Mutation Research 729 (2012) 35–40 Contents lists available at SciVerse ScienceDirect Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis jo ur n al hom ep a ge: www.elsevier.com/locate/molmut C om mun i ty a ddress: www.elsevier.com/locate/mutres Nuclear envelope defects impede a proper response to micronuclear DNA lesions Mariona Terradas, Marta Martín, Laia Hernández, Laura Tusell, Anna Genescà ∗ Department of Cell Biology, Physiology and Immunology, Universitat Autònoma de Barcelona, 08193 Bellaterra, Spain a r t i c l e i n f o Article history: Received 12 April 2011 Received in revised form 7 September 2011 Accepted 9 September 2011 Available online 16 September 2011 Keywords: Micronuclei DNA Damage response UV photolesions NER pathway Nuclear envelope a b s t r a c t When damage is inflicted in nuclear DNA, cells activate a hierarchical plethora of proteins that consti- tute the DNA damage response machinery. In contrast to the cell nucleus, the ability of micronuclear DNA lesions to activate this complex network is controversial. In order to determine whether the DNA contained in micronuclei is protected by the cellular damage response system, we studied the recruit- ment of excision repair factors to photolesions inflicted in the DNA of radiation-induced micronuclei. To perform this analysis, primary human dermal fibroblasts were exposed to UV-C light to induce pho- tolesions in nuclear and micronuclear DNA. By means of immunofluorescence techniques, we observed that most micronuclei were devoid of NER factors. We conclude that UV photoproducts in micronuclei are mostly unable to generate an effective DNA damage response. We observed that the micronuclear envelope structure is a determinant factor that influences the repair of the DNA lesions inside micronu- clei. Therefore, our results allow us to conclude that photolesions in radiation-induced micronuclei are poorly processed because the repair factors are unable to reach the micronuclear chromatin when a micronucleus is formed or after a genotoxic insult. © 2011 Elsevier B.V. All rights reserved. 1. Introduction DNA double strand breaks (DSBs) are considered one of the most hazardous lesions because they can result in DNA fragment loss or in chromosome rearrangements. Among the latter, ring and dicentric chromosomes are of special interest as they can ini- tiate breakage-fusion-bridge (BFB) cycles that promote the main genomic alterations observed in cancer cells [1,2]. When the two centromeres of a dicentric chromatid or two interlaced ring chro- matids are pulled to opposite poles, a chromatin bridge is formed at anaphase [3]. Bridged chromatids can break or detach from spindle poles [4] resulting in the formation of micronuclei containing acen- tric fragments or whole chromosomes at the end of mitosis [5]. In accordance with the key role played by BFB-dependent genomic instability in tumorigenesis, anaphase bridges and micronuclei have been observed in a great variety of cancer cells at higher fre- quencies than the baseline in normal cells [2]. Moreover, increased frequencies of micronuclei have been observed in peripheral blood lymphocytes of healthy subjects with a higher risk of cancer [6]. For the above reasons, micronuclei are considered as indicators of chromosome instability and exposure to mutagenic agents. When DNA damage is inflicted, cells activate a hierarchical cas- cade of proteins composed of sensors, transducers and effectors, widely known as the DDR (DNA damage response). In contrast ∗ Corresponding author. Tel.: +34 935811498; fax: +34 935812295. E-mail addresses: [email protected], [email protected] (A. Genescà). to the DNA damage present in the cell nucleus, which efficiently activates the DDR response, the ability of lesions sequestered in the micronuclear DNA to activate this complex signalling network is controversial. The reports published on the presence of DDR proteins in micronuclei include studies of factors involved in the DSB repair pathway. In 1999, Haaf et al. detected the homolo- gous recombination repair protein RAD51 and the single-stranded DNA-binding Replication Protein A (RPA) in radiation-induced micronuclei [7]. Both proteins belong to the homologous recom- bination pathway and are able to bind areas of single strand DNA that result from DSB resection during DNA replication and repair. Later on, Medvedeva and colleagues detected the phosphorylated form of the histone H2AX (H2AX) in most of the newly formed micronuclei after irradiation [8]. These authors also observed par- tial micronuclear co-localisation of H2AX with ATM, a central player that triggers the essential events of the cell response to DSBs. However, no co-localisation was detected for the presence of 53BP1, RAD50 and MRE11, which are involved in the pathways acti- vated by ATM. Similarly, in cancer cells Yoshikawa et al. observed that only 10% of H2AX positive micronuclei were ATM positive and also a very low co-localisation of phosphorylated 53BP1 or DNA-PKcs with H2AX in micronuclei [9]. These data, together with our co-localization studies on DDR proteins in radiation-induced micronuclei in human foreskin fibroblasts [10], lead us to speculate about the possibility that trafficking defects through the micronuclear envelope [5] could potentially compromise the recruitment of DNA damage-response factors inside micronuclei. This possibility is reinforced by the 0027-5107/$ – see front matter © 2011 Elsevier B.V. All rights reserved. doi:10.1016/j.mrfmmm.2011.09.003

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

N
MD
a
ARRAA
KMDUNN
1
mlatgcmaptaihqflFc
cw
0d
Mutation Research 729 (2012) 35– 40
Contents lists available at SciVerse ScienceDirect
Mutation Research/Fundamental and MolecularMechanisms of Mutagenesis
jo ur n al hom ep a ge: www.elsev ier .com/ locate /molmutC om mun i ty a ddress : www.elsev ier .com/ locate /mutres
uclear envelope defects impede a proper response to micronuclear DNA lesions
ariona Terradas, Marta Martín, Laia Hernández, Laura Tusell, Anna Genescà ∗
epartment of Cell Biology, Physiology and Immunology, Universitat Autònoma de Barcelona, 08193 Bellaterra, Spain
r t i c l e i n f o
rticle history:eceived 12 April 2011eceived in revised form 7 September 2011ccepted 9 September 2011vailable online 16 September 2011
eywords:icronuclei
a b s t r a c t
When damage is inflicted in nuclear DNA, cells activate a hierarchical plethora of proteins that consti-tute the DNA damage response machinery. In contrast to the cell nucleus, the ability of micronuclearDNA lesions to activate this complex network is controversial. In order to determine whether the DNAcontained in micronuclei is protected by the cellular damage response system, we studied the recruit-ment of excision repair factors to photolesions inflicted in the DNA of radiation-induced micronuclei.To perform this analysis, primary human dermal fibroblasts were exposed to UV-C light to induce pho-tolesions in nuclear and micronuclear DNA. By means of immunofluorescence techniques, we observed
NA Damage responseV photolesionsER pathwayuclear envelope
that most micronuclei were devoid of NER factors. We conclude that UV photoproducts in micronucleiare mostly unable to generate an effective DNA damage response. We observed that the micronuclearenvelope structure is a determinant factor that influences the repair of the DNA lesions inside micronu-clei. Therefore, our results allow us to conclude that photolesions in radiation-induced micronuclei arepoorly processed because the repair factors are unable to reach the micronuclear chromatin when amicronucleus is formed or after a genotoxic insult.
. Introduction
DNA double strand breaks (DSBs) are considered one of theost hazardous lesions because they can result in DNA fragment
oss or in chromosome rearrangements. Among the latter, ringnd dicentric chromosomes are of special interest as they can ini-iate breakage-fusion-bridge (BFB) cycles that promote the mainenomic alterations observed in cancer cells [1,2]. When the twoentromeres of a dicentric chromatid or two interlaced ring chro-atids are pulled to opposite poles, a chromatin bridge is formed at
naphase [3]. Bridged chromatids can break or detach from spindleoles [4] resulting in the formation of micronuclei containing acen-ric fragments or whole chromosomes at the end of mitosis [5]. Inccordance with the key role played by BFB-dependent genomicnstability in tumorigenesis, anaphase bridges and micronucleiave been observed in a great variety of cancer cells at higher fre-uencies than the baseline in normal cells [2]. Moreover, increasedrequencies of micronuclei have been observed in peripheral bloodymphocytes of healthy subjects with a higher risk of cancer [6].or the above reasons, micronuclei are considered as indicators ofhromosome instability and exposure to mutagenic agents.
When DNA damage is inflicted, cells activate a hierarchical cas-ade of proteins composed of sensors, transducers and effectors,idely known as the DDR (DNA damage response). In contrast
∗ Corresponding author. Tel.: +34 935811498; fax: +34 935812295.E-mail addresses: [email protected], [email protected] (A. Genescà).
027-5107/$ – see front matter © 2011 Elsevier B.V. All rights reserved.oi:10.1016/j.mrfmmm.2011.09.003
© 2011 Elsevier B.V. All rights reserved.
to the DNA damage present in the cell nucleus, which efficientlyactivates the DDR response, the ability of lesions sequestered inthe micronuclear DNA to activate this complex signalling networkis controversial. The reports published on the presence of DDRproteins in micronuclei include studies of factors involved in theDSB repair pathway. In 1999, Haaf et al. detected the homolo-gous recombination repair protein RAD51 and the single-strandedDNA-binding Replication Protein A (RPA) in radiation-inducedmicronuclei [7]. Both proteins belong to the homologous recom-bination pathway and are able to bind areas of single strand DNAthat result from DSB resection during DNA replication and repair.Later on, Medvedeva and colleagues detected the phosphorylatedform of the histone H2AX (�H2AX) in most of the newly formedmicronuclei after irradiation [8]. These authors also observed par-tial micronuclear co-localisation of �H2AX with ATM, a centralplayer that triggers the essential events of the cell response toDSBs. However, no co-localisation was detected for the presence of53BP1, RAD50 and MRE11, which are involved in the pathways acti-vated by ATM. Similarly, in cancer cells Yoshikawa et al. observedthat only 10% of �H2AX positive micronuclei were ATM positiveand also a very low co-localisation of phosphorylated 53BP1 orDNA-PKcs with �H2AX in micronuclei [9].
These data, together with our co-localization studies on DDRproteins in radiation-induced micronuclei in human foreskin
fibroblasts [10], lead us to speculate about the possibility thattrafficking defects through the micronuclear envelope [5] couldpotentially compromise the recruitment of DNA damage-responsefactors inside micronuclei. This possibility is reinforced by the
3 ion Re
o�lwHpiicFcisamboe
rnfTtttswtafnwmUeiDai
2
2
csc
2
2i6efi
2
n(VtiSwai
6 M. Terradas et al. / Mutat
bservance that only a small fraction of micronuclei with discreteH2AX foci co-localised with 53BP1 and MRE11, while total co-
ocalisation was observed in the cell nucleus and at nuclear blebs,hich are structures connected physically to the cell nucleus [10].owever, alternative explanations exist. According to Costes et al., aossible explanation for the inconsistent detection of DDR proteins
n �H2AX foci in micronuclei could be that the histone foci observedn micronuclei may not reflect the existence of DSBs but rather ahromatin modification status [11], like scars left after DNA repair.urthermore, works published deal with the presence in micronu-lei of factors related to DSB repair. As this kind of lesion is involvedn the micronucleus formation per se, one cannot exclude the pos-ibility that micronuclei contain DDR factors that were alreadyttached to the damaged chromatin before the formation of theicronuclear envelope. According to this view, micronuclei would
e completely unable to activate the DDR, thus, the DDR factorsbserved in some micronuclei would only be the remainders of anarlier response.
In the light of all these possibilities, we decided to analyse theecruitment of DDR factors to micronuclear DNA lesions that areot related to micronucleus formation. To this end, we induced the
ormation of micronuclei by exposing human fibroblasts to �-rays.he micronucleated cells were then exposed to UV light in ordero induce cyclobutane pyrimidine dimmers (CPD) and (6-4) pho-oproducts ((6-4) PPs), among other less frequent lesions [12]. Wehen studied the presence of factors involved in processing photole-ions by the nucleotide excision repair (NER) pathway. Specifically,e determined the presence or absence of xeroderma pigmen-
osum group C (XPC), which is involved in the detection step,nd xeroderma pigmentosum group A (XPA), which is responsibleor the stabilisation of the unwound DNA helix so that damageducleotides can be removed [13,14]. This approach is a fairly gooday of checking whether DDR factors are recruited or not intoicronuclear bodies because factors that participate in repairingV-induced damage are not related to the initial damage thatngenders the micronuclei. Here, we document that DNA lesionsnduced in micronuclei are generally unable to generate an effectiveDR, and demonstrate that the micronuclear envelope structure is
determinant factor that influences the repair of the DNA lesionsnside micronuclei.
. Materials and methods
.1. Cell culture
Human primary fibroblasts (AG01522D, Coriel Cell Culture Repository) wereultivated in monolayers in MEM with Earle Salts and NEAA (GIBCO, BRL) andupplemented with 15% foetal bovine serum and 1% Glutamax-1x. The incubationonditions were set at 37 ◦C and 5% CO2 atmosphere.
.2. UV- and �-irradiation
To induce micronuclei, cells were seeded onto coverslips and irradiated with Gy of �-rays using IBL-437C R-137Cs (dose-rate: 5.5 Gy/min). At 48 h after �-
rradiation, cells were washed with PBS and irradiated with UV-C light at a dose of0 J/m2. Then cells were incubated in supplemented MEM before fixation. In somexperiments, to induce local UV damage, cells were covered with a polycarbonatelter (5 �m pore size; Millipore) while they were exposed to UV-C.
.3. Immunofluorescence
For the immunofluorescence analysis of XPC (sc-30156, Santa Cruz Biotech-ology, Inc.), XPA (GTX72316, GeneTex, Inc.) and cyclobutane pyrimidine dimersMC-062, Kamiya Biomedical Company), we followed the protocol developed byolker et al. [15]. To detect cyclin D1 (ab16663, Abcam), nuclear pore complex pro-
eins (ab24609, Abcam) and LaminB1 (ab16048, Abcam), we fixed the cells for 15 min
n 4% paraformaldehyde and permeabilised them in 0.5% Triton X-100 for 15 min.lides were blocked in 2% foetal calf serum (FCS) – 0.1% Tween-20 for 30 min. Cellsere then incubated for 2 h with primary antibodies and 90 min with secondaryntibodies, which were linked to Alexa488 (Invitrogen) or to Cy3 (GE Healthcare). Allncubations were performed at room temperature. Dried coverslips were mounted
search 729 (2012) 35– 40
on microscope slides using the Vectashield Mounting Medium for fluorescence (Vec-tor Laboratories, Inc.) supplemented with 0.25 �g/ml 4,6-diamino-2-phenylindole(DAPI). Specimens were viewed with an epifluorescent microscope (BX61; Olym-pus) equipped with a Plan-Apochromat 100× NA 1.4 oil objective. Images wereacquired at room temperature using a digital camera (ER3339; Applied Imaging) andthe Cytovision software (version 3.93.2; Applied Imaging). Brightness and contrastadjustments were applied to the whole image using Adobe Photoshop CS version8.0.1 (Adobe Systems).
2.4. Criteria for scoring micronuclei
To score micronuclei we followed the criteria established by Fenech [16]. There-fore, those structures that were morphologically identical to but smaller than thecell nucleus were considered to be micronuclei. Moreover, their diameter shouldnot be larger than one third of the diameter of the cell nucleus and the micronuclearboundary should be distinguishable from the nuclear one.
2.5. Statistical analysis
The statistical analysis was performed using the Fisher’s exact test to deter-mine significant differences in the results obtained. In order to study whether thebehaviour of XPC and XPA differed in relation to the time after UV-irradiation, weapplied the Poisson regression model followed by the �2 test. This model considersthe total recount as an offset. Tests and table contents were obtained using Graph-Pad Instant v3.05 (GraphPad Software Inc.) and SAS v9.1.3 (SAS Institute Inc.), andgraphics were obtained with Microsoft Office Excel 2007.
3. Results
3.1. NER factors are properly recruited to UV-induced CPDs in thecell nucleus but not in micronuclei
UV photoproducts are repaired by the NER pathway, which is avery versatile repair system as it allows different types of single-strand lesions and bulky DNA adducts to be removed [13]. To inducelocal DNA damage, normal human fibroblasts were covered with apolycarbonate filter while they were exposed to UV-C. UV-damageinduction was confirmed by scoring the presence of CPD labelledspots in the cell nucleus (Fig. 1a). First of all, we performed an exper-iment to ascertain the most appropriate UV-C dose for detecting thepresence of XPC at the UV-damaged sites. Under the immunoflu-orescence conditions applied in our study, we established that60 J/m2 was the most suitable dose for detecting XPC at CPD labelledsites in normal human fibroblasts. In this experiment we also deter-mined that at 15 min after UV-C exposure 95% of cells (n = 100) hadrecruited XPC to the nuclear CPD labelled spots (Fig. 1b). We thenstudied XPA recruitment to the nuclear photolesions by means of aco-localisation study with XPC. At the same post-irradiation time,we observed that 98% of cells (n = 100) recruited XPA properly to thenuclear XPC labelled sites (Fig. 1c). These results show that, in thiscell system and under the present experimental conditions, XPCand XPA factors rapidly accumulate to locally UV-exposed areaswithin the cell nucleus. According to Houtsmuller et al. [17] andRademakers et al. [18], this accumulation results from collisionsbetween the NER proteins that move around within the cell nucleusand photolesions [17,18].
As the aim of this study was to determine the capacity of theDNA damage response machinery to detect and repair micronu-clear DNA lesions, we analysed the presence of XPC and XPA insideradiation-induced micronuclei. To determine whether performinga tridimensional analysis of the samples would improve data col-lection we examined the samples with both an epifluorescent anda confocal microscope. However, no qualitative differences werefound (data not shown), and the amount of protein detected withthe epifluorescent microscope did not differ from that detectedwith the confocal microscope. For these reasons, we decided to per-
form all the analyses with conventional epifluorescent microscopy.In these experiments we avoided the use of polycarbonate fil-ters in order to increase the number of micronuclei exposed toUV light. In agreement with this irradiation pattern, 15 min after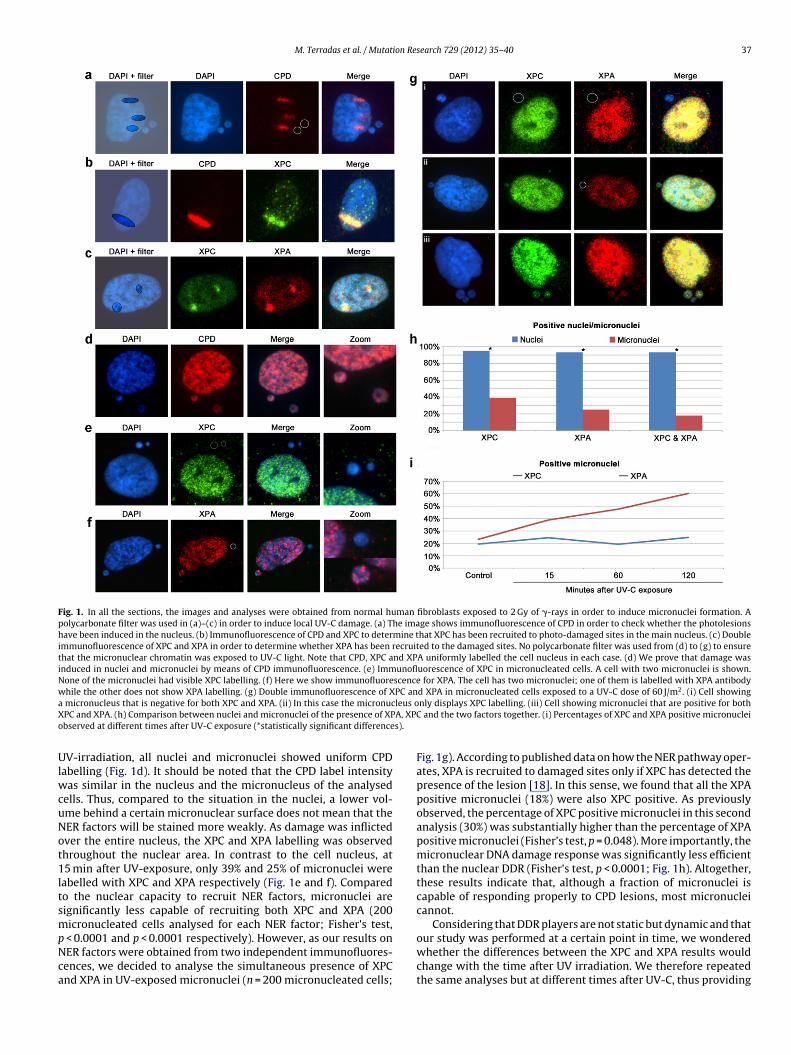
M. Terradas et al. / Mutation Research 729 (2012) 35– 40 37
Fig. 1. In all the sections, the images and analyses were obtained from normal human fibroblasts exposed to 2 Gy of �-rays in order to induce micronuclei formation. Apolycarbonate filter was used in (a)–(c) in order to induce local UV-C damage. (a) The image shows immunofluorescence of CPD in order to check whether the photolesionshave been induced in the nucleus. (b) Immunofluorescence of CPD and XPC to determine that XPC has been recruited to photo-damaged sites in the main nucleus. (c) Doubleimmunofluorescence of XPC and XPA in order to determine whether XPA has been recruited to the damaged sites. No polycarbonate filter was used from (d) to (g) to ensurethat the micronuclear chromatin was exposed to UV-C light. Note that CPD, XPC and XPA uniformly labelled the cell nucleus in each case. (d) We prove that damage wasinduced in nuclei and micronuclei by means of CPD immunofluorescence. (e) Immunofluorescence of XPC in micronucleated cells. A cell with two micronuclei is shown.None of the micronuclei had visible XPC labelling. (f) Here we show immunofluorescence for XPA. The cell has two micronuclei; one of them is labelled with XPA antibodywhile the other does not show XPA labelling. (g) Double immunofluorescence of XPC and XPA in micronucleated cells exposed to a UV-C dose of 60 J/m2. (i) Cell showinga leus oX A, XPCo s).
UlwcuNot1ltsmpNca
micronucleus that is negative for both XPC and XPA. (ii) In this case the micronucPC and XPA. (h) Comparison between nuclei and micronuclei of the presence of XPbserved at different times after UV-C exposure (*statistically significant difference
V-irradiation, all nuclei and micronuclei showed uniform CPDabelling (Fig. 1d). It should be noted that the CPD label intensity
as similar in the nucleus and the micronucleus of the analysedells. Thus, compared to the situation in the nuclei, a lower vol-me behind a certain micronuclear surface does not mean that theER factors will be stained more weakly. As damage was inflictedver the entire nucleus, the XPC and XPA labelling was observedhroughout the nuclear area. In contrast to the cell nucleus, at5 min after UV-exposure, only 39% and 25% of micronuclei were
abelled with XPC and XPA respectively (Fig. 1e and f). Comparedo the nuclear capacity to recruit NER factors, micronuclei areignificantly less capable of recruiting both XPC and XPA (200icronucleated cells analysed for each NER factor; Fisher’s test,
< 0.0001 and p < 0.0001 respectively). However, as our results onER factors were obtained from two independent immunofluores-ences, we decided to analyse the simultaneous presence of XPCnd XPA in UV-exposed micronuclei (n = 200 micronucleated cells;
nly displays XPC labelling. (iii) Cell showing micronuclei that are positive for both and the two factors together. (i) Percentages of XPC and XPA positive micronuclei
Fig. 1g). According to published data on how the NER pathway oper-ates, XPA is recruited to damaged sites only if XPC has detected thepresence of the lesion [18]. In this sense, we found that all the XPApositive micronuclei (18%) were also XPC positive. As previouslyobserved, the percentage of XPC positive micronuclei in this secondanalysis (30%) was substantially higher than the percentage of XPApositive micronuclei (Fisher’s test, p = 0.048). More importantly, themicronuclear DNA damage response was significantly less efficientthan the nuclear DDR (Fisher’s test, p < 0.0001; Fig. 1h). Altogether,these results indicate that, although a fraction of micronuclei iscapable of responding properly to CPD lesions, most micronucleicannot.
Considering that DDR players are not static but dynamic and that
our study was performed at a certain point in time, we wonderedwhether the differences between the XPC and XPA results wouldchange with the time after UV irradiation. We therefore repeatedthe same analyses but at different times after UV-C, thus providing
3 ion Re
kmmaniceiaXplttrlstpsttbtwn
mDsimsat
3f
atocwmlwCtcoetmrisodspmlm
8 M. Terradas et al. / Mutat
inetic modelling of the dynamics of XPA and XPC to reachicronuclear photolesions. For each protein, three groups, with 200icronucleated cells in each, were analysed at 15, 60 and 120 min
fter exposure to UV-C. An additional sample of 200 micronucleatedon-UV-exposed cells was used as the control group. This exper-
ment allowed us to observe that, under these external stressfulonditions, XPC and XPA accumulation in micronuclei was differ-nt (�2 test, p = 0.0002). Although slight, a progressive increasen XPC positive micronuclei was observed during the first 2 hfter UV exposure (�2 test, p < 0.001). In contrast, the number ofPA-positive micronuclei remained constant over time (�2 test,
= 0.8170; Fig. 1i). This difference might be due to the subcellu-ar location of the two proteins and how this changes in responseo stressful conditions. Under unchallenged conditions, XPC con-inuously shuttles between the nucleoplasm and the cytosol. Inesponse to UV irradiation, the export of XPC is impeded and thiseads to a temporary increase in XPC in the nucleus [19]. Thistress-dependent retention of XPC in micronuclei would favourhe observed accumulation and its subsequent collision with DNAhotolesions generated in micronuclei. Unlike XPC, XPA does nothuttle between the nucleus and the cytoplasm; therefore, in ordero collide and accumulate at DNA damage sites, it needs to be inhe nucleoplasm before UV irradiation [18]. Consequently it muste imported into the nucleus after nuclear envelope reorganisa-ion at the end of mitosis with the bulk of the nuclear proteins. Thisould explain why the number of XPA positive micronuclei doesot increase over time.
Overall, the results demonstrate that lesions induced in theicronuclear DNA are much less capable of triggering an adequateDR than nuclear injuries are. In this sense, although it has been
hown that XPC is more frequently found in micronuclei than XPA,n terms of protein recruitment to the photolesions these factors
obilise similarly in the nucleus and in the micronucleus. Con-equently, as only a small fraction of micronuclei show properccumulation of these repair factors to photolesions, it is possiblehat there are structural micronuclear deficiencies.
.2. An impaired micronuclear DNA damage response resultsrom an abnormal micronuclear envelope
Morphological observations of micronuclei in cancer cell linesnd of those that have arisen due to aneugenic treatments showedhat the envelope of micronuclei is abnormal compared to thatf nuclei [5,20,21]. To check whether defective DDR in micronu-lei could be related to alterations in their envelope structure,e decided to study some morphological features of the nuclearembrane of radiation-induced micronuclei. The nuclear enve-
ope is composed of two lipid bilayers and the nuclear lamina,hich consists of intermediate filaments (lamins A, B1, B2 and) that participate in nuclear stability and chromatin organisa-ion. The two nuclear membranes fuse to form the nuclear poreomplexes (NPC), composed of multiple copies of different nucle-porins (Nups), which mediate macromolecule traffic through thenvelope. Out of all these nuclear envelope proteins, we studiedhe presence of Lamin-B1 and some Nups (for the latter we used a
onoclonal antibody that recognises the conserved domain FXFGepeats in nucleoporins; Fig. 2a). As expected, all nuclei were pos-tive for both markers. One hundred micronucleated cells wereelected to analyse micronuclear envelope features. While 86.4%f micronuclei showed nucleus-like Lamin-B1 labelling, only 32.2%isplayed Nups labelling with an intensity similar to the nuclei. Ithould also be noted that all the micronuclei observed to be Nups
ositive were also Lamin-B1 labelled. These results show that likeicronuclei originated by chemical treatments or in cancer cellines, radiation-induced micronuclei generally have an abnormalicronuclear envelope; in some cases this is due to the lack of
search 729 (2012) 35– 40
Lamin-B1, but it is mainly due to the absence or reduced density ofNPCs.
To obtain insight into the functionality of the micronuclearenvelope, we checked for the presence of cyclin D1 inside micronu-clei (Fig. 2b). This protein has to be imported into the nucleus,and therefore into the micronucleus, to commit the cell to DNAreplication. Once cells enter S-phase, cyclin D1 is exported to thecytoplasm to be degraded. In this experiment, two groups of irra-diated and non-irradiated cells, each with 200 cells, were checked.Whereas 67.5% of nuclei were positive in the non-irradiated group,85% were cyclin D1 labelled at 48 h after ionising-irradiation. Thisincrease reflects the cell-cycle delay caused by irradiation. To checkthe presence of cyclin D1 in micronuclei, only the irradiated groupof cells was examined. Two hundred micronucleated cells dis-playing cyclin D1 in the nucleus were analysed. Out of a total of237 micronuclei, 54 were positive and 183 were devoid of cyclinD1 labelling; thus, only 22.8% of micronuclei showed cyclin D1.This percentage is not significantly different from that of XPC/XPApositive micronuclei (Fisher’s test, p = 0.4839). Therefore, theseresults demonstrate that abnormalities in protein transport intomicronuclei affect proteins that need to be shuttled through themicronuclear envelope.
The observed reduction or total absence of Nups in themicronuclear envelope could be responsible for the impaired NERprotein recruitment observed in micronuclei subjected to UV-irradiation. To test this possibility, we performed a XPC/Nupsdouble immunodetection on two groups of non-exposed and UV-exposed micronucleated cells (n = 200; Fig. 2c). The fraction of XPCpositive micronuclei increased from 17.83% to 23.95% after UV-Cexposure (Fig. 2d and e). As outlined above, this increase couldbe due to the accumulation of XPC in micronuclei under externalgenotoxic stress. As expected, the number of micronuclei showingnuclear-like Nups labelling was similar in the UV-irradiated andnon-irradiated groups (51.7% and 56.1% respectively; Fisher’s test,p = 0.35). However, and more importantly, the fraction of micronu-clei devoid of Nups labelling did not show XPC labelling eitherin the irradiated or in the non-irradiated cells. This result indi-cates that a defect in the structure of the micronuclear envelopecauses the observed deficiency in the cell response to micronuclearUV-C photolesions. Moreover, not all the Nups positive micronu-clei were also XPC positive (39.1% and 28.6% of UV-C exposedmicronuclei with visible Nups labelling lacked XPC in the controland in the UV-irradiated group respectively). These results indi-cate that even if the structural proteins of NPCs are visible undera fluorescence microscope, they do not necessarily form functionalpore complexes. In conclusion, the results demonstrate that NPCsof most micronuclei are scarce or not functional, which explainswhy the NER factors are not found in most of the UV-exposedmicronuclei.
4. Discussion
To restore genome integrity, the cell must overcome impor-tant hurdles in space and time to rapidly sense and initiate thecorrect signalling and repair programs [22]. For instance, the con-densed nature of heterochromatin acts as a barrier to factors thatprocess DNA lesions, and therefore some chromatin decondensa-tion steps are necessary for DSB repair in heterochromatin [23,24].The work presented here shows that the lesions produced in themicronuclear DNA are almost incapable of generating an effec-tive response to DNA damage. While XPC and XPA are properly
recruited in photolesions induced in the nucleus, they are hardlyever found in micronuclei exposed to UV light. This suggests that aneffective cell response for maintaining genome integrity not onlydepends on the proper coordination in space and time of the DDR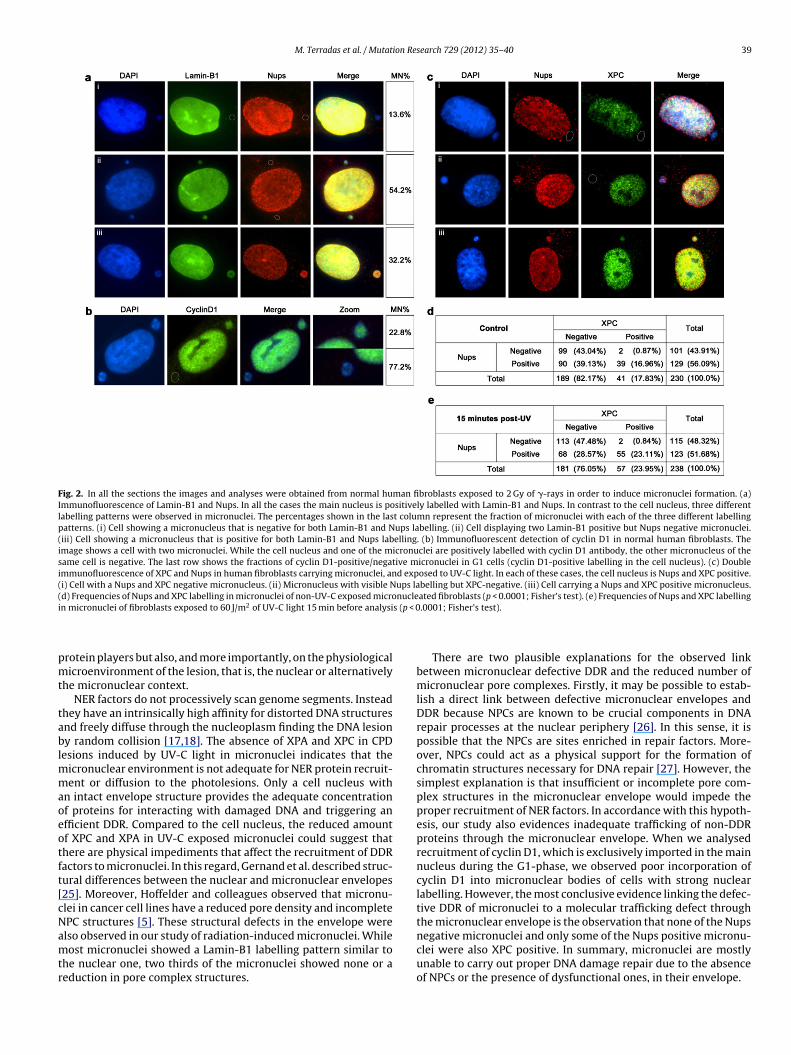
M. Terradas et al. / Mutation Research 729 (2012) 35– 40 39
Fig. 2. In all the sections the images and analyses were obtained from normal human fibroblasts exposed to 2 Gy of �-rays in order to induce micronuclei formation. (a)Immunofluorescence of Lamin-B1 and Nups. In all the cases the main nucleus is positively labelled with Lamin-B1 and Nups. In contrast to the cell nucleus, three differentlabelling patterns were observed in micronuclei. The percentages shown in the last column represent the fraction of micronuclei with each of the three different labellingpatterns. (i) Cell showing a micronucleus that is negative for both Lamin-B1 and Nups labelling. (ii) Cell displaying two Lamin-B1 positive but Nups negative micronuclei.(iii) Cell showing a micronucleus that is positive for both Lamin-B1 and Nups labelling. (b) Immunofluorescent detection of cyclin D1 in normal human fibroblasts. Theimage shows a cell with two micronuclei. While the cell nucleus and one of the micronuclei are positively labelled with cyclin D1 antibody, the other micronucleus of thesame cell is negative. The last row shows the fractions of cyclin D1-positive/negative micronuclei in G1 cells (cyclin D1-positive labelling in the cell nucleus). (c) Doubleimmunofluorescence of XPC and Nups in human fibroblasts carrying micronuclei, and exposed to UV-C light. In each of these cases, the cell nucleus is Nups and XPC positive.(i) Cell with a Nups and XPC negative micronucleus. (ii) Micronucleus with visible Nups labelling but XPC-negative. (iii) Cell carrying a Nups and XPC positive micronucleus.(d) Frequencies of Nups and XPC labelling in micronuclei of non-UV-C exposed micronucleated fibroblasts (p < 0.0001; Fisher’s test). (e) Frequencies of Nups and XPC labellingi s (p < 0
pmt
tablmmaoeotft[cNamtr
n micronuclei of fibroblasts exposed to 60 J/m2 of UV-C light 15 min before analysi
rotein players but also, and more importantly, on the physiologicalicroenvironment of the lesion, that is, the nuclear or alternatively
he micronuclear context.NER factors do not processively scan genome segments. Instead
hey have an intrinsically high affinity for distorted DNA structuresnd freely diffuse through the nucleoplasm finding the DNA lesiony random collision [17,18]. The absence of XPA and XPC in CPD
esions induced by UV-C light in micronuclei indicates that theicronuclear environment is not adequate for NER protein recruit-ent or diffusion to the photolesions. Only a cell nucleus with
n intact envelope structure provides the adequate concentrationf proteins for interacting with damaged DNA and triggering anfficient DDR. Compared to the cell nucleus, the reduced amountf XPC and XPA in UV-C exposed micronuclei could suggest thathere are physical impediments that affect the recruitment of DDRactors to micronuclei. In this regard, Gernand et al. described struc-ural differences between the nuclear and micronuclear envelopes25]. Moreover, Hoffelder and colleagues observed that micronu-lei in cancer cell lines have a reduced pore density and incompletePC structures [5]. These structural defects in the envelope were
lso observed in our study of radiation-induced micronuclei. Whileost micronuclei showed a Lamin-B1 labelling pattern similar tohe nuclear one, two thirds of the micronuclei showed none or aeduction in pore complex structures.
.0001; Fisher’s test).
There are two plausible explanations for the observed linkbetween micronuclear defective DDR and the reduced number ofmicronuclear pore complexes. Firstly, it may be possible to estab-lish a direct link between defective micronuclear envelopes andDDR because NPCs are known to be crucial components in DNArepair processes at the nuclear periphery [26]. In this sense, it ispossible that the NPCs are sites enriched in repair factors. More-over, NPCs could act as a physical support for the formation ofchromatin structures necessary for DNA repair [27]. However, thesimplest explanation is that insufficient or incomplete pore com-plex structures in the micronuclear envelope would impede theproper recruitment of NER factors. In accordance with this hypoth-esis, our study also evidences inadequate trafficking of non-DDRproteins through the micronuclear envelope. When we analysedrecruitment of cyclin D1, which is exclusively imported in the mainnucleus during the G1-phase, we observed poor incorporation ofcyclin D1 into micronuclear bodies of cells with strong nuclearlabelling. However, the most conclusive evidence linking the defec-tive DDR of micronuclei to a molecular trafficking defect throughthe micronuclear envelope is the observation that none of the Nups
negative micronuclei and only some of the Nups positive micronu-clei were also XPC positive. In summary, micronuclei are mostlyunable to carry out proper DNA damage repair due to the absenceof NPCs or the presence of dysfunctional ones, in their envelope.
4 ion Re
ccpmfmhrtUtetisrDbcs
i[ccimntaibccot
C
A
lGME
tAaf
R
[
[
[
[
[
[
[[
[
[
[
[
[
[
[
[
[
0 M. Terradas et al. / Mutat
Inconsistent results have been reported so far regarding theapacity of micronuclear DSBs to recruit DDR factors. These dis-repancies have been attributed to the dynamics between theseroteins and the damaged DNA, which might be different inicronuclei and nuclei [8], or to the residual presence of these
actors in the damaged chromatin before it is wrapped insideicronuclear bodies [9]. Here we demonstrate that, although they
ave low efficiency compared to the nucleus, the NER factors can beecruited inside micronuclear bodies after damage induction, sincehe number of positive XPC micronuclei increase over time afterVC exposure. But most importantly, here we evidence that pho-
olesions induced in micronuclei are mostly unable to induce anffective DNA damage response, and demonstrate that the struc-ure of the micronuclear envelope is a determinant factor thatnfluences the repair of chromatin entrapped in micronuclei. Inummary, our results demonstrate that the DDR machinery is noteady for action anytime or anywhere. In the case of micronuclearNA lesions, the chromatin encapsulated in micronuclei does notenefit from the intricate and efficient web of DDR players of theell and thus mutagenesis might be favoured under these circum-tances.
It has recently been observed that some cells carrying radiation-nduced micronuclei are able to undergo a successful cell division28]. Moreover, these authors also observed that micronucleatedells have a predisposition to produce micronucleated daughterells after cell division. Therefore, considering this scenario, its possible that inefficient repair of DNA contained in persisting
icronuclei could contribute to carcinogenesis. This statement isot surprising when epidemiological studies have already shownhat the presence of micronuclei in a healthy population indicates
higher risk of cancer development in the future [6]. Therefore, its possible that on some occasions micronuclei, which have mainlyeen considered as indicators of ongoing chromosome instability,ould also be a source of instability. In this sense, whereas canceran be attributed to an accumulation of mutations in the genomef actively replicating cells, our results reveal a new dimension inhe significance of micronucleation in carcinogenic processes.
onflicts of interest statement
The authors declare that there are no conflicts of interest.
cknowledgements
This work was supported by grants from Instituto de Salud Car-os III, Grant number RD06/0020/1020; Generalitat de Catalunya,rant number 2009 SGR-282 and Consejo de Seguridad Nuclear..T. is recipient of a predoctoral fellowship from the Ministerio de
ducación y Ciencia (Grant number FPU2006-01805).We thank Jordi Surrallés’ group for help with UV-C irradia-
ion, the Language Advisory & Translation Unit at the Universitatutònoma de Barcelona Language Service for editing the manuscriptnd the Statistics’ Service at the Universitat Autònoma de Barcelonaor carrying out the tests of XPA and XPC kinetics.
eferences
[1] S.E. Artandi, R.A. DePinho, A critical role for telomeres in suppressing and facil-itating carcinogenesis, Curr. Opin. Genet. Dev. 10 (2000) 39–46.
[2] D. Gisselsson, J. Bjork, M. Hoglund, F. Mertens, P. Dal Cin, M. Akerman, N. Man-dahl, Abnormal nuclear shape in solid tumors reflects mitotic instability, Am.J. Pathol. 158 (2001) 199–206.
[
[
search 729 (2012) 35– 40
[3] B. McClintock, The stability of broken ends of chromosomes in zea mays, Genet-ics 26 (1941) 234–282.
[4] J. Pampalona, D. Soler, A. Genesca, L. Tusell, Whole chromosome loss is pro-moted by telomere dysfunction in primary cells, Genes Chromosomes Cancer49 (2010) 368–378.
[5] D.R. Hoffelder, L. Luo, N.A. Burke, S.C. Watkins, S.M. Gollin, W.S. Saunders,Resolution of anaphase bridges in cancer cells, Chromosoma 112 (2004)389–397.
[6] S. Bonassi, A. Znaor, M. Ceppi, C. Lando, W.P. Chang, N. Holland, M.Kirsch-Volders, E. Zeiger, S. Ban, R. Barale, M.P. Bigatti, C. Bolognesi, A. Cebulska-Wasilewska, E. Fabianova, A. Fucic, L. Hagmar, G. Joksic, A. Martelli, L. Migliore,E. Mirkova, M.R. Scarfi, A. Zijno, H. Norppa, M. Fenech, An increased micronu-cleus frequency in peripheral blood lymphocytes predicts the risk of cancer inhumans, Carcinogenesis 28 (2007) 625–631.
[7] T. Haaf, E. Raderschall, G. Reddy, D.C. Ward, C.M. Radding, E.I. Golub, Sequestra-tion of mammalian Rad51-recombination protein into micronuclei, J. Cell Biol.144 (1999) 11–20.
[8] N.G. Medvedeva, I.V. Panyutin, I.G. Panyutin, R.D. Neumann, Phosphorylationof histone H2AX in radiation-induced micronuclei, Radiat. Res. 168 (2007)493–498.
[9] T. Yoshikawa, G. Kashino, K. Ono, M. Watanabe, Phosphorylated H2AX foci intumor cells have no correlation with their radiation sensitivities, J. Radiat. Res.(Tokyo) 50 (2009) 151–160.
10] M. Terradas, M. Martin, L. Tusell, A. Genesca, DNA lesions sequestered inmicronuclei induce a local defective-damage response, DNA Repair (Amst.) 8(2009) 1225–1234.
11] S.V. Costes, I. Chiolo, J.M. Pluth, M.H. Barcellos-Hoff, B. Jakob, Spatiotempo-ral characterization of ionizing radiation induced DNA damage foci and theirrelation to chromatin organization, Mutat. Res. 704 (2010) 78–87.
12] G.P. Pfeifer, Y.H. You, A. Besaratinia, Mutations induced by ultraviolet light,Mutat. Res. 571 (2005) 19–31.
13] L.C. Gillet, O.D. Scharer, Molecular mechanisms of mammalian global genomenucleotide excision repair, Chem. Rev. 106 (2006) 253–276.
14] C. Dinant, A.B. Houtsmuller, W. Vermeulen, Chromatin structure and DNA dam-age repair, Epigenetics Chromatin. 1 (2008) 9.
15] M. Volker, M.J. Mone, P. Karmakar, A. van Hoffen, W. Schul, W. Vermeulen,J.H. Hoeijmakers, R. van Driel, A.A. van Zeeland, L.H. Mullenders, Sequentialassembly of the nucleotide excision repair factors in vivo, Mol. Cell 8 (2001)213–224.
16] M. Fenech, The in vitro micronucleus technique, Mutat. Res. 455 (2000) 81–95.17] A.B. Houtsmuller, S. Rademakers, A.L. Nigg, D. Hoogstraten, J.H. Hoeijmakers,
W. Vermeulen, Action of DNA repair endonuclease ERCC1/XPF in living cells,Science 284 (1999) 958–961.
18] S. Rademakers, M. Volker, D. Hoogstraten, A.L. Nigg, M.J. Mone, A.A. Van Zeeland,J.H. Hoeijmakers, A.B. Houtsmuller, W. Vermeulen, Xeroderma pigmentosumgroup A protein loads as a separate factor onto DNA lesions, Mol. Cell Biol. 23(2003) 5755–5767.
19] D. Hoogstraten, S. Bergink, J.M. Ng, V.H. Verbiest, M.S. Luijsterburg, B. Geverts, A.Raams, C. Dinant, J.H. Hoeijmakers, W. Vermeulen, A.B. Houtsmuller, VersatileDNA damage detection by the global genome nucleotide excision repair proteinXPC, J. Cell Sci. 121 (2008) 2850–2859.
20] B. Labidi, M. Gregoire, S. Frackowiak, D. Hernandez-Verdun, M. Bouteille, RNApolymerase activity in PtK1 micronuclei containing individual chromosomes.an in vitro and in situ study, Exp. Cell Res. 169 (1987) 233–244.
21] G. Geraud, F. Laquerriere, C. Masson, J. Arnoult, B. Labidi, D. Hernandez-Verdun,Three-dimensional organization of micronuclei induced by colchicine in PtK1cells, Exp. Cell Res. 181 (1989) 27–39.
22] C. Lukas, J. Bartek, J. Lukas, Imaging of protein movement induced by chromo-somal breakage: tiny ‘local’ lesions pose great ‘global’ challenges, Chromosoma114 (2005) 146–154.
23] M. Falk, E. Lukasova, S. Kozubek, Chromatin structure influences the sen-sitivity of DNA to gamma-radiation, Biochim. Biophys. Acta 1783 (2008)2398–2414.
24] M. Falk, E. Lukasova, S. Kozubek, Higher-order chromatin structure in DSBinduction, repair and misrepair, Mutat. Res. 704 (2010) 88–100.
25] D. Gernand, T. Rutten, A. Varshney, M. Rubtsova, S. Prodanovic, C. Bruss, J. Kum-lehn, F. Matzk, A. Houben, Uniparental chromosome elimination at mitosis andinterphase in wheat and pearl millet crosses involves micronucleus forma-tion, progressive heterochromatinization, and DNA fragmentation, Plant Cell17 (2005) 2431–2438.
26] P. Therizols, C. Fairhead, G.G. Cabal, A. Genovesio, J.C. Olivo-Marin, B. Dujon,E. Fabre, Telomere tethering at the nuclear periphery is essential for efficientDNA double strand break repair in subtelomeric region, J. Cell Biol. 172 (2006)189–199.
27] K. Mekhail, D. Moazed, The nuclear envelope in genome organization, expres-sion and stability, Nat. Rev. Mol. Cell Biol. 11 (2010) 317–328.
28] Y. Huang, H. Hou, Q. Yi, Y. Zhang, D. Chen, E. Jiang, Y. Xia, M. Fenech, Q. Shi, Thefate of micronucleated cells post X-irradiation detected by live cell imaging,DNA Repair (Amst.) 10 (2011) 629–638.
Related Documents











