44)
Container index (CI) 28.4% 30.6% 21.4% 14.3%
House index (HI) 53.9% 56.8% 37.2% 29.5%
Breteau index (BI) 120.5 125.0 64.1 34.1
Pupae per person 0.37 0.45 0.46 0.40
* The first and second surveys were conducted in October 2007 and July 2008, respectively, in an urban neighbourhood in District No. 8 of Ho Chi Minh City. A water supply system was installed in the 78 premises 3–6 months before the second survey (i.e. intervention premises), whereas the system had been already installed in 44 premises when the first survey was conducted (i.e. control premises).

164 Dengue Bulletin – Volume 33, 2009
Water supply system installation and distribution of containers and abundance of Ae. aegypti in Viet Nam
Table 2: Number of water-holding containers and presence of pre-adult Ae. aegypti in 122 premises before and after water supply installation in Ho Chi Minh City, Viet Nam
Container types
1st survey (before) 2nd survey (after)
No. of containers
(%)
No. of pre-adult positive
containers (%)
No. of pupae collected
(%)
No. of containers
(%)
No. of pre-adult positive
containers (%)
No. of pupae collected
(%)
Intervention (n = 78)
Plastic buckets 111 (33.5) 21 (22.3) 34 (24.3) 32 (13.7) 9 (18.9) 41 (23.2)
Water jars 61 (18.4) 21 (22.3) 49 (35.0) 46 (19.7) 19 (38.0) 103 (58.2)
Concrete basins* 20 (6.0) 10 (10.6) 18 (12.9) 20 (8.5) 6 (12.0) 7 (4.0)
Others‡ 139 (42.0) 42 (44.7) 39 (27.9) 136 (58.1) 16 (32.0) 26 (14.7)
Total 331 (100) 94 (100) 140 (100) 234 (100) 50 (100) 177 (100)
Control (n = 44)
Plastic buckets 61 (33.9) 14 (25.5) 3 (2.8) 27 (25.7) 1 (6.7) 25 (26.6)
Water jars 29 (16.1) 12 (21.8) 39 (36.8) 16 (15.2) 3 (20.0) 0 (0)
Concrete basins* 17 (9.4) 9 (16.4) 52 (49.1) 17 (16.2) 9 (60.0) 62 (66.0)
Others‡ 73 (40.6) 20 (36.3) 12 (11.3) 45 (42.9) 2 (13.3) 7 (7.4)
Total 180 (100) 55 (100) 106 (100) 105 (100) 15 (100) 94 (100)
* Built-in concrete basins in kitchens, gardens and bathrooms were included.‡ Others included flower vases, ant-traps, outdoor discards, used tyres and plant saucers.
storage containers responsible for most of the pupal population in the intervention premises were 72.1% and 85.3% in the first and second survey, respectively; and in control premises were 88.0% and 92.6% in the first and second survey, respectively.
Although the number of plastic buckets and water jars per premises during the second survey had drastically decreased compared with the first survey (plastic buckets: P<0.001 and P<0.001 in the intervention and control premises, respectively; water jars: P=0.05 and P<0.05 in the intervention and control premises, respectively), no significant difference
was observed between the intervention and control premises (Table 3). We found only Ae. aegypti pupae in the water-holding containers throughout the survey period in our study area.
DiscussionOur results indicated that installation of a water supply system in residential premises may lead to a reduction in the number of different water storage containers, such as plastic buckets and water jars, within a short period. However, other factors such as the

Dengue Bulletin – Volume 33, 2009 165
Water supply system installation and distribution of containers and abundance of Ae. aegypti in Viet Nam
reliability of the supply system may have a greater impact on reducing the number of water-holding containers. In the 78 premises that had private pump-wells before the water system was installed, water was intermittently pumped up from the wells and stored in plastic buckets, water jars and concrete basins. After the installation of the water supply system, residents could directly use the water from taps, thereby avoiding the need to store water. Hence, the number of water storage containers, except for concrete basins, in the study area reduced within a short period. We also observed that the distribution of the water storage containers in the control premises was similar to their distribution in the intervention premises and the number of water storage containers had drastically reduced in the second survey. This may be because of the construction of a water supply system 3–6 months before the second survey, which increased the reliability of the system not only for the residents in the intervention premises but also for those in the control premises (i.e. increased water pressure in a water supply system and less frequent suspension of water supply).
The first survey may have also resulted in a change in the water storage habits of
the residents (i.e. people recognized Ae. aegypti breeding sites in their premises during the house-to-house entomological survey). Although we do not know about the seasonal changes in the water-storing behaviour of residents, almost all the water storage containers in which water was stored artificially were located indoors or under eaves in our study area, and seasonal changes in the number of water storage containers might not be influenced by rainfall. The number of concrete basins before and after water supply system installation did not change because they were fixed structures in the houses (i.e. built-in type).
The number of plastic buckets and water jars was reduced in the second survey; however, these containers were still an important source of Ae. aegypti pupae. A high mean temperature promotes the growth of Ae. aegypti.[6] Different weather conditions between the first and second surveys might be the primary reason for the difference in Ae. aegypti abundance. Furthermore, after installation of the water supply system, the less frequently used water-storage containers and the resultant standstill water in them may have also favoured larval breeding (i.e. fewer chance of water exchange and more nutrition
Table 3: Average number of plastic buckets and water jars in premises before and after water supply installation, Ho Chi Minh City, Viet Nam*
1st survey (before) 2nd survey (after)
Intervention (mean (SD))
Control (mean (SD)) P Intervention
(mean (SD))Control
(mean (SD)) P
Plastic buckets 1.94 (1.18) 1.74 (1.34) 0.5 0.56 (0.68) 0.69 (0.83) 0.2
Water jars 2.03 (1.35) 2.23 (1.09) 0.6 1.33 (1.37) 1.08 (1.38) 0.6
*Average number of containers in the premises that owned plastic buckets and water jars in the first survey was calculated (i.e. 57 and 35 of the intervention and control premises, respectively, had plastic buckets; 30 and 13 of the intervention and control premises, respectively, had water jars).

166 Dengue Bulletin – Volume 33, 2009
Water supply system installation and distribution of containers and abundance of Ae. aegypti in Viet Nam
in the water). Thus, more pupae may have been produced from fewer numbers of plastic buckets and water jars.
It was not clear why some residents stored water in jars and plastic buckets even after the water supply system had been installed; we speculate that this could be because residents felt using water from the well was economical (i.e. no extra cost incurred, except for the electricity used by the motor pump), or because they were still doubtful about an assured water supply system given the fact that the system had been newly installed in the area.
It is difficult to immediately change the attitude/habits of residents; in the meantime, water storage containers such as plastic buckets, water jars and concrete basins may become
important breeding sites for Ae. aegypti. To effectively control dengue infection, it is necessary to conduct entomological monitoring considering the seasonal changes in vector abundance and dengue transmission threshold[7] even after installation of a water supply system. We believe that a follow-up survey in the succeeding year and/or a survey spanning more residential areas would be helpful in reaching a definitive conclusion.
AcknowledgmentsWe express our deep gratitude to Cuc Vu, Sy Ngo, Lap Nguyen, Tung Le, Khanh Ly, Van Nguyen and Thai Nguyen at the Ho Chi Minh Pasteur Institute and the District No. 8 Health Office for field assistance.
ReferencesHalstead SB. Dengue virus-mosquito [1] i n t e r a c t i o n s . A n n u R e v E n t o m o l 2008;53:273-91.
Webster DP, Farrar J, Rowland-Jones S. Progress [2] towards a dengue vaccine. Lancet Infect Dis 2009;9(11):678-87.
Guzman MG, Kouri G. Dengue: an update. [3] Lancet Infect Dis 2002;2(1):33-42.
Tsuzuki A, Huynh T, Tsunoda T, Luu L, [4] Kawada H, Takagi M. Effect of existing practices on reducing Aedes aegypti pre-adults in key breeding containers in Ho Chi Minh City, Vietnam. Am J Trop Med Hyg 2009;80(5):752-7.
Focks DA, Haile DG, Daniels E, Mount [5] GA. Dynamic life table model for Aedes
aegypti (Diptera: Culicidae): simulation results and validation. J Med Entomol 1993;30(6):1018-28.
Focks DA, Haile DG, Daniels E, Mount GA. [6] Dynamic life table model for Aedes aegypti (Diptera: Culicidae): analysis of the literature and model development. J Med Entomol 1993;30(6):1003-17.
Focks DA, Alexander N. [7] Multicountry study of Aedes aegypti pupal productivity survey methodology: finding and recommendations. TDR/TRM/Den/06.1 Geneva: WHO, 2006. http://apps.who.int/tdr/publications/tdr-research-publications/multicountry-study-aedes-aegypti/pdf/aedes_aegypti.pdf – accessed 02 July 2010.

Dengue Bulletin – Volume 33, 2009 167
Evaluation of premise condition index in the context of Aedes aegypti control in Marília, São Paulo, Brazil
Maria Teresa Macoris Andrighetti, Karen Cristina Galvani and Maria de Lourdes da Graça Macoris#
Superintendência de Controle de Endemias – SUCEN, Serviço Regional 11, Av. Santo Antônio, 1627. 17 506 040 Bairro Somenzari, Marília, Brazil
Abstract
The dengue control programme in Brazil involves house-to-house inspection in each municipality, on a bimonthly basis, for vector control and education activities. The sustainability of this strategy has been questioned due to its failure in interrupting dengue transmission and also by the exhaustion of field workers and lack of active cooperation by populations due to the tedious routine of bimonthly visits to all houses. An evaluation of the infestation of Aedes aegypti by larval, pupal and adult collection, and classification of houses with the premise condition index (PCI) in conjunction with residents’ housing characteristics was performed in 11 575 houses in Marilia, Sao Paulo state, Brazil. The results showed that the surveillance tool and associated data on resident housing characteristics were useful in identifying heavily infested premises. The index could thus provide a basis for prioritizing control measures within communities during routine vector control activities.
Keywords: Aedes aegypti surveillance; premise condition index; housing characteristics; vector control..
#E-mail: [email protected]; Phone/fax 55 14 34331080
IntroductionIn Brazil, the National Programme for Dengue Control[1] (PNCD) has been ongoing since 2002 and integrates eight components: epidemiological surveillance, vector control, medical assistance, integration with the primary health-care system, environmental sanitation, education and social mobilization, social communication, and training and legislation. For vector control, the strategy recommends the bimonthly inspection
of all houses in each municipality where infestation by Aedes aegypti is registered. During each visit, the vector control agent looks for breeding sites, gives educational information to residents that explains how to prevent mosquito foci, and uses larvicides where environmental management methods are not feasible.
Eyebrows have been raised against this strategy as it has failed to prevent dengue transmission, and also due to the growing

168 Dengue Bulletin – Volume 33, 2009
Evaluation of premise condition index in the context of Ae. aegypti control in Marília, São Paulo, Brazil
attrition and exhaustion of both vector control workers and residents due to the tedious and routine nature of bimonthly visits to all houses.
A stratified approach to the problem was described by Tun-Lin,[2] who proposed a new indicator, the premise condition index (PCI), based on certain environmental characteristics of the house, yard and shading of premises in which there was an evident relationship between the PCI and the risk of harbouring breeding foci of the vector. There have been a number of subsequent studies corroborating these findings.[3-6] In accordance with this approach, there is growing evidence about the influence of socioeconomic and environmental conditions as expressed by the condition of the premises on the prevalence of dengue. The construction of maps based on social conditions has identified areas for intervention with the possibility of ranking geographical points for different vector control actions.[7-14]
With the objective of evaluating how this new tool could be used within the routine of the PNCD, a study was carried out to compare infestation of Ae. aegypti using several entomological parameters and PCI.
Methods
Study area
The study was conducted in Marília, a city in the western area of the state of Sao Paulo, in Brazil. The city with 223 000 inhabitants has been infested by Ae. aegypti since 1989. However, due to an efficient vector control programme, dengue transmission has been kept in check.[15] The urban area covers 42 sq. km. with about 88 000 premises. There is regular refuse collection and water supply in the city.
Field mosquito collection
A sample size of 1200 houses for each field survey was determined.[16] The surveys were performed bimonthly for two and a half years (October 2006 to March 2009). With the definition of the number of houses to be inspected, the choice of blocks where all houses should be inspected was defined by probabilistic raffling using the census tract as the data aggregation unit. As the census lists all blocks in the city, the sampling involved the entire urban area.
At the premises, the inspection was performed by a team of three field workers having skills in entomology. During house inspection all possible breeding sites were inspected for the presence of larvae and pupae. When the presence of immature forms of mosquitoes was found, a sample of larvae and the total number of pupae in each breeding site was collected. Adult collection was performed with a motorized backpack aspirator.[17] Data from field collections were used for the estimation of Ae. aegypti indices and also new indices as proposed by Fox and Chadee.[18-21]
All field data was registered in Access databank and analysed with Epi-info 3.3, 2004 software. Single linear regression was used to calculate the correlation between variables related to infestation and the PCI.
PCI scoring
Each premise that was inspected for the presence of Ae. aegypti was scored according to the method developed by Tun-Lin et al.[2] to obtain its PCI value. The degree of shade, the condition of the house and the condition of the yard in all premises were visually assessed and recorded during the inspection. For each of the three variables scores ranged from

Dengue Bulletin – Volume 33, 2009 169
Evaluation of premise condition index in the context of Ae. aegypti control in Marília, São Paulo, Brazil
1 to 3, with their sum, representing the PCI, as a value from 3 to 9. The lowest ranking PCI represented conditions that were considered the least favourable for the vector and the highest ranking PCI the most favourable.
Variables regarding the person in charge of household responsibilities
Considering our experience on the importance of the residents’ adoption of control measures on infestation,[19-21] a survey on each premise was performed during the visit to identify selected characteristics of the person who was in charge of household responsibilities. Having obtained agreement from the interviewee, the variables recorded using a standard questionnaire were age, profession, educational level, knowledge of someone who already had dengue, water supply periodicity and knowledge about the vector control agent in the area. Residents’
educational levels were classified 1 for no formal education, 2 for elementary level, 3 for second-grade or intermediate level and 4 for university level.
Results
Distribution of positive premises by PCI ranking
During the period of the study a total of 11 575 premises were inspected for the presence of immature and adult Ae. aegypti and also classified according to their PCI score. The number and per cent distribution of the premises according to their PCI ranking is shown in Table 1.
There were 2444 premises which could not be classified according to the PCI. These were non-residential premises, vacant lots and construction sites. As those premises
Table 1: Number and per cent distribution of inspected premises by PCI ranking
PCI Number of premises Percentage
Number of premises
with larvae
Positivity of premises for larvae
Number of premises
with pupae
Positivity of premises for pupae
0* 2444 21.1 49 2.0 26 1.1
3 283 2.4 3 1.1 0 0.0
4 925 8.0 14 1.5 3 0.3
5 2144 18.5 37 1.7 13 0.6
6 3805 32.9 129 3.4 58 1.5
7 1293 11.2 98 7.6 48 3.7
8 499 4.3 71 14.2 34 6.8
9 182 1.6 40 22.0 18 9.9
Total 11 575 100 441 3.8 200 1.7
*Non-residential premises, vacant lots and construction sites; not classified by PCI.

170 Dengue Bulletin – Volume 33, 2009
Evaluation of premise condition index in the context of Ae. aegypti control in Marília, São Paulo, Brazil
were included on the blocks that had been identified for survey, they were also inspected for the presence of Aedes indices and coded as PCI “zero” as it was not possible to use the criteria proposed for classification. The decision to include the survey on these premises was also based on the vector control routine where workers must visit all premises, including non-residential ones.
The majority of premises were ranked with an intermediate PCI score of 6, whereas those with the lowest and highest scores (PCI 3 and PCI 9) represented only 2.4% and 1.6% of the premises respectively.
The positivity for larvae, pupae and adult of Ae. aegypti was highly correlated to PCI with a “r” value of 0.90, 0.93 and 0.96 (p<0.05) respectively for each stage (Tables 1 and 3).
Among the non-residential premises, vacant lots and construction sites (PCI “0”), 2.0%, 1.1% and 6.0% were infested with larvae, pupae and adult of Ae. aegypti respectively.
Pupal positivity by PCI ranking
Seventy-four per cent of pupae were found on premises with PCI values ≥6, while 20.9% of all Ae. aegypti pupae were found in zero category (Table 2).
Distribution of adults in premises without breeding
The distribution of adults was not restricted to the premises where they were reared. Table 3 includes data on the number of Ae. aegypti in premises where immature stages were not detected.
Table 2: Number of pupae and per cent distribution by PCI ranking
PCI Number of pupae
Per cent
Cumulative per cent
0* 397 20.9 20.9
3 0 0 0
4 37 1.9 22.8
5 58 3.0 25.9
6 429 22.5 48.4
7 374 19.7 68.1
8 478 25.1 93.2
9 130 6.8 100
Total 1903 100 100
* Non-residential premises, vacant lots and construction sites; not classified by PCI.
Pupae per person, indices by age groups of householders
Of all the premises that were inspected, we had the agreement for questionnaire response from the residents of 7800. An analysis of the mean distribution of pupae per person (Table 4) showed a direct relationship with the increasing age of the residents in charge of the households. Pupae per person was the lowest at 0.03 in the age group <20 years while it reached the maximum of 0.10 in the age group >59 years.
Pupae per person by educational level
Among the characteristics of the person in charge of the households that were collected in the interviews, the educational level was the only one which was correlated with the infestation. The mean number of pupae per

Dengue Bulletin – Volume 33, 2009 171
Evaluation of premise condition index in the context of Ae. aegypti control in Marília, São Paulo, Brazil
Table 3: Distribution of adult Ae. aegypti by PCI ranking and presence of immature foci
PCINumber of
premises with adults
Total number of mosquitoes
Number of mosquitoes in premises without
immature forms
Per cent from the total number of
mosquitoes
0* 146 834 558 66.9
3 22 30 25 83.3
4 171 473 436 92.2
5 363 813 702 86.4
6 742 2336 1783 76.3
7 318 952 688 72.3
8 134 702 306 43.6
9 57 288 175 60.8
Total 1953 6428 4673 72.7
*Non-residential premises, vacant lots and construction sites; not classified by PCI.
Table 4: Number of pupae distributed by age of person in charge of household duties among the respondents to the questionnaire
Age (years)
Number of respondents Number of pupae Percent of pupae Number of pupae
person
Less than 20 474 51 4.2 0.03
21–39 2345 147 12.2 0.02
40–59 2976 518 43.0 0.05
Over 59 2009 489 40.6 0.10
Total 7804 1205 100 0.06
premise (Table 5) was negatively correlated with the educational level (Pearson Correlation coefficient = 0.81).
Discussion
The use of PCI has proved to be an effective tool to classify premises according to the risk of harbouring mosquito breeding sites and,
in this sense, a tool to prioritize houses for vector control measures. However, training field teams for PCI ranking was not easy as it took a while for the teams to standardize the method of classification. This difficulty was also reported by Nogueira.[5]
The evaluation of PCI on a large scale in a whole city for a significant length of time gave important information on the immature

172 Dengue Bulletin – Volume 33, 2009
Evaluation of premise condition index in the context of Ae. aegypti control in Marília, São Paulo, Brazil
The majority of premises (51.4% of the total) were classified on the intermediate scores of 5 to 6 which could be read as the “mid-ranking” of scores.
Other important limitation of the PCI methodology is the fact that non-residential premises, vacant lots and construction worksites could not be ranked and that they comprised 21% of the premises studied. As they harboured 11.6% of larvae, 20.9% of pupae and 20.8% of adults, their contribution to the infestation was of great relevance.
The presence of foci in vacant lots is of operational significance because in this kind of premises no resident looks after the control measures; hence this task must be performed by the vector control teams.
The low positivity for all forms of the vector, illustrated in Figures 1 – 3, show how relevant it is to focus vector control on a small number of premises. Considering the high correlation between PCI and vector infestation, this tool appears to be effective if operational vector control measures are targeted or stratified. These findings are
Table 5: Mean number of pupae per premise, according to educational level of residents
Formal education Pupae per premise
No information 7.3
1 14.5
2 4.2
3 2.9
4 3.8
Total 5.4
1 = no formal education; 2 = elementary level; 3 = intermediate level; 4 = university.
Figure 1: Percentage of premises with Ae. aegypti larvae according to PCI score
and adult distribution of the vector species, and also raised important questions to be addressed in the implementation of this strategy for vector control.
The first observation is related to the uneven distribution of premises. As listed in Table 1, a small number of premises are classified on the extreme scores (3 and 9), corresponding to 2.4% and 1.6% of the total number of premises inspected respectively.

Dengue Bulletin – Volume 33, 2009 173
Evaluation of premise condition index in the context of Ae. aegypti control in Marília, São Paulo, Brazil
Figure 2: Percentage of premises with Ae. aegypti pupae according to PCI
Figure 3: Percentage of premises with Ae. aegypti adult according to PCI score
consistent with those from the studies by Nogueira[5] and Maciel de Freitas[6] when working with key houses in Brazil.
In our study, data showed that if only 17.1% or 5.9% of the premises that were classified on the worse PCI scores of 7–9 or 8–9 are responsible for the production of
51.6% or 31.9% of pupae, efforts for source reduction could be directed at those premises. The choice of thresholds in PCI to plan vector control activities will depend on field resources and also on the epidemiological situation.
Another possibility that this tool provides to the Brazilian Dengue Control Programme

174 Dengue Bulletin – Volume 33, 2009
Evaluation of premise condition index in the context of Ae. aegypti control in Marília, São Paulo, Brazil
is to schedule the vector control teams’ visits with different frequencies based on PCI scores. As there is a routine of visiting 100% of premises, one PCI survey per year may be enough for strategic planning. Intense vector control on those premises might impact vector density with lower costs than the current methodology.
Community participation is crucial for Ae. aegypti control since domestic breeding sites play a major role in mosquito infestation.
Considering that there are barriers for people adopting vector control attitudes, we investigated the characteristics of the residents during the surveys in order to identify what was the profile of the person charged with taking care of the house. From all the variables collected, age and educational level were found to be important criteria for identifying premises with the presence of pupae. This information should be considered important for devising appropriate communication and educational strategies and also for vector control planning.
ReferencesFundação Nacional da Saúde. Programa [1] Nacional de Controle da Dengue (PNCD). Brasília, Funasa; 2002.
Tun-Lin W, Kay BH, Barnes A. The premise [2] condition index: a tool for streamlining surveys of Aedes aegypti. Am J Trop Med Hyg 1995;53(6):591-4.
Moloney JM, Skelly C, Weinstein P, Maguire [3] M, Ritchie S. Domestic Aedes aegypti breeding site surveillance: limitations of remote sensing as a predictive surveillance tool. Am J Trop Med Hyg 1998;59:261-264.
Gomez FE, Surez CMM, Cardenas RC. Factores [4] que modifican los índices larvarios de Aedes aegypti en Colima, México. Rev Panam Salud Publica 2001;10:6-12.
Nogueira LA, Gushi LT, Miranda JE, Madeira [5] NG, Ribolla PEM. Application of an alternative Aedes species (Diptera: Culicidae) surveillance method in Botucatu city, Sao Paulo, Brazil. Am J Trop Med Hyg 2005;73:309-311.
Maciel-de-Freitas R, Peres RC, Souza-Santos [6] R, Lourenco de Oliveira. Occurrence, productivity and spatial distribution of key premises in two dengue-endemic areas of Rio de Janeiro and their role in adult Aedes aegypti
spatial infestation pattern. Trop Med Int Health 2008;13(12):1488-1494.
Akerman M, Stephens C, Campanario P, Maia [7] PB. Saúde e meio ambiente: uma análise de diferenciais intra-urbanos enfocando o Município de São Paulo, Brasil. Rev Saúde Pública 1994;28(4):320-325.
Akerman M, Campanario P, Maia PB. Saúde e [8] meio ambiente: análise de diferenciais intra-urbanos, Município de São Paulo, Brasil. Rev Saúde Pública 1996;30(4):372-382.
Costa AIP da, Natal D. Distribuição espacial da [9] dengue e determinantes socioeconômicos em localidade urbana no Sudeste do Brasil. Rev Saúde Pública 1998;32(3):232-236.
Lacerda J, Calvo M, Freitas S. Diferenciais [10] intra-urbanos no Município de Florianópolis, Santa Catarina, Brasil: potencial de uso para o planejamento em saúde. Cad Saúde Pública 2002;18(5):1331-1338.
Oliveira CDL, Assunção R, Reis IA, Fernando [11] AP. Spatial distribution of human and canine visceral leishmaniasis in Belo Horizonte, Minas Gerais State, Brasil, 1994-1997. Cad Saúde Pública 2001;17(5):1231-1239.

Dengue Bulletin – Volume 33, 2009 175
Evaluation of premise condition index in the context of Ae. aegypti control in Marília, São Paulo, Brazil
Ximenes RA, Martelli CM, Souza WV, Lapa TM, [12] Albuquerque M, Andrade AL, Morais Neto OL, Silva SA, Lima ML, Portugal JL. Vigilância de doenças endêmicas em áreas urbanas: a interface entre mapas de setores censitários e indicadores de morbidade. Cad Saúde Pública 1999;15(1):53-62.
Marmot MG, Kogevinas M, Elston MA. Social/[13] economic status and disease. Annu Rev Public Health 1987;8:111-135.
Barker DJP, Osmond C. Infant mortality, [14] childhood nutrition, and ischaemic heart disease in England and Wales. Lancet 1986;327:1077-1081.
Dengue.[15] Casos autóctones por DIR e município. Centro de Vigilância Epidemiológica. Secretaria de Estado da Saúde. São Paulo. Avaiable at: http: http://www.cve.saude.sp.gov.br/htm/zoo/Den_dir06.htm. Accessed on March 13th, 2010.
Manual de vigilância Entomológica de [16] Aedes aegypti, Superintendência de Controle de Endemias, Secretaria de Estado da Saúde, 1997.
Clark GG, Seda H, Gubler DJ. Use of the “CDC [17] backpack aspirator” for surveillance of Aedes aegypti in San Juan, Puerto Rico. J Am Mosq Control Assoc 1994;10(1):119-24.
Focks DA, Chadee DD. Pupal survey: an [18] epidemiologically significant surveillance method for Aedes aegypti: an example using data from Trinidad. Am J Trop Med Hyg 1997; 56(2):159-67.
Focks DA. [19] A review of entomological sampling methods and indicators for dengue vectors. Document no. TDR/IDE/ Den/03.1. Geneva: World Health Organization, 2002.
Strickman D, Kittayapong P. Dengue and its [20] vectors in Thailand: calculated transmission risk from total pupal counts of Aedes aegypti and association of wing-length measurements with aspects of the larval habitat. Am J Trop Med Hyg 2003;68(2):209-17.
Nathan MB, Focks DA, Kroeger A. Pupal [21] demographic surveys to inform dengue-vector control. Ann Trop Med Parasitol 2006;Suppl. 1:S1-S3.
Mazine CAB, Macoris MLG, Andrighetti MTM, [22] Yasumaro S, Silva ME, Nelson MJ, Winch PJ. Disposable containers as larval habitats for Aedes aegypti in a city wit regular refuse collection: A study in Marilia, Sao Paulo State, Brazil. Acta Tropica 1996;62:1-13.
Macoris MLG, Mazine CAB, Andrighetti MTM, [23] Yasumaro S, Silva ME, Nelson MJ, Winch PJ. Factors favoring houseplant container infestation with Aedes aegypti larvae in Marilia, Sao Paulo, Brazil. Pan Am Jour Public Health 1997;1(4):280-286.
Yasumaro S, Silva ME, Mazine CAB, Andrighetti [24] MTM, Macoris MLG, Winch P. Community involvement in a dengue prevention project in Marilia, São Paulo State, Brazil. Human Organization 1998;57(2):209-214.

176 Dengue Bulletin – Volume 33, 2009
The control of Aedes aegypti for water access in households: Case studies towards a school-based
education programme through the use of net coversJoão Bosco Jardima#, Héliton da Silva Barrosa, Caroline Macedo Gonçalvesb,
Paulo Filemon Paolucci Pimentab and Virgínia T. Schalla#
aLaboratory of Health and Environmental Education, René Rachou Research Center, Fundação Oswaldo Cruz (Fiocruz Minas), Belo Horizonte, Minas Gerais, Brazil
bLaboratory of Medical Entomology, René Rachou Research Center, Fundação Oswaldo Cruz (Fiocruz Minas), Belo Horizonte, Minas Gerais, Brazil
Abstract
We report the progress made so far towards developing a school-based education programme for controlling Aedes aegypti oviposition in household flowerpots through the use of a net cover (evidengue®) to seal off the flowerpot saucer. A core feature of this programme is the association of evidengue® delivery with a basic package of oral and written information on dengue in classrooms. The flowerpot saucer is one of the most common type of water-bearing containers positive for the larvae of Ae. aegypti in the south-eastern region of Brazil. We present the results of a preliminary laboratory efficacy evaluation of evidengue® and of an inter-group, experimental, exploratory trial in which the evidengue® delivery was associated with educational information by means of a lecture and/or a leaflet in a school situated in a dengue-endemic area. The results are encouraging in both cases: (i) evidengue® has shown to be an efficacious tool to prevent ovipositing female access to flowerpot saucers in the laboratory; and (ii) despite the small numbers of students involved in the trial, one of the experimental groups yielded 85.7% of evidengue®-user households among those students whose households had flowerpot saucers. Use of evidengue® was maintained for at least 60 days, the period of data collection.
Keywords: Aedes aegypti; dengue prevention; flowerpot saucer; evidengue®, correctness of use; health education; school-based programme.
#E-mail: [email protected], [email protected]
IntroductionIndoor plant growing is a valued and prevalent habit in Brazil, and it is rare to find a home in urban and suburban areas of the country’s south-eastern region (states
of São Paulo, Rio de Janeiro, Espírito Santo and Minas Gerais) without a plant flowerpot and its associated saucer to collect water under it. Household surveys have frequently identified the flowerpot saucer as one of the most common water-bearing containers

Dengue Bulletin – Volume 33, 2009 177
The control of Aedes aegypti for water access in households
positive for the larvae of Aedes aegypti in this region.[1,2]
Mosquito-proof net covers have been evaluated as physical barriers to prevent ovipositing Ae. aegypti access to water storage containers,[3,4,5] buckets of rainwater[6] and flowerpot saucers.[7] Among a variety of sanitation measures for dengue prevention,[8] net covers constitute a basic method to control oviposition and hence prevent the development of this mosquito vector in water-bearing containers.[9] The implementation of their use as prevention tools in households of dengue-endemic areas, however, is far from simple. Not only must the efficacy of a candidate cover be previously demonstrated in laboratory and field trials, but also an efficient scheme of educational information on dengue must be associated with its distribution in order to stimulate residents to: (i) use it collectively and extensively; (ii) use it correctly; and (iii) maintain its use over a long period of time. Correct use of this device is an essential skill for its implementation.[7]
The present paper describes the progress made so far towards developing a school-based health education programme for controlling Ae. aegypti oviposition in household flowerpot saucers through the use of a net cover (evidengue®) designed to seal off the
same. Highlights include: (i) main features of evidengue®; (ii) an account of preliminary laboratory efficacy evaluations of the cover; and (iii) an experimental, exploratory field trial in a dengue-endemic area in Brazil. The trial aimed at quantifying students’ households as users of evidengue® after associating its delivery with a basic package of oral and/or written information about the disease in a classroom. Lastly, we discuss the educational nature of the programme and its efficacy as a complement to vector control initiatives at the household level.
Evidengue® – the net cover
Evidengue® is a circular mosquito-proof net cover, manufactured with a polyester resin mesh equal to or smaller than 2 mm x 1 mm (Figure 1a). Its upper structure has a frill along the aperture brim, where a strap of the same material and a rubber band are embedded. The strap allows the user to adjust tightly the cover to the flowerpot wall, whereas the rubber band helps to keep the brim fastened. When used correctly, evidengue® seals off the saucer completely without leaving gaps for access of ovipositing Ae. aegypti (Figure 1b). Evidengue®*, then, can be characterized as a sealing cover for flowerpot saucers.
Figure: (a) Evidengue® cover; (b) flowerpot saucer being sealed up with the cover; (c) correct use; (d) partially-correct; (e) incorrect use
* A prototype of the cover was registered as a utility model at the Brazilian National Institute for Industrial Property in 2003. This prototype is now being used at the Oswaldo Cruz Foundation in the state of Minas Gerais as an experi-mental model of a research line dedicated to simultaneous development of tools and educational programmes for protecting household water containers.

178 Dengue Bulletin – Volume 33, 2009
The control of Aedes aegypti for water access in households
Preliminary laboratory evaluations
A full account of a preliminary efficacy evaluation of evidengue® was published elsewhere.[7] In short, two black plastic flowerpot saucers, each containing 220 ml of unchlorinated water, were individually wrapped in the cover and placed with their respective pots in two entomological cages (A and B), one in each cage. One identical set of a flowerpot and a saucer, with the same amount of unchlorinated water, was placed without evidengue® in a third cage (C). Twenty gravid female Ae. aegypti, bred in the laboratory’s insectary, were placed into each cage four days after receiving a blood meal from an anaesthetized mouse. Upon the opening of the cages, no female was found inside the evidengues®. In cage C, there were eggs on the portion of the pot wall immediately above the saucer’s waterline.
A n o t h e r l a b o r a t o r y e v a l u a t i o n (unpublished data) was performed with five identical sets of a flowerpot and a saucer. This time, in addition to replicating the previous procedure with a greater number of saucers, we also evaluated evidengue®’s preventive efficacy under conditions of skilled and unskilled use. Four identical black plastic flowerpot saucers 11 cm in diameter, each containing 200 ml of unchlorinated water, were individually wrapped with the evidengue® and placed with their respective pots in four (A to D) entomological cages (40 cmx40 cmx40 cm), one in each cage.
In cages A and B, the strap of the aperture brim was fastened tightly; in cages C and D, the strap-fitting left gaps between the brim and the pot wall that could allow ovipositing females
access to the saucers. In cage C, although the aperture brim was fastened to the pot wall by the rubber band, the strap did not form a tight knot to seal completely the cover to the vessel; in cage D, the strap was completely loose. Such an arrangement allowed to compare evidengue® efficacy under three conditions of use in the laboratory: correct use (tightly-fastened strap) in cages A and B (Figure 1c); partially-correct use (strap without tight knot) in cage C (Figure 1d); and incorrect use (loose strap) in cage D (Figure 1e). A control set of flowerpot and saucer with the same amount of unchlorinated water was placed without the evidengue® in the fifth cage (E) and, as before, 20 gravid female Ae. aegypti were released into each cage four days after receiving a blood meal from an anaesthetized mouse. Eggs were not counted.
The results of this evaluation can be summarized as follows: no female was found inside the evidengues® in cages A and B; one female was found in the saucer of cage C, and ten in the saucer of cage D; in cage E (control), four females were found in the saucer. All other females were found dispersed either on the plant leaves, on the pot soil or on the bottom of the cages, except for cages A and D, where four and one females, respectively, were found on the external surface of evidengue®.
Taken together, the results of both evaluations showed that evidengue® is 100% effective to prevent ovipositing Ae. aegypti access to flowerpot saucers. Yet, the second evaluation showed that the preventive efficacy of the cover depends not only on design and structure factors, but also on the correctness of its use. Further investigations are needed to better quantify oviposition and to evaluate denier and texture factors.

Dengue Bulletin – Volume 33, 2009 179
The control of Aedes aegypti for water access in households
Exploratory field trial
An exploratory field trial was conducted as part of a Masters dissertation[10] which sought to determine the domicile use of evidengue® by students, after associating its delivery with a package of educational information about the disease in a classroom. The focus of the study was on three modalities of association of evidengue® delivery with oral (lecture) and/or written (leaflet) information on dengue.
Design of field trial
The trial was carried out in four classes (three 9th grade and one 8th grade) of a public school in the Venda Nova district in the city of Belo Horizonte, Minas Gerais. The students’ households were distributed across a large low-income peri-urban dengue-endemic region. A recent household survey had shown that the flowerpot saucer is the second most frequent breeding site for Ae. aegypti in the study area.[11] The sampling comprised 115 students regularly attending classes. Their average age was 16.2 years. Prior to the study, ethical clearance and informed consent were obtained.
The programme comprised three components: lecture on dengue (LD), delivery of evidengues® (EV) and delivery of an information leaflet (LF) regarding sealing of water-bearing containers. The components were differentially associated in classes 1, 2 and 3 (hereafter called experimental groups), whereas the fourth class was considered the control group. The modalities of association were as follows: Group 1: LD+EV+LF; Group 2: LD+EV; Group 3: EV+LF; Group 4: LD. The modality of association assigned to each group was defined by the drawing of lots. The lecture instructor (see below) was not informed of the outcome of the draw.
Lecture on dengue
The 20-minute lecture was given once to each group, during regular school hours and in the students’ own classrooms. The instructor (second author) had no link whatsoever with the school. The lecture content comprised six topics: (i) concept of dengue; (ii) symptoms of the disease; (iii) forms of clinical manifestation; (iv) transmission; (v) life cycle of Ae. aegypti; and (vi) prevention. Short texts (maximum 94 characters) and pictures on these topics were projected onto a screen using the following multimedia: (i) 35 powerpoint slides; (ii) five segments of a professional video about the vector;[12] and (iii) a 70-second black-and-white domestic video in which the instructor demonstrated the correct placement of evidengue® on a violet flowerpot. The texts and pictures related to the cover were part of the topic of prevention. There was no practice of evidengue® placement. All of the instructor’s lines referred to the texts and images showed on the screen, and most of the times the strict function performed by him was of pointing out or complementing relevant aspects of the visual information. No instruction was given about any other measure to control Ae. aegypti oviposition in household flowerpots. After the lecture, the students returned to their school routine. There was no questionnaire or discussion on the lecture topics.
The instructor’s performance was registered by four independent observers, according to a proper direct observation data collection system. The instructor’s five behaviour categories were recorded: (i) follow orally the script (say the lines of a script: see below); (ii) point out using a laser pointer (manipulate the device so as to make it project a red light on the screen); (iii) look at the group (make eye contact and/or head movement either from the screen, the keyboard or the instructor’s

180 Dengue Bulletin – Volume 33, 2009
The control of Aedes aegypti for water access in households
script towards one or more students); (iv) call attention (single out some aspect shown on screen using one of the following verbs: notice, see, observe, look); and (v) change the slide (handle the computer mouse or keyboard and substitute the slide projected on the screen). The four observers were previously trained to record these categories. Records were entered in two distinct paper-and-pencil observation forms. Two of the observers, working independently, recorded those speech lines which identified the instructor’s compliance with a pre-established sequence of topics for each slide according to a printed script of the lecture topics, slide by slide. The same pair of observers recorded the time the instructor spent on each slide. The other pair, also working independently, recorded the frequency of the other categories.
Delivery of evidengues®
Four evidengues® were delivered to each student of the experimental groups, irrespective of the number of flowerpots that their households could have. The evidengues®
were numbered in series, which allowed their identification during data collection. To increase the use likelihood, evidengues® were delivered in three sizes: two small (aperture diametre of about 18 cm), one medium (35 cm), one large (45 cm). These sizes encompass the majority of flowerpot saucers found in Brazilian homes. In groups 1 and 2, evidengues® were delivered at the end of the lecture; in group 3 (not submitted to LD component), delivery was made at the end of a regular class after the lecture was given to the other groups. Group 4 was not informed as to where to get the evidengue®. In the event of a question in this sense, the instructor plainly replied that the cover was on test and therefore not available.
Delivery of the leaflet
The leaflet (15 cm x 21 cm, four colours) stressed the importance of sealing up household water containers. In its upper half, a three-line text heading enunciated: “It is dangerous to leave pots uncovered. To prevent dengue, it is necessary to seal up. It is not enough just to cover water-bearing containers.” Underneath, three photos exemplified situations of uncovered containers, partially (incorrectly) covered, and correctly sealed. In the lower half, a photo of an empty evidengue®, sided by a secondary text (“Evidengue® seals up flowerpot saucers completely”), presented the cover to students. Underneath, three photos exemplified the situations of containers uncovered, partially covered and correctly sealed in three household flowerpots wrapped with evidengue®.
Data collection
Evidengue® use was verified by direct observation in the students’ households. Use was defined as the observation of a flowerpot wrapped with an evidengue®. Only flowerpots wrapped with the cover were registered, and observation of at least one cover in use (that is, irrespective of the number of covers provided in the classroom) was enough to consider the household as an evidengue® user. Residents were not questioned about who in the household was responsible for placing the cover(s), or about any other behaviour related to its use.
Verifications were done by a trained male health worker and an assistant researcher (the researcher acted as independent observer in 30% of households). Both of them had fieldwork experience. Two series of verification visits were conducted: the first one started

Dengue Bulletin – Volume 33, 2009 181
The control of Aedes aegypti for water access in households
five days after classroom intervention and lasted for three weeks; the second started 60 days after intervention and lasted for two weeks. Each household was visited once in each series. Evidengue® use was registered on a standardized paper-and-pencil form. Upon introducing himself, the health worker informed the resident that the home had been chosen among the addresses of the students registered in the school office and that he would check household interventions regarding dengue prevention. Knowledge, attitudes and other practices relating to vector control were not surveyed.
Results
The main outcome measure was the number of evidengue® user households among those households that had flowerpot saucers. Of the 115 households that made up the sample, 98 were visited in the first verification and 46 (46.9%) of them had flowerpot(s) with saucer (of those which were not visited, 13 had incorrect addresses and four were empty).
In the second verification, 94 households were visited and 44 (46.8%) of them had flowerpot(s) with saucer (four other were empty in addition to the 17 not previously visited). The Table shows, for each group, the number of students who received evidengues®, the number of households in which at least one flowerpot with saucer was verified, and the number of evidengue® user households in both verifications. It can be seen that the number of user households is higher in groups 1 and 3 (modalities LD + LF + EV and EV + LF, respectively) in which there was delivery of leaflet. In the second verification, the number of user households increased in all three experimental groups, reaching 85.7% (12 out of 14) of those households with saucer in group 1. None of the households discontinued the use of the cover between verifications, which means a period of maintenance of use of 60 days or more. The great majority of flowerpots with saucer (82.7%) was observed on verandas (average 6.4 per household). Overall, the education programme resulted in 34.8% evidengue® user households in the first verification (16 out of 46 households
Table: Number of students who received evidengues®, number of households with saucer, and number of user households in each group in two verifications of cover use
GroupStudents
with evidengue®
1st verification 2nd verification
Households with saucers
User households
Households with saucer
User households
1 (LD+EV+LF) 30 10 5 14 12
2 (LD+EV) 33 13 4 13 8
3 (EV+LF) 34 15 7 11 9
4 (LD) – 8 – 6 –
Total 97 46 16 (34.8%) 44 29 (65.9%)
LD = lecture on dengue EV = delivery of evidengues® LF = delivery of the leaflet

182 Dengue Bulletin – Volume 33, 2009
The control of Aedes aegypti for water access in households
with saucer) and 65.9% in the second (29 out of 44 households with saucer). Data collected on correct use were not reliable and were discarded. Nonetheless, some level of incorrect use (5% to 10% in the three experimental groups) may be assumed.
The data on the instructor’s performance resulted in indexes of agreement between the observers (number of agreed-upon records divided by the sum of agreements and disagreements) of above 90% in the three modalities of association of the LD component. Therefore, such indexes do not indicate a trend in the instructor’s performance toward any of the groups submitted to these modalities.
Discussion
Despite the small numbers of students involved in each group, the results of this exploratory trial reflect the potential of the education programme in stimulating the use of a net cover for flowerpot saucers in households of students. Evidengue® use was verified in varied proportions in all three experimental groups irrespective of the modality of association of the components of the programme. Moreover, all early user households maintained the cover in use for a period of 60 days at least. Additional research is obviously needed to clarify the specific influence of each modality of association of the components of the programme. The results of groups 1 and 3 suggest that the leaflet may have differentially influenced the use of evidengue®; however, replication of the current modalities of the association in larger samples is required. The maintenance of use for periods longer than the 60-day inter-verification interval utilized in this study also needs further investigation.
The results refer to user households of evidengue® and do not constitute, as is obvious, entomological indicators for the evaluation of oviposition control in the whole set of flowerpot saucers that a house might have. Rather, results indicate merely the occurrence, in students’ households, of a behaviour class suitable and specific to this control through the use of evidengue®. A post-hoc analysis of the behaviours constituting this class would include the receipt of the cover by the student in school, the transportation, the handling, the wrapping of a flowerpot saucer with it, and the exposure (or re-exposure) of the container together with the vase in a place susceptible to the presence of Ae. aegypti. Although one cannot identify objectively the agent(s) of all behaviours of such a sequence, it is logically plausible to infer that at least part of them may be attributed to the student. We could not find prior school-based dengue control research addressing students’ mediation, at the household level, through the use of a net cover. The lack of prior studies explains why our exploratory field trial contained two information components, given that we could not anticipate the probability of obtaining a successful outcome with any one or the other.
A relevant aspect in the evaluation of the present results concerns the matching of the sizes of evidengues® delivered in the classroom to the sizes of the existing flowerpot saucers in households. The study did not measure a baseline by means of which this matching could have been made. In such a circumstance, it is highly likely that a number of user households express some size incompatibility between the covers delivered to students and the flowerpot saucers in their homes, to the extent that inadequate sizes of evidengue® reduced opportunities of use which otherwise would have materialized.

Dengue Bulletin – Volume 33, 2009 183
The control of Aedes aegypti for water access in households
Other limitations of the study were the short time-span of the trial, which did not allow a greater number of verifications and, therefore, was insufficient to see any change, positive or negative, in the trends of evidengue® use; and the relatively small number of households with flowerpot saucers in all experimental groups, which precluded a statistical assessment of the effects of the different modalities of association or comparisons between these trends. Moreover, the possibility exists that the previous visit had contributed to the enhancement of evidengue® use in the second verification, a limitation that the experimental procedure could not overcome.
General discussion
Although the need for health education fo r behav ioura l research i s wide ly acknowledged,[8,13] current research practices on dengue prevention have been inadequate in producing educational programmes capable of fostering effective preventive behaviours at the level of the household.[14,15] Net covers are probably the most efficacious way to prevent access of female Ae. aegypti to the interior of water-bearing domestic containers, and evidengue® seems to be a convenient method to control vector oviposition in flowerpot saucers. A variety of educational programmes have sought to establish dengue prevention behaviours among residents of endemic areas of the disease.[13,16] Although aimed at residents in their homes, some of these programmes are implemented in schools.
It is assumed that educational interventions in schools can somehow influence the behaviour of residents, using the student as mediator.[16] However, notwithstanding the presumed cost-effectiveness of such a strategy,[17] behavioural research data
on health education are insufficient to empirically support this conjecture. In fact, at the present stage of behavioural research on dengue prevention, the pursuit of educational procedures that are able to establish, through school, a minimum of preventive behaviours at the level of the household, is an area fit for detailed empirical investigations. It is in this context of relative absence of data that evidengue® has been used at the Oswaldo Cruz Foundation as an experimental model of a research line conjointly dedicated to the development of tools and school-based education programmes for protecting household water containers.
In Brazil and other dengue-endemic countries, most residents are passive participants in vector control initiatives. The simple argument we make in this paper is that a school-based programme can better educate students to actively co-participate in those initiatives by offering them a domestic tool for vector control rather than merely advising them to undertake preventive measures. The data set we presently have is, of course, too incipient to draw any conclusion about the efficaciousness of such a programme, but building on the research results in the domain of behaviour modification for injury prevention,[18] it is reasonable to argue that if a resident (student or other) behaves in ways that effectively avoid a threatening consequence, such as when he/she makes a correct use of a net cover for vector control, there may be an increased probability that such behaviour will in turn induce other preventive behaviours in the household. Our next study will test the possibility that the use of evidengue® in flowerpot saucers might be an opportunity for the generalization of the net cover use to other kinds of water-bearing containers, including larger tanks that are productive for Ae. aegypti pupae.[19] Also, maintenance of use will be investigated for longer periods.

184 Dengue Bulletin – Volume 33, 2009
The control of Aedes aegypti for water access in households
One critical aspect to be assessed is the stability of evidengue® at a height of the flowerpot which is far enough from the saucer to prevent possible lowering of the net which would enable Ae. aegypti to oviposition in puddles formed by water coming out through the mesh. On another front, field investigations of the correctness of use will be a crucial aspect for the evaluation of the efficacy of evidengue®. The importance of correct net cover use can be illustrated by a study that evaluated the efficacy of an insecticide-treated cover for large water storage containers in Cambodia.[3] Albeit a follow-up of this study found a great proportion of containers to be duly protected by the cover, 16.5% of them were still infested, an outcome that puts into question the efficacy with which residents were using the tool to seal off the container.
Evidengue® can be produced in a variety of colours and manufactured with ornamental fastenings. This may encourage residents to use them in verandas and gardens as well as inside their homes. Polyester nets are highly durable and resistant to moisture and temperature changes. This could increase their protection factor. They could be reasonably priced if manufactured in large numbers (we are presently paying to our evidengue® manufacturer the equivalent of US$ 1 for each cover) and may be widely commercialized and distributed through gardening shops and supermarkets.
We intend to keep developing the programme outlined in this paper with simultaneous further laboratory and field tests of evidengue®. The development of an efficacious education programme takes time and the evidengue® efficacy itself
depends, to a large degree, on understanding
the behavioural processes that result in its adoption and use. Looking at the matter of efficaciousness in a different perspective, one might go so far as to presume, as one reviewer of this manuscript did, that perhaps evidengue® should be insecticide-treated. This view, however, does not diminish the need to investigate the efficacy of this and other vector control tools in behavioural terms. Perhaps training methods should be used, but it seems clear that this is an area of research in health education and health behaviour domains that needs more attention.
It is curious that health education programmes are generally provident and categorical when addressing the need to use seat-belts in vehicles, condoms in sexual relations, special clothing in risk tasks and other individual and collective safety devices in ordinary circumstances of prevention. Interestingly enough, despite the weak evidence that educational information is able to convert itself into behaviours for vector control of dengue in households,[14,15] one does not see the same providence and determination to recommend the use of preventive tools when it comes to the safe use of domestic water containers by residents of endemic areas of the disease.
Acknowledgements The authors are grateful to the reviewers for their appropriate and constructive suggestions for improving the manuscript. Gratitude is extended to Daniele Abrão, Isabela de Brito Ferreira, Pedro Mafia, Stéphanie Riveros, Amanda Fonseca, Alberto Mesaque Martins and Maíra Neves Santos for their assistance in conducting our laboratory and field work.

Dengue Bulletin – Volume 33, 2009 185
The control of Aedes aegypti for water access in households
ReferencesMaciel-de-Freitas R, Marques WA, Peres RC, [1] Cunha SP, Lourenço-de-Oliveira R. Variation in Aedes aegypti (Diptera: Culicidae) container productivity in a slum and a suburban district of Rio de Janeiro during dry and wet seasons. Mem Inst Oswaldo Cruz 2007;102:489-96.
Macoris MLG, Mazine CAB, Andrighetti [2] MTM, Yasumaro S, Silva ME, Nelson MJ et al. Factors favoring houseplant container infestation with Aedes aegypti larvae in Marília, São Paulo, Brazil. Rev Panam Salud Publica 1997;1:280-286.
Kittayapong P, Strickman D. Three simple [3] devices for preventing development of Aedes aegypti larvae in water jars. Am J Trop Med Hyg 1993;49:158-165.
Socheat D, Chantha N, Setha T, Hoyer S, [4] Chang MS, Nathan MB. The development and testing of water storage jar covers in Cambodia. Dengue Bull 2004;28 Suppl:8-12.
Seng CM, Setha T, Nealon J, Chanta N, Socheat [5] D, Nathan MB. The effect of long-lasting insecticidal water container covers on field populations of Ae. aegypti (L.) mosquitoes in Cambodia. J Vector Ecol 2008;33:333-341.
Chuang HY, Huang JJ, Huang YC, Liu PL, [6] Chiu YW, Wang MC. The use of fine nets to prevent the breeding of mosquitoes on dry farmland in southern Taiwan. Acta Trop 2009;110:35-37.
Schall VT, Barros HS, Jardim JB, Secundino [7] NFC, Pimenta PFP. Dengue prevention at the household level: Preliminary evaluation of a mesh cover for flowerpot saucers. Rev Saude Publica 2009;43:895-897.
Renganathan E, Parks W, Lloyd L, Nathan [8] MB, Hosein E, Odugleh A, Clark GG, Gubler DJ, Prasittisuk C, Palmer K, San Martin J-L. Towards sustaining behavioural impact in dengue prevention and control. Dengue Bull 2003;27:6-12.
Jardim JB, Schall VT. Dengue prevention: [9] focus on proficiency. Cad Saude Publica 2009;25:2529-2530.
Barros HS. [10] Investigação de conhecimentos sobre a dengue e do índice de adoção de um recurso preventivo (capa evidengue®) no domicílio de estudantes, associados a uma ação educativa em ambiente escolar [masters dissertation]. Rio de Janeiro, RJ. Instituto Oswaldo Cruz; 2007. http://www.bdtd.cict.fiocruz.br/tedesimplificado/tde_busca/arquivo.php?codArquivo=120 – accessed 2 July 2010.
Portal PBH. Prefeitura de Belo Horizonte. [11] Sala de Notícias. Retrieved on December 17, 2009 from http://portalpbh.pbh.gov.br/pbh/ecp/comunidade.do?app=salanoticias.
Vieira G, Perin L, Romeu R, Dinis HN, Labarthe [12] NV. O mundo macro e micro do Aedes aegypti. Rio de Janeiro: Editora Fiocruz; 2006.
Elder J, Lloyd LS. [13] Achieving behaviour change for dengue control: methods, scaling-up, and sustainability. World Health Organization, Scientific Working Group, Report on Dengue 1-5 2006. http://www.who.int/tdr/publications/publications/swg_dengue_2.htm – accessed 2 July 2010.
Claro LBL, Kawa H, Cavalini LT, Rosa MLG. [14] Community participation in dengue control in Brazil. Dengue Bull 2006;30:214-221.
Heintze C, Garrido MV, Kroeger A. What do [15] community-based dengue control programmes achieve? A systematic review of published evaluations. Trans R Soc Trop Med Hyg 2007;101:317-325.
Khun S, Manderson L. Community and school-[16] based health education for dengue control in rural Cambodia: a process evaluation. PLoS Negl Trop Dis 2007;1:e143.

186 Dengue Bulletin – Volume 33, 2009
The control of Aedes aegypti for water access in households
World Bank. [17] World development report: investing in health. Washington, DC: Oxford University Press, 1993.
Geller ES, Berry TD, Ludwig TD, Evans RE, [18] Gilmore MR, Clarke. A conceptual framework for developing and evaluating behaviour change interventions for injury control. Health Educ Res 1990;5:125-137.
Focks DA, Chadee DD. Pupal survey: an [19] epidemiologically significant surveillance method for Aedes aegypti: an example using data from Trinidad. Am J Trop Med Hyg 1997;56:159-167.

Dengue Bulletin – Volume 33, 2009 187
Container survey of mosquito breeding sites in a university campus in Kuala Lumpur, Malaysia
C.D. Chena#, H.L. Leeb, S.P. Stella-Wonga, K.W. Laua and M. Sofian-Aziruna
aCenter for Tropical Biodiversity Research, Institute of Biological Sciences, Faculty of Science, University of Malaya, 50603 Kuala Lumpur, Malaysia
bMedical Entomology Unit, WHO Collaborating Centre for Vectors, Institute for Medical Research, Jalan Pahang, 50588 Kuala Lumpur, Malaysia
Abstract
A survey on container breeding sites of Aedes spp. and Culex spp. larvae was conducted in the campus of the University of Malaya, Kuala Lumpur. Sampling was carried out by dipping using pipette or dipper depending on container types. All breeding sources of mosquito larvae were grouped into eight different container types: plastic container, plastic pail, bottle, earthen plate, natural container, vase, can and concrete tank. A total of 262 containers were identified as potential breeding sites. However, only 65 containers (86.15% outdoors and 13.85% indoors) were found containing larvae. Among all types of containers, 50.00% of the total surveyed natural containers were positive with mosquito larvae, followed by plastic containers (32.77%), plastic pails (23.81%), concrete tanks (20.00%), vases (18.75%), bottles (14.71%), cans (13.33%) and earthen plates (11.90%). The collected mosquito larvae were those of Aedes albopictus (86.96%), Aedes niveus group (5.80%) and Culex quinquefasciatus (7.24%). No Aedes aegypti was found in any container in the study site. This study indicated that Ae. albopictus was capable of breeding in a wide range of container types. To control these mosquitoes, the elimination of artificial and natural containers or alteration of breeding sites in and around the campus areas should be taken into consideration. Towards this goal, the university management should practise Integrated Vector Management (IVM) in the campus.
Keywords: Aedes albopictus; Aedes niveus gp.; Culex quinquefasciatus; container; university campus; Malaysia.
# E-mail: [email protected]
Introduction
Developmental activities, especially urban development associated with rapid growth of townships, have accentuated the problem of vector-borne diseases, especially dengue, which is basically an urban disease.[1] With regard to dengue vector proliferation,
human ecology is responsible for the creation of a mosquitogenic environment. Man is directly or indirectly creating such a situation.[1] Containers are probably the most important factor determining the breeding of Aedes spp. since artificial containers are the major larval habitats in and near human habitation.[2]

188 Dengue Bulletin – Volume 33, 2009
Container survey of mosquito breeding sites in a university campus in Kuala Lumpur, Malaysia
In Malaysia, Culex quinquefasciatus is one of the mosquito species that are most studied because of their potential as vector of filariasis and also for the continuous biting nuisance caused to those living close to their larval habitats.[3]
The outbreaks of dengue are always reported in the campuses of higher learning institutions in Malaysia; few larval surveys, if any, are conducted in these campuses, and because of this very little information on the prevalence of Aedes breeding is available. Such lack of information also hampers dengue control efforts.
The present study was therefore conducted to determine the container breeding preferences of mosquitoes by larval survey in the campus of the University of Malaya, Kuala Lumpur. Such information can be used to design an effective control programme for campuses.
Materials and methods
The container survey was conducted from 15 September 2006 to 31 December 2006 in the campus of the University of Malaya (N03°07.466’, E101°39.281’), Kuala Lumpur. The campus is located in the suburb of Lembah Pantai, south-west of Kuala Lumpur. The campus has an area of 750 acres (3.0 sq. km.) and most of the main administrative buildings and faculties are located here. The surroundings of the campus have been planted with vegetation and trees, providing ideal resting sites for mosquitoes. During the survey, all the containers and reachable tree holes were examined. Larvae collection was carried out indoors and outdoors by dipping, using pipette or dipper depending on container type and location. In this study, “indoors” refers to the interior of the building while “outdoors” refers to the outside of the building but confined to
its immediate vicinity. The number, type and water condition of containers that may serve as potential breeding sites were examined and recorded. The collected larvae and pupae were kept in the laboratory for adult emergence. The emerged adult mosquitoes were then pinned and identified.
Results and discussionThe Table presents the container preferences and water quality in containers found positive with mosquito larvae, while the Figure shows the percentage of different types of containers that were positive. A total of 262 containers were identified as potential breeding sites, of which 45 were indoor and 217 were outdoor containers. However, only 65 containers (9 indoors and 56 outdoors) were found containing larvae, with plastic containers being the most dominant (number of containers = 39), followed by plastic pails (5), bottles (5), earthen plates (5), natural containers (5), vases (3), cans (2) and concrete tank (1). Out of the total number of positive containers, 86.15% were outdoors while only 13.85% were indoors, indicating thereby that containers stored with water indoors were seldom found positive and containers found outdoors were probably filled with rainwater, thus providing ideal breeding sites for mosquitoes.
Among all types of containers, 50.00% of the total surveyed natural containers were positive with mosquito larvae, followed by plastic containers (32.77%), plastic pails (23.81%), concrete tanks (20.00%), vases (18.75%), bottles (14.71%), cans (13.33%) and earthen plates (11.90%). This indicated that natural containers, especially tree holes, were the dominant breeding sites for mosquitoes in the campus. The collected mosquito larvae were those of Aedes albopictus Skuse 1894 (86.96%), Aedes niveus gp. (Eichwald, 1837) (5.80%) and Culex quinquefasciatus Say 1823 (7.24%).

Dengue Bulletin – Volume 33, 2009 189
Container survey of mosquito breeding sites in a university campus in Kuala Lumpur, Malaysia
Tabl
e: C
onta
iner
pre
fere
nces
and
wat
er q
ualit
y in
con
tain
ers
posit
ive
with
mos
quito
larv
ae in
a u
nive
rsity
cam
pus,
Kua
la L
umpu
r
Cont
aine
r ty
pes
No.
co
ntai
ners
su
rvey
ed
No.
of p
ositi
ve
cont
aine
rs
Spec
ies
of m
osqu
ito la
rvae
Wat
er q
ualit
y in
con
tain
ers
posi
tive
with
mos
quito
la
rvae
Aede
s al
bopi
ctus
Aede
s niv
eus
gp.
Cule
x
quin
quef
asci
atus
Mix
ed
bree
ding
Clea
r wat
erTu
rbid
wat
erPo
llute
d w
ater
N%
N%
N%
N%
N%
N%
N%
N%
Plas
tic
cont
aine
r11
939
32.7
735
53.8
4–
–2
3.08
2a3.
0831
47.6
95
7.69
3c4.
62
Plas
tic p
ail
215
23.8
15
7.69
––
––
––
57.
69–
––
–
Bottl
e34
514
.71
57.
69–
––
––
–5
7.69
––
––
Earth
en p
late
425
11.9
05
7.69
––
––
––
46.
161
1.54
––
Nat
ural
co
ntai
ner
105
50.0
01
1.54
23.
08–
–2b
3.08
23.
073
4.62
––
Vase
163
18.7
53
4.61
––
––
––
34.
62–
––
–
Can
152
13.3
32
3.08
––
––
––
23.
07–
––
–
Conc
rete
tank
51
20.0
0–
––
–1
1.54
––
––
––
11.
54
Tota
l26
265
24.8
156
86.1
42
3.08
34.
624
6.16
5279
.99
913
.85
46.
16
N =
num
ber o
f con
tain
ers
– =
nil
a Mix
ed b
reed
ing
of A
e. a
lbop
ictu
s an
d C
x. Q
uinq
uefa
scia
tus
b Mix
ed b
reed
ing
of A
e. a
lbop
ictu
s an
d Ae
. niv
eus
gp.
c Tw
o co
ntai
ners
with
pol
lute
d w
ater
wer
e fo
und
bree
ding
with
Cx.
qui
nque
fasc
iatu
s on
ly; w
hile
one
con
tain
er w
ith p
ollu
ted
wat
er w
as fo
und
with
mix
ed b
reed
ing
of A
e. a
lbop
ictu
s an
d C
x. q
uinq
uefa
scia
tus.

190 Dengue Bulletin – Volume 33, 2009
Container survey of mosquito breeding sites in a university campus in Kuala Lumpur, Malaysia
Our study indicated that Ae. albopictus was capable of breeding in a wide range of container types, except in concrete tanks with polluted water. Thavara et al.,[4] Preechaporn et al.[5] and Thenmozhi et al.[6] also reported that Ae. albopictus larvae were found in a wide range of artificial containers (plastic containers, earthen jars, used cans, tyres, metal boxes, latex collecting cups, mud pots, etc.) and natural containers (tree holes, coconut shells, fruit peels and plant axils). On the other hand, the vector of filarial parasite Wuchereria bancrofti, Ae. niveus gp. was found breeding only in tree holes. The occurrence of this mosquito in the university campus may be due to a small preserved forested area in the campus. Ae. niveus gp. has been reported as forest breeder in tree holes and bamboo stumps in Thailand.[7] This mosquito also plays an important role in the sylvatic cycle of dengue in Malaysia.[8] Cx. quinquefasciatus was found breeding only in plastic containers in this study. Besides plastic containers, Hribar et al.[9] and Preechaporn et al.[10] also reported that Cx. quinquefasciatus bred in several types
of water containers including ceramic vessels, metal vessels, tucker boxes, plastic and metal water barrels, and concrete water tanks.
The species’ preference was not only for the vessels in question but also for the water quality they were found breeding in. In this study, water quality was categorized into three types: clear water, turbid water, and polluted water. Clear water is defined as water free from opaqueness; turbid water as water having sediments and suspended foreign particles; and polluted water as water containing wastes and oily matter.
The condition of water in the majority of surveyed containers was clear (79.99%), followed by turbid water (13.85%) and polluted water (6.16%). Almost all the Ae. albopictus and Ae. niveus gp. larvae were found in the clear and turbid waters, indicating that Aedes larvae required clear, but not necessarily clean, water to grow and develop. This finding was similar to the findings reported by Lee and Hishamudin[11] and Chen et al.[12]
Figure: Percentage of different types of containers with mosquito larvae in a university campus, Kuala Lumpur

Dengue Bulletin – Volume 33, 2009 191
Container survey of mosquito breeding sites in a university campus in Kuala Lumpur, Malaysia
On the other hand, Cx. quinquefasciatus was found breeding only in plastic containers with polluted water. This was similarly reported by Hidayati et al.,[13] in which Cx. quinquefasciatus usually breeds in polluted and stagnant water with high organic content. However, some researchers reported that Cx. quinquefasciatus also preferred habitats with turbid water where turbidity was caused by organic matter.[14,15,16]
Besides, this study also found that the percentage of mixed breeding of Cx. quinquefasciatus and Ae. niveus gp. with Ae. albopictus accounted for 6.16% of the total examined positive containers with larvae. Mixed breeding of Aedes mosquitoes with Cx. quinquefasciatus has been reported in ovitraps by Yap[17] and Chen et al.,[18] but in very low frequency. Although mixed breeding of Ae. albopictus and Ae. aegypti was also often reported by many researchers,[18-21] no mixed breeding of Ae. albopictus and Ae. aegypti was found in this study.
No Ae. aegypti was found in any containers in our study site. This was confirmed by the ovitrap surveillance conducted by Wan-Norafikah et al.[22] in the same campus of the University of Malaya. Their study indicated that Ae. aegypti mosquitoes were found neither indoors nor outdoors in the student hostels and the surroundings of the campus, probably due to the ecology comprising mostly of vegetation, bushes and trees which are typical breeding foci of Ae. albopictus.
Ae. albopictus is a ubiquitous mosquito found breeding outdoors in Malaysia. As such, it is very likely that this mosquito is not only found breeding inside the campus but may also be found in the periphery of the campus and thus having migrated into the campus. In Malaysia, the flight range of Ae. albopictus females is similar to Ae. aegypti, which is less
than 200 metres (Lee, per. Comm.). Thus, the dispersal of Ae. albopictus is presumably quite similar to that of Ae. aegypti.
In Malaysia, rainfall has been shown to correlate with Ae. aegypti breeding and dengue transmission.[23] The relationship between these variables was partly due to the creation of more breeding sites for Ae. aegypti. Therefore, seasonal rainfall is an important attribute in determining Aedes breeding.
Water-holding containers produced by humans are the main important larval habitats for Aedes mosquito. The quality of water as well as conditions of water containers seemed to contribute to the abundance of Aedes species in the study site. Besides, water chemistry of aquatic habitats may also play a critical role in determining the survival rate of mosquitoes. The ability of gravid mosquito females to distinguish among potential oviposition sites that will or will not support the growth, development and survival of their offspring are critical to the maintenance of the mosquito population.[24]
Conclusion
Source reduction is an effective way for the community to manage the populations of many kinds of mosquitoes.[25] The elimination of artificial and natural containers or alteration of breeding sites in and around living/working areas, such as university campuses, should be taken into consideration since the presence of water in containers is probably the most important factor in determining the breeding of mosquitoes, especially Aedes. Container management should be taken up as a priority by the campus community.
The university management should practise Integrated Vector Management (IVM)

192 Dengue Bulletin – Volume 33, 2009
Container survey of mosquito breeding sites in a university campus in Kuala Lumpur, Malaysia
recommended by WHO[26] in the campus. An IVM approach takes into account the available health infrastructure and resources, and integrates all available and effective measures, whether chemical, biological or environmental. IVM also encourages effective coordination of the control activities of all sectors that have an impact on vector-borne diseases, including health, water, and solid waste and sewage disposal. An IVM approach is evidence-based and its essential feature is the development of capacity to generate local data on disease epidemiology and vector ecology. IVM integrates all available resources to achieve maximum impact on vector-borne diseases.[26] However, integration of IVM at the
level required is not a simple task. Commitment is needed from the university management to integrate IVM within the campus community and local health authorities to coordinate their efforts.
Acknowledgements
The authors thank the staff of the Medical Entomology Unit, Institute for Medical Research, Kuala Lumpur, for their field assistance. This study was supported by a research grant from the Institute of Biological Sciences, Faculty of Science, University of Malaya, Malaysia.
ReferencesDutta P, Mahanta J. Potential vectors of dengue [1] and the profile of dengue in the North-Eastern Region of India: An epidemiological perspective. Dengue Bulletin 2006;30:234-42.
Lee HL. A nationwide resurvey of the factors [2] effecting the breeding of Aedes aegypti (L.) and Aedes albopictus Skuse (Diptera: Culicidae) in urban towns of Peninsular Malaysia – 1988-1989. Tropical Biomedicine 1991;8:151-60.
Vythilingam I, Tan CH, Nazni WA. Transmission [3] potential of Wuchereria bancrofti by Culex quinquefasciatus in urban areas of Malaysia. Tropical Biomedicine 2005; 22: 83-85.
Thavara U, Tawatsin A, Chompoosri J. [4] Evaluation of attractants and egg-laying substrate preference for oviposition by Aedes albopictus (Diptera: Culicidae). Journal of Vector Ecology 2004;29(1):66-72.
Preechaporn W, Jaroensutas inee M, [5] Jaroensutasinee K. The larval ecology of Aedes aegypti and Aedes albopictus in three topographical areas of Southern Thailand. Dengue Bulletin 2006;30:204-13.
Thenmozhi V, Hiriyan JG, Tewari SC, Samuel [6] PP, Paramasivan R, Rajendran R, Mani TR, Tyagi BK. Natural vertical transmission of dengue virus in Aedes albopictus (Diptera: Culicidae) in Kerala, a Southern Indian State. Japanese Journal of Infectious Diseases. 2007;60:245-9.
Walsh JF, Molyneux DH, Bir ley MH. [7] Deforestation: effects on vector-borne diseases. Journal of Parasitology 1993;106:55-65.
Rudnick A, Lim TW. Dengue fever studies [8] in Malaysia. Bullet in of Inst i tute for Medical Research, Kuala Lumpur, Malaysia 1986;23:241.
Hriber LJ, Smith JM, Vlach JJ, Verna TN. Survey [9] of container-breeding mosquitoes from the Florida keys, Monroe county, Florida. Journal of the American Mosquito Control Association 2001;17:245-248.

Dengue Bulletin – Volume 33, 2009 193
Container survey of mosquito breeding sites in a university campus in Kuala Lumpur, Malaysia
Preechaporn W, Jaroensutas inee M, [10] Jaroensutasinee K, Saeton J. Seasonal and topographical factors affecting breeding sites of Culex larvae in Nakhon Si Thammarat, Thailand. Walailak Journal of Science & Technology 2007;4(1):23-39.
Lee HL, Hishamudin M. Nationwide [11] Aedes larval survey in urban towns of Peninsular Malaysia (1988–1989). Tropical Biomedicine 1990;7:185-8.
Chen CD, Seleena B, Masri MS, Chiang YF, [12] Lee HL, Nazni WA, Sofian-Azirun M. Dengue vector surveillance in urban residential and settlement areas in Selangor, Malaysia. Tropical Biomedicine 2005;22(1):39-43.
Hidayati H, Sofian-Azirun M, Nazni WA, [13] Lee HL. Insecticide resistance development in Culex quinquefasciatus Say, Aedes aegypti (L.) and Aedes albopictus Skuse larvae against malathion, permethrin and temephos. Tropical Biomedicine 2005;22(1):45-52.
Asimeng E, Mutinga, M. Effect of rice husbandry [14] on mosquito breeding at Mwea Rice Irrigation Scheme with reference to biocontrol strategies. Journal of the American Mosquito Control Association 1993;9:17-22.
Maxwell CA, Mohammed K, Kisumku U, [15] Curtis CF. Can vector control play a useful supplementary role against Bancroftian filariasis? Bulletin of the World Health Organization 1999;77:138-143.
Muturi EJ, Shililu JI, GU W, Jacob BG, Githure JI, [16] Novak RJ. Larval habitat dynamics and diversity of Culex mosquitoes in rice agro-ecosystem in Mwea, Kenya. American Journal of Tropical Medicine and Hygiene 2007;76(1):95-102.
Yap HH. Distribution of [17] Aedes aegypti (Linnaeus) and Aedes albopictus (Skuse) in small towns and villages of Penang Island, Malaysia – an ovitrap survey. Southeast Asian Journal of Tropical Medicine and Public Health 1975;6(4):519-24.
Chen CD, Nazni WA, Lee HL, Seleena B, [18] Masri MS, Chiang YF, Sofian-Azirun M. Mixed breeding of Aedes aegypti (L.) and Aedes albopictus Skuse in four dengue endemic areas in Kuala Lumpur and Selangor, Malaysia. Tropical Biomedicine 2006;23(2):224-7.
Chan KL, Chan YC, Ho BC. [19] Aedes aegypti (L.) and Aedes albopictus (Skuse) in Singapore city. IV. Competition between species. Bulletin of the World Health Organization 1971;44:643-9.
Yap HH, Thiruvengadam V. Relative abundance [20] of Aedes aegypti (Linnaeus) and Aedes albopictus (Skuse) in different habitats. Medical Journal of Malaysia 1979;34:76-9.
Chang MS, Jute N. Breeding of [21] Aedes aegypti (L.) and Aedes albopictus (Skuse) in urban housing of Sibu Town, Sarawak. Southeast Asian Journal of Tropical Medicine and Public Health 1994;25(3):543-8.
Wan-Norafikah O, Chen CD, Soh HN, Lee [22] HL, Nazni WA, Sofian-Azirun M. Surveillance of Aedes mosquitoes in a university campus in Kuala Lumpur, Malaysia. Tropical Biomedicine 2009;26(2):206-15.
Foo LC, Lim TW, Lee HL, Fang R. Rainfall, [23] abundance of Aedes aegypti and dengue infection in Selangor, Malaysia. Southeast Asian Journal of Tropical Medicine and Public Health 1985;16:560-8.
Chen CD, Nazni WA, Seleena B, Moo JY, [24] Azizah M, Lee HL. Comparative oviposition preferences of Aedes (Stegomyia) aegypti (L.) to water from storm water drains and seasoned tap water. Dengue Bulletin 2007;31:124-30.
Dame D, Fasulo TR. [25] Mosquitoes. In: Public-health pesticide applicator training manual for USA and its territories. Gainesville: University of Florida, 2003.
World Health Organization. [26] Global strategic framework for integrated vector management. Geneva, 2004. Document No. WHO/CDS/CPE/PVC/2004.10.

194 Dengue Bulletin – Volume 33, 2009
Detection of insecticide resistance in Aedes aegypti to organophosphate and synthetic pyrethroid compounds
in the north-east of ThailandS. Pimsamarna, W. Sornpengb#, S. Akksilpb, P. Paepornc
and M. Limpawitthayakulb
aDepartment of Plants and Agricultural Resources, Khon Kaen University, Thailand
bOffice of Diseases Prevention and Control 7th, Ubon Ratchathani Province, Ministry of Public Health, Thailand
cNational Institute of Health, Ministry of Public Health, Thailand
Abstract
Aedes aegypti is a vector of dengue fever, which is a public health problem in the north-east of Thailand. During epidemics, emphasis is laid on the use of insecticides for the control of the disease. A survey of insecticides used was carried out in eight provinces, namely Amnat Charoen, Kalasin, Mukdahan, Nakhon Phanom, Sakon Nakhon, Si Sa Ket, Ubon Ratchathani and Yasothon. The study indicated that temephos was the only chemical used for the control of mosquito larvae while cypermethrin was used as space spray for the control of adult mosquitoes. Based on the results of resistance ratio of Ae. aegypti in comparison to the susceptible Bora Bora strain, Ae. aegypti showed greater resistance to deltamethrin and permethrin at a high level, with a 10-fold resistance ratio. Regarding the resistance in all study areas of Ae. aegypti to malathion and fenitrothion, it varied from low to medium levels. With biochemical assay, the results demonstrated that monooxygenase and esterase activity of Ae. aegypti had increased in all strains. Nevertheless, the glutathione-S-transferase activity had increased in seven strains. Therefore, monooxygenase, esterase and glutathione-S-transferase activity seems to be associated with mechanisms for creating pyrethroid-resistance in Ae. aegypti. These chemicals were used for the control of adult mosquitoes, which showed a high resistance ratio in these areas. Consequently, the continued use of these insecticides would lead to operational failure.
Keywords: Aedes aegypti; insecticide; north-east Thailand; resistance.
#E-mail: [email protected]
IntroductionDengue fever, transmitted by Aedes aegypti, is a public health problem in the north-east of Thailand. Vector control by reducing
Ae. aegypti population is at present the only viable option available to control the disease. Strategies to do so include environmental management and biological and chemical control. The north-east of Thailand has

Dengue Bulletin – Volume 33, 2009 195
Detection of insecticide-resistance in Ae. aegypti in the north-east of Thailand
relied mainly on the use of organophosphate and synthetic pyrethroids. During dengue epidemics, deltamethrin and cypermethrin were the main synthetic pyrethroids used to control adult Aedes mosquitoes through mass spraying,[1] which resulted in the vector’s resistance to pyrethroids. This is a worldwide phenomenon which is making control of vector-borne diseases difficult.
The increase in the development of mosquito resistance is the primary concern of all control programmes. Common insecticide-resistance mechanisms include alteration of target sites and increased enzyme activities of non-specific esterase, glutathione-S-transferase (GSTs) and P450-mediated monooxygenases. [2]
The major metabolic enzymes involved in resistance against pyrethroids in insects include P450-mediated monooxygenases, evaluated non-specific esterase, and reduced sensitivity of sodium ion channels along nerve axons. Recently, several cases of field–associated resistance have been reported in Ae. aegypti against pyrethroid products.[3] Resistance to pyrethroids, including permethrin and deltamethrin in Ae. aegypti, were documented in the north-east provinces of Thailand (see Figure). An attempt was made to lay bare the determinants, the level of resistance of Ae. aegypti to chemicals by susceptibility test, and study the changes in the level of enzymes that are related to the reduction and detoxification of chemical toxins of Ae. aegypti.
Figure: Map of study area in north-east Thailand

196 Dengue Bulletin – Volume 33, 2009
Detection of insecticide-resistance in Ae. aegypti in the north-east of Thailand
Materials and methods
Survey of insecticides
Urban respondents from eight sites, Amnat Charoen, Kalasin, Mukdahan, Nakhon Phanom, Sakon Nakhon, Si Sa Ket, Ubon Ratchathani and Yasothon, were interviewed by using a questionnaire in a cross-sectional survey on the type of insecticides used to control larvae and adult Ae. aegypti. The key informants were the officials who were responsible for the control of contagious diseases at municipal offices and district administration offices in these areas. The survey took place between May and July 2007.
Study of resistance to insecticides
Testing of susceptibility of Ae. aegypti to chemicals was carried out and the results were interpreted, as recommended by the World Health Organization.[4] The adult mosquitoes used were reared from larvae collected from the study areas to generation F1. Fully-grown females aged 3–5 days were fed on 5% sugar before testing, and then placed inside an exposure tube that was lined with chemically-treated paper. Each tube contained 25 mosquitoes and there were four tubes in all. The mosquitoes were left in the tubes for 15, 30, 60 and 120 minutes with the temperature at 25±2 °C. After the exposure time, the test mosquitoes were transferred to the holding tubes and cotton pad soaked in 10% sugar solution was provided. The mortality was recorded after 24 hours.
Interpretation of the results of the ••bioassay data were pooled and LT50 was obtained by probit analysis.[5]
Resistance ratio (RR•• 50) was calculated by comparing LT50 with LT50 of sus-ceptible strain.
LT50 resistance strainResistance ratio, RR50 = LT50 susceptible strain*
Note: *The susceptible strain (Bora Bora) from the Faculty of Tropical Medicine, Mahidol University.
Biochemical assay
Protein assay
The total protein content of individual Ae. aegypti mosquitoes was determined using a commercial protein (Bio Rad) assay system. Results were compared with a derived standard curve. The plates were read after five minutes using an ELISA plate reader at 570 nm wavelength.
Monooxygenase assay
The procedure described by Valule et al. (1999)[6]
was followed with only minor modifications. Fresh individual mosquitoes were homogenized in 50 ml distilled water in a 1 ml plastic vial. Homogenates were diluted with additional 150 µl distilled water. 20 µl of each homogenate was transferred to a microplate followed by the addition of 80 µl 0.0625 M potassium phosphate buffer (PPB) at pH 7.0. A solution of 0.01 g of 3,3’,5,5’-tetramethylbenzidine (TMBZ) in 5 ml methanol was prepared and a 0.25 M sodium acetate buffer (pH 5.0) was added. Following this, 200 µl of TMBZ solution was added with the 100 µl of mosquito homogenate plus PPB in each well followed by 25 µl of 3% hydrogen peroxide. The plates were read after 5 and 10 minutes using an ELISA plate reader at 620 nm wavelength.

Dengue Bulletin – Volume 33, 2009 197
Detection of insecticide-resistance in Ae. aegypti in the north-east of Thailand
Esterase assay
In the method of Peiris and Hemingway (1990),[7] the test could begin by adding 30 mM -naphthyl acetate that had been previously prepared at a fixed quantity in each well and leaving it at a temperature of 30 °C for 15 minutes. Following that, 0.1% fast blue B-SDS was added and then a 20 µl sample of the mosquito that had been separated was added to each well and it was left at room temperature at 25±2 °C. The test was then carried out using a microplate reader and the absorbance value was read at 620 nm.
Glutathione-S-transferase (GST) assay
GST activity was assayed following Habig et al. (1974).[8] The working solutions were mixed at a ratio of 125 µl CDNB to 2.5 ml GSH. In the test, 20 µl of the prepared liquid was added to each well. Following that, a 20 µl sample of the mosquito that had been separated was
added to each well and it was left at room temperature (25±2 °C) for 15 minutes. The test was then carried out using a microplate reader and the absorbance value was read at 340 nm.
The analysis was carried out using ANOVA, which was used to compare the protein content and enzyme expression levels within and between populations. All levels of statistical significance were determined at 95% confidence limit.
Results
Survey of insecticides
A survey indicated that the only chemical used in controlling mosquito larvae was temephos while the chemical most commonly used in spraying to control adult mosquitoes was cypermethrin (Table 1).
Table 1: Types of chemicals used in all eight study provinces
Province Number of offices
Chemical used in controlling larvae
Per cent used
Chemical used in controlling adult
mosquitoes
Per cent used
Amnat Charoen 22 Temephos 100 Cypermethrin
Zetamethrin+ Dichlorvos
95.45
4.55
Total 100 Total 100
Kalasin 69 Temephos 100 Cypermethrin
Alpha-cypermethrin
Zeta-cypermethrin
Deltamethrin
Etofenprox
Zetamethrin+ Dichlorvos
71.01
1.45
1.45
5.80
1.45
18.84
Total 100 Total 100

198 Dengue Bulletin – Volume 33, 2009
Detection of insecticide-resistance in Ae. aegypti in the north-east of Thailand
Province Number of offices
Chemical used in controlling larvae
Per cent used
Chemical used in controlling adult
mosquitoes
Per cent used
Mukdahan 21 Temephos 100 Cypermethrin 100
Total 100 Total 100
Nakorn Phanom 33 Temephos 100 Cypermethrin
Deltamethrin
Zetamethrin + Dichlorvos
90.91
6.06
3.03
Total 100 Total 100
Sakon Nakorn 66 Temephos 100 Cypermethrin
Deltamethrin
Etofenprox
Zetamethrin + Dichlorvos
90.90
6.06
1.52
1.52
Total 100 Total 100
Si Sa Ket 88 Temephos 100 Cypermethrin
Alpha-cypermethrin
Bifenthrin
Dichlorvos
Fenitrothion
Sumithion
93.18
2.27
1.14
1.14
1.14
1.14
Total 100 Total 100
Ubon Ratchathani
81 Temephos 100 Cypermethrin
Alpha-cypermethrin
Zeta-cypermethrin
Deltamethrin
Fenitrothion
Etofenprok
88.89
2.47
2.47
3.70
1.23
1.23
Total 100 Total 100
Yasothon 57 Temephos 100 Cypermethrin
Alpha-cypermethrin
Zeta-cypermethrin
Dichlorvos
87.72
8.77
1.75
1.75
Total 100 Total 100

Dengue Bulletin – Volume 33, 2009 199
Detection of insecticide-resistance in Ae. aegypti in the north-east of Thailand
Table 2: Resistance ratios of the eight strains of Ae. aegypti to four types of chemically-treated paper in the north-east of Thailand
Ae. aegypti strains
Chemicals
0.5% deltamethrin 0.75% permethrin 5% malathion 1% fenitrothion
LT50* (minute) RR50** LT50
(minute) RR50 LT50 (minute) RR50 LT50
(minute) RR50
Bora Bora 1.0821 1 6.3388 1 2.7756 1 5.9366 1
Amnat Charoen 21.9124 20.25 67.0799 10.58 21.6527 7.80 25.5521 4.30
Kalasin 15.9024 13.95 95.8781 15.13 12.4080 4.47 19.6806 3.32
Mukdahan 25.9294 23.96 80.6522 12.72 17.8287 6.42 25.1353 4.23
Nakhon Phanom 21.5179 19.88 68.3064 10.78 14.3295 5.16 21.7699 3.67
Sakon Nakhon 22.6224 20.91 70.8879 11.18 16.5196 5.95 28.9672 4.88
Si Sa Ket 26.8370 24.80 64.6004 10.19 13.8454 4.99 27.6615 4.66
Ubon Ratchathani
24.6451 22.78 74.2632 11.72 10.0989 3.64 9.1136 1.54
Yasothon 20.4058 18.86 64.3992 10.16 6.7857 2.44 11.5399 1.94
* the LT50 value of the Ae. aegypti to chemically-treated paper.** the resistance ratio (RR50) = LT50 field strain / LT50 susceptible strain.
Resistance level of Ae. aegypti to insecticides
The results of the study of Ae. aegypti’s resistance level to insecticides are included in Table 2. It may be seen that Ae. aegypti, in every area, showed high resistance to deltamethrin and permethrin. Ae. aegypti from Si Sa Ket had the highest resistance ratio to 0.05% deltamethrin at 24.80-fold. Next in order were Ae. aegypti from Mukdahan, Ubon Ratchathani and Sakon Nakhon with resistance ratios of 23.96-, 22.78-, and 20.91-fold respectively. With regard to the study of the resistance rate to treated paper with 0.75% permethrin, Ae. aegypti from Kalasin had the highest resistance ratio at
15.13-fold. Regarding the chemicals, namely 5% malathion and 1% fenitrothion, Ae. aegypti from all areas had resistance ratios that varied from low to medium.
Biochemical study
The results of testing the amount of proteins and enzymes in Ae. aegypti in all eight provinces are included in Table 3. The investigations indicated that the monooxygenase, esterase and glutathione-S-transfease enzymes had shown a tendency to increase when compared with the Bora Bora susceptible strain. Nevertheless, the glutathione-S-transferase activity had shown an increasing trend in seven strains.

200 Dengue Bulletin – Volume 33, 2009
Detection of insecticide-resistance in Ae. aegypti in the north-east of Thailand
Discussion
Dengue fever has been a public health problem in the north-east of Thailand for more than 50 years. Control of the disease in the epidemic season warrants the use of chemicals to eliminate the Ae. aegypti vector mosquito both at the larval and adult stages. Temephos 1% has been recommended for larval control because of its low oral toxicity in mammals (8600 mg/kg in male rats), and it is one of the few insecticides recommended for potable water.[9] The chemical that was mostly used to spray adult mosquitoes was cypermethrin.
The present study on the level of resistance of Ae. aegypti in all eight provinces, in comparison with that of the Bora Bora susceptible strains, indicated high resistance to deltamethrin and permethrin.[10] The
resistance ratio was found to be more than ten-fold.[11] With regard to other chemicals, viz. malathion and fenitrothion, the resistance ratio in Ae. aegypti varied from low to medium. Sathantriphop,[12] who studied the resistance phenomenon to various insecticides from four groups in a field strain of Ae. aegypti in the Baan Suan community, in Nonthaburi province, Thailand, also detected its high resistance to deltamethrin and permethrin but the vector was 100% susceptible to fenitrothion.
The results of the study by biochemical assay of the amount of protein, monooxygenase, esterase and glutathione-S-transfease enzyme had shown a tendency to increase when compared with the Bora Bora susceptible strain. This suggested that monooxygenase, esterase and glutathione-S-transfease were
Table 3: Levels of proteins and enzymes for all eight strains of Ae. aegypti in the north-east of Thailand when compared with susceptible strain
Strains (n=40 mosquitoes
for each)
Total proteinMean±SD
(mg protein/ml per mosquito (n))
MFOs (nmol product/min/
mg protein)
Esterase (nmol naphthol/min/mg protein
GSTs (nmol CDNB/min/
mg protein)
Bora Bora 0.6569+0.0919 c* 0.0561+0.3552 c 0.9175+0.6176 c 0.9701+0.7335 c
Amnat Charoen 0.8252+0.0952 ab 0.1285+0.1174 bc 1.4279+0.2225 a 1.2642+0.0481 b
Kalasin 0.6747+0.0936 bc 0.1054+0.0646 b 1.3357+0.1307 b 0.9832+0.0824 c
Mukdahan 0.8604+0.1340 a 0.3846+0.1913 a 1.3962+0.1315 b 1.4897+0.0843 a
Nakhon Phanom 0.8613+0.1254 a 0.2257+0.1623 bc 1.3409+0.1234 b 1.4941+0.0614 a
Sakon Nakhon 0.8236+0.1311 ab 0.1184+0.0957 b 1.3970+0.0815 b 0.9577+0.0741 c
Si Sa Ket 0.7143+0.1386 b 0.0573+0.0139 a 1.5420+0.0498 a 1.1241+0.0348 b
Ubon Ratchathani 0.6891+0.1129 bc 0.0697+0.0446 a 1.4553+0.0844 a 1.1076+0.0357 b
Yasothon 0.7774+0.1008 b 0.1331+0.0927 bc 1.3223+0.1346 b 1.2661+0.0249 b
*In every column that ends with the same letter, there is no statistical difference from a comparison using the Duncan multiple range test at a confidence level of 95%.

Dengue Bulletin – Volume 33, 2009 201
Detection of insecticide-resistance in Ae. aegypti in the north-east of Thailand
associated with permethrin and deltamethrin resistance in Ae. aegypti. Resistance to insecticides usually appears to be unstable, and is associated with a genetic cost in the absence of selection.[13,14] This has occurred due to other resistance-creating mechanisms apart from the increase of enzymes that detoxify the aforementioned insecticidal toxins. Regular monitoring of the mosquitoes’ susceptibility to the most widely used insecticides is necessary to ensure an appropriate choice of chemicals. Introduction of inappropriate insecticides without a proper understanding of
the prevailing resistance mechanisms may lead to operational control failure. Moreover, early detection and knowledge of the resistance status as well as the underlying mechanisms in vector mosquitoes are essential for effective long-term control of Ae. aegypti.
Acknowledgements
This research was supported in part by the graduate school, Khon Kaen University, and the WHO Country Office, Thailand.
ReferencesDepartment of Communicable Disease [1] Control, Ministry of Public Health, Thailand. Vector borne disease annual report 2002-2003. Nonthaburi: Bureau of Vector Borne Diseases, 2003.
Hemingway J, Ranson H. Insecticide resistance [2] in insect vectors of human disease. Annual Review of Entomology 2000;45:371-91.
Scott JA. The molecular genetics of resistance: [3] resistance as a response to stress. Symposium on pesticide resistance. Insect biochemistry and molecular biology 1999;29:757-77.
World Health Organizat ion. [4] Dengue hemorrhagic fever: diagnosis, treatment and control. Geneva, 1986.
Finney JD. [5] Probit analysis. 3rd ed. Cambridge: The University Press, 1971.
Vulule JM, Beach RF, Atieli FK, McAllister [6] JC, Brogdon WG, Roberts JM, et al. Elevated oxidase and esterase levels associated with permethrin tolerance in Anopheles gambiae from Kenyan villages using permethrin impregnated nets. Medical Veterinary and Entomology 1999;13:239-44.
Peiris HTR, Hemingway J. Temephos resistance [7] and the associated cross resistance spectrum in a strain of Culex quinquefasciatus from Peliyagoda, Sri Lanka. Bulletin of Entomological Research 1990;80:49-55.
Habig WH, Padst MJ, Jakody WB. Glutathione [8] S-transferase the first enzymatic step in mercapturic acid formation. The Journal of Biological Chemistry 1974;249(22):7130-9.
World Health Organziation, Regional Office [9] for the Pan American. Dengue and dengue hemorrhagic fever in the Americas: guidelines for prevention and control. Washington, DC: PAHO, 1994. pp. 548.
Paeporn P, Supaphathom K, Srisawat S, [10] Boonyabancha, Phanurai P. Efficacy of aerosol insecticide product. Bull Dept Med Sci 1996;38:37-43.
Mazzari MB, Georghiou GP. Characterization [11] of resistance to organophoshate, carbamate, and pyrethroid insecticides in field populations of Aedes aegypti from Venezuela. Journal of the American Mosquito Control Association 1995;11:315-22.

202 Dengue Bulletin – Volume 33, 2009
Detection of insecticide-resistance in Ae. aegypti in the north-east of Thailand
Sathantriphop S, Paeporn P, Supaphathom [12] K. Detection of insecticides resistance status in Culex quinquefasciatus and Aedes aegypti to four major groups of insecticides. Tropical Biomedicine 2006;23(1):97-101.
Hemingway J, Hawkes NJ, McCarroll L, [13] Ranson H. The molecular basis of insecticide resistance in mosquitoes. Insect biochemistry and molecular biology 2004;34:653-65.
World Health Organization. [14] Instructions for determining the susceptibility or resistance of adult mosquitoes to organochlorine, organophosphate and carbamate insecticide-diagnostic test. Document no. WHO/VBC/81-806. Geneva: WHO, 1981.

Dengue Bulletin – Volume 33, 2009 203
Evaluation of a “fogging” canister for indoor elimination of adult Aedes aegypti
Pang Sook Chenga, Foo Siew Yoonga, Png Ah Baha, Deng Lua, Lam-Phua Sai Geka, Tang Choon Siangb and Ng Lee Chinga#
aEnvironmental Health Institute, National Environment Agency, 11 Biopolis Way #06-05/08, Helios Block, Singapore 138667
bEnvironmental Health Department, National Environment Agency (EHD), 40 Scotts Road, Environment Building, #21-00, Singapore 228231
Abstract
Aedes aegypti is the primary vector for dengue transmission in Singapore. In response to dengue transmission, ultra-low volume (ULV) spraying of insecticides in apartments in transmission areas is used to supplement source-reduction effort. This study seeks to determine the efficacy of Mozzie Zap, a “fogger” in an aerosol canister, which contains 1.7% cypermethrin and 3.4% piperonyl butoxide (PBO, a pesticide synergist). The product, which has the unique continual release mechanism, was tested using two canisters in a two-bedroom apartment block. Caged female adult mosquitoes and magnesium oxide slides were positioned prior to the release of the chemicals. All bioassays and slides were exposed for 30 minutes before removal. Subsequent residual activity tests were carried out using WHO contact bioassay cones on horizontal (floor) and vertical (wall) surfaces. Mozzie Zap’s fog produced fog of effective droplet size (average VMD=24.32 µm), and achieved 100% mortality against Ae. aegypti. However, no residual activity on wall surfaces (3.1% on Day 2) was observed.
Keywords: Aedes aegypti; ULV spray; apartments; Singapore.
# E-mail: [email protected]; Tel.: +65 6771 9108; Fax: +65 6777 8029
Introduction
Aedes aegypti is the primary vector of dengue transmission in Singapore. A programme based on source reduction through public education backed by law enforcement was set in place in 1970.[1] This led to a period of low dengue incidence that lasted for more than a decade. However, from the 1990s, the incidence of dengue has increased despite the
control measures. Possible contributory factors include increase in human population and their activities, lowered immunity levels and increased frequency of travel. To address the dengue challenge, Singapore’s dengue control programme has evolved into an integrated programme that focuses on inter-epidemic field and laboratory surveillance, coupled with intersectoral cooperation for source reduction. The programme is continuously evolving

204 Dengue Bulletin – Volume 33, 2009
Evaluation of a “fogging” canister for indoor elimination of adult Aedes aegypti
through operational research and exploration, and the application of innovative products which are accepted by the community and can be used easily.
In response to dengue transmission, ultra-low volume (ULV) spraying of insecticides in apartments where transmission takes place is used to supplement source-reduction efforts.[2] However, ULV spraying causes inconvenience. The equipment requires a power source as well as trained officers to operate. The inconvenience caused and the negative perceptions about fogging often lead to resistance by residents.
We thus explored the effectiveness of Mozzie Zap, which offers an alternative insecticide delivery mechanism: continuous release of chemicals through an aerosol canister. The canister contains 1.7% cypermethrin and 3.4% piperonyl butoxide (PBO) with citrus scent. PBO, a pesticide synergist, is typically added to synthetic pyrethroids and natural pyrethrins to negate the effect of resistance to the insecticides. It is a well-known inhibitor of enzymes that are necessary for detoxification of chemicals and transformation of hormones.[3] This study aimed to determine the feasibility of using Mozzie Zap, which has the unique design of continual release mechanism, in Singapore’s typical housing units for the control of dengue and chikungunya vector, Ae. aegypti.
Materials and methods
Site
Three two-bedroom, government-built apartments, each having an area of 71.27 sq. metre, were used as test sites for both efficacy and residual tests. The walls within each unit are made of painted plastered cement while floors are made of wooden parquet, ceramic
tiles and marble tiles. Two units were used for Mozzie Zap spraying and one unit was left as control.
Mosquitoes
Ae. aegypti larvae, collected throughout the island of Singapore, were reared into adults in the insectary. Unfed, 3–5-day-old F2 progeny of the mosquitoes were used to test the efficacy and residual effect of Mozzie Zap.
Efficacy test
Mosquitoes were transferred into cylindrical mesh cages (5 cm diameter × 15 cm height) prior to the spraying day. Two cans of Mozzie Zap were used for each flat as recommended by the supplier (Trojan Hospitality Asia Pte Ltd). Cages were hung at 11 randomly selected points, approximately 1.2 m high from ground level, before the canister was triggered to release the chemicals. All windows and the main door leading to the corridor were shut during the entire exposure period. Thirty minutes after the release of the insecticide, mosquitoes were removed from the apartments and transferred instantaneously into clean cups with 10% sucrose as food source. Per cent mortalities at removal, 10 minutes, 1 hour and 24 hours were scored. Control was carried out in a similar manner except that no insecticide was released in the apartment.
Residual test
WHO guidelines for testing mosquito adulticides for indoor residual spraying and treatment of mosquito nets were adopted for this study with little modification.[4] The residual test was done two days after the spraying. Bioassay cones were attached to sixteen points identified for each apartment – 11 points on

Dengue Bulletin – Volume 33, 2009 205
Evaluation of a “fogging” canister for indoor elimination of adult Aedes aegypti
the wall surfaces and five points on the floor surfaces. Ten mosquitoes were introduced into each bioassay cone and aspirated out after 30 minutes. Ten per cent sucrose was supplied as the food source and the mortality was scored 24 hours post-exposure. The test was discontinued if the mortality dropped below 70%.[5]
Droplet size analysis
Insecticidal droplets were sampled at five points within a unit using magnesium oxide-coated slides. Distances from canister to slides ranged from 0.81 m to 6.58 m. Four slides were placed at each point, with three slides placed horizontally at 0 m (ground level), 1.2 m and 2.3 m, and one slide was placed parallel to the wall surface at a height of 1.2 m. At least 100 droplets were measured at a magnification of 100 times under a compound microscope. Droplets-size data were analysed by Sofield and Kent (1984) and expressed in terms of Volume Median Diameter (VMD), Number Median Diameter (NMD) and ratio of VMD and NMD.
Results and discussion
Mozzie Zap attained a high efficacy of 100% mortality at 24 hours post-exposure in the typical Singapore apartment. Of 220
mosquitoes exposed to Mozzie Zap aerosols, 201 mosquitoes were found moribund or dead after 30 minutes of exposure (Table 1). Knockdown effect is a unique characteristic of pyrethriods.[6] A subsequent observation of the mortality was made to ensure that the actual mortality was recorded. The mortality was observed to increase to 99.5% after one hour of post-exposure. As the cages of mosquitoes were placed as far away as 6.04 m from the canister, the total kill of mosquitoes in the room showed that aerosols from the canister were able to reach out to at least 6.04 m to kill Ae. aegypti adults.
Magnesium oxide slides were placed at allocated points, with some points in close proximity to the mosquito cages. Slides were adhered vertically (wall surfaces) and horizontally (floor surfaces) prior to spraying to capture insecticidal droplets. The results indicated that the slides placed vertically captured only a few droplets, averaging 0.4 droplets per optical field. In contrast, there were more than 200 droplets per optical field impinged on the slides that were placed horizontally at a distance between 1–2 m. Table 2 shows the inverse relation between the number of droplets and the distance from the dispensing canister. The droplet size of the aerosols ranged from 18.84 to 30.16 µm, with an average VMD value of 24.32 µm. They fall within the range considered to be the most effective droplet size for killing
Table 1: Efficacy results of Mozzie Zap against Ae. aegypti
Treatment N
No. of mosquitoes knocked down upon
removal (% knockdown)
No. of mosquitoes knocked down at 1 hr
post exposure (% knockdown)
No. of mosquitoes moribund or dead
(24-hrs % mortality)
Mozzie Zap treated 220 201 (91.4%) 219 (99.5%) 220 (100%)
Control 110 0 (0%) 0 (0%) 0 (0%)

206 Dengue Bulletin – Volume 33, 2009
Evaluation of a “fogging” canister for indoor elimination of adult Aedes aegypti
adult mosquitoes.[7] It is apparent that the droplet size upon dispensing is critical for an effective coverage of sprayed areas as large insecticidal droplets are uneconomical and have the tendency to deposit on the ground near the dispenser while droplets that are too small contain insufficient dosage to kill a mosquito. [7,8]
It is interesting to note that Mozzie Zap was very effective despite a previous finding that Ae. aegypti from Singapore, in general, has developed resistance against cypermethrin with an average RR50 and RR95 values of 53.8 and 62.3 respectively (unpublished data). This could be explained by the synergist effect of the mixture of cypermethrin and PBO included in Mozzie Zap, thus increasing the mortality rate. A significant increase in mortality rate (Synergist Ratio= 21.22) was
reported after the addition of four parts of PBO into one part of cypermethrin.[6] However, the study also showed that the level of synergism of PBO with synthetic pyrethoids depends greatly on the types of insecticide resistance mechanisms for individual insecticides, and multiple resistance mechanisms in mosquitoes increased the complexity further.
A study is underway to further understand the resistance and the effect of PBO on local mosquitoes. Nevertheless, the mixture, continuously dispensed through the novel nozzle, was found to be effective in killing local mosquitoes. Being handy, user-friendly and less intimidating, Mozzie Zap’s design may increase the acceptability of insecticide treatment of homes by residents. It will also reduce manpower needs. Approximately 80% of Singapore residents live in similarly built
Table 2: Comparison between droplet size and residual activities for floor and wall surfaces
Distance from
canister (m)Orientation
24 hours % mortality VMD (µm)
VMD/NMD
No. of droplets/
optical fieldDay 2 Day 3 Wk 1 Wk 2
0.81 Floor 90 100 100 95 30.16 1.23 243a
Wall –d – – – NAc NAc 0b
2.00 Floor 100 100 100 85 24.05 1.31 263.33b
Wall 0 – – – 11.60 1.00 2b
3.10 Floor 90 80 50 25 18.84 1.64 188a
Wall 0 – – – NAc NAc 0b
4.70 Floor 100 85 75 15.6 24.48 1.00 191a
Wall 0 – – – NAc NAc 0b
6.58 Floor 70 37.1 12.5 0 24.07 1.10 89.67a
Wall –d – – – NAc NAc 0b
a Average number of droplets per optical field on floor surfaces is 195.b Average number of droplets per optical field on wall surfaces is 0.4.c No droplet was collected on slide.d No bioassay done at these distances.

Dengue Bulletin – Volume 33, 2009 207
Evaluation of a “fogging” canister for indoor elimination of adult Aedes aegypti
government apartments, and the protocol for its usage in these apartments would be simple and straightforward.
The results of the residual activities of the insecticides on horizontal and vertical surfaces are presented in Table 3. Residual effects on vertical surfaces attained only 1.1% mortality after a two-day post-spraying for all 11 bioassay cones in both apartments. Residual test for vertical bioassay cones was stopped after Day 2 since the results were consistently low between the two apartments. Bioassays on floor surfaces showed residual activity of the insecticide three days post-spraying, with 75.4% mortality. The mortality was found to increase with a decrease in the distance from the canister (see Figure).
The behaviour of the mosquitoes in the WHO contact bioassay cones was observed during the exposure periods. No significant sign of avoidance of insecticide-treated surface
was noticed. However, rapid-flight escape to areas or surfaces without insecticides was observed, demonstrating significant contact irritancy of Ae. aegypti. The observations were consistent with other studies on Aedes response to synthetic pyrethroids.[9,10]
Table 3: Residual activities of Mozzie Zap against Ae. aegypti
SurfacesDays post-
spraying
Average % mortality at 24
hours (± standard deviation)
Wall D2 1.6 (± 2.25)
Floor
D2 91.7 (± 7.07)
D3 75.4 (± 25.42)
Wk1 64.6 (± 10.01)
Wk2 39.9 (± 9.00)
Figure : Residual activities of Mozzie Zap on different floor surfaces against Ae. aegypti
0
10
20
30
40
50
60
70
80
90
100
Day 2 Day 3 Week 1 Week 2
24 hr % mortality
6.58 m3.10 m
4.63 m
2.00 m
0.81 m

208 Dengue Bulletin – Volume 33, 2009
Evaluation of a “fogging” canister for indoor elimination of adult Aedes aegypti
Despite good residual activities shown on the floor surface, the feasibility of using Mozzie Zap’s continuous release mechanism for residual spray is uncertain, as frequent cleaning of floors and horizontal surfaces is commonly practised by most residents. Nevertheless, Mozzie Zap has the application potential for eliminating indoor Ae. aegypti in an apartment, which is especially important for dengue outbreak areas.
Acknowledgements
We would like to thank Mr Paranjothy Charles Joseph from M/s Trojan Hospitality Asia Pte. Ltd. for generously providing us with the supply of insecticides for this study. Also, we wish to extend our gratitude to the South-West Regional Office of the National Environment Agency for their assistance, and the Housing Development Board (HDB) for providing the facilities for this study.
in three strains of Aedes aegypti (Linn.) (Diptera: Culicidae) on insecticide permethrin, cypermethrin and D-allethrin. Bulletin Penelitian Kesehatan 2005;33(2):73-79.
Lofgren CS. Ultralow volume (ULV) application [7] of insecticides. The American Journal of Tropical Medicine and Hygiene 1972;21(5):819-824.
Hoffman WC, Walker TW, Smith VL, Martin [8] DE, Fritz BK. Droplet-size characterization of handheld atomization equipment typically used in vector control. Journal of American Mosquito Control Association 2007;23(3):15-32.
Chareonviriyaphap S, Monthathip K, Bangs MJ, [9] Sunaiyana S, Vithee M, Atchariya P, Wannapa S, Pongthep A. Influence of nutritional and physiological status on behavioral responses of Aedes aegypti (Diptera: Culicidae) to deltamethrin and cypermethrin. Journal of Vector Ecology 2006;31(1):81-101.
Paepron P, Supapthathom K, Sathantriphop S, [10] Chareonviriyaphap T, Yaicharoen R. Behavioral responses of deltamethrin- and permethrin-resistant strains of Aedes aegypti when exposed to permethrin in an excito-repellency test system. Dengue Bulletin 2007;31:153-159.
References
Chan KL. [1] Singapore’s dengue haemorrhagic fever control programme: a case study on the successful control of Aedes aegypti and Aedes albopictus using mainly environmental measures as a part of integrated vector control. Tokyo: Southeast Asian Medical Information Centre, 1985.
Ministry of Health, Singapore. [2] Epidemiological News Bulletin 2006:32(1):1-20.
Cox C. Insecticide synergist factsheet - [3] Piperonyl butoxide. Journal of Pesticide Reform 2002;22(2):12-20.
World Health Organization. [4] WHO guidelines for testing mosquito adulticides for indoor residual spraying and treatment of mosquito nets. Document No. WHO/CDS/NTD/WHOPES/GCDPP/2006.3. Geneva, 2006.
National Institute of Malarial Research, India. [5] Protocols for uniform evaluation of insecticide for use in vector control. http://mrcindia.org/com_pro/comm_prot.htm – accessed 30 June 2010.
Astari S, Ahmad I. Insecticide resistance and [6] effect of piperonyl butoxide as a synergist

Dengue Bulletin – Volume 33, 2009 209
Oviposition behaviour of Aedes albopictus in temephos and Bacillus thuringiensis israelensis-treated ovitraps
W.A. Naznia#, H.L. Leea, W.M. Wan Rozitab, A.C. Liana, C.D. Chena, A.H. Azaharia and I. Sadiyaha
aMedical Entomology Unit, Infectious Diseases Research Centre, Institute for Medical Research, Jalan Pahang 50588, Kuala Lumpur, Malaysia
bEpidemiology Unit, Medical Resource Research Centre, Institute for Medical Research, Jalan Pahang 50588, Kuala Lumpur, Malaysia
Abstract
This study was conducted to determine the response of dengue vectors to ovitraps treated with temephos and Bacillus thuringensis israelensis (Bti) compared with untreated ovitraps which only contained seasoned tap water. The study was conducted at two sites: the natural breeding habitats where the ovitraps were exposed directly to sunshine and rain, and under shelter beneath a cabin without direct exposure. The ovitraps were placed for a period of five days at six different sites each in both conditions. The experiments were replicated three times. The results indicated that all mosquitoes breeding in the ovitraps were Aedes albopictus. There was no significant difference in the ovipositioning behaviour in the natural habitat, but there was a significant difference in the number of eggs laid in the ovitraps placed under the cabins. Though the mean number of eggs laid in traps in response to each treatment varied among the sites, the overall ovipositioning activity was not significantly different between sites when egg densities in the test ovitraps were averaged over the six-week ovitrapping period (site versus treatment effect) at p<0.05. The study also indicated that there was no ovicidal effect of both control agents since hatching of eggs occurred. Although eggs hatched, all larvae died eventually. The male to female ratio was 1:1 for both the sites. The study also showed that larval index is as effective as pupal index and can be used as a surveillance tool for the dengue vectors. It is concluded that containers treated with larvicide are not repellent to Aedes.
Keywords: Aedes albopictus; oviposition; repellent; temephos; Bacillus thuringiensis israelensis.
#E-mail: [email protected]; Tel.: +60-3-26162687, Fax: +60-3-26162688
IntroductionDengue as well as chikungunya fever pose serious public health problems in Malaysia. Ae. aegypti is the predominant vector of dengue,
a mosquito-borne arborvirus belonging to the family flaviviridae which is capable of causing dengue fever, dengue haemorrhagic fever (DHF) and dengue shock syndrome (DSS).[1] During 2008, a total of 36 991 cases with

210 Dengue Bulletin – Volume 33, 2009
Oviposition behaviour of Aedes albopictus in temephos and Bacillus thuringiensis israelensis-treated ovitraps
81 deaths due to dengue were reported in Malaysia till October.[2] In addition, there were 1703 cases of chikungunya reported from 12 August to 25 September that year.[3] With the rapid movement of people the number of cases may only increase with time.
Ae. albopictus is also a known vector of dengue and chikungunya fever in Malaysia. The chikungunya virus, as a potential menace and responsible for epidemics transmitted by Ae. aegypti and Ae. albopictus, has been documented since 1960.[4] Current measures for the control of Ae. aegypti and Ae. albopictus are based on: (i) the physical elimination of larval development sites; and (ii) the application of 1% sand granules formulation of temephos to artificial containers. Recently, as an alternative to the use of temephos, the biological larvicide such as Bacillus thuringiensis israelensis (Bti) is also used as a bio-larvicide.
The appropriateness of larvicides for use in vector control programmes depends on multiple factors such as persistency of larvicide in the breeding sites and the behavioural responses of vectors against the larvicide. It is in this context that the present study aimed to assess the possible repellent effect of temephos and Bti on the oviposition of gravid females that may influence vector control effectiveness for dengue and chikungunya control programmes in Malaysia.[5]
Materials and methods
Study site
This study was conducted in the vicinity of the Institute for Medical Research, Kuala Lumpur (latitude: 3°10.190’ N and longitude: 101°41.950’ E). The area consisted of several cabins (10 ft X 30 ft X 8 ft) with many trees,
grass and bushes between them. The cabins are placed in the midst of a natural habitat. The temperature and humidity of the site throughout the study period was 25.9 °C±0.68 and 77.7%±4.51 RH for the natural habitat and 25.8 °C±0.57 and 77.50%±4.95 RH for the cabins.
Placement of ovitrap
The ovitrap used in this study was based on the specifications as described by Lee.[6] Ovitraps were placed in: (i) open natural breeding habitat of Aedes sp with minimal exposure to sunshine and rain; and (ii) underneath the cabins without direct exposure to sunlight and rain. The studies were conducted concurrently.
Concentration of temephos and Bacillus thuringiensis
Six spots (three in each habitat) were identified for the placement of ovitraps for the tests. In each site, three ovitraps were placed with one containing water treated with temephos (ABATE) sand granule formulation at the operational dosage of 1 mg/l , and another water treated with Bti (a Vectobac WG, wettable granule formulation with 3000 ITU/mg) at the recommended concentration of 0.008 g/l. the third ovitrap contained only seasoned tap water serving as control. All these ovitraps were placed adjacent to each other and remained in position for five days. In total, for each experiment, 18 ovitraps were placed in six spots. The experiment was replicated three times.
All ovitraps were collected after five days and brought to the laboratory. A five-day duration was used because longer trapping periods increase the risk of egg loss through

Dengue Bulletin – Volume 33, 2009 211
Oviposition behaviour of Aedes albopictus in temephos and Bacillus thuringiensis israelensis-treated ovitraps
predation and hatching which could lead to the disruption of production of adults.[7] The number of positive ovitraps were recorded and the ovitrap index was determined. In the laboratory, the paddle sticks from the ovitraps were dried and the number of eggs laid was counted under a dissecting microscope. The total number of eggs per ovitrap was also calculated. The container was screened for larvae and the number was recorded. The paddle stick was transferred into the same plastic container (14.5 cm x 8.5 cm x 7.5 cm) and the water from the respective ovitraps was poured into the plastic container. A pinch of beef-liver powder was added into the containers. Observations were made for a period of 14 days post-collection. The species of the mosquito was identified at the third instar stage for the control ovitraps. The total number of eggs, the hatchability rate, the pupation and the adult emergence were counted and recorded.
Data and statistical analysis
Field data was analysed by the analysis of variance (ANOVA) method. The preference of Aedes mosquito to oviposit on temephos, Bti or seasoned tap water was evaluated based on the positive ovitrap index (OI) (% positive ovitraps). The oviposition active index (OAI) was used to determine the Aedes attractant to the treated water compared with the control.[8,9] Kramer and Mulla[8] suggested that compounds with an OAI of +0.3 and above are considered as attractants, while those with –0.3 and below are considered as repellents.
Results and discussion
The oviposition responses of the Aedes females to the treated ovitraps placed in the natural habitat showed that there was no significant
difference in the mean number of eggs laid in traps placed at different sites (site main effect; F=0.34; P=0.89), demonstrating that the oviposition activity was comparable among all sites that we used. Although the mean number of eggs laid in traps in response to each treatment varied among the sites, the overall oviposition activity was not significantly different between sites when egg densities in the test ovitraps were averaged over the six-week ovitrapping period (site versus treatment effect; F=0.67, P=0.75). Egg densities in ovitraps with water only were similarly varied, but found to be not significantly different (P>0.05) within or between sites. On the other hand, ovitraps placed under the cabin showed a significant difference in the mean number of eggs laid in traps at different sites (site main effect; F=4.44; P=0.003). Similarly, the overall oviposition activity was not significantly altered between sites when egg densities in the test ovitraps were averaged over the six-week ovitrapping period (site versus treatment effect; F=1.45, P=0.20).
The ovitrap index and the oviposition active index are presented in Table 1. The positive ovitrap index was in the range of 67%–89%. Mosquitoes laid eggs on both treated and untreated paddle sticks. The highest number of eggs laid was in the ovitrap treated with Bti with 425 eggs while the highest number of eggs laid in the control and temephos-treated ovitraps were 400 and 306 eggs respectively. More eggs were laid in the ovitrap placed in the natural breeding habitat compared to the ovitrap placed under the cabin as shown in Table 1. This could be due to the ovitraps placed in the natural breeding sites being exposed to sunshine and rain which could have diluted the treatment effect of the larvicides, but was still equally effective in eliminating larvae that had hatched in the temephos- and Bti-treated ovitraps over the five-day period. Another factor could be that

212 Dengue Bulletin – Volume 33, 2009
Oviposition behaviour of Aedes albopictus in temephos and Bacillus thuringiensis israelensis-treated ovitraps
in the natural breeding site for Ae. abopictus, the high oviposition rate might be due to the high mosquito population in the sites as well as easy accessibility to the ovitrap by the oviposition females.
All mosquitoes from the collected eggs were of the Ae. albopictus species. The mean number of eggs per ovitrap for temephos, Bti and control placed in the natural breeding habitat were 17.00±4.99, 23.61±6.85 and 22.22±5.4 respectively as shown in Table 2. On the other hand, the mean number of eggs per ovitrap placed under the cabin for temephos, Bti and control were 14.72±5.03, 11.00±2.93 and 17.56±4.37 respectively. No significant differences were found in the number of eggs laid in the ovitraps with the treatments and control placed in the natural breeding habitat and those placed under the cabin (P=1.000). In some ovitraps there was no oviposition activity and this accounted for 26 out of 108 (24.07%) ovitrap containers.
A study by Craig[10] has stated that conspecific eggs provide an attractive cue for gravid Ae. aegypti because they are a signal of a suitable breeding site. The authors considered an Allee effect[11,12] in which the presence of conspecifics is attractive and presumably beneficial to egg-laying up to a particular density.
Mosquitoes integrate a wide range of stimuli prior to the act of ovipositioning. Photoperiod, colour and optical density of the water, oviposition substrate texture and moisture, temperature and reflectance, volatile and contact chemical cues have all been shown to affect the choice of oviposition site by mosquitoes.[13] An additional factor affecting the endpoint of oviposition behaviour is known as “skip oviposition”,[14] which occurs when females lay their eggs in several containers as opposed to laying their entire clutch in one container.[15,16,17] This behaviour increases the distribution of eggs in an area
Table 1: Ovitrap index and oviposition response of Ae. albopictus to field conditions with ovitraps treated with temephos and Bacillus thuringiensis israelensis
Treatment Total no. of ovitraps
Positive ovitraps (OI in %)
Total no. of eggs laid
Mean no. of eggs per
ovitrap ± SD
OAI of baited ovitraps
aOT placed in natural breeding sites
Temephos 18 12 (66.67) 306 17.00±4.99 –0.13
Bti 18 14 (77.78) 425 23.61±6.85 +0.04
Control 18 16 (88.89) 400 22.22±5.40
aOT placed under a cabin
Temephos 18 12 (66.67) 265 14.72±5.03 –0.09
Bti 18 14 (77.78) 198 11.00±2.93 –0.23
Control 18 14 (77.78) 316 17.56±4.37aOT = ovitrap

Dengue Bulletin – Volume 33, 2009 213
Oviposition behaviour of Aedes albopictus in temephos and Bacillus thuringiensis israelensis-treated ovitraps
and may be increased by the tendency of gravid females to avoid ovipositing in sites where eggs of conspecific females had been laid.[17,18,19] The oviposition attractive index for the Ae. albopictus at the natural breeding habitat and under the cabin with treatment of temephos and Bti were –0.13, +0.04, –0.09 and –0.23 respectively as shown in Table 1. This showed that there was no attraction for the gravid female mosquitoes to oviposit in either treatment, indicating that neither temephos nor Bti appears to influence the choice of ovitrap for oviposition site.
However, the number of eggs laid showed that Bti has some influence on egg-laying. According to Gubler,[20] ovipositing female Ae. albopictus are attracted to dark-coloured water. Hence, the visual stimulus might have been a factor in oviposition site selection, rather than the presence of Bti or any other chemical.[21] Furthermore, very
few chemicals have been found to attract ovipositing Ae. albopictus. Studies by Sharma[22] showed that compounds such as hexadecyl pentanoate, tetradecyl heptanoate and tridecyl octanoate presented significant oviposition repellent activity against the two mosquito species. However, the compound propyl octadecanoate was found to attract Ae. aegypti to the treated oviposition substrate. Previous studies[23,24,25] have indicated that temephos was not repellent for oviposition behaviour of Ae. aegypti and this was in agreement with our study. The emergence ratio of male to female in the natural breeding habitat and under the cabin was 1:1 and no significant difference was observed in the emergence for both sites for sex (p=0.923, t=–0.103 and p=0.974, t=0.034, respectively).
The life-table of the development of the Ae. albopictus in both the sites is shown in Table 2. There is no significant difference
Table 2: Mean number of eggs, larvae, pupae and adults obtained from the ovitraps placed in the natural breeding site and under a cabin
Treatment
Mean number ± SD
Eggs Larvae PupaeAdults
Male Female Total
aOT placed in natural breeding sites
Temephos 17.0±8.28 10.06±6.77 0 0 0 0
Bti 23.61±9.03 12.39±6.59 0 0 0 0
Control 22.22±9.06 22.17±9.01 21.34±8.86 9.50±9.40 9.78±10.79 19.28±8.16
aOT placed under a cabin
Temephos 14.72±9.02 9.28±6.33 0 0 0 0
Bti 11.00±5.14 7.33±4.09 0 0 0 0
Control 17.56±7.20 16.84±7.21 16.45±7.17 7.78±8.24 7.67±9.07 15.50±6.94aOT = ovitrap

214 Dengue Bulletin – Volume 33, 2009
Oviposition behaviour of Aedes albopictus in temephos and Bacillus thuringiensis israelensis-treated ovitraps
(p>0.05) for egg production for the temephos-treated ovitraps and control ovitraps placed either in the natural breeding habitat or under cabin. More eggs were laid in ovitraps treated with Bti in the natural habitat compared to ovitraps under a cabin. In a study conducted by Santos,[26] it was suggested that Bti did influence the number of eggs and that this effect may depend on the infusion concentration. The resultant metabolites from the bacterial growth within the ovitrap may also be a contributing factor for increased oviposition and ovitrap attractiveness.[27] It could also be due to the exposure to sunshine which activates the Bti spores to yield the aroma that attracts ovipositioning. It is interesting to note that all larvae that hatched from the temephos- and Bti-treated ovitraps died due to the toxic effect of the larvicide. This indicated that both larvicides did not exhibit ovicidal properties
because eggs did hatch but the larvae were completely eliminated upon hatching.
An extended analysis in the non-treated group (control), with ovitraps placed in natural habitat and under cabin, showed that there was no significant difference in the developmental stages from larvae to adults (P=0.69; P=0.83, respectively), as well as from pupae to adults (P=0.77; P=0.88, respectively). Hence, the number of ovitraps positive for larvae (larval index) is still as effective as the number of ovitraps positive for pupae (pupal index). Since it is quite difficult in surveys to find containers with pupae, containers with larvae can also be a proxy for the number of adults that emerge, as shown in Figures 1 and 2 for ovitraps placed in the two conditions. The result from this study is a model for what can transpire in an environment.
Figure 1: Survival of Ae. albopictus in each developmental stage in natural breeding sites

Dengue Bulletin – Volume 33, 2009 215
Oviposition behaviour of Aedes albopictus in temephos and Bacillus thuringiensis israelensis-treated ovitraps
Table 3: Percentage mortality and survival of Ae. albopictus in the ovitraps placed in natural habitat and under a cabin
TreatmentNo. of survival cases in each developmental stage (percentage)
Eggs Larvae Pupae Adults
Natural breeding sites
Temephos 306 181 (59.18) 0 0
Bti 425 223 (52.48) 0 0
Control 400 399 (99.77) 96.25 90.34
Under a cabin
Temephos 265 167 (63.00) 0 0
Bti 198 132 (66.64) 0 0
Control 316 303 (95.90) 97.68 94.22
Figure 2: Survival of Ae. albopictus in each developmental stage in ovitraps placed under a cabin

216 Dengue Bulletin – Volume 33, 2009
Oviposition behaviour of Aedes albopictus in temephos and Bacillus thuringiensis israelensis-treated ovitraps
ReferencesGubler DJ. Dengue and dengue hemorrhagic [1] fever: Its history and resurgence as a global public health problem. In: Gubler DJ, Kuno G, eds. Dengue and dengue haemorrhagic fever. Wallingford: CAB International, 1997. pp. 1-22.
Yao. [2] Dengue Fever claims 81 lives in Malaysia. China View. 2008 Oct 25. http://news.xinhuanet.com/english/2008-10/25/content_10249418.htm – accessed 30 June 2010.
Centre of Disease Control. [3] Dengue fever 2007. http://www.svinfectologia.org/Dengue%20CDC%202007%5B1%5D.doc – accessed 30 June 2010.
Marchette NJ, Rudnick A, Garcia R. Alphaviruses [4] in Peninsular Malaysia: II. Serological evidence of human infection. Southeast Asian Journal of Tropical Medicine and Public Health 1980;11(1):14-23.
Canyon D. [5] Irritancy and repellency of Aedes aegypti (Diptera: Culicidae) to insecticides and implications for vector control operations. (http://www.tropmed.org/rreh/rrehi.htm – accessed 30 June 2010).
Lee HL.[6] Aedes ovitrap and larval survey in several suburban communities in Selangor, Malaysia. Mosquito Borne Diseases Bulletin 1992;9(1):9-15.
Ritchie SA. The production of [7] Aedes aegypti by a weekly ovitrap survey. Mosquito News 1984;44(1):77-79.
Kramer WL, Mulla MS. Oviposition attractants [8] and repellents of mosquitoes: oviposition responses of Culex mosquitoes to organic infusions. Environmental Entomology 1979;8:1111-1117.
Geetha I, Paily KP, Padmanaban V, Balaraman [9] K. Oviposition response of the mosquito, Culex quinquefasciatus to the secondary metabolite(s) of the fungus, Trichoderma viride. Mem Inst Oswaldo Cruz 2003;98(2):223-226.
Craig RW, Katherine JL, Natasha JW, Veronica [10] RS. The Allee effect in site choice behaviour of egg-laying dengue vector mosquitoes. Tropical Biomedicine 2008;25(2):140-144.
Allee WC. [11] Animal aggregations, a study in general sociology. Chicago: University of Chicago Press, 1931.
This study also showed that the survival rate into adult stages in breeding containers in natural habitat and in semi-covered buildings was 90.34% and 94.22% respectively (Table 3), which is a good survival rate in natural environmental conditions, indicating that dengue or chikungunya transmission can be highly efficient and effective.
Our study also showed that Ae. albopictus was not repelled by the treatment of either temephos or Bti. Hence, we can conclude that if Bti was used in the control programme, dispersing Bti by ultra-low volume (ULV) would not influence the oviposition behaviour of
Ae. albopictus in Malaysia. This, in fact, will also serve as a good control measure because containers which contain the dispersed Bti will be attracted to oviposition and larvae that hatch will eventually be killed by Bti.
Acknowledgements
We would like to thank the Director-General, Ministry of Health, Malaysia, and the Director, Institute for Medical Research, Kuala Lumpur, for allowing us to publish this paper. Thanks are also due to the staff of the Medical Entomology Unit for their assistance.

Dengue Bulletin – Volume 33, 2009 217
Oviposition behaviour of Aedes albopictus in temephos and Bacillus thuringiensis israelensis-treated ovitraps
Stephens PA, Sutherland WJ, Freckleton [12] RP. What i s the Al lee ef fect? Oikos 1999;87:185-190.
Bentley MD, Day JF. Chemical ecology [13] and behavioral aspects of mosquito oviposition. Annual Review of Entomology 1989;34:401-21.
Mogi M, Mokry J. Distribution of [14] Wyeomyia smithii (Diptera: Culicidae) eggs in pitcher plants in Newfoundland, Canada. Tropical Medicine 1980;22:1-12.
Fay RW, Perry AS. Laboratory studies of the [15] ovipositional preferences of Aedes aegypti. Mosquito News 1965;26:531-535.
Rozeboom LE, Rosen L, Ikeda J. Observations on [16] oviposition by Aedes (S.) albopictus Skuse and A. (S.) polynesienses Marks in nature. Journal of Medical Entomology 1973;4:397-399.
Apostol BL, Black WC, Reiter P, Miller BR. [17] Use of randomly amplified polymorphic DNA amplified by polymerase chain reaction markers to estimate the number of Aedes aegypti families at oviposition sites in San Juan, Puerto Rico. American Journal of Tropical Medicine and Hygiene 1994;51:89-97.
Kitron U, Webb DDW, Novak RJ. Oviposition [18] behavior of Aedes triseriatus (Diptera: Culicidae): prevalence, intensity, and aggregation of eggs in oviposition traps. Journal of Medical Entomology 1989;26:462-467.
Chadee DD, Corbet PS, Greenwood JJD. [19] Egg-laying yellow fever mosquitoes avoid sites containing eggs laid by themselves or by conspecifics. Entomologia Experimentalis et Applicata 1990;57:295-298.
Gubler DJ. Studies on the comparative [20] oviposition behavior of Aedes (Stegomyia)
albopictus and Aedes (Stegomyia) polynesiensis Marks. Journal of Medical Entomology 1971;8:675-682.
Trexler JD, Apperson CS, Gemeno C, Perich [21] MJ, Carlson D, Schal C. Field and laboratory evaluations of potential oviposition attractants for Aedes albopictus (Diptera: Culicidae). Journal of the American Mosquito Control Assocication 2003;19:228-234.
Sharma KR, Seenivasagan T, Rao AN, [22] Ganesan K, Agarwal OP, Malhotra RC, Prakash S. Oviposition responses of Aedes aegypti and Aedes albopictus to certain fatty acid esters. Parasitology Research 2008;103(5):1065-1073.
Mather TN, DeFoliart GR Repellency and initial [23] toxicity of Abate and Dursban formulations to Aedes triseriatus in oviposition sites. Mosquito News 1983;43:474-479.
Beehler JW, Mulla MS. Effect of the insect [24] growth regulator methoprene on the ovipositional behavior of Aedes aegypti and Culex quinquefasciatus. Journal of the American Mosquito Control Association 1993;9:13-16.
Pates H, Curtis C. Mosquito behavior and [25] vector control. Annual Review of Entomology 2005;50:53-70.
Santos SRA, Melo-Santos MAV, Regis [26] L, Albuquerque CMR. Field evaluation consociated with grass infusion and Bacillus thuringiensis var. israelensis to determine oviposition rates of Aedes aegypti. Dengue Bulletin 2003;27:156-162.
Benzon GL, Apperson CS. Reexamination of [27] chemically mediated oviposition behavior in Aedes aegypti (L.) (Diptera: Culicidae). Journal of Medical Entomology 1988;25(3):158-164.

218 Dengue Bulletin – Volume 33, 2009
Book review
Dengue guidelines for diagnosis, treatment, prevention and control, 2009 (WHO/HTM/NTD/DEN/2009.1)
Since the second edit ion of Dengue haemorrhagic fever: diagnosis, treatment, prevention and control was published by the World Health Organization (WHO) in 1997, the magnitude of the dengue problem has increased dramatically and has extended geographically to many previously unaffected areas. It was then, and remains today, the most important arthropod-borne viral disease of humans.
Activities undertaken by WHO regarding dengue are most recently guided at the global policy level by World Health Assembly resolution WHA55.17 (adopted by the Fifty-fifth World Health Assembly in 2002) and at the regional level by resolution CE140.R17 of the Pan American Sanitary Conference (2007), resolution WPR/RC59.R6 of the WHO Regional Committee for the Western Pacific (2008) and resolution SEA/RC61/R5 of the WHO Regional Committee for South-East Asia (2008).
This new edition has been produced to make widely available to health practitioners, laboratory personnel, those involved in vector control and other public health officials, a concise source of information of worldwide relevance on dengue. The guidelines provide updated practical information on the clinical management and delivery of clinical services; vector management and delivery of vector
control services; laboratory diagnosis and diagnostic tests; and surveillance, emergency preparedness and response. Looking ahead, some indications of new and promising avenues of research are also described. Additional and more detailed specific guidance on the various specialist areas related to dengue are available from other sources in WHO and elsewhere, some of which are cited in the references.
The contributions of, and review by, many experts both within and outside WHO have facilitated the preparation of this publication through consultative and peer review processes. All contributors are gratefully acknowledged, a list of whom appears under “Acknowledgements”. These guidelines are the result of collaboration between the WHO Department of Control Neglected Tropical Diseases, the WHO Department of Epidemic and Pandemic Alert and Response, and the Special Programme for Research and Training in Tropical Diseases.
This publication is intended to contribute to prevention and control of the morbidity and mortality associated with dengue and to serve as an authoritative reference source for health workers and researchers. These guidelines are not intended to replace national guidelines but to assist in the development of national or regional guidelines. They are expected

Dengue Bulletin – Volume 33, 2009 219
Dengue guidelines for diagnosis, treatment, prevention and control, 2009
to remain valid for five years (until 2014), although developments in research could change their validity, since many aspects of the prevention and control of dengue are currently being investigated in a variety of studies. The guidelines contain the most up-to-date
information at the time of writing. However, the results of studies are being published regularly and should be taken into account. To address this challenge, the guide is also available on the Internet and will be updated regularly by WHO.

220 Dengue Bulletin – Volume 33, 2009
Instructions for contributors
Dengue Bulletin welcomes all original research papers, short notes, review articles, letters to the Editor and book reviews which have a direct or indirect bearing on dengue fever/dengue haemorrhagic fever prevention and control, including case management. Papers should not contain any political statement or reference.
Manuscripts should be typewritten in English in double space on one side of white A4-size paper, with a margin of at least one inch on either side of the text and should not exceed 15 pages. The title should be as short as possible. The name of the author(s) should appear after the title, followed by the name of the institution and complete address. The e-mail address of the corresponding author should also be included and indicated accordingly.
References to published works should be listed on a separate page at the end of the paper. References to periodicals should include the following elements: name and initials of author(s); title of paper or book in its original language; complete name of the journal, publishing house or institution concerned; and volume and issue number, relevant pages and date of publication, and place of publication (city and country). References should appear in the text in the same numerical order (Arabic numbers in parenthesis) as at the end of the article. For example:
Nimmannitaya S. (1) Clinical spectrum and management o f dengue haemorrhagic fever. The Proceedings of the International Conference on Dengue Haemorrhagic Fever, Kuala Lumpur, September 1-3, 1983:16-26.
Gubler DJ. Dengue and dengue (2) haemorrhagic fever: Its history and resurgence as a global public health problem. In: Gubler DJ, Kuno G (ed.), Dengue and dengue haemorrhagic fever. CAB International, New York, NY, 1997, 1-22.
Nguyen Trong Lan, Nguyen Thanh (3) Hung, Do Quang Ha, Bui Thi Mai Phuong, Le Bich Lien, Luong Anh Tuan, Vu Thi Que Huong, Lu Thi Minh Hieu, Tieu Ngoc Tran, Le Thi Cam and Nguyen Anh Tuan. Treatment of dengue haemorrhagic fever at Children’s Hospital N.1, Ho Chi Minh City, 1991-1996. Dengue Bulletin. 1997; 22: 150-161.
Figures and tables (Arabic numerals), with appropriate captions and titles, should be included on separate pages, numbered consecutively, and included at the end of the text with instructions as to where they belong. Abbreviations should be avoided or explained at the first mention. Graphs or figures should be clearly drawn and properly labelled, preferably using MS Excel, and all data clearly identified.
Articles should include a self-explanatory abstract at the beginning of the paper of not more than 300 words explaining the need/gap in knowledge and stating very briefly the area and period of study. The outcome of the research should be complete, concise and focused, conveying the conclusions in totality. Appropriate keywords and a running title should also be provided.
Articles submitted for publication should be accompanied by a statement that they have not already been published, and, if accepted

Dengue Bulletin – Volume 33, 2009 221
Instructions for contributors
for publication in the Bulletin, will not be submitted for publication elsewhere without the agreement of WHO, and that the right of republication in any form is reserved by the WHO Regional Offices for South-East Asia (SEARO) and the Western Pacific (WPRO).
One hard copy of the manuscript with original and clear figures/tables and a computer diskette/CD-ROM indicating the name of the software should be submitted to:
The Editor Dengue Bulletin WHO Regional Office for South-East Asia Indraprastha Estate Mahatma Gandhi Road New Delhi 110002, India Telephone: 91-11-23370804 Fax: 91-11-23379507, 23370972 E-mail: [email protected]
Manuscripts received for publication are subjected to in-house review by professional experts and are peer-reviewed by experts in the respective disciplines. Papers are accepted on the understanding that they are subject to editorial revision, including, where necessary, condensation of the text and omission of tabular and illustrative material.
Original copies of articles submitted for publication will not be returned. The principal author will receive 10 reprints of the article published in the Bulletin. A pdf file can be supplied on request.

Related Documents











