Veronica: Chemical characters for the support of phylogenetic relationships based on nuclear ribosomal and plastid DNA sequence data Dirk C. Albach a,1, * , Søren R. Jensen b , Fevzi O ¨ zgo ¨ kce c , Rene ´ e J. Grayer d a Department of Higher Plant Systematics, Institute of Botany, University of Vienna, Rennweg 14, A-1030 Vienna, Austria b Department of Chemistry, The Technical University of Denmark, DK-2800 Lyngby, Denmark c Department of Biology, Faculty of Science and Arts, Yu ¨ zu ¨ ncu ¨ Yil University, 65080 Van, Turkey d Royal Botanic Gardens, Kew, Richmond, Surrey TW9 3AB, UK Received 15 December 2004; accepted 6 June 2005 Abstract Molecular phylogenetic analyses have revealed many relationships in Veronica (Plantagi- naceae) never anticipated before. However, phytochemical characters show good congruence with DNA-based analyses. We have analysed a combined data set of 49 species and subspecies derived from the nuclear ribosomal ITS-region (18 new sequences) and the plastid trnL-F region (12 new sequences) of Veronica emphasizing subgenera Chamaedrys and Pocilla and separate analyses of subgenera Pentasepalae (ITS only) and Pseudolysimachium. Results for subgenus Chamaedrys show that Eur opean and Asi an perennial species are monophyletic sister groups with the annual species consecutive sisters to them. All species of Veronica that contain cornoside are found in this subgenus, although some species seem to have secondarily lost the abi lity to produce thi s compound. Subgenera Pocilla and Pentasepalae are wel l supported sister groups characterized by the occurrence of 8-hydroxyflavones. The traditional subsection Biloba of subgenus Pocilla is biphyl eti c with Veronic a intercedens being clearly * Corresponding author. Tel.: C49 6131 39 23169; fax: C49 6131 39 23524. E-mail address: [email protected] (D.C. Albach). 1 Present address: Institute of Special Botany, Johannes Gutenberg-Universita ¨ t Mainz, Bentzelweg 9b, 55099 Mainz, Germany. 0305-1978/$ - see front matter Ó 2005 Elsevier Ltd. All rights reserved. doi:10.1016/j.bse.2005.06.002 Biochemical Systematics and Ecology 33 (2005) 1087 e1106 www.elsevier.com/locate/biochemsyseco

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

8/14/2019 BSE Chemtax
http://slidepdf.com/reader/full/bse-chemtax 1/20
Veronica : Chemical characters for the supportof phylogenetic relationships based on nuclear
ribosomal and plastid DNA sequence data
Dirk C. Albacha ,1 ,
*, Søren R. Jense nb, Fevzi O ¨zgo kce
c,
Rene e J. Grayer d
a Department of Higher Plant Systematics, Institute of Botany, University of Vienna,Rennweg 14, A-1030 Vienna, Austria
b Department of Chemistry, The Technical University of Denmark, DK-2800 Lyngby, Denmarkc Department of Biology, Faculty of Science and Arts, Yu ¨ zu ¨ ncu ¨ Yil University, 65080 Van, Turkey
d Royal Botanic Gardens, Kew, Richmond, Surrey TW9 3AB, UK
Received 15 December 2004; accepted 6 June 2005
Abstract
Molecular phylogenetic analyses have revealed many relationships in Veronica (Plantagi-naceae) never anticipated before. However, phytochemical characters show good congruencewith DNA-based analyses. We have analysed a combined data set of 49 species and subspeciesderived from the nuclear ribosomal ITS-region (18 new sequences) and the plastid trnL-F region (12 new sequences) of Veronica emphasizing subgenera Chamaedrys and Pocilla andseparate analyses of subgenera Pentasepalae (ITS only) and Pseudolysimachium . Results for
subgenus Chamaedrys show that European and Asian perennial species are monophyleticsister groups with the annual species consecutive sisters to them. All species of Veronica thatcontain cornoside are found in this subgenus, although some species seem to have secondarilylost the ability to produce this compound. Subgenera Pocilla and Pentasepalae are wellsupported sister groups characterized by the occurrence of 8-hydroxyavones. The traditionalsubsection Biloba of subgenus Pocilla is biphyletic with Veronica intercedens being clearly
* Corresponding author. Tel.: C 49 6131 39 23169; fax: C 49 6131 39 23524.E-mail address: [email protected] (D.C. Albach).
1 Present address: Institute of Special Botany, Johannes Gutenberg-Universita t Mainz, Bentzelweg 9b,55099 Mainz, Germany.
0305-1978/$ - see front matter Ó 2005 Elsevier Ltd. All rights reserved.doi:10.1016/j.bse.2005.06.002
Biochemical Systematics and Ecology 33 (2005) 1087 e 1106
www.elsevier.com/locate/biochemsyseco

8/14/2019 BSE Chemtax
http://slidepdf.com/reader/full/bse-chemtax 2/20
separate from the rest of the group. This result is mirrored by the unusual phytochemicalarsenal of V. intercedens , which is the only species in the genus analysed to date to containmelittoside and globularifolin. Subgenus Pentasepalae appears to be a clade of diverse lineages
from southwestern Asia and a single European clade. Species shown to have 6-hydroxyavones do not form a monophyletic group. Subgenus Pseudolysimachium seems tohave originated in Eastern Asia. 6-Hydroxyavones acylated with phenolic acids are commonin this subgenus but may have originated only later in the evolution of the group. Possiblechemotaxonomic markers for other groups are discussed.Ó 2005 Elsevier Ltd. All rights reserved.
Keywords: Veronica ; Plantaginaceae; Chemosystematics; Phylogenetic analysis; ITS; trnL-F region
1. Introduction
Veronica is a genus of the Plantaginaceae sensu APG (1998) formerly placed inScrophulariaceae with about 500 species ( Albach et al., 2004a ). It is distributed overmost of the Northern Hemisphere and in many parts of the Southern Hemisphere, andis ecologically highly diverse with species growing in aquatic to dry steppe habitatsfrom sea level to high alpine regions. This diversity and the fact that many species havebeautiful blue owers may explain the interest Veronica has drawn for a long time.Extensive breeding programs andmorphological studies between1900 and1955, i.e. byWatzl (1910) and Lehmann and his pupils (e.g., Lehmann, 1908, 1937; Ro ¨mpp, 1928;Ha rle, 1932; Riek, 1935; Schlenker, 1936; Riek-Ha ußermann, 1943 ) have beenfollowed by cytological and morphological analyses by M.A. Fischer (1967, 1972,1975, 1987) , Martı nez-Ortega ( Martı nez-Ortega et al., 2000; Martı nez-Ortega andRico, 2001 ) and oristic surveys (e.g., Borissova, 1955; Hartl, 1966 e 1968; M.A.Fischer, 1978, 1981 ). Cladistic analyses incorporating much of the morphological datahave been publishedby Hong(1984) and Kampny and Dengler (1997) . Phytochemistryof Veronica and relatedspeciesof the Northern Hemisphere has been studiedby severalauthors with an emphasis on iridoid compounds (e.g., Grayer-Barkmeijer, 1973;Lahloub et al., 1993; Taskova et al., 2002, 2004 ) and avonoid compounds (e.g.,
Grayer-Barkmeijer, 1978, 1979; Peev, 1982; Albach et al., 2003 ).Recently, molecular techniques and phylogenetic analyses have been applied toVeronica and related genera ( Wagstaff and Garnock-Jones, 1998; Albach and Chase,2001; Albach et al., 2004b,c ). These studies have helped revolutionizing our ideasabout the evolution of the genus and led to a new, phylogenetic infragenericclassication of Veronica (Albach et al., 2004a ). Combined with the vast amount of information from other aspects of their biology, we now have a much betterunderstanding of how major groups in Veronica are delimited and related, and howthey have evolved. This information combined in a phylogenetic framework opensa new perspective for character evolution in the genus and helps focus on those
clades that have not been studied intensively in the past.One area of the study of plant evolution that has shown strong correlations with the
results of DNA sequence analyses is chemotaxonomy ( Grayer et al., 1999 ). The
1088 D.C. Albach et al. / Biochemical Systematics and Ecology 33 (2005) 1087 e 1106

8/14/2019 BSE Chemtax
http://slidepdf.com/reader/full/bse-chemtax 3/20
objective of the present paper is therefore to investigate the distribution of phytochemical compounds, which may be useful as an independent test of relation-ships suggested in DNA sequence analyses. Taskova et al. (2004) have compared the
distribution of iridoid compounds in the genus Veronica with dendrograms based onDNA sequences. However, interesting chemotaxonomic characters can also be foundamong avonoids and verbascoside-like compounds of Veronica . In two accompany-ing studies, we investigate the phytochemistry of additional species focusing onunusual avonoids (acylated 6- and 8-hydroxylated avone glycosides, Albach et al.,2005 ) and the iridoid glucoside, ajugol, and on the non-iridoid compound, cornoside,in the genus ( Jensen et al., 2005 ). The distribution of these characters is discussed herein the light of a phylogenetic hypothesis derived from DNA sequences from the nuclearribosomal internal spacer region (ITS 1, 5.8 S rDNA, ITS 2) and the plastid trnL-F region ( trn L intron, trn L 3# exon, trnL-F spacer).
2. Materials and methods
2.1. Plant sampling
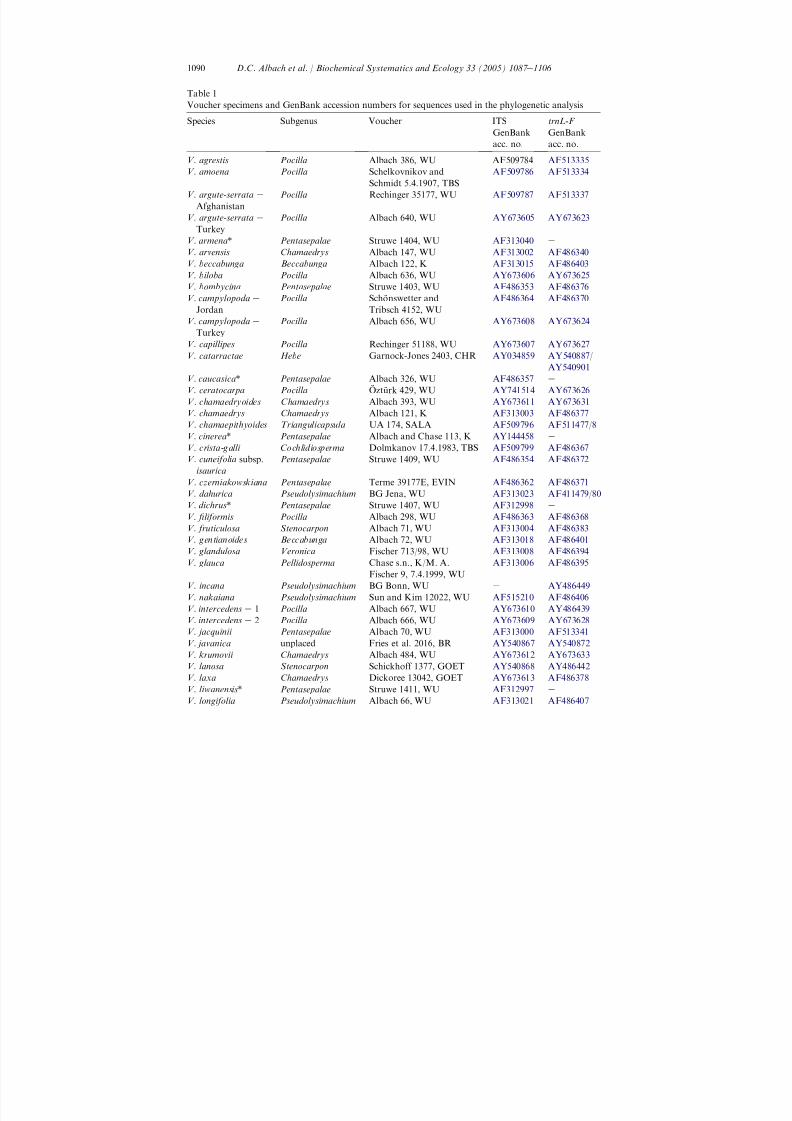
For a larger analysis, 49 accessions were sequenced for the ITS- and trnL-F regionwith an emphasis on subgenera Chamaedrys and Pocilla , which appeared as mostinteresting in the phytochemical analyses ( Albach et al., 2005 ). Taxa from all othersubgenera sensu Albach et al. (2004a) were added to the data matrix ( Table 1 ).Veronica montana and Veronica beccabunga were designated as outgroups based onprevious analyses ( Albach and Chase, 2001; Albach et al., 2004b,c ). Due to limitedvariation in the trnL-F region of subgenus Pentasepalae , a smaller matrix of 15 ITS-sequences including eight new sequences ( Table 1 ) for species of subgenusPentasepalae was analysed separately and rooted with Veronica czerniakowskianabased on previous analyses ( Albach et al., 2004b,c ). Finally, nine ITS- and ve trnL-F -sequences available from subgenus Pseudolysimachium (Table 1 ) were analysed inseparate analyses with Veronica glandulosa , V. beccabunga and Veronica gentianoidesas outgroups based on previous analyses ( Albach et al., 2004b,c ). Eleven of the trnL-F
and 19 of the ITS-sequences are reported here for the rst time. Voucher specimenswere made for all plants used in this study ( Table 1 ).
2.2. DNA extraction, amplication and sequencing
The protocol followed previous studies ( Albach and Chase, 2001; Albach et al.,2004b ). Total genomic DNA was extracted from herbarium material and silica gel-dried leaf samples according to the 2x CTAB procedure of Doyle and Doyle (1987) .The trnL intron, 3 # exon, and trnL-F spacer (hereafter trnL-F ) were amplied withprimers c and f of Taberlet et al. (1991) . ITS-sequences were amplied and sequenced
using the primers 17SE ( Sun et al., 1994 ) and ITS4 ( White et al., 1991 ). PCRproducts were run on a 1.0% TBE-agarose gel, cut out of the gel, and cleaned usingQIAquick Ô PCR purication and gel extraction kit (Qiagen GmbH, Hilden,
1089D.C. Albach et al. / Biochemical Systematics and Ecology 33 (2005) 1087 e 1106
8/14/2019 BSE Chemtax
http://slidepdf.com/reader/full/bse-chemtax 4/20
Table 1Voucher specimens and GenBank accession numbers for sequences used in the phylogenetic analysis
Species Subgenus Voucher ITS
GenBankacc. no.
trnL-F
GenBankacc. no.
V. agrestis Pocilla Albach 386, WU AF509784 AF513335V. amoena Pocilla Schelkovnikov and
Schmidt 5.4.1907, TBSAF509786 AF513334
V. argute-serrata e
AfghanistanPocilla Rechinger 35177, WU AF509787 AF513337
V. argute-serrata e
TurkeyPocilla Albach 640, WU AY673605 AY673623
V. armena * Pentasepalae Struwe 1404, WU AF313040 e
V. arvensis Chamaedrys Albach 147, WU AF313002 AF486340V. beccabunga Beccabunga Albach 122, K AF313015 AF486403V. biloba Pocilla Albach 636, WU AY673606 AY673625V. bombycina Pentasepalae Struwe 1403, WU AF486353 AF486376V. campylopoda e
JordanPocilla Scho nswetter and
Tribsch 4152, WUAF486364 AF486370
V. campylopoda e
TurkeyPocilla Albach 656, WU AY673608 AY673624
V. capillipes Pocilla Rechinger 51188, WU AY673607 AY673627V. catarractae Hebe Garnock-Jones 2403, CHR AY034859 AY540887 /
AY540901V. caucasica * Pentasepalae Albach 326, WU AF486357 e
V. ceratocarpa Pocilla O ¨ztu rk 429, WU AY741514 AY673626
V. chamaedryoides Chamaedrys Albach 393, WU AY673611 AY673631V. chamaedrys Chamaedrys Albach 121, K AF313003 AF486377V. chamaepithyoides Triangulicapsula UA 174, SALA AF509796 AF511477 /8V. cinerea * Pentasepalae Albach and Chase 113, K AY144458 e
V. crista-galli Cochlidiosperma Dolmkanov 17.4.1983, TBS AF509799 AF486367V. cuneifolia subsp.
isauricaPentasepalae Struwe 1409, WU AF486354 AF486372
V. czerniakowskiana Pentasepalae Terme 39177E, EVIN AF486362 AF486371V. dahurica Pseudolysimachium BG Jena, WU AF313023 AF411479 /80V. dichrus * Pentasepalae Struwe 1407, WU AF312998 e
V. liformis Pocilla Albach 298, WU AF486363 AF486368V. fruticulosa Stenocarpon Albach 71, WU AF313004 AF486383
V. gentianoides Beccabunga Albach 72, WU AF313018 AF486401V. glandulosa Veronica Fischer 713/98, WU AF313008 AF486394V. glauca Pellidosperma Chase s.n., K/M. A.
Fischer 9, 7.4.1999, WUAF313006 AF486395
V. incana Pseudolysimachium BG Bonn, WU e AY486449V. nakaiana Pseudolysimachium Sun and Kim 12022, WU AF515210 AF486406V. intercedens e 1 Pocilla Albach 667, WU AY673610 AY486439V. intercedens e 2 Pocilla Albach 666, WU AY673609 AY673628V. jacquinii Pentasepalae Albach 70, WU AF313000 AF513341V. javanica unplaced Fries et al. 2016, BR AY540867 AY540872V. krumovii Chamaedrys Albach 484, WU AY673612 AY673633V. lanosa Stenocarpon Schickhoff 1377, GOET AY540868 AY486442V. laxa Chamaedrys Dickoree 13042, GOET AY673613 AF486378V. liwanensis * Pentasepalae Struwe 1411, WU AF312997 e
V. longifolia Pseudolysimachium Albach 66, WU AF313021 AF486407
1090 D.C. Albach et al. / Biochemical Systematics and Ecology 33 (2005) 1087 e 1106
8/14/2019 BSE Chemtax
http://slidepdf.com/reader/full/bse-chemtax 5/20
Germany) following the manufacturer’s protocols. Sequencing reactions (10 ml) werecarried out using the Taq DyeDeoxy Terminator Cycle Sequencing mix (AppliedBiosystems Inc.). Reactions were run out on a Prism 377 automated sequencer(Applied Biosystems Inc.), and both strands were sequenced. Sequences wereassembled and edited using Sequence Navigator Ô (Applied Biosystems Inc.).Assembled sequences were manually aligned prior to analysis. Aligned sequencematrices are available from DCA by request.
2.3. Sequence analysis
Insertions and deletions are frequent in both DNA regions. We have scored those
gaps that are neither found in mononucleotide repeats nor are only 1 bp long aspresent/absent to preserve the information included in them. All matrices wereanalysed with PAUP* 4.0b10 ( Swofford, 1998 ) using heuristic parsimony methods.
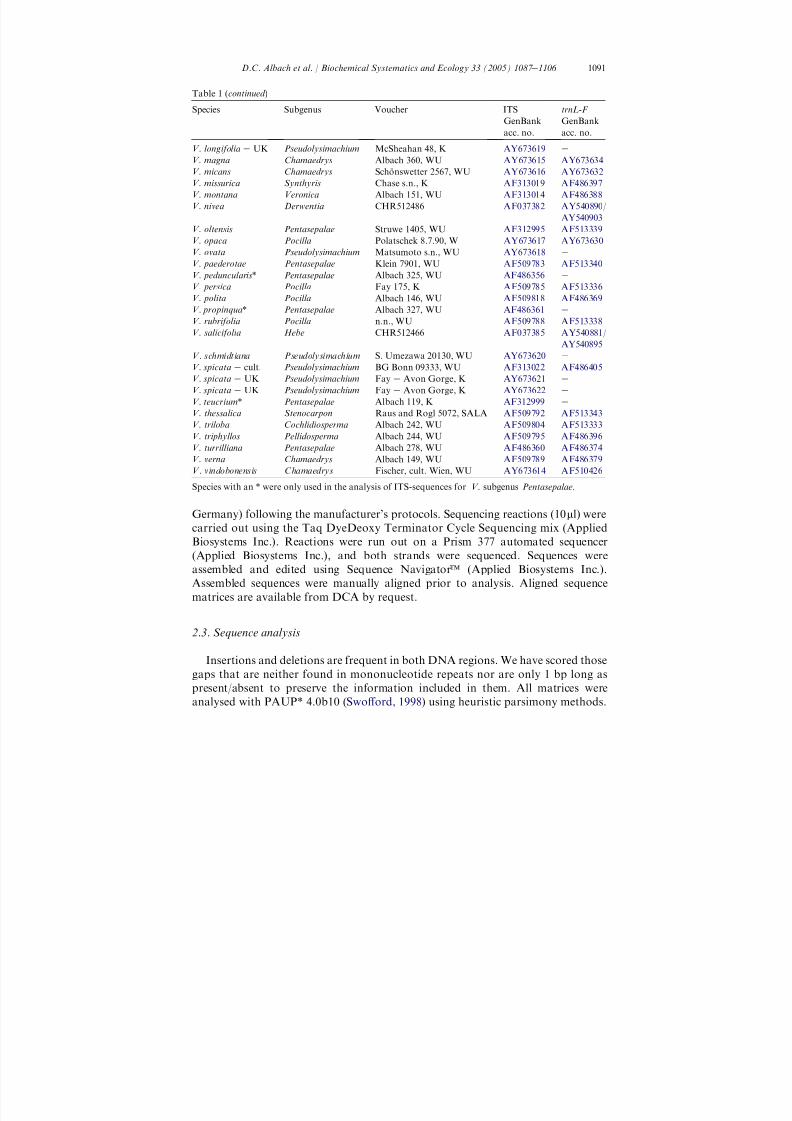
Table 1 ( continued )
Species Subgenus Voucher ITSGenBank
acc. no.
trnL-F GenBank
acc. no.V. longifolia e UK Pseudolysimachium McSheahan 48, K AY673619 e
V. magna Chamaedrys Albach 360, WU AY673615 AY673634V. micans Chamaedrys Scho nswetter 2567, WU AY673616 AY673632V. missurica Synthyris Chase s.n., K AF313019 AF486397V. montana Veronica Albach 151, WU AF313014 AF486388V. nivea Derwentia CHR512486 AF037382 AY540890 /
AY540903V. oltensis Pentasepalae Struwe 1405, WU AF312995 AF513339V. opaca Pocilla Polatschek 8.7.90, W AY673617 AY673630V. ovata Pseudolysimachium Matsumoto s.n., WU AY673618 e
V. paederotae Pentasepalae Klein 7901, WU AF509783 AF513340V. peduncularis * Pentasepalae Albach 325, WU AF486356 e
V. persica Pocilla Fay 175, K AF509785 AF513336V. polita Pocilla Albach 146, WU AF509818 AF486369V. propinqua * Pentasepalae Albach 327, WU AF486361 e
V. rubrifolia Pocilla n.n., WU AF509788 AF513338V. salicifolia Hebe CHR512466 AF037385 AY540881 /
AY540895V. schmidtiana Pseudolysimachium S. Umezawa 20130, WU AY673620 e
V. spicata e cult. Pseudolysimachium BG Bonn 09333, WU AF313022 AF486405V. spicata e UK Pseudolysimachium Fay e Avon Gorge, K AY673621 e
V. spicata e UK Pseudolysimachium Fay e Avon Gorge, K AY673622 e
V. teucrium * Pentasepalae Albach 119, K AF312999 eV. thessalica Stenocarpon Raus and Rogl 5072, SALA AF509792 AF513343V. triloba Cochlidiosperma Albach 242, WU AF509804 AF513333V. triphyllos Pellidosperma Albach 244, WU AF509795 AF486396V. turrilliana Pentasepalae Albach 278, WU AF486360 AF486374V. verna Chamaedrys Albach 149, WU AF509789 AF486379V. vindobonensis Chamaedrys Fischer, cult. Wien, WU AY673614 AF510426
Species with an * were only used in the analysis of ITS-sequences for V. subgenus Pentasepalae .
1091D.C. Albach et al. / Biochemical Systematics and Ecology 33 (2005) 1087 e 1106

8/14/2019 BSE Chemtax
http://slidepdf.com/reader/full/bse-chemtax 6/20
Ten runs of random taxon addition (10 replicates each) using tree bisectionreconnection (TBR) were conducted with MulTrees (keeping multiple shortest trees)in effect and no tree limit. Bootstrap percentages were assessed using 1000 replicates
and TBR-branch swapping with a maximum of 100 trees per replicate.
3. Results and discussion
3.1. ITS
Aligned sequences of the ITS-region included 759 characters including 4 gapsscored as present/absent (one having a CI ! 1). One hundred and ninety-sevencharacters were potentially parsimony informative. We found 121 most parsimoni-ous trees, which required 943 steps ( Fig. 1 ; CI Z 0.46, RI Z 0.71). Relationshipsamong the subgenera are not well supported except for the clade consisting of subgenera Chamaedrys , Pocilla and Pentasepalae (72 BP). However, monophyly of subgenera is well supported. Within subgenus Chamaedrys (99 BP), the annualspecies Veronica arvensis and Veronica verna are consecutive sisters to the perennialspecies. Among the latter, the Asian species, Veronica laxa and Veronica magna , aresister to the European species (75 BP). Veronica chamaedrys is most closely related toVeronica micans (90 BP). Within subgenus Pocilla (100 BP), two clades are stronglysupported that resemble subsection Agrestes in the circumscription of Lehmann(1908; 96 BP) and subsection Biloba sensu Ro ¨mpp (1928; 100 BP) . Within subsectionAgrestes , Veronica polita , Veronica persica and Veronica opaca form one moderatelysupported clade (87 BP), as does the group of Veronica ceratocarpa , Veronica liformis , and Veronica agrestis . A position of Veronica amoena close to subsect.Agrestes has hitherto not been suggested. Within subsection Biloba , Veronica argute-serrata is not monophyletic, one sequence being more closely related than the otherto Veronica campylopoda . Veronica biloba and Veronica capillipes are stronglysupported sisters (99 BP). Veronica intercedens , usually considered part of subsectionBiloba , does not seem to belong to the group in a strict sense.
3.2. trnL-F
Aligned sequences of the trnL-F region included 1067 characters including 16 gapsscored as present/absent with 136 characters potentially parsimony informative. Westopped saving trees when 1000 most parsimonious trees that required 475 steps werefound ( Fig. 2 ; CI Z 0.79, RI Z 0.83). Repeating the search ve times did not result insearches nding shorter trees. Relationships among subgenera do nd somebootstrap support above 50%. Within subgenus Chamaedrys (83 BP), the branchingpattern differs from that in the ITS-analysis by V. verna switching its place withV. magna and V. laxa , which renders the perennial species of the subgenus non-
monophyletic. Veronica vindobonensis is sister to V. micans (59 BP) rather than foundin a clade together with Veronica krumovii and Veronica chamaedryoides as in theITS-analysis. Within subgenus Pocilla (90 BP), subsection Biloba is paraphyletic with
1092 D.C. Albach et al. / Biochemical Systematics and Ecology 33 (2005) 1087 e 1106

8/14/2019 BSE Chemtax
http://slidepdf.com/reader/full/bse-chemtax 7/20

8/14/2019 BSE Chemtax
http://slidepdf.com/reader/full/bse-chemtax 8/20
Fig. 2. One of more than 1000 most parsimonious trees from the analysis of the trnL-F region for 49 taxa of Veronica . Numbers above branch and before the slash are branch lengths, numbers behind the slash arebootstrap percentages. Branches not present in strict consensus tree are dotted with branch lengths in italics.
1094 D.C. Albach et al. / Biochemical Systematics and Ecology 33 (2005) 1087 e 1106

8/14/2019 BSE Chemtax
http://slidepdf.com/reader/full/bse-chemtax 9/20
respect to subsection Agrestes and Veronica rubrifolia . Within subsection Agrestes ,V. amoena is sister to the rest of the subsection. Subgenus Pentasepalae (97 BP) ischaracterized by a lack of resolution due to low sequence divergence except for the
sister-group relationship of V. czerniakowskiana to the rest of the subgenus (75 BP).
3.3. Combined data set
The complete data set of both regions combined included 1826 charactersincluding 20 gaps scored as present/absent with 333 characters potentially parsimonyinformative. We found 384 most parsimonious trees that required 1441 steps ( Fig. 3 ;CI Z 0.56, RI Z 0.73). The analysis differs from that of the single data sets in anoverall increase in bootstrap support in all relationships. Branching pattern amongsubgenera Pocilla , Pentasepalae , and Chamaedrys resembles that in the analysis of ITS. In subgenus Chamaedrys (100 BP), the annual species are paraphyletic withrespect to the perennials (71 BP). Branching pattern in subgenus Pocilla (100 BP) issimilar to that of the trnL-F analysis with subsection Biloba paraphyletic tosubsection Agrestes and V. rubrifolia and V. amoena sister to the rest of subsectionAgrestes . Due to the lack of variation in the trnL-F data set, relationships withinsubgenus Pentasepalae (95 BP) are the same as in the analysis of ITS.
3.4. ITS-data set for subgenus Pentasepalae
The 15 sequences for subgenus Pentasepalae included 36 potentially parsimony-informative characters but no potentially parsimony-informative indel. We foundthree most parsimonious trees that required 133 steps ( Fig. 4 ; CI Z 0.78, RI Z 0.66).Bootstrap support is generally low with the exception of the European species fromsubsection Pentasepalae and the association of Veronica peduncularis and Veronicacaucasica .
3.5. Analyses of subgenus Pseudolysimachium
The analysis of nine ITS-sequences from six species from subgenus Pseudoly-
simachium included 76 potentially parsimony-informative characters and resultedin two most parsimonious trees ( Fig. 5 A) with 175 steps (CI Z 0.89, RI Z 0.86).The analysis of ve trnL-F -sequences from ve species included 24 potentiallyparsimony-informative characters and resulted in a single most parsimonious tree(Fig. 5 B) with 71 steps (CI Z 0.99, RI Z 0.97). The most parsimonious trees fromthe analysis of ITS are reasonably well resolved, but the one from the analysis of trnL-F is not, because of limited sequence variability. Both analyses agree inVeronica nakaiana from Eastern Asia being sister to the rest of the subgenus. Inthe analysis of ITS-sequences, V. nakaiana is consecutively followed by two otherEast Asian species, Veronica dahurica and Veronica schmidtiana . However,
bootstrap support is relatively low except for the monophyly of the subgenus, of those two species where more than one sequence was available, and of the cladecomprising the European Veronica spicata and Veronica longifolia together with
1095D.C. Albach et al. / Biochemical Systematics and Ecology 33 (2005) 1087 e 1106

8/14/2019 BSE Chemtax
http://slidepdf.com/reader/full/bse-chemtax 10/20
Fig. 3. One of 384 most parsimonious trees from the combined analysis of the ITS-region and the trnL-F region for 49 taxa of Veronica . Numbers above branch and before the slash are branch lengths, numbersbehind the slash are bootstrap percentages. Branches not present in strict consensus tree are dotted withbranch lengths in italics. Branches with bootstrap support above 70% appear in double thickness, thoseabove 95% in triple thickness.
1096 D.C. Albach et al. / Biochemical Systematics and Ecology 33 (2005) 1087 e 1106

8/14/2019 BSE Chemtax
http://slidepdf.com/reader/full/bse-chemtax 11/20
Fig. 4. One of three most parsimonious trees. The other two show Veronica cuneifolia either as sister toVeronica cinerea or as sister to V. subsect. Pentasepalae . Numbers above the branches indicate branch
lengths, those below the branch bootstrap support.
1097D.C. Albach et al. / Biochemical Systematics and Ecology 33 (2005) 1087 e 1106

8/14/2019 BSE Chemtax
http://slidepdf.com/reader/full/bse-chemtax 12/20
Fig. 5. Phylograms for sequence analyses of subgenus Pseudolysimachium . Branch lengths (before the dash) and bootstrap dash) are indicated. (A) Strict consensus and one of two most parsimonious trees of the analysis of the ITS-region (the other most aV. spicata and V. longifolia with V. ovata as sister to both). (B) Most parsimonious tree of the analysis of the trnL-F

8/14/2019 BSE Chemtax
http://slidepdf.com/reader/full/bse-chemtax 13/20
Veronica ovata . Sequence variability within V. spicata is surprisingly high. Eventwo individuals within one English population differ by three nucleotidesubstitutions, which is also supported by strong differentiation in AFLP-ngerprint
patterns (Albach, unpubl.).
3.6. Discussion of the DNA sequence analyses
The present analysis of DNA sequence data focuses on two subgenera (subg.Pocilla and Chamaedrys ), which appeared especially interesting regarding theirphytochemical arsenal. Relationships between subgenera have been the focus of previous publications ( Albach and Chase, 2004; Albach et al., 2004b,c ). SubgenusPocilla is sister to subgenus Pentasepalae in analyses of both nuclear ITS and plastidtrnL-F sequences with weak support in the separate analyses (ITS: 57 BP; trnL-F : 68BP) but moderate support in the combined analysis (84 BP). This sister-grouprelationship has also been found in all previous publications ( Albach and Chase,2001, 2004; Albach et al., 2004b,c ). The present analysis extends taxon sampling insubgenus Pocilla , allowing for the rst time a discussion of relationships in thissubgenus. As mentioned before, subgenus Pocilla includes species belonging to theAgrestis -group sensu Lehmann (1908) and ‘‘Verwandtschaftsgruppe’’ Biloba sensuRo ¨mpp (1928) . The analysis supports subsection Agrestes in the circumscription of Lehmann (1908) including V. agrestis , V. liformis , V. ceratocarpa , V. polita ,V. persica , and V. opaca . Apart from the two strictly European tetraploid speciesV. agrestis and V. opaca , all other species originate from the hyrcanian-caucasianregion with three of them ( V. ceratocarpa , Veronica francispetae e both diploid;Veronica siaretensis e unknown ploidy) restricted to this region. V. polita ,V. liformis and V. persica (the rst two diploid, the latter tetraploid) have becomecosmopolitan weeds at different times. The occurrence of V. liformis in this group ispeculiar because it differs from all other species of the subgenus in being perennial andself-incompatible ( Lehmann, 1944 ), whereas all other species are annuals and at leastthose of subsection Agrestes self-compatible ( Lehmann and Schmitz-Lohner, 1954 ).Self-incompatibility in subsection Biloba is unlikely. Morphological differencesbetween the species of subsection Agrestes have been discussed by Thaler (1951) and
Fischer (1987) . Several publications have dealt with the origin of the polyploid speciesin this subsection. Based on karyological and biogeographical evidence, Beatus (1936)hypothesized that all three tetraploid species ( V. agrestis , V. opaca , V. persica ) areautopolyploids, V. persica derived from V. liformis and the other two derived fromV. polita . Lehmann (1940) contradicted Beatus with respect to V. persica , which heconsidered to be also derived from V. polita . Fischer (1987) however, argued based onmorphological intermediacy that V. persica is an allopolyploid species derived froma cross of V. polita and V. ceratocarpa . Evidence presented here from bothmaternally- and biparently-inherited DNA sequences supports a contribution of V. polita to the genomes of V. persica and V. opaca , although it cannot be conrmed
whether they are autopolyploid or allopolyploids, because ITS-sequence could beconverging towards the maternal parent. The position of V. agrestis as sister toV. liformis is rather surprising as a close relationship has not been proposed before
1099D.C. Albach et al. / Biochemical Systematics and Ecology 33 (2005) 1087 e 1106

8/14/2019 BSE Chemtax
http://slidepdf.com/reader/full/bse-chemtax 14/20
based on morphological evidence, although their avonoid proles are very similar(Grayer-Barkmeijer, 1979 ). Further investigations are necessary to elucidate theancestry of the polyploid species in this group.
Subsection Biloba is characterized by pairwise fusion of the calyces, more or lessrecurved pedicels when fruiting and in most cases strongly bilobed capsules. Ouranalyses ( Figs. 1 e 3) do not support this subsection to be monophyletic in its originalcircumscription by Lehmann (1910 ; see also Ro ¨mpp, 1928 ). All analyses indicateV. intercedens to be more closely related to V. rubrifolia than to the rest of thesubsection. V. intercedens was split from subsection Biloba by Elenevsky (1977)together with Veronica cardiocarpa to subsection Cardiocarpae based on theverticillate cauline leaves in these species. This is also supported by the unusualiridoid phytochemistry of V. intercedens , which is the only species in the genus tocontain melittoside and globularifolin ( Albach et al., 2003 ). V. cardiocarpa has notyet been investigated phytochemically. V. rubrifolia does not share any of thecharacters of subsection Biloba or subsection Cardiocarpae mentioned above and itsposition in the phylogeny remains doubtful until further annual species fromsouthwest Asia have been included in the analyses.
Analysis of the subgenus Pentasepalae supports the sister-group relationship of V. czerniakowskiana to the rest of the subgenus ( Figs. 1 e 4) found in previousanalyses ( Albach et al., 2004b,c ). However, using cpDNA, subgenus Chamaedrys issister to a larger clade of subgenera Pentasepalae and Pocilla , subgenus Stenocarponand the Hebe complex in analyses of the plastid trnL-F region ( Fig. 2, Albach et al.,2004b,c ) and the plastid rps16 intron ( Albach and Chase, 2004 ).
The association of the perennial V. chamaedrys with the annual V. arvensis wasrst noted by Albach and Chase (2001) . Both species have never been consideredclosely related because the distinction between annuals with a terminal inorescenceand those perennial species that only have lateral inorescences was considered themajor division in the genus ( Bentham, 1846; Ro ¨mpp, 1928 ). Apart from this,V. arvensis and V. verna have seeds that differ strongly from V. chamaedrys andrelatives (Martı nez-Ortega and Albach, in preparation). Phytochemical similarity of these species (absence of catalpol esters in both species, Taskova et al., 2004 ; Jensenet al., 2005 ) is therefore valuable support for the results of DNA sequence analyses.
The results for subgenus Chamaedrys presented here ( Figs. 1 e 3) show that theperennial species are further divided into an Asian group ( V. magna , V. laxa ) anda European group. The European group can be termed V. chamaedrys sensu latobecause all species have been included in V. chamaedrys at one time or another.Apart from the tetraploid V. chamaedrys all other species sampled are diploid. Nodenite answer can be given regarding the ancestors of V. chamaedrys but it isnoteworthy that in all analyses V. chamaedrys seems to be closer to the CentralEuropean species V. vindobonensis and V. micans than to the strictly Balkanianspecies V. krumovii and V. chamaedryoides . An origin of V. chamaedrys in CentralEurope seems therefore more likely. However further diploid populations of
V. chamaedrys in Central and Eastern Europe are known ( Fischer, 1973 ; M.A.Fischer, pers. comm.). Those populations so closely resemble the tetraploidV. chamaedrys that they have not been separated from it at any taxonomic rank.
1100 D.C. Albach et al. / Biochemical Systematics and Ecology 33 (2005) 1087 e 1106

8/14/2019 BSE Chemtax
http://slidepdf.com/reader/full/bse-chemtax 15/20
Due to this fact they should be considered in future analyses investigating theancestors of the tetraploid form of V. chamaedrys .
Subgenus Pentasepalae is a species-rich group with its 70 species mostly found in
Turkey and Iran. Its geographical outliers, subsect. Pentasepalae in Europe andsubsect. Petraeae in the Greater Caucasus, are the only well supported groups (97 BPand 83 BP, respectively; Fig. 4 ) in this subgenus. A larger analysis with moreinformative characters is needed to resolve the relationships of this group.
Subgenus Pseudolysimachium is a highly diverse group with approximately 25species currently accepted despite approximately 400 names and combinationsavailable. This taxonomic complexitydemonstrates the high morphological variabilitywithin the subgenus and explains the difficulties for taxon sampling in molecularanalyses. Another problem in systematic analyses of this subgenus is the probablywidespread interfertility and hybridisation between taxa (e.g. Ha rle, 1932 ). The currentanalyses are therefore only intended as broad guidelines in analyses of AFLP-ngerprints (Albach, unpubl.). Nevertheless, one important result is the paraphyly of East Asian species and the derived position of European species, which indicates anEast Asian origin of the subgenus. This result is important because it points to thosespecies that may resemble most the ancestral species of this subgenus. Among thosespecies is V. schmidtiana , which is the only species that was not associated with thissubgenus before 1968 ( Yamazaki, 1968 ), because it has a attened capsule, which iscommon amongother species of Veronica but only found in a few Eastern Asian speciesof subgenus Pseudolysimachium . V. schmidtiana is also distinct from the rest of thesubgenus in pollen morphology ( Hong, 1984 ) and its rosette habit, which is frequentlyfound amongother species of Veronica (e.g. subgenus Synthyris ) but only rarely amongthe more derived members of subgenus Pseudolysimachium . V. schmidtiana , however,shares with the rest of the subgenus the unusual chromosome number of x Z 17 (e.g.Sakai, 1935 ). Its inorescence is long and terminal but not as dense and spike-like asthose of most other species. The loosely owered inorescence is associated with short-tubed corollas and is found in several Eastern Asian species of this subgenus but not inits derived members. An emphasis of future studies should therefore be put on the EastAsian speciesand not only study its relationship based on DNA-markers but also studyits phytochemistry (see below), seed and pollen.
3.7. Review of chemotaxonomic markers in Veronica
Molecular systematics has often given results that were hitherto unsuspected,which in turn has reinitiated research in other elds of systematics (e.g. Williams andFriedman, 2004 ). Phytochemistry is a prime example for this. Sometimestaxonomically useful characters were overlooked and only later appreciated ( Grayeret al., 1999 ) but much too often phytochemistry is only incompletely known, whichprevents a full assessment of its use in classifying plants. Veronica is a fortunateexception to this. A comparison of our analyses of ITS- and plastid trnL-F sequence
data with published reports on iridoids, avonoids and verbascoside-like compoundsreveals several compounds that are good indicators of relationships, several of thempreviously not recognized. Due to the lack of information on iridoids in the Hebe
1101D.C. Albach et al. / Biochemical Systematics and Ecology 33 (2005) 1087 e 1106

8/14/2019 BSE Chemtax
http://slidepdf.com/reader/full/bse-chemtax 16/20
complex and the complexity of information on avonoids in the Hebe complex(Bayly et al., 2000, 2002 ), we focus here on species from the Northern Hemisphere.Comparisons reveal not only synapomorphies for clades, so far exclusively
characterized by nucleotide differences but also reveal much about the evolutionof these phytochemical compounds. Convergence in phytochemical arsenal seems tobe widespread. The convergence in the production of acetylated 8-hydroxyavoneallosyl glycosides between Veronica (Grayer-Barkmeijer, 1979; Toma s-Barbera net al., 1988; Albach et al., 2003 ) and Stachys (Lamiaceae, Lenherr et al., 1984;Toma s-Barbera n et al., 1988 ) is just one example. Reversals to ancestral conditionsseem to be frequent despite long divergence from ancestors that also contained thesecompounds. This supports the idea of Grayer et al. (1999) that those genesresponsible for the biosynthesis of such compounds may be retained in the genomedespite lack of expression. Alternatively, these reversals may occur if additional stepsin the synthesis of secondary compounds are secondarily lost again. Analyses of genes underlying the biosynthetic pathways such as those in the avonoidbiosynthetic pathway ( Shirley, 1996; Quattrocchio et al., 1999; Zufall and Rausher,2004 ) will be necessary to determine the fate of the underlying genes.
Among those compounds that are chemotaxonomically useful in Veronica areiridoids, avonoids and verbascoside-like compounds. Mussaenoside is one of thefew iridoid compounds in Veronica not derived from aucubin and catalpol. It wasrst detected by Grayer-Barkmeijer (1973, 1979) and then identied by A-Yazarand Sticher (1981) . Subsequent studies showed that it is almost exclusively found ina clade comprising Veronica officinalis , Veronica aphylla and Veronica urticifolia(axillary inorescences) and Veronica alpina and Veronica bellidioides (terminalinorescences) ( Grayer-Barkmeijer, 1979; Taskova et al., 2002, 2004 ), which usedto be classied in two different sections. Grayer-Barkmeijer (1973, 1979) alreadynoted that species producing mussaenoside also share the same basic chromosomenumber ( x Z 9) and have similar seeds. However, the clade was not formallyrecognized until recently (subgenus Veronica , Albach et al., 2004a ), because formerintrageneric classications of Veronica relied on the presence or absence of a terminal inorescence, an unstable character in this group. Two other iridoidsthat are likely to be chemotaxonomically useful are melittoside and globularifolin,
which are found in V. intercedens (Albach et al., 2003 ). Investigation of relatedspecies is necessary to evaluate their distribution, but occurrence in Veronicatenuissima and V. cardiocarpa seems likely because they also do not blacken whiledrying, an indication that aucubin is lacking. Finally, ajugol has been found inmembers of Veronica subsection Biloba (Jensen et al., 2005 ), and as a dominantcompound is a chemotaxonomic marker for this subsection, although it is alsofound in trace amounts in V. arvensis (subgenus Chamaedrys ) (Jensen et al., 2005 ).Other species of Veronica contain aucubin, catalpol and its esters, whereas thespecic combination of esters may be signicant for various groups at varioustaxonomic levels ( Taskova et al., 2002, 2004 ), no ester is conned to a group at
subgeneric level or below.Among avonoids, 8-hydroxyavones are the most prominent chemotaxonomic
marker supporting the sister-group relationship of subgenus Pocilla and subgenus
1102 D.C. Albach et al. / Biochemical Systematics and Ecology 33 (2005) 1087 e 1106

8/14/2019 BSE Chemtax
http://slidepdf.com/reader/full/bse-chemtax 17/20
Pentasepalae (Albach et al., 2005 ). Whereas subgenus Pocilla contains annualspecies, subgenus Pentasepalae is exclusively perennial. Life history (annuals versusperennials) has been considered an essential character for the classication (e.g.
Ro ¨mpp, 1928 ), but annuals have been shown to be derived in parallel multiple timeswithin Veronica (Albach et al., 2004b ). Within these two subgenera, a reversal tothe production of 6-hydroxyavones is also inferred several times ( Albach et al.,2005 ).
Less important chemotaxonomic markers among the avonoids of Veronica are6-hydroxyavones acylated with phenolic acids ( Albach et al., 2005 ). Thesecompounds, which we called spicosides A e F ( Albach et al., in press ), occur in veout of eight investigated species of subgenus Pseudolysimachium . They are lackingin V. dahurica and V. nakaiana , inferred to be the earliest diverging species in thesubgenus ( Fig. 5 ), which contrasts with a possible presence of these compounds inthe ancestor of the subgenus. Two of the spicosides have also been found inV. thymoides ssp. pseudocinerea (Saracoglu et al., 2004 ) belonging to subgenusPentasepalae , where this character is thought to have originated independently(Albach et al., 2005 ). Other avonoid compounds that may be chemotaxonomicmarkers still need closer investigation. Grayer-Barkmeijer (1979) noticed therestricted occurrence of luteolin 7,3 # -O -glucuronide in Veronica fruticans , Veronica fruticulosa and Veronica saturejoides . Previously these species were thought tobelong to two different subsections, but they are now considered more closelyrelated ( Albach et al., 2004b ). Scutellarein-glycosides may be a fourth avonoidchemotaxonomic marker in Veronica being only found in members of sectionBeccabunga with the exception of Veronica peregrina (Grayer-Barkmeijer, 1979 ).The position of V. peregrina in section Beccabunga was unrecognised until recently(Albach and Chase, 2001; Albach et al., 2004c ). Although the absence of scutellarein-glycosides may argue against its inclusion, the overall similarity of theavonoid arsenal ( Grayer-Barkmeijer, 1979 ) is clearly in support of the DNA-based results. This highlights the trend already observed among iridoids that whilespecic chemotaxonomic markers may be lacking, the overall similar combinationof compounds is generally supporting results from DNA-based molecularsystematics.
Finally, cornoside, a compound related to verbascoside, has been found in bothannual and perennial species of subgenus Chamaedrys , but not outside this subgenusin Veronica (Jensen et al., 2005 ). It is therefore an important character supportinga clade that until now has only been distinguished by DNA sequence data ( Figs. 1 e 3;Albach et al., 2004b,c ).
Phytochemical characters may also be important as chemosystematic markers ata lower taxonomic level in Veronica , e.g. the presence or absence of a diosmetinglycoside in the species-complex of Veronica hederifolia (Peev, 1982 ). Furtherexamples have been given by Grayer-Barkmeijer (1973, 1978, 1979) and Taskovaet al. (1997, 2002) , but many more may be found after closer inspection. Much
chemotaxonomic work is currently being carried out on the Hebe complex in NewZealand, where several examples of chemosystematic markers useful at the specieslevel have been found (e.g. Bayly et al., 2002 ).
1103D.C. Albach et al. / Biochemical Systematics and Ecology 33 (2005) 1087 e 1106

8/14/2019 BSE Chemtax
http://slidepdf.com/reader/full/bse-chemtax 18/20
Acknowledgments
We thank the Studienstiftung des deutschen Volkes for a doctoral scholarship to
D.C.A. and nancial support by FWF (Fonds zur Fo rderung der wissenschaftlichenForschung) project P15336.
References
A-Yazar, F.U ¨., Sticher, O., 1981. Ladroside ( Z 6# -caffeoyl-mussaenoside), a new iridoid glucoside fromVeronica officinalis L. (Scrophulariaceae) and the elucidation of the absolute conguration at C (8) of mussaenoside. Helv. Chim. Acta 64, 16 e 24.
Albach, D.C., Chase, M.W., 2001. Paraphyly of Veronica (Veroniceae; Scrophulariaceae): evidence fromthe internal transcribed spacer (ITS) sequences of nuclear ribosomal DNA. J. Plant Res. 114, 9 e 18.
Albach, D.C., Chase, M.W., 2004. Incongruence in Veroniceae (Plantaginaceae): evidence from twoplastid and a nuclear region. Mol. Phylogenet. Evol. 32, 183 e 197.
Albach, D.C., Grayer, R.J., Jensen, S.R., O ¨zgo kce, F., Veitch, N.C., 2003. Acylated avone glycosidesfrom Veronica . Phytochemistry 64, 1295 e 1301.
Albach, D.C., Martı nez-Ortega, M.M., Fischer, M.A., Chase, M.W., 2004a. A new classication of thetribe Veroniceae e problems and a possible solution. Taxon 53, 429 e 452.
Albach, D.C., Martı nez-Ortega, M.M., Chase, M.W., 2004b. Veronica : parallel morphological evolutionand phylogeography in the Mediterranean. Plant Syst. Evol. 246, 177 e 194.
Albach, D.C., Martı nez-Ortega, M.M., Fischer, M.A., Chase, M.W., 2004c. Evolution of Veroniceae:a phylogenetic perspective. Ann. Missouri Bot. Gard. 91, 275 e 302.
Albach, D.C., Grayer, R.J., Kite, G.C., Jensen, S.R., 2005. Veronica : acylated avone glycosides as
chemosystematic markers. Biochem. Syst. Ecol. 33 (11), 1167 e 1177.Angiosperm Phylogeny Group, 1998. An ordinal classication for the families of owering plants. Ann.Missouri Bot. Gard. 85, 531 e 553.
Bayly, M.J., Garnock-Jones, P.J., Mitchell, K.A., Markham, K.R., Brownsey, P.J., 2000. A taxonomicrevision of the Hebe parviora complex (Scrophulariaceae), based on morphology and avonoidchemistry. N.Z. J. Bot. 38, 165 e 190.
Bayly, M.J., Kellow, A.V., Mitchell, K.A., Markham, K.R., deLange, P.J., Harper, G.E., Garnock-Jones, P.J., Brownsey, P.J., 2002. Descriptions and avonoid chemistry of new taxa in Hebe sect.Subdistichae (Scrophulariaceae). N.Z. J. Bot. 40, 571 e 602.
Beatus, R., 1936. Die Veronica -Gruppe Agrestis der Sektion Alsinebe Griseb., ein Beitrag zum Problem derArtbildung. Z. indukt. Abstammungs- Vererbungsl. 71, 353 e 381.
Bentham, G., 1846. Scrophulariaceae. In: de Candolle, A. (Ed.), Prodromus Systematis Naturalis Regni
Vegetabilis. Victor Masson, Paris, pp. 448 e 491.Borissova, A.G., 1955. Veronica. In: Shishkin, B.K., Bobrov, E.G. (Eds.), Flora SSSR. Izdatel’stvo
Akademii Nauk SSSR, Moskva, Leningrad, pp. 373 e 557.Doyle, J.J., Doyle, J.L., 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue.
Phytochem. Bull. Bot. Soc. Am. 19, 11 e 15.Elenevsky, A.G., 1977. Sistema roda Veronca L. Bjull. Moskovsk. Obsc. Isp. Prir., Otd. Biol. 82, 149 e 160.Fischer, M., 1967. Beitra ge zur Cytotaxonomie der Veronica hederifolia -Gruppe (Scrophulariaceae).
O ¨sterr. Bot. Z. 114, 189 e 233.Fischer, M., 1972. Neue Taxa, Chromosomenzahlen und Systematik von Veronica subsect. Acinifolia
(Ro ¨mpp). Stroh. O ¨sterr. Bot. Z. 121, 413 e 437.Fischer, M., 1975. Untersuchungen u ber den Polyploidkomplex Veronica cymbalaria agg. (Scrophular-
iaceae). Plant Syst. Evol. 123, 97 e 105.Fischer, M.A., 1973. Notizen zur Systematik, Chromosomenzahl und Verbreitung einiger Veronica -Sippen
in Ka rnten. Carinthia 83, 379 e 388.
1104 D.C. Albach et al. / Biochemical Systematics and Ecology 33 (2005) 1087 e 1106

8/14/2019 BSE Chemtax
http://slidepdf.com/reader/full/bse-chemtax 19/20
Fischer, M.A., 1978. Veronica L. In: Davis, P.H. (Ed.), Flora of Turkey and the East Aegean IslandsUniversity Press, Edinburgh, pp. 689 e 753.
Fischer, M.A., 1981. Veronica . In: Rechinger, K.H. (Ed.), Flora des iranischen Hochlandes und derumrahmenden Gebirge. Akademische Druck- und Verlagsanstalt, Graz, pp. 52 e 165.
Fischer, M.A., 1987. On the origin of Veronica persica (Scrophulariaceae) e a contribution to the historyof a neophytic weed. Plant Syst. Evol. 155, 105 e 132.
Grayer, R., Chase, M.W., Simmonds, M.S.J., 1999. A comparison between chemical and molecularcharacters for the determination of phylogenetic relationships among plant families: an appreciation of Hegnauer’s ‘‘Chemotaxonomie der Panzen’’. Biochem. Syst. Ecol. 27, 369 e 393.
Grayer-Barkmeijer, R.J., 1979. Chemosystematic investigations in Veronica L. (Scrophulariaceae) andrelated genera. PhD thesis, University of Leiden.
Grayer-Barkmeijer, R.J., 1973. A chemosystematic study of Veronica : iridoid glucosides. Biochem. Syst.Ecol. 1, 101 e 110.
Grayer-Barkmeijer, R.J., 1978. Flavonoids in Parahebe and Veronica : a chemosystematic study. Biochem.Syst. Ecol. 6, 131 e 137.
Ha rle, A., 1932. Die Arten und Formen der Veronica -Sektion Pseudolysimachia Koch auf Grundsystematischer und experimenteller Untersuchungen. Bibl. Bot. 26, 1 e 86.Hartl, D., 1966 e 1968. Scrophulariaceae. In: Hartl, D.G.W. (Ed.), Gustav Hegi, Illustrierte Flora von
Mitteleuropa. Verlag Paul Parey, Berlin Hamburg, pp. 16 e 469.Hong, D.-Y., 1984. Taxonomy and evolution of the Veroniceae (Scrophulariaceae) with special reference
to palynology. Opera Bot. 75, 5 e 60.Jensen, S.R., Albach, D.C.., Ohno, T., Grayer, R.J., 2005. Veronica : iridoids and cornoside as
chemotaxonomic markers. Biochem. Syst. Ecol. 33, 1031 e 1047.Kampny, C.M., Dengler, N.G., 1997. Evolution of ower shape in Veroniceae. Plant Syst. Evol. 205, 1 e 25.Lahloub, M.-F., Zaghloul, M.G., A, M.S., Sticher, O., 1993. Iridoid glucosides from Veronica anagallis-
aquatica . Phytochemistry 33, 401 e 405.Lehmann, E., 1908. Geschichte und Geographie der Veronica -Gruppe Agrestis . Bull. Herb. Boissier 8,
229 e 660.Lehmann, E., 1910. U ¨ber Merkmalseinheiten in der Veronica -Sektion Alsinebe . Z. Bot. 2, 577 e 602.Lehmann, E., 1937. Die Gattung Veronica in entwicklungsgeschichtlicher Betrachtung. Cytologia Fuji
Jubilaei Volumen, 903 e 919.Lehmann, E., 1940. Polyploidie und geographische Verbreitung der Arten der Gattung Veronica . Jahrb.
Wiss. Bot. 89, 461 e 542.Lehmann, E., 1944. Veronica liformis Sm., eine selbststerile Panze. Jahrb. Wiss. Bot. 91, 395.Lehmann, E., Schmitz-Lohner, M., 1954. Entwicklung und Polyploidie in der Veronica -Gruppe Agrestis .
Z. indukt. Abstammungs- Vererbungsl. 86, 1 e 34.Lenherr, A., Lahloub, M.F., Sticher, O., 1984. Three avonoid glycosides containing acetylated allose
from Stachys recta . Phytochemistry 23, 2343 e 2345.Martı nez-Ortega, M.M., Rico, E., 2001. Seed morphology and its systematic signicance in some Veronica
species (Scrophulariaceae) mainly from the western Mediterranean. Plant Syst. Evol. 228, 15 e 32.Martı nez-Ortega, M.M., Sanchez Sanchez, J., Rico, E., 2000. Palynological study of Veronica Sect.
Veronica and Sect. Veronicastrum (Scrophulariaceae) and its taxonomic signicance. Grana 39, 21 e 31.Peev, D.R., 1982. Different substitution tendencies of leaf avones in the Veronica hederifolia group
(Scrophulariaceae). Plant Syst. Evol. 140, 235 e 242.Quattrocchio, F., Wing, J., van der Woude, K., Souer, E., de Vetten, N., Mol, J., Koes, R., 1999.
Molecular analysis of the anthocyanin2 gene of Petunia and its role in the evolution of ower color.Plant Cell 11, 1433 e 1444.
Riek, R., 1935. Systematische und panzengeographische Untersuchungen in der Veronica -SektionChamaedrys Griseb. Repert. Spec. Nov. Regni Veg. Beih. 79, 1 e 68.
Riek-Ha ußermann, C., 1943. Vergleichend-anatomische und entwicklungsgeschichtliche Untersuchungenu ber Samen in der Gattung Veronica . Beih. Bot. Centralbl. 62, 1 e 60.
Ro ¨mpp, H., 1928. Die Verwandtschaftsverha ltnisse in der Gattung Veronica . Repert. Spec. Nov. RegniVeg. Beih. 50, 1 e 171.
1105D.C. Albach et al. / Biochemical Systematics and Ecology 33 (2005) 1087 e 1106

8/14/2019 BSE Chemtax
http://slidepdf.com/reader/full/bse-chemtax 20/20
Sakai, K.-I., 1935. Studies on the chromomosome number in alpine-plants. II. Jap. J. Genet. 11, 68 e 73.Saracoglu, I., Varel, M., Harput, U.S., Nagatsu, A., 2004. Acylated avonoids and phenol glycosides from
Veronica thymoides subsp. pseudocinerea . Phytochemistry 65, 2379 e 2385.Schlenker, G., 1936. Systematische Untersuchungen u ber die Sektion Beccabunga der Gattung Veronica .
Repert. Spec. Nov. Regni Veg. Beih. 90, 1 e 40.Shirley, B.W., 1996. Flavonoid biosynthesis: ‘new’ functions for an ‘old’ pathway. Trends Plant Sci. 1,
377e 382.Sun, Y., Skinner, D.Z., Liang, G.H., Hulbert, S.H., 1994. Phylogenetic analysis of Sorghum and related
taxa using internal transcribed spacers of nuclear ribosomal DNA. Theor. Appl. Genet. 89, 26 e 32.Swofford, D.L., 1998. PAUP ) Phylogenetic Analysis using Parsimony ( ) and Other Methods). Sinauer
Associates, Sunderland, Massachusetts.Taberlet, P., Gielly, L., Pautou, G., Bouvet, J., 1991. Universal primers for amplication of three non-
coding regions of chloroplast DNA. Plant Mol. Biol. 17, 1105 e 1109.Taskova, R., Peev, D., Handjieva, N., Baranovska, I., 1997. A taxonomic study of some Veronica species.
Phytol. Balcanica 3, 79 e 84.
Taskova, R., Peev, D., Handjieva, N., 2002. Iridoid glucosides of genus Veronica s.l. and their systematicsignicance. Plant Syst. Evol. 231, 1 e 17.Taskova, R.M., Albach, D.C., Grayer, R.J., 2004. Phylogeny of Veronica e a combination of molecular
and chemical evidence. Plant Biol. 6, 673 e 682.Thaler, I., 1951. Morphologisches u ber Veronica liformis Smith und ihre Verwandten. Phyton 3,
216e 226.Toma s-Barbera n, F.A., Grayer-Barkmeijer, R.J., Gil, M.I., Harborne, J.B., 1988. Distribution of
6-hydroxy-, 6-methoxy- and 8-hydroxyavone glycosides in the Labiatae, the Scrophulariaceae andrelated families. Phytochemistry 27, 2631 e 2645.
Wagstaff, S.J., Garnock-Jones, P.J., 1998. Evolution and biogeography of the Hebe complex(Scrophulariaceae) inferred from ITS sequences. N.Z. J. Bot. 36, 425 e 437.
Watzl, B., 1910. Veronica prostrata L., V. teucrium L. und V. austriaca L. nebst einem Anhang u ber deren
na chste Verwandte. Abh. zool.-botan. Ges. Wien. 5, 1 e 94.White, T.J., Bruns, T., Lee, S., Taylor, J., 1991. Amplication and direct sequencing of fungal ribosomal
RNA genes for phylogenetics. In: Innis, M., Gelfand, D., Sninsky, J., White, T. (Eds.), PCR Protocols:A Guide to Methods and Applications. Academic Press, San Diego, pp. 315 e 322.
Williams, J.H., Friedman, W.E., 2004. The four-celled female gametophyte of Illicium (Illiciaceae:Austrobaileyales): implication for understanding the origin and early evolution of monocots,eumagnoliids, and eudicots. Am. J. Bot. 91, 332 e 351.
Yamazaki, T., 1968. On the genus Pseudolysimachion . J. Jap. Bot. 43, 405 e 412.Zufall, R.A., Rausher, M.D., 2004. Genetic changes associated with oral adaptation restrict future
evolutionary potential. Nature 428, 847 e 850.
1106 D.C. Albach et al. / Biochemical Systematics and Ecology 33 (2005) 1087 e 1106
Related Documents











